

Abstract
Suboptimal choice behavior, or behavior that leads to a loss of resources over time, has been observed in a laboratory setting from multiple species. A procedure commonly used to capture this effect involves presenting two alternatives during choice trials, one of which is optimal whereas the other is suboptimal. The optimal alternative yields reinforcement more often than the suboptimal alternative, but often does not produce signals that indicate whether reinforcement will occur. The suboptimal alternative produces less reinforcement than the optimal alternative but may include reinforcement-predictive stimuli that indicate to the organism whether reinforcement will occur. This procedural framework has consistently produced a preference for the suboptimal alternative in pigeons and, to a lesser extent, rats. However, human participants have demonstrated preference for the optimal alternative. Following a review of past suboptimal choice research, we applied the reinforcement-derived definition of optimality to two sets of our previously published human data. We found that under multiple conditions, human choice behavior was consistent with what was predicted by the proportion of obtained reinforcement, thus supporting that the behavior was optimal. However, we found that participants in two conditions chose the suboptimal alternative more than expected. This finding could be considered as a demonstration of suboptimal choice in humans. We propose that comparing choice behavior to what past obtained reinforcement outcomes would predict might be a more accurate view of whether patterns of choice are within the parameters of this task.
Keywords: Decision making, Suboptimal choice, Matching law
Suboptimal behavior can be defined as making choices that result in a relative net loss of resources over time. A key component of this definition is the emphasis on the net result over the long term, rather than on the outcome following any single instance. Thus, the focus is on a pattern of choice across time (Baum, 2002) that results in a net loss relative to another pattern of choice. Human gambling behavior is one example of suboptimal behavior in that a single trial, a pull on a slot machine, for example, may have a winning outcome. However, over many trials, individuals frequently lose more than they gain by choosing to gamble.
Although it is appropriate to acknowledge that some contributing factors to human gambling behavior may be related to social influences (Russell et al., 2018; Sirola et al., 2021) and economic status (Fu et al., 2021), the puzzle becomes more complex when we find that nonhuman animals have been observed engaging in similar patterns of suboptimal behavior. Thus, any explanation for suboptimal behavior that is uniquely applicable to humans may be insufficient to be the sole mechanism.
In the laboratory, a suboptimal pattern of behavior has been defined by the selection of the lower-payoff alternative more often than expected by chance. That is, given two alternatives that result in differing amounts of reinforcement, choosing the lower-payoff alternative more than 50% of the time has been described as suboptimal performance (e.g., Gipson et al., 2009; Stagner & Zentall, 2010). One presumes that the definition of an optimal pattern is selecting the higher-payoff alternative more often than expected by chance. By this definition, pigeons have been found to choose suboptimally (e.g., Stagner & Zentall, 2010; Zentall & Stagner, 2011). In contrast, human participants have consistently chosen the optimal alternative more than expected by chance (McDevitt et al., 2019; Stagner et al., 2020; Bodily et al., 2024). We suggest that the chance-based definitions of optimal and suboptimal patterns of behavior are imprecise categories for capturing patterns of behavior that produce a net loss relative to other patterns. Instead, we propose comparing the observed pattern of choice to that which would be expected based on upon reinforcement obtained within the task. If the suboptimal alternative is selected more than would be predicted from reinforcement earned, then this behavior could be considered suboptimal.
To explore this, we will first look at a choice task design used for testing suboptimal choice behavior, results from a variety of species on this task, some of the manipulations that have been done to this task, and their effects on choice behavior. We will then focus on human choice behavior observed within this task. In particular, we will examine obtained human choice data in comparison to predictions based on reinforcement outcomes experienced.
Suboptimal Choice Task
Pigeons have shown a strong preference for reinforcement-predictive stimuli over those that do not predict reinforcement, even when there was no additional food available for doing so (Roper & Zentall, 1999). Much of the subsequent choice research with pigeons has focused on whether a suboptimal preference for reinforcement-predictive stimuli would still be observed if there was a cost for doing so.
To assess the strength of the preference for reinforcement predictors, Stagner and Zentall (2010) used a concurrent chains procedure that manipulated the probabilities of reinforcement associated with the reinforcement-predictive and -unpredictive alternatives (see Fig. 1). The reinforcement-predictive alternative, also referred to as the suboptimal alternative, was associated with an S+ terminal stimulus on 20% of trials and an S- terminal stimulus on 80% of trials. Thus, reinforcement occurred on 20% of suboptimal alternative trials. On the other hand, the reinforcement-unpredictive alternative, also referred to as the optimal alternative, was associated with two terminal stimuli that were followed by reinforcement 50% of the time. Thus, reinforcement occurred on 50% of optimal trials. Pigeons were given both forced experience trials for each alternative as well as choice trials in which they could select either alternative. Pigeons demonstrated an almost exclusive preference for the suboptimal reinforcement-predictive alternative in this choice task, the mechanism behind which seems to be the predictive nature of the terminal stimuli because pigeons were indifferent between the two alternatives when neither provided reinforcement-predictive terminal links (see Unpredictive condition in Fig. 1). This pattern of suboptimal choice in the predictive terminal links condition resulted in a net loss of two and a half times the amount of food available (Stagner & Zentall, 2010). This finding has been reproduced, affirming that pigeons readily choose suboptimally under similar conditions (Spetch et al., 1990; Spetch et al., 1994; Zentall & Stagner, 2011; McDevitt et al., 2019).
Fig. 1.
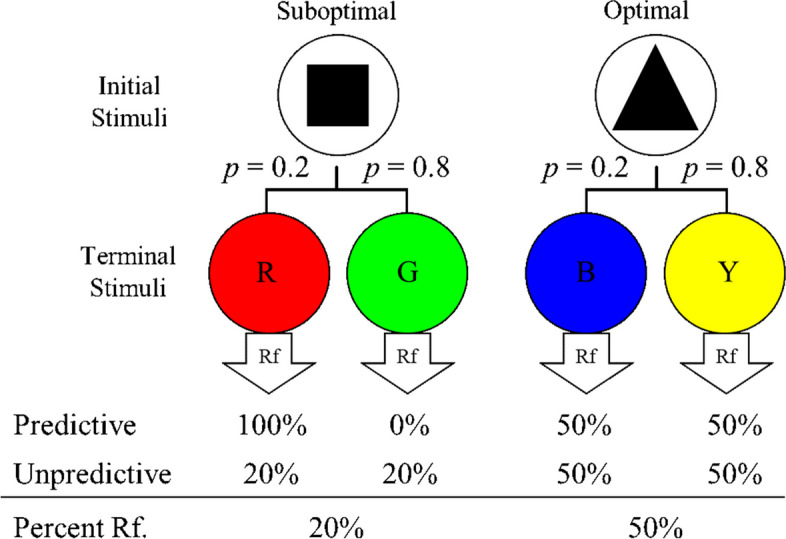
Stagner and Zentall (2010) Procedure. Note. Initial stimuli were on a FR1 response requirement. Terminal stimuli were counterbalanced. Terminal stimuli remained on for 10 s and followed by either a 10-s ITI on nonreinforced trials, or for 2-s reinforcement and 10-s ITI for reinforcement trials
Suboptimal choice has also been observed in rats (Chow et al., 2017; Cunningham & Shahan, 2019, 2020) and nonhuman primates (Blanchard et al., 2015; Smith et al., 2017), demonstrating that suboptimal behavior patterns can be observed in a variety of species under certain conditions. Recent studies have further explored factors that influence engagement in suboptimal choice behavior. Delay to reinforcement could be a contributing factor, and studies have been done that have manipulated both the response requirement and durations of the initial links (Kendall, 1974; Dunn & Spetch, 1990, Experiment 3; Zentall et al., 2017; Pisklak et al., 2019; Cunningham & Shahan, 2020) and the terminal links (Spetch et al., 1990; Belke & Spetch, 1994; Spetch et al., 1994). Although findings vary across studies, most results seem to support that extending the duration or increasing the response requirement of the initial link may reduce suboptimal preference in pigeons (Kendall, 1974; Pisklak et al., 2019; Zentall et al., 2017). In addition, these studies also observed an increase in intersubject variability when initial links required more responses or were on a higher VI/FI schedule. Work with rats suggests that when similar modifications are made to the initial links, resulting in delayed onset of food predictive terminal links, that suboptimal preference is not affected (Cunningham & Shahan, 2020).
Further, other studies with nonhumans have removed the predictive value from terminal links associated with the suboptimal alternative. That is, all terminal links in these tasks provided no additional information about reinforcement beyond what the initial links did (see Unpredictive condition in Fig. 1). When terminal links had no reinforcement predictive value, then pigeons (Spetch et al., 1990; Stagner & Zentall, 2010; Zentall & Stagner, 2011) as well as rhesus macaques (Smith et al., 2017; Smith & Beran, 2020) have been found to choose the optimal alternative. Similar studies, one with pigeons and one with monkeys, aimed at observing preferences for reinforcement-predictive stimuli have found preference for those predictors, even when there was no additional reinforcement for doing so (with pigeons, Roper & Zentall, 1999; with monkeys, Blanchard et al., 2015).
Human Performance on Suboptimal Choice Tasks
Although suboptimal behavior has been readily found in pigeons and other nonhuman species under certain conditions, one species that seems resistant to the propensity to choose suboptimally when given this task has been humans. Human participants prefer the optimal alternative regardless of whether the terminal links have reinforcement-predictive value (McDevitt et al., 2019; Stagner et al., 2020; Bodily et al., 2024), or the context in which the task is presented (Bodily et al., 2024).
To begin our investigation of human performance in more detail, we will first consider Molet et al. (2012). College students were selected to participate in this study based upon their tendencies toward gambling behavior that they self-reported on a screening survey in their introductory psychology course. Participants that had reported engaging in higher levels of gambling behavior selected the suboptimal alternative 56.5% of the time, whereas participants that reported no/low levels of gambling behavior chose the suboptimal alternative 23% of the time. Thus, although Molet et al. (2012) found that participants that had higher self-reported rates of gambling chose suboptimally more often than participants that did not self-report gambling behavior, these participants selected the suboptimal alternative a modest 56.5% of the time.
In a more direct replication of the design used to study pigeons by Zentall and Stagner (2011), McDevitt et al. (2019) tested college student participants on a computerized version of the task. In this study, there was a Signaled condition and an Unsignaled condition. In the Signaled condition, both the suboptimal and optimal alternatives provided reinforcement-predictive terminal links. In particular, the optimal alternative was associated with two terminal links that signaled a 3-point reward. The suboptimal alternative was associated with an S+ that occurred on 20% of suboptimal trials and yielded a 10-point reward, whereas the S- occurred on 80% of suboptimal trials and provided no points. For the Unsignaled condition, the suboptimal alternative was associated with a single terminal link that either ended with a 10-point reward on 20% of suboptimal trials or ended with no reward on 80% of suboptimal trials. The optimal alternative was associated with two terminal links, but each provided a 3-point reward on every optimal trial. Thus, for both Signaled and Unsignaled conditions, the reinforcement associated with the suboptimal and optimal alternatives was the same. Participants were split into one of four conditions: receiving the Signaled condition only, receiving the Unsignaled condition only, receiving the Signaled condition first followed the Unsignaled condition, or receiving the Unsignaled condition first followed by the Signaled condition. The main question was whether the predictive signals for reinforcement would affect human performance, as it has been found that pigeons will choose optimally if the suboptimal alternative does not have reinforcement-predictive terminal links (Stagner & Zentall, 2010; Zentall & Stagner, 2011). McDevitt et al. (2019) found that, consistent with Molet et al. (2012), human participants chose the optimal alternative on 74% of choice trials on average. In addition, there was no significant difference in choice trials between participants in the four conditions. Thus, neither the reinforcement-predictive signals, nor the order in which participants received the Unsignaled or Signaled procedures, influenced preference for the optimal alternative.
The role of reinforcement-predictive stimuli was further explored by Stagner et al. (2020, Experiment 2) with the removal of terminal links altogether. In particular, college student participants were presented with a computerized version of a suboptimal task similar to Stagner and Zentall’s (2010) procedure. Participants were split into three conditions: reinforcement-predictive terminal stimuli, reinforcement-unpredictive terminal stimuli, and no terminal stimuli. Stagner et al. (2020) found that there was no significant difference between these three groups, with participants demonstrating a preference for the optimal alternative on 62.5% of choice trials. This study reproduced the preference for the optimal alternative by humans found by Molet et al (2012) and McDevitt et al. (2019). Taken together, the findings suggest that unlike pigeons, human choice behavior in this task may not be influenced by signals for reinforcement.
It is possible that the stimuli used in this task, colors and shapes, are too abstract and not relevant enough for human participants to attend to them. Previous research has found that participants are more successful solving problems when they are presented using a social context rather than an abstract one. In particular, participants were found to solve a problem more readily when given stimuli that applied to a real-world construct (e.g., legal drinking age) than when they were abstract stimuli such as numbers and colors (Griggs & Cox, 1982). To explore this phenomenon in the context of the suboptimal choice task, Bodily et al. (2024) investigated whether framing the task in a real-world scenario might influence human choice. Participants experienced either the Abstract procedure with or without reinforcement-predictive stimuli (see Fig. 1), or a Narrative procedure with or without reinforcement-predictive stimuli (see Fig. 2). Thus, the groups were as follows: Abstract Predictive, Abstract Unpredictive, Narrative Predictive, and Narrative Unpredictive (see Fig. 2 for narrative conditions). With exception of the Narrative Unpredictive participants, there was a clear preference for the optimal alternative (Bodily et al., 2024). Participants in the Narrative Unpredictive condition were at chance performance, demonstrating no preference for either alternative. Findings from Bodily et al. (2024) add further support that the signals for reinforcement do not affect human choice behavior, as well as illuminating that applying a more real-world context to the task did not fundamentally affect choice behavior.
Fig. 2.
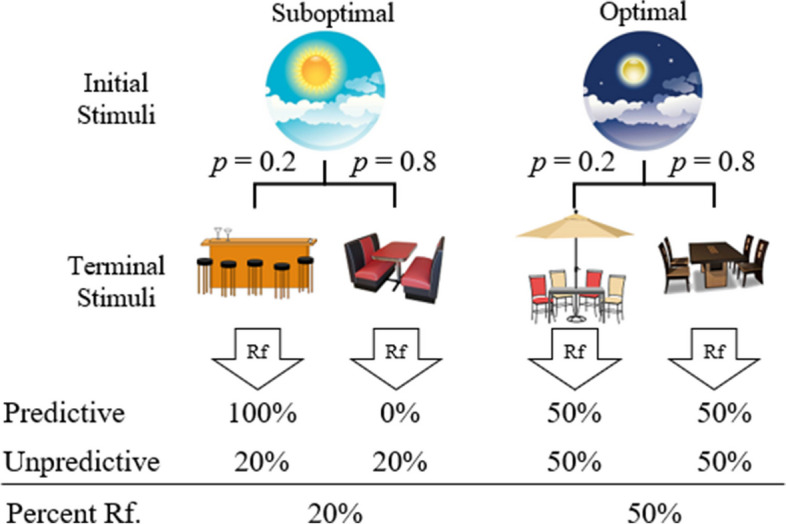
Bodily et al. (2024) Narrative Procedure. Note. Narrative Unpredictive and Narrative Predictive conditions are represented. Abstract Unpredictive and Abstract Predictive conditions used colors and shapes rather than picture stimuli, much like the Stagner and Zentall (2010) procedure shown in Fig. 1
Overall, humans have been more resistant to choosing suboptimally when compared to other species (Molet et al., 2012; McDevitt et al., 2019; Stagner et al., 2020; Bodily et al., 2024). However, although humans demonstrate a preference for the optimal alternative, they do not choose the optimal alternative exclusively. Depending on the experimental conditions, humans choose the suboptimal alternative 25%–45% of the time, which reduces the overall amount of reward possible to be obtained in the task.
This raises the question of what exactly optimal choice would look like within the context of this task. In some previous research, choice performance was compared to chance. Responding to one of the alternatives with sufficient preference that performance differed from chance was evidence of “optimal” or “suboptimal” preference (e.g., Hinnenkamp et al. 2017; Cunningham & Shahan, 2019; Stagner et al., 2020; Bodily et al., 2024). Pigeons show a clear preference for the suboptimal alternative using this metric. However, human participants show a preference for the optimal alternative when compared to chance. Perhaps optimal choice within this task might be defined in a way that could address the different preferences observed from human and nonhuman subjects by first exploring models that predict choice behavior.
There are existing models that have had some success in accounting for choice behavior but appear to be limited regarding human performance. For example, the signal for good news (SiGN) model predicts that choice behavior is reinforced when the delay to reward is reduced (for full review, see Dunn et al., 2024). The SiGN model assumes that conditioned stimuli, or reinforcement-predictive stimuli, play a role in choice behavior. However, McDevitt et al. (2019), Stagner et al. (2020), and Bodily et al. (2024) found that predictors of reinforcement did not affect human performance. In addition, the SiGN model also includes a temporal component in its calculation to account for the delay to reinforcement. Although pigeon research has found evidence that the duration of the terminal stimuli in the task can affect pigeons’ preference, there has been a different observation made from human participants. In Experiment 1 from Stagner et al. (2020), the duration of the terminal stimuli was manipulated such that one group received 2 s terminal durations, one received 8s terminal durations, and one received 20 s terminal durations. Participants in these three conditions all showed a modest preference for the optimal alternative, and most interestingly, there was no significant difference between the three conditions (Stagner et al., 2020). That is, the duration of the terminal link produced no systematic differences in human choice behavior. As mentioned previously, the SiGN model includes both reinforcement predictors and temporal information into its calculation, other models such as Fantino’s (1969) delay reduction hypothesis and Cunningham and Shahan’s (2018) temporal modeling also include one or both as well. This presents a challenge for application of these models to human data in that existing research has not found evidence that terminal-link durations and reinforcement predictors affect human suboptimal choice behavior. In addition, models like the SiGN model use programmed rates of reinforcement in the calculation. In doing so, this model does not allow for the incorporation of an individual participants’ reinforcement experience to predict choice behavior. Due to these factors, we decided to pursue examination of human performance in a different way.
If suboptimal choice is defined as a pattern of responding that produces a net loss relative to another pattern, there may be a more precise approach than establishing preference relative to chance. To define suboptimality, one must first define optimality. An extreme definition of optimality would be exclusive choice of the alternative associated with the higher probability (or proportion) of reward (see Molet et al., 2012). From that definition, any significant deviation from exclusive preference of the “optimal” alternative would be a suboptimal pattern. A more conservative definition of optimality is to consider the history of reinforcement for choosing each alternative. Optimality may be defined by the proportion of reinforcement obtained following choice of the “optimal” alternative. That is, the number of reinforcers following “optimal” choices is divided by the total number of reinforcers earned (e.g., Herrnstein, 1970). Choosing the optimal alternative less often than the obtained proportion reinforcement would be evidence of a suboptimal pattern of behavior. That is, the pattern of behavior would be suboptimal relative to the proportion of reinforcement experienced by the participant. Viewing choice behavior in this way has merit as it takes reinforcement effects following both forced and choice trials into consideration. Including forced trial reinforcement outcomes seems to be of importance as a recent study found that pigeons that experienced forced trials developed a strong preference for the suboptimal alternative whereas pigeons that did not have any forced trial experience showed no such preference (McDevitt et al., 2022).
To reexamine the optimality of human choice behavior, we applied the reinforcement-derived definition of optimality to two of our sets of human data (Stagner et al., 2020; Bodily et al., 2024). For each participant, the number of reinforcers earned following an “optimal” response was divided by the total number of reinforcers earned in each block of trials (including choice and forced-experience trials). The resulting proportion is the prediction for optimality for the succeeding block (see Baum, 1974, Equation 1). That is, the proportion of “optimal” reinforcers earned in Block 1 served as the predicted response allocation for Block 2. Therefore, there were no predictions for Block 1. In addition, reinforcers were accumulated across blocks. That is, reinforcers from Block 1 and Block 2 were included in the calculation of Block 3 optimal prediction. Plotting these proportions provided a comparison line against which we could compare the obtained choice data. First, we will focus on data from Stagner et al. (2020, Experiment 2), which manipulated presence or absence of terminal links (predictive/nonpredictive signals).
The presence or absence of terminal links did not affect preference for the optimal alternative (Stagner et al., 2020). Figure 3 shows that although group means of “optimal” responding were all greater than chance (50%), the means were also lower than reinforcement-derived optimality would predict. We can take a closer look at these data by comparing obtained choice to optimality to see if participants were choosing as optimally as past reinforcement experience would predict. Table 1 presents paired-sample t-tests (obtained choice against matching prediction) for Block 6. We found that obtained choice did not differ from matching prediction for participants that were given either reinforcement predictive or reinforcement nonpredictive terminal links. However, participants that did not receive terminal links chose suboptimally more often than would be expected. That is, despite the overall preference for the optimal alternative observed, the analysis revealed that choice allocation to the optimal alternative was less than expected by their experience with reinforcement outcomes within the task when participants did not receive terminal link stimuli.
Fig. 3.
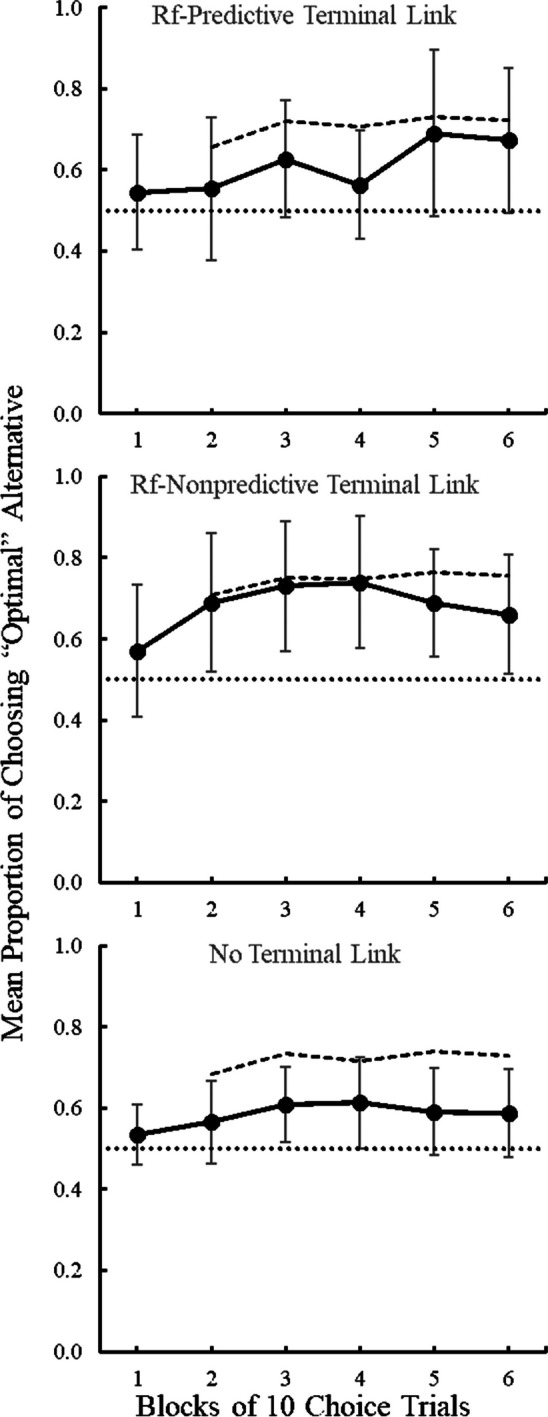
Optimality Predictions and Obtained Choice Data from Stagner et al. (2020). Note. The solid line represents the mean proportion of obtained choices to the alternative associated with the higher probability of reinforcement (i.e., optimal) across 10-trial blocks of choice trials. Error bars represent 95% Confidence Intervals. The dotted line represents chance (0.5). The dashed line represents the mean choice allocation as predicted by proportion of obtained reinforcers across forced and choice trial types. There was no statistical difference between Reinforcement-Predictive (top), Reinforcement Nonpredictive (middle), and No Terminal Link (bottom) conditions
Table 1.
Paired-Sample T-test Comparing Obtained Optimal Choice Performance to Predicted Optimal Choice Performance: Data from Stagner et al. (2020) Block 6
| Condition | Mean | t-score | df | p-value | d |
|---|---|---|---|---|---|
| Rf-Predictive Terminal Link | 0.673 | -0.585 | 10 | 0.572 | 0.176 |
| Rf-Nonpredictive Terminal Link | 0.660 | -1.595 | 9 | 0.145 | 0.504 |
| No Terminal Link | 0.587 | -2.875 | 22 | 0.009* | 0.599 |
Next, we applied the reinforcement-derived definition of optimality to our other published set of human choice data (Bodily et al., 2024). In this study, all participants were given the suboptimal task framework with manipulations to the predictive value of the terminal links (predictive vs. unpredictive) and to the type of stimuli used (colors or pictures that represented a story narrative, see Fig. 3 for narrative conditions). In Fig. 4, we have plotted obtained choice and reinforcement-derived optimality as a function of trial block.
Fig. 4.

Matching Predictions and Actual Choice Data from Bodily et al. (2024). Note. Mean proportion of obtained choices to the alternative associated with the higher probability of reinforcement (i.e., optimal) across 10-trial blocks of choice trials. The dotted line represents chance (0.50). The dashed line represents the mean choice allocation as predicted by proportion of obtained reinforcers across forced and choice trial types. There was no statistical difference between Reinforcement-Predictive (top), Reinforcement Nonpredictive (middle), and No Terminal Link (bottom) conditions
The common finding from all four conditions is that most choices made by participants corresponded with the reinforcement previously obtained following those stimuli. We can see further support for the strong impact previous reinforcement has on choice behavior in this task in Table 2.
Table 2.
Paired-Sample T-test Comparing Obtained Optimal Choice Performance to Predicted Optimal Choice Performance: Data from Bodily et al. (2024) Block 6
| Condition | Mean | t-score | df | p-value | d |
|---|---|---|---|---|---|
| Abstract Predictive | 8.822 | 2.015 | 22 | 0.056 | 0.42 |
| Narrative Predictive | 0.704 | -1.117 | 22 | 0.276 | -0.233 |
| Abstract Nonpredictive | 0.657 | -1.651 | 22 | 0.113 | -0.344 |
| Narrative Nonpredictive | 0.575 | -2.968 | 23 | 0.007 | -0.606 |
For three of the four conditions, choosing the optimal alternative did not significantly differ from reinforcement-derived optimality by Block 6. Thus, participants that received abstract reinforcement-predictive terminal stimuli (Abstract Predictive), abstract reinforcement-nonpredictive terminal stimuli (Abstract Nonpredictive), and reinforcement-predictive terminal stimuli with a story narrative context (Narrative Predictive) all chose optimally by the end of the test session. However, participants that were given the story narrative context in combination with reinforcement-nonpredictive terminal stimuli (Narrative Nonpredictive) continued to choose the suboptimal alternative more than what past reinforcement would predict. Thus, we could say that participants in this group demonstrated suboptimal behavior.
Although our discussion of reinforcement-derived optimality has been limited to human participants and to our data from two previous studies, it seems likely that comparable results would be found if optimality predictions were calculated using data from other studies with human participants. Of particular interest to us is the use of the proportion of obtained reinforcement as the reference point for optimality as opposed to definitions relative to chance. Past human work has found that human participants choose the optimal alternative in this task more often than by chance (Molet et al., 2012; McDevitt et al., 2019; Stagner et al., 2020; Bodily et al., 2024), but further examination using past reinforcements to predict choice behavior may illuminate a bias towards suboptimal choice. We have found that by examining our data sets that indeed most of the time, human participants choose optimally, or choose the optimal alternative as often as the proportion of obtained reinforcement would predict. However, we found that the absence of terminal links and the presence of a narrative context with reinforcement-nonpredictive terminal stimuli both produced more engagement with the suboptimal alternative than obtained reinforcement would predict. Thus, using past reinforcements to predict and define optimality, we have found that humans demonstrated suboptimal choice in this task under certain conditions. Perhaps then, we might consider the proportion of reinforcement obtained following optimal choices to be a reference point. Choice of the optimal alternative above matching would then be considered optimal whereas choice of the optimal alternative below matching would be considered suboptimal. To explore this finding further, future suboptimal work with both human and nonhuman subjects may benefit from applying this revised definition of optimal choice when interpreting choice data.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Declarations
Conflicts of Interest
We have no known conflicts of interest to disclose.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Baum, W. M. (1974). On two types of deviation from the matching law: Bias and undermatching. Journal of the Experimental Analysis of Behavior,22(1), 231–242. 10.1901/jeab.1974.22-231 10.1901/jeab.1974.22-231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baum, W. M. (2002). From molecular to molar: A paradigm shift in behavior analysis. Journal of the Experimental Analysis of Behavior,78(1), 95–116. 10.1901/jeab.2002.78-95 10.1901/jeab.2002.78-95 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belke, T. W., & Spetch, M. L. (1994). Choice between reliable and unreliable reinforcement alternatives revisited: Preference for unreliable reinforcement. Journal of the Experimental Analysis of Behavior,62(3), 353–366. 10.1901/jeab.1994.62-353 10.1901/jeab.1994.62-353 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanchard, T. C., Hayden, B. Y., & Bromberg-Martin, E. S. (2015). Orbitofrontal cortex uses distinct codes for different choice attributes in decisions motivated by curiosity. Neuron,85(3), 602–614. 10.1016/j.neuron.2014.12.050 10.1016/j.neuron.2014.12.050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bodily, J. S., Bodily, K. D., Southern, R. A., Baum, E. E., & Edwards, V. M. (2024). Narrative framing may increase human suboptimal choice behavior. Learning & Behavior,52, 162–169. 10.3758/s13420-023-00587-z. Online ahead of print. 10.3758/s13420-023-00587-z [DOI] [PubMed] [Google Scholar]
- Chow, J. J., Smith, A. P., Wilson, A. G., Zentall, T. R., & Beckmann, J. S. (2017). Suboptimal choice in rats: Incentive salience attribution promotes maladaptive decision-making. Behavioural Brain Research,320, 244–254. 10.1016/j.bbr.2016.12.013 10.1016/j.bbr.2016.12.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham, P. J., & Shahan, T. A. (2018). Suboptimal choice, reward-predictive signals, and temporal information. Journal of Experimental Psychology: Animal Learning & Cognition,44(1), 1–22. 10.1037/xan0000160 10.1037/xan0000160 [DOI] [PubMed] [Google Scholar]
- Cunningham, P. J., & Shahan, T. A. (2019). Rats engage in suboptimal choice when the delay to food is sufficiently long. Journal of Experimental Psychology: Animal Learning & Cognition,45(3), 301–310. 10.1037/xan0000211 10.1037/xan0000211 [DOI] [PubMed] [Google Scholar]
- Cunningham, P. J., & Shahan, T. A. (2020). Delays to food-predictive stimuli do not affect suboptimal choice in rats. Journal of Experimental Psychology: Animal Learning & Cognition,46(4), 385–397. 10.1037/xan0000245 10.1037/xan0000245 [DOI] [PubMed] [Google Scholar]
- Dunn, R., & Spetch, M. L. (1990). Choice with uncertain outcomes: Conditioned reinforcement effects. Journal of the Experimental Analysis of Behavior, 53(2), 201–218. 10.1901/jeab.1990.53-201 [DOI] [PMC free article] [PubMed]
- Dunn, R. M., Pisklak, J. M., McDevitt, M. A., & Spetch, M. L. (2024). Suboptimal choice: A review and quantification of the signal for good news (SiGN) model. Psychological Review,131(1), 58–78. 10.1037/rev0000416 10.1037/rev0000416 [DOI] [PubMed] [Google Scholar]
- Fantino, E. (1969). Choice and rate of reinforcement. Journal of Experimental Analysis of Behavior,12(5), 723–730. 10.1901/jeab.1969.12-723 10.1901/jeab.1969.12-723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu, H.-N., Monson, E., & Otto, A. R. (2021). Relationships between socio-economic status and lottery gambling across lottery types: Neighborhood-level evidence from a large city. Addiction,116(5), 1256–1261. 10.1111/add.15252 10.1111/add.15252 [DOI] [PubMed] [Google Scholar]
- Gipson, C. D., Alessandri, J. J. D., Miller, H. C., & Zentall, T. R. (2009). Preference for 50% over 75% reinforcement by pigeons. Learning & Behavior,37(4), 289–298. 10.3758/LB.37.4.289 10.3758/LB.37.4.289 [DOI] [PubMed] [Google Scholar]
- Griggs, R. A., & Cox, J. R. (1982). The elusive thematic-materials effect in Wason’s selection task. British Journal of Psychology,73, 407–420. 10.1111/j.2044-8295.1982.tb01823.x 10.1111/j.2044-8295.1982.tb01823.x [DOI] [Google Scholar]
- Herrnstein, R. J. (1970). On the law of effect. Journal of Experimental Analysis of Behavior,13(2), 243–266. 10.1901/jeab.1970.13-243 10.1901/jeab.1970.13-243 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinnenkamp, J. E., Shahan, T. A., & Madden, G. J. (2017). How suboptimal is suboptimal choice? Journal of Experimental Analysis of Behavior,107(1), 136–150. 10.1002/jeab.239 10.1002/jeab.239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kendall, S. B. (1974). Preference for intermittent reinforcement. Journal of the Experimental Analysis of Behavior,21(3), 463–473. 10.1901/jeab.1974.21-463 10.1901/jeab.1974.21-463 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDevitt, M. A., Pisklak, J. M., Spetch, M., & Dunn, R. (2018). The influence of outcome delay on suboptimal choice. Behavioural Processes,157, 279–285. 10.1016/j.beproc.2018.10.008 10.1016/j.beproc.2018.10.008 [DOI] [PubMed] [Google Scholar]
- McDevitt, M. A., Diller, J. W., & Pietrzykowski, M. O. (2019). Human and pigeon suboptimal choice. Learning & Behavior,47(4), 334–343. 10.3758/s13420-019-00391-8 10.3758/s13420-019-00391-8 [DOI] [PubMed] [Google Scholar]
- McDevitt, M. A., Pisklak, J. M., Dunn, R. M., & Spetch, M. (2022). Forced-exposure trials increase suboptimal choice. Psychonomic Bulletin & Review,29, 1514–1523. 10.3758/s13423-022-02092-2 10.3758/s13423-022-02092-2 [DOI] [PubMed] [Google Scholar]
- Molet, M., Miller, H. C., Laude, J. R., Kirk, C., Manning, B., & Zentall, T. R. (2012). Decision making by humans in a behavioral task: Do humans, like pigeons, show suboptimal choice? Learning & Behavior,40, 439–447. 10.3758/s13420-012-0065-7 10.3758/s13420-012-0065-7 [DOI] [PubMed] [Google Scholar]
- Pisklak, J. M., McDevitt, M. A., Dunn, R. M., & Spetch, M. L. (2019). Suboptimal choice and initial-link requirement. Journal of the Experimental Analysis of Behavior,112(2), 242–253. 10.1002/jeab.553 10.1002/jeab.553 [DOI] [PubMed] [Google Scholar]
- Roper, K. L., & Zentall, T. R. (1999). Observing behavior in pigeons: The effect of reinforcement probability and response cost using a symmetrical choice procedure. Learning & Motivation,30(3), 201–220. 10.1006/lmot.1999.1030 10.1006/lmot.1999.1030 [DOI] [Google Scholar]
- Russell, A. M. T., Langham, E., & Hing, N. (2018). Social influences normalize gambling-related harm among higher risk gamblers. Journal of Behavioral Addictions,7(4), 1100–1111. 10.1556/2006.7.2018.139 10.1556/2006.7.2018.139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sirola, A., Kaakinen, M., Savolainen, I., Paek, H.-J., Zych, I., & Oksanen, A. (2021). Online identities and social influence in social media gambling exposure: A four-country study on young people. Telematics & Informatics,60, 101582. 10.1016/j.tele.2021.101582 10.1016/j.tele.2021.101582 [DOI] [Google Scholar]
- Smith, T. R., & Beran, M. J. (2020). Outcome expectancy and suboptimal risky choice in nonhuman primates. Learning & Behavior,48, 301–321. 10.3758/s13420-019-00406-4 10.3758/s13420-019-00406-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith, T. R., Beran, M. J., & Young, M. E. (2017). Gambling in rhesus macaques (macaca mulatta): The effect of cues signaling risky choice outcomes. Learning & Behavior,45(3), 288–299. 10.3758/s13420-017-0270-5 10.3758/s13420-017-0270-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spetch, M. L., Belke, T. W., Barnet, R. C., Dunn, R., & Pierce, W. D. (1990). Suboptimal choice in a percentage-reinforcement procedure: Effects of signal condition and terminal-link length. Journal of the Experimental Analysis of Behavior,53(2), 219–234. 10.1901/jeab.1990.53-219 10.1901/jeab.1990.53-219 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spetch, M. L., Mondloch, M. V., & Belke, T. W. (1994). Determinants of pigeons’ choice between certain and probabilistic outcomes. Animal Learning & Behavior,22(3), 239–251. 10.3758/BF03209832 10.3758/BF03209832 [DOI] [Google Scholar]
- Stagner, J. P.., Edwards, V. M., Bond, S. R., Jasmer, J. A., Southern, R. A., & Bodily, K. D. (2020). Human choice predicted by obtained reinforcers, not by reinforcement predictors. Frontiers in Psychology, 11.10.3389/fpsyg.2020.01631 [DOI] [PMC free article] [PubMed]
- Stagner, J. P., & Zentall, T. R. (2010). Suboptimal choice behavior by pigeons. Psychonomic Bulletin & Review,17(3), 412–416. 10.3758/PBR.17.3.412 10.3758/PBR.17.3.412 [DOI] [PubMed] [Google Scholar]
- Zentall, T. R., & Stagner, J. P. (2011). Maladaptive choice behaviour by pigeons: An animal analogue and possible mechanism for gambling (sub-optimal human decision-making behaviour). Proceeding of the Royal Society B,278, 1203–1208. 10.1098/rspb.2010.1607 10.1098/rspb.2010.1607 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zentall, T. R., Andrews, D. M., & Case, J. P. (2017). Prior commitment: Its effect on suboptimal choice in a gambling-like task. Behavioural Processes,145, 1–9. 10.1016/j.beproc.2017.09.008 10.1016/j.beproc.2017.09.008 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.