

Abstract
Afterimages are illusory, visual conscious perceptions. A widely accepted theory is that afterimages are caused by retinal signaling that continues after the physical disappearance of a light stimulus. However, afterimages have been reported without preceding visual, sensory stimulation (e.g. conditioned afterimages and afterimages induced by illusory vision). These observations suggest the role of top-down brain mechanisms in afterimage conscious perception. Therefore, some afterimages may share perceptual features with sensory-independent conscious perceptions (e.g. imagery, hallucinations, and dreams) that occur without bottom-up sensory input. In the current investigation, we tested for a link between the vividness of visual imagery and afterimage conscious perception. Participants reported their vividness of visual imagery and perceived sharpness, contrast, and duration of negative afterimages. The afterimage perceptual features were acquired using perception matching paradigms that were validated on image stimuli. Relating these perceptual reports revealed that the vividness of visual imagery positively correlated with afterimage contrast and sharpness. These behavioral results support shared neural mechanisms between visual imagery and afterimages. However, we cannot exclude alternative explanations, including demand characteristics and afterimage perception reporting inaccuracy. This study encourages future research combining neurophysiology recording methods and afterimage paradigms to directly examine the neural mechanisms of afterimage conscious perception.
Keywords: afterimages, imagery, conscious perception
Introduction
Afterimages are illusory visual perseverations—lasting seconds to minutes—that often follow light stimulation but lack the original inducing light source. Analogous perceptual phenomena are reported in other human senses, including auditory afterimages or “aftersounds” (Rosenblith et al. 1947, Zwicker 1964, Wiegrebe et al. 1995). Afterimages have been a source of intrigue for centuries because of their apparent ubiquity, including among non-human animals (e.g. macaques, cats, and pigeons) and its unique insight into the physiological mechanisms of vision (Goethe 1970, Williams 1974, Duysens et al. 1985, McLelland et al. 2009). In fact, afterimages helped debunk emission theories of vision that explained conscious sight by the projection of light or aether rays from the eyes. Afterimages share perceptual characteristics with aftereffects (e.g. the McCollough effect; McCollough 1965, Smith et al. 1969) and filling-in illusions (e.g. Kanizsa or occluded stimuli illusions; Kanizsa 1976). However, afterimages are distinct because they do not require concurrent visual input (e.g. they can appear in total darkness).
A widely accepted theory is that afterimages are sourced by retinal signaling following the physical disappearance of a light stimulus. Likewise, afterimages have been described as painted or photographed on the retina (Helmholtz 1968, Favreau and Corballis 1976). Support of afterimage retinal mechanisms includes evidence of fatigue or bleaching of retinal photoreceptors that persistently signal in the absence of physical light stimulation, forming the appearance of an afterimage (Feinbloom 1938, Brindley 1962, Miller 1966, Rushton and Henry 1968, Goethe 1970, MacLeod and Hayhoe 1974, Favreau and Corballis 1976, Georgeson and Turner 1985). Likewise, direct recordings from retinal ganglion cells find a post-receptor rebound response following inducer stimulation that may originate from photoreceptor signaling (Zaidi et al. 2012). A similar retinal-based process is suggested by the opponent-process theory of chromatic afterimages, whereby complementary color afterimages are perceived according to the inducer color (e.g. a yellow inducer forms a blue afterimage), predicted by opponent visual pairs—black–white, red–green, and blue–yellow—so that adaptation to one half of an opponent pair will drive its opposite shade or hue in the subsequent afterimage (Hurvich and Jameson 1957, Favreau and Corballis 1976).
Research also finds that afterimages are not fully explained by retinal mechanisms. This evidence includes that the bleaching of photoreceptors is not a necessary condition for the formation of afterimages (Marriott 1965, Long and Kling 1983, McLelland et al. 2009). In fact, afterimages can emerge without previous photoreceptor stimulation, as in afterimages by illusory vision (e.g. a perceptually filled-in image) Shimojo et al. (2001). Likewise, studies find that color spreading in afterimages can extend beyond the boundary of the preceding inducer stimulus (van Lier et al. 2009, Hamburger et al. 2012). There are also reports of afterimages evoked by dreams, imagery, and hallucinations (Downey 1901, Weiskrantz 1950, Oswald 1957, Barber 1959). Similarly, conditioned afterimages are reported by pairing tones and inducer stimuli and then withholding the anticipated inducer, yet participants still report seeing afterimages without preceding visual stimulation (Davies 1974). These results support that some afterimages are comparable to sensory-independent conscious perceptions (e.g. imagery, hallucinations, and dreams) that are sourced by top-down brain mechanisms without bottom-up sensory input.
If some afterimages emerge without retinal signaling, a possible implication is these afterimages are perceptually linked to sensory-independent conscious perceptions that are formed by top-down brain mechanisms. Following a similar logic, previous research studied a relationship between the vividness of imagery and the occurrence of hallucinations and dreams (Barrett 1993, Dawes et al. 2020). While consideration of afterimages as a kind of sensory-independent conscious perception has been discussed, there is limited research on this topic (Kolev 1991). In fact, there is no previous study comparing the perception of afterimages and imagery. The nearest instances include rare reports of imagery inducing afterimages, afterimages induced by stimuli that were reported as challenging to imagine, and a note by William James (1842–1910) that his visual imagery could be subliminally driven by afterimages (James 1890, Downey 1901, Ingle 2005).
To address this gap in the literature, we investigated a possible perceptual link between visual imagery and afterimages. Specifically, we tested if the vividness of visual imagery (i.e. the ability to evoke lifelike visual perception by imagination) correlates with the sharpness, contrast, and duration of afterimages. We hypothesized that if the brain mechanisms involved in visual imagery are shared with those of afterimage conscious perception, then the vividness of visual imagery and afterimages may be linked. For example, people with more vivid visual imagery may also experience more vivid afterimages. Interrogating the perceptual relationship between afterimages and imagery is significant toward resolving the long-standing query for the role of bottom-up retinal versus top-down brain mechanisms in afterimage conscious perception (Oswald 1957).
Methods
Participants
Healthy, adult participants (N = 62; males = 22; mean age: 28.90 years; age standard deviation (SD): 10.31 years; mean education = 16.34 years; education SD: 1.91 years) were recruited from the local Bethesda, Maryland, United States community. Two additional participants who completed the study were excluded from the analyses because of poor behavioral performance or a corrupted behavioral file. All participants were recruited and consented following protocols approved by the Institutional Review Board of the National Institute of Mental Health. Inclusion criteria included: (1) being between the ages of 18 and 65 years old at the time of experimentation, (2) a healthy physical examination completed by a nurse practitioner within a year of the study session, and (3) ability to give informed consent. Exclusion criteria included: (1) previous or current histories of neurological or psychiatric disorder, (2) low vision (corrected normal vision was acceptable), and (3) head injuries (e.g. loss of consciousness for >30 min and three or more concussive injuries). Prior to each testing session, a nurse practitioner completed a health exam for each participant, including recording temperature, vitals, and assessment for COVID-19 symptoms.
Afterimage induction
Afterimages were elicited using an inducer stimulus: a black silhouette image of a human face in frontal view (presentation duration = 4 s; visual angle = 4.60 × 8.47 degrees; maximum luminance = ∼ 4 cd/m2; https://creazilla.com/nodes/2524-face-silhouette; Fig. 1c, “Inducer Stimulus”; Supplementary data, Movie 1). The inducer stimulus resulted in negative afterimages that appeared as white or light gray versions of the inducer. A face image inducer is unique from previous studies, which typically use flashes of light or geometric shaped images to induce afterimages (e.g. Shimojo et al. 2001, Sperandio et al. 2012).
Figure 1.
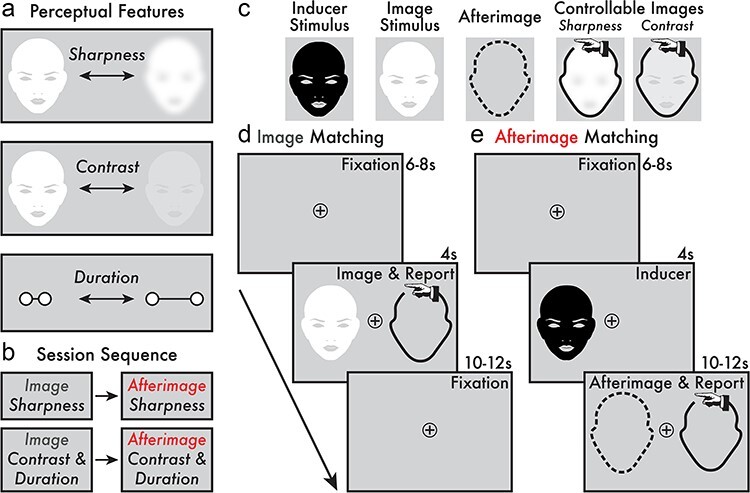
Target perceptual features, session sequence, and perception matching paradigms. (a) The target image and afterimage perceptual features were: (1) sharpness, (2) contrast, and (3) duration. (b) Participants completed four task phases in the following order: (1) image and (2) afterimage sharpness perception matching and (3) image and (4) afterimage contrast and duration perception matching. (c) The stimuli and controllable images presented in the perception matching tasks. The afterimage perception is depicted as a dashed outline because no image was physically presented—the afterimage is an illusory conscious perception. Depending on the task phase, the controllable image allowed participants to manually adjust its sharpness or contrast. The controllable image is depicted with a hand icon (not present during the task) to indicate that participants manually adjusted these images with key presses. (d) The main trial phases of the image perception matching task (Supplementary data, Movies 2 and 4). Each trial began with a fixation interval (6–8 s). When the image stimulus appeared (4 s) on either the left or right side of the central fixation, participants were instructed to immediately adjust the controllable image using key presses to match with the image stimuli according to the target perceptual feature (i.e. sharpness and contrast/duration; see “Image Sharpness and Contrast” and “Duration Perception Matching Paradigm” in “Methods” section). A subsequent fixation interval (10–12 s) followed the “Image & Report” stage prior to initiating the next trial. (e) The main trial events of the afterimage perception matching task (Supplementary data, Movies 3 and 5). Each trial began with a jittered fixation interval (6–8 s). Next, the inducer stimulus was shown (4 s) on either the left or right side of the central fixation and, subsequently, an afterimage might appear. If an afterimage was perceived, participants were instructed to immediately adjust the controllable image to match with the target perceptual feature of their afterimage (i.e. sharpness and contrast/duration; see “Afterimage Sharpness” and “Contrast and Duration Perception Matching Paradigm” in “Methods” section). The “Afterimage & Report” stage completed when the participant no longer perceived their afterimage, and the remaining duration of time (10–12 s) was a fixation interval prior to initiating the next trial.
In pilot testing, it was observed that some participants perceived an instantaneous, illusory, crisp white version of the inducer stimulus at the moment of its disappearance. This experience was sometimes confused with the subsequent negative afterimage that was typically delayed from the offset of the inducer, less sharp than the inducer, and lasted for several seconds. To limit the occurrence of this flashbulb-like conscious perception at the sudden offset of the full contrast inducer, in the first and last second of the inducer presentation, the inducer contrast was gradually ramped up and down to full contrast and no contrast, respectively. Thus, the inducer appeared at full contrast for a total of 2 s. In pilot testing (data not shown), the inducer contrast ramping suppressed the perceived offset flash without reducing the occurrence of afterimages.
During initial task instructions, participants were repeatedly shown the inducer to determine their susceptibility for perceiving afterimages. If there was confusion regarding what parts of their visual experience constituted the afterimage, clarifying instructions were provided by the experimenter to guide when and what parts of their visual perception following the inducer constituted the afterimage conscious perception.
Image and afterimage perceptual vividness
Participants were asked to report on three target perceptual features that contribute to the overall perceived strength or vividness of conscious vision: (1) sharpness (i.e. crisp versus blurry), (2) contrast (i.e. bright versus dim), and (3) duration (Fig. 1a). Sharpness, contrast, and duration are previously interrogated as markers for the vividness of afterimages (e.g. Brindley 1962, Cerf-Beare 1984). Here, participants made judgements on these perceptual features for both image and afterimage conscious perception (Fig. 1d and e). These perceptual reports were acquired using paradigms where participants adjusted the appearance of an on-screen image—a “controllable image” (Fig. 1c, “Controllable Images”)—to match in real time with the perceived sharpness, contrast, and duration of images and afterimages. Note that the contrast and duration reports were acquired simultaneously (see “Contrast and Duration Perception Matching” section). The current perception matching approach builds on previous methods for reporting the perceptual features of afterimages (e.g. Anderson and Deffenbacher 1971, Virsu and Laurinen 1977, Georgeson and Turner 1985, Lupyan 2015). Before completing the perception matching tasks, participants were administered instructions and a practice session (see “Sharpness” and “Contrast and Duration Perception Matching” sections). Only when participants felt comfortable with the reporting procedure did they continue to the image and afterimage perception matching tasks (Fig. 1b, d and e).
Sharpness perception matching paradigm
Participants were asked to notice and report on the maximum perceived sharpness of images and afterimages. Sharpness was reported using a controllable, on-screen image that participants volitionally manipulated using key presses. See full details below.
Image sharpness perception matching
Participants completed an image sharpness perception matching task (Fig. 1d; Supplementary data, Movie 2). The image stimulus was a white version of the afterimage inducer stimulus (presentation duration = 4 s; visual angle = 4.60 × 8.47 degrees; Fig. 1c, “Image Stimulus”). The stimulus was shown at a contrast of 0.3 (luminance = ∼ 63 cd/m2). In the first and last second of presentation, the image stimulus gradually increased and decreased its sharpness, respectively. When the image reached its maximum sharpness, it maintained this value for 2 s. This dynamic of increasing and decreasing sharpness was programmed according to pilot testing (data not shown) that suggested this trend matched the perception of afterimage sharpness by the current inducer stimulus.
The sharpness values applied to the image stimulus ranged from 0 (no blurring) to 25 (maximum blurring) in increments of 1, with each value representing the number of pixels (px) in the radius of a Gaussian kernel used to blur the image stimulus (blurred image size = 600 × 800 px; Gaussian blur; Illustrator, Adobe, Inc.). Three maximum sharpness values were tested: 10, 15, and 20 px. In the analyses and figures (Figs 2c, 3d and e, 4a), the sharpness values were inverted so that 0 px indicated the blurriest perception and 25 px the sharpest. Inverting the sharpness px scale was implemented because it corresponded with the contrast and duration scales, where larger values indicated more vivid images and afterimages. Thus, all sharpness values and accompanying figures are reported along the inverted px scale.
Figure 2.
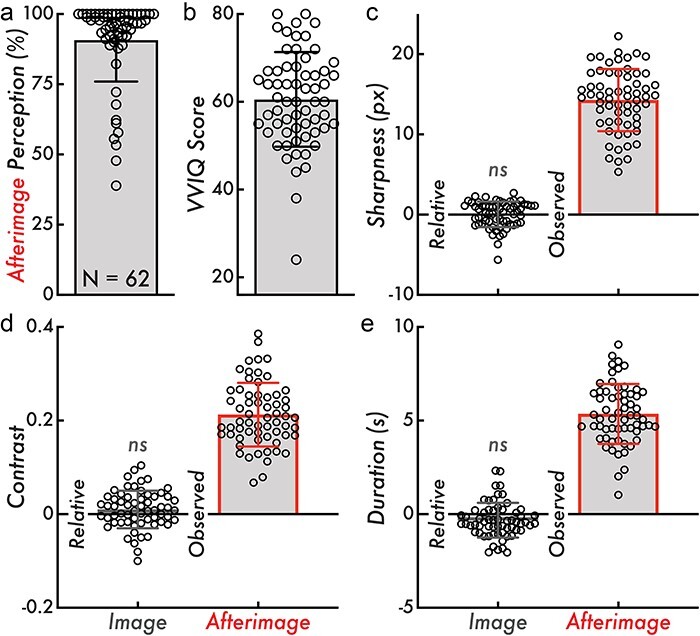
Afterimage perception rate, VVIQ score, and relative image and observed afterimage sharpness, contrast, and duration. (a) Afterimage perception rate calculated as the percentage of inducers where a subsequent afterimage was reported across all trials of the afterimage perception matching tasks (90 trials total). The bar graph indicates the mean afterimage perception percentage across participants (90.79%), and the error bar displays standard deviation (SD; 14.84%). (b) The VVIQ score calculated as the sum of scores across all questionnaire items within participant (score range: 16–80; larger values indicating more vivid visual imagery). The bar graph indicates the mean VVIQ score (60.55), and the error bars display the SD (10.78). (c) Relative image and observed afterimage reported maximum sharpness in px. The relative image sharpness is compared on a trial level against the true image sharpness (true values: 10, 15, or 20 px). The bar height indicates the group mean (“Relative” = 0.033 px; “Observed” = 14.27 px), and the error bars display SD (“Relative” = 1.60 px; “Observed” = 3.88 px). (d) Relative image and observed afterimage reported maximum contrast. The relative image contrast is compared against the true maximum image contrast (0.25). The bar height indicates the group mean (“Relative” = 0.01; “Observed” = 0.21), and the error bars display SD (“Relative” = 0.04; “Observed” = 0.068). (e) Relative image and observed afterimage reported duration in seconds (s). The relative image contrast is compared against the true image duration (4 s). The bar height indicates the group mean (“Relative” = −0.33 s; “Observed” = 5.35 s), and the error bars display SD (“Relative” = 0.93 s; “Observed” = 1.60 s). Comparing the relative image contrast, sharpness, and duration values from zero was not statistically significant (ns; Wilcoxon rank-sum tests, P > 0.05). In all subplots, the open circles represent individual participants (N = 62)
Figure 3.
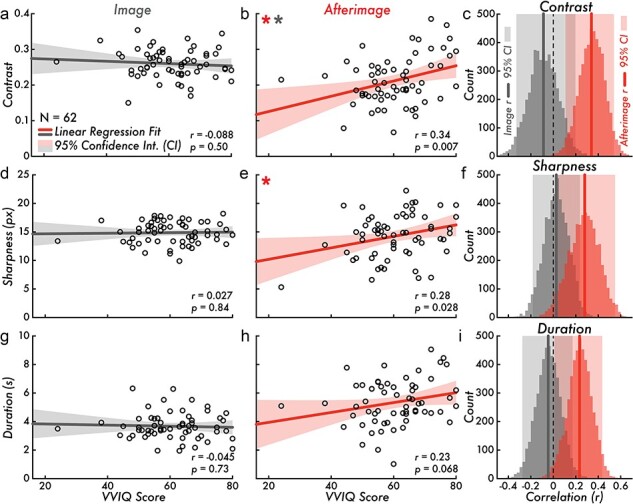
VVIQ score versus image and afterimage contrast, sharpness, and duration. (a) VVIQ score versus image contrast (correlation is not statistically significant; Pearson correlation coefficient [r] = −0.088, P = 0.50). (b) VVIQ score versus afterimage contrast (correlation is statistically significant red *; r = 0.34, P = 0.007). The VVIQ score versus afterimage contrast regression fit slope is significantly different from the VVIQ score versus image contrast regression fit slope (a; gray *; F[1, 120] = 7.45, P = 0.0073). (c) Bootstrapped image and afterimage VVIQ score and contrast correlation distributions and estimated 95% CI [image: (−0.33, 0.20); afterimage: (0.11, 0.54)]. (d) VVIQ score versus image sharpness (correlation is not statistically significant; r = 0.027, P = 0.84). (e) VVIQ score versus afterimage sharpness (correlation is statistically significant *; r = 0.28, P= 0.028). (f) Bootstrapped image and afterimage VVIQ score and sharpness correlation distributions and estimated 95% CI [image: (−0.18, 0.24); afterimage: (0.041, 0.55)]. (g) VVIQ score versus image duration (correlation is not statistically significant; r = −0.045, P = 0.73). (h) VVIQ score versus afterimage duration (correlation is not statistically significant; r = 0.23, P = 0.068). (i) Bootstrapped image and afterimage VVIQ score and duration correlation distributions and estimated 95% CI [image: (−0.28, 0.17); afterimage: (0.01, 0.44)]. Subplots a, b, d, e, g, and h, display the VVIQ score along the horizontal axis (score range: 16–80; larger values indicating more vivid visual imagery). The gray and red lines draw the linear regression fit of VVIQ score versus image or afterimage contrast, sharpness, and duration. The shaded area on either side of the main trend line is the 95% CI of the linear regression fit. The open circles represent individual participants (N = 62). In subplots c, f, and i, the gray and red vertical lines draw the Pearson correlation coefficient r value of the VVIQ score versus image or afterimage contrast, sharpness, and duration. The shaded area behind the bootstrap correlation distributions is the estimated 95% CI
Figure 4.
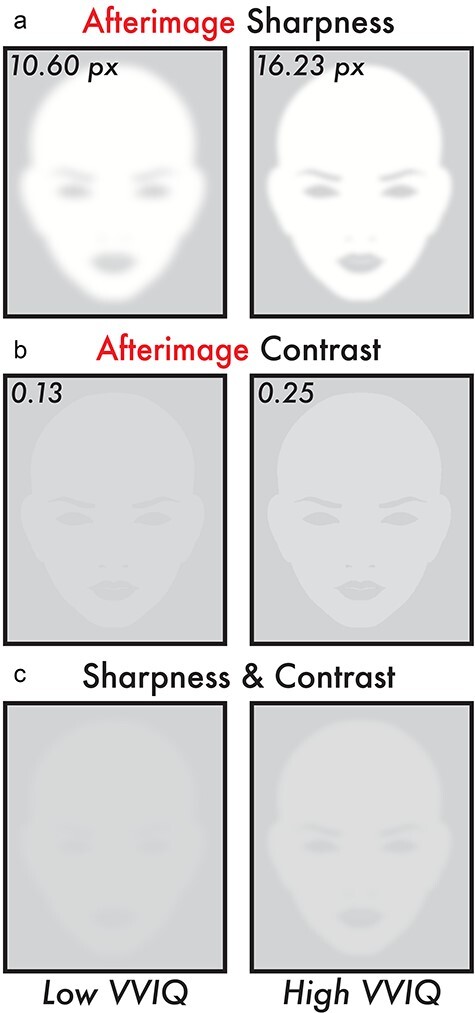
Image reconstruction of the estimated perceived afterimage sharpness and contrast for low and high VVIQ score participants. All subplots display image reconstructions of the estimated perceived afterimage sharpness and contrast values according to the fitted linear regression trend lines (Fig. 3b and e) for low and high VVIQ scores. The low and high VVIQ scores are the minimum (24) and maximum (80) VVIQ scores reported among participants (Fig. 2b). (a) Image reconstruction of the estimated perceived afterimage sharpness for low (10.60 px) and high (16.23 px) VVIQ scores. (b) Image reconstruction of the estimated perceived afterimage contrast for low (0.13) and high (0.25) VVIQ scores. (c) Image reconstruction combining the estimated perceived afterimage sharpness (a) and contrast (b) for low (10.60 px and 0.13) and high (16.23 px and 0.25) VVIQ scores. Image reconstructions show apparent differences in overall visibility and facial feature details for the estimated perceived sharpness and contrast of afterimages between low and high VVIQ score participants
Participants were instructed to report the maximum sharpness of the image stimulus in real time. This was achieved by the following steps within each image sharpness perception matching task trial (Supplementary data, Movie 2):
(1) Participants were instructed to fixate on a central plus sign inside an open circle (visual angle = 1.33 × 1.33 degrees) on a blank gray screen throughout the experiment (Fig. 1d, “Fixation” phase).
(2) After a jittered prestimulus interval (6–8 s), an image stimulus appeared (see image stimulus description above; Fig. 1d, “Image & Report” phase). The image would appear at random and in equal proportion either to the left or right of the fixation point along the midline (image stimulus location from central fixation = 5.88 degrees).
(3) Immediately upon perceiving the image stimulus, participants manually adjusted the sharpness of a controllable stimulus to match with the perceived maximum sharpness of the image stimulus using two keys: one increasing and the other decreasing the controllable image sharpness in increments of 1 px (Fig. 1c and d, “Controllable Images—Sharpness and Image & Report” phase). The controllable image was absent from the screen until the participant made their first key press to adjust its sharpness. The controllable image appeared at a random initial sharpness value (0–25 px) and was shown on the opposite side of the screen from where the image stimulus appeared.
(4). Once participants completed adjusting the sharpness of the controllable stimulus, they were instructed to press a third key to record their selection. While participants were encouraged to report the maximum sharpness of the image while the stimulus was still present on-screen, participants had a minimum of 10 s and a maximum of 12 s from the image offset (i.e. 10–12 s jittered postimage interval) to adjust the controllable image and make their perceived maximum sharpness selection (Fig. 1d, post-Image & Report “Fixation” phase). Otherwise, the trial was automatically aborted and no response was logged. A total of 20 trials of the image sharpness perception matching task were completed for each participant.
Afterimage sharpness perception matching
Participants completed an afterimage sharpness perception matching task (Fig. 1e; Supplementary data, Movie 3). The goal was for participants to report the maximum sharpness of their perceived afterimages. The reporting method and trial phases were identical to the image sharpness perception matching task (i.e. manually updating the sharpness of a controllable image with key presses to match with the perceived afterimage maximum sharpness; see “Image Sharpness Perception Matching” section). The main difference between the image and afterimage sharpness perception matching task phases was that participants were first shown the inducer stimulus in the afterimage condition (see “Afterimage Induction” section; Fig. 1e, “Inducer” phase). When the inducer disappeared, the participants might see an afterimage on the blank gray screen and were instructed to immediately adjust the controllable image using the same reporting procedure as the image sharpness perception matching task (Fig. 1e, “Afterimage & Report” phase). If participants did not see an afterimage, they were instructed to not press any keys and wait until the next trial began automatically. A total of 30 trials of the afterimage sharpness matching task were completed for each participant.
Contrast and duration perception matching paradigm
Participants were asked to report on the perceived contrast of images and afterimages overtime (i.e. to follow the change in the image and afterimage contrast throughout its perception). Previous studies have used perceptual cancellation to assess afterimage contrast (i.e. overlaying a physical image over the afterimage location and having participants adjust that physical image until the afterimage percept disappears; e.g. Georgeson and Turner 1985). In the current investigation, contrast was reported using a controllable, on-screen image that participants volitionally manipulated using key presses. See full details below.
Image contrast and duration perception matching
Participants completed an image contrast and duration perception matching task (Fig. 1d; Supplementary data, Movie 4). The image was the same stimulus used in the image sharpness perception matching task (presentation duration = 4 s; visual angle = 4.60 × 8.47 degrees; Fig. 1c, “Image Stimulus”). In the first second of image presentation, the stimulus gradually increased its contrast to a maximum contrast of 0.25 (luminance = ∼ 60 cd/m2) and then gradually decreased its contrast until the stimulus disappeared. The maximum contrast value (0.25) was selected according to pilot testing (data not shown) that suggested this contrast was similar to the maximum contrast of afterimages that appeared by the current inducer stimulus. There were three increasing contrast intervals (1, 1.5, and 2 s from the image stimulus onset until the image reached maximum contrast) and three decreasing contrast intervals (2.5, 3, and 3.5 s from the image stimulus onset until the image contrast began decreasing to full disappearance at 4 s). Thus, the image stimulus maintained the maximum contrast for a varied interval of 0.5, 1, 1.5, 2, and 2.5 s. The ramping contrast intervals were selected to approximate the contrast dynamic of the afterimage conscious perceptions reported in pilot testing (data not shown) from the current inducer stimulus.
Participants were instructed to report in real time the contrast of the image stimulus throughout its presentation. This was achieved by the following steps within each image contrast perception matching trial (Supplementary data, Movie 4):
(1) Participants were instructed to fixate on a central plus sign inside an open circle (visual angle = 1.33 × 1.33 degrees) on a blank gray screen throughout the experiment (Fig. 1d, “Fixation” phase).
(2) After a jittered pre-stimulus interval (6–8 s), an image stimulus appeared (see image stimulus description above; Fig. 1d, “Image & Report” phase). The image would appear at random and in equal proportion, either to the left or right of the fixation point along the midline (image stimulus location from central fixation = 5.88 degrees).
(3) Immediately upon perceiving the image stimulus, participants manually adjusted the contrast of a controllable image to match with the perceived contrast of the image stimulus overtime using two keys: one increasing and the other decreasing the controllable image contrast in increments of 0.025 (Fig. 1c, “Controllable Images—Contrast”). Participants could also use a third key that would immediately set the controllable image contrast to 0. Thereby, participants could report the perception of an immediate disappearance. Critically, participants were instructed to manipulate the controllable image to match with the image stimulus contrast throughout its presentation, so that at any given moment both the image and controllable image appeared with identical contrast. The controllable image appeared on the opposite side of the screen from the image stimulus.
The reported duration of the images was acquired by measuring the length of time participants manipulated the controllable image (i.e. the time when participants first reported a perceived image with greater than 0 contrast and its subsequent disappearance time; see the “Duration” subsection under “Statistical Analyses” section). While participants were encouraged to report the contrast of the image while the image stimulus was still present on-screen, participants had a minimum of 10 s and a maximum of 12 s from the image offset (i.e. 10–12 s jittered post-image interval) to continue adjusting the controllable image, after which the trial was automatically aborted and the responses made in the preceding interval were logged (Fig. 1d, post-Image & Report “Fixation” phase). A total of 18 trials of the image contrast and duration matching task were completed for each participant.
Afterimage contrast and duration perception matching
Participants completed an afterimage contrast and duration perception matching task (Fig. 1e; Supplementary data, Movie 5). The goal was for participants to report the change in contrast overtime of their perceived afterimages. The reporting method and trial phases were identical to the image contrast and duration perception matching task (i.e. manually updating the contrast of a controllable image with key presses to match with the perceived afterimage contrast throughout its conscious perception; see “Image Contrast and Duration Perception Matching” section). The main difference between the image and afterimage contrast and duration perception matching task phases was that in the afterimage condition, participants were first shown the inducer stimulus (see “Afterimage Induction” section; Fig. 1e, “Inducer” phase). When the inducer disappeared, the participants might see an afterimage on the blank gray screen and were instructed to immediately display and adjust the controllable image using the same reporting procedure as the image contrast and duration perception matching task (Fig. 1e, “Afterimage & Report” phase). If participants did not see an afterimage, they were instructed to not press any keys and wait until the next trial began automatically. A total of 60 trials of the afterimage contrast and duration perception matching task were completed for each participant.
Visual imagery vividness
Visual imagery vividness was acquired with the 16-item, self-reported Vividness of Visual Imagery Questionnaire (VVIQ) (Marks 1973). The questionnaire asks participants to imagine people, objects, and scenes (e.g. the contour of a familiar face, the front of a shop, and a sun rise) and then introspect on how vivid that imagined content appears in their visual imagery on a 5-point scale between “no image at all” to “perfectly clear and as vivid as normal vision.” Participants were instructed to complete the VVIQ with their eyes open and were given no time constraint in completing the questionnaire. The VVIQ was displayed on a computer monitor and participants used a mouse click to select their answers for each questionnaire item. The VVIQ was administered at either the beginning (∼80% of participants) or end of the study session (∼20% of participants).
Equipment, software, and facility
The behavioral study was completed in a single 2-h study session in a windowless behavioral testing room. The room lighting was set to a consistent brightness level for all participants. The experimenter was present in the testing room but positioned out of sight of the participant to monitor behavior and deliver task instructions. The behavioral paradigm was coded in Python and run with PsychoPy (v2022.2.4; Open Science Tools Ltd) on a behavioral laptop (MacBook Pro; 13-inch; 2560 × 1600 px, 2019; Mac OS Catalina v10.15.7; Apple, Inc.; Peirce 2007). The behavioral laptop monitor was mirrored by DVI cable to a VIEWPixx monitor (1920 × 1200 px; VPixx Technologies, Inc.) on which the participants viewed the experimental paradigms and the VVIQ. The participants were positioned ∼56 cm from the center of the display monitor. The viewing distance was fixed using a table mounted head-chin rest (SR Research Head Support; SR Research Ltd). All participants used their right hand (regardless of handedness) to make key presses during the task with a keyboard positioned on a table in front of the participant.
Statistical analyses
All analyses were completed in MATLAB v2022b (MathWorks, Inc.) and Prism v10 (GraphPad Software, Inc.). Figures were generated and edited in MATLAB v2022b (MathWorks, Inc.), Prism v10 (GraphPad Software, Inc.), and Illustrator (Adobe, Inc.).
Afterimage perception rate
Afterimage perception rate measures how often afterimages were perceived by each participant following the inducer stimulus. The perception rate was calculated by finding the percentage of inducer presentations during which an afterimage was perceived across the sharpness, contrast, and duration perception matching tasks—a total of 90 trials (i.e. the number of perceived afterimage trials in the sharpness perception matching task plus the number of perceived afterimage trials in the contrast and duration perception matching task divided by the total number of trials across all tasks). Perception rate values were multiplied by 100 to convert from units of fraction to percentage.
VVIQ Score
The VVIQ score for each participant was calculated by taking the sum of all scores across the questionnaire items. Each item was scored on a scale from 1 (no image) to 5 (perfectly clear). The minimum and maximum VVIQ score was 16 and 80, respectively, where larger values indicate more vivid visual imagery.
Sharpness
Calculating reported sharpness
Participants reported the perceived maximum sharpness of images and afterimages (see subsection “Sharpness Perception Matching Paradigm” in “Methods” section). The participant image and afterimage sharpness values were calculated by averaging all trial sharpness values within participant and image and afterimage sharpness perception matching tasks. Trials without a sharpness value (e.g. response timeout or afterimage was not perceived) were excluded from consideration in calculating the participant maximum sharpness value. The sharpness value scale was inverted, so that larger values correspond with a sharper perception. This scale inversion was achieved by taking the absolute value of the participant mean sharpness value minus the maximum sharpness value (25; i.e. the largest px radius of the blurring Gaussian kernel).
Calculating reported image sharpness accuracy
Participant-reported image sharpness accuracy was calculated by subtracting the reported maximum image sharpness from the true maximum image sharpness (10, 15, or 20 px) across trials. Next, all subtracted or “relative” sharpness trial values were averaged within participant. A positive relative sharpness value indicated the image was reported as sharper than its true maximum sharpness, while a negative relative sharpness indicated the image was reported as blurrier than its true maximum sharpness, where a value of 0 indicated a perfect match between the reported and true image maximum sharpness (Fig. 2c, “Relative”). To statistically test the reporting accuracy of the image maximum sharpness, a Wilcoxon rank-sum test (P < 0.05) was applied on the relative sharpness values and tested against 0. If the relative image sharpness was found to be no different from 0, these suggested participants were accurate in reporting the maximum sharpness of the image stimulus.
Calculating the correlation between VVIQ score and sharpness
The relationship between VVIQ scores and the reported maximum image and afterimage sharpness were statistically tested using a two-tailed, Pearson correlation test (P < 0.05; Fig. 3d and e). Correlation analyses were applied in two comparisons: (1) VVIQ score versus image sharpness and (2) VVIQ score versus afterimage sharpness. A linear regression fit was applied to model the trend and 95% CI for each of the comparisons. The regression fit slopes were directly compared between VVIQ score versus image sharpness and VVIQ score versus afterimage sharpness using an analysis of covariance (P < 0.05). In addition, the bootstrap resampling method (5000 samples) estimated the 95% CI for the correlation between VVIQ score and image and afterimage maximum sharpness (Fig. 3f). A CI that includes 0 suggests no correlation at the 5% significance level.
Image reconstruction of the afterimage sharpness
The perceived maximum sharpness of the afterimage was reconstructed for low (24) and high (80) VVIQ scores, representing the minimum and maximum VVIQ score recorded among participants (Fig. 2b). Reconstruction was achieved by finding the sharpness value for the low and high VVIQ scores along the VVIQ score versus afterimage sharpness linear regression fit trend line and creating images (Gaussian blur; Illustrator; Adobe, Inc.) that matched with these estimated sharpness values (Fig. 4a and c).
Contrast
Calculating reported contrast
Participants reported the perceived contrast of images and afterimages overtime (see subsection “Contrast and Duration Perception Matching Paradigm” in “Methods” section). The participant image and afterimage contrast values were calculated by finding the maximum contrast value reported in each image and afterimage contrast and duration perception matching task trial. Next, the maximum contrast values were averaged across trials within the image and afterimage conditions for each participant. Any trial with less than two reported contrast values or a maximum contrast value of 0 (e.g. an afterimage was not perceived) was ignored from calculating the participant image and afterimage contrast value.
Calculating reported image contrast accuracy
Participant-reported image contrast accuracy was calculated by subtracting the reported maximum image contrast values from the known maximum image contrast value (0.25) across trials. Next, all subtracted or “relative” contrast trial values were averaged within participant. A positive relative contrast indicated the image was reported as brighter than its true maximum contrast, while a negative relative contrast indicated the image was reported as dimmer than its true maximum contrast, where a value of 0 indicated a perfect match between the participant reports and the true image maximum contrast (Fig. 2d, “Relative”). To statistically test the reporting accuracy of the image maximum contrast, a Wilcoxon rank-sum test (P < 0.05) was applied on the relative contrast values and tested against 0. If the relative image contrast was found to be no different from 0, these suggested participants were accurate in reporting on the contrast of the image stimulus.
Calculating the correlation between VVIQ score and contrast
The relationship between VVIQ scores and the reported maximum image and afterimage contrast were statistically tested using a two-tailed, Pearson correlation test (P < 0.05; Fig. 3a and b). Correlation analyses were applied in two comparisons: (1) VVIQ score versus image contrast and (2) VVIQ score versus afterimage contrast. The regression fit slopes were directly compared between VVIQ score versus image contrast and VVIQ score versus afterimage contrast using an analysis of covariance (P < 0.05). A linear regression fit was applied to model the trend and 95% CI for each of the comparisons. In addition, the bootstrap resampling method (5000 samples) estimated the 95% CI for the correlation between VVIQ score and image and afterimage maximum contrast (Fig. 3c). A CI that includes 0 suggests no correlation at the 5% significance level.
Image reconstruction of the afterimage contrast
The perceived maximum contrast of the afterimage was reconstructed for low (24) and high (80) VVIQ scores, representing the minimum and maximum VVIQ score recorded among participants (Fig. 2b). Reconstruction was achieved by finding the contrast value for the low and high VVIQ scores along the VVIQ score versus afterimage contrast linear regression fit trend line and creating images (Illustrator; Adobe, Inc.) that matched with these estimated contrast values (Fig. 4b and c).
Duration
Calculating reported duration
Image and afterimage durations were calculated from the contrast and duration perception matching tasks (see subsection “Contrast and Duration Perception Matching Paradigm” in “Methods” section). Contrast and duration perception matching task trials were considered valid by the same criteria for calculating the reported maximum contrast of images and afterimages (see subsection “Contrast” in “Statistical Analyses” section). The duration was measured as the time between the initial and final key press participants made to adjust the controllable image to match with the perceived contrast of the images and afterimages or when the participant reported the image or afterimage had a contrast of zero, whichever occurred first.
Calculating reported image duration accuracy
Participant reported image duration accuracy was calculated by subtracting the reported image duration across trials within participant from the true image duration (4 s). A positive relative duration indicated participants reported on average that the image was presented longer than its true duration, while a negative relative duration indicated that participants reported on average that the image was briefer, where a value of 0 indicated a perfect match between the reported and true image duration (Fig. 2e, “Relative”). To statistically test how accurate participants were in reporting the image duration, a Wilcoxon Rank Sum test (P < 0.05) was applied on the relative duration values and tested against 0. If the relative image duration was found no different from 0, this suggested participants were accurate in reporting on the duration of the image stimulus.
Calculating the correlation between VVIQ score and duration
The relationship between VVIQ scores and the reported image and afterimage duration were statistically tested using a two-tailed, Pearson correlation test (P < 0.05; Fig. 3g and h). Correlation analyses were applied in two comparisons: (1) VVIQ score versus image duration and (2) VVIQ score versus afterimage duration. A linear regression fit was applied to model the trend and 95% CI for each of the comparisons. The regression fit slopes were directly compared between VVIQ score versus image duration and VVIQ score versus afterimage duration using an analysis of covariance (P < 0.05). In addition, the bootstrap resampling method (5000 samples) estimated the 95% CI for the correlation between VVIQ score and image and afterimage duration (Fig. 3i). A CI that includes 0 suggests no correlation at the 5% significance level.
Results
Afterimage perception rate and VVIQ score
The inducer stimulus consistently induced afterimages in most participants: a mean afterimage perception rate of 90.79% (SD = 14.84%; minimum participant afterimage perception rate = 38.89%; maximum participant afterimage perception rate = 100%; Fig. 2a). The mean VVIQ score was 60.55 (SD = 10.78; minimum participant VVIQ score = 24; maximum participant VVIQ score = 80; Fig. 2b). One participant was below the ∼ 30-score threshold that is commonly used to designate “aphantasia”—the near or total inability to form visual imagery—estimated to account for less than 5% of the general population (Dance et al. 2022).
Image and afterimage perceptual features
Participants reported on their perceived sharpness, contrast, and duration of images and afterimages. The mean image and afterimage maximum sharpness values were 14.87 px (SD = 1.91 px) and 14.27 px (SD = 3.88 px), respectively (Fig. 2c, “Observed”; image condition observed not shown). The mean image and afterimage maximum contrast values were 0.26 (SD = 0.04) and 0.21 (SD = 0.068), respectively (Fig. 2d, “Observed”; image condition observed not shown). The mean image and afterimage duration values were 3.67 s (SD = 0.93 s) and 5.35 s (SD = 1.60 s), respectively (Fig. 2e, “Observed”; image condition observed not shown). These results revealed broad individual variability for the perceived sharpness, contrast, and duration of afterimages.
Accuracy of the reported image sharpness, contrast, and duration
A key validation was whether participants could accurately report on their perceptual experiences using the current perception matching paradigms (see subsections “Sharpness Perception Matching Paradigm” and “Contrast and Duration Perception Matching Paradigm” in “Methods” section). Image stimuli with known sharpness, contrast, and duration were used to validate the accuracy of the perceptual reports using the perception matching paradigms. In support of the perception matching paradigms and their resulting perceptual reports, there was no statistically significant difference (P > 0.05) between the reported and true image sharpness, contrast, and duration. Specifically, the reported sharpness, contrast, and duration minus the true image sharpness, contrast, and duration were not statistically different from 0 (i.e. a perfect match between the reported and true image sharpness, contrast, and duration; Fig. 2c, d and e, “Relative”; see “Methods” section for image features and statistical testing details).
VVIQ score versus image sharpness, contrast, and duration
Task-based factors, including motor and cognitive processes involved in reporting on the perceptual features of conscious perception could explain a relationship between VVIQ score and reported image and afterimage sharpness, contrast, and duration. This alternative hypothesis was tested by comparing VVIQ score with the reported image sharpness, contrast, and duration. There was no statistically significant correlation for VVIQ score versus image contrast [Pearson correlation coefficient (r) = −0.088, P = 0.50; Fig. 3a], VVIQ score versus image sharpness (r = 0.027, P = 0.84; Fig. 3d), and VVIQ score versus image duration (r = −0.045, P = 0.73; Fig. 3g). The bootstrap estimated 95% CIs of the correlation distributions confirmed that there was no relationship between VVIQ score and image contrast [(−0.33, 0.20); Fig. 3c], sharpness [(−0.18, 0.24); Fig. 3f], and duration [(−0.28, 0.17); Fig. 3i].
VVIQ score versus afterimage sharpness, contrast, and duration
To test for a link between the vividness of visual imagery and afterimages, VVIQ scores were correlated with the afterimage sharpness, contrast, and duration. A statistically significant, moderate positive correlation was found between VVIQ score and afterimage contrast (r = 0.34, P = 0.007; linear regression fit trend line equation: Y = 0.0021*X + 0.083; Fig. 3b). A statistically significant, weak positive correlation was found between VVIQ score and afterimage sharpness (r = 0.28, P = 0.028; linear regression fit trend line equation: Y = 0.10*X + 8.19; Fig. 3e). There was no statistically significant correlation between VVIQ score and afterimage duration (r = 0.23, P = 0.068; Fig. 3h). The VVIQ score versus image and VVIQ score versus afterimage regression fit slopes were significantly different for contrast (F[1, 120] = 7.45, P = 0.0073). In short, the correlation between VVIQ score and afterimage contrast is significantly stronger than the correlation between VVIQ score and image contrast. However, the VVIQ score versus image and VVIQ score versus afterimage regression fit slopes were not significantly different for sharpness (F[1, 120] = 3.65, P = 0.058) nor duration (F[1, 120] = 3.16, P = 0.078). Finally, the bootstrap estimated 95% CIs of the correlation distributions supported a positive correlation between VVIQ score and afterimage contrast [(0.11, 0.54); Fig. 3c], sharpness [(0.041, 0.55); Fig. 3f], and duration [(0.01, 0.44); Fig. 3i].
The estimated sharpness value according to the linear regression fit trend line for a low VVIQ score (minimum participant VVIQ score = 24) was 10.60 px and a high VVIQ score (maximum participant VVIQ score = 80) was 16.23 px (Fig. 4a and c). The estimated contrast value according to the linear regression fit trend line for a low VVIQ score (minimum participant VVIQ score = 24) was 0.13 and a high VVIQ score (maximum participant VVIQ score = 80) was 0.25 (Fig. 4b and c).
When the recruited participant with aphantasia (see “Afterimage Perception Rate and VVIQ Score” section) is removed from the dataset, the correlation between VVIQ score and afterimage contrast and duration become stronger (contrast: r = 0.38, P = 0.0026; duration: r = 0.25, P = 0.052). Meanwhile, because this participant reported the least sharp (i.e. blurriest) afterimages of all participants, when removed from analyses the correlation between VVIQ score and afterimage sharpness maintained the same trend but was no longer statistically significant (r = 0.17, P = 0.18). These results support a statistically robust correlation between VVIQ score and afterimage contrast. Meanwhile, additional research is required (e.g. testing participants with aphantasia) to confirm a possible relationship between VVIQ score and afterimage sharpness and duration.
Discussion
The current investigation is the first study relating the perception of imagery and afterimages. Our main finding was evidence for a perceptual link between visual imagery and negative afterimages. To interrogate this relationship, we developed novel perception matching paradigms where participants manipulated the appearance of on-screen images to report their perceived afterimage sharpness, contrast, and duration. The efficacy of these reporting methods was validated by testing the participant reporting accuracy for image stimuli with known sharpness, contrast, and duration (Fig. 2c, d and e, “Relative”). These perceptual reports revealed variability in the perceived sharpness, contrast, and duration of afterimages across participants (Fig. 2c, d and e, “Observed”). Moreover, we discovered a statistically significant, moderate positive correlation between visual imagery vividness and the perceived afterimage contrast (Fig. 3b and c). In short, people who reported more vivid visual imagery tended to report brighter afterimages. There was also a weak positive correlation between visual imagery vividness and afterimage sharpness. However, this statistical result was sensitive to a single participant with aphantasia (Fig. 3e and f; see subsection “Afterimage Perception Rate and VVIQ Score” in “Results” section).
Bolstering these results was the specificity of the relationship of visual imagery vividness to afterimage conscious perception. Meanwhile, no correlation was found between VVIQ score and image contrast, sharpness, and duration in the same participants (Fig. 3a, c, d, f, g and i). Moreover, reconstructions of the estimated afterimage conscious perceptions for low and high VVIQ scores were visibly distinct—the high VVIQ score afterimage reconstruction revealed an apparently brighter image with sufficient sharpness to discern facial features that were absent in the low VVIQ afterimage reconstruction (Fig. 4c, “Low” versus “High VVIQ”).
The relationship between the vividness of visual imagery and afterimage conscious perception is a novel source of behavioral evidence that imagery and afterimages may share neural mechanisms. Specifically, the afterimages induced in the current investigation may involve top-down brain mechanisms in common with visual imagery that helps explain the observed individual variability in the perceived afterimage sharpness, contrast, and duration. This interpretation of the current results is corroborated by observations that afterimages can form without bottom-up retinal stimulation (e.g. Davies 1974, Shimojo et al. 2001). However, these behavioral findings can only speculate on neural mechanisms. Thereby, our results encourage combining afterimage paradigms with neuroimaging and other neurophysiology recording methods to directly examine the neural mechanisms of afterimage conscious perception.
Alternative explanations and limitations
An alternative explanation for these findings is the influence of mediating sensory, cognitive, or behavioral variables required by the perception reporting tasks (e.g. reaction time and sensory sensitivity) and reporting on visual imagery vividness. This account is dampened because reporting sharpness, contrast, and duration of images and afterimages versus visual imagery vividness involved orthogonal tasks (see subsection “Image and Afterimage Perceptual Vividness” in “Methods” section). Specifically, reporting on image and afterimage conscious perception required adjusting a controllable image with key presses. Meanwhile, visual imagery vividness was inquired using a self-paced questionnaire (i.e. VVIQ) that involved marking responses with a mouse click. Therefore, sensory, cognitive, and behavioral ability are excluded as likely factors influencing the current findings. Indeed, if mediating variables linked to task requirements explained these results, we would also expect a relationship between the vividness of visual imagery and image conscious perception, as identical perception reporting procedures were involved for both images and afterimages. Thus, task requirements involved in perception reporting are unlikely to explain the current findings. Still, we cannot rule out the influence of other unknown factors that may be shared across reporting methods and unique to afterimages (e.g. metacognitive or introspection ability).
In addition, we cannot exclude demand characteristics—cues or expectations about the research objective that can influence participant behavior and perceptual experience (Orne 1962, Corneille and Béna 2023, Corneille and Lush 2023). The unknown influence of demand characteristics undermines conclusions about experimental effects. For example, numerous perceptual and psychological phenomena (e.g. behavioral priming, hypnosis, and illusions) have been found subject to demand characteristics, thus questioning the validity of the purported mechanisms for these effects (Damaser et al. 1963, Doyen et al. 2012, Lush et al. 2020). In the current experiment, participants may have formed assumptions about the relationship between their responses on the VVIQ and image and afterimage reporting tasks that influenced their performance on these experimental assessments. If demand characteristics were a significant factor, our findings may be driven by participant expectations regarding imagery and image and afterimage conscious perception.
Experimenter bias demand characteristics were partly controlled in the current experiment using scripted task instructions that were read verbatim to each participant to ensure consistent instructions and limit off-script instructional cues that could hint at the experimental objectives and hypothesized effects. Moreover, the experimenter was unaware of the participant VVIQ score prior to the main task, thus limiting the influence of experimenter priming of participant behavior (e.g. Doyen et al. 2012). Also, the participants did not receive feedback during the study to suggest their imagery and afterimage perception profiles (i.e. participants were not told if their responses indicated dull or vivid imagery ability or afterimage perception).
Still, experimental design demand characteristics may be present independent of experimenter bias and despite VVIQ and image and afterimage perception reporting involving orthogonal tasks. A direct approach to study the role of experimental design demand characteristics is an instruction-based replication. For example, in the context of the current experiment, new participants could be explicitly instructed of the opposite experimental inference (i.e. vivid imagery predicts dull afterimages) prior to completing perception reporting. If participant imagery and afterimage perception reports correspond with the instructed inference, this would evidence the influence of demand characteristics. Alternatively, a conceptual replication might involve instructing new participants on the current experimental design and asking them to simulate the responses of the participants who completed the current experiment. For example, participants could be asked to predict VVIQ responses after viewing the image and afterimage perception matching performance of a participant tested in the current study. Without an instruction-based replication, we cannot rule out experimental design demand characteristics as a significant factor in the current experiment.
Finally, a challenge for the current investigation is assuring accuracy of the afterimage perceptual reports. Previous studies have manipulated the inducer stimulus to predictably change afterimage conscious perception to determine the veracity of perceptual reports (Georgeson and Turner 1985). Here, we used images with known sharpness, contrast, and duration to validate reporting on the same perceptual features of afterimages. In support of the perception reporting methods and reporting accuracy, participants were accurate for indicating the sharpness, contrast, and duration of images (Fig. 2c, d and e). Importantly, the image stimuli were designed to approximate the perceptual features of the afterimages to match the reporting difficultly between images and afterimages. Still, reporting on-screen images versus illusory afterimages may be distinct (e.g. more challenging to report on illusory conscious perceptions). Therefore, it is possible that image reporting accuracy may not predict afterimage reporting accuracy. Future studies may address this limitation by recording physiological markers of conscious perception (e.g. pupil size) that may serve as a covert measure of afterimage perceptual vividness to corroborate the veracity of overt perceptual reports. Likewise, pupil size has been shown to validate self-report of visual imagery vividness (Kay et al. 2022).
Future directions
The current findings motivate future study on the relationship between afterimages and other categories of sensory-independent conscious perception (e.g. hallucinations and dreams) in healthy physiology. Furthermore, it would be valuable to research how afterimages may change in psychiatric and neurologic disorders that impact sensory and sensory-independent conscious perception. Supporting this research aim, a study found differences in afterimage onset latency among people with brain injury (Ruesch 1944). In addition, afterimages are altered in people with autistic traits and schizophrenia (Hartman 1962, Sperandio et al. 2017, Thakkar et al. 2019). These findings suggest afterimages may offer translational value, as previously suggested in posterior cortical atrophy—a variant of Alzheimer’s disease—and Parkinson’s disease where afterimage perception have also been found to be modified relative to healthy individuals (Chan et al. 2001, Khadjevand et al. 2010, Crutch et al. 2011).
Conclusion
Afterimages have long been a source of curiosity and implemented as a perceptual tool to interrogate vision and the neural mechanisms of consciousness. In the current investigation, we studied a possible link between visual imagery and afterimage conscious perception. We developed novel perception matching paradigms that allowed for the acquisition of various perceptual features of image and afterimage conscious perception. Our main result was a correlation between the perceived vividness of visual imagery and negative afterimages. This represents a new source of evidence that some afterimages may share neural mechanisms with visual imagery. Still, we cannot exclude alternative explanations, including demand characteristics and perceptual reporting inaccuracy. This study motivates future research to directly examine the precise neural mechanisms of afterimage conscious perception.
Supplementary Material
Acknowledgements
We thank members of the Section on Functional Imaging Methods for their constructive feedback.
Contributor Information
Sharif I Kronemer, Section on Functional Imaging Methods, Laboratory of Brain and Cognition, National Institute of Mental Health (NIMH), National Institutes of Health (NIH), Bethesda, MD 20892, United States.
Micah Holness, Section on Functional Imaging Methods, Laboratory of Brain and Cognition, National Institute of Mental Health (NIMH), National Institutes of Health (NIH), Bethesda, MD 20892, United States.
A Tyler Morgan, Section on Functional Imaging Methods, Laboratory of Brain and Cognition, National Institute of Mental Health (NIMH), National Institutes of Health (NIH), Bethesda, MD 20892, United States; Functional Magnetic Resonance Imaging Core Facility, NIMH, NIH, Bethesda, MD 20892, United States.
Joshua B Teves, Section on Functional Imaging Methods, Laboratory of Brain and Cognition, National Institute of Mental Health (NIMH), National Institutes of Health (NIH), Bethesda, MD 20892, United States.
Javier Gonzalez-Castillo, Section on Functional Imaging Methods, Laboratory of Brain and Cognition, National Institute of Mental Health (NIMH), National Institutes of Health (NIH), Bethesda, MD 20892, United States.
Daniel A Handwerker, Section on Functional Imaging Methods, Laboratory of Brain and Cognition, National Institute of Mental Health (NIMH), National Institutes of Health (NIH), Bethesda, MD 20892, United States.
Peter A Bandettini, Section on Functional Imaging Methods, Laboratory of Brain and Cognition, National Institute of Mental Health (NIMH), National Institutes of Health (NIH), Bethesda, MD 20892, United States; Functional Magnetic Resonance Imaging Core Facility, NIMH, NIH, Bethesda, MD 20892, United States.
Author contributions
Sharif I. Kronemer (Conceptualization, Methodology, Software, Formal analysis, Investigation, Data curation, Visualization, Supervision, Project administration, and Writing—original draft), Micah Holness (Methodology, Software, Investigation, and Writing—review and editing), A. Tyler Morgan (Conceptualization, Methodology, and Writing—review and editing), Joshua B. Teves (Methodology, Software, and Writing—review and editing), Javier Gonzalez-Castillo (Formal analysis and Writing—review and editing), Daniel A. Handwerker (Conceptualization, Methodology, Supervision, Formal analysis, and Writing—review and editing), Peter A. Bandettini (Conceptualization, Methodology, Supervision, Project administration, Funding acquisition, and Writing—review and editing).
Supplementary data
Supplementary data is available at Neuroscience of Consciousness online.
Conflict of interest
None declared.
Funding
This research was made possible by the support of the National Institute of Mental Health Intramural Research Program (ZIAMH002783). The study was completed in compliance with the National Institutes of Health Clinical Center protocol ID 93-M-0170 (ClinicalTrials.gov ID: NCT00001360).
Data availability
Data and analysis scripts are available at https://github.com/nimh-sfim/Afterimage-Imagery-Behavioral.
References
- Anderson RH, Deffenbacher KA. Effect of sound stimulation on visual afterimages. Percept Mot Skills 1971;32:343–6.doi: 10.2466/pms.1971.32.2.343 [DOI] [PubMed] [Google Scholar]
- Barber TX. The afterimages of “hallucinated” and “imagined” colors. J Abnorm Psychol 1959;59:136–9.doi: 10.1037/h0047334 [DOI] [PubMed] [Google Scholar]
- Barrett TR. Verbal hallucinations in normals-II: self-reported imagery vividness. Pers Individ Dif 1993;15:61–7. [Google Scholar]
- Brindley GS. Two new properties of foveal after-images and a photochemical hypothesis to explain them. J Physiol 1962;164:168–79.doi: 10.1113/jphysiol.1962.sp007011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cerf-Beare A. Pattern and orientation effects on afterimage duration. Perception 1984;13:443–53.doi: 10.1068/p130443 [DOI] [PubMed] [Google Scholar]
- Chan D, Crutch SJ, Warrington EK. A disorder of colour perception associated with abnormal colour after-images: a defect of the primary visual cortex. J Neurol Neurosurg Psychiatry 2001;71:515–7.doi: 10.1136/jnnp.71.4.515 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corneille O, Béna J. Instruction-based replication studies raise challenging questions for psychological science. Collabra-Psychol 2023;9:1–14.doi: 10.1525/collabra.82234 [DOI] [Google Scholar]
- Corneille O, Lush P. Sixty years after orne’s American Psychologist article: a conceptual framework for subjective experiences elicited by demand characteristics. Pers Soc Psychol Rev 2023;27:83–101.doi: 10.1177/10888683221104368 [DOI] [PubMed] [Google Scholar]
- Crutch SJ, Lehmann M, Gorgoraptis N et al. Abnormal visual phenomena in posterior cortical atrophy. Neurocase 2011;17:160–77.doi: 10.1080/13554794.2010.504729 [DOI] [PubMed] [Google Scholar]
- Damaser EC, Shor RE, Orne MT. Physiological effects during hypnotically requested emotions. Psychosom Med 1963;25:334–43.doi: 10.1097/00006842-196307000-00002 [DOI] [PubMed] [Google Scholar]
- Dance CJ, Ipser A, Simner J. The prevalence of aphantasia (imagery weakness) in the general population. Conscious Cogn 2022;97:103243.doi: 10.1016/j.concog.2021.103243 [DOI] [PubMed] [Google Scholar]
- Davies P. Conditioned after-images. I. Br J Psychol 1974;65:191–204.doi: 10.1111/j.2044-8295.1974.tb01394.x [DOI] [PubMed] [Google Scholar]
- Dawes AJ, Keogh R, Andrillon T et al. A cognitive profile of multi-sensory imagery, memory and dreaming in aphantasia. Sci Rep 2020;10:10022.doi: 10.1038/s41598-020-65705-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Downey JE. An experiment on getting an after-image from a mental image. Psychol Rev 1901;8:42–55. [Google Scholar]
- Doyen S, Klein O, Pichon CL et al. Behavioral priming: It’s all in the mind, but whose mind? PLoS One 2012;7:e29081.doi: 10.1371/journal.pone.0029081 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duysens J, Orban GA, Cremieux J et al. Visual cortical correlates of visible persistence. Vision Res 1985;25:171–8.doi: 10.1016/0042-6989(85)90110-5 [DOI] [PubMed] [Google Scholar]
- Favreau OE, Corballis MC. Negative aftereffects in visual perception. Sci Am 1976;235:42–8.doi: 10.1038/scientificamerican1276-42 [DOI] [PubMed] [Google Scholar]
- Feinbloom W. A Quantitative Study of the Visual After-image. Archives of psychology 1938. [Google Scholar]
- Georgeson MA, Turner RSE. Afterimages of sinusoidal, square-wave and compound gratings. Vision Res 1985;25:1709–20.doi: 10.1016/0042-6989(85)90143-9 [DOI] [PubMed] [Google Scholar]
- Goethe JWV. Theory of Colours. Cambridge, Massachusetts, London: The MIT Press, 1970. [Google Scholar]
- Hamburger K, Geremek A, Spillmann L. Perceptual filling-in of negative coloured afterimages. Perception 2012;41:50–6.doi: 10.1068/p7066 [DOI] [PubMed] [Google Scholar]
- Hartman AM. The apparent size of after-images in delusional and non-delusional schizophrenics. Am J Psychol 1962;75:587–95.doi: 10.2307/1420281 [DOI] [PubMed] [Google Scholar]
- Helmholtz HV. Helmholtz on Perception: Its Physiology and Development. Warren RM and Warren RP (eds), New York, London, Sydney: John Wiley & Sons, 1968. [Google Scholar]
- Hurvich LM, Jameson D. An opponent-process theory of color vision. Psychol Rev 1957;64:384–404.doi: 10.1037/h0041403 [DOI] [PubMed] [Google Scholar]
- Ingle D. Central visual persistences: I. Visual and kinesthetic interactions. Perception 2005;34:1135–51.doi: 10.1068/p5408 [DOI] [PubMed] [Google Scholar]
- James W. The Principles of Psychology. Vol. 2. New York: Henry Holt and Company, 1890. [Google Scholar]
- Kanizsa G. Subjective contours. Sci Am 1976;234:48–53. [DOI] [PubMed] [Google Scholar]
- Kay L, Keogh R, Andrillon T et al. The pupillary light response as a physiological index of aphantasia, sensory and phenomenological imagery strength. Elife 2022;11:1–17.doi: 10.7554/eLife.72484 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khadjevand F, Shahzadi S, Abbassian A. Reduction of negative afterimage duration in Parkinson’s disease patients: a possible role for dopaminergic deficiency in the retinal interplexiform cell layer. Vision Res 2010;50:279–83.doi: 10.1016/j.visres.2009.09.026 [DOI] [PubMed] [Google Scholar]
- Kolev OI. Comparative investigation of vestibular influence on visual hallucinations and flash induced afterimage. Acta Otolaryngol Suppl 1991;481:262–5.doi: 10.3109/00016489109131397 [DOI] [PubMed] [Google Scholar]
- Long GM, Kling SC. Positive and negative afterimages from brief target gratings. Vision Res 1983;23:959–63.doi: 10.1016/0042-6989(83)90005-6 [DOI] [PubMed] [Google Scholar]
- Lupyan G. Object knowledge changes visual appearance: semantic effects on color afterimages. Acta Psychol (Amst) 2015;161:117–30.doi: 10.1016/j.actpsy.2015.08.006 [DOI] [PubMed] [Google Scholar]
- Lush P, Botan V, Scott RB et al. Trait phenomenological control predicts experience of mirror synaesthesia and the rubber hand illusion. Nat Commun 2020;11:4853.doi: 10.1038/s41467-020-18591-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacLeod DIA, Hayhoe M. Rod origin of prolonged afterimages. Science 1974;185:1171–2.doi: 10.1126/science.185.4157.1171 [DOI] [PubMed] [Google Scholar]
- Marks DF. Visual imagery differences in the recall of pictures. Br J Psychol 1973;64:17–24.doi: 10.1111/j.2044-8295.1973.tb01322.x [DOI] [PubMed] [Google Scholar]
- Marriott FH. Thresholds for negative after-images. J Physiol 1965;180:888–92.doi: 10.1113/jphysiol.1965.sp007738 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCollough C. Color adaptation of edge-detectors in the human visual system. Science 1965;149:1115–6.doi: 10.1126/science.149.3688.1115 [DOI] [PubMed] [Google Scholar]
- McLelland D, Ahmed B, Bair W. Responses to static visual images in macaque lateral geniculate nucleus: implications for adaptation, negative afterimages, and visual fading. J Neurosci 2009;29:8996–9001.doi: 10.1523/JNEUROSCI.0467-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller ND. Positive afterimage following brief high-intensity flashes. J Opt Soc Am 1966;56:802–6.doi: 10.1364/josa.56.000802 [DOI] [PubMed] [Google Scholar]
- Orne MT. On the social psychology of the psychological experiment: with particular reference to demand characteristics and their implications. Am Psychol 1962;17:776–83.doi: 10.1037/h0043424 [DOI] [Google Scholar]
- Oswald I. After-images from retina and brain. Q J Exp Psychol 1957;9:88–100.doi: 10.1080/17470215708416225 [DOI] [Google Scholar]
- Peirce JW. PsychoPy—Psychophysics software in Python. J Neurosci Methods 2007;162:8–13.doi: 10.1016/j.jneumeth.2006.11.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenblith WA, Miller GA, Egan JP et al. An auditory afterimage? Science 1947;106:333–5.doi: 10.1126/science.106.2754.333 [DOI] [PubMed] [Google Scholar]
- Ruesch J. Dark adaptation, negative after images, tachistoscopic examinations and reaction time in head injuries. J Neurosurg 1944;1:243–51.doi: 10.3171/jns.1944.1.4.0243 [DOI] [Google Scholar]
- Rushton WAH, Henry GH. Bleaching and regeneration of cone pigments in man. Vision Res 1968;8:617–31.doi: 10.1016/0042-6989(68)90040-0 [DOI] [PubMed] [Google Scholar]
- Shimojo S, Kamitani Y, Nishida S. Afterimage of perceptually filled-in surface. Science 2001;293:1677–80.doi: 10.1126/science.1060161 [DOI] [PubMed] [Google Scholar]
- Smith GJ, Fries I, Andersson AL. Individual consistencies in adaptation to negative afterimages and spiral aftereffects. Percept Mot Skills 1969;28:767–75.doi: 10.2466/pms.1969.28.3.767 [DOI] [PubMed] [Google Scholar]
- Sperandio I, Chouinard PA, Goodale MA. Retinotopic activity in V1 reflects the perceived and not the retinal size of an afterimage. Nat Neurosci 2012;15:540–2.doi: 10.1038/nn.3069 [DOI] [PubMed] [Google Scholar]
- Sperandio I, Unwin KL, Landry O et al. Size constancy is preserved but afterimages are prolonged in typical individuals with higher degrees of self-reported autistic traits. J Autism Dev Disord 2017;47:447–59.doi: 10.1007/s10803-016-2971-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thakkar KN, Silverstein SM, Brascamp JW. A review of visual aftereffects in schizophrenia. Neurosci Biobehav Rev 2019;101:68–77.doi: 10.1016/j.neubiorev.2019.03.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Lier R, Vergeer M, Anstis S. Filling-in afterimage colors between the lines. Curr Biol 2009;19:R323–324.doi: 10.1016/j.cub.2009.03.010 [DOI] [PubMed] [Google Scholar]
- Virsu V, Laurinen P. Long-lasting afterimages caused by neural adaptation. Vision Res 1977;17:853–60.doi: 10.1016/0042-6989(77)90129-8 [DOI] [PubMed] [Google Scholar]
- Weiskrantz L. An unusual case of after-imagery following fixation of an “imaginary” visual pattern. Q J Exp Psychol 1950;2:170–5. [Google Scholar]
- Wiegrebe L, Kössl M, Schmidt S. Auditory sensitization during the perception of acoustical negative afterimages: analogies to visual processing? Naturwissenschaften 1995;82:387–9.doi: 10.1007/BF01134569 [DOI] [PubMed] [Google Scholar]
- Williams JL. Evidence of complementary afterimages in the pigeon. J Exp Anal Behav 1974;21:421–4.doi: 10.1901/jeab.1974.21-421 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaidi Q, Ennis R, Cao D et al. Neural locus of color afterimages. Curr Biol 2012;22:220–4.doi: 10.1016/j.cub.2011.12.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zwicker E. “Negative afterimage” in hearing. J Acoust Soc Am 1964;36:2413–5.doi: 10.1121/1.1919373 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data and analysis scripts are available at https://github.com/nimh-sfim/Afterimage-Imagery-Behavioral.