

Abstract
Background
Monocytes and macrophages play an indispensable role in maintaining intestinal homeostasis and modulating mucosal immune responses in inflammatory bowel disease (IBD). Although numerous studies have described macrophage properties in IBD, the underlying mechanisms whereby the monocyte-macrophage lineage modulates intestinal homeostasis during gut inflammation remain elusive.
Main body
In this review, we decipher the cellular and molecular mechanisms governing the generation of intestinal mucosal macrophages and fill the knowledge gap in understanding the origin, maturation, classification, and functions of mucosal macrophages in intestinal niches, particularly the phagocytosis and bactericidal effects involved in the elimination of cell debris and pathogens. We delineate macrophage-mediated immunoregulation in the context of producing pro-inflammatory and anti-inflammatory cytokines, chemokines, toxic mediators, and macrophage extracellular traps (METs), and participating in the modulation of epithelial cell proliferation, angiogenesis, and fibrosis in the intestine and its accessory tissues. Moreover, we emphasize that the maturation of intestinal macrophages is arrested at immature stage during IBD, and the deficiency of MCPIP1 involves in the process via ATF3-AP1S2 signature. In addition, we confirmed the origin potential of IL-1B+ macrophages and defined C1QB+ macrophages as mature macrophages. The interaction crosstalk between the intestine and the mesentery has been described in this review, and the expression of mesentery-derived SAA2 is upregulated during IBD, which contributes to immunoregulation of macrophage. Moreover, we also highlight IBD-related susceptibility genes (e.g., RUNX3, IL21R, GTF2I, and LILRB3) associated with the maturation and functions of macrophage, which provide promising therapeutic opportunities for treating human IBD.
Conclusion
In summary, this review provides a comprehensive, comprehensive, in-depth and novel description of the characteristics and functions of macrophages in IBD, and highlights the important role of macrophages in the molecular and cellular process during IBD.
Keywords: Macrophage, Inflammatory bowel disease, Intestinal mucosa, Susceptibility genes, Homeostasis, Immunopathology
Introduction
Inflammatory bowel disease (IBD) is a group of chronic relapsing inflammatory diseases that affect the entire gastrointestinal tract, including Crohn’s disease and ulcerative colitis [1, 2]. During gut inflammation, the integrity of the intestinal mucosal barrier is compromised, leading to exposure to commensal microbiota and pathogens [1, 3]. Recent studies have highlighted that the destruction of intestinal mucosal barrier integrity and the disruption of intestinal mucosal immune homeostasis are strongly associated with the dysregulated immune response to commensal microbiota [3–5]. Therefore, it is highly desired to investigate the great significance and complex mechanisms whereby macrophages regulate mucosal homeostasis during IBD.
Intestinal resident macrophages are at the front line of host defense at the mucosal area and form a paramount nexus in maintaining intestinal homeostasis and modulating inflammatory response in gut mucosa [2, 6, 7]. Increasing lines of evidence has indicated that macrophages in the intestine are involved in the generation of cytokines, chemokines, and macrophage extracellular traps (METs), the modulation of immune cell–cell crosstalk, the activation of phagocytosis and bactericidal effects, the response to exogenous antigens and bacteria, and process of epithelial cell proliferation, angiogenesis, tissue repair and fibrosis [6–9]. In line with our recent report [10], perturbation of the intestinal homeostasis contributes to the dramatic changes in the compositions of the intestinal monocyte-macrophage lineages in the pathology of IBD, characterized by the replacement of tissue-resident macrophages by infiltrating proinflammatory monocytes-macrophages [11]. Specifically, excessive or prolonged activation of proinflammatory macrophages results in the impairment of tissue repair and the enhancement of fibrosis, which is associated with an aggravation of surgical risks (e.g., stenosis and intestinal obstruction) [12]. In addition, our recent data have identified that macrophages-derived serum amyloid A2 (SAA2) is positively correlated with the microbial transporters and modulates immune homeostasis in the mesentery, mesenteric lymph nodes and mesenteric adipose tissues [13].
Previous studies have highlighted that copious amounts of IBD susceptibility genes are related to the monocyte-macrophage lineages like nucleotide binding oligomerization domain containing 2 (NOD2), autophagy related 16 like 1 (ATG16L1), C-X3-C motif chemokine receptor 1 (CX3CR1), Janus kinase 2 (JAK2), and signal transducer and activator of transcription 3 (STAT3) [14–17]. The latest breakthrough from our team has illuminated the newly founded genetic characteristics of IBD in East Asian ancestry, including macrophage-related genes like RUNX family transcription factor 3 (RUNX3), IL21R, general transcription factor Iii (GTF2I), and leukocyte immunoglobulin like receptor B3 (LILRB3) [17]. Subsequently, a reduction in pro-inflammatory macrophage polarization ascribed to anti-TNF treatment (e.g., infliximab) and a declination in monocyte accumulation on account of anti-α4β7 integrin therapy (e.g., vedolizumab) lead to clinical remission in IBD [18, 19]. Thus, integrating the definite properties of intestinal monocyte-macrophage lineage under physiological and pathological conditions and translating these findings into future intervention strategies have essential implications for better understanding the pathogenesis and alleviating the clinical manifestations of IBD.
In this review, we discuss the origin and location, the maturation and differentiation, and the function of macrophages in intestinal mucosa sequentially. Importantly, we leverage recently developed technologies, including single-cell RNA-sequencing (scRNA-seq), fate mapping techniques, genome-wide association studies (GWAS), and ImmunoChip, which are instrumental in improving our understanding of the biology and pathology in macrophages. Moreover, we also characterize the features of macrophages during IBD and delineate the relationship between IBD susceptibility gene-driven macrophage activation and potential therapeutics, which concomitantly provide vast connotations for our understanding of macrophage immunoregulation and tissue repair in IBD.
Origin and location of macrophage in the gut
Macrophages in the intestine contribute to a robust payload of immune regulation, which is markedly distinct from circulating monocytes, and the intestine is the largest reservoir of macrophages in adult tissues [11, 20]. In terms of the anatomical distribution of intestinal macrophages, accumulating lines of evidence have observed that macrophages are strategically localized at the different layers of the entire GI tract, ranging from the lamina propria (LP), which closes proximity to the epithelial monolayer, to the submucosa plexus and muscularis externa [3, 7, 11, 21]. They are also connected to the crypt base within the villi right around lymphoid tissues, the enteric neurons, and blood vessels and are involved in regulating intestinal motility [11, 21, 22]. Interestingly, the number of macrophages changes gradually in the mouse intestine from the proximal to distal ends while this concept is not applicable for macrophages in the human intestine, where they appear to distribute evenly [11].
In the early 1970s, it was described that embryonic precursor macrophages colonizing the tissues in the embryonic stage emanate from either Yolk sac (YS) erythro-myeloid precursors (EMP) or fetal-liver precursors. These lifelong tissue-resident macrophages obtain minimal supplement from adult hematopoietic cells [23, 24]. However, tissue-resident macrophages in the fetal and neonatal intestine, although colonized by embryonic precursors from E8.5 onwards, are reported to have a 3-week half-life and unable to last into adulthood due to their poor proliferative capacity and the exposure to the intestinal microbiota and its metabolites [7, 8, 11, 25, 26]. Therefore, continuous recruitment and replenishment of CCR2-dependent circulating Ly6Chi monocytes, which arise from postnatal hematopoiesis and mature into tolerogenic IL-10+ macrophages, are of great importance for the immune homeostasis of intestinal mucosal under physiological and pathological conditions with a unique turnover rate [7, 25, 27]. Intestinal macrophages are distinguished from other immune cells in the gastrointestinal tract with their expression of colony-stimulating factor 1 receptor (CSF1R), CD68, and FcγRI (also known as CD64) [28, 29]. Flow cytometric analysis reveals that the heterogeneous phenotypes of intestinal macrophages differ from circulating Ly6Chi monocytes based on specific surface markers [20]. By combining flow cytometric analysis with principle component analysis, infiltrated monocytes are observed to fail to differentiate into macrophages but remain in an activated status in steady states [30]. A subsequent study has identified a unique genetic signature of intestinal macrophages which differentiate from circulating monocytes with global transcriptomic analysis [31].
Since the twenty-first century, the notion that circulating monocytes have the potential to apply to all adult tissue macrophages has been challenged [23]. The recently developed technologies, including fate mapping techniques and scRNA-seq, have shed some light on understanding the mononuclear phagocyte system, and additional subsets of the monocyte-macrophage population have been deciphered [27, 32, 33]. scRNA-seq enables the differentiation of macrophages from intestinal LP into 4 subpopulations with distinction in their unique localization and functions [34]. Seminal studies have further revealed that several subpopulations of macrophages arising from embryonic precursors in the gut niches maintain locally reflected by their distinct transcriptional profiles and contribute to ontogeny as well as intestinal homeostasis independently of replenishment by circulating Ly6Chi monocytes [23, 33–35]. Additionally, other studies have proposed that most of these tissue-resident macrophages arising from embryonic precursors are constituted by late c-Myb+ EMP-derived fetal liver monocytes rather than early EMP-derived YS macrophages [27, 33]. However, an opposite paradigm uncovered that fetal hematopoietic stem cell (HSC) wave, which originates from aorta-gonad-mesonephros (AGM) but not YS EMPs, is the origin of intestinal resident macrophages [36]. In particular, distinct expression of Tim-4 and CD4 could separate intestinal macrophages into three subsets, including macrophages locally maintained (Tim-4+CD4+), macrophages turnover from circulating monocytes slowly (Tim-4-CD4+), and macrophages with the high monocyte-replenishment rate (Tim-4-CD4-) [35]. On the contrary, Tim4 is absent in the intestinal macrophages of human [29]. Moreover, macrophages from the colon LP cells can also be divided into several subsets based on the expression of F4/80 and CD11b. F4/80hi macrophages are considered intestinal resident macrophages, while CD11bhi macrophages are thought of as infiltrating macrophages with replenishment by circulating monocytes [36, 37].
The gradual replenishment and maturation of these hematopoietic stem cell-derived progenitors in the intestinal mucosa as well as the formation of their distinctive surface markers are shaped by the environmental element, including the high commensal burden, and pro-inflammatory and anti-inflammatory signatures [25, 31, 34, 35]. The surrounding environment has a concept that is also known as the macrophage ‘niche’. Signals in the niche involve generating niche-specific phenotypes and functional plasticity of macrophages by activating specific transcriptional profiles within tissue resident macrophages [7]. On the other hand, the death of macrophages in the niche facilitates the recruitment and maturation of monocytes to fill the niches [38]. Previous work has described that the monocyte-macrophages are remarkably downregulated in the colon of germ-free (GF) mice compared with those in conventional control mice. In particular, markedly fewer Ly6ChiMHC II- and Ly6C+MHC II+ cells are observed in the colon of GF mice compared with those in conventional control mice [25]. In addition, the proportion of intestinal macrophages, in particular CD11c+CD206intCD121b+ and CD11c-CD206hiCD121b- macrophages, is selectively decreased in GF mice compared with that in SPF mice [39]. Moreover, Enterobacteriaceae, a specific microbiota, is involved in the recruitment of Ly6ChiCCR2+ monocytes from the circulation into intestinal mucosa in dextran sulfate sodium salt (DSS)-induced colitis model [40], indicating the critical role of microbiota in modulating the composition of intestinal mucosal macrophages [25, 39]. Evidence has also shown that the generation of tissue-resident macrophages deriving from either HSC-dependent or independent fetal-liver precursors relies on c-Myb activity [33]. The differentiation of intestinal monocytes-macrophages is also modulated by environmental factors like IL-10, TGF-β, colony-stimulating factor 1 (CSF1), and oxysterol [2]. Accordingly, IL-10 is required to suppress the hyperactivation of tissue-resident macrophages in the gut [41]. The IL-10–IL-10R signaling blockage leads to the remodeling of intestinal macrophage phenotypes by restricting the activation of STAT3 [41]. TGF-β, a tolerogenic signal, is released from intestinal epithelia and indirectly induced by gut microbiota, and the TGF-βR1-dependent signaling is involved in the terminal monocyte-macrophage maturation by orchestrating the genetic signature of intestinal macrophages, which is associated with an enhancement of genes including CX3CR1, IL-10, and CCDC23 [2, 20, 31]. The mechanisms underlying the TGF-βR1-mediated signaling in regulating the constitution of the intestinal macrophage pool appear to be distinct from those used by IL-10 [31]. CSF1, the primary regulator of the mononuclear phagocyte lineage, participates in the proliferation of intestinal macrophage maturation. Consistently, deficiency of CSF1 in mice leads to the suppression of macrophage differentiation [42]. Silico trajectory analysis uncovers that the localization and function of intestinal macrophages are modulated by extravasated monocytes and key transcription factors [8, 29]. Depletion of these tissue-resident macrophages leads to the Csf-1R/Csf-2 signaling-dependent recovery of intestinal macrophages and reduction in intestinal motility [23, 34]. As for oxysterol levels, it seems that the enhancement of oxysterol subsequently recruits GPR183+ inflammatory circulating monocyte-derived cells into inflamed intestine [38].
Taken together, cumulative data have confirmed that embryonic original tissue-resident macrophages as well as peripheral-derived macrophages co-existed in the intestinal mucosa, and their compositions and distribution are tremendously dependent on signatures in the niches.
Macrophage maturation
The process of macrophage maturation
As described above, intestinal-resident macrophages lack the ability of self-sustainment, and the continuous recruitment and maturation of circulating monocytes are required to fill the intestinal niches and maintain the homeostasis of intestinal mucosal immunity in a steady state [11, 38, 43]. This process is triggered by the release of pro-inflammatory mediators, cytokines, and chemokines (e.g., CCL2, CCL8) [6]. After entering into the LP of intestinal mucosa, CCR2+Ly6C+ monocytes initiate a series well-defined process of maturation, which acquire major histocompatibility complex II (MHC II) firstly, followed by losing the expression of Ly6C and CCR2, and the enhancement of F4/80, CD64, CX3CR1 and CD206 in mice, or the enhancement of CD64, CD68, CD206 and CD163 in humans [2, 6, 7, 11, 25, 28, 43]. By using flow cytometry, the process of the monocyte-to-macrophage maturation continuum is identified in both mouse and human intestines [24, 31]. Gating strategy is exemplified: Monocytes and macrophages filled the LP of the intestinal mucosa are gated according to the high expression of CD45, CD11b, and CD64 [28]. After that, these gated cells are divided into four populations according to their maturation status. Newly infiltrated monocytes are defined as population 1 (P1) and characterized by a unique phenotype that expresses a high level of Ly6C, but the expression of MHC‧II is absent (namely, Ly6ChiMHC‧II+). Subsequently, monocytes that gradually acquire MHC‧II expression are reckoned as immature macrophages and named population 2 (P2, Ly6C+ MHC‧II+) to “bridge” monocytes and mature macrophages. Finally, immature macrophages lose their expression of Ly6C and then obtain the characteristics of mature macrophages, namely population 3 (P3, Ly6C-MHC II+CX3CR1int). Mature macrophages fill the intestinal niche and are classified as population 4 (P4, Ly6-CMHC II+CX3CR1hi) [3, 24, 28, 31, 43]. Given that the P3 and P4 cells are only distinguished by differential expression of CX3CR1, they are considered as a single population 3/4 (P3/P4, Ly6C-MHC II+CD64+) in mice [28]. This entire maturation process is called monocyte-waterfall, which usually takes almost 5–7 days [11, 24]. In addition, flow cytometry also distinguishes macrophages from DCs in the LP of intestinal mucosa from both mice and humans through the expression of CD64 [28]. These mature macrophages from the LP of mouse intestinal mucosa develop characteristic functions like the enrichment of receptors associated with phagocytic and bactericidal activity, including TIM4, CD36, and αvβ5 integrin, IL-10 production, apoptotic cell elimination, as well as the tolerance of TLR stimulation [3, 11, 31, 43–45].
The application of unbiased scRNA-seq reveals the differences in transcriptomes among monocytes, immature macrophages, and macrophages from intestinal mucosa [8, 31, 46, 47]. Several subsets of mRNA transcripts involved in an extravasation process and TLR and pro-inflammatory associated processes are significantly downregulated during the maturation process from the newly infiltrated monocytes (P1) to mature macrophages (P4) in the LP of intestinal mucosa, including Ly6c1, Itgb7, Sell, Itga1, Ccr2, Gpr35, Myd88, Irak3, Trem1, and Il6 [31]. In contrast, mRNA transcripts associated with phagocytosis (Mertk, Mrc1 (CD206), Cd36, Gas6, Axl, Itgav, Itgb5, Cd9, Cd81, C1qa-c, Klf2, Stab1), metalloproteinases (Mmp2, Mmp9, Mmp12, Mmp13), TGFβR signaling (Tgfbr1, Tgfbr2, Smad7, Serpine1), and monocyte chemoattractants (Ccl7 and Ccl8) are upregulated during maturation process from the P1 to P4 in the LP of intestinal mucosa [8, 31, 46]. These observations emphasize that mature macrophages are more involved in phagocytosis, anti-inflammation, and tissue repair and are consistent with previous studies described above. In addition, the mRNA transcriptions of S100A8, S100A9, CD209, CD163, RXRA, MPO, TRAF3IP3, PFKP, FCN1, and VCAN are restricted, while the mRNA transcriptions of CD169, MHC II, ACP5, C1Q, ICA1, GOPC, STOML1, KIFAP3, and KIF3C are enhanced during the maturation from the newly infiltrated monocytes to mature macrophages in the LP of intestinal mucosa [8, 46, 47].
Regulation of macrophage maturation
The maturation of monocytes to macrophages mainly depends on the tolerogenic environment in intestinal niches and the integrity of the intestinal epithelial barrier [6, 11, 24, 38, 48]. For instance, commensal microbiota and its metabolites in the intestinal mucosa orchestrate the recruitment of circulating monocytes and constantly fine-tune the fulfillment of mature macrophages in intestinal niches under both physiological and pathological conditions [2, 28, 49]. Chemokines and their receptors also participate in the process of the monocyte-macrophage maturation. CX3CR1 is known as a TGF-β-dependent chemokine receptor, bound by CX3CL1, thus indicating the maturation of macrophages [8, 31]. The niche environment in the gut instructs an upregulation of CX3CR1 in monocytes-macrophages since macrophages in other tissues lack the expression of CX3CR1 [11]. Thereafter, CX3CR1hi macrophages in the gut secrete high levels of CCL2 and CCL7 and consequently fine-tune their replenishment [11, 50, 51]. Published data have also revealed that the monocyte-macrophage maturation is largely dependent on the CSF1/CSF1R signaling, and multiple growth factors like flt3L, CSF1R, CSF1, and CSF2 are also involved [8, 11, 24, 40, 52–54]. It should be noted that CSF1 functions in a concentration-dependent manner since a high level of CSF1 facilitates macrophage proliferation while a low level of CSF1 participates in macrophage survival [7, 55–57]. In contrast, anti-CSF-1R monoclonal antibody treatment significantly interferes with the monocyte-macrophage maturation but does not influence the infiltration of pro-inflammatory monocytes [57, 58]. In the local environment of the gut, a large amount of IL-10 and IFN-γ secreted by Treg cells, and epithelium-derived mediators, including CX3CL1, IL-33, and IL-25 also involved in the modulation of the monocyte-macrophage maturation [6, 8, 59–61]. IL-10 has been confirmed to upregulate the expression of CD206 and CD163, and guarantee the generation of MHC II, leading to the monocyte-macrophage maturation [11]. Interestingly, the generation of IL-10 is regulated by macrophages, which contribute to a potent payload in maintaining the immunity homeostasis of intestinal niches [6]. Cumulative lines of evidence have demonstrated that several transcription factors are involved in the development of macrophages in the colon, exemplified by triggering receptors expressed on myeloid cells 1 (TREM1), which acts as an amplifier of pro-inflammatory responses [8, 62]. Additionally, our recent data have uncovered that monocyte chemotactic protein-1-induced protein 1 (MCPIP1) orchestrates monocyte to macrophage maturation in the intestine via a activating transcription factor 3 (ATF3)-adaptor related protein complex 1 subunit sigma 2 (AP1S2)-dependent manner and contributes to the restraint of mucosal inflammation [10]. Evidence also verifies that zinc finger E-box-binding homeobox 2 (ZEB2) is involved in maintaining the tissue-specific identities of macrophages in the intestine [8, 63]. In addition, the circadian clock also participates in the process of macrophage maturation [38]. Collectively, intestinal niches exert a crucial influence on shaping the fate of the monocyte-macrophage lineage.
Macrophage maturation during colitis
Evidence has shown that monocyte maturation to macrophages is arrested at an immature state (P2, Ly-6Cint MHC II+CD64+) during inflammation, including IBD [3, 10, 24, 28, 31, 43, 50, 60, 64]. Circulating monocytes (Ly6ChiMHC II-, equivalent to CD14hiCD11chiCCR2+CX3CR1+ monocytes in humans) infiltrate into intestinal mucosa massively, while the process of its maturation into mature macrophages (Ly6C-MHC II+CD64+, equivalent to CD14hiCD11c-CCR2-CX3CR1- macrophages in humans) is disrupted compared with that at steady state, leading to the enrichment of immature macrophages (Ly6C+MHC II+CD64+, equivalent to CD14hiCD11cCCR2-CX3CR1+ immature macrophages in humans) and the exhaustion of mature macrophages [6, 10, 24, 28]. These immature macrophages are different from ‘classical macrophages’ and highly responsive to the TLR signaling [24, 43, 65], contributing to an upregulation of pro-inflammatory mediators (TNF-⍺, IL-1β, IL-6, IL-12, IL-23, iNOS, and OSM) and downregulation of IL-10, leading to monocyte infiltration, tissue damage and function deterioration [6, 24, 28, 38, 66]. Taken together, these inflammation-associated signals further skew the process of the monocyte-macrophage maturation toward a disordered status [38].
Macrophage heterogeneity
Previous studies have roughly divided macrophages into pro-inflammatory macrophages (M1) and anti-inflammatory macrophages (M2). However, with the advanced technology, especially scRNA-seq, macrophages are now further divided into several subpopulations based on distinct classification dimensions (Table 1).
Table 1.
Macrophage heterogeneity in the intestine
| Population | Markers | Characteristics | References |
|---|---|---|---|
| M1 | inos, IL-1β, IL-6, CCL5, CXCL9, MMP9 | 1) Activated by LPS and/or IFN- | [67–71] |
| 2) Metabolized by glycolysis | |||
| 3) Deregulate tight junction proteins | |||
| 4) Increase apoptosis of epithelial cell | |||
| 5) Disrupt epithelial barrier | |||
| M2 | CD206, CD163, CCL17, IRF4, ARG-1, Retnla, Chi3l3 | 1) Activated by IL-4, or IL-13 | |
| 2) Repair damaged tissue | |||
| 3) Regulated IL-10 and TGF-β1 producing Treg cells | |||
| 4) Produce IL-10 and ARG-1 | |||
| 5) Exacerbate pathological fibrosis | |||
| Tim-4+CD4+ macrophages | Tim-4+CD4+ | Rarely replaced by infiltrated monocytes | [35] |
| Tim-4-CD4+ macrophages | Tim-4-CD4+ | Slowly replaced by infiltrated monocytes | |
| Tim-4-CD4- macrophages | Tim-4-CD4− | High replenishment rate from infiltrated monocyte | |
| Mϕ1 | CD11bhi CD64+ MerTK+ | Similar to peripheral blood monocytes phenotypically | [47] |
| CD163+ CD115+ CX3CR1+ | |||
| CD206- CD1c- CD103− | |||
| CCR2+ calprotectin+ | |||
| Mϕ2 | CD11b+ CD64+ MerTK+ | ||
| CD163med CD115+ | |||
| CX3CR1med CD206+ CD1c+ | |||
| CD103+ CCR2+ calprotectin+ | |||
| Mϕ3 | CD11b- CD64low MerTK+ | Similar to mature macrophages phenotypically | |
| CD163+ CD115+ CX3CR1- | |||
| CD206+ CD1c− CD103 | |||
| - CCR2- calprotectin- | |||
| Mϕ4 | CD11b+ CD64low MerTK+ | ||
| CD163+ CD115low CX3CR1low | |||
| CD206+ CD1c- | |||
| CD103-CCR2- calprotectin- | |||
| monocyte-like cells | CD11chigh CCR2+ | 1) Share characteristics with circulating CD14+ monocytes | [90] |
| CX3CR1+ | 2) Increased in the inflamed intestinal mucosa from IBD patients | ||
| 3) Produce IL-1β | |||
| macrophage-like cells | CD11c- CCR2- CX3CR1- | 1) macrophage-like tissue resident counterparts | |
| 2) Produce IL-10 |
Pro- and anti-inflammatory macrophages
Mature macrophages can polarize into ‘classically activated’/pro-inflammatory macrophages (M1) or ‘alternatively activated’/anti-inflammatory macrophages (M2), characterized by surface receptor expression, mediator secretion, and cell functions [2, 67–70]. With the activation of LPS (TLR ligand) and IFN-ℽ, macrophages are polarized into M1-like macrophages dependent on the TLR4 activation and NF-ĸB facilitation, which are metabolized by glycolysis and identified through the expression of inos (NOS2), IL-1β, IL-6, CCL5, CXCL9, and MMP9 [67–71]. These pro-inflammatory cytokines and inos lead to epithelial barrier disruption and tissue damage due to the deregulation of tight junction proteins and upregulation of epithelial cell apoptosis, predisposing to IBD [72]. The accumulation of iron, cholesterol, and other sterile inflammatory signals in pro-inflammatory macrophages enhances TNF-α production and further impairs wound-healing [57, 73].
Additionally, macrophages stimulated with IL-4 or IL-13 can polarize into M2-like macrophages, committing to oxidative phosphorylation (OXPHOS) and expression of CD206, CD163, CCL17, IRF4, ARG-1, Retnla and Chi3l3 [2, 67, 71]. These M2-like macrophages are involved in tissue repair during inflammation directly or through the regulation of IL-10- and TGF-β1-producing Treg cells [57, 74]. Interestingly, endotoxin-tolerant macrophages also produce IL-10 and arginase 1, participating in tissue repair and immune regulation [75]. However, overactivation of M2-like macrophages has been found to exacerbate pathological fibrosis [57, 76]. Previous studies have demonstrated that cytokines like IL-6, IL-10, and IL-21 facilitate anti-inflammatory function and M2 polarization of macrophages by enhancing IL-4 receptor expression, as characterized by the upregulation of tissue repair and regeneration [40, 57, 77, 78]. In contrast, TNF-α production interferes with M2 polarization of macrophages and thus counteracts tissue repair [57, 73].
Evidence has uncovered that several biological processes are also involved in regulating macrophage polarization. Methyltransferase Setdb2 has been found to facilitate macrophage polarization from inflammatory phenotype to a reparative status by trimethylating histone 3 at NF-κB binding sites on inflammatory cytokine gene promoters [79]. Suppression of DNA methylation through the deletion of DNA methyltransferase 1 (DNMT1) results in the enhancement of alternatively activated macrophages [80]. PGE2 has been demonstrated to downregulate the secretion of pro-inflammatory cytokines and promote the activation of M2 polarization, which is modulated by the cyclic AMP (cAMP)-protein kinase A pathway, cAMP-responsive element-binding protein and Kruppel-like factor 4 [81]. However, other studies suggest that the proliferation of M2-like polarization of macrophages and the expression of M2-related marker genes is suppressed by PGE2 [82]. Besides, PGE2 inhibits oxidative phosphorylation activity via the downregulation of tricarboxylic acid-cycle intermediates [82]. Moreover, macrophage polarization is also modulated by a variety of micro-RNAs (miRs). miR-720 [83], miR-223 [84], miR-127 [85], and miR-155 [86] have been found to contribute to the M1 polarization by targeting GATA3, C/EBPβ, and BCL6, respectively, while miR-378–3p [87], miR-511–3p [88], and miR-146a [87] facilitate the M2 polarization by targeting PI3K/AKT1, ROCK2, Notch1, and PPAR-ℽ, respectively [70].
Unclassical classification criterion
In addition to the classification criterion that identifies macrophages into M1-like and M2-like macrophages, emerging evidence distinguishes macrophages based on the expression of Tim-4 and CD4 [35]. Tim-4, highly expressed by Kupffer cells, acts as an upstream in the apoptotic cell-eradication process [35, 89]. Tim-4+CD4+ macrophages in intestinal mucosa are observed to be maintained locally and rarely replaced by circulating monocytes. In contrast, accumulated monocytes replenish intestinal mucosa via the replacement of Tim-4-CD4- macrophages. Tim-4-CD4+ macrophages ‘bridge’ these two phenotypes and can be slowly interchanged by circulating monocytes [35].
Synthesized analysis of surface markers has manifested the heterogeneity of macrophages and classified human intestinal macrophages into several subsets. Bujko et al. have identified small intestinal macrophages into four subsets according to surface markers, including CD14, CD11b, HLA-DR, and CD11c with the utilization of flow cytometric analysis [6, 47], namely Mϕ1 (CD11bhiCD64+MerTK+CD163+CD115+CX3CR1+CD206-CD1c-CD103-CCR2+calprotectin+), Mϕ2 (CD11b+CD64+MerTK+CD163medCD115+CX3CR1medCD206+CD1c+CD103+CCR2+ calprotectin+), Mϕ3 (CD11b-CD64lowMerTK+CD163+CD115+CX3CR1-CD206+CD1c-CD103-CCR2-calprotectin-), and Mϕ4 (CD11b+CD64lowMerTK+CD163+CD115lowCX3CR1lowCD206+CD1c-CD103-CCR2-calprotectin-). Among them, Mϕ1 and Mϕ2 equip the comparable characteristics as circulating monocytes, while Mϕ3 and Mϕ4 share the same features of mature macrophages [47]. In addition, the locations of these populations are distinct from each other. Mϕ3 is predominantly located in the intestinal villus and forms a network in the LP, whereas Mϕ4 is mainly in the submucosa [47].
Additionally, human intestinal macrophages can also be phenotyped into 2 subsets based on the expression of CD11c, CCR2, and CX3CR1 [90]. Macrophages are identified as CD45+HLA-DR+CD14+CD64+ cells, and the subsets are divided into monocyte-like cells (CD11chiCCR2+CX3CR1+), macrophage-like tissue-resident cells (CD11c-CCR2-CX3CR1-). LPS triggers the expression of IL-1β in monocyte-like cells but suppresses the expression of IL-10 in macrophage-like cells. It should be noted that the enrichment of monocyte-like cells in intestinal mucosa may contribute to the retention of inflammation in IBD patients, while macrophage-like tissue-resident cells are generated during resolution [90].
With the assistance of scRNA-seq, our recent study has unraveled a proinflammatory population (Ccr2+Il-1β+Tlr2+Cx3cr1-Cd163-Mrc1-Ly6c+) of the monocyte/macrophage lineage from LP CD11b+ cells. During intestinal inflammation in Mcpip1∆Mye mice, the maturation from monocytes to macrophages is predominantly arrested, characterized by the enrichment of proinflammatory monocytes subset (Ccr2+Il-1β+Tlr2+Cx3cr1-Cd163-Mrc1-Ly6c+), leading to the aggravation of gut inflammation via an Atf3-Ap1s2 axis-dependent manner [10]. After that, a subset of monocyte-macrophage lineage equipped with unique marker genes (e.g., Cx3cr1, Cd163, Mrc1) is identified as mature macrophages, while newly recruited monocytes are identified according to their unique marker genes (e.g., Ccr2, Il-1β, Tlr2, Ly6c).
The classification criterion shown above is far from exhaustion, and we believe that the analysis of macrophage phenotypes will continue to be improved with advanced technologies (e.g., scRNA-Seq) in the future.
The functions of macrophages
The monocyte-macrophage lineage in intestinal LP is indispensable in maintaining intestinal mucosal homoeostasis [6, 7, 11, 24, 35]. Macrophages located snugly under the epithelial layer possess high phagocytic and bactericidal activity and can capture and eliminate pathogenic bacteria that penetrate the lining of the gut [2, 7, 11, 20, 41, 91–93]. Specifically, after being stimulated by pathogenic bacteria or its derivates, hyporesponsive macrophages generate transepithelial dendrites, which can capture Salmonella organisms as well as soluble antigens in the lumen [2, 11, 94, 95]. Accordingly, the antigens are then presented to neighboring migratory dendritic cells, thus contributing to subsequent immunomodulation [2]. Besides pathogenic microorganisms, macrophages can also eliminate apoptotic or senescent cells and cell debris, contributing to host defense of intestinal mucosal homoeostasis [2, 11, 31, 35, 57, 91, 92, 96]. In contrast, eradication of mature macrophage leads to the augmentation of intestinal permeability and epithelial cell death, thus driving the aggravation of intestinal inflammation [6, 97].
Increasing lines of evidence have shown that macrophages produce several types of pro-inflammatory or anti-inflammatory cytokines, chemokines, TLRs, METs, and lipid mediators, which regulate intestinal immune homeostasis [2, 3, 11, 40]. Of note, IL-10 is the most critical cytokine responsible for the modulation of the responses of macrophages to pattern recognition receptor triggering as well as the maintenance of the amplification of FoxP3+ Treg cells and ILC3 in the intestinal mucosa [3, 34, 41, 78, 93, 98, 99]. Macrophage-derived IL-1β is also involved in the survival of IL-17-producing CD4+ T cells and CSF2 secretion by ILC3 [3, 100, 101]. Moreover, macrophages are one of the primary sources of chemokines, including CCL2, CCL8, CXCL1, and CXCL2, leading to the recruitment of neutrophils, monocytes, and T cells [3, 31, 50]. Macrophages are also required to maximize the bactericidal activity of neutrophils by facilitating suicidal NETosis, a process through which neutrophils form and release neutrophil extracellular traps (NETs) to capture and eliminate bacteria. Simultaneously, NETs promote the phagocytosis and antibacterial activity of macrophages synergistically [102]. Similar to the NETs, METs, composed of cellular DNA, citrullinated histone H3, and MPO, are released from macrophages to capture, immobilize, and kill microorganisms [103, 104]. METs have been clarified to be involved in several pathological processes, including rhabdomyolysis-induced acute kidney injury, iron overload-related liver ischemia/reperfusion injury, and the phagocytosis of Candida albicans [103, 105]. However, the role of METs in orchestrating intestinal mucosa immunity has hitherto not been elucidated. Besides, a full range of TLRs is generated by intestinal tissue-resident macrophages [43] and responsible for the downregulation of several adapter molecules, including CD14, MyD88, TRAF-6, MD2, TRIF, and IRAK1 [20, 50], leading to the hyporesponsiveness of these macrophages. Concomitantly, TLR signaling and NF-ĸB activation are suppressed by IRAK-M and IkBNS [50], which are overexpressed in intestinal-resident macrophages [11]. It should be noticed that macrophages also interfere with cell metabolism, facilitate apoptosis, deteriorate ischemic injury through producing reactive oxygen species (ROS) and toxic mediators, and subsequently impede mucosa healing [40].
Macrophages in intestinal LP have also been revealed to be involved in fundamental physiological processes, including maintaining intestinal barrier integrity, proliferation of epithelial cells, angiogenesis, and tissue repair. Through generating mediators like hepatocyte growth factor (HGF) [106], PGE2 [64, 107, 108], WNT ligands [109, 110], and metalloproteinases [31] as well as signaling pathways including NOX1 signaling activated by annexin A1 [111, 112] and CREB-WISP1 signaling activated by IL-10 [113], intestinal macrophages contribute to the renewal of intestinal stem cells in intestinal crypts, proliferation of epithelial progenitors, and promotion of arteriogenesis, leading to the preservation integrity of the mucosa and recovery of ischemic tissue with minimal collateral damage [2, 3, 6, 11, 29, 31, 35, 40, 57, 107]. Besides, MyD88 signaling secreted from myeloid cells also drives intestinal epithelial repair [11, 114]. Interestingly, GeneChip analysis combined with immunostaining and electron microscopy has pointed out that macrophages can connect to colonic epithelial stem cells directly near the crypt base and thus induce epithelial proliferation [7, 107]. Previous studies have confirmed that macrophages synthesize polyamines via a mTORC1-arginase-1-dependent manner that is being taken up by the epithelial cells, contributing to the triggering of metabolic reprogramming and the enhancement of proliferation at a steady or inflammatory state [115]. In addition to epithelial cells, intestinal macrophages also regulate Paneth and goblet cell generation, since macrophage depletion by anti-CSF1R treatment interferes with the differentiation of LGR5+ intestinal stem cells into intestinal epithelial cells, Paneth cells and goblet cells [2, 6, 116]. Additionally, deficiency of intestinal macrophages leads to the depletion of VE-cadherin+ blood vessels and deterioration of the submucosal vascular network [7, 34, 40, 117, 118].
Emerging evidence demonstrated a specialized function of resident macrophages of the muscularis externa (MMϕ) in the enteric nervous system (ENS). ENS, composed of the submucous plexus and the myenteric plexus, controls different intestine processes independently of the brain or spinal cord [6]. MMϕ has been demonstrated to play an indispensable role in meliorating the nascent ENS by pruning synapses and phagocytosing enteric neurons. MMϕ deficiency causes a caspase3-dependent neuronal loss and thus contributes to peristalsis alteration, intestinal secretion downregulation, and intestinal transit abnormity [6, 34, 119]. However, the regulation of intestinal contractility and motility is a consequence of the paracrine release of PGE2 from intestinal macrophages in a neuron-independent manner [120]. In addition, MMϕ are feedback-regulated by the ENS. Contacting with ENS and ENS-derived CSF1 or TGF-β maintains MMϕ homeostasis and facilitates its differentiation into a neurosupportive, long-lived and self-maintained phenotype, contributing to the formation of ENS and maintenance of intestinal transit [6, 21, 34, 119]. Swift activation of extrinsic sympathetic neurons in cooperation with norepinephrine signaling to β2 adrenergic receptors on MMϕ enhances tissue-protective programs [21].
Apart from the data mentioned above, intestinal macrophages exert an indispensable role in intestinal fibrosis [6, 15, 40, 57, 121, 122]. Matrix metalloproteinases (MMPs) derived from macrophages are involved in the regulation of fibrin and collagen turnover through the degradation of extracellular matrix (ECM) proteins [57]. Macrophages generate mediators including TGF-β1, IL-36, platelet-derived growth factor (PDGF), insulin-like growth factor 1 (IGF-1), and connective tissue growth factor (CTGF) to directly activate fibroblasts, leading to the development of fibrosis and facilitation of wound healing [1, 15, 40, 57, 121–124]. In contrast, macrophages also participate in the inhibition of fibrosis directly via the production of IL-10, RELMα, and ARG1 [125–127] and indirectly through the suppression of CD4+ T cell proliferation and fibroblast activation [40, 121]. However, the dysregulated immune response of intestinal macrophages leads to inefficient tissue wound-healing in the intestinal mucosa, contributing to chronic tissue injuries or chronic inflammation characterized by an infiltration of immune cells and pathological fibrosis or scarring. This aberrant repair culminates with organ failure and death [40, 57, 76]. Consequently, the accumulation, activation, and elimination of monocytes and macrophages are tightly regulated during gut inflammation.
Macrophages during gut inflammation
Intestinal pathological encounters, including IBD, predispose to a massive infiltration of CD14+ monocytes (equivalent as Ly6Chi monocytes in mice) through the CCL3-CCR1 chemokine axis, which differentiate into macrophages to regulate intestinal immune homeostasis and resolve mucosal wound or inflammation [3, 8, 11, 26, 29, 35, 128–130]. The dysregulation of intestinal macrophages results in intolerance to commensal bacteria and food antigens, and concomitantly contributes to chronic pathological disorders in the gut, including IBD [2, 24, 35, 43, 131, 132]. Here, we introduce the divergent functions of the monocyte-macrophage lineage during IBD (Fig. 1, graphical abstract).
Fig. 1.

Potential roles of macrophages in intestinal mucosa. The infiltration of circulating monocytes is triggered by a CCL2-CCR2 axis, which consistently maturate into macrophages to modulate intestinal immunity and homeostasis. Macrophages involve in phagocytosis and bactericidal effects, and response to exogenous antigens and microbiotas. Macrophage extracellular traps (METs) are released to capture, immobilize and kill microorganisms. In addition, macrophages upregulate the production of cytokines (e.g., IL-1b, IL-6, IL-10, IL-12, IL-23, TGF-b, and TNF-a) and chemokines (e.g., CCL8, CXCL1, CXCL2, CXCL9, and CXCL10), resulting in the regulation of proliferation, differentiation and immune response of neutrophils and T cells. Additionally, macrophages also contribute to tissue repair with the enhancement of epithelial cell proliferation, angiogenesis and fibrosis
The maturation of macrophages during IBD
Evidence has shown that monocyte maturation to macrophages is arrested at P2 stage (immature macrophage, P2, Ly6C+MHC II+) during gut inflammation, leading to the enhancement of proinflammatory monocytes (P1, Ly6ChiMHC II-) and immature macrophage (P2, Ly6C+MHC II+), but exhaust mature macrophages (P3/P4, Ly6C-MHC II+CD64+) [10, 28, 30, 43]. During IBD, infiltrated monocytes and immature macrophages in inflamed mucosa stay responsive to TLRs, contributing to an upregulation of pro-inflammatory mediators (e.g., IL-1β, IL-6, IL-12, TNF-α, iNOS) and bolster respiratory burst activity, and downregulation of IL-10, leading to monocyte infiltration, tissue damage and function deterioration [11, 24, 28, 40, 45, 50, 133, 134]. Mechanically, these pathological processes contribute to the exacerbation of intestinal immune inflammation. Relevant mediators and immune cells in intestinal niches are involved in regulating the monocyte-macrophage maturation process under inflammatory conditions. Since the infiltration and development of monocytes are strictly dependent on CCR2 expression, the enhancement of myeloid cell-derived CCR2 expression in intestinal niches interacts with their ligand CCL2 during inflammation to induce the recruitment of monocytes and immature macrophages and fills the empty niche [28]. Additionally, an upregulation of macrophage maturation in the intestine has been demonstrated after neutralizing with an anti-granulocyte–macrophage colony-stimulating factor (GM-CSF) antibody [135]. In line with this, an IFNℽ-STAT1 axis is required for the maturation of monocytes in the intestine of DSS-challenged mice by triggering histone acetylation at the promoter regions of the Tnf and Nos2 loci [61]. According to our recent study, MCPIP1 deficiency enhances the infiltration of circulating monocytes and restricts monocyte-macrophage maturation via the ATF3-AP1S2 pathway in the intestinal mucosa, leading to the deterioration of intestinal mucosal inflammation upon DSS insult [10]. Synchronously, depletion of innate lymphoid cells (ILC) has been found to restrain the maturation of intestinal monocyte-macrophage lineage, but the number of monocytes remains unchanged in the DSS-induced colitis model [135]. Except for the proinflammatory mediators and immune cells, microbiota and its metabolites in the intestinal niches also participate in the recruitment and maturation of monocyte-macrophage lineage during inflammation [49]. Nevertheless, the retardation of macrophage maturation during intestinal inflammation causes the attenuation of bactericidal and phagocytic ability [2, 136, 137]. In DSS-induced murine colitis model, a transient loss of resident macrophages is uncovered, resulting in an empty macrophage niche in inflammatory intestinal mucosa [138, 139], which is partly ascribed to the affection of sulfated polysaccharides [139]. In addition, the elimination of microbiota in the intestinal downregulates replenishing the “emptied” niches during gut inflammation, suggesting a commensal-dependent refilling manner in the niche [49].
Phenotypic changes of macrophages during IBD
To further classify intestinal macrophages from IBD patients, we retrieved the publicly available scRNA-seq data, including 11 samples from Crohn's disease patients [128] and 18 samples from ulcerative colitis patients [129]. We painted the landscapes of monocyte-macrophages and identified 5 distinct subsets including monocytes, C1QB+ macrophages, HSPA1B+ macrophages, IL-1B+ macrophages, and TMSB4X + macrophages in the intestinal mucosa from Crohn’s disease patients (Fig. 2a). Notably, TMSB4X, RPLP1, and RPS6 are found to be highly expressed in both C1QB+ macrophages and TMSB4X+ macrophages from Crohn’s disease patients, while CCL3, CCL4 and CCL3L3 are highly expressed in both IL-1B+ macrophages and HSPA1B+ macrophages, showing a similar convergence in biological characteristics and functions between these two subpopulations (Fig. 2b). Moreover, we also portrayed 5 subpopulations of monocytes, C1QB+ macrophages, CALD1+ macrophages, IL-1B+ macrophages, and SERPINA1+ macrophages in the intestinal mucosa from ulcerative colitis patients (Fig. 2c). Interestingly, APOE, C1QB, and matrix metalloproteinase 12 (MMP12) are highly expressed in C1QB+ macrophages, IL-1B+ macrophages and CALD1+ macrophages from ulcerative colitis patients (Fig. 2d). Additionally, we then leveraged a monocle2 analysis approach and confirmed the origin of IL-1B+ macrophages among monocyte-macrophages from both Crohn’s disease and ulcerative colitis patients. In parallel, C1QB+ macrophages appear to be mature macrophages (Fig. 2e-f).
Fig. 2.

Identification and characteristics of the monocyte-macrophage lineage in intestinal mucosa from IBD patients. (A, C) UMAP plots defines 8 clusters in CD (A) and UC (C) patients, respectively. (B, D) Dot plots showing expression levels of selected signature genes of the monocyte-macrophage lineage subsets in intestinal mucosa from CD (B) and UC (D) patients. Dot size indicates fraction of expressing cells, colored based on the relative expression of specific gene. (E, F) The developmental trajectory of the monocyte-macrophage lineage subsets in CD patients (E) and UC patients (F) inferred by Monocle2
Macrophages modulate intestinal and extraintestinal homeostasis during IBD
Dysregulation of intestinal immune response results in an increase in myeloid cell-derived cytokines, chemokines as well as other pro-inflammatory mediators, which further influence the fates and functions of immune cells. GM-CSF, an ILC3-derived or IL-23-stimulated mediator, has been found to contribute to a powerful payload in elevating the expression of IL-1β and IL-23 by monocytes-macrophages lineage within the context of C. rodentium infection or DSS-induced colitis [8, 40, 135, 140]. TNF-α and iNOS are considered to be representative colitogenic mediators derived from monocyte-macrophage lineages that contribute to the development of colitis [61, 141]. Infiltrated-monocytes and immature macrophages are also enriched of pro-inflammatory mediators including TREM1, TREM2, S100A8, S100A9, CXCL2, CXCL8, SPP1, GPNMB, AREG, HBEGF, NRG1 and SOD2, leading to the persistence of intestinal inflammation [10, 38, 130]. Evidence has been emerging that colitis-associated monocytes upregulate the expression of TREM1, which is reckoned as a potent amplifier of pro-inflammatory responses [45, 62, 142]. Consistently, TREM1 deficiency compromises pro-inflammatory features of the monocyte-macrophage lineages, thus culminating in the inhibition of colitis in mice [8]. However, intestinal mucosal inflammation also induces anti-inflammatory features of the monocyte-macrophage lineage, which is instrumental in the secretion of anti-inflammatory mediators, including IL-10, TGF-β, vascular endothelial growth factor (VEGF), and metalloproteinases with the stimulation of inhibitory mediators, thus participating in the process of wound healing and tissue repair [11, 28, 41, 43, 45, 50, 57, 78, 113, 143]. Notably, macrophage cell surface receptors, including PD-L1 and PD-L2, are involved in the suppression of inflammation and ultimately delay the injury repair [57, 78]. Nonetheless, evidence has shown that anti-inflammatory cytokines are transferred to lysosomes and degraded rather than exerted functions in the intracytoplasmic of Crohn’s disease macrophages, and this dysregulated process is triggered by E. coli or TLR agonists [136]. These data suggest that compromised macrophage cytokine secretion underlies acute inflammation in Crohn’s disease.
Apart from cytokine secretion, the monocyte-macrophage lineage contributes to the process of epithelial cell proliferation, angiogenesis, fibrosis, and tissue repair, while tissue repair fails due to the domination of proinflammatory macrophages and the inappropriate functions of macrophages in the inflamed intestinal mucosa [57, 144]. After stimulating with inflammatory signaling, intestinal macrophages are also preferentially endowed with a feature of wound healing by regulating the generation of WNT [109, 110], MMPs [31], PGE2 [108], VEGF [40], HGF [106], as well as the signaling pathways like NOX1 signaling activated by annexin A1 (ANXA1) [111, 112], mTORC1 − arginase-1 axis [115], CSF1R-related signaling [2, 6, 116], CREB-WISP1 axis [113], and MyD88 signaling [11, 114], which are involved in the intestinal stem cell proliferation and its differentiation into intestinal epithelial cells, Paneth cells and goblet cells, thus contributing to tissue repair of inflamed intestine. Previous data have uncovered that macrophages from Crohn’s disease patients produce less HGF than healthy controls, leading to ineffective epithelial repair [106]. Hypoxic macrophages produce WNT in a HIF-1-dependent manner, which impairs epithelial autophagy in the intestinal mucosa of IBD patients [109]. The activation of WNT signaling during TNBS-induced colitis also contributes to wound healing in a STAT6-dependent manner [110]. Macrophage-derived PGE2 activates the WNT/β-catenin signaling in intestinal stem cells and facilitates stem cell self-renewal by binding to its receptors EP1/EP4. This process can be further amplified after a combination of 5-hydroxytryptamine and its receptors HTR2A/3A in macrophages [108]. During gut inflammation, the expression of VEGF from macrophages is upregulated, leading to the activation of perivascular fibroblasts and the reduction of CXCR7 expression [40]. ANXA1 and ROS have been found to be increased in the intestinal epithelia cells and infiltrating immune cells in the inflamed mucosa of patients with ulcerative colitis, which contribute to tissue repair during intestinal inflammation in a NOX1-dependent manner [111, 112]. Moreover, intestinal microbiota orchestrates the expression of NOX1 by TLR signaling to maintain intestinal immune homeostasis [112], and intestinal macrophages can also produce polyamines to synergistically induce intestinal epithelial cell proliferation via the mTORC1 − arginase-1 axis [115]. It is noteworthy that the CSF/CSF1R signaling acts as a modulator of intestinal epithelial differentiation, except for its affection for the intestinal monocyte-macrophage maturation. Conversely, macrophage depletion through CSF1R blockage leads to the failure of intestinal epithelial cell differentiation [116]. An upregulation of macrophage-generated IL-10 in the context of intestinal inflammation activates epithelial CREB and facilitates synthesis and secretion of the pro-repair WISP-1 subsequently [113]. On the contrary, IL-10 deficiency promotes the apoptosis of intestinal epithelial cells in mice, which is reversed via the neutralization of TNF-α and iNOS [61, 141]. Additionally, evidence has emerged that Myd88 signaling pathway is indispensable in the enrichment of intestinal Ptsg2-expressing stromal cells and the activation of colonic epithelial response to inflammation during DSS-induced mouse colitis [114]. With the application of novel technologies, there is potential to further explore the underlying mechanisms whereby macrophages are involved in tissue repair and intestinal epithelial proliferation.
Accumulating lines of evidence have suggested that fibrosis contributes to the intestinal tissue repair after an inflammatory attack, but dysregulation of intestinal mucosal fibrosis leads to pathological fibrosis or even scarring [6, 8, 34, 38, 40, 57, 144]. Mechanically, macrophages are involved in the recruitment and differentiation of tissue fibroblasts into myofibroblasts and upregulate the synthesis of ECM components [57, 121] via the production of MMPs, tissue inhibitor of metalloproteinases 1 (TIMP1), PDGF, TGFβ1, IGF-1 and CTGF [57, 121, 122]. MMPs participate as enzymes for the degradation of ECM proteins or triggers for fibrosis [57]. For example, macrophage-derived MMP9 and MMP12 are markedly induced in the context of IL-13 stimulation or CCL4 and thioacetamide-induced liver fibrosis and subsequently promote fibrosis [145, 146]. The TGFβ-dependent activation of MMP2 is negatively regulated by BMP7 [147]. In contrast, macrophage-derived TIMP1 contributes to the normalization of the pro-fibrotic hematopoietic-vascular niche, and these macrophages are recruited by endothelial-produced endocrine chemokines [148]. PDGF and TGFβ1 have long been recognized as macrophage-derived growth factors that promote angiogenesis and fibrosis, thus playing an indispensable role in tissue rehabilitation [149]. S Typhimurium infection results in chronic colitis, as evidenced by transmural ECM deposition within the intestine and the activation of fibrotic response, concomitantly with the enhancement expression of TGFβ1 and IGF-1 [150]. CTGF, identified as a fibrotic marker, has been found to be upregulated during inflammatory diseases, which further facilitates the expression of pro-inflammatory cytokines and chemokines to participate in the fibrotic process. On the contrary, the elimination of CTGF in mice restrains the expression of pro-inflammatory mediators and fibrosis-associated biological processes [150]. Moreover, IL-36α+ macrophages and immature macrophages in the intestinal mucosa of IBD patients have been reported to be strongly associated with inefficient tissue wound healing and contribute to the infliction of pathological fibrosis or scarring in the intestinal mucosa, leading to chronic tissue injuries or chronic inflammation, and finally culminating with the occurrence of intestinal stricture, stenosis or obstruction [124, 151]. Through the integration of single-cell transcriptomics and spatial transcriptomics, a recent study has provided compelling evidence to show that hepatocyte growth factor activator (HGFAC) Arg509His (R509H), a risk variant for Crohn’s disease activated by thrombin protease activity, restricts fibroblast-mediated tissue reestablishment in the inflammatory intestine on account of impairing proteolytic activation of the growth factor macrophage-stimulating protein (MSP). Notably, a reduction of growth factors causes an impairment of wound-associated epithelial cell differentiation and retinoic acid (RA) generation, leading to delayed repair [152].
Evidence shows that the functions and maturation process of intestinal macrophages are consistent among animals and humans in both steady and inflammatory states, and prompts that animal models contribute to a potent payload of exploring the definite properties of macrophages in intestinal mucosa [11]. Animal models of intestinal inflammation, such as the DSS-induced colitis model, 2,4,6-trinitrobenzene sulfonic acid (TNBS)-induced colitis model and transfer of radio-labeled autologous blood monocytes into mice, have verified an influx of monocytes as well as eosinophils, and the suppression of macrophage maturation during these situation [10, 11, 43, 50]. In addition, published data have also elucidated that specific microbiota participates in the modulation of macrophage formation and functions in DSS-induced colitis. For example, the enrichment of Enterobacteriaceae microbiota in the inflammatory enteric cavity leads to the accumulation of circulating monocytes. Bacterial hemolysin is instrumental in persisting the activation of macrophage NOD-like receptor thermal protein domain associated protein 3 (NLRP3) inflammasomes, which mediates the excessive expression of IL-1β triggered by the pathogen Salmonella [40]. Synchronously, Helicobacter hepaticus-induced colitis enlightens the paramount role of GM-CSF in recruiting monocytes [40, 140].
Apart from the regulatory processes in the intestine, macrophages are also involved in the modulation of extraintestinal homeostasis in IBD patients, which is ascribed to the interaction crosstalk between the intestine and its accessory tissues, including mesenteric adipose tissue, mesentery, and mesenteric lymph nodes [153, 154]. By analyzing integrative multi-omics between proteomic and microbiome, our recent data have identified that macrophages-derived serum amyloid A2 (SAA2) is involved in the regulation of adaptive immunity in mesentery and mesenteric lymph nodes, and Th17 immunity in mesenteric adipose tissue, which is further confirmed in both serum and fecal samples as a potential diagnostic biomarker [13]. Previous studies have demonstrated that SAA2 is illustrated as a significant inducer to macrophages- or Th17 cell-mediated immunopathology [155]. Our study thus indicates that SAA2 is positively correlated with the microbial transporters ugpE and ugpC and short-chain fatty acids-producing genera, including Dorea and Butyricicoccus. In contrast, the negative correlation between SAA2 and microbial gerKC and flaG is also identified [13]. In addition, macrophage-associated gene arginase-2 (ARG2) is increased in inflamed intestinal mucosa as well as intestinal accessory tissues (i.e., mesenteric adipose tissue, mesentery, and mesenteric lymph nodes) from Crohn’s disease patients [13], and it is found to be localized at the mitochondria and upregulated by the IL-10/miR-155 axis in pro-inflammatory macrophages, thus contributing to the enhancement of oxidative phosphorylation via the suppression of HIF-1α and IL-1β in inflammatory macrophages [156]. Taken together, these conceptions open new horizons for prospects and emphasize a potential future for scrutinizing the feature of macrophages in intestinal accessory tissues from IBD patients (Fig. 3).
Fig. 3.
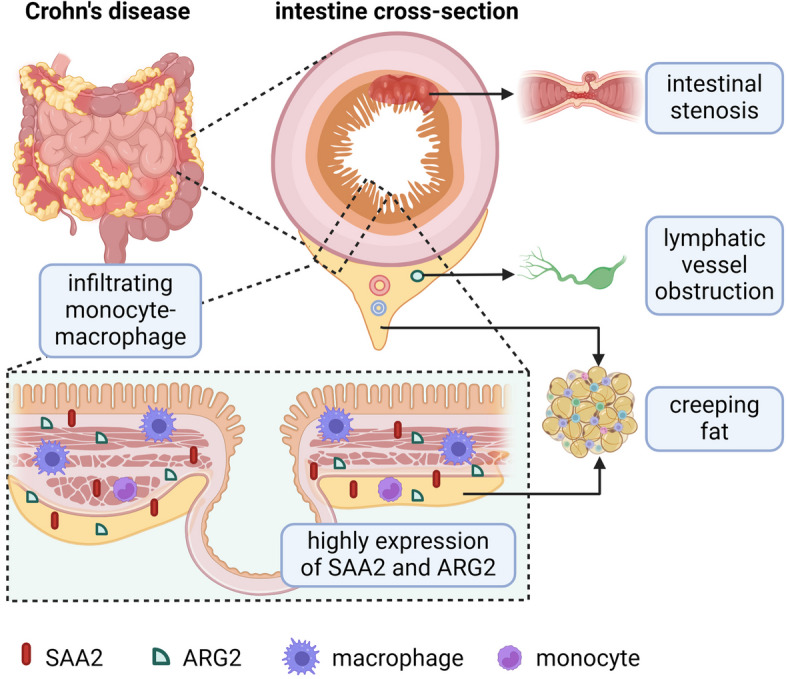
Macrophages modulate mesentery immune homeostasis in IBD patients. The development of creeping fat, mesenteric fibrosis as well as lymphatic vessel obstruction in the mesentery are reckoned as the characteristics of IBD patients. Macrophages infiltrate in the mesentery and involve in the modulation of extraintestinal immune homeostasis in IBD patients via macrophage-associated genes including SAA2 and ARG2, which are highly expressed in the mesentery and intestinal mucosa
Susceptibility genes are associated with the diverse functions of macrophages in IBD
In addition to the delicate immune regulations of intestinal macrophages and the underlying mechanisms as mentioned above, more susceptibility genes have gradually been discovered in IBD patients, which are ascribed to the advancement of nucleic acid sequencing technology, like GWAS and ImmunoChip [14, 16, 17, 157, 158]. Copious amounts of susceptibility genes are found to be related to the maturation and function of monocyte-macrophages lineage (Table 2). Susceptibility loci related to intestinal macrophage functions have been unraveled in Crohn’s disease patients, including NOD2, ATG16L1, CX3CR1, IL12p40, IL23R, JAK2, STAT3, and protein tyrosine phosphatase non-receptor type 2 (PTPN2) [14, 16, 17, 158–160]. NOD2 is the first conferred and one of the most important susceptibility genes of Crohn’s disease [161, 162], which participates in regulating inflammatory response to bacterial antigens [2, 6, 159, 160, 163]. NOD2 recognizes the bacterial-derived muramyl dipeptide by suppressing TLR2-mediated activation of NF-ĸB [164], and the mutation of NOD2 results in the enhancement of TLR2-driven activation of NF-ĸB [165]. By using the DSS-induced colitis model in NOD2-deficient zebrafish, the deficiency of NOD2 is observed to contribute to an impairment in the monocyte-macrophage lineage, while the mutation of NOD2 leads to the overexpression of collagen, all of which are associated with pathological fibrosis [6]. Together with ATG16L1, another susceptibility locus for Crohn’s disease [16, 157], NOD2 triggers the process of autophagy by accumulating at the site where bacteria gather [14, 160, 166, 167]. Autophagy-related mediators, including RIPK2, ATG5, and ATG7 participate in this process [160, 168, 169]. Impaired antifungal responses due to the missense mutation of another IBD-related gene CX3CR1 in myeloid cells lead to an increase in intestinal and extraintestinal inflammatory diseases [2, 170]. Several components of the IL-23 signaling pathway are also confirmed as IBD susceptibility genes, including IL23R, IL12B, STAT3, and JAK2, which help to explain the regulatory effect of these mediators on intestinal mucosal immune homeostasis in IBD patients [160, 167, 171–173]. IL-23 is derived from macrophages and DCs, and induces the expression of IL-17 from T cells, which participates in the regulation of intestinal immune homeostasis [174]. In addition, the JAK2-STAT3 signaling is involved in the modulation of macrophage proliferation [14, 175]. More studies, assisted by scRNA-seq analysis, have revealed the enhancement of proliferation and glycolytic metabolism of inflammatory macrophages in individuals with JAK2 mutation, resulting in the activation of AIM2 inflammasome and the aggravation of inflammation [175]. On the other hand, PTPN2 has been observed as a susceptibility gene to ulcerative colitis [176, 177], which participates in the regulation of the JAK2-STAT3 signaling, modulation of IL-2 responsiveness, and maintenance of intestinal epithelial barrier integrity [176, 178].
Table 2.
IBD-related susceptibility genes are associated with macrophage functions
| Gene name | Functions |
|---|---|
| NOD2 | 1) Regulate inflammatory response to bacterial antigens [2, 6, 159, 160, 163] |
| 2) Recognize bacterial-derived muramyl dipeptide [164] | |
| 3) Suppress TLR2-mediated activation of NF--B [164, 165] | |
| 4) Maintain the homeostasis of monocyte-macrophage lineage [6] | |
| 5) Regulate collagen production and fibrosis [6] | |
| 6) Trigger autophagy process [160, 166, 167] | |
| ATG16L1 | Trigger autophagy process [160, 167] |
| CX3CR1 | Involve in invading pathogen process and antifungal responses [2, 170] |
| JAK2 | 1) Modulate macrophage proliferation [14, 175] |
| 2) Induce T cell-mediated IL-17 [174] | |
| STAT3 | 1) Modulate macrophage proliferation [14, 175] |
| 2) Induce T cell-mediated IL-17 [174] | |
| IL23R | Induce T cell-mediated IL-17 [174] |
| PTPN2 | 1) Regulate the JAK2-STAT3 signaling [176] |
| 2) Maintain the integrity of intestinal epithelium [176] | |
| 3) Modulate IL-2 responsiveness [178] | |
| LILRB3 | Enhance M2-like macrophage polarization [179] |
| IL21R | Amplify macrophage activation [180] |
| GTF2I | Modulate M2-like macrophage polarization [181] |
| RUNX3 | 1) Maintain of the expression of CD4 and CD14 in the monocyte-macrophage lineage [182] |
| 2) Orchestrate macrophage maturation [182] |
NOD2 nucleotide binding oligomerization domain containing 2, ATG16L1 autophagy related 16 like 1, CX3CR1 C-X3-C motif chemokine receptor 1, JAK2 Janus kinase 2, STAT3 signal transducer and activator of transcription 3, IL23R interleukin 23 receptor, PTPN2 protein tyrosine phosphatase non-receptor type 2, LILRB3 leukocyte immunoglobulin like receptor B3, IL21R interleukin 21 receptor, GTF2I general transcription factor Iii, RUNX3 RUNX family transcription factor 3
Recently, we have verified 54 new IBD-associated genetic loci in East Asian ancestry, like RUNX3, ADAP1, IL21R, GTF2I, and LILRB3 [17]. Notably, deficiency of LILRB3 facilitates macrophage polarization into an pro-inflammatory M1-like phenotype, contributing to the worsening of intestinal inflammation [179]. IL21R has been identified to promote macrophage activation, participate in pathogen-induced Th2 responses, and play a paramount role in inflammation and chronic fibrotic diseases [180]. The mutation of GTF2I fails to the induction of anti-inflammatory M2-like polarization in macrophages [181]. RUNX3 is a CXCL12-dependent transcription factor that is involved in the maintenance of the expression of CD4 and CD14 in the monocyte-macrophage lineage, which orchestrates the maturation of macrophages in the intestine [182].
Macrophage-directed therapeutics for IBD
Given the paramount role and heterogeneity of macrophages in the development of IBD [130], we therefore interrogate the macrophage-directed IBD therapeutics. Based on the mechanisms underlying immunoregulation of macrophages on intestinal immune homeostasis, therapeutic approaches focus on modulating processes including phagocytosis, bactericidal effect, secretion of mediators, involvement in tissue repair and fibrosis, and the maturation of macrophage [2]. Classical drugs for treating IBD directly inhibit inflammation of macrophages in the intestinal mucosa. Corticosteroids suppress the activation of NF-ĸB and activator protein 1 (AP-1), thus contributing to monocyte maturation to macrophage [183, 184]. Mutually, the activation of NF-ĸB is also downregulated by 5-aminosalicylate [185]. In parallel, methotrexate is reckoned as an inhibitor of thymidylate synthase, which involves pro-inflammatory cytokine expression from macrophages [186]. In addition, azathioprine and 6-mercaptopurine are involved in the reduction of JUN N-terminal kinase (JNK) phosphorylation, which is ascribed to the metabolized 6-thioguanine triphosphates-dependent Ras-related C3 botulinum toxin substrate 1 (Rac1) activity [187]. With the increasing application scenarios and diversified types of biologics, the immunomodulatory effects of biological therapies have been investigated in IBD. Evidence has demonstrated that administration of anti-TNF mAb (e.g., infliximab) could facilitate maturation of CD68+CD206+ macrophages in the intestinal mucosa of IBD patients and concomitantly inhibiting T cell proliferation [6, 188]. Moreover, previous studies have also elucidated that another anti-TNF mAb (e.g., adalimumab) enables to constrain immature macrophage (CD14+HLADRint) infiltration in inflamed intestine of Crohn’s disease patients [6, 189]. Since the JAK2-STAT3 signaling is involved in the modulation of macrophage maturation and proliferation [14, 175], a target therapeutic approach using JAK inhibitors has become a hot topic worldwide. Tofacitinib, as a JAK 1/3 inhibitor, appears to suppress pro-inflammatory M1 macrophages and promotes anti-inflammatory M2 macrophages and M2-associated markers, thus contributing to maintaining intestinal barrier function through regulating the expression of tight junction protein, constraining the JAK2-STAT3 signaling, and downregulating the secretion of IL-6 and IL-22 [176, 190]. Taken together, therapeutic strategies through suppressing monocyte infiltration, downregulating the expression or the activation of pro-inflammation cytokines, bolster the maturation and M2-like polarization of macrophages, and upregulating the secretion of anti-inflammation cytokines may contribute to the resolution of intestinal inflammation in IBD.
Conclusion
Emerging evidence has highlighted the notion that macrophages possess a striking degree of plasticity, heterogeneity, and adaptation, which makes it necessary to define macrophage subpopulations in the intestine according to their features and functions. Importantly, these data could provide benefits for a precise directional effect on translational study, diagnosis, and precision medicine in IBD [2, 35, 47, 67, 90]. Except for intestinal macrophages, extraintestinal monocyte-macrophages also gain attention from recent studies, especially mesenteric monocyte-macrophages in IBD [13, 153, 154]. SAA2 and ARG2 have been observed to upregulate in inflamed intestinal mucosa and intestinal accessory tissues, being associated with immunoregulation on intestinal macrophages [13]. Given the plasticity and heterogeneity of intestinal macrophages, as well as the diverse functions performed by extraintestinal monocyte-macrophages, which may also influence the immune homeostasis in IBD, the simplex paradigm has been challenged, improved, and enlarged. To better illustrate the precise molecular interactions between macrophages and other immune cells and dissect the potential immunoregulation of the newly identified susceptibility genes on macrophages in intestinal mucosa, cell–cell crosstalk and niche-specific functions of intestinal macrophages have gained a diverse array of attention, and an in-depth investigation on the underlying mechanisms is warranted [7, 8, 13, 29].
Decades ago, studies on IBD patients were limited by the acquisition of samples and the reconstruction of the intestinal microenvironment. It is encouraging and increasingly apparent that tremendous strides made in research of intestinal macrophage biology are a consequence of novel technology advances, including scRNA-seq [10, 29, 128], spatial transcriptomics (ST) [191–193], CosMx Spatial Molecular Imaging [130], seq-scope [194], GeoMx Digital Spatial Profiler (DSP) [195, 196], spatially-resolved transcript amplicon readout mapping (STARmap) [197], spatially enhanced resolution omics-sequencing (Stereo-seq) [198], and omics technologies [13]. The development of novel technologies responsive to the unmet need of researches will help us to understand better the pathogenesis of IBD, the relationship between macrophages and the intestinal accessory tissues in IBD, and the features of the newly founded susceptibility genes of IBD, which is foreseeable to get rid of limitations and emphasize prospects for potential therapeutic intervention.
Acknowledgements
We would like to express our gratitude to physicians and PhD students from the Center for IBD Research, Departments of Gastroenterology and Pathology at the Shanghai Tenth People’s Hospital of Tongji University.
Abbreviations
- AGM
Aorta-gonad-mesonephros
- ARG2
Arginase-2
- AP-1
Activator protein 1
- CSF1
Colony-stimulating factor 1
- CSF1R
Colony-stimulating factor 1 receptor
- CTGF
Connective tissue growth factor
- DSP
Digital Spatial Profiler
- DSS
Dextran Sulfate Sodium Salt
- ECM
Extracellular matrix
- EMP
Erythro-myeloid precursors
- GF
Germ-free
- GWAS
Genome-wide association studies
- GM-CSF
Granulocyte–macrophage colony stimulating factor
- HGF
Hepatocyte growth factor
- HSCs
Hematopoietic stem cells
- IBD
Inflammatory bowel disease
- IGF-1
Insulin-like growth factor 1
- JNK
JUN N-terminal kinase
- LP
Lamina propria
- METs
Macrophage extracellular traps
- MHC II
Major histocompatibility complex II
- MMPs
Matrix metalloproteinases
- NLRP3
NOD-like receptor thermal protein domain associated protein 3
- OXPHOS
Oxidative phosphorylation
- PDGF
Platelet-derived growth factor
- PGE2
Prostaglandin E2
- Rac1
Ras-related C3 botulinum toxin substrate 1
- ROS
Reactive oxygen species
- SAA2
Serum amyloid A2
- scRNA-seq
Single-cell RNA-sequencing
- ST
Spatial transcriptomics
- STARmap
Spatially-resolved transcript amplicon readout mapping
- Stereo-seq
Spatial enhanced resolution omics-sequencing
- TNBS
2,4,6-Trinitrobenzene sulfonic acid
- TREM1
Triggering receptor expressed on myeloid cells 1
- TS
Thymidylate synthase
- YS
Yolk sac
- ZEB2
Zinc finger E-box-binding homeobox 2
Authors’ contributions
ZL conceived the study; HL conception and design of the review, selected studies for inclusion, and drafted the manuscript; SZ, JL and YC interpreted the data; HL and SZ prepared figures. All authors approved the final draft.
Funding
This work was sponsored by grants from the National Natural Science Foundation of China (82300585, 82370532, 82341219), Shanghai Sailing Program (23YF1432700).
Availability of data and materials
No datasets were generated or analysed during the current study.
Declarations
Ethics approval and consent to participate
Ethical approval was obtained from Ethics Committee of the Shanghai Tenth People’s Hospital of Tongji University (SHSY-IEC-4.0/19–52/01).
Consent for publication
The authors declare that they have no competing interests.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Huiying Lu, Zhimin Suo and Jian Lin contributed equally to this work.
References
- 1.Schett G, McInnes IB, Neurath MF. Reframing immune-mediated inflammatory diseases through signature cytokine hubs. N Engl J Med. 2021;385:628–39. 10.1056/NEJMra1909094 [DOI] [PubMed] [Google Scholar]
- 2.Na YR, Stakenborg M, Seok SH, Matteoli G. Macrophages in intestinal inflammation and resolution: a potential therapeutic target in IBD. Nat Rev Gastroenterol Hepatol. 2019;16:531–43. 10.1038/s41575-019-0172-4 [DOI] [PubMed] [Google Scholar]
- 3.Mowat AM, Scott CL, Bain CC. Barrier-tissue macrophages functional adaptation to environmental challenges. Nat Med. 2017;23:1258–70. 10.1038/nm.4430 [DOI] [PubMed] [Google Scholar]
- 4.He C, Yu T, Shi Y, Ma C, Yang W, Fang L, et al. MicroRNA 301A promotes intestinal inflammation and colitis-associated cancer development by inhibiting BTG1. Gastroenterology. 2017;152:1434-48.e15. 10.1053/j.gastro.2017.01.049 [DOI] [PubMed] [Google Scholar]
- 5.Zhou G, Yu L, Fang L, Yang W, Yu T, Miao Y, et al. CD177+ neutrophils as functionally activated neutrophils negatively regulate IBD. Gut. 2018;67:1052–63. 10.1136/gutjnl-2016-313535 [DOI] [PubMed] [Google Scholar]
- 6.Delfini M, Stakenborg N, Viola MF, Boeckxstaens G. Macrophages in the gut: masters in multitasking. Immunity. 2022;55:1530–48. 10.1016/j.immuni.2022.08.005 [DOI] [PubMed] [Google Scholar]
- 7.Viola MF, Boeckxstaens G. Niche-specific functional heterogeneity of intestinal resident macrophages. Gut. 2021;70:1383–95. 10.1136/gutjnl-2020-323121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hegarty LM, Jones GR, Bain CC. Macrophages in intestinal homeostasis and inflammatory bowel disease. Nat Rev Gastroenterol Hepatol. 2023;20:538–53. 10.1038/s41575-023-00769-0 [DOI] [PubMed] [Google Scholar]
- 9.Mass E, Nimmerjahn F, Kierdorf K, Schlitzer A. Tissue-specific macrophages: how they develop and choreograph tissue biology. Nat Rev Immunol. 2023;23:563–79. 10.1038/s41577-023-00848-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lu H, Zhang C, Wu W, Chen H, Lin R, Sun R, et al. MCPIP1 restrains mucosal inflammation by orchestrating the intestinal monocyte to macrophage maturation via an ATF3-AP1S2 axis. Gut. 2023;72:882–95. 10.1136/gutjnl-2022-327183 [DOI] [PubMed] [Google Scholar]
- 11.Bain CC, Mowat AM. Macrophages in intestinal homeostasis and inflammation. Immunol Rev. 2014;260:102–17. 10.1111/imr.12192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Heresbach D. Frequency and significance of granulomas in a cohort of incident cases of Crohn’s disease. Gut. 2005;54:215–22. 10.1136/gut.2004.041715 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gao X, Sun R, Jiao N, Liang X, Li G, Gao H, et al. Integrative multi-omics deciphers the spatial characteristics of host-gut microbiota interactions in Crohn’s disease. Cell Rep Medicine. 2023;4: 101050. 10.1016/j.xcrm.2023.101050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cleynen I, González JR, Figueroa C, Franke A, McGovern D, Bortlík M, et al. Genetic factors conferring an increased susceptibility to develop Crohn’s disease also influence disease phenotype: results from the IBDchip European Project. Gut. 2013;62:1556–65. 10.1136/gutjnl-2011-300777 [DOI] [PubMed] [Google Scholar]
- 15.Gundra UM, Girgis NM, Ruckerl D, Jenkins S, Ward LN, Kurtz ZD, et al. Alternatively activated macrophages derived from monocytes and tissue macrophages are phenotypically and functionally distinct. Blood. 2014;123:e110–22. 10.1182/blood-2013-08-520619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hampe J, Franke A, Rosenstiel P, Till A, Teuber M, Huse K, et al. A genome-wide association scan of nonsynonymous SNPs identifies a susceptibility variant for Crohn disease in ATG16L1. Nat Genet. 2006;39:207–11. 10.1038/ng1954 [DOI] [PubMed] [Google Scholar]
- 17.Liu Z, Liu R, Gao H, Jung S, Gao X, Sun R, et al. Genetic architecture of the inflammatory bowel diseases across East Asian and European ancestries. Nat Genet. 2023;55:796–806. 10.1038/s41588-023-01384-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vos ACW, Wildenberg ME, Duijvestein M, Verhaar AP, van den Brink GR, Hommes DW. Anti-tumor necrosis factor-α antibodies induce regulatory macrophages in an Fc region-dependent manner. Gastroenterology. 2011;140:221-30.e3. 10.1053/j.gastro.2010.10.008 [DOI] [PubMed] [Google Scholar]
- 19.Schleier L, Wiendl M, Heidbreder K, Binder M-T, Atreya R, Rath T, et al. Non-classical monocyte homing to the gut via α4β7 integrin mediates macrophage-dependent intestinal wound healing. Gut. 2020;69:252–63. 10.1136/gutjnl-2018-316772 [DOI] [PubMed] [Google Scholar]
- 20.Smythies LE, Sellers M, Clements RH, Mosteller-Barnum M, Meng G, Benjamin WH, et al. Human intestinal macrophages display profound inflammatory anergy despite avid phagocytic and bacteriocidal activity. J Clin Investig. 2005;115:66–75. 10.1172/JCI200519229 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gabanyi I, Muller PA, Feighery L, Oliveira TY, Costa-Pinto FA, Mucida D. Neuro-immune interactions drive tissue programming in intestinal macrophages. Cell. 2016;164:378–91. 10.1016/j.cell.2015.12.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Muller PA, Koscso B, Rajani GM, Stevanovic K, Berres ML, Hashimoto D, et al. Crosstalk between muscularis macrophages and enteric neurons regulates gastrointestinal motility. Cell. 2014;158:300–13. 10.1016/j.cell.2014.04.050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hashimoto D, Chow A, Noizat C, Teo P, Beasley MB, Leboeuf M, et al. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity. 2013;38:792–804. 10.1016/j.immuni.2013.04.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Scott CL, Henri S, Guilliams M. Mononuclear phagocytes of the intestine, the skin, and the lung. Immunological Review. 2014;262:9–24. 10.1111/imr.12220 [DOI] [PubMed] [Google Scholar]
- 25.Bain CC, Bravo-Blas A, Scott CL, Gomez Perdiguero E, Geissmann F, Henri S, et al. Constant replenishment from circulating monocytes maintains the macrophage pool in the intestine of adult mice. Nature Immunology. 2014;15:929–37. 10.1038/ni.2967 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yona S, Kim KW, Wolf Y, Mildner A, Varol D, Breker M, et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity. 2013;38:79–91. 10.1016/j.immuni.2012.12.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ginhoux F, Guilliams M. Tissue-resident macrophage ontogeny and homeostasis. Immunity. 2016;44:439–49. 10.1016/j.immuni.2016.02.024 [DOI] [PubMed] [Google Scholar]
- 28.Tamoutounour S, Henri S, Lelouard H, de Bovis B, de Haar C, van der Woude CJ, et al. CD64 distinguishes macrophages from dendritic cells in the gut and reveals the Th1-inducing role of mesenteric lymph node macrophages during colitis. Eur J Immunol. 2012;42:3150–66. 10.1002/eji.201242847 [DOI] [PubMed] [Google Scholar]
- 29.Domanska D, Majid U, Karlsen VT, Merok MA, Beitnes AR, Yaqub S, et al. Single-cell transcriptomic analysis of human colonic macrophages reveals niche-specific subsets. J Exp Med. 2022;219: e20211846. 10.1084/jem.20211846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jakubzick C, Gautier EL, Gibbings SL, Sojka DK, Schlitzer A, Johnson TE, et al. Minimal differentiation of classical monocytes as they survey steady-state tissues and transport antigen to lymph nodes. Immunity. 2013;39:599–610. 10.1016/j.immuni.2013.08.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Schridde A, Bain CC, Mayer JU, Montgomery J, Pollet E, Denecke B, et al. Tissue-specific differentiation of colonic macrophages requires TGFbeta receptor-mediated signaling. Mucosal Immunol. 2017;10:1387–99. 10.1038/mi.2016.142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Paul F, Arkin Y, Giladi A, Jaitin DA, Kenigsberg E, Keren-Shaul H, et al. Transcriptional heterogeneity and lineage commitment in myeloid progenitors. Cell. 2015;163:1663–77. 10.1016/j.cell.2015.11.013 [DOI] [PubMed] [Google Scholar]
- 33.Hoeffel G, Chen J, Lavin Y, Low D, Almeida FF, See P, et al. C-Myb(+) erythro-myeloid progenitor-derived fetal monocytes give rise to adult tissue-resident macrophages. Immunity. 2015;42:665–78. 10.1016/j.immuni.2015.03.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.De Schepper S, Verheijden S, Aguilera-Lizarraga J, Viola MF, Boesmans W, Stakenborg N, et al. Self-maintaining gut macrophages are essential for intestinal homeostasis. Cell. 2018;175:400-15 e13. 10.1016/j.cell.2018.07.048 [DOI] [PubMed] [Google Scholar]
- 35.Shaw TN, Houston SA, Wemyss K, Bridgeman HM, Barbera TA, Zangerle-Murray T, et al. Tissue-resident macrophages in the intestine are long lived and defined by Tim-4 and CD4 expression. J Exp Med. 2018;215:1507–18. 10.1084/jem.20180019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sheng J, Ruedl C, Karjalainen K. Most tissue-resident macrophages except microglia are derived from fetal hematopoietic stem cells. Immunity. 2015;43:382–93. 10.1016/j.immuni.2015.07.016 [DOI] [PubMed] [Google Scholar]
- 37.Schulz C, Perdiguero EG, Chorro L, Szabo-Rogers H, Cagnard N, Kierdorf K, et al. A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science. 2012;336:86–90. 10.1126/science.1219179 [DOI] [PubMed] [Google Scholar]
- 38.Park MD, Silvin A, Ginhoux F, Merad M. Macrophages in health and disease. Cell. 2022;185:4259–79. 10.1016/j.cell.2022.10.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kang B, Alvarado LJ, Kim T, Lehmann ML, Cho H, He J, et al. Commensal microbiota drive the functional diversification of colon macrophages. Mucosal Immunol. 2020;13:216–29. 10.1038/s41385-019-0228-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Vannella KM, Wynn TA. Mechanisms of organ injury and repair by macrophages. Annu Rev Physiol. 2017;79:593–617. 10.1146/annurev-physiol-022516-034356 [DOI] [PubMed] [Google Scholar]
- 41.Zigmond E, Bernshtein B, Friedlander G, Walker CR, Yona S, Kim KW, et al. Macrophage-restricted interleukin-10 receptor deficiency, but not IL-10 deficiency, causes severe spontaneous colitis. Immunity. 2014;40:720–33. 10.1016/j.immuni.2014.03.012 [DOI] [PubMed] [Google Scholar]
- 42.Dai X-M, Zong X-H, Sylvestre V, Stanley ER. Incomplete restoration of colony-stimulating factor 1 (CSF-1) function in CSF-1–deficient Csf1op/Csf1op mice by transgenic expression of cell surface CSF-1. Blood. 2004;103:1114–23. 10.1182/blood-2003-08-2739 [DOI] [PubMed] [Google Scholar]
- 43.Bain CC, Scott CL, Uronen-Hansson H, Gudjonsson S, Jansson O, Grip O, et al. Resident and pro-inflammatory macrophages in the colon represent alternative context-dependent fates of the same Ly6Chi monocyte precursors. Mucosal Immunol. 2013;6:498–510. 10.1038/mi.2012.89 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Smith PD, Smythies LE, Shen R, Greenwell-Wild T, Gliozzi M, Wahl SM. Intestinal macrophages and response to microbial encroachment. Mucosal Immunol. 2011;4:31–42. 10.1038/mi.2010.66 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Weber B, Saurer L, Schenk M, Dickgreber N, Mueller C. CX3CR1 defines functionally distinct intestinal mononuclear phagocyte subsets which maintain their respective functions during homeostatic and inflammatory conditions. Eur J Immunol. 2011;41:773–9. 10.1002/eji.201040965 [DOI] [PubMed] [Google Scholar]
- 46.Fenton TM, Wulff L, Jones GR, Vandamme J, Jørgensen PB, Bain CC, et al. Single-cell characterisation of mononuclear phagocytes in the human intestinal mucosa. bioRxiv. 2021.
- 47.Bujko A, Atlasy N, Landsverk OJB, Richter L, Yaqub S, Horneland R, et al. Transcriptional and functional profiling defines human small intestinal macrophage subsets. J Exp Med. 2018;215:441–58. 10.1084/jem.20170057 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Liu Z, Gu Y, Chakarov S, Bleriot C, Kwok I, Chen X, et al. Fate mapping via Ms4a3-expression history traces monocyte-derived cells. Cell. 2019;178(1509–25): e19. [DOI] [PubMed] [Google Scholar]
- 49.Chen Q, Nair S, Ruedl C. Microbiota regulates the turnover kinetics of gut macrophages in health and inflammation. Life Sci Alliance. 2021;5: e202101178. 10.26508/lsa.202101178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zigmond E, Varol C, Farache J, Elmaliah E, Satpathy AT, Friedlander G, et al. Ly6C hi monocytes in the inflamed colon give rise to proinflammatory effector cells and migratory antigen-presenting cells. Immunity. 2012;37:1076–90. 10.1016/j.immuni.2012.08.026 [DOI] [PubMed] [Google Scholar]
- 51.Serbina NV, Pamer EG. Monocyte emigration from bone marrow during bacterial infection requires signals mediated by chemokine receptor CCR2. Nat Immunol. 2006;7:311–7. 10.1038/ni1309 [DOI] [PubMed] [Google Scholar]
- 52.Hume DA, Freeman TC. Transcriptomic analysis of mononuclear phagocyte differentiation and activation. Immunol Rev. 2014;261:74–84. 10.1111/imr.12211 [DOI] [PubMed] [Google Scholar]
- 53.Dai XM, Ryan GR, Hapel AJ, Dominguez MG, Russell RG, Kapp S, et al. Targeted disruption of the mouse colony-stimulating factor 1 receptor gene results in osteopetrosis, mononuclear phagocyte deficiency, increased primitive progenitor cell frequencies, and reproductive defects. Blood. 2002;99:111–20. 10.1182/blood.V99.1.111 [DOI] [PubMed] [Google Scholar]
- 54.Schlitzer A, McGovern N, Teo P, Zelante T, Atarashi K, Low D, et al. IRF4 transcription factor-dependent CD11b+ dendritic cells in human and mouse control mucosal IL-17 cytokine responses. Immunity. 2013;38:970–83. 10.1016/j.immuni.2013.04.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Jenkins SJ, Ruckerl D, Thomas GD, Hewitson JP, Duncan S, Brombacher F, et al. IL-4 directly signals tissue-resident macrophages to proliferate beyond homeostatic levels controlled by CSF-1. J Exp Med. 2013;210:2477–91. 10.1084/jem.20121999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hume DA, MacDonald KPA. Therapeutic applications of macrophage colony-stimulating factor-1 (CSF-1) and antagonists of CSF-1 receptor (CSF-1R) signaling. Blood. 2012;119:1810–20. 10.1182/blood-2011-09-379214 [DOI] [PubMed] [Google Scholar]
- 57.Wynn TA, Vannella KM. Macrophages in tissue repair, regeneration, and fibrosis. Immunity. 2016;44:450–62. 10.1016/j.immuni.2016.02.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.MacDonald KP, Palmer JS, Cronau S, Seppanen E, Olver S, Raffelt NC, et al. An antibody against the colony-stimulating factor 1 receptor depletes the resident subset of monocytes and tissue- and tumor-associated macrophages but does not inhibit inflammation. Blood. 2010;116:3955–63. 10.1182/blood-2010-02-266296 [DOI] [PubMed] [Google Scholar]
- 59.Barnes MJ, Powrie F. Regulatory T cells reinforce intestinal homeostasis. Immunity. 2009;31:401–11. 10.1016/j.immuni.2009.08.011 [DOI] [PubMed] [Google Scholar]
- 60.Rivollier A, He J, Kole A, Valatas V, Kelsall BL. Inflammation switches the differentiation program of Ly6Chi monocytes from antiinflammatory macrophages to inflammatory dendritic cells in the colon. J Exp Med. 2012;209:139–55. 10.1084/jem.20101387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Nakanishi Y, Sato T, Takahashi K, Ohteki T. IFN-gamma-dependent epigenetic regulation instructs colitogenic monocyte/macrophage lineage differentiation in vivo. Mucosal Immunol. 2018;11:871–80. 10.1038/mi.2017.104 [DOI] [PubMed] [Google Scholar]
- 62.Caer C, Gorreja F, Forsskahl SK, Brynjolfsson SF, Szeponik L, Magnusson MK, et al. TREM-1+ macrophages define a pathogenic cell subset in the intestine of Crohn’s disease patients. J Crohns Colitis. 2021;15:1346–61. 10.1093/ecco-jcc/jjab022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Scott CL, T’Jonck W, Martens L, Todorov H, Sichien D, Soen B, et al. The transcription factor ZEB2 is required to maintain the tissue-specific identities of macrophages. Immunity. 2018;49:312-25 e5. 10.1016/j.immuni.2018.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Grainger JR, Wohlfert EA, Fuss IJ, Bouladoux N, Askenase MH, Legrand F, et al. Inflammatory monocytes regulate pathologic responses to commensals during acute gastrointestinal infection. Nature Medicine. 2013;19:713–21. 10.1038/nm.3189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Hausmann M, Kiessling S, Mestermann S, Webb G, Spottl T, Andus T, et al. Toll-like receptors 2 and 4 are up-regulated during intestinal inflammation. Gastroenterology. 2002;122:1987–2000. 10.1053/gast.2002.33662 [DOI] [PubMed] [Google Scholar]
- 66.West NR, Hegazy AN, Owens BMJ, Bullers SJ, Linggi B, Buonocore S, et al. Oncostatin M drives intestinal inflammation and predicts response to tumor necrosis factor-neutralizing therapy in patients with inflammatory bowel disease. Nat Med. 2017;23:579–89. 10.1038/nm.4307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Murray PJ, Allen JE, Biswas SK, Fisher EA, Gilroy DW, Goerdt S, et al. Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity. 2014;41:14–20. 10.1016/j.immuni.2014.06.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol. 2008;8:958–69. 10.1038/nri2448 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Shapouri-Moghaddam A, Mohammadian S, Vazini H, Taghadosi M, Esmaeili SA, Mardani F, et al. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol. 2018;233:6425–40. 10.1002/jcp.26429 [DOI] [PubMed] [Google Scholar]
- 70.Locati M, Curtale G, Mantovani A. Diversity, mechanisms, and significance of macrophage plasticity. Annu Rev Pathol. 2020;15:123–47. 10.1146/annurev-pathmechdis-012418-012718 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ip WKE, Hoshi N, Shouval DS, Snapper S, Medzhitov R. Anti-inflammatory effect of IL-10 mediated by metabolic reprogramming of macrophages. Science. 2017;356:513–9. 10.1126/science.aal3535 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Lissner D, Schumann M, Batra A, Kredel LI, Kühl AA, Erben U, et al. Monocyte and M1 macrophage-induced barrier defect contributes to chronic intestinal inflammation in IBD. Inflamm Bowel Dis. 2015;21:1297–305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kroner A, Greenhalgh AD, Zarruk JG, Passos dos Santos R, Gaestel M, David S. TNF and increased intracellular iron alter macrophage polarization to a detrimental M1 phenotype in the injured spinal cord. Neuron. 2014;83:1098–116. 10.1016/j.neuron.2014.07.027 [DOI] [PubMed] [Google Scholar]
- 74.Soroosh P, Doherty TA, Duan W, Mehta AK, Choi H, Adams YF, et al. Lung-resident tissue macrophages generate Foxp3+ regulatory T cells and promote airway tolerance. J Exp Med. 2013;210:775–88. 10.1084/jem.20121849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol. 2010;11:889–96. 10.1038/ni.1937 [DOI] [PubMed] [Google Scholar]
- 76.Wynn TA, Ramalingam TR. Mechanisms of fibrosis: therapeutic translation for fibrotic disease. Nat Med. 2012;18:1028–40. 10.1038/nm.2807 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Mauer J, Chaurasia B, Goldau J, Vogt MC, Ruud J, Nguyen KD, et al. Signaling by IL-6 promotes alternative activation of macrophages to limit endotoxemia and obesity-associated resistance to insulin. Nat Immunol. 2014;15:423–30. 10.1038/ni.2865 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Shouval DS, Biswas A, Goettel JA, McCann K, Conaway E, Redhu NS, et al. Interleukin-10 receptor signaling in innate immune cells regulates mucosal immune tolerance and anti-inflammatory macrophage function. Immunity. 2014;40:706–19. 10.1016/j.immuni.2014.03.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Kimball AS, Davis FM, denDekker A, Joshi AD, Schaller MA, Bermick J, et al. The histone methyltransferase Setdb2 modulates macrophage phenotype and uric acid production in diabetic wound repair. Immunity. 2019;51:258-71.e5. 10.1016/j.immuni.2019.06.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wang X, Cao Q, Yu L, Shi H, Xue B, Shi H. Epigenetic regulation of macrophage polarization and inflammation by DNA methylation in obesity. JCI Insight. 2016;1:e87748. 10.1172/jci.insight.87748 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Luan B, Yoon Y-S, Le Lay J, Kaestner KH, Hedrick S, Montminy M. CREB pathway links PGE2 signaling with macrophage polarization. Proc Natl Acad Sci. 2015;112:15642–7. 10.1073/pnas.1519644112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Sanin DE, Matsushita M, Klein Geltink RI, Grzes KM, van Teijlingen Bakker N, Corrado M, et al. Mitochondrial membrane potential regulates nuclear gene expression in macrophages exposed to prostaglandin E2. Immunity. 2018;49:1021-33.e6. 10.1016/j.immuni.2018.10.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zhong Y, Yi C. MicroRNA-720 suppresses M2 macrophage polarization by targeting GATA3. Bioscience Reports. 2016;36:e00363. 10.1042/BSR20160105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zhou H, Xiao J, Wu N, Liu C, Xu J, Liu F, et al. MicroRNA-223 regulates the differentiation and function of intestinal dendritic cells and macrophages by targeting C/EBPβ. Cell Rep. 2015;13:1149–60. 10.1016/j.celrep.2015.09.073 [DOI] [PubMed] [Google Scholar]
- 85.Ying H, Kang Y, Zhang H, Zhao D, Xia J, Lu Z, et al. MiR-127 modulates macrophage polarization and promotes lung inflammation and injury by activating the JNK pathway. J Immunol. 2015;194:1239–51. 10.4049/jimmunol.1402088 [DOI] [PubMed] [Google Scholar]
- 86.Cai X, Yin Y, Li N, Zhu D, Zhang J, Zhang C-Y, et al. Re-polarization of tumor-associated macrophages to pro-inflammatory M1 macrophages by microRNA-155. J Mol Cell Biol. 2012;4:341–3. 10.1093/jmcb/mjs044 [DOI] [PubMed] [Google Scholar]
- 87.Rückerl D, Jenkins SJ, Laqtom NN, Gallagher IJ, Sutherland TE, Duncan S, et al. Induction of IL-4Rα–dependent microRNAs identifies PI3K/Akt signaling as essential for IL-4–driven murine macrophage proliferation in vivo. Blood. 2012;120:2307–16. 10.1182/blood-2012-02-408252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Squadrito ML, Pucci F, Magri L, Moi D, Gilfillan GD, Ranghetti A, et al. miR-511–3p modulates genetic programs of tumor-associated macrophages. Cell Reports. 2012;1:141–54. 10.1016/j.celrep.2011.12.005 [DOI] [PubMed] [Google Scholar]
- 89.Scott CL, Zheng F, De Baetselier P, Martens L, Saeys Y, De Prijck S, et al. Bone marrow-derived monocytes give rise to self-renewing and fully differentiated Kupffer cells. Nat Commun. 2016;7: 10321. 10.1038/ncomms10321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Bernardo D, Marin AC, Fernandez-Tome S, Montalban-Arques A, Carrasco A, Tristan E, et al. Human intestinal pro-inflammatory CD11c(high)CCR2(+)CX3CR1(+) macrophages, but not their tolerogenic CD11c(-)CCR2(-)CX3CR1(-) counterparts, are expanded in inflammatory bowel disease. Mucosal Immunol. 2018;11:1114–26. 10.1038/s41385-018-0030-7 [DOI] [PubMed] [Google Scholar]
- 91.Ortega-Gómez A, Perretti M, Soehnlein O. Resolution of inflammation: an integrated view. EMBO Mol Med. 2013;5:661–74. 10.1002/emmm.201202382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Fullerton JN, Gilroy DW. Resolution of inflammation: a new therapeutic frontier. Nat Rev Drug Discovery. 2016;15:551–67. 10.1038/nrd.2016.39 [DOI] [PubMed] [Google Scholar]
- 93.Hadis U, Wahl B, Schulz O, Hardtke-Wolenski M, Schippers A, Wagner N, et al. Intestinal tolerance requires gut homing and expansion of FoxP3+ regulatory T cells in the lamina propria. Immunity. 2011;34:237–46. 10.1016/j.immuni.2011.01.016 [DOI] [PubMed] [Google Scholar]
- 94.Kim K-W, Vallon-Eberhard A, Zigmond E, Farache J, Shezen E, Shakhar G, et al. In vivo structure/function and expression analysis of the CX3C chemokine fractalkine. Blood. 2011;118:e156–67. 10.1182/blood-2011-04-348946 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Niess JH, Brand S, Gu X, Landsman L, Jung S, McCormick BA, et al. CX3CR1-mediated dendritic cell access to the intestinal lumen and bacterial clearance. Science. 2005;307:254–8. 10.1126/science.1102901 [DOI] [PubMed] [Google Scholar]
- 96.Cummings RJ, Barbet G, Bongers G, Hartmann BM, Gettler K, Muniz L, et al. Different tissue phagocytes sample apoptotic cells to direct distinct homeostasis programs. Nature. 2016;539:565–9. 10.1038/nature20138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Chikina AS, Nadalin F, Maurin M, San-Roman M, Thomas-Bonafos T, Li XV, et al. Macrophages maintain epithelium integrity by limiting fungal product absorption. Cell. 2020;183:411-28.e16. 10.1016/j.cell.2020.08.048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Glocker E-O, Kotlarz D, Boztug K, Gertz EM, Schäffer AA, Noyan F, et al. Inflammatory bowel disease and mutations affecting the interleukin-10 receptor. N Engl J Med. 2009;361:2033–45. 10.1056/NEJMoa0907206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Murai M, Turovskaya O, Kim G, Madan R, Karp CL, Cheroutre H, et al. Interleukin 10 acts on regulatory T cells to maintain expression of the transcription factor Foxp3 and suppressive function in mice with colitis. Nat Immunol. 2009;10:1178–84. 10.1038/ni.1791 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Shaw MH, Kamada N, Kim Y-G, Núñez G. Microbiota-induced IL-1β, but not IL-6, is critical for the development of steady-state TH17 cells in the intestine. J Exp Med. 2012;209:251–8. 10.1084/jem.20111703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Mortha A, Chudnovskiy A, Hashimoto D, Bogunovic M, Spencer SP, Belkaid Y, et al. Microbiota-dependent crosstalk between macrophages and ILC3 promotes intestinal homeostasis. Science. 2014;343: 1249288. 10.1126/science.1249288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Liu JZ, Sommeren SV, Huang H, Ng SC, Alberts R, Takahashi A, et al. Association analyses identify 38 susceptibility loci for inflammatory bowel disease and highlight shared genetic risk across populations. Nat Genet. 2015;47:979–86. 10.1038/ng.3359 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Doster RS, Rogers LM, Gaddy JA, Aronoff DM. Macrophage extracellular traps: a scoping review. J Innate Immun. 2018;10:3–13. 10.1159/000480373 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Okubo K, Kurosawa M, Kamiya M, Urano Y, Suzuki A, Yamamoto K, et al. Macrophage extracellular trap formation promoted by platelet activation is a key mediator of rhabdomyolysis-induced acute kidney injury. Nat Med. 2018;24:232–8. 10.1038/nm.4462 [DOI] [PubMed] [Google Scholar]
- 105.Hartl D. Macrophages and platelets join forces to release kidney-damaging DNA traps. Nat Med. 2018;24:128–9. 10.1038/nm.4486 [DOI] [PubMed] [Google Scholar]
- 106.D’Angelo F, Bernasconi E, Schäfer M, Moyat M, Michetti P, Maillard MH, et al. Macrophages promote epithelial repair through hepatocyte growth factor secretion. Clin Exp Immunol. 2013;174:60–72. 10.1111/cei.12157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Pull SL, Doherty JM, Mills JC, Gordon JI, Stappenbeck TS. Activated macrophages are an adaptive element of the colonic epithelial progenitor niche necessary for regenerative responses to injury. Proc Natl Acad Sci U S A. 2005;102:99–104. 10.1073/pnas.0405979102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Zhu P, Lu T, Wu J, Fan D, Liu B, Zhu X, et al. Gut microbiota drives macrophage-dependent self-renewal of intestinal stem cells via niche enteric serotonergic neurons. Cell Res. 2022;32:555–69. 10.1038/s41422-022-00645-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Ortiz-Masiá D, Cosín-Roger J, Calatayud S, Hernández C, Alós R, Hinojosa J, et al. Hypoxic macrophages impair autophagy in epithelial cells through Wnt1 relevance in IBD. Mucosal Immunol. 2014;7:929–38. 10.1038/mi.2013.108 [DOI] [PubMed] [Google Scholar]
- 110.Cosín-Roger J, Ortiz-Masiá D, Calatayud S, Hernández C, Esplugues JV, Barrachina MD. The activation of Wnt signaling by a STAT6-dependent macrophage phenotype promotes mucosal repair in murine IBD. Mucosal Immunol. 2016;9:986–98. 10.1038/mi.2015.123 [DOI] [PubMed] [Google Scholar]
- 111.Leoni G, Alam A, Neumann P-A, Lambeth JD, Cheng G, McCoy J, et al. Annexin A1, formyl peptide receptor, and NOX1 orchestrate epithelial repair. J Clin Investig. 2012;123:443–54. 10.1172/JCI65831 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.van der Post S, Birchenough GMH, Held JM. NOX1-dependent redox signaling potentiates colonic stem cell proliferation to adapt to the intestinal microbiota by linking EGFR and TLR activation. Cell Rep. 2021;35: 108949. 10.1016/j.celrep.2021.108949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Quiros M, Nishio H, Neumann PA, Siuda D, Brazil JC, Azcutia V, et al. Macrophage-derived IL-10 mediates mucosal repair by epithelial WISP-1 signaling. J Clin Investig. 2017;127:3510–20. 10.1172/JCI90229 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Malvin NP, Seno H, Stappenbeck TS. Colonic epithelial response to injury requires Myd88 signaling in myeloid cells. Mucosal Immunol. 2012;5:194–206. 10.1038/mi.2011.65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Fritsch SD, Sukhbaatar N, Gonzales K, Sahu A, Tran L, Vogel A, et al. Metabolic support by macrophages sustains colonic epithelial homeostasis. Cell Metab. 2023;35:1931-43.e8. 10.1016/j.cmet.2023.09.010 [DOI] [PubMed] [Google Scholar]
- 116.Sehgal A, Donaldson DS, Pridans C, Sauter KA, Hume DA, Mabbott NA. The role of CSF1R-dependent macrophages in control of the intestinal stem-cell niche. Nat Commun. 2018;9:1272–88. 10.1038/s41467-018-03638-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Krishnasamy K, Limbourg A, Kapanadze T, Gamrekelashvili J, Beger C, Häger C, et al. Blood vessel control of macrophage maturation promotes arteriogenesis in ischemia. Nat Commun. 2017;8:952–65. 10.1038/s41467-017-00953-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Heil M, Schaper W. Influence of mechanical, cellular, and molecular factors on collateral artery growth (arteriogenesis). Circ Res. 2004;95:449–58. 10.1161/01.RES.0000141145.78900.44 [DOI] [PubMed] [Google Scholar]
- 119.Viola MF, Chavero-Pieres M, Modave E, Delfini M, Stakenborg N, Estévez MC, et al. Dedicated macrophages organize and maintain the enteric nervous system. Nature. 2023;618:818–26. 10.1038/s41586-023-06200-7 [DOI] [PubMed] [Google Scholar]
- 120.Luo J, Qian A, Oetjen LK, Yu W, Yang P, Feng J, et al. TRPV4 channel signaling in macrophages promotes gastrointestinal motility via direct effects on smooth muscle cells. Immunity. 2018;49:107–19. 10.1016/j.immuni.2018.04.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Murray PJ, Wynn TA. Protective and pathogenic functions of macrophage subsets. Nat Rev Immunol. 2011;11:723–37. 10.1038/nri3073 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Wynn TA. Cellular and molecular mechanisms of fibrosis. J Pathol. 2007;214:199–210. 10.1002/path.2277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Akhurst RJ, Hata A. Targeting the TGFβ signalling pathway in disease. Nat Rev Drug Discovery. 2012;11:790–811. 10.1038/nrd3810 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Scheibe K, Backert I, Wirtz S, Hueber A, Schett G, Vieth M, et al. IL-36R signalling activates intestinal epithelial cells and fibroblasts and promotes mucosal healing in vivo. Gut. 2017;66:823–38. 10.1136/gutjnl-2015-310374 [DOI] [PubMed] [Google Scholar]
- 125.Wilson MS, Elnekave E, Mentink-Kane MM, Hodges MG, Pesce JT, Ramalingam TR, et al. IL-13Rα2 and IL-10 coordinately suppress airway inflammation, airway-hyperreactivity, and fibrosis in mice. J Clin Investig. 2007;117:2941–51. 10.1172/JCI31546 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Kazura JW, Pesce JT, Ramalingam TR, Mentink-Kane MM, Wilson MS, El Kasmi KC, et al. Arginase-1–expressing macrophages suppress Th2 cytokine-driven inflammation and fibrosis. PLoS Pathog. 2009;5:e1000371. 10.1371/journal.ppat.1000371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Mansfield JM, Pesce JT, Ramalingam TR, Wilson MS, Mentink-Kane MM, Thompson RW, et al. Retnla (Relmα/Fizz1) suppresses helminth-induced Th2-type immunity. PLoS Pathog. 2009;5: e1000393. 10.1371/journal.ppat.1000393 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Martin JC, Chang C, Boschetti G, Ungaro R, Giri M, Grout JA, et al. Single-cell analysis of crohn’s disease lesions identifies a pathogenic cellular module associated with resistance to anti-TNF therapy. Cell. 2019;178:1493-508.e20. 10.1016/j.cell.2019.08.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Smillie CS, Biton M, Ordovas-Montanes J, Sullivan KM, Burgin G, Graham DB, et al. Intra- and inter-cellular rewiring of the human colon during ulcerative colitis. Cell. 2019;178:714-30.e22. 10.1016/j.cell.2019.06.029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Garrido-Trigo A, Corraliza AM, Veny M, Dotti I, Melón-Ardanaz E, Rill A, et al. Macrophage and neutrophil heterogeneity at single-cell spatial resolution in human inflammatory bowel disease. Nat Commun. 2023;14:4506. 10.1038/s41467-023-40156-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Mowat AM. Anatomical basis of tolerance and immunity to intestinal antigens. Nat Rev Immunol. 2003;3:331–41. 10.1038/nri1057 [DOI] [PubMed] [Google Scholar]
- 132.Kamada N, Hisamatsu T, Okamoto S, Chinen H, Kobayashi T, Sato T, et al. Unique CD14+ intestinal macrophages contribute to the pathogenesis of Crohn disease via IL-23/IFN-γ axis. J Clin Investig. 2008;118:2269–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Seo S-U, Kamada N, Muñoz-Planillo R, Kim Y-G, Kim D, Koizumi Y, et al. Distinct commensals induce interleukin-1β via NLRP3 inflammasome in inflammatory monocytes to promote intestinal inflammation in response to injury. Immunity. 2015;42:744–55. 10.1016/j.immuni.2015.03.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Rugtveit J, Haraldsen G, Høgåsen AK, Bakka A, Brandtzaeg P, Scott H. Respiratory burst of intestinal macrophages in inflammatory bowel disease is mainly caused by CD14+L1+ monocyte derived cells. Gut. 1995;37:367–73. 10.1136/gut.37.3.367 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Castro-Dopico T, Fleming A, Dennison TW, Ferdinand JR, Harcourt K, Stewart BJ, et al. GM-CSF calibrates macrophage defense and wound healing programs during intestinal infection and inflammation. Cell Rep. 2020;32: 107857. 10.1016/j.celrep.2020.107857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Smith AM, Rahman FZ, Hayee B, Graham SJ, Marks DJ, Sewell GW, et al. Disordered macrophage cytokine secretion underlies impaired acute inflammation and bacterial clearance in Crohn’s disease. J Exp Med. 2009;206:1883–97. 10.1084/jem.20091233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Schwerd T, Pandey S, Yang H-T, Bagola K, Jameson E, Jung J, et al. Impaired antibacterial autophagy links granulomatous intestinal inflammation in Niemann-Pick disease type C1 and XIAP deficiency with NOD2 variants in Crohn’s disease. Gut. 2017;66:1060–73. 10.1136/gutjnl-2015-310382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Ginhoux F, Bleriot C, Lecuit M. Dying for a cause: regulated necrosis of tissue-resident macrophages upon infection. Trends Immunol. 2017;38:693–5. 10.1016/j.it.2017.05.009 [DOI] [PubMed] [Google Scholar]
- 139.Soncin I, Sheng J, Chen Q, Foo S, Duan K, Lum J, et al. The tumour microenvironment creates a niche for the self-renewal of tumour-promoting macrophages in colon adenoma. Nat Commun. 2018;9:582. 10.1038/s41467-018-02834-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Pearson C, Thornton EE, McKenzie B, Schaupp AL, Huskens N, Griseri T, et al. ILC3 GM-CSF production and mobilisation orchestrate acute intestinal inflammation. eLife. 2016;5:e10066. 10.7554/eLife.10066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Nakanishi Y, Sato T, Ohteki T. Commensal Gram-positive bacteria initiates colitis by inducing monocyte/macrophage mobilization. Mucosal Immunol. 2015;8:152–60. 10.1038/mi.2014.53 [DOI] [PubMed] [Google Scholar]
- 142.Desalegn G, Pabst O. Inflammation triggers immediate rather than progressive changes in monocyte differentiation in the small intestine. Nat Commun. 2019;10:3229. 10.1038/s41467-019-11148-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Leoni G, Neumann PA, Sumagin R, Denning TL, Nusrat A. Wound repair: role of immune–epithelial interactions. Mucosal Immunol. 2015;8:959–68. 10.1038/mi.2015.63 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Wynn TA, Chawla A, Pollard JW. Macrophage biology in development, homeostasis and disease. Nature. 2013;496:445–55. 10.1038/nature12034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Madala SK, Pesce JT, Ramalingam TR, Wilson MS, Minnicozzi S, Cheever AW, et al. Matrix metalloproteinase 12-deficiency augments extracellular matrix degrading metalloproteinases and attenuates IL-13–dependent fibrosis. J Immunol. 2010;184:3955–63. 10.4049/jimmunol.0903008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Pellicoro A, Aucott RL, Ramachandran P, Robson AJ, Fallowfield JA, Snowdon VK, et al. Elastin accumulation is regulated at the level of degradation by macrophage metalloelastase (MMP-12) during experimental liver fibrosis. Hepatology. 2012;55:1965–75. 10.1002/hep.25567 [DOI] [PubMed] [Google Scholar]
- 147.Wang S, Hirschberg R. BMP7 antagonizes TGF-beta -dependent fibrogenesis in mesangial cells. Am J Physiol Renal Physiol. 2003;284:F1006-1013. 10.1152/ajprenal.00382.2002 [DOI] [PubMed] [Google Scholar]
- 148.Chen Y, Pu Q, Ma Y, Zhang H, Ye T, Zhao C, et al. Aging reprograms the hematopoietic-vascular niche to impede regeneration and promote fibrosis. Cell Metab. 2021;33:395-410.e4. 10.1016/j.cmet.2020.11.019 [DOI] [PubMed] [Google Scholar]
- 149.Shimokado K, Raines EW, Madtes DK, Barrett TB, Benditt EP, Ross R. A significant part of macrophage-derived growth factor consists of at least two forms of PDGF. Cell. 1985;43:277–86. 10.1016/0092-8674(85)90033-9 [DOI] [PubMed] [Google Scholar]
- 150.Grassl GA, Valdez Y, Bergstrom KS, Vallance BA, Finlay BB. Chronic enteric salmonella infection in mice leads to severe and persistent intestinal fibrosis. Gastroenterology. 2008;134:768–80. 10.1053/j.gastro.2007.12.043 [DOI] [PubMed] [Google Scholar]
- 151.Scheibe K, Kersten C, Schmied A, Vieth M, Primbs T, Carlé B, et al. Inhibiting interleukin 36 receptor signaling reduces fibrosis in mice with chronic intestinal inflammation. Gastroenterology. 2019;156:1082-97.e11. 10.1053/j.gastro.2018.11.029 [DOI] [PubMed] [Google Scholar]
- 152.Nakata T, Li C, Mayassi T, Lin H, Ghosh K, Segerstolpe Å, et al. Genetic vulnerability to Crohn’s disease reveals a spatially resolved epithelial restitution program. Sci Transl Med. 2023;15:eadg5252. 10.1126/scitranslmed.adg5252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Coffey JC, O’Leary DP. The mesentery: structure, function, and role in disease. The Lancet Gastroenterology & Hepatology. 2016;1:238–47. 10.1016/S2468-1253(16)30026-7 [DOI] [PubMed] [Google Scholar]
- 154.Coffey JC, Byrnes KG, Walsh DJ, Cunningham RM. Update on the mesentery: structure, function, and role in disease. The Lancet Gastroenterology & Hepatology. 2022;7:96–106. 10.1016/S2468-1253(21)00179-5 [DOI] [PubMed] [Google Scholar]
- 155.Lee J-Y, Hall JA, Kroehling L, Wu L, Najar T, Nguyen HH, et al. Serum amyloid A proteins induce pathogenic Th17 cells and promote inflammatory disease. Cell. 2020;180:79-91.e16. 10.1016/j.cell.2019.11.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Dowling JK, Afzal R, Gearing LJ, Cervantes-Silva MP, Annett S, Davis GM, et al. Mitochondrial arginase-2 is essential for IL-10 metabolic reprogramming of inflammatory macrophages. Nat Commun. 2021;12:1460. 10.1038/s41467-021-21617-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Rioux JD, Xavier RJ, Taylor KD, Silverberg MS, Goyette P, Huett A, et al. Genome-wide association study identifies new susceptibility loci for Crohn disease and implicates autophagy in disease pathogenesis. Nat Genet. 2007;39:596–604. 10.1038/ng2032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.de Lange KM, Moutsianas L, Lee JC, Lamb CA, Luo Y, Kennedy NA, et al. Genome-wide association study implicates immune activation of multiple integrin genes in inflammatory bowel disease. Nat Genet. 2017;49:256–61. 10.1038/ng.3760 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Rieder F, Fiocchi C, Rogler G. Mechanisms, management, and treatment of fibrosis in patients with inflammatory bowel diseases. Gastroenterology. 2017;152:340-50.e6. 10.1053/j.gastro.2016.09.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Lees CW, Barrett JC, Parkes M, Satsangi J. New IBD genetics: common pathways with other diseases. Gut. 2011;60:1739–53. 10.1136/gut.2009.199679 [DOI] [PubMed] [Google Scholar]
- 161.Ogura Y, Bonen DK, Inohara N, Nicolae DL, Chen FF, Ramos R, et al. A frameshift mutation in NOD2 associated with susceptibility to Crohn’s disease. Nature. 2001;411:603–6. 10.1038/35079114 [DOI] [PubMed] [Google Scholar]
- 162.Hugot JP, Chamaillard M, Zouali H, Lesage S, Cézard JP, Belaiche J, et al. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn’s disease. Nature. 2001;411:599–603. 10.1038/35079107 [DOI] [PubMed] [Google Scholar]
- 163.Hampe J, Grebe J, Nikolaus S, Solberg C, Croucher PJP, Mascheretti S, et al. Association of NOD2 (CARD 15) genotype with clinical course of Crohn’s disease: a cohort study. Lancet. 2002;359:1661–5. 10.1016/S0140-6736(02)08590-2 [DOI] [PubMed] [Google Scholar]
- 164.Watanabe T, Kitani A, Murray PJ, Strober W. NOD2 is a negative regulator of Toll-like receptor 2–mediated T helper type 1 responses. Nat Immunol. 2004;5:800–8. 10.1038/ni1092 [DOI] [PubMed] [Google Scholar]
- 165.Vermeire S, Pierik M, Hlavaty T, Claessens G, van Schuerbeeck N, Joossens S, et al. Association of organic cation transporter risk haplotype with perianal penetrating Crohn’s disease but not with susceptibility to IBD. Gastroenterology. 2005;129:1845–53. 10.1053/j.gastro.2005.10.006 [DOI] [PubMed] [Google Scholar]
- 166.Marks DJB, Harbord MWN, MacAllister R, Rahman FZ, Young J, Al-Lazikani B, et al. Defective acute inflammation in Crohn’s disease: a clinical investigation. Lancet. 2006;367:668–78. 10.1016/S0140-6736(06)68265-2 [DOI] [PubMed] [Google Scholar]
- 167.Fowler EV, Doecke J, Simms LA, Zhao ZZ, Webb PM, Hayward NK, et al. ATG16L1 T300A shows strong associations with disease subgroups in a large Australian IBD population: further support for significant disease heterogeneity. Am J Gastroenterol. 2008;103:2519–26. 10.1111/j.1572-0241.2008.02023.x [DOI] [PubMed] [Google Scholar]
- 168.Cooney R, Baker J, Brain O, Danis B, Pichulik T, Allan P, et al. NOD2 stimulation induces autophagy in dendritic cells influencing bacterial handling and antigen presentation. Nat Med. 2009;16:90–7. 10.1038/nm.2069 [DOI] [PubMed] [Google Scholar]
- 169.Travassos LH, Carneiro LAM, Ramjeet M, Hussey S, Kim Y-G, Magalhães JG, et al. Nod1 and Nod2 direct autophagy by recruiting ATG16L1 to the plasma membrane at the site of bacterial entry. Nat Immunol. 2009;11:55–62. 10.1038/ni.1823 [DOI] [PubMed] [Google Scholar]
- 170.Leonardi I, Li X, Semon A, Li D, Doron I, Putzel G, et al. CX3CR1+ mononuclear phagocytes control immunity to intestinal fungi. Science. 2018;359:232–6. 10.1126/science.aao1503 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Brand S. Crohn’s disease: Th1, Th17 or both? The change of a paradigm: new immunological and genetic insights implicate Th17 cells in the pathogenesis of Crohn’s disease. Gut. 2009;58:1152–67. 10.1136/gut.2008.163667 [DOI] [PubMed] [Google Scholar]
- 172.Duerr RH, Taylor KD, Brant SR, Rioux JD, Silverberg MS, Daly MJ, et al. A genome-wide association study dentifies IL23R as an inflammatory bowel disease gene. Science. 2006;314:1461–3. 10.1126/science.1135245 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Barrett JC, Hansoul S, Nicolae DL, Cho JH, Duerr RH, Rioux JD, et al. Genome-wide association defines more than 30 distinct susceptibility loci for Crohn’s disease. Nat Genet. 2008;40:955–62. 10.1038/ng.175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Shichita T, Sugiyama Y, Ooboshi H, Sugimori H, Nakagawa R, Takada I, et al. Pivotal role of cerebral interleukin-17–producing γδT cells in the delayed phase of ischemic brain injury. Nat Med. 2009;15:946–50. 10.1038/nm.1999 [DOI] [PubMed] [Google Scholar]
- 175.Fidler TP, Xue C, Yalcinkaya M, Hardaway B, Abramowicz S, Xiao T, et al. The AIM2 inflammasome exacerbates atherosclerosis in clonal haematopoiesis. Nature. 2021;592:296–301. 10.1038/s41586-021-03341-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Spalinger MR, Sayoc-Becerra A, Ordookhanian C, Canale V, Santos AN, King SJ, et al. The JAK inhibitor tofacitinib rescues intestinal barrier defects caused by disrupted epithelial-macrophage interactions. J Crohns Colitis. 2021;15:471–84. 10.1093/ecco-jcc/jjaa182 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Parkes M, Barrett JC, Prescott NJ, Tremelling M, Anderson CA, Fisher SA, et al. Sequence variants in the autophagy gene IRGM and multiple other replicating loci contribute to Crohn’s disease susceptibility. Nat Genet. 2007;39:830–2. 10.1038/ng2061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Long SA, Cerosaletti K, Wan JY, Ho JC, Tatum M, Wei S, et al. An autoimmune-associated variant in PTPN2 reveals an impairment of IL-2R signaling in CD4+ T cells. Genes Immun. 2010;12:116–25. 10.1038/gene.2010.54 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Ma G, Pan PY, Eisenstein S, Divino CM, Lowell CA, Takai T, et al. Paired immunoglobin-like receptor-B regulates the suppressive function and fate of myeloid-derived suppressor cells. Immunity. 2011;34:385–95. 10.1016/j.immuni.2011.02.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Pesce J. The IL-21 receptor augments Th2 effector function and alternative macrophage activation. J Clin Investig. 2006;116:2044–55. 10.1172/JCI27727 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Liu W, Yang H-S, Zheng S-Y, Weng J-H, Luo H-H, Lei Y-Y, et al. Thymic epithelial tumors: examining the GTF2I mutation and developing a novel prognostic signature with LncRNA pairs to predict tumor recurrence. BMC Genomics. 2022;23:656. 10.1186/s12864-022-08880-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Sánchez-Martín L, Estecha A, Samaniego R, Sánchez-Ramón S, Vega MÁ, Sánchez-Mateos P. The chemokine CXCL12 regulates monocyte-macrophage differentiation and RUNX3 expression. Blood. 2011;117:88–97. 10.1182/blood-2009-12-258186 [DOI] [PubMed] [Google Scholar]
- 183.Ehrchen J, Steinmüller L, Barczyk K, Tenbrock K, Nacken W, Eisenacher M, et al. Glucocorticoids induce differentiation of a specifically activated, anti-inflammatory subtype of human monocytes. Blood. 2006;109:1256–74. [DOI] [PubMed] [Google Scholar]
- 184.Oakley RH, Cidlowski JA. The biology of the glucocorticoid receptor: new signaling mechanisms in health and disease. Journal of Allergy and Clinical Immunology. 2013;132:1033–44. 10.1016/j.jaci.2013.09.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Bantel H, Berg C, Vieth M, Stolte M, Kruis W, Schulze-Osthoff K. Mesalazine inhibits activation of transcription factor NF-kappaB in inflamed mucosa of patients with ulcerative colitis. Am J Gastroenterol. 2000;95:3452–7. 10.1111/j.1572-0241.2000.03360.x [DOI] [PubMed] [Google Scholar]
- 186.Municio C, Soler Palacios B, Estrada-Capetillo L, Benguria A, Dopazo A, García-Lorenzo E, et al. Methotrexate selectively targets human proinflammatory macrophages through a thymidylate synthase/p53 axis. Annals of the Rheumatic Diseases. 2016;75:2157–65. 10.1136/annrheumdis-2015-208736 [DOI] [PubMed] [Google Scholar]
- 187.Marinković G, Hamers AA, de Vries CJ, de Waard V. 6-Mercaptopurine reduces macrophage activation and gut epithelium proliferation through inhibition of GTPase Rac1. Inflamm Bowel Dis. 2004;20:1487–95. 10.1097/MIB.0000000000000122 [DOI] [PubMed] [Google Scholar]
- 188.Vos ACW, Wildenberg ME, Arijs I, Duijvestein M, Verhaar AP, de Hertogh G, et al. Regulatory macrophages induced by infliximab are involved in healing in vivo and in vitro. Inflamm Bowel Dis. 2012;18:401–8. 10.1002/ibd.21818 [DOI] [PubMed] [Google Scholar]
- 189.Dige A, Magnusson MK, Öhman L, Hvas CL, Kelsen J, Wick MJ, et al. Reduced numbers of mucosal DRintmacrophages and increased numbers of CD103+dendritic cells during anti-TNF-α treatment in patients with Crohn’s disease. Scand J Gastroenterol. 2016;51:692–9. 10.3109/00365521.2015.1134649 [DOI] [PubMed] [Google Scholar]
- 190.De Vries LCS, Duarte JM, De Krijger M, Welting O, Van Hamersveld PHP, Van Leeuwen-Hilbers FWM, et al. A JAK1 selective kinase inhibitor and tofacitinib affect macrophage activation and function. Inflamm Bowel Dis. 2019;25:647–60. 10.1093/ibd/izy364 [DOI] [PubMed] [Google Scholar]
- 191.Tian L, Chen F, Macosko EZ. The expanding vistas of spatial transcriptomics. Nat Biotechnol. 2022;41:773–82. 10.1038/s41587-022-01448-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Chen J, Suo S, Tam PPL, Han J-DJ, Peng G, Jing N. Spatial transcriptomic analysis of cryosectioned tissue samples with Geo-seq. Nat Protoc. 2017;12:566–80. 10.1038/nprot.2017.003 [DOI] [PubMed] [Google Scholar]
- 193.Ståhl PL, Salmén F, Vickovic S, Lundmark A, Navarro JF, Magnusson J, et al. Visualization and analysis of gene expression in tissue sections by spatial transcriptomics. Science. 2016;353:78–82. 10.1126/science.aaf2403 [DOI] [PubMed] [Google Scholar]
- 194.Cho C-S, Xi J, Si Y, Park S-R, Hsu J-E, Kim M, et al. Microscopic examination of spatial transcriptome using Seq-Scope. Cell. 2021;184:3559-72.e22. 10.1016/j.cell.2021.05.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Merritt CR, Ong GT, Church SE, Barker K, Danaher P, Geiss G, et al. Multiplex digital spatial profiling of proteins and RNA in fixed tissue. Nat Biotechnol. 2020;38:586–99. 10.1038/s41587-020-0472-9 [DOI] [PubMed] [Google Scholar]
- 196.Cabrita R, Lauss M, Sanna A, Donia M, Skaarup Larsen M, Mitra S, et al. Tertiary lymphoid structures improve immunotherapy and survival in melanoma. Nature. 2020;577:561–5. 10.1038/s41586-019-1914-8 [DOI] [PubMed] [Google Scholar]
- 197.Wang X, Allen WE, Wright MA, Sylwestrak EL, Samusik N, Vesuna S, et al. Three-dimensional intact-tissue sequencing of single-cell transcriptional states. Science. 2018;361: eaat5691. 10.1126/science.aat5691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Chen A, Liao S, Cheng M, Ma K, Wu L, Lai Y, et al. Spatiotemporal transcriptomic atlas of mouse organogenesis using DNA nanoball-patterned arrays. Cell. 2022;185:1777-92.e21. 10.1016/j.cell.2022.04.003 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
No datasets were generated or analysed during the current study.