

Abstract
Background
Gene transfer studies in primary T cells have suffered from the limitations of conventional viral transduction or transfection techniques. Replication-defective adenoviral vectors are an attractive alternative for gene delivery. However, naive lymphocytes are not readily susceptible to infection with adenoviruses due to insufficient expression of the coxsackie/adenovirus receptor.
Results
To render T cells susceptible to adenoviral gene transfer, we have developed three new murine transgenic lines in which expression of the human coxsackie/adenovirus receptor (hCAR) with a truncated cytoplasmic domain (hCARΔcyt) is limited to thymocytes and lymphocytes under direction of a human CD2 mini-gene. hCARΔcyt.CD2 transgenic mice were crossed with DO11.10 T cell receptor transgenic mice (DO11.hCARΔcyt) to allow developmental studies in a defined, clonal T cell population. Expression of hCARΔcyt enabled adenoviral transduction of resting primary CD4+ T cells, differentiated effector T cells and thymocytes from DO11.hCARΔcyt with high efficiency. Expression of hCARΔcyt transgene did not perturb T cell development in these mice and adenoviral transduction of DO11.hCARΔcyt T cells did not alter their activation status, functional responses or differentiative potential. Adoptive transfer of the transduced T cells into normal recipients did not modify their physiologic localization.
Conclusion
The DO11.hCARΔcyt transgenic model thus allows efficient gene transfer in primary T cell populations and will be valuable for novel studies of T cell activation and differentiation.
Background
Gene transfer into developing and mature primary T cells has been limited by the lack of efficient techniques for ex vivo or in vivo transfection or transduction. The adaptation of retroviral vectors for gene delivery into lymphocytes has gained favor recently [1,2], but has several features that limit its use. First, retroviral gene transduction requires cell cycle progression of the cellular host to permit viral genome integration, limiting studies of regulatory genes that are operative early in T cell activation or differentiation. Second, because the integration site of the viral genome is variable, there can be local, host chromatin effects on exogenous gene expression and unpredictable silencing of viral integrants. Third, efficiencies of transduction with retroviral vectors are often limiting. Finally, the retroviral genome is small, limiting the size of exogenous genes that can be packaged.
Replication-defective adenoviral vectors offer several advantages that make them attractive for gene delivery [3-6]. Adenoviral vectors are able to transduce proliferating or quiescent cells; they can accommodate dual, large insert sizes and can deliver additional genes by superinfection; and stable, high-titer viral stocks are easily generated [7]. Further, since the adenoviral genome is maintained extra-chromosomally, insertion site-dependent effects of the host genome are minimized, and exogenous genes can be reliably expressed at high levels. Unfortunately, naive lymphocytes are not susceptible to infection with adenoviruses due to lack of detectable expression of a viral receptor, such as the coxsackie/adenovirus receptor (CAR), which is required for efficient viral attachment [8-10]. The CAR protein contains two extra-cellular immunoglobulin-like (Ig) domains, the first of which appears sufficient for binding to the adenoviral fiber knob protein [11,12]. Expression of either human or murine CAR in various cell lines has been shown to facilitate gene transfer by recombinant human adenoviruses [8-10,13]. Stable transfection of transformed murine B and T cells lines with human CAR (hCAR) lacking its cytoplasmic domain rendered them susceptible to adenoviral transfection, establishing that lymphoid cells can be permissive hosts for adenoviral transduction and demonstrating that the cytoplasmic domain of CAR is not required for viral entry [13]. More recently, mice transgenic for full length [14] or truncated hCAR [15] have been described in which adenoviral transduction of primary lymphocytes has been demonstrated.
We describe here three new transgenic (Tg) mouse lines in which expression of human CAR with a truncated cytoplasmic domain (hCARΔcyt) is limited to thymocytes and lymphocytes under direction of a human CD2 mini-gene [16]. The three lines are similar in having transgene expression restricted to the lymphoid lineage, but differ in the pattern of hCARΔcyt expression profiles within lymphoid populations. In each transgenic line, expression of the hCARΔcyt transgene imparted susceptibility to efficient adenoviral transduction that was independent of cellular activation. By crossing the hCARΔcyt.CD2 Tg mice with DO11.10 TCR Tg mice, we have generated an antigen-specific model with which to study the effects of adenovirally delivered genes on T cell activation and differentiation. Transduction of hCARΔcyt-positive naïve, Th1 or Th2 DO11.10 T cells with a recombinant adenoviral vector resulted in high-level reporter gene expression without detectable activation of transduced cells or deleterious effect on their antigenic responses.
Results
hCARΔcyt.CD2 transgenic mice: characterization of distinct hCAR expression profiles in three different founder lines
To develop a transgenic model with high-level expression of hCAR on murine T cells, a transgene was constructed based on a human CD2 mini-gene described previously [16] (Fig. 1A). This mini-gene includes the 5' promoter region of the hCD2 gene as well as the hCD2 LCR, which is reported to direct transgene expression to T cells in a position-independent, copy number-dependent manner [17,18]. Since the function of CAR is undefined, and deletion of the cytoplasmic tail of hCAR does not adversely affect adenoviral transduction [13,19], a cDNA encoding a cytoplasmic tail-truncated variant of hCAR (hCARΔcyt) was generated and inserted into the CD2 mini-gene to eliminate any potential signaling capacity of hCAR. Of eight founder mice bearing an intact transgene, all demonstrated lymphoid lineage-specific expression of hCARΔcyt, with three distinct expression profiles represented by founder lines 6, 13, and 18 (Figure 1B and data not shown). To develop an antigen-specific T cell system with which to deliver genes of interest, each of these lines was then crossed with DO11.10 TCR Tg mice [20] to generate the DO11.hCARΔcyt double transgenic lines shown (Figure 1B &1C).
Figure 1.

hCARΔcyt transgene map and expression in lymph node cells and thymocytes of hCAR transgenic mice.A. An 807 bp fragment (hatched box) encoding the extracellular and transmembrane region with a truncated cytoplasmic tail (TM-RKKKRR*) of the human CAR was cloned into the EcoRI/SmaI site of VA hCD2. The hCARΔcyt coding region was inserted between intron 1 of hCD2 and the poly A1 site from the hCD2 3' untranslated region. All potential ATG start codons in exon 1 of hCD2 have been mutated [16]. B. Lymphoid cells were isolated from three different transgenic lines (6, 13 and 18) and stained with RmcB anti-human CAR mAb and anti-mouse CD4, CD8 or CD19 before analysis on a FACSscan. The quadrants were set based on the staining of a hCARΔcyt negative littermate control (data not shown). The graphs represent two-color dot plots of lymph node cells gated on viable lymphoid gate; the percentage of cells in each quadrant is indicated. C. Two-color dot plots of thymocytes stained with anti-CD4 and anti-CD8 mAbs. The staining with the anti-hCAR RmcB mAb is represented as a histogram for each corresponding quadrant with the percentage of positive cells.
Lymph node cells were isolated from mice in DO11.hCARΔcyt lines 6, 13 and 18, stained with antibodies to CD4, CD8, CD19 and RmcB (anti-hCAR) and analyzed by flow cytometry (Fig. 1B). All of the CD4+ and CD8+ T cells from line 6 and 18 expressed high levels of the hCARΔcyt transgene, while CD4+ and CD8+T cells from line 13 had a bimodal pattern of expression, with approximately 35% of both CD4+ and CD8+ negative for hCARΔcyt. While both lines 13 and 18 had negligible expression in the B cell compartment, the vast majority of B cells from line 6 expressed the hCARΔcyt transgene, although at a lower level than the T cells (mean fluorescence intensity (MFI): CD19+ B cells, 64; CD4+ T cells, 193). hCARΔcyt was not detected in non-lymphoid tissues nor on lymphocytes from non-transgenic littermate controls (data not shown).
To determine at what developmental stage the hCARΔcyt transgene is expressed in T cells, thymocytes were isolated from 4-week-old mice and the expression of hCARΔcyt relative to the expression of CD4 and CD8 was evaluated (Fig. 1C). Consistent with the expression of hCARΔcyt on mature CD4+ and CD8+ T cells from lines 6 and 18, all of the thymocytes in the CD4+CD8+ double positive (DP) and CD4+ or CD8+ single positive (SP) fractions were positive for hCARΔcyt. In line 13, approximately 68% of the DP and SP cells expressed hCARΔcyt, consistent with the expression of hCARΔcyt on T cells in the periphery. A lower percentage of hCARΔcyt positive cells was observed in the CD4-CD8- double negative fraction in all lines studied, which likely reflects the heterogeneous nature of the double negative population. The skewed pattern of thymocyte development toward CD4+ single positive in each of the lines was similar to that observed in DO11.10.hCARΔcyt- control littermates (data not shown). Thus, in the three distinct lines studied, the CD2 promoter/LCR/enhancer directed the expression of the hCARΔcyt transgene to the T cell and in one of the lines, the B cell lineage. Line 18 was chosen for further study as the recipient of adenoviral-mediated gene delivery due to its uniform, T cell-specific expression of the hCARΔcyt transgene.
Primary T cells and thymocytes from DO11.hCARΔcyt mice are efficiently transduced by adenoviral vectors
To determine whether lymphocytes from hCARΔcyt.CD2 transgenic mice were susceptible to adenoviral transduction, lymph node cells were isolated from line DO11.hCARΔcyt.18 and incubated with adenoviral vectors designed to express GFP under the control of the CMV5 or ubiquitin (UbP) promoters (Fig. 2A). Seventy percent of resting CD4+ cells that were transduced with the Ad5.UbP.GFP expressed the GFP reporter without activation. In contrast, only 36% of resting CD4+ cells transduced with the Ad5.CMV5.GFP were GFP+, and the mean fluorescence intensity was lower, suggesting an inability of the CMV5 promoter to induce strong GFP expression in resting T cells. Following activation with PMA and ionomycin, CMV5-driven expression of GFP increased to 81% of the CD4+ T cells. A small percentage of CD4- cells, representing the hCARΔcyt+ CD8+ fraction, were transduced with both the UbP and CMV5 viruses as well.
Figure 2.

Transduction with Ad5.UbP.GFP and Ad5.CMV5.GFP of lymph node cells and thymocytes from line 18 mice. Lymph node cells and thymocytes from line 18 mice were isolated and transduced with Ad5.UbP.GFP or Ad5.CMV5.GFP adenoviral reporter vectors at a MOI of 10 for 30' and incubated for 24 h at 37°C/5%CO2. Half of the cells transduced with Ad5.CMV5.GFP were stimulated for 4 h with PMA/ionomycin before analysis. Lymph node cells were then stained with anti-CD4, anti-CD25 and anti-CD62L; thymocytes were stained with anti-CD4 and anti-CD8. The quadrants were set based on the transduction of hCARΔcyt negative littermate controls (data not shown). A. Two color dot-plots of lymph nodes cells transduced with Ad5.Ubp.GFP or Ad5.CMV5.GFP with or without activation with PMA/ionomycin. The top panel represent the expression of hCARΔcyt and CD4 detected in the viable lymphoid gate while the two bottom panel were gated on CD4 positive cells only (rectangular gate). The percentage of cells in each quadrant is indicated. B. Two-color dot plot of CD4 and CD8 expression of line 18 thymocytes transduced with Ad5.UbP.GFP. The histograms represent the GFP expression for each thymocyte subset defined by their CD4 and CD8 expression. The percentage of GFP positive cells is indicated.
To determine whether adenoviral transduction changed their activation status, resting CD4+ T cells were examined for expression of the early activation markers CD62L (L-selectin) and CD25 (IL-2Rα). After transduction with Ad5.UbP.GFP or Ad5.CMV5.GFP without PMA/ionomycin, the GFP positive and the GFP negative fractions of CD4+ cells were both CD62L high and CD25 low, confirming that the cells were initially of a resting phenotype and that adenoviral transduction did not activate them. Upon activation with PMA/ionomycin, the CD4+ T cells down-regulated CD62L and up-regulated CD25 (Fig. 2A). Control T cells from hCARΔcyt transgene-negative littermates had indistinguishable flow cytometric profile (data not shown).
Because the expression of hCARΔcyt was detected early in thymocyte development (Fig. 1C), the ability of these cells to be transduced by adenovirus was also determined. Thymocytes from line 18 were incubated with Ad5.UbP.GFP as stated above, and were evaluated by flow cytometry for their expression of CD4, CD8, and GFP (Fig. 2B). Seventy-five percent of the CD4 SP cells were transduced for GFP expression, consistent with the transduction frequency of CD4+ T cells in the periphery. Approximately half of the CD8 SP could be transduced as well. Interestingly, whereas a high percentage of the DN thymocytes expressed GFP after transduction (75%), the DP fraction was relatively refractory to transduction even though all of the DP cells expressed hCARΔcyt; less than 10% of the transduced DP cells were GFP-positive irrespective of the MOI tested (Fig. 2B and data not shown).
Adenoviral transduction of Th1 and Th2 lines derived from DO11.hCARΔcyt transgenic mice
To examine the susceptibility of effector T cell populations to adenoviral transduction, Th1 and Th2 cells were derived from DO11.hCARΔcyt.18 mice by in vitro culture under polarizing cytokine conditions [21]. The resulting Th1 and Th2 lines were incubated with Ad5.UbP.GFP or Ad5.CMV5.GFP, with or without activation with PMA/ionomycin, and expression of GFP was measured by flow cytometry 24 hours later (Fig. 3 and Table 1). Similar to naïve T cells, resting Th1 and Th2 cells were readily transduced by Ad5.UbP.GFP, and PMA/ionomycin activation had little effect on either the frequency or expression levels of GFP positive cells. In contrast, both the frequencies and levels of GFP expression were increased in Ad5.CMV5.GFP-transduced Th1 and Th2 lines following activation with PMA/ionomycin, anti-CD3 or OVA peptide. Further, the activation-induced expression of GFP by cells transduced with the Ad5.CMV5 vector yielded significantly better expression levels than those induced by the Ad5.UbP vector. Thus, both Th1 and Th2 effectors derived from DO11.hCARΔcyt mice are competent hosts for adenoviral transduction, and were equivalent to or better than naïve DO11.hCARΔcyt cells with respect to their transduction efficiencies and/or reporter expression levels.
Figure 3.

Transduction with Ad5.UbP.GFP and Ad5.CMV5.GFP of Th1 and Th2 lines derived from line 18 mice. Th1 and Th2 lines were derived from line 18 DO11.hCARΔcyt double transgenic mice by antigenic stimulation in polarizing conditions for 3 weeks. One week after the last antigenic stimulation, the resting Th1 and Th2 lines were transduced for 30' with Ad5.UbP.GFP or Ad5.CMV5.GFP at a MOI of 4 and incubated for 24 h at 37°C/5%CO2. Half of the cells transduced with Ad5.CMV5.GFP were stimulated for 4 h with PMA/ionomycin before analysis. The histograms represent the GFP expression of the Th1 (left panels) and Th2 (right panels) lines transduced with the indicated adenovirus with or without stimulation with PMA/ionomycin. The percentage of positive cells and mean fluorescence intensity are indicated in Table 1.
Table 1.
Transduction of Th1 and Th2 lines with Ad5.CMV5.GFP and Ad5.UbP.GFP
| Stimulation* | ||||
| T cell Line/Ad Vector | None | Anti-CD3 | OVA peptide | PMA/Ionomycin |
| Th1+Ad5.UbP.GFP | 51% (117)a | NDb | ND | 54% (139) |
| Th1+Ad5.CMV5.GFP | 20% (112) | 27% (212) | 45% (529) | 73% (1122) |
| Th2+Ad5.UbP.GFP | 60% (212) | ND | ND | 70% (269) |
| Th2+Ad5.CMV5.GFP | 43% (242) | 66% (1274) | 70% (753) | 75% (1421) |
*Th1 and Th2 lines derived from line 18 DO11.10/hCARΔcyt transgenic mice were transduced and GFP expression was analyzed as described in Fig. 4. The lines were activated by incubation on anti-CD3 coated plates (10 μg/ml), addition of irradiated BALB/c splenocytes and 5 μg/ml of OVA peptide or 50 ng/ml PMA and 5 μM ionomycin for 24 h at 37°C/5% CO2. aPercentage of GFP positive cells (and mean fluorescence intensity of the GFP positive population) are indicated. bND – not determined.
Adenoviral transduction does not alter the functional responses or differentiative potential of primary T cells
Although adenoviral transduction did not induce detectable activation of resting, naïve CD4+ T cells (Figure 2), we wished to further determine whether transduction altered the activation-induced functions of this population. The proliferative responses and potential for Th1 and Th2 phenotype development were therefore examined following adenoviral transduction. CD4+ T cells isolated from DO11.hCARΔcyt.18 mice were transduced with Ad5.UbP.GFP, and were sorted after 24 hours based on their expression of GFP. The sorted cells were cultured with antigen presenting cells and increasing concentrations of OVA peptide for 3 days and then pulsed with [3H]thymidine for 24 hours. Figure 4A shows that the GFP positive population of Ad-transduced cells had a comparable proliferative response to the non-transduced GFP negative cells across the entire antigen dose range. Thus, adenoviral transduction did not significantly affect primary CD4 T cell proliferation.
Figure 4.

Transduction of CD4+ T cells from line 18 mice does not affect proliferation or cytokine production. CD4+ T cells were isolated from spleen and lymph nodes of line DO11.hCARΔcyt.18 double transgenic mice, transduced for 30' with Ad5.UbP.GFP at a MOI of 10 and incubated for 24 h at 37°C/5%CO2. The GFP positive and GFP negative population were then sorted on a FACSvantage cell sorter before assay. A. Proliferative responses of the GFP- and GFP+ population stimulated for 72 h with increasing amount of OVA peptide in the presence of irradiated BALB/c splenocytes. 50 μCi/ml [3H]thymidine was added to the cultures for 48 h before harvesting the cells and count of the incorporated radioactivity. The curves represent the dose-dependant proliferative responses (as indicated by the [3H]thymidine incorporation in CPM) of the GFP+ (closed circles) and the GFP- (open diamond) populations. B. Cytokine production of the transduced CD4+ cells. The GFP+ and GFP- sorted population were stimulated with irradiated BALB/c splenocytes and 5 μg/ml ovalbumin peptide in conditions driving them toward a Th1 or a Th2 phenotype. One week later, the culture supernatant were collected and analyzed by ELISA for the production of IFN-γ, IL-4 and IL-10. The histogram indicates the concentration of the different cytokines produced by the indicated populations. The relative concentrations were calculated using serial dilution of a known standard for each cytokine as a reference curve.
Transduced, sorted GFP+ and GFP- CD4 T cells were cultured under Th1-/Th2-polarizing conditions and examined for cytokine production following re-stimulation to determine potential effects of adenoviral transduction on effector phenotype development (Figure 4B). Following one week of polarization, Th1 and Th2 cells were tested by ELISA for their ability to produce cytokines indicative of the two different T cell subsets (Figure 4B). Th1- and Th2-polarized cells from both the transduced (GFP+) and non-transduced (GFP-) populations produced comparable amounts of Th1-specific (IFN-γ) or Th2-specific (IL-4 and IL-10) cytokines, respectively. Cytokine production by Th1- and Th2-polarized DO11.10 T cells from hCARΔcyt-negative littermate controls was not significantly different (data not shown). Thus, neither hCAR expression nor adenoviral transduction alters the naïve T cell's potential for Th1 or Th2 phenotype development.
In vivo trafficking of Ad-transduced CD4+ T cells
To further investigate the effects of adenoviral transduction on T cell biology, and to examine the possibility that hCAR expression (an Ig superfamily member of undetermined function) might alter the in vivo distribution of transduced T cells, we examined the tissue distribution of DO11.hCARΔcyt T cells following adoptive transfer into BALB/c recipients. For these experiments, we used line 13 DO11.hCARΔcyt Tg mice that have a bimodal distribution of hCAR expression to provide an internal control, "sham transfection" population. CD4+ cells were isolated from DO11.hCARΔcyt.13 lymph and spleen cells and transduced with Ad5.UbP.GFP. Twenty-four hours later, transduced T cells were recovered and examined for GFP expression by flow cytometry prior to transfer of 5 × 106 cells into each recipient mouse. In a representative experiment shown in Figure 5, 45% percent of the pre-transfer CD4+ cells expressed the clonotypic marker KJ-126 and of these cells, 47.5% had been transduced for GFP expression (Fig. 5A, top panel).
Figure 5.
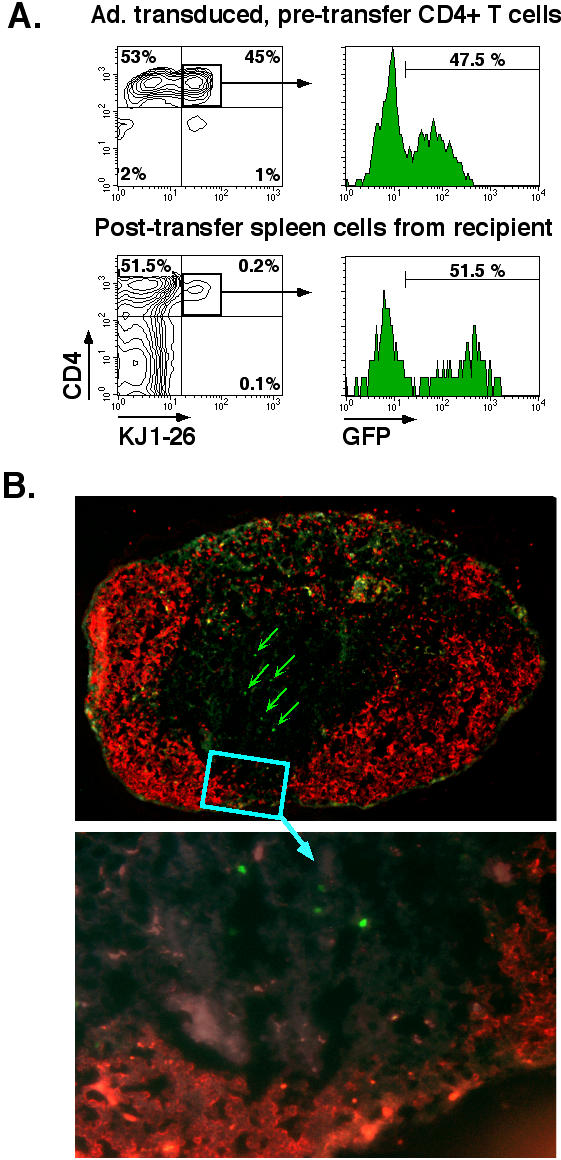
Adoptive transfer of CD4+ T cells from line 18 mice in BALB/c recipient. CD4+ T cells were isolated from spleen and lymph nodes of DO11.hCARΔcyt line 13 transgenic mice, transduced for 30' with Ad5.UbP.GFP at a MOI of 10 and incubated for 24 h at 37°C/5%CO2. 5 × 106 transduced cells were injected in the tail vein of BALB/c mice. The mice were sacrificed after 48 h and the inguinal, axillary, submandibular and mesenteric lymph nodes, the spleen and the Peyer's Patches were isolated. A. Flow cytometric analysis of GFP expression of transduced CD4+ cells before and after adoptive transfer. Recovered, pooled lymph node cells from BALB/c recipients were stained with anti-CD4 and anti-DO11.10 TCR clonotypic mAb, KJ-126, and analyzed on a FACScan. The panels depict two-color dot plots of the transduced CD4+ cells before transfer (top panel) and the pooled lymph node cells from a representative BALB/c recipient (lower panel) and histograms representing the GFP expression for each of the CD4+KJ126+ population (square gate). The percentage of positive cells is indicated. B. Immunohistochemical analysis of a peripheral lymph node of Ad5.UbP.GFP-transduced CD4 T cell adoptive transfer recipient. An inguinal lymph node was processed and stained with anti-B22O (red) as described in Material and Methods. The red fluorescent areas define primary B cell follicles. GFP positive cells (green) are indicated (arrows) in parafollicular, T cell areas of a representative lymph node (magnification: top, 10×; bottom, 40×).
Forty-eight hours after I.V. injection of the transduced (DO11.hCARΔcyt.13) or control (DO11.10) cells, recipient mice were sacrificed and secondary lymphoid tissues (lymph nodes, spleen and Peyer's patches) were divided for detection of the transferred KJ1-26+ and GFP+ T cells by flow cytometry (Fig. 5A, lower panel) or immuno-histochemistry (Fig. 5B). Flow cytometric analysis determined that the transferred population of Ad-transduced cells from DO11.hCARΔcyt Tg donors retained the pre-transfer frequencies of GFP+ and GFP- cells within the DO11.10 T cell (CD4+KJ1-26+) fraction and were equivalent to control DO11.10 cells with respect to the relative distribution between secondary lymphoid organs (Fig. 5A &5B; data not shown). Thus, 51.5% of the recovered KJ1-26 positive cells isolated from pooled lymph nodes expressed GFP, compared to a pre-transfer frequency of 47.5%. Similar results were obtained in recovered splenic tissues, and at 72 and 96 hours post-transfer (data not shown). There was an increase in the level of GFP reporter expression by Ad-transduced cells in the interval between transfer and recovery (pre-transfer MFI: 75; post-transfer MFI: 320), suggesting continued GFP transcription in vivo. Thus, there was no preferential loss or increase of the Ad-transduced cells in vivo, nor were there gross differences in trafficking to secondary lymphoid tissues.
To determine if adenoviral transduction or GFP reporter expression altered T cell trafficking to tissue microenvironments, recovered lymphoid tissues were sectioned and processed for fluorescent microscopic analysis (Figure 5B and Table 2). A double-label immunofluorescent protocol was developed to permit in situ localization of GFP+ or KJ1-26+ T cells and B220+ B cells in fixed, frozen tissue sections (Figure 5B, and data not shown). Irrespective of the population of T cells transferred (DO11.10 wild-type control or Ad5.UbC.GFP-transduced DO11.hCARΔcyt), a similar distribution of KJ1-26+ and GFP+ T cells was found within the secondary lymphoid tissue microenvironments. In the spleen, KJ1-26+ or GFP+ cells were primarily located within the periarterial lymphoid sheaths (PALS), with fewer positive cells in the marginal zones, and only scattered positive cells in the red pulp (Table 2). Relatively few positive cells were detected within the B cell follicles, although positive cells could be found in the PALS, at the border with B cell follicles. Similarly, in lymph nodes and Peyer's patches, the wild-type and Ad-transduced transgenic T cells localized primarily to paracortical T cell areas. Collectively, these data indicate that neither hCARΔcyt or GFP expression nor adenoviral transduction affects survival or trafficking of adoptively transferred cells for at least 4 days in vivo.
Table 2.
Anatomic Distribution of Ad5.UbP.GFP-transduced DO11.hCARΔcyt versus Control DO11.10 T Cell Populations following Adoptive Transfer
| % of Total KJ1-26+ Cells in Each Lymphoid Tissue Microenvironmenta | |||
| Tissue/Microenvironent | DO11.10 | GFP-positive DO11.hCARΔcyt | |
| Spleen | |||
| PALS | 92b | 91 | |
| Primary Nodules/Follicles | 3 | 5 | |
| Marginal Zone | 4 | 2 | |
| Red Pulp | 1 | 3 | |
| Inguinal LN | |||
| Diffuse Cortex/Paracortex | 98 | 97 | |
| Primary Nodules/Follicles | 2 | 3 | |
| Medullary Cords | 0 | 0 | |
| Mesenteric LN | |||
| Diffuse Cortex/Paracortex | 97 | 96 | |
| Primary Nodules/Follicles | 3 | 4 | |
| Medullary Cords | 0 | 0 | |
| Peyer's Patch | |||
| Paranodular zones (T cell) | 99 | 98 | |
| Nodules (B cell) | 1 | 2 | |
a Two days following transfer of 5 × 106 DO11.10 or Ad5.UbP.GFP-transduced DO11.hCARΔcyt CD4+ T cells into BALB/c recipients, animals were sacrificed and the recovered lymphoid tissues were processed for microscopic analysis as described in Material and Methods. b Numbers are the fraction of the total cell population in each tissue. Definition of the lymphoid tissue microenvironments follows the nomenclature of Weiss. Data are the mean from 3–4 animals per group (standard deviations <16%).
Discussion
The introduction of transgenic systems adapted to take advantage of adenoviral gene delivery technology offers an attractive adjunct to existing gene delivery strategies for studies of lymphocyte biology. In an effort to facilitate gene transfer into primary lymphocytes, we and others [14,15] have employed different strategies to engineer transgenic mice that express the human adenoviral receptor, hCAR, on murine lymphocytes. Here we report the characterization of three new transgenic lines that express a cytoplasmic-tail truncated hCAR under control of hCD2 gene regulatory elements, and demonstrate their utility for gene transfer studies of primary T cells and thymocytes. In agreement with previous studies using human CD2 mini-gene expression cassette [16-18], the truncated hCAR transgene was expressed in all cells of the T cell lineage, and except for transgenic line 6 (hCARΔcyt.CD2.6), in which there was expression on mature B cells as well as T cells, hCAR was not expressed by non-T cells. Importantly, the expression of hCARΔcyt by T cells was stable and high-level at all stages of development, from double-negative thymocytes to Th1 and Th2 effectors. With the possible exception of double positive thymocytes (see below), reporter expression by transduced hCARΔcyt-positive cells at each of these developmental stages was very efficient. Thus, it is possible to target adenoviral transduction to the T cell compartment in vitro and in vivo. Of particular note, the level of expression of hCARΔcyt was not significantly lower for double-negative and double-positive thymocyte populations in line hCARΔcyt.CD2.18 mice, contrary to what was observed previously [15]. The high-level expression of hCARΔcyt by double-negative (CD4-CD8-) thymocytes rendered this population readily transducible, suggesting that these early thymocytes express sufficient integrin co-receptors (e.g. αvβ3 or αvβ5) for adenoviral entry and making this a useful model for early thymocyte gene targeting
Despite expression of levels of hCARΔcyt comparable to that of double-negative and single-positive thymocytes, double-positive thymocytes were relatively refractory to reporter expression following transduction with either the Ad5.UbP.GFP or Ad5.CMV5.GFP vector. The lack of reporter expression could reflect poor entry of adenovirus in this population or poor transcriptional activity of the promoters used (UbP and CMV5), among other possibilites. Given the comparable levels of hCARΔcyt expression by double-positive and double-negative or single positive thymocytes, it is unlikely that viral entry is limiting in these cells, although this has not been formally tested. We favor the possibility that the poor reporter expression reflects inefficient transcriptional activity of these promoters at this developmental stage. In this same vein, expression of the GFP reporter by freshly isolated mature T cells transduced with the Ad5.UbP.GFP vector was very efficient, while the Ad5.CMV5.GFP vector required a post-transduction activation of T cells to induce optimal expression of GFP [14,15]. On the contrary, the CMV5 promoter yielded better reporter expression in activated, transduced Th1 and Th2 cells. Thus, the choice of promoter will need to be optimized for each adenoviral vector depending on the developmental stage and specific purpose for its use. Studies with other promoter elements (e.g. CD2) are in progress and should be informative.
Interestingly, the bimodal pattern of transgene expression by T cells and thymocytes from line hCARΔcyt.CD2.13 suggested some positional effect variegation (PEV) of the transgene, an effect not previously reported in trangenes that include the entire CD2 LCR [17,18]. Further, we have noticed that the percentage of hCARΔcyt positive T cells in the line hCARΔcyt.CD2.13 increased with successive generations of back-cross of the original hCARΔcyt founder (B6xSJL F1) onto a BALB/c background, reflecting a strain background effect of the PEV (data not shown). Although the mechanism of this expression heterogeneity is not clear, it is fortuitous in providing different lines that offer distinct targeting profiles. Accordingly, it should prove possible to target both B and T cells in line hCARΔcyt.CD2.6 mice, whereas line hCARΔcyt.CD2.13 mice can be exploited to perform transduction studies in which there is an hCARΔcyt negative population to serve as an internal control (Figure 5).
An important consideration at the outset of these studies concerned the possibility that ectopic hCAR expression or adenoviral transduction might affect the normal development and/or function of T cells. For this reason, we chose to use a truncated variant of hCAR to preclude possible signaling by this molecule [13]. As evidenced by the functional data reported herein, and those from previous reports [14,15], there does not appear to be an adverse effect of either hCARΔcyt expression or adenoviral transduction on T cell activation, development or function. The antigen-induced proliferative responses of naïve DO11.hCARΔcyt T cells were comparable to those of non-hCAR DO11.10 littermates, and the differentiative potential of transduced naïve T cells into Th1 or Th2 effectors was not compromised. While at higher multiplicities of adenoviral infection (MOIs) than those required for efficient transduction there were deleterious effects on naïve T cell viability (data not shown), at the MOIs used we saw no decrease in cell viability or function. Further, adenoviral transduction of resting, naïve T cells exerted no detectable intrinsic activation of these cells, suggesting that adenoviral delivery of genes of interest to non-cycling T cells will permit loss or gain of function studies prior to antigen- or mitogen-driven stimulation.
Ad-transduced DO11.hCARΔcyt trafficked normally to secondary lymphoid organs following adoptive transfer, indicating that neither hCARΔcyt nor adenoviral transduction altered normal in vivo migration patterns. This affords the opportunity to examine the in vivo responses of transduced, naïve or effector T cells following ex vivo gene transfer. Limitations in previous studies of in vivo adenoviral transduction have been the poor stability of adenoviral transduction in dividing cells and the induction of cytotoxic immune responses to adenoviral gene products. We observed no decrement in GFP expression by adoptively transferred cells as long as 4 days following transfer (data not shown), but have not yet examined later time points. Wan et al. reported accelerated loss of Ad-transduced cells relative to controls via an immune-mediated mechanism [15] suggesting limitations for long-term adoptive transfer studies into immune-competent recipients. It is unclear at present how immune responses to Ad-transduced T cells might affect their in vivo function, but the ability to examine early in vivo responses following adoptive transfers for even 3–4 days should prove valuable, and transfers into immune-deficient recipients should facilitate long-term in vivo studies. Further, continued advances in the development of "gutless" adenoviral vectors that avoid expression of cytotoxic antigenic targets may alleviate immune clearance problems [22-24].
Conclusions
The hCARΔcyt.CD2 transgenic lines reported here offer unique properties that will be advantageous for many T cell transduction studies. These models offer several advantages over existing approaches for short-term gene delivery into primary T cells, principal of which is the capacity for high-efficiency transduction of T cells in G0 or G1 of cell cycle. The introduction of the hCARΔcyt.CD2 transgene into TCR transgenic lines, such as the DO11.10 model reported here, will provide a powerful system for T cell developmental studies. Due to their high immunogenicity and their extra-chromosomal replication, the current adenoviral vectors may not be appropriate for long-term transduction in-vivo but improvement in vector design and adoptive transfer into immunodeficient hosts may facilitate experiments in the future. The development of newer recombinant adenoviruses containing elements for epichromosomal replication may alleviate limitations that prevent more stable in vivo expression in dividing lymphoid cells derived from hCAR transgenic mice.
Materials and methods
Mice
DO11.10 TCR transgenic mice specific for ovalbumin peptide residues 323–339 (OVA323-339) and restricted by I-Ad[20] were backcrossed onto BALB/cJ background (>16 generations). Mice were screened for TCR transgene expression by two-color flow cytometric analysis of peripheral blood lymphocytes stained with anti-CD4 and the anti-DO11.10 TCR mAb, KJ1-26 [25]. hCARΔcyt transgenic mice were crossed with BALB/c DO11.10 TCR Tg mice for at least 5 generations and used at 4–6 weeks of age. BALB/cJ mice were purchased from the Jackson Laboratory (Bar Harbor, ME) and were used at 6–10 weeks of age. All mice were maintained in a specific pathogen-free facility at the University of Alabama at Birmingham and treated according to the Animal Care Guidelines adopted by this institution.
Antibodies and reagents
The RmcB (anti-hCAR) (ATCC, [26]) and the KJ1-26 monoclonal antibodies (anti-DO11.10 TCR) were purified from ascites and conjugated with fluorescein isothiocyanate (FITC), biotin or phycoerythrin (PE) by Dr. Roger Lallone (Brookwood Biomedical; Birmingham, AL). Purified anti-CD3 (17A2), anti-IL-4 (11B11 and BVD6-24G2), anti-IL-10 (JES5-2A5 and JES5-16E3), anti-IL-12 (C17.8) and anti-IFN-γ, PE-conjugated anti-CD4 (GK1.5), biotin-conjugated anti-CD19 (1D3), anti-CD45R/B220 (RA3-6B2), anti-CD8 (53-6.72), anti-CD25 (7D4) and anti-CD62L/L-selectin (MEL-14), Cy-Chrome and PE-labeled streptavidin were purchased from PharMingen (San Diego, CA). Recombinant murine IL-4 and IL-12 were purchased from R&D Systems Inc. (Minneapolis, MN). Phorbol 12-myristate 13-acetate (PMA) and ionomycin were purchased from Sigma (St. Louis, MO).
Generation of hCARΔcyt transgenic mice
DNA encoding the extracellular and transmembrane domain of the hCAR was obtained by PCR amplification of a human CAR cDNA [9] kindly provided by Dr. Jeffrey Bergelson, (Children's Hospital of Philadelphia, Philadelphia, PA). The forward primer, 5'ACTGACGAATTCAGCCACCATGGCGCTCCTGCTGTGC3' introduced an EcoRI site and the reverse primer, 5'GTCAGTCCCGGGTCAGTCAGCTACTTTTTACGACAGCAAAAGAT3' introduced stop codons in three reading frames and a SmaI site. The resulting fragment (hCARΔcyt) was restriction endonuclease digested and subcloned into the EcoRI/SmaI site of the VA human CD2 mini-gene containing the human CD2 promoter and 3' LCR [16]. Automated sequencing (Applied Biosystems, Foster City, CA; University of Alabama at Birmingham Department of Microbiology sequencing facility) of the entire construct confirmed its accuracy. A KpnI/XbaI digest liberated the CD2/hCARΔcyt transgene from the vector backbone and the transgene was subsequently microinjected into fertilized oocytes by the UAB ES/Transgenic facility.
Adenoviruses
Ad5.CMV5.GFP adenovirus, with an improved cytomegalovirus promoter (CMV5) expressing enhanced green fluorescent protein (GFP), was purchased from Quantum Biotechnology (Qbiogen, Carlsbad, CA). The Ad5.UbP.GFP expressing an enhanced GFP under the control of a human ubiquitin promoter (UbP) [27] was kindly provided by Dr. J. DeGregori (University of Colorado Health Science Center, Denver, CO). After large-scale production, adenoviruses were purified by double cesium chloride gradient [7], dialyzed in 10% glycerol/10 mM HEPES/0.5 mM MgCl2 and stored in viral preservation medium (25% glycerol/5 mM Tris pH8/50 mM NaCl/0.05% BSA) until use. Viral titers were determined by OD measurement and limiting dilution analysis of GFP expression by transduced 293 cells following Quantum's AdEasy TCID50 protocol.
Flow cytometric analysis
Splenocytes or lymph node cells were stained for flow cytometry with the PE- or biotin-conjugated monoclonal antibodies indicated in the appropriate figure legends as previously described [28]. Cells stained with biotinylated primary antibody were detected with streptavidin-Cy-Chrome (Pharmingen). GFP expression was detected in the FL1 channel. For analytical flow cytometry, 10,000 events with forward and side scatter properties of lymphocytes were collected on a FACScan flow cytometer and analyzed using CellQuest® software (Becton Dickinson; San Diego, CA).
Primary and secondary cell cultures
CD4+ T cells were isolated from spleen and lymph nodes from DO11.10 TCR × hCARΔcyt double transgenic mice by positive sorting using anti-CD4 magnetic beads (Dynal AS, Oslo, Norway). Greater than 95% of the resulting cells were CD4 positive and were plated at a ratio of 1:5 with irradiated BALB/c splenocytes and 5 μg/ml ovalbumin peptide 323–339. The addition of 50 U/ml IL-12 and 10 μg/ml anti-IL-4 was used to generate cells with a Th1 phenotype, while 1000 U/ml IL-4 and 10 μg/ml anti-IL-12 was used to generate cells with a Th2 phenotype [29]. After the initial isolation of the naïve CD4+ T cells, cultures were treated with the above cytokine/antibody cocktail weekly for three consecutive weeks to ensure complete polarization.
Adenoviral transduction
For adenoviral transduction, lymph nodes and spleen cells were isolated, treated with ACK lysing buffer for 3 min and washed in DMEM/10 mM HEPES (pH 7.2). Cells (50 × 106/ml) were incubated with adenovirus at a multiplicity of infection (MOI) of 10 in DMEM/10 mM HEPES at 37°C under 5% CO2 for 30 min with gentle mixing every 10 min. The cells were then washed with PBS and cultured in RP10 (RPMI 1640(GIBCO/BRL)/10% FBS (HyClone)/0.1 mM 2-mercaptoethanol/1% penicillin-streptomycin) at 37°C/5% CO2 until analysis. For some experiments, the transduced cells were then activated with 5 μM ionomycin and 50 ng/ml PMA for 4 h or stimulated for 18 h by immobilized anti-CD3 (pre-coated at 10 μg/ml in PBS) or 5 μg/ml OVA peptide with antigen-presenting cells before analysis.
Cytokines and proliferation assays
CD4+ T cells from DO11.10 TCR × hCARΔcyt double transgenic mice (DO11.hCARΔcyt) were isolated and transduced as described above. After 24 h, the GFP positive and GFP negative fractions of the transduced lymphocytes were sorted using a Becton Dickinson FACSvantage and cultured under Th1 or Th2 polarizing conditions as described above. For cytokines assays, 1 × 106 transduced cells were incubated on anti-CD3 coated plates for 72 h at 37°C/5% CO2. Culture supernatants were then harvested and an ELISA assay was performed to measure the production of IL-4, IL-10 and IFN-γ as previously described [30]. To assay proliferation, the transduced cells were plated in duplicate in microtiter plates (5 × 105/well) at a ratio of 1:5 with irradiated BALB/c splenocytes and the indicated doses of OVA peptide at 37°C/5% CO2. After 72 h, 50 μCi/ml [3H]thymidine (Amersham, Piscataway, NJ) was added to the well. Cells were further incubated for 24 h and harvested. The incorporation of [3H]thymidine was measured using a LKB-Wallac (Gaithersburg, MD) scintillation counter.
Adoptive transfers
CD4+ T cells from hCARΔcyt transgenic mice were isolated and transduced as described above. After 24 h in culture, 5 × 106 cells were washed in DMEM, resuspended in 100 μl of DMEM and injected in the tail vein of 6 weeks old BALB/c mice. The mice were sacrificed after 48 h and the inguinal, axillary, submandibular and mesenteric lymph nodes, the spleen and the Peyer's patches were isolated. Half of the spleen and lymph nodes were used for flow cytometric analysis. The remaining tissues were fixed for 18 h in 4% formalin in PBS, then washed in PBS and frozen embedded in OCT (Sakura Finetek, Torrance, CA). Frozen sections of the spleen, lymph nodes and Peyer's patches were incubated with 10% normal horse serum in PBS, counterstained with biotin-labeled B220 antibody followed by PE-labeled streptavidin, analyzed using a Nikon E800 fluorescent microscope with a SPOT imaging system (Diagnostic instruments, Sterling Heights, MI) and counted as previously described [28].
Statistical analyses
Data from experimental and control groups were evaluated by the Student t-test. Differences were considered significant when p < 0.05.
List of abbreviations
Ad Adenovirus
hCAR human coxsackie/adenovirus receptor
MFI mean fluorescence intensity
MOI multiplicity of infection
OVA ovalbumin
PEV positional effect variegation
Tg transgenic
Authors' contributions
V.H. carried out the characterization of the transgenic mice, the analysis of the virally transduced cells (flow cytometric analysis, cytokine and proliferation assays, adoptive transfers) and drafted the manuscript. R.D.-H. conceived and carried out the generation of the transgenic mice (construct, sequencing), the primary and secondary cell cultures (production of Th lines) and corrected the manuscript. J.O. carried out the breeding of the transgenic mice and their genotyping. R.J.M. participated in the design of the study and corrected the manuscript. C.T.W. conceived the study, performed the microscopic and statistical analysis of the transferred cells and finalized the manuscript.
Acknowledgments
Acknowledgments
The authors thank Dr. Roger Lallone for antibody purifications and conjugations, Dr. Carl Pinkert for his expert assistance in generation of the hCARΔcyt.CD2 transgenic mice, Enid Keyser for flow cytometric cell sorting, Drs. Jeffrey Bergelson, Dimitris Kioussis and James DeGregori for provision of the hCAR cDNA, CD2 mini-gene and the Ad5.UbP.GFP adenovirus, respectively. We also thank Noelle Nicholls, Joyce Pike, Henrietta Turner and Fred Wagner for their precious help. This work was supported by grants AI 35783, AR 43272 and DK44240 from the National Institutes of Health (C.T.W.), a grant from Sankyo Co., LTD (C.T.W.), and a Research Career Development Fellowship Grant from The Wellcome Trust (R.J.M.).
Contributor Information
Vincent Hurez, Email: vhurez@uab.edu.
Robin Dzialo-Hatton, Email: rdzialo@path.uab.edu.
James Oliver, Email: joliver@path.uab.edu.
R James Matthews, Email: MatthewsRJ@Cardiff.ac.uk.
Casey T Weaver, Email: cweaver@path.uab.edu.
References
- Nolan GP, Shatzman AR. Expression vectors and delivery systems. Curr Opin Biotechnol. 1998;9:447–50. doi: 10.1016/S0958-1669(98)80027-X. [DOI] [PubMed] [Google Scholar]
- Costa GL, Benson JM, Seroogy CM, Achacoso P, Fathman CG, Nolan GP. Targeting rare populations of murine antigen-specific T lymphocytes by retroviral transduction for potential application in gene therapy for autoimmune disease. J Immunol. 2000;164:3581–90. doi: 10.4049/jimmunol.164.7.3581. [DOI] [PubMed] [Google Scholar]
- Berkner KL. Development of adenovirus vectors for the expression of heterologous genes. Biotechniques. 1988;6:616–29. [PubMed] [Google Scholar]
- Haddada H, Cordier L, Perricaudet M. Gene therapy using adenovirus vectors. Curr Top Microbiol Immunol. 1995;199:297–306. doi: 10.1007/978-3-642-79586-2_14. [DOI] [PubMed] [Google Scholar]
- Chartier C, Degryse E, Gantzer M, Dieterle A, Pavirani A, Mehtali M. Efficient generation of recombinant adenovirus vectors by homologous recombination in Escherichia coli. J Virol. 1996;70:4805–10. doi: 10.1128/jvi.70.7.4805-4810.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He TC, Zhou S, da Costa LT, Yu J, Kinzler KW, Vogelstein B. A simplified system for generating recombinant adenoviruses. Proc Natl Acad Sci U S A. 1998;95:2509–14. doi: 10.1073/pnas.95.5.2509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevins JR, DeGregori J, Jakoi L, Leone G. Functional analysis of E2F transcription factor. Methods Enzymol. 1997;283:205–19. doi: 10.1016/s0076-6879(97)83017-0. [DOI] [PubMed] [Google Scholar]
- Tomko RP, Xu R, Philipson L. HCAR and MCAR: the human and mouse cellular receptors for subgroup C adenoviruses and group B coxsackieviruses. Proc Natl Acad Sci U S A. 1997;94:3352–6. doi: 10.1073/pnas.94.7.3352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergelson JM, Cunningham JA, Droguett G, Kurt-Jones EA, Krithivas A, Hong JS, Horwitz MS, Crowell RL, Finberg RW. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science. 1997;275:1320–3. doi: 10.1126/science.275.5304.1320. [DOI] [PubMed] [Google Scholar]
- Bergelson JM, Krithivas A, Celi L, Droguett G, Horwitz MS, Wickham T, Crowell RL, Finberg RW. The murine CAR homolog is a receptor for coxsackie B viruses and adenoviruses. J Virol. 1998;72:415–9. doi: 10.1128/jvi.72.1.415-419.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freimuth P, Springer K, Berard C, Hainfeld J, Bewley M, Flanagan J. Coxsackievirus and adenovirus receptor amino-terminal immunoglobulin V-related domain binds adenovirus type 2 and fiber knob from adenovirus type 12. J Virol. 1999;73:1392–8. doi: 10.1128/jvi.73.2.1392-1398.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bewley MC, Springer K, Zhang YB, Freimuth P, Flanagan JM. Structural analysis of the mechanism of adenovirus binding to its human cellular receptor, CAR. Science. 1999;286:1579–83. doi: 10.1126/science.286.5444.1579. [DOI] [PubMed] [Google Scholar]
- Leon RP, Hedlund T, Meech SJ, Li S, Schaack J, Hunger SP, Duke RC, DeGregori J. Adenoviral-mediated gene transfer in lymphocytes. Proc Natl Acad Sci U S A. 1998;95:13159–64. doi: 10.1073/pnas.95.22.13159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt MR, Piekos B, Cabatingan MS, Woodland RT. Expression of a human coxsackie/adenovirus receptor transgene permits adenovirus infection of primary lymphocytes. J Immunol. 2000;165:4112–9. doi: 10.4049/jimmunol.165.7.4112. [DOI] [PubMed] [Google Scholar]
- Wan YY, Leon RP, Marks R, Cham CM, Schaack J, Gajewski TF, DeGregori J. Transgenic expression of the coxsackie/adenovirus receptor enables adenoviral-mediated gene delivery in naive T cells. Proc Natl Acad Sci U S A. 2000;97:13784–13789. doi: 10.1073/pnas.250356297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhumabekov T, Corbella P, Tolaini M, Kioussis D. Improved version of a human CD2 minigene based vector for T cell-specific expression in transgenic mice. J Immunol Methods. 1995;185:133–40. doi: 10.1016/0022-1759(95)00124-S. [DOI] [PubMed] [Google Scholar]
- Kioussis D, Festenstein R. Locus control regions: overcoming heterochromatin-induced gene inactivation in mammals. Curr Opin Genet Dev. 1997;7:614–619. doi: 10.1016/S0959-437X(97)80008-1. [DOI] [PubMed] [Google Scholar]
- Greaves DR, Wilson FD, Lang G, Kioussis D. Human CD2 3'-flanking sequences confer high-level, T cell-specific, position-independent gene expression in transgenic mice. Cell. 1989;56:979–986. doi: 10.1016/0092-8674(89)90631-4. [DOI] [PubMed] [Google Scholar]
- Wang X, Bergelson JM. Coxsackievirus and adenovirus receptor cytoplasmic and transmembrane domains are not essential for coxsackievirus and adenovirus infection. J Virol. 1999;73:2559–62. doi: 10.1128/jvi.73.3.2559-2562.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy KM, Heimberger AB, Loh DY. Induction by antigen of intrathymic apoptosis of CD4+CD8+TCRlo thymocytes in vivo. Science. 1990;250:1720–1723. doi: 10.1126/science.2125367. [DOI] [PubMed] [Google Scholar]
- Bucy RP, Karr L, Huang G-Q, Li J, Carter D, Honjo K, Lemons JA, Murphy KM, Weaver CT. Single cell analysis of cytokine gene co-expression during CD4+ T-cell phenotype development. Proc Natl Acad Sci U S A. 1995;92:7565–7569. doi: 10.1073/pnas.92.16.7565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher KJ, Choi H, Burda J, Chen SJ, Wilson JM. Recombinant adenovirus deleted of all viral genes for gene therapy of cystic fibrosis. Virology. 1996;217:11–22. doi: 10.1006/viro.1996.0088. [DOI] [PubMed] [Google Scholar]
- Kochanek S, Clemens PR, Mitani K, Chen HH, Chan S, Caskey CT. A new adenoviral vector: Replacement of all viral coding sequences with 28 kb of DNA independently expressing both full-length dystrophin and beta-galactosidase. Proc Natl Acad Sci U S A. 1996;93:5731–6. doi: 10.1073/pnas.93.12.5731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parks RJ, Chen L, Anton M, Sankar U, Rudnicki MA, Graham FL. A helper-dependent adenovirus vector system: removal of helper virus by Cre-mediated excision of the viral packaging signal. Proc Natl Acad Sci U S A. 1996;93:13565–70. doi: 10.1073/pnas.93.24.13565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haskins K, Kubo R, White J, Pigeon M, Kappler J, Marrack P. The major histocompatibility complex-restricted antigen receptor on T cells. I. Isolation with a monoclonal antibody. J Exp Med. 1983;157:1149–1169. doi: 10.1084/jem.157.4.1149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu KH, Lonberg-Holm K, Alstein B, Crowell RL. A monoclonal antibody specific for the cellular receptor for the group B coxsackieviruses. J Virol. 1988;62:1647–52. doi: 10.1128/jvi.62.5.1647-1652.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schorpp M, Jager R, Schellander K, Schenkel J, Wagner EF, Weiher H, Angel P. The human ubiquitin C promoter directs high ubiquitous expression of transgenes in mice. Nucleic Acids Res. 1996;24:1787–8. doi: 10.1093/nar/24.9.1787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers WO, Weaver CT, Kraus LA, Li J, Li L, Bucy RP. Visualization of antigen-specific T cell activation and cytokine expression in vivo. J Immunol. 1997;158:649–57. [PubMed] [Google Scholar]
- Szabo SJ, Jacobson NG, Dighe AS, Gubler U, Murphy KM. Developmental commitment to the Th2 lineage by extinction of IL-12 signaling. Immunity. 1995;2:665–675. doi: 10.1016/1074-7613(95)90011-x. [DOI] [PubMed] [Google Scholar]
- Bucy RP, Panoskaltsis-Mortari A, Huang G-Q, Li J, Karr L, Ross M, Russell JH, Murphy KM, Weaver CT. Heterogeneity of single-cell cytokine gene expression in clonal T cell populations. J Exp Med. 1994;180:1251–1262. doi: 10.1084/jem.180.4.1251. [DOI] [PMC free article] [PubMed] [Google Scholar]