

SUMMARY
In February 2008, three white-nosed coatis (Nasua narica) were found dead in a recreational park in Cancun, Mexico. The diagnosis of rabies virus (RABV) infection was confirmed by direct immunofluorescence test. The phylogenetic analysis performed with the complete RABV nucleoprotein gene positioned this isolate close to a sequence of a human rabies case reported during 2008 from Oaxaca, Mexico, sharing 93% similarity. In turn, these two variants are related to another variant found in rabid Tadarida brasiliensis mexicana bats across North America. Anti-RABV neutralizing activity (1.3 IU/ml) was found in the serum of one white-nosed coati captured with another five that cohabited with the dead animals. Enhanced rabies surveillance and pathogenesis studies should be conducted in coatis and insectivorous bats of the region to clarify the role of these species as potential emergent or long-term unidentified RABV reservoirs.
Keywords: Insectivorous bats, outbreak, rabies, white-nosed coatis
Rabies is an extremely lethal encephalitis caused by all members of the Lyssavirus genus. Rabies virus (RABV) is the most widespread member of the 11 major lyssavirus species identified worldwide [1]. This genotype has great diversity in the Americas and is maintained enzootic by specific associations within different species of mammalian carnivores and bats.
Rabies reservoirs should be distinguished from other mammals that are susceptible to RABV infection but do not play a key role in the perpetuation of the disease in nature (e.g. ruminants, rodents, humans, etc.) [2].
White-nosed coatis (Nasua narica) belong to the family Procyonidae. These social carnivores are commonly found in different neo-tropical park reserves from Central America to the Southwestern USA. Exposure of coatis to RABV has been documented from incidental surveillance reports in the USA, and suggested from ecological studies in Northwestern Mexico [3]. However, these animals are rarely found rabid, and have not been reported as a source of RABV transmission to humans. In February 2008, rabies was detected in three white-nosed coatis at Kabah National Park in Cancun, Quintana Roo, Mexico. Given the low frequency of rabies in this species, the objective was to identify and characterize the RABV variant affecting these animals. Three young female white-nosed coatis were found dead on 11, 18 and 19 February. The last one presented clear signs of having been scavenged by unidentified animals. The brain samples were recovered by the Secretaría del Medio Ambiente y Recursos Naturales (SEMARNAT) Mexico (Environment and Natural Resources Ministry) and submitted to two rabies reference laboratories in Mexico City. Rabies was confirmed by the standard direct florescence test (DFA) (http://www.cdc.gov/ncidod/dvrd/rabies/Professional/publications/DFA_diagnosis/DFA_protocol-b.htm) performed in brain tissues at the Laboratorio Estatal de Salud Pública (LESP) (State Health Authorities) in Quintana Roo, Mexico. Brain tissues were kept in a buffered PBS glycerol solution at −70 °C until used for typing.
Virus isolation was performed using the mouse inoculation test (MIT) as described by the WHO [4]. Clinical signs in mice occurred between days 15 and 21 with a fatality rate of 95.6%. The presence of RABV antigen in mice brains was confirmed by the standard DFA.
Antigenic typing of isolated RABV was performed using an indirect fluorescent antibody technique with a panel of eight monoclonal antibodies standardized by the Centers for Disease Control and Prevention (CDC) in Atlanta, Georgia, to identify major RABV circulating in Latin American countries and the Caribbean [5]. The RABV isolates were identified as antigenic variant 9 (AgV 9), which is associated with enzootic rabies in Tadarida brasiliensis mexicana bats in the USA and Mexico.
Genetic typing was performed by sequencing the complete RABV nucleoprotein of the isolated virus from the coati brains and was compared with sequences from a diversity of rabid animals obtained throughout Mexico [6]. To corroborate if the RABV isolated from these coatis was a new RABV variant, an overall phylogeny was reconstructed using a Bayesian Markov Chain Monte Carlo (BMCMC) method (v. 3.2) [7]. The phylogenetic reconstruction was based on a dataset representative of RABV diversity in the Americas (including all terrestrial mammals [6] and bat RABV variants [8]). Nucleotide substitutions were modelled under GTR +Γ4+I, and allowed adequate generations of MCMC sampling in order that the effective sample size for each estimated parameter would exceed 200. The phylogenetic reconstruction placed RABV into two clear phylogroups, one associated with mammalian carnivores and the other encompassing primarily bat-associated lineages, with two extraordinary inclusions: one raccoon lineage and the other of at least three skunk lineages (Fig. 1). An Arizona skunk lineage has been recently identified as a spillover event from Eptesicus fuscus with further adaptation to skunks [9]. The RABV variants isolated from rabid coatis in Quintana Roo, Mexico segregated as an independent highly supported sublineage, within a cluster grouping two other RABV variants typed as antigenic AgV 9. These two variants are associated with enzootic rabies in T. b. mexicana in Mexico and the USA (TbNA), and a recently discovered variant obtained from two human rabies cases (in 2005 and 2008) in Oaxaca, Mexico [8]. The putative RABV reservoir for the Oaxaca variant has not been identified to date. The antigenic typing corroborated that these RABV variants are related. The phylogenetic reconstruction contributed to better define the suggestion that these viruses, despite sharing common ancestry, have evolved as independent RABV variants. The sequence similarity in the three isolates from Kabah was 100% along the entire RABV nucleoprotein gene sequence. The genetic distances between the new coati variant and the TbNA RABV and Oaxaca variants were 6.9% and 6.6%, respectively. In addition a conserved asparagine at position 397 was found in association with RABV variants obtained from these coatis.
Fig. 1.
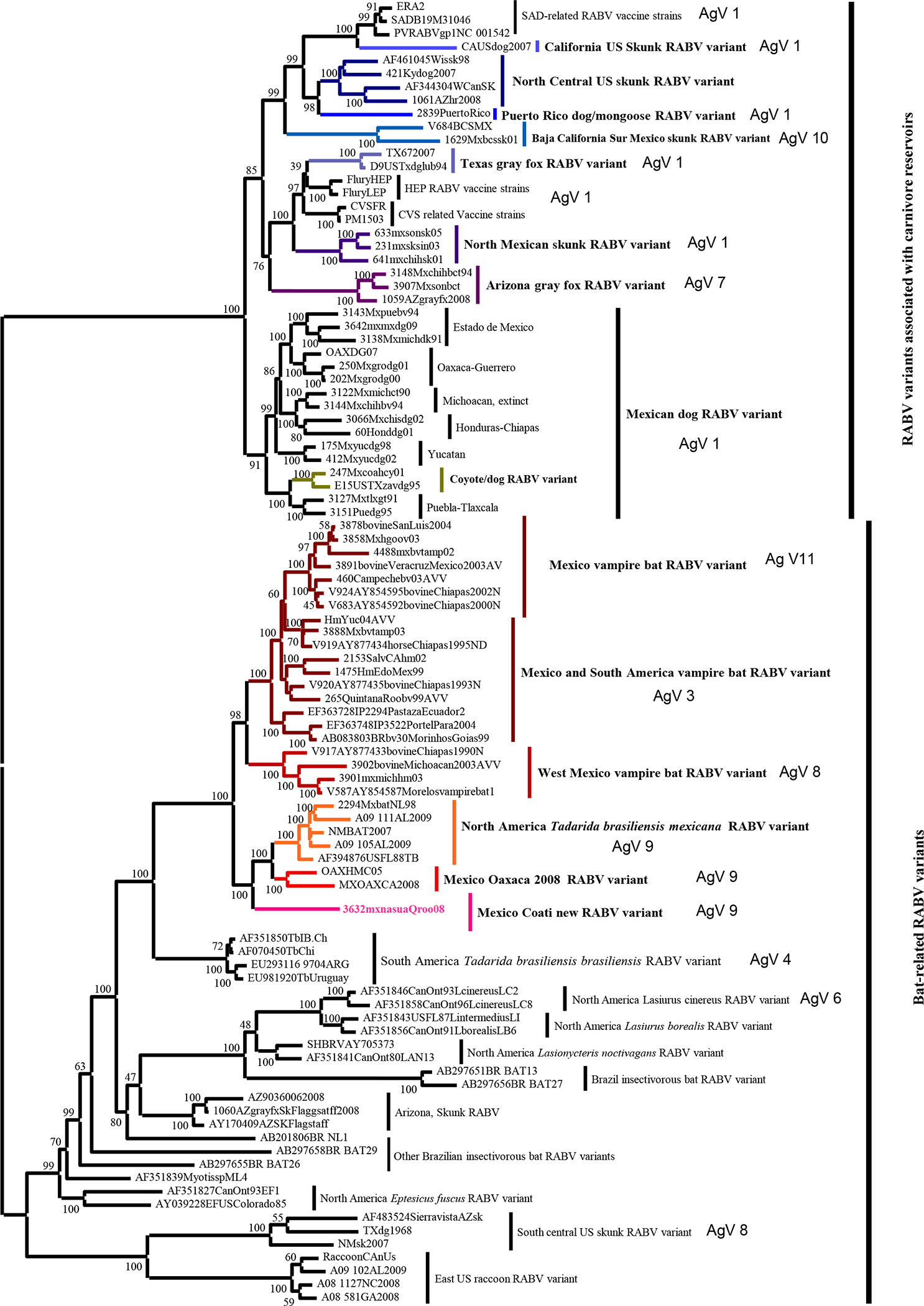
Phylogenetic tree of the complete Lyssavirus nucleoprotein gene, comparing the new variant isolated from coatis with representative antigenic variants of rabies virus circulating in Mexico and North America in mammalian carnivores and bat reservoirs.
Similar to the Oaxaca human cases, the primary origin of the virus reported here may be related with RABV variants currently circulating in insectivorous bats, belonging to the family Molossidae [8]. The current reported geographical distribution for T. b. mexicana does not include the Yucatan Peninsula [10]. Alternatively, this case may be representative of putative host-switching events, similar to the cases reported in Arizona [9]. However, in the present case, the RABV more likely originated from a molossid bat virus that consequently adapted to coatis in the region. Nowadays, the main natural rabies reservoirs described in the Yucatan Peninsula are domestic dogs and haematophagous bats. Non-haematophagous bats can act as a source of infection to humans and other animals. Carnivores prey upon dead, injured, or sick bats, and subsequent infections with bat RABV are not uncommon [6, 8, 9].
The discovery of all three cases just few days apart would suggest that all animals may have been infected by the same source or by different rabid animals within the same focus, which is supported by the phylogenetic and genetic analyses.
Six wild white-nosed coatis from Kabah Park were captured within a hectare using Tomahawk live-traps 5 days after confirmation of the cases, to examine the extent of the outbreak. The animals were kept under observation for 30 days, looking for clinical signs of rabies and were released thereafter. Procedures of manipulating and sampling wild-living animals were made according to the Ley General de Vida Silvestre (General Law of Wild life) Mexico. Blood samples were obtained from jugular or femoral veins to test for the presence of RABV neutralizing antibodies (VNA). None of these coatis (five females and one male) showed clinical signs of rabies. The VNA were determined using the rapid fluorescent focus inhibition test (RFFIT) [11]. The serum corresponding to the male coati presented VNA (1.3 IU/ml). This finding supports the concept that males may be more prone to RABV infection given their more solitary social behaviour.
There is a paucity of information on the possible role of white-nosed coatis in the epidemiology of RABV. Coatis belong to the family Procyonidae, the same as raccoons (Procyon lotor), which are known to be a natural reservoir of rabies. More studies are needed to clarify the coatis’ role.
Coatis are charismatic species, common in parks and tourist areas. Besides the risk factor they could represent to tourists in Mexico, white-nosed coatis could pose a rabies risk to other species, since they are part of the diet of large felids, including jaguars (Panthera onca) and pumas (Puma concolor) [3, 12].
Infectious diseases should be considered and prioritized within long-term wildlife management plans, if conservation objectives are implicit. Enhancement of rabies surveillance in bats and wild carnivores within protected and recreational parks should be undertaken to better document the role of potential reservoirs of this new RABV variant and to determine if new patterns of host adaptation have occurred.
Supplementary Material
ACKNOWLEDGEMENTS
We thank Verónica Torres of the Laboratorio de Variación Biológica y Evolución of the ENCB-IPN for the technical support. This work was partially supported by CONACyT’s scholarship awarded to Nidia Aréchiga. The findings and conclusions in this report are those of the authors and do not necessarily represent the views of the funding agency. Use of trade names and commercial sources are for identification only and do not imply endorsement by the US Department of Health and Human Services.
Footnotes
DECLARATION OF INTEREST
None.
Supplementary material accompanies this paper on the Journal’s website (http://journals.cambridge.org/hyg).
RERERENCES
- 1.International Committee on Taxonomy of Viruses. ICTV Files and Discussions, ICTV Master Species List 2009 – Version 3 (http://talk.ictvonline.org/files/ictv_documents/m/ms//1231.aspx). Accessed January 2010.
- 2.Smith JS. Rabies virus epitopic variation: use in ecologic studies. Advances in Virus Research 1989; 36: 215–253. [DOI] [PubMed] [Google Scholar]
- 3.Valenzuela D Natural history of the white-nosed coati, Nasua narica, in a tropical dry forest of Western Mexico. Revista Mexicana de Mastozoología 1998; 3: 26–44. [Google Scholar]
- 4.Koprowsky H The mouse inoculation test. In: Meslin FX, Kaplan MM, Koprowsky H, eds. Laboratory Techniques in Rabies. Geneva: World Health Organization, 1996, pp. 80–87. [Google Scholar]
- 5.Diaz AM, et al. Antigenic analysis of rabies-virus isolates from Latin American and the Caribbean. Journal of Veterinary Medicine Series B 1994; 41: 153–160. [DOI] [PubMed] [Google Scholar]
- 6.Velasco-Villa A, et al. Enzootic rabies elimination from dogs and reemergence in wild terrestrial carnivores, United States. Emerging Infectious Diseases 2008; 14: 1849–1854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ronquist F, Huelsenbeck JP. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003; 19: 1572–1574. [DOI] [PubMed] [Google Scholar]
- 8.Velasco-Villa A, et al. Identification on a new rabies virus variant in a Mexican immigrant. Emerging Infectious Diseases 2008; 12: 1906–1908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Leslie MJ, et al. Bat-associated rabies virus in skunks. Emerging Infectious Diseases 2006; 12: 1274–1277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hall ER. The Mammals of North America, 2nd edn. New York: John Wiley & Sons Inc. 1981, pp. 173–176, 240–247, 1013–1018. [Google Scholar]
- 11.Smith JS, Yager PA, Baer GM. A rapid reproducible test for determining rabies neutralizing antibodies. Bulletin of the World Health Organization 1973; 48: 535–541. [PMC free article] [PubMed] [Google Scholar]
- 12.Valenzuela D, Ceballos G, Garcia A. Mange epizootic in white-nosed coatis in Western Mexico. Journal of Wildlife Diseases 2000; 36: 56–63. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.