

Abstract
Background
Plants have evolved various defense mechanisms against insect herbivores, including the formation of physical barriers, the synthesis of toxic metabolites, and the activation of phytohormone responses. Although plant-associated microbiota influence plant growth and health, whether they play a role in plant defense against insect pests in natural ecosystems is unknown.
Results
Here, we show that leaves of beetle-damaged weeping willow (Salix babylonica) trees are more resistant to the leaf beetle Plagiodera versicolora (Coleoptera) than those of undamaged leaves. Bacterial community transplantation experiments demonstrated that plant-associated microbiota from the beetle-damaged willow contribute to the resistance of the beetle-damaged willow to P. versicolora. Analysis of the composition and abundance of the microbiome revealed that Pseudomonas spp. is significantly enriched in the phyllosphere, roots, and rhizosphere soil of beetle-damaged willows relative to undamaged willows. From a total of 49 Pseudomonas strains isolated from willows and rhizosphere soil, we identified seven novel Pseudomonas strains that are toxic to P. versicolora. Moreover, re-inoculation of a synthetic microbial community (SynCom) with these Pseudomonas strains enhances willow resistance to P. versicolora.
Conclusions
Collectively, our data reveal that willows can exploit specific entomopathogenic bacteria to enhance defense against P. versicolora, suggesting that there is a complex interplay among plants, insects, and plant-associated microbiota in natural ecosystems.
Graphical Abstract

Video Abstract
Supplementary Information
The online version contains supplementary material available at 10.1186/s40168-024-01884-z.
Keywords: Willow, Plagiodera versicolora, Pseudomonas, Colonization, Defense
Background
Plants face various environmental and biotic challenges during their lifetime. Among these challenges, insect herbivores represent a pervasive threat and are responsible for about 20% of the loss in total crop production worldwide annually [1]. Plants have evolved versatile defense strategies that help them withstand insect attacks, including the establishment of physical barriers, production of toxic metabolites, and activation of phytohormone responses [2].
Plants and their associated microbiomes have co-evolved for more than 400 million years and have formed a “holobiont” consisting of the plant host and its symbionts [3]. Plant-associated microbiota are also referred to as the plant’s second genome and play fundamental roles in plant growth and stress tolerance [4]. Accumulating evidence indicates that plants can recruit certain beneficial microbes to suppress the growth of soil-borne pathogens [5]. For example, the roots of pathogen-infected wheat [6, 7], sugar beet [8], and Arabidopsis [9] plants can attract groups of beneficial microbes that protect the next generation of plants. Pseudomonas spp. are bacteria that play an important role in promoting plant growth and pathogen resistance by producing various antimicrobial metabolites [10]. A greater richness of related Pseudomonas spp. has been demonstrated to be correlated with pathogen inhibition in plants and is required for plant health [11, 12]. Some strains of Pseudomonas fluorescens group, such as P. protegens and P. chlororaphis, also cause systemic infections and the eventual death of several Lepidopteran, Dipteran, Coleopteran, and Hemipteran insects [13–16]. However, whether plant-associated bacteria contribute to plant defenses against insects in natural ecosystems is unknown.
Plagiodera versicolora is one of the most damaging pest species to Salicaceae plants such as willow (Salix) and poplar (Populus). These insects are widely distributed across northern Africa, America, Europe, and Asia. Both P. versicolora larvae and adults feed on the leaves of willow and poplar, especially during the summer. During the winter season, adult P. versicolora individuals burrow into the soil to undergo overwintering, posing a continued threat to plants in the following year [17]. The life cycle of P. versicolora consists of several stages, including an egg stage lasting 5–6 days, a larval stage lasting 14 days, a pupal stage lasting 4–5 days, and an adult stage that spans 3–4 weeks [18, 19]. Since Salicaceae plants are the natural hosts of P. versicolora, the Salix-P. versicolora association has been utilized as a model system to investigate the evolution adaptations of herbivorous insects in terms of their resource-utilization traits [20].
In this study, using the weeping willow (Salix babylonica)-P. versicolora association as a model system, we revealed that insect-damaged willows can enrich their microbiota in specific microbes that enhance their resistance against further damage by the insect pests. We demonstrate that insect-damaged willows are more resistant to P. versicolora than healthy willows in the field. We further identified certain novel Pseudomonas strains with insecticidal activities that are enriched in the microbiota of willows that were damaged by P. versicolora. Our results suggest a mechanism by which plants exploit specific entomopathogenic bacteria for defense against insect pests in nature.
Methods
Sampling
Samples were collected from two willow (Salix babylonica) field trials. Field site 1 (S1) is located at the Shahu Lake (30° 34′ 17″ N, 114° 20′ 4″ E), and field site 2 (S2) is located at the Nanhu lake (30° 28′ 47″ N, 114° 21′ 33″ E) in Wuhan, Hubei province, China (SI Appendix, Fig. S1A and 1B). The two sampling sites are located in a temperate climate zone with the same annual mean temperature of 22 °C and annual mean precipitation of 1343.5 mm. The leaves, roots, and rhizosphere soil of willows were sampled on August 11–12, 2021, as previously described [21]. The rhizosphere soil was defined as the soil particles adhering to the roots [21]. At each site, willows without beetle damage were classified as healthy (SI Appendix, Fig. S1C). Willows damaged by beetles were classified as sick (SI Appendix, Fig. S1D). The healthy leaves of healthy willows will be abbreviated as HL-HW, the healthy leaves of sick willows as HL-SW, and the sick leaves of sick willows as SL. Ten replicates from healthy and sick willows were collected from ten adjacent plots at each site. The distance between healthy and sick willows was about 0.5 km (SI Appendix, Fig. S1E). The samples were transported with dry ice and stored at − 80 °C until analysis.
Processing of samples
Willow leaves (three leaves per sample) were surface-sterilized with 75% (v/v) ethanol for 1 min and washed with sterile Millipore water three times [22]. Root samples were rinsed with sterile water to wash off the attached soil, surface-sterilized with 2.5% NaClO (w/v) for 5 min and subsequently with 70% (v/v) ethanol for 1 min, and washed with sterile Millipore water three times [23]. The treated plant samples (leaves and roots) were then chopped into small pieces using a sterile scalpel and homogenized with a sterile pestle under axenic conditions. Rhizosphere soil and homogenized plant samples were stored at − 80 °C prior to DNA extraction.
Preparation of axenic willow leaves
Preparation of axenic willow leaves was conducted as described previously [24–26]. Briefly, sick leaves (SL) and healthy leaves (HL) from sample sites were washed under running tap water for 3 min to remove all the dirt particles and impurities, and subsequently soaked in 75% alcohol for 1 min. After rinsed with sterile water three times, the leaves were immersed in 0.1% HgCl2 for 5 min, and rinsed with sterile water three times. Removal of bacteria was confirmed by a colony-forming unit assay and PCR analysis using conserved primers for the 16S rRNA gene of bacteria (Fig. S2).
DNA extraction
Approximately 250 mg of rhizosphere soil was used for DNA extraction using a PowerSoil DNA Isolation Kit following the manufacturer’s instructions (MoBio, Carlsbad, CA, USA). DNA extraction from willow leaves and roots (100 mg per sample) was performed using a MoBioPowerPlant Pro DNA Isolation Kit (Mo Bio Laboratories) according to the manufacturer’s protocol.
Analysis of microbiota
The bacterial communities from all samples were determined based on their 16S rRNA amplicon profiles [22, 27] on an Illumina NovaSeq platform. Sequencing libraries from bacterial DNA were generated with a NEBNext® Ultra™ IIDNA Library Prep Kit (Cat No. E7645). Raw sequences were split according to their unique barcodes and trimmed off the adaptors and primer sequences using QIIME [28]. Paired-end reads were merged using FLASH (Version 1.2.11, http://ccb.jhu.edu/software/FLASH/). Quality filtering of the raw tags was performed using fastp software (version 0.20.0) to obtain high-quality clean tags, which were compared to the reference database (Silva database https://www.arbsilva.de/) to detect and remove chimeric sequences, yielding final effective tags [29]. Denoising was performed with DADA2 in QIIME2 software (Version QIIME2-202,006) to obtain initial amplicon sequence variants (ASVs). The sequences annotated as chloroplast were removed. ASVs with an abundance of < 5 reads were removed [30]. Microbial diversity and community composition were analyzed using vegan packages in R (version 3.5.3).
Non-metric multidimensional scaling (NMDS) based on Bray–Curtis dissimilarities was used to identify differences between microbial communities. Compositional differences in NMDS between healthy and diseased samples were assessed using ANOSIM based on the Bray–Curtis distance (Table S3–8). Linear discriminant analysis (LDA) of effect size (LEfSe) was applied to the ASV table to identify differentially abundant bacterial taxa from the phyllosphere, root, and rhizosphere soil samples between beetle-damaged and healthy willows. Absolute LDA scores (> 2) were used to analyze statistical significance.
Genome sequencing and assembly
Samples were sequenced using multiplexed libraries on a Novaseq 6000 instrument to produce paired-end reads with lengths of 150 bp. For each sample, read quality was assessed with the FastQC tool (http://www.bioinformatics.babraham.ac.uk/projects /fastqc/) and low-quality reads were removed by Quake [31]. All genome assemblies were performed with ABySS [32] using a k-mer size of 21. The best assembly for each strain, with the largest scaffold N50, was annotated by Prokka (v1.14.6) [33]. The final assembly genome information is shown in Table S9.
Phylogenetic analysis
Phylogenetic trees were reconstructed using the maximum likelihood method in MEGA version 7.0 [34]. Bootstrap support values were calculated from 1000 replicates. The phylogenetic tree was visualized with iTOL [35]. The 14 reference strains were as follows: P. quercus (MT036111), P. coleopterorum (KM888184), P. koreensis (AF468452), P. neuropathica (LR797591), P. iridis (LR797554), P. piscium (LR797558), P. parafulva (FNYJ01000011), P. chlororaphis (BBQB01000031), P. alloputida (LT718459), P. flavescens (FNDG01000047), P. alcaligenes (BATI01000076), P. resinovorans (Z76668), P. aeruginosa (BAMA01000316), and P. psychrotolerans (FMWB01000061). Acetobacter ascendens LMG 1590 (CP015164) was selected as an outgroup. The accession numbers of the 16S rRNA sequences are provided within the parentheses.
Quantification of Pseudomonas spp. colony-forming units, strain isolation, and identification
The density of Pseudomonas spp. from the phyllosphere, root, and rhizosphere soil samples was tested by cultivation-based methods. Pseudomonas colony-forming units (CFUs) were determined via serial dilutions. Three aliquots (100 µL) per dilution were spread onto CFC (cephaloridine fucidin cetrimide) medium designed for Pseudomonas selection [36]. The number of colonies was recorded after incubation at 28 °C for 3 days [12]. Pseudomonas isolates were purified, and genomic DNA from each Pseudomonas culture was extracted using a MiniBEST Bacteria Genomic DNA Extraction kit (TaKaRa, China) following the manufacturer’s protocol. The 16S rRNA (1,466 bp) gene was amplified with primer pair 27F and 1492R (SI Appendix, Table S1) and sequenced. The 16S rRNA gene sequence of Pseudomonas was searched against the EzBioCloud Database [37], a well-curated database of 16S rRNA sequences and bacterial genomes.
Insect rearing
P. versicolora adults were collected from willow trees at Sha Lake Park in Wuhan, Hubei Province in China (30.35° N, 114.33 E). P. versicolora was regularly reared by feeding with detached fresh willow leaves at 28 °C and 60 ± 5% relative humidity under a 16-h light/8-h dark photoperiod in transparent plastic boxes (40 cm × 24 cm × 10 cm). Fresh willow leaves were replaced daily in transparent plastic boxes to serve as food for P. versicolora [38]. Newly laid eggs or hatched larvae were collected from willow leaves to conduct insect bioassays (at 28 °C, 60% ± 5% relative humidity, 16-h light/8-h dark photoperiod) [17]. They have been maintained since 2022. The colony is refreshed with wild-type stock every 3 months.
Insect bioassays on detached leaves
The leaves of beetle-damaged and healthy willows were fed to first-instar P. versicolora larvae (n = 30, divided into three experimental groups, with 10 larvae in each group). For each feeding experiment, synchronized larvae were selected, weighed individually, and divided into three groups (each group containing 10 larvae). To calculate the consumed leaf area, the leaves were photographed after feeding by P. versicolora larvae, and the consumed area was determined using ImageJ software (https://imagej.nih.gov/ij/). Pupation rate and eclosion rate were monitored as described [39]. The entire experiment was replicated three times.
The pathogenicity test of Pseudomonas strains against insects employed the previously described methods of Vacheron et al. [14]. First-instar larvae were fed with fresh willow leaves (8 cm2) that had been painted with 5 µL of 5.0 × 106 bacterial cells in sterile phosphate-buffered saline (PBS). Willow leaves painted with the equivalent volume of PBS solution were used as a negative control. Larvae were divided into three groups (for three biological replicates), and each group had 10 larvae per treatment. Willow leaves painted with an equivalent volume of PBS were used as negative control. Treated leaves were exchanged daily, and insect survival was recorded daily for seven consecutive days. The entire experiment was replicated three times.
Synthetic community of leaf bacteria (SynCom)
To generate SynCom, sick leaves (SL) and healthy leaves (HL) from sample sites were harvested (6 leaves were sampled from three individual plants), surface-sterilized with 75% ethanol for 1 min, and washed with sterile water three times. Leaves were ground in sterile water, and bacterial suspensions were diluted to 10−2 and plated on R2A plates for 4 days at 22 °C [22]. About 80 colonies from SL and HL were randomly picked to constitute SynCom-SL or SynCom-HL, respectively. SynCom-SL or SynCom-HL were prepared and the final OD600 was adjusted to 0.06 (~ 2 × 107 CFU/mL), and 10 µL of the SynCom-SL or SynCom-HL suspension was painted to each 1 cm2 of willow leaf.
Introduction of Pseudomonas strains into willow plants
Willow plants were grown in a growth chamber under controlled conditions (25 °C under a 16-h light/8-h dark photoperiod) [40]. Bacterial re-introduction was conducted as described previously [41]. Briefly, Pseudomonas strains were incubated overnight at 28 °C in an orbital shake. Bacterial cells were collected by centrifugation at 4000 rpm for 5 min to remove the supernatant, washed with sterile PBS twice, resuspended in sterile PBS to a cell density of 108 cells/ml, and then inoculated onto the leaves of willow plants. SynCom-mix7 is a mixture of PSE-30, PSE-32, PSE-34, PSE-37, PSE-38, PSE-39, and PSE-49 (1 × 108 cells/ml for total SynCom-7 mixtures; 1.4 × 107 cells/ml for each strain), and used to inoculate leaves of willow plants. Willow plants treated with PBS served as control. The CFU of Pseudomonas strains on the leaves was determined by a serial-dilution method on the CFC plate [36].
Insect bioassays on whole plants
For bioassays with whole plants, first-instar P. versicolora larvae (n = 30, divided into three experimental groups, with 10 larvae in each group) were allowed to feed on the healthy willow plants reintroduced with SynCom-mix7 or Pseudomonas strain PSE37/38. Larvae were divided into three groups, with 10 larvae in each group. Survival rates were recorded daily. The entire experiment was replicated three times.
Statistical analysis of data
Prior to statistical analysis, the normality of data distribution was assessed using the Shapiro–Wilk test (P > 0.05). All statistical analyses were performed using SPSS version 19.0 software. Survival curves of first-instar P. versicolora larvae (n = 30) were analyzed using the Kaplan–Meier method, and the differences in survival curves were determined using the log-rank test with a significance level at P < 0.05. The damaged leaf area, pupation rate, eclosion rate, and larval weight were analyzed using one-way analysis of variance (ANOVA). Data comprising two groups were analyzed using a Student’s t-test for unpaired comparisons, and data consisting of more than two groups were analyzed using one-way analysis of variance. Alpha diversity indices (e.g., Shannon index and Chao1 index) and beta-diversity were calculated using QIIME 2.
Results
Beetle-induced activation of plant defense against P. versicolora
To examine whether plants employ defensive tactics when repeatedly exposed to leaf beetles, we fed P. versicolora larvae with the sick leaves of insect-damaged sick willows (SL), the healthy leaves of healthy willows (HL-HW), and sick willows (HL-SW) that were sampled from two field sites in Wuhan, China (SI Appendix, Fig. S1). We observed that P. versicolora larvae cause less damage to SL than HL-HW or HL-SW, based on the area consumed by the larvae (Fig. 1A and B). Moreover, compared to that with HL, feeding with SL significantly reduced the growth of P. versicolora larvae from day 2 onward (Fig. 1E) and substantially delayed pupation (Fig. 1C) and eclosion rate (Fig. 1D). These results suggest that beetle damage can activate plant defenses against further damage by pests.
Fig. 1.

Feeding assays of P. versicolora larvae with leaves from beetle-damaged sick willows (SW) and healthy willows (HW). A Representative bioassay with detached sick leaves (SL) and healthy leaves (HL) from two sampling sites. Leaves were fed to first-instar P. versicolora larvae and replaced with fresh young leaves daily. The photographs were taken on day 3. Red arrows: the leaf sites damaged by beetle. B Leaf area consumed by first-instar P. versicolora larvae in A. C, D Pupation rate (C) and eclosion rate (D) of P. versicolora feeding with SL and HL from two sampling sites. E Larval weight after the indicated number of days of feeding on SL and HL from two sampling sites. S1, field site 1; S2, field site 2. HL-HW: healthy leaves of healthy willows; HL-SW: healthy leaves of sick willows; SL: sick leaves of sick willows. Significant differences between two groups were determined by Student’s t-test. *P < 0.05, **P < 0.01, ***P < 0.001
Contribution of plant-associated microbiota in plant defense against P. versicolora
To assess the contribution of plant-associated microbiota in the resistance of the beetle-damaged willows to P. versicolora, we assembled two synthetic microbial communities from sick leaves (SynCom-SL) and healthy leaves (SynCom-HL) of sick and healthy willow plants, respectively, and introduced them onto the axenic sick or healthy leaves that had been in vitro sterilized (Fig. 2 and Fig. S3). Compared to the axenic healthy or sick leaves that were reintroduced with SynCom-HL, reintroduction of SynCom-SL significantly reduced the damage to willow leaves (Fig. 2A and B), and substantially delayed pupation (Fig. 2C), eclosion rate (Fig. 2D), and larval weight (Fig. 2E) of P. versicolora. These results demonstrated that plant-associated microbiota from the beetle-damaged willow contribute to the resistance of the beetle-damaged willow to P. versicolora.
Fig. 2.

Feeding assays of P. versicolora larvae with the detached axenic leaves that had reintroduced with SynCom-SL or SynCom-HL. A Representative bioassays with the detached axenic sick leaves (SL-axenic) or healthy leaves (HL-axenic) that had reintroduced with SynCom-SL or SynCom-HL. Leaves were fed to first-instar P. versicolora larvae and refreshed with the axenic leaves reintroduced with SynCom-SL or SynCom-HL daily. The photographs were taken on day 3. B Leaf area consumed by first-instar P. versicolora larvae in A. Pupation rate (C), eclosion rate (D), and larval weight (E) of P. versicolora feeding with the axenic leaves reintroduced with SynCom-SL or SynCom-HL
Pseudomonas spp. is enriched in the phyllosphere of beetle-damaged willows
We next investigated the composition of the bacterial community within the phyllosphere of SL and HL by deep sequencing of amplified bacterial 16S rRNA sequences. Non-metric multidimensional scaling (NMDS) based on Bray–Curtis dissimilarity indicated that bacterial communities from SL and HL were significantly different in samples from both Shahu Lake and Nanhu Lake (Figs. 3A and 4A, B, ANOSIM, P < 0.01). We estimated alpha diversity based on the Shannon index and Simpson index and revealed a significant difference for the α-diversity indices between phyllosphere bacteria from SL and HL from Shahu Lake (Fig. 3B, C, P< 0.01).
Fig. 3.

Assembly of willow bacterial communities. A NMDS of bacterial communities in leaf, root, and rhizosphere soil of willow plants (3 sections). Solid and hollow represent two different sampling sites: Shahu Lake (S1) and Nanhu Lake (S2). Different symbols represent three different compartments: leaf (triangle), root (circle), soil (diamond). Different colors correspond to healthy (blue) and beetle-damaged sick (orange) willows, respectively. B Shannon diversity index of bacterial community in healthy and beetle-damaged sick willows. C Simpson diversity index of bacterial community in healthy and beetle-damaged sick willows
Fig. 4.

Compositional differences in NMDS between healthy and diseased samples were assessed using ANOSIM based on the Bray–Curtis distance. NMDS plots depict the bacterial communities in leaves from Shahu Lake (A) and Nanhu Lake (B), roots from Shahu Lake (C) and Nanhu Lake (D), and rhizosphere soil from Shahu Lake (E) and Nanhu Lake (F). S1, field site 1 located in Shahu Lake; S2, field site 2 located in the Nanhu Lake
Linear discriminant analysis of effect size (LefSe) showed that the genus Pseudomonas was significantly more abundant in the phyllosphere of SL compared to HL (Fig. 5A and B). Similarly, we thus isolated Pseudomonas from SL and HL; indeed, more Pseudomonas species were present in SL samples than in HL (Fig. 5C). These results suggest that Pseudomonas spp. is enriched in willow leaves damaged by beetles.
Fig. 5.

Pseudomonas spp. is enriched in leaves from beetle-damaged sick willows. A and B Linear discriminant analysis (LDA) showing the enrichment patterns of bacteria in beetle-damaged sick leaves (SL) compared to healthy leaves (HL) in different sampling sites S1 (A) and S2 (B) at the genus (g) levels. C Number of Pseudomonas spp. isolated from SL and HL, in CFUs per gram leaf fresh weight. Significant differences between two groups were determined by Student’s t-test. ***P < 0.001
Enrichment of Pseudomonas in the root and rhizosphere soil samples of beetle-damaged willows
Several recent studies have indicated that infections with aboveground pathogens also alter the rhizosphere microbial community [13]. We thus tested whether leaf damage by beetles also influenced the bacterial community in root and rhizosphere soil samples by 16S rRNA amplicon sequencing. NMDS based on Bray–Curtis dissimilarity revealed that bacterial communities from beetle-damaged sick root (SR) and healthy root (HR) were significantly different in samples from both Nanhu and Shahu Lake (Figs. 3A and 4C, D, ANOSIM, P < 0.01). Bacterial communities from beetle-damaged sick willows (SS) and healthy willows (HS) were significantly different in samples from both Nanhu and Shahu Lake (Figs. 3A and 4E, F, ANOSIM, P < 0.05). The alpha diversity of the bacterial community in the SR has no significant difference from that in the HR in terms of Shannon diversity index (Fig. 3B, C, P > 0.05). We observed no significant difference for α-diversity indices between rhizosphere soil bacteria from SS and those from HS (Fig. 3B, C, P > 0.05).
LefSe showed that the genus Pseudomonasis was significantly more abundant in SR compared to HR samples (Fig. 6A and B). Similarly, Pseudomonas bacteria were significantly more abundant in SS compared to HS samples (Fig. 6C and D).
Fig. 6.

Pseudomonasare enriched in roots and rhizosphere soils of beetle-damaged sick willows. A and B LDA showing the enrichment patterns of bacteria in SR compared to HR in different sampling sites S1 (A) and S2 (B) at the genus (g) levels. C and D LDA showing the enrichment patterns of bacteria in SS compared to HS in different sampling sites S1 (C) and S2 (D) at the genus (g) levels. E Number of Pseudomonas strains isolated from rhizosphere root samples from beetle-damaged or healthy willows. F Number of Pseudomonas strains isolated from rhizosphere soil samples from beetle-damaged or healthy willows. HR, root of heathy willow; SR, root of beetle-damaged sick willow; SS, rhizosphere soil of beetle-damaged sick willow. HS, rhizosphere soil of healthy willow. Significant differences between two groups were determined by Student’s t-test. **P < 0.01
We next isolated Pseudomonas from root and rhizosphere soil samples collected from beetle-damaged and healthy willows. We determined that the abundance of Pseudomonas isolated from SR or SS samples is significantly higher than that from HR or HS samples (Fig. 6E and F). These results suggest that Pseudomonas spp. is also enriched in the roots and rhizosphere soil of beetle-damaged willows, similar to the phyllosphere results above.
The Pseudomonas enriched on beetle-damaged plants show insecticidal activity against beetles
We isolated and identified 49 Pseudomonas strains from the phyllosphere, root, and rhizosphere soil samples collected from beetle-damaged and healthy willows. Based on a phylogenetic analysis of these strains, we established that they represent distinct phylogenetic groups based on 16S rRNA gene sequences. Importantly, we detected nine P. psychrotolerans (Pp) strains specifically on samples collected from SL (Fig. 7).
Fig. 7.

Phylogenetic relationships among the Pseudomonas isolated from beetle-damaged and healthy willows. From the innermost to the outside circles: (1) species-level taxonomy of Pseudomonas spp.; (2) Pseudomonas spp. detected in the phyllosphere, root, or rhizosphere soil; (3) Pseudomonas spp. enriched in beetle-damaged willows or healthy willows. The phylogenetic tree was constructed based on bacterial 16S rRNA gene sequences using the maximum likelihood method. Fourteen reference sequences are marked in bold. Acetobacter ascendens LMG 1590 (CP015164) is chosen as the outgroup. The numbers at the nodes indicate the percentage levels of bootstrap support (%) based on analysis of 1000 re-sampled datasets (only values higher than 0.5 are shown)
To test whether isolated P. psychrotolerans strains have insecticidal activity, we allowed P. versicolora larvae to feed on healthy willow leaves that had been painted with a cell suspension of these strains. We scored the survival rate of larvae. Two Pseudomonas strains, P. chlororaphis PcS1-2 isolated from SS and PcR3-3 isolated from SR, were used as positive control. We discovered that, of the nine P.psychrotolerans strains tested, seven resulted in a higher mortality rate for P. versicolora larvae (Fig. 8A). Moreover, P. versicolora larvae caused less damage to willow leaves painted with these P. psychrotolerans strains compared to those painted with phosphate-buffered saline (PBS) as control (Fig. 8B and C). These results demonstrate that some of the Pseudomonas strains that are enriched in the beetle-damaged willows exhibit insecticidal activity against P. versicolora.
Fig. 8.

Insecticidal activity of Pseudomonas strains enriched on willows. A Mortality curves of P. versicolora in response to different isolated Pseudomonas strains. Kaplan–Meier survival curves of P. versicolora larvae (n = 30) after feeding with Pseudomonas-painted willow leaves. Feeding of PBS-painted willow leaves was used as negative control (CK). P. chlororaphis PcS1-2 or PcR3-3 that showed insecticidal activity toward P. versicolora was used as positive control. The survival curves were analyzed by the log-rank test. B Leaf area consumed by first-instar P. versicolora larvae in A. C Representative photographs of a bioassay with detached leaves from healthy willows painted with different isolated Pseudomonas. Leaves were fed to first-instar P. versicolora larvae and replaced with fresh young leaves daily. The photographs were taken after 3 days of feeding
To identify potential genes associated with the observed insecticidal activities, we performed genome sequencing and assembly of P. psychrotolerans strains PSE37 or PSE38. Our analysis revealed the absence of genes encoding insecticidal proteins such as Fit toxin [42, 43], IPD072 [16], Chitinase C, and Phospholipase C [13], which are commonly present in P. protegens and P. chlororaphis [13], in P. psychrotolerans strains PSE37 or PSE38 genome (SI Appendix, Table S9).
Inoculation of a synthetic microbial community (SynCom) enhances willow resistance to beetles on whole plants
To measure the contribution of the enrichment of Pseudomonas strains in the beetle-damaged willows to the resistance to P. versicoloraon whole plants, we reconstructed a synthetic microbial community (SynCom-mix7) composed of seven P. psychrotolerans (Pp) strains isolated from SL. Re-inoculation of SynCom-mix7 or Pseudomonas strain PSE37/38 to the healthy willow brought out 56% mortality of P. versicolora larvae compared to those inoculated with PBS control (Fig. 9).
Fig. 9.
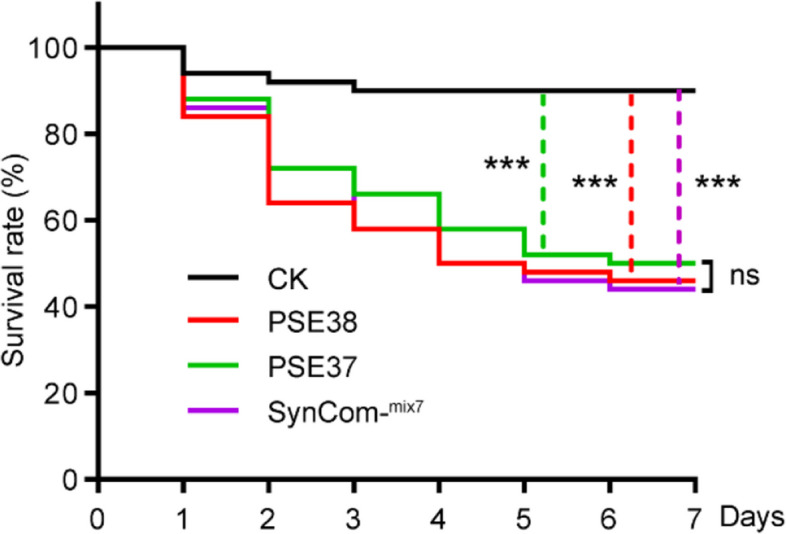
Whole plant bioassays with SynCom-mix7 or Pseudomonas strain PSE37/38. Survival rates of P. versicolora larvae (n = 30) fed on the healthy willows introduced with SynCom-mix7, Pseudomonas strain PSE37 and PSE38. Healthy willows treated with PBS solution (without bacteria) served as negative control (CK). The log-rank test was used to assess the significance of differences between two survival curves
Discussion
Our data show that Pseudomonas strains become enriched in the microbiota of willows damaged by insect herbivores. We further determined that some of these enriched Pseudomonas strains have insecticidal activity, thus contributing to the protection of willows from further attacks by P. versicolora. We were able to reconstruct a synthetic microbial community (SynCom) of Pseudomonas that could enhance willow resistance to P. versicolora. It has been reported that SynComs have been reconstructed for the purpose of biocontrol. An example of this is the SynCom consisting of Flavobacterium and Chitinophaga, which has been shown to enhance sugar beet resistance to fungal root disease [8]. These findings provide new insights into the functional significance of insect damage-induced enrichment of Pseudomonas communities in willows. Consistent with this idea, plants inoculated with rhizobacterium decreased the growth of caterpillars after 4 days of feeding compared to non-inoculated control plants [44]. Similarly, recent studies showed that fungal infection of plant roots can lead to the assemblage of bacterial groups with disease-suppressive functions [8]. However, it should be noted that changes in plant nutrition, metabolite biosynthesis and secretion, and phytohormone contents can also significantly contribute to the plant resistance against insect pests. For example, introgression of the 7-epizingiberene biosynthetic pathway from wild tomato (Solanum habrochaites) to tomato (Solanum lycopersicum) cultivars produced plants that can synthesize this chemical that is toxic to spider mites while also making them less attractive to whiteflies [45].
We showed that the aboveground insect damage can also induce the assemblage of an enriched community of Pseudomonas in the roots and rhizosphere soil of willows (Fig. 3), suggesting that plant microbiome assembly and functions in the below and aboveground compartments under insect damage may be systemically linked. In agreement with this notion, several recent studies have also suggested that the pathogen infection of plants aboveground can induce the assemblage of a plant-beneficial bacterial consortium in the root or soil [8, 46–48]. For example, Berendsen et al. [9] reported that upon foliar infection with the oomycete Hyaloperonospora arabidopsidis, three bacterial taxa are specifically enriched in the rhizosphere of Arabidopsis plants, induce systemic resistance against pathogens, and promote plant growth. It remains to be determined whether Pseudomonas spp. enriched in the rhizosphere soil also confer resistance against willow pathogens or increase the growth of willows.
In this study, we identified several novel Pseudomonas strains from the phyllosphere with insecticidal activities against P. versicolora. However, the underlying mechanism and the genes responsible for these insecticidal activities are unknown. For example, P. psychrotolerans strains PSE37 or PSE38 were shown to be toxic to P. versicolora but lack the genes encoding insecticidal proteins (SI Appendix, Table S9). Therefore, the novel insecticidal Pseudomonas identified in this study could provide new insecticidal genes. Although a group of insecticidal Pseudomonas strains was shown to be enriched in the phyllosphere of willows (Figs. 5 and 7), their origin in the phyllosphere is unclear. It has been suggested that the rhizosphere soil could be a major source of bacteria detected in the phyllosphere [49–52]. Whether insecticidal Pseudomonas strains can migrate from the rhizosphere soil to the phyllosphere needs to be investigated experimentally.
Conclusions
In summary, this study illuminates the complex interplay between plants, insects, and plant-associated microbiota in a natural system. By using willow, willow leaf beetle, and the associated microbiota as a model, we have uncovered a natural mechanism whereby upon damage by insect herbivory, plants can enrich their phyllosphere with insecticidal Pseudomonasto raise their defense against further damage by insect pests, providing new insight on plant defense against pests.
Supplementary Information
Additional file 1. Fig. S1–S3 and Table S1–S8.
Authors’ contributions
F. Z. and J.Z. conceived and designed the project; H.W., Y.Z., Y.Z., and M.W. conducted the experiments; H.W., F. Z. Y.Z. and J.Z. analyzed the data; F. Z. and J.Z. wrote the article with contributions from all other authors.
Funding
This research was supported by the National Key R&D Program of China (2023YFC2607000), the National Natural Science Foundation of China (32271912 and 32271546), and Key Program in AGIS under Grand No. AGIS-ZDXM-202304.
Availability of data and materials
No datasets were generated or analysed during the current study.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Haitao Wang and Fengjuan Zhang contributed equally to this work.
References
- 1.Divekar PA, Narayana S, Divekar BA, Kumar R, Gadratagi BG, Ray A, et al. Plant secondary metabolites as defense tools against herbivores for sustainable crop protection. Int J Mol Sci. 2022;23:2690. 10.3390/ijms23052690 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mostafa S, Wang Y, Zeng W, Jin B. Plant responses to herbivory, wounding, and infection. Int J Mol Sci. 2022;23:7931. 10.3390/ijms23137031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Trivedi P, Leach JE, Tringe SG, Sa T, Singh BK. Plant-microbiome interactions: from community assembly to plant health. Nat Rev Microbiol. 2020;18:607–21. 10.1038/s41579-020-0412-1 [DOI] [PubMed] [Google Scholar]
- 4.Li H, La S, Zhang X, Gao L, Tian Y. Salt-induced recruitment of specific root-associated bacterial consortium capable of enhancing plant adaptability to salt stress. ISME J. 2021;15:2865–82. 10.1038/s41396-021-00974-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gao M, Xiong C, Gao C, Tsui CKM, Wang MM, Zhou X, et al. Disease-induced changes in plant microbiome assembly and functional adaptation. Microbiome. 2021;9:187. 10.1186/s40168-021-01138-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Liu H, Li J, Carvalhais LC, Percy CD, Prakash Verma J, Schenk PM, et al. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytol. 2021;229:2873–85. 10.1111/nph.17057 [DOI] [PubMed] [Google Scholar]
- 7.Yin C, Casa Vargas JM, Schlatter DC, Hagerty CH, Hulbert SH, Paulitz TC. Rhizosphere community selection reveals bacteria associated with reduced root disease. Microbiome. 2021;9:86. 10.1186/s40168-020-00997-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Carrión VJ, Perez-Jaramillo J, Cordovez V, Tracanna V, de Hollander M, Ruiz-Buck D, et al. Pathogen-induced activation of disease-suppressive functions in the endophytic root microbiome. Science. 2019;366:606–12. 10.1126/science.aaw9285 [DOI] [PubMed] [Google Scholar]
- 9.Berendsen RL, Vismans G, Yu K, Song Y, de Jonge R, Burgman WP, et al. Disease-induced assemblage of a plant-beneficial bacterial consortium. ISME J. 2018;12:1496–507. 10.1038/s41396-018-0093-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hansen ML, Wibowo M, Jarmusch SA, Larsen TO, Jelsbak L. Sequential interspecies interactions affect production of antimicrobial secondary metabolites in Pseudomonas protegens DTU9.1. ISME J. 2022;16:2680–90. [DOI] [PMC free article] [PubMed]
- 11.Sun X, Xu Z, Xie J, Hesselberg-Thomsen V, Tan T, Zheng D, et al. Bacillus velezensis stimulates resident rhizosphere Pseudomonas stutzeri for plant health through metabolic interactions. ISME J. 2022;16:774–87. 10.1038/s41396-021-01125-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tao C, Li R, Xiong W, Shen Z, Liu S, Wang B, et al. Bio-organic fertilizers stimulate indigenous soil Pseudomonas populations to enhance plant disease suppression. Microbiome. 2020;8:137. 10.1186/s40168-020-00892-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Flury P, Aellen N, Ruffner B, Péchy-Tarr M, Fataar S, Metla Z, et al. Insect pathogenicity in plant-beneficial pseudomonads: phylogenetic distribution and comparative genomics. ISME J. 2016;10:2527–42. 10.1038/ismej.2016.5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Vacheron J, Péchy-Tarr M, Brochet S, Heiman CM, Stojiljkovic M, Maurhofer M, et al. T6SS contributes to gut microbiome invasion and killing of an herbivorous pest insect by plant-beneficial Pseudomonas protegens. ISME J. 2019;13:1318–29. 10.1038/s41396-019-0353-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Vesga P, Flury P, Vacheron J, Keel C, Croll D, Maurhofer M. Transcriptome plasticity underlying plant root colonization and insect invasion by Pseudomonas protegens. ISME J. 2020;14:2766–82. 10.1038/s41396-020-0729-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Schellenberger U, Oral J, Rosen BA, Wei JZ, Zhu G, Xie W, et al. A selective insecticidal protein from Pseudomonas for controlling corn rootworms. Science. 2016;354:634–7. 10.1126/science.aaf6056 [DOI] [PubMed] [Google Scholar]
- 17.Xu L, Xu S, Sun L, Zhang Y, Luo J, Bock R, et al. Synergistic action of the gut microbiota in environmental RNA interference in a leaf beetle. Microbiome. 2021;9:98. 10.1186/s40168-021-01066-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wade MJ, Breden F. Life history of natural populations of the imported willow leaf beetle, Plagiodera versicolora (Coleoptera: Chrysomelidae). Ann Entomol Soc Am. 1986;79:73–9. 10.1093/aesa/79.1.73 [DOI] [Google Scholar]
- 19.Ishihara M, Hayashi T, Ohgushi TJES. Life cycle of the willow leaf beetle, Plagiodera versicolora (Coleoptera: Chrysomelidae) in Ishikari (Hokkaido, Japan). Entomol Sci. 1999;2:57–60. [Google Scholar]
- 20.Utsumi S, Ando Y, Ohgushi T. Evolution of feeding preference in a leaf beetle: the importance of phenotypic plasticity of a host plant. Ecol Lett. 2009;12:920–9. 10.1111/j.1461-0248.2009.01349.x [DOI] [PubMed] [Google Scholar]
- 21.Beckers B, Op De Beeck M, Weyens N, Boerjan W, Vangronsveld J. Structural variability and niche differentiation in the rhizosphere and endosphere bacterial microbiome of field-grown poplar trees. Microbiome. 2017;5:25. 10.1186/s40168-017-0241-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chen T, Nomura K, Wang X, Sohrabi R, Xu J, Yao L, et al. A plant genetic network for preventing dysbiosis in the phyllosphere. Nature. 2020;580:653–7. 10.1038/s41586-020-2185-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dove NC, Veach AM, Muchero W, Wahl T, Stegen JC, Schadt CW, et al. Assembly of the populus microbiome is temporally dynamic and determined by selective and stochastic factors. mSphere. 2021;6:e01316–20. 10.1128/mSphere.01316-20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ugur R, Research E. Development of in vitro sterilization protocol for DO-1 (Prunus domestica) rootstock. Appl Ecol Environ Res. 2020;18:2339–49. 10.15666/aeer/1802_23392349 [DOI] [Google Scholar]
- 25.Asmono SL, Wardana R. Optimization of the sterilization method for leaf explant Robusta BP 308 coffee in vitro. IOP Conference Series: Earth Environ Sci. 2022;980:012001. [Google Scholar]
- 26.Barpanda S, Beura S, Rout S, Jagadev P. Studies on in vitro regeneration of Sandalwood (Santalum album Linn) from leaf disc explant. J Pharmacogn Phytochemistry. 2017;6:892–6. [Google Scholar]
- 27.Bai Y, Müller DB, Srinivas G, Garrido-Oter R, Potthoff E, Rott M, et al. Functional overlap of the Arabidopsis leaf and root microbiota. Nature. 2015;528:364–9. 10.1038/nature16192 [DOI] [PubMed] [Google Scholar]
- 28.Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods. 2010;7:335–6. 10.1038/nmeth.f.303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Haas BJ, Gevers D, Earl AM, Feldgarden M, Ward DV, Giannoukos G, et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011;21:494–504. 10.1101/gr.112730.110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li M, Shao D, Zhou J, Gu J, Qin J, Chen W, et al. Signatures within esophageal microbiota with progression of esophageal squamous cell carcinoma. Chin JCancer Res. 2020;32:755–67. 10.21147/j.issn.1000-9604.2020.06.09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kelley DR, Schatz MC, Salzberg SL. Quake: quality-aware detection and correction of sequencing errors. Genome Biol. 2010;11:R116. 10.1186/gb-2010-11-11-r116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Simpson JT, Wong K, Jackman SD, Schein JE, Jones SJ, Birol I. ABySS: a parallel assembler for short read sequence data. Genome Res. 2009;19:1117–23. 10.1101/gr.089532.108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30:2068–9. 10.1093/bioinformatics/btu153 [DOI] [PubMed] [Google Scholar]
- 34.Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–4. 10.1093/molbev/msw054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Letunic I, Bork P. Interactive Tree Of Life (iTOL) v4: recent updates and new developments. Nucleic Acids Res. 2019;47:W256–9. 10.1093/nar/gkz239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mead GC, Adams BW. A selective medium for the rapid isolation of Pseudomonads associated with poultry meat spoilage. Br Poult Sci. 1977;18:661–70. 10.1080/00071667708416418 [DOI] [PubMed] [Google Scholar]
- 37.Chun J, Lee JH, Jung Y, Kim M, Kim S, Kim BK, et al. EzTaxon: a web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences. Int J Syst Evol Mic. 2007;57:2259–61. 10.1099/ijs.0.64915-0 [DOI] [PubMed] [Google Scholar]
- 38.Wang H, Zhang Y, Dai D, Fu J, Sung Kim D, Li S, et al. Genomic insight into the insecticidal potential of a new Pseudomonas chlororaphis isolate. J Econ Entomol. 2024;117:82–92. 10.1093/jee/toad232 [DOI] [PubMed] [Google Scholar]
- 39.Ohsawa M. Life cycle, ecological characteristics, and control of Trachys yanoi (Coleoptera: Buprestidae), an important pest of Zelkova serrata. Insects. 2017;8:35. 10.3390/insects8020035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhu LH, Xu W, Huang L, Ye JR, Li DW. Pathogenicity and biological characteristics of Septotinia populiperda causing leaf blotch of willow. Plant Dis. 2022;106:1262–70. 10.1094/PDIS-07-21-1537-RE [DOI] [PubMed] [Google Scholar]
- 41.Wilson M, Lindow SE. Inoculum density-dependent mortality and colonization of the phyllosphere by Pseudomonas syringae. Appl Environ Microbiol. 1994;60:2232–7. 10.1128/aem.60.7.2232-2237.1994 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Péchy-Tarr M, Bruck DJ, Maurhofer M, Fischer E, Vogne C, Henkels MD, et al. Molecular analysis of a novel gene cluster encoding an insect toxin in plant-associated strains of Pseudomonas fluorescens. Environ Microbiol. 2008;10:2368–86. 10.1111/j.1462-2920.2008.01662.x [DOI] [PubMed] [Google Scholar]
- 43.Ruffner B, Péchy-Tarr M, Höfte M, Bloemberg G, Grunder J, Keel C, et al. Evolutionary patchwork of an insecticidal toxin shared between plant-associated pseudomonads and the insect pathogens Photorhabdus and Xenorhabdus. BMC Genomics. 2015;16:1–14. 10.1186/s12864-015-1763-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Pangesti N, Reichelt M, van de Mortel JE, Kapsomenou E, Gershenzon J, van Loon JJ, et al. Jasmonic acid and ethylene signaling pathways regulate glucosinolate levels in plants during rhizobacteria-induced systemic resistance against a leaf-chewing herbivore. J Chem Ecol. 2016;42:1212–25. 10.1007/s10886-016-0787-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bleeker PM, Mirabella R, Diergaarde PJ, VanDoorn A, Tissier A, Kant MR, et al. Improved herbivore resistance in cultivated tomato with the sesquiterpene biosynthetic pathway from a wild relative. Proc Natl Acad Sci USA. 2012;109:20124–9. 10.1073/pnas.1208756109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yuan J, Zhao J, Wen T, Zhao M, Li R, Goossens P, et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome. 2018;6:156. 10.1186/s40168-018-0537-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Thomashow LS, LeTourneau MK, Kwak YS, Weller DM. The soil-borne legacy in the age of the holobiont. Microb Biotechnol. 2019;12:51–4. 10.1111/1751-7915.13325 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bakker P, Pieterse CMJ, de Jonge R, Berendsen RL. The soil-borne legacy. Cell. 2018;172:1178–80. 10.1016/j.cell.2018.02.024 [DOI] [PubMed] [Google Scholar]
- 49.Massoni J, Bortfeld-Miller M, Widmer A, Vorholt JA. Capacity of soil bacteria to reach the phyllosphere and convergence of floral communities despite soil microbiota variation. Proc Natl Acad Sci USA. 2021;118:e2100150118. 10.1073/pnas.2100150118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zarraonaindia I, Owens SM, Weisenhorn P, West K, Hampton-Marcell J, Lax S, et al. The soil microbiome influences grapevine-associated microbiota. mBio. 2015;6:e02527-14. 10.1128/mBio.02527-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Tkacz A, Bestion E, Bo Z, Hortala M, Poole PS. Influence of plant fraction, soil, and plant species on microbiota: a multikingdom comparison. mBio. 2020;11:02785–19. 10.1128/mBio.02785-19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Copeland JK, Yuan L, Layeghifard M, Wang PW, Guttman DS. Seasonal community succession of the phyllosphere microbiome. Mol Plant Microbe Interact. 2015;28:274–85. 10.1094/MPMI-10-14-0331-FI [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1. Fig. S1–S3 and Table S1–S8.
Data Availability Statement
No datasets were generated or analysed during the current study.