

Abstract
Snake venom is an ecologically relevant functional trait directly linked with a snake’s fitness and survival, facilitating predation and defence. Snake venom variation occurs at all taxonomic levels, but the study at the intraspecific level is still in its early stages. The common adder (Vipera berus) exhibits considerable variation in colour phenotypes across its distribution range. Melanistic (fully black) individuals are the subject of myths and fairytales, and in German folklore such ‘hell adders’ are considered more toxic than their normally coloured conspecifics despite any formal investigation. Here, we provide the first comparative analysis of venoms from melanistic and normally coloured common adders. Specifically, we compared the venom profiles by sodium dodecylsulfate polyacrylamide gel electrophoresis and reversed-phase high-performance liquid chromatography and tested the venoms’ protease, phospholipase A2 and cytotoxic activities. Phospholipase A2 activity was similar in both phenotypes, whereas general protease activity was higher in the melanistic venom, which was also more cytotoxic at two concentrations (6.25 and 12.5 µg ml−1). These minor differences between the venoms of melanistic and normally coloured adders are unlikely to be of clinical relevance in the context of human envenomation. In light of our results, the claim that melanistic adders produce more toxic venom than their normally coloured conspecifics appears rooted entirely in folklore.
Keywords: Vipera berus, snake venom, venom variation, melanism, colour phenotypes, folklore
1. Introduction
Venom has evolved multiple times independently in the animal kingdom, and serves the three major functions of hunting, defence and intraspecific competition [1–3]. Concerning snakes, evidence suggests that snake venom evolved to mainly aid predation [4–6]. However, selection of snake venom composition resulting from it being used for defensive purposes has also been demonstrated [7]. Snake venoms are commonly defined as complex mixtures of biomolecules, described as toxins, most of which are proteins or peptides [1]. Initially, snake venoms were considered to be well defined and conserved within species and between closely related species. However, several studies have revealed the dynamic nature of this ecologically relevant trait, highlighting both interspecific [8] and intraspecific differences in venom composition [9]. In the latter case, venom profiles have been shown to differ according to sex [10], life stage [11] and geographical origin [12]. Although this phenomenon of venom variation provides insight into the evolutionary ecology of snakes and other venomous animals, it also makes the treatment of envenomation more challenging [9]. If the symptoms of envenomation differ according to the individual animal, this affects both clinical decision making and the efficacy of any anti-venom that is administered [9]. Venom variation is particularly important in snakes because snakebite is classified by the World Health Organization as a priority 1 neglected tropical disease [13]. Up to 2.7 million cases are estimated to occur every year, leading to hundreds of thousands of fatalities and/or permanent disabilities [14,15].
In Europe, the most widespread venomous snake is the common adder (Vipera berus). The taxonomy of this species is not completely resolved, with several subspecies being the subject of ongoing discussion [16–19]. Adult specimens are on average 50−70 cm in length, rarely exceeding 80 cm [20]. The neonates feed primarily on small amphibians and reptiles, although adults will also prey on birds and smaller mammals [20–22]. One characteristic trait of common adders is their wide range of colour and pattern phenotypes [20,21]. The most common phenotype is a pale grey (males) or brown (females) base colour with a dark dorsal zigzag pattern that forms a V-shaped motif on the head. These normally coloured individuals are often also referred to as ‘cryptic’; however, some authors argue for an aposematic function of this normal colouration [23–27], while others proposed that negative frequency-dependent selection maintains colour polymorphism of adders [28]. The dorsal zigzag pattern varies in width, and the body colour can shift towards reddish brown or even blue. The most unique phenotypes are copper-coloured and black (melanistic) specimens. Based on our own observations, their phenotype can change over the course of maturation. For instance, most of the melanistic adders are born with a cryptic pattern and change to the melanistic phenotype, while relatives of the same litter retain a cryptic phenotype. Interestingly, colour phenotypes may have ecological repercussions in adders. For example, melanistic adders occur in many populations but vary in abundance even though they are easier for predators to see [25,29]. The fitness trade-off may involve the higher sunlight-to-body-temperature conversion rate of melanistic compared to cryptic forms, particularly in colder temperatures and during shorter days, helping to increase metabolic activity and foraging, which could be translated into earlier maturation, an earlier start to the annual mating season, shorter egg development times and a better fitness overall [23,25,29]. A selection of colour phenotypes of the common adder is presented in figure 1.
Figure 1.

Colour and dorsal pattern variation in German Vipera berus: (a) specimen with classic colour phenotype, exhibiting greyish background colouration, conspicuous black dorsal zigzag pattern and a clear V-shaped head marking; (b–d) specimens exhibiting red/brown/copper background colouration and different levels of intensity in the dorsal pattern and V-shaped marking; (e) specimen with blueish background colouration, less marked zigzag pattern and inconspicuous V-shaped head marking; (f) melanistic specimen, with fully black colouration and unnoticeable dorsal pattern and V-shaped head marking. Created with BioRender.
Compared to other European vipers, human envenomation by V. berus is rarely fatal, but the incidence is high because the species is broadly distributed, including colder regions that are not inhabited by other venomous snakes [30–32]. Due to this high bite incidence, the common adder ranks among the clinically most relevant venomous snakes [30–32]. The venom profile of V. berus has been analysed (in a few cases using modern venomics technologies) and seems to match the typical viperine profile dominated by members of the phospholipase A2 (PLA2) family, snake venom metalloproteases (svMPs), serine proteinases (svSPs) and C-type lectins (CTLs), including snaclecs and C-type lectin-related proteins [33–35]. Vipera berus also appears to show intraspecific venom variation because the proportion and abundance of dominant and lesser toxin families vary between studies and according to the origin [8] and assigned subspecies [36] of each specimen.
Interestingly, the common adder and particularly its venom are mentioned in European folklore, especially across German-speaking countries. Here, melanistic adders have a reputation for greater aggression and toxicity than adders exhibiting lighter colour phenotypes, and were initially considered a separate species, earning the name ‘hell adder’ [21,37]. This largely reflects superstitions connected to dark animals, including cats, dogs and ravens, which have been integrated into myths and folklore, often as symbols of bad luck, treachery, evil and witchcraft [38]. While it is now understood that ‘hell adders’ are simply melanistic animals, in certain parts of Europe, such as the more rural parts of Germany, black-coloured adders are still feared as more toxic in the general public, despite lacking scientific basis. Phenotype-dependent venom variation is rarely investigated [39], but a comparison of venom from a rare melanistic rattlesnake (Crotalus durissus terrificus) and its conspecifics provided some support for the hypothesis [40]. However, a single specimen does not provide enough data to draw firm conclusions.
Here we set out to clarify whether or not melanistic adders may indeed have distinct venom profiles or increased toxicity in comparison to normally coloured conspecifics. Therefore, we carried out the first comparative analysis of venom composition and venom bioactivity in adders of different phenotypes too. We compared the venom profiles, enzymatic activities and cytotoxicity of venom from normally coloured (cryptic, CRY) and melanistic (MEL) phenotypes. Our work provides insights into venom variation in adders and could be used as the basis for future comparative venomics studies in snakes and to facilitate adder conservation.
2. Material and methods
2.1. Venom
Venom was donated by members of the German Society of Herpetology and Herpetoculture, and was sourced from captive individuals of German origin from North-Rhine Westphalia and Bavaria. Venom was pooled from nine adult males of each phenotype (MEL or CRY) and lyophilized. Only male individuals of German origin were used for this study. Since V. berus has smaller amounts of crude venom and only a limited number of specimens were available, we opted for pooling the venom prior to lyophilization and laboratory analyses. Lyophilized venoms were redissolved in double-distilled water and aliquots were stored at −20°C.
2.2. Compositional venom profiling
Compositional venom profiling was carried out as previously described [41] by combining reducing and non-reducing sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-PAGE) with 5 µg of each sample and reversed-phase high-performance liquid chromatography (RP-HPLC) with 125 µg of each sample. Chromatograms (220 nm) were subtracted by an initial equilibration run to reduce noise. Raw gel image is provided in electronic supplementary material, figure S1.
2.3. Phospholipase A2 assay
Phospholipase activity was measured using the EnzChek Phospholipase A2 Assay Kit (Invitrogen, cat. no. E10217) for 96-well plates. Venoms were redissolved in 1 × Reaction Buffer and added in triplicates at final concentrations of 3.125, 6.25, 12.5, 25 and 50 µg ml−1. Autolytic activity was minimized by handling all samples on ice and reducing the pre-incubation time. After incubation for 30 min at room temperature, the plates were transferred to a Synergy H4 Hybrid Microplate Reader (BioTek) and analysed using Gen 5 v. 2.09 software (BioTek). The signal was detected at 515 nm following excitation at 470 nm. Values were averaged and normalized against the positive control (5 U ml−1 purified bee venom phospholipase A2, 100%) and negative control (1× Reaction Buffer, 0%). Normalized raw data from the phospholipase A2 activity assays are provided in electronic supplementary material, table S1.
2.4. Protease activity assay
We used a non-specific Protease Activity Assay Kit (Calbiochem, cat. no. 539125) for 96-well plates as previously described [41]. Venoms were redissolved in double-distilled water and added in triplicates at final concentrations of 25, 50, 100, 200 and 400 μg ml−1, as described above. The reactions were incubated for 2 h at 37°C, shaking at 120 rpm on a Multitron device (Infors HT) and the OD492 was measured in an Eon microplate reader (BioTek). The signals were averaged and normalized to the positive control (166 µg ml−1 trypsin, 100%) and negative control (double-distilled water, 0%). Normalized raw data from the protease activity assay are provided in electronic supplementary material, table S2.
2.5. Cell viability assay
The cytotoxicity of venoms was assessed in Madin-Darby canine kidney II (MDCKII) cells using the CellTiter-Glo Luminescent Cell Viability Assay (Promega, cat. no. G7570) as previously reported [42]. The venoms were redissolved in cultivation medium and added to final concentrations of 1.56, 3.125, 6.25, 12.5 and 25 µg ml−1. Luminescence was measured using a Synergy H4 Hybrid Microplate Reader, and mean values for each treatment were normalized against the positive control (100 µM ionomycin, 0% growth) and negative control (cultivation medium, 100% growth). Normalized raw data from the cell viability assay are provided in electronic supplementary material, table S3.
2.6. Haemolytic assay
The haemolytic activity was based on a previously described method [43] that was adjusted to a 96-well format. To purify horse erythrocytes, whole blood was washed in Alsever buffer until the supernatant prepared by centrifugation (804g, 5 min, 4°C) was clear. The pellet was resuspended using a cut 1000 µl tip to reduce shear forces. Purified erythrocytes were diluted in Alsever buffer to a final concentration of 1% (w/v). Venom was redissolved to final assay concentrations of 5, 10, 20, 40 and 80 µg ml−1 and added in triplicate to the erythrocyte suspension at a 1 : 1 ratio in V-bottom 96-well plates. The signal was detected at OD405 and mean values were normalized to the positive control (1% Triton X-100, 100% lysis) and negative control (Alsever buffer, 0% lysis). Normalized raw data from the haemolytic activity assay are provided in electronic supplementary material, table S4.
3. Results
3.1. Compositional profiling
We separated 5 µg of each venom by reducing SDS-PAGE (figure 2a ), resulting in protein denaturation and the separation of monomeric proteins, and also by non-reducing SDS-PAGE (figure 2b ), to preserve any disulfide bonds holding together multimeric complexes. This provided a comprehensive overview of the protein composition to facilitate our analysis of putative toxin families. Reducing SDS-PAGE revealed protein mass ranges of 12−70 kDa for both phenotypes. MEL venom featured slightly more intense bands at approximately 14, 23, 31 and 47−70 kDa, with a distinct band at approximately 55 kDa (figure 2a ). Non-reducing SDS-PAGE revealed a mass range of 12−115 kDa for both phenotypes. MEL venom featured more intense bands at 76–80, approximately 90 and 115 kDa compared to CRY venom, but the latter featured a more intense band at 32−34 kDa (figure 2b ).
Figure 2.

Analysis of Vipera berus venom samples from melanistic (MEL) and cryptic (CRY) phenotypes by SDS-PAGE under (a) reducing and (b) non-reducing conditions. Putative toxin families were predicted according to described Viperidae toxin families, their polymerization potential and the previously reported composition of V. berus venom [33–36]. Abbreviations: 5N, 5′-nucleotidase; AP, aminopeptidase; CTL, C-type lectin including snaclec and C-type lectin-related proteins; DC-Fragments, disintegrin-like/cysteine-rich protein (fragments); HYAL, hyaluronidase; LAAO, l-amino acid oxidase; NGF, nerve-growth factor; PLA2, phospholipase A2; PLB, phospholipase B-like; PDE, phosphodiesterase; QC, glutaminyl cyclase; svMP, snake venom metalloprotease; svSP, snake venom serine proteinase; VEGF, vascular endothelial growth factor. Created with BioRender.
We then separated 125 µg of each venom by RP-HPLC (figure 3) revealing peaks over a retention time of 7−27 min, with the highest peak for CRY venom at around 20 min and for MEL venom at around 18 min. Clusters of peaks were observed for both venoms at 7−14, 14.45−19, 19.30−23 and 24−27 min, although the peaks at 8.30 min, 16.45 min and in the last two clusters were overall smaller in MEL than CRY venom. The smaller peaks and peak cluster were mostly similar in appearance, but the small peak cluster at 13 min and the peak at 16 min were much less apparent in CRY venom. While some differences in peak intensity seem to be present between samples, their venom profile (i.e. chromatographic landscape) appears largely similar.
Figure 3.

Chromatograms of Vipera berus venoms. The chromatograms (220 nm) correspond to venoms obtained from melanistic (orange line) and cryptic (blue line) specimens, eluted and measured using a gradient of acetonitrile (dashed line) over a 30 min time frame. Differences in peak intensities are highlighted by colouring the area between the traces, using the colour code of the venom exhibiting the more intense signals.
3.2. Bioactivity profiling
Next, we tested the bioactivity of the venoms in enzymatic assays covering the dominant venom toxin families of V. berus (PLA2 and the svMP and svSP families) and in cytotoxicity assays against the model mammalian cell line MDCKII and equine erythrocytes.
PLA2 activity (figure 4) in both venoms was assessed at concentrations of 3.125, 6.25, 12.5, 25 and 50 µg ml−1. The observed activity ranged between 38.86% (CRY, 3.125 µg ml−1) and 101.47% (MEL, 50 µg ml−1), relative to assay buffer (0%) and 5 U ml−1 bee venom PLA2 (100%). PLA2 activity was similar between MEL venom (3.125 µg ml−1, 41.87%; 6.25 µg ml−1, 62.33%; 12.5 µg ml−1, 81.06%; 25 µg ml−1, 101.37%; 50 µg ml−1, 101.47%) and CRY venom (3.125 µg ml−1, 38.86%; 6.25 µg ml−1, 64.32%; 12.5 µg ml−1, 85.15%; 25 µg ml−1, 98.09%; 50 µg ml−1, 99.76%), with standard deviations exceeding relative activity disparities at most concentrations.
Figure 4.
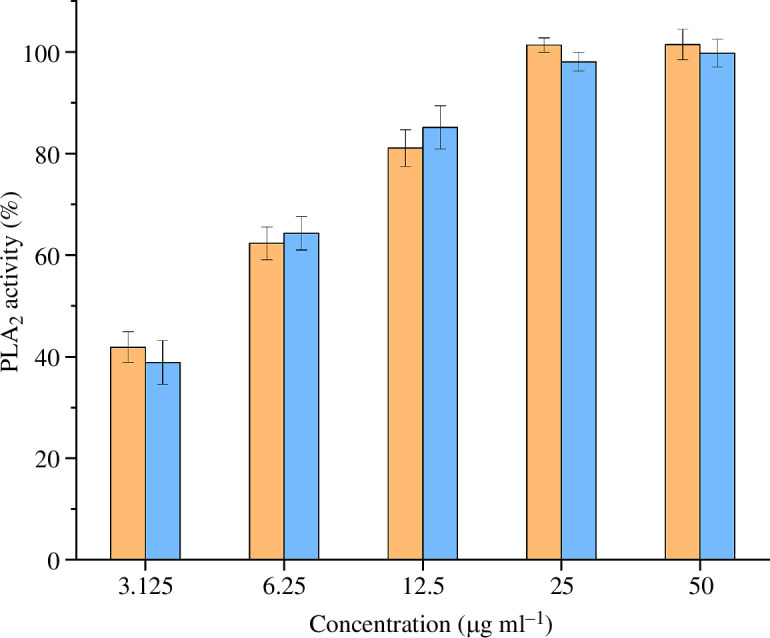
Phospholipase A2 (PLA2) activity of the two Vipera berus venoms. Venoms from melanistic and cryptic specimens are represented in orange and blue, respectively. The graph represents the normalized (positive control: bee venom PLA2, 100%; negative control: buffer, 0%) results of the five concentrations tested (i.e. 3.125, 6.25, 12.5, 25 and 50 µg ml−1). Data are means ± s.d. of technical replicates (n = 3).
In the protease activity assay, we applied the MEL and CRY venoms at concentrations of 25, 50, 100, 200 and 400 µg ml−1. The protease activity (figure 5) ranged between 1.57% (CRY, 25 µg ml−1) and 46.00% (MEL, 400 µg ml−1) relative to the negative control (0%) and the positive control (100%). At every tested concentration, the MEL venom (25 µg ml−1, 7.46%; 50 µg ml−1, 10.80%; 100 µg ml−1, 22.49%; 200 µg ml−1, 33.83%; 400 µg ml−1, 46.00%) showed higher activity than the CRY venom (25 µg ml−1, 1.57%; 50 µg ml−1, 3.41%; 100 µg ml−1, 8.21%; 200 µg ml−1, 15.60%; 400 µg ml−1, 28.18%). Thus, the MEL venom showed a tendency for higher protease activity compared to CRY venom.
Figure 5.
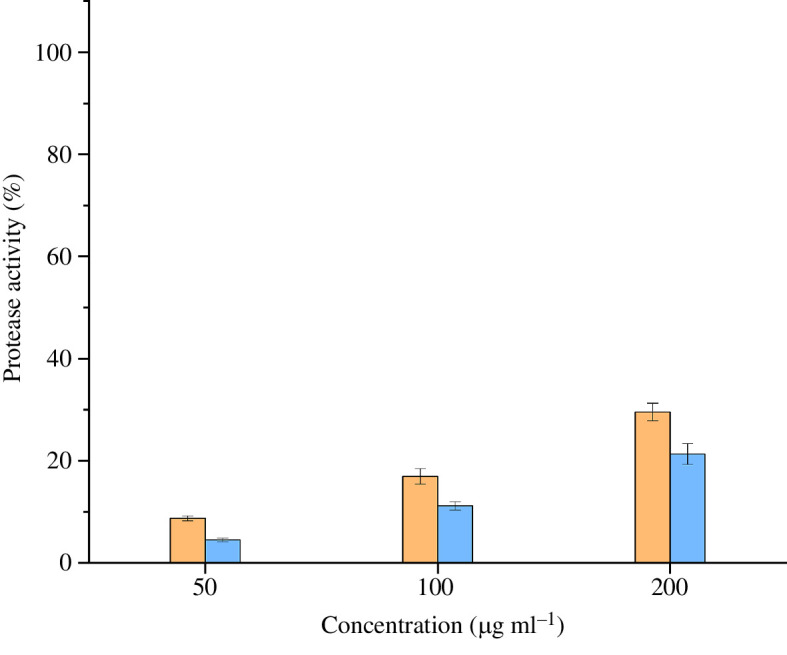
Protease activity of the two Vipera berus venoms. Venoms from melanistic and cryptic specimens are represented in orange and blue, respectively. The graph represents the normalized (positive control: trypsin, 100%; negative control: ddH2O, 0%) results of the five concentrations tested (i.e. 25, 50, 100, 200 and 400 µg ml−1). Data are means ± s.d. of technical replicates (n = 3).
The cytotoxic effects of the venoms were tested at concentrations of 1.56, 3.125, 6.25, 12.5 and 25 µg ml−1 against canine MDCKII cells (figure 6). Strong effects on cell viability were observed regardless of the venom type at a concentration of 25 µg ml−1 (MEL 0.49%, CRY 0.35%), but no effects were detected at 3.125 µg ml−1 (MEL −101.50%, CRY 124.55%) or 1.56 µg ml−1 (MEL 115.96%, CRY 119.04%). At 6.25 µg ml−1 (MEL 86.27%, CRY 114.26%) and 12.5 µg ml−1 (MEL 27.54%, CRY 67.76%), the decrease in cell viability caused by the venom from melanistic animals was higher than that of the CRY venom, although the absence of biological replicates renders our data to be insufficient for statistical comparison.
Figure 6.
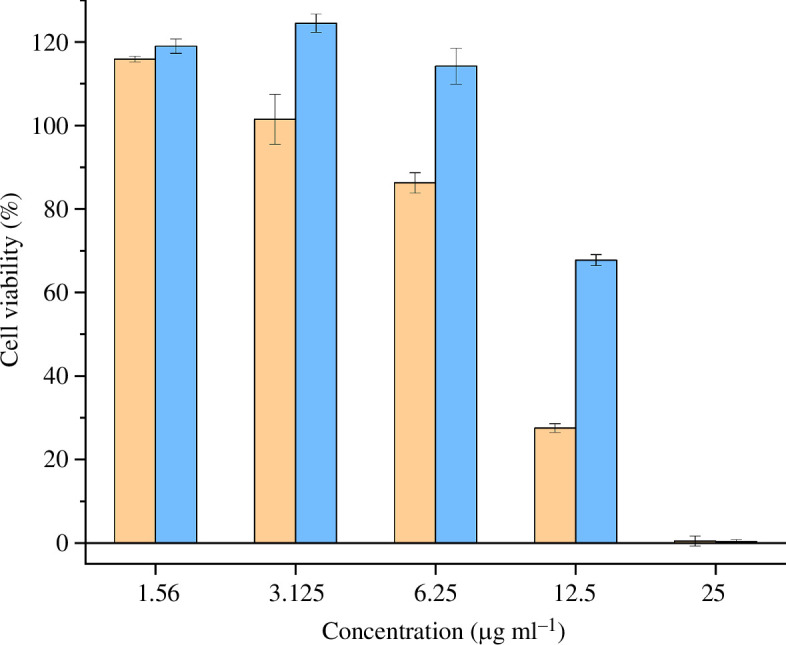
Normalized cell viability of the two Vipera berus venoms. Venoms from melanistic and cryptic specimens are represented in orange and blue, respectively. The graph represents the normalized (positive control: ionomycin, 0% growth; negative control: cultivation media, 100%) results of the five concentrations tested (i.e. 1.56, 3.125, 6.25, 12.5 and 25 µg ml−1) against MDCKII cell lines. Data are means ± s.d. of technical replicates (n = 3).
We also tested the venom against purified horse erythrocytes at concentrations of 5, 10, 20, 40 and 80 µg ml−1, but no haemolysis was detected regardless of the venom type or its concentration (electronic supplementary material, figure S4).
4. Discussion
Melanistic common adders have a reputation across Europe for being more toxic than normally coloured ones. Although this perception appears to be based on folklore and superstition rather than empirical evidence, it was never tested scientifically. To our knowledge, this is the first work formally investigating the presence of differences between the venoms of specimens of the two phenotypes in terms of composition and biological activities.
The comparison of CRY and MEL by SDS-PAGE and RP-HPLC venom profiles revealed qualitative similarities (i.e. number of bands and peaks did not differ between the two venoms) but quantitative differences (i.e. bands and peaks presented different intensities between the two venoms). This variation partly translated into differences in enzymatic activity among the dominant toxin families, with MEL venom showing a trend for higher protease (svMP and svSP) activity, whereas PLA2 activity was comparable between the samples. However, we observed little difference in cytotoxicity between the venoms when tested against a canine (MDCKII) cell line, and there was no haemolytic activity against equine erythrocytes.
The analysis of the venom profiles through SDS-PAGE at reducing and non-reducing conditions allowed us to identify putative venom toxins based on known Viperidae venom components, their potential to form multimeric complexes and the previously reported venom composition of V. berus [33–36]. The MEL banding pattern was generally more intense than its CRY counterpart, especially at approximately 31 and 47−70 kDa in reducing gels and at 76−115 kDa in non-reducing gels. Based on the analysis by Al-Shekhadat et al. [33], these bands likely represent dominant V. berus venom components such as svMP and svSP, suggesting that they are more abundant in the MEL venom. Previous assessments of V. berus venom proteases showed strong activity for fractions in the range 50−66 kDa in reducing gels [44]. In the same size range of this proteolytically active fraction, the bands were more intense in MEL venom, potentially explaining differences observed in the bioactivity profiling.
Indeed, although the analysis of enzyme activity reflecting the dominant components of V. berus venom revealed no consistent differences in PLA2 activity, it suggested an increased protease activity in MEL venoms. Significant differences in protease activity between individual V. berus specimens but consistent levels of PLA2 activity have been reported before [44]. In light of this, it is possible that the composition and activity of V. berus venom could vary in terms of proteases, but remain relatively stable in the context of PLA2 activity within populations and between individuals and phenotypes.
The cytotoxicity of the MEL venom appeared higher than that of the CRY venom, but only when tested at 6.25 or 12.5 µg ml−1. This limited effect in a narrow concentration range is unlikely to be clinically relevant because adult adders inject 10−18 mg of venom during human envenomation [33]. State-of-the-art in vivo assays to assess the potency of venom usually test for necrosis, lethality, neurotoxicity and other effects in mice or rats which is difficult to translate into human symptomatics [44–46]. Similarly, our cytotoxicity data cannot directly predict the symptoms of envenomation or the lethal dosage. That said, it is a useful approximation towards the potential for degenerative necrosis in the affected area, with differences occurring in a limited range of concentrations. Further, we observed no evidence of haemolytic activity against equine erythrocytes at the concentrations we tested. Case reports mention haemoconcentration, which is often caused by haemolytic activities, as symptoms of V. berus envenomation in only 5% and 10% of incidents, respectively, suggesting haemolysis as a minor effect [31].
The analysis of intraspecific venom variation is relatively novel in the field of toxicology, and the role of a snake’s phenotype in its occurrence has rarely been considered relevant thus far, resulting in a lack of data. However, a rare case of melanism in the pit viper C. d. terrificus prompted a comparison of its venom profile with a normally coloured specimen [40]. The analysis of protein bands revealed a core venom composition, albeit with some quantitative variation, as well as unique bands in each specimen. Although the analysis of a single specimen is not representative, the study nevertheless showed a similar trend to our observations, with the venom from the melanistic snake yielding more intense bands corresponding to larger toxins. However, the venom composition of V. berus berus specimens from Russia [33,35] and the Slovakian Republic [34], as well as one V. berus barani specimen from Türkiye [36], revealed remarkable quantitative diversity for PLA2 (10% [35], 17.9% [36], 25.3% [33], 59% [34]), as well as svMP (0.2% [36], 3.15% [34], 17.2% [33], 19% [35]) and svSP (15% [34], 16.2% [33], 31% [35], 46.1% [36]). In relation to this extreme compositional variation across the sampling sites in previous studies, the venom variation suggested by our SDS-PAGE and RP-HPLC appeared marginal and only affected the abundance of certain fractions rather than their overall diversity. Also, recent studies in related species of the subfamily Viperinae suggest ontogenetic shifts, diet and environmental conditions as important drivers of venom variation, which are certainly more dominant than phenotype dependencies [39,47–50]. Previous studies of intra-population venom variation in V. berus from Hungary revealed individual venom variation similar in magnitude to our observations, with quantitative as well as qualitative differences [44]. Accordingly, the few quantitative differences suggested by our SDS-PAGE and RP-HPLC profiling as well as the functional differences determined in our bioassays may represent only the normal biological variability of V. berus venom instead of an attribute of a specific colour phenotype.
In light of that, our exploratory investigation is limited by the restricted number of specimens and low sample volumes, forcing us to pool samples and resulting in the lack of biological replicates. Therefore, we have been unable to run meaningful statistical comparisons. Accordingly, our results need to be interpreted with caution and more detailed follow-up investigations should be carried out. Thus, considering the great extent of biological variation within V. berus and given the many factors that can influence venom composition, these future studies should be based on larger sample sizes. In addition, they should account for relevant confounding factors such as the life stage, sex and diet of the specimens, the population, season and probably even individual variations. This will allow the statistical validation of our herein presented findings to definitively address the conundrum of phenotype-dependent venom variation.
5. Conclusion
Several colour phenotypes of common adders (Vipera berus) are known in Europe. Particularly, the melanistic specimens are subject to myth and folklore, where they are said to be of higher toxicity than normally coloured conspecifics. Here we provide the first compositional and functional determination of phenotype-dependent venom variation in common adders, using melanistic and normally coloured individuals as a model system. By rigorous implementation of chemical profiling methods (SDS-PAGE and RP-HPLC), we unveiled that venoms of both phenotypes contain fundamentally the same components yet not necessarily at the same quantities. As for functional differences, which we investigated via in vitro bioassays targeting important viperine venom activities, we detected some differences. Venoms from melanistic specimens seem to display higher protease activity and higher cytotoxicity, albeit only at a narrow concentration range. On a first glance, these results support a conceptual difference between venoms of both phenotypes. However, as they only correspond to a few factors tested and, especially for the cytotoxicity assays, are only detected at lower concentrations they are unlikely to be of clinical relevance. Considering the tremendous extent of venom variation reported in V. berus across its distribution range, the differences observed in our experiments may represent the normal biological variability within this species instead of a trait of melanistic animals. We recommend further investigations of that topic using larger sample sizes and additional assays to fully resolve this question. However, interpreting our data in light of this known venom variability, the limited range of experiments returning significant differences and the magnitude of differences measured, it seems that the reputation of MEL phenotypes is not based on human experience of envenomation and is probably an irrational superstition after all.
Acknowledgements
The authors thank Tanja Berghöfer (Fraunhofer Institute for Molecular Biology and Applied Ecology, Giessen) for technical assistance, Maik Damm and Ignazio Avella (Institute for Insect Biotechnology, Justus Liebig University Giessen, Giessen) for providing insightful comments on a preliminary version of the manuscript, and Eva Böttcher-Friebertshäuser (Institut of Virology, Philipps University, Marburg) for providing the MDCKII cells.
Contributor Information
Lennart Schulte, Email: lennart.schulte@ime.fraunhofer.de.
Lilien Uhrig, Email: lilien.uhrig@ime.fraunhofer.de.
Johanna Eichberg, Email: Johanna.Eichberg@ime.fraunhofer.de.
Michael Schwartze, Email: michaschwartze@t-online.de.
Ingve Auth, Email: ingve.auth@gmail.com.
Miriam Schulz, Email: miriam.schulz@posteo.de.
Thomas Lindner, Email: thomas.tl.lindner@outlook.de.
Paul Hien, Email: kaltblut.video@googlemail.com.
Kornelia Hardes, Email: kornelia.hardes@ime.fraunhofer.de.
Andreas Vilcinskas, Email: andreas.vilcinskas@ime.fraunhofer.de.
Tim Lüddecke, Email: tim.lueddecke@ime.fraunhofer.de.
Ethics
This work did not require ethical approval from a human subject or animal welfare committee.
Data accessibility
All relevant data have been deposited on Dryad [51].
Supplementary material is available online [52].
Declaration of AI use
We have not used AI-assisted technologies in creating this article.
Authors’ contributions
L.S.: data curation, formal analysis, investigation, methodology, visualization, writing—original draft; L.U.: data curation, formal analysis, investigation, methodology, writing—original draft; J.E.: investigation, methodology, writing—review and editing; M.S.: investigation, methodology, writing—review and editing; I.A.: investigation, methodology, writing—review and editing; M.S.: investigation, methodology, writing—review and editing; T.L.: investigation, methodology, writing—review and editing; P.H.: investigation, methodology, writing—review and editing; K.H.: data curation, formal analysis, resources, supervision, writing—review and editing; A.V.: conceptualization, project administration, resources, supervision, writing—review and editing; T.L.: conceptualization, formal analysis, funding acquisition, project administration, resources, supervision, writing—original draft.
All authors gave final approval for publication and agreed to be held accountable for the work performed therein.
Conflict of interest declaration
We declare we have no competing interests.
References
- 1. Casewell NR, Wüster W, Vonk FJ, Harrison RA, Fry BG. 2013. Complex cocktails: the evolutionary novelty of venoms. Trends Ecol. Evol. 28 , 219–229. ( 10.1016/j.tree.2012.10.020) [DOI] [PubMed] [Google Scholar]
- 2. Fry BG, et al. 2009. The toxicogenomic multiverse: convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics Hum. Genet. 10 , 483–511. ( 10.1146/annurev.genom.9.081307.164356) [DOI] [PubMed] [Google Scholar]
- 3. Schendel V, Rash LD, Jenner RA, Undheim EAB. 2019. The diversity of venom: the importance of behavior and venom system morphology in understanding its ecology and evolution. Toxins 11 , 666. ( 10.3390/toxins11110666) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Barlow A, Pook CE, Harrison RA, Wüster W. 2009. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. R. Soc. B 276 , 2443–2449. ( 10.1098/rspb.2009.0048) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Daltry JC, Wüster W, Thorpe RS. 1996. Diet and snake venom evolution. Nature 379 , 537–540. ( 10.1038/379537a0) [DOI] [PubMed] [Google Scholar]
- 6. Holding ML, et al. 2021. Phylogenetically diverse diets favor more complex venoms in North American pitvipers. Proc. Natl Acad. Sci. USA 118 , e2015579118. ( 10.1073/pnas.2015579118) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Kazandjian TD, et al. 2021. Convergent evolution of pain-inducing defensive venom components in spitting cobras. Science 371 , 386–390. ( 10.1126/science.abb9303) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Damm M, Hempel BF, Süssmuth RD. 2021. Old World vipers—a review about snake venom proteomics of Viperinae and their variations. Toxins 13 , 427. ( 10.3390/toxins13060427) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Casewell NR, Jackson TNW, Laustsen AH, Sunagar K. 2020. Causes and consequences of snake venom variation. Trends Pharmacol. Sci. 41 , 570–581. ( 10.1016/j.tips.2020.05.006) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Menezes MC, Furtado MF, Travaglia-Cardoso SR, Camargo ACM, Serrano SMT. 2006. Sex-based individual variation of snake venom proteome among eighteen Bothrops jararaca siblings. Toxicon 47 , 304–312. ( 10.1016/j.toxicon.2005.11.007) [DOI] [PubMed] [Google Scholar]
- 11. Cipriani V, et al. 2017. Correlation between ontogenetic dietary shifts and venom variation in Australian brown snakes (Pseudonaja). Comp. Biochem. Physiol. C 197 , 53–60. ( 10.1016/j.cbpc.2017.04.007) [DOI] [PubMed] [Google Scholar]
- 12. Zancolli G, et al. 2019. When one phenotype is not enough: divergent evolutionary trajectories govern venom variation in a widespread rattlesnake species. Proc. R. Soc. B 286 , 20182735. ( 10.1098/rspb.2018.2735) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Chippaux JP. 2017. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 23 , 38. ( 10.1186/s40409-017-0127-6) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Gutiérrez JM, Calvete JJ, Habib AG, Harrison RA, Williams DJ, Warrell DA. 2017. Snakebite envenoming. Nat. Rev. Dis. Primers 3 , 1–21. ( 10.1038/nrdp.2017.63) [DOI] [PubMed] [Google Scholar]
- 15. Kasturiratne A, Wickremasinghe AR, de Silva N, Gunawardena NK, Pathmeswaran A, Premaratna R, Savioli L, Lalloo DG, de Silva HJ. 2008. The global burden of snakebite: a literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 5 , e218. ( 10.1371/journal.pmed.0050218) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Geniez P. 2018. Snakes of Europe, North Africa and the Middle East: a photographic guide. Princeton, NJ: Princeton University Press. [Google Scholar]
- 17. Oskyrko O, Mi C, Meiri S, Du W. 2024. ReptTraits: a comprehensive dataset of ecological traits in reptiles. Sci. Data 11 , 243. ( 10.1038/s41597-024-03079-5) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Speybroeck J, Beukema W, Bok B, Voort JVD. 2016. Field guide to the amphibians and reptiles of Britain and Europe. London, UK: Bloomsbury Wildlife. [Google Scholar]
- 19. Speybroeck J, et al. 2020. Species list of the European herpetofauna—2020 update by the Taxonomic Committee of the Societas Europaea Herpetologica. Amphib.-Reptil. 41 , 139–189. ( 10.1163/15685381-bja10010) [DOI] [Google Scholar]
- 20. Mallow D, Ludwig D, Nilson G. 2003. True vipers: natural history and toxinology of Old World vipers. Malabar, FL: Krieger Publishing Company. [Google Scholar]
- 21. Otte N, Bohle D, Thiesmeier B. 2020. Die Kreuzotter—ein Leben in ziemlich festen Bahnen, 2nd edn. Bielefeld, Germany: Laurenti-Verlag. [Google Scholar]
- 22. Samsonov S, Grudinskaya V, Grabovsky A, Makarova T, Shitikov D. 2022. A hidden threat in abandoned fields: frequent nest predation by common adder on ground-nesting passerines. Eur. J. Wildl. Res. 68 , 11. ( 10.1007/s10344-022-01559-w) [DOI] [Google Scholar]
- 23. Forsman A. 1995. Heating rates and body temperature variation in melanistic and zigzag Vipera berus: does colour make a difference? Ann. Zool. Fenn. 32 , 365–374. [Google Scholar]
- 24. Martínez-Freiría F, Pérez i de Lanuza G, Pimenta AA, Pinto T, Santos X. 2017. Aposematism and crypsis are not enough to explain dorsal polymorphism in the Iberian adder. Acta Oecol. 85 , 165–173. ( 10.1016/j.actao.2017.11.003) [DOI] [Google Scholar]
- 25. Martínez-Freiría F, Toyama KS, Freitas I, Kaliontzopoulou A. 2020. Thermal melanism explains macroevolutionary variation of dorsal pigmentation in Eurasian vipers. Sci. Rep. 10 , 16122. ( 10.1038/s41598-020-72871-1) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Valkonen J, Niskanen M, Björklund M, Mappes J. 2011. Disruption or aposematism? Significance of dorsal zigzag pattern of European vipers. Evol. Ecol. 25 , 1047–1063. ( 10.1007/s10682-011-9463-0) [DOI] [Google Scholar]
- 27. Wüster W, et al. 2004. Do aposematism and Batesian mimicry require bright colours? A test, using European viper markings. Proc. R. Soc. Lond. B 271 , 2495–2499. ( 10.1098/rspb.2004.2894) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Madsen T, Stille B, Ujvari B, Bauwens D, Endler JA. 2022. Negative frequency-dependent selection on polymorphic color morphs in adders. Curr. Biol. 32 , 3385–3388.( 10.1016/j.cub.2022.05.060) [DOI] [PubMed] [Google Scholar]
- 29. Andrén C, Nilson G. 1981. Reproductive success and risk of predation in normal and melanistic colour morphs of the adder, Vipera berus. Biol. J. Linnean Soc. 15 , 235–246. ( 10.1111/j.1095-8312.1981.tb00761.x) [DOI] [Google Scholar]
- 30. Di Nicola MR, Pontara A, Kass GEN, Kramer NI, Avella I, Pampena R, Mercuri SR, Dorne JLCM, Paolino G. 2021. Vipers of major clinical relevance in Europe: taxonomy, venom composition, toxicology and clinical management of human bites. Toxicology 453 , 152724. ( 10.1016/j.tox.2021.152724) [DOI] [PubMed] [Google Scholar]
- 31. Hermansen MN, Krug AH, Tjønnfjord E, Brabrand M. 2019. Envenomation by the common European adder (Vipera berus): a case series of 219 patients. Eur. J. Emerg. Med. 26 , 362–365. ( 10.1097/MEJ.0000000000000577) [DOI] [PubMed] [Google Scholar]
- 32. Paolino G, et al. 2020. Vipera snakebite in Europe: a systematic review of a neglected disease. J. Eur. Acad. Dermatol. Venereol. 34 , 2247–2260. ( 10.1111/jdv.16722) [DOI] [PubMed] [Google Scholar]
- 33. Al-Shekhadat RI, Lopushanskaya KS, Segura Á, Gutiérrez JM, Calvete JJ, Pla D. 2019. Vipera berus berus venom from Russia: venomics, bioactivities and preclinical assessment of microgen antivenom. Toxins 11 , 90. ( 10.3390/toxins11020090) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Bocian A, Urbanik M, Hus K, Łyskowski A, Petrilla V, Andrejčáková Z, Petrillová M, Legath J. 2016. Proteome and peptidome of Vipera berus berus venom. Molecules 21 , 1398. ( 10.3390/molecules21101398) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Latinović Z, et al. 2016. Venomics of Vipera berus berus to explain differences in pathology elicited by Vipera ammodytes ammodytes envenomation: therapeutic implications. J. Proteomics 146 , 34–47. ( 10.1016/j.jprot.2016.06.020) [DOI] [PubMed] [Google Scholar]
- 36. Damm M, Kariş M, Petras D, Nalbantsoy A, Göçmen B, Süssmuth RD. 2024. Venomics and peptidomics of palearctic vipers: a clade-wide analysis of seven taxa of the genera Vipera, Montivipera, Macrovipera, and Daboia across Türkiye. J. Proteome Res. 23 , 3524–3541. ( 10.1021/acs.jproteome.4c00171) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Blum I. 1888. Die Kreuzotter und ihre Verbreitung in Deutschland. Frankfurt am Main. ( 10.5962/bhl.title.16086) [DOI] [Google Scholar]
- 38. Byghan Y. 2020. Sacred and mythological animals: a worldwide taxonomy. Jefferson, NC: McFarland. [Google Scholar]
- 39. Avella I, Damm M, Freitas I, Wüster W, Lucchini N, Zuazo Ó, Süssmuth RD, Martínez-Freiría F. 2023. One size fits all—venomics of the Iberian adder (Vipera seoanei, Lataste 1878) reveals low levels of venom variation across its distributional range. Toxins 15 , 371. ( 10.3390/toxins15060371) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. da Silva R, Fontes M, Rodrigues R, Bruder E, Stein M, Sipoli G, Pinhão R, Lopes C. 1999. A report on a case of melanism in a specimen of Crotalus durissus terrificus (Laurenti, 1768). J. Venom. Anim. Toxins 5 , 91–97. ( 10.1590/S0104-79301999000100008) [DOI] [Google Scholar]
- 41. Schulte L, et al. 2023. Venomics of the milos viper (Macrovipera schweizeri) unveils patterns of venom composition and exochemistry across blunt-nosed viper venoms. Front. Mol. Biosci. 10 , 1254058. ( 10.3389/fmolb.2023.1254058) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Hurka S, Lüddecke T, Paas A, Dersch L, Schulte L, Eichberg J, Hardes K, Brinkrolf K, Vilcinskas A. 2022. Bioactivity profiling of in silico predicted linear toxins from the ants Myrmica rubra and Myrmica ruginodis. Toxins 14 , 846. ( 10.3390/toxins14120846) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Sæbø IP, Bjørås M, Franzyk H, Helgesen E, Booth JA. 2023. Optimization of the hemolysis assay for the assessment of cytotoxicity. Int. J. Mol. Sci. 24 , 2914. ( 10.3390/ijms24032914) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Malina T, et al. 2017. Individual variability of venom from the European adder (Vipera berus berus) from one locality in eastern Hungary. Toxicon 135 , 59–70. ( 10.1016/j.toxicon.2017.06.004) [DOI] [PubMed] [Google Scholar]
- 45. Calderón L, Lomonte B, Gutiérrez JM, Tarkowski A, Hanson LA. 1993. Biological and biochemical activities of Vipera berus (European viper) venom. Toxicon 31 , 743–753. ( 10.1016/0041-0101(93)90380-2) [DOI] [PubMed] [Google Scholar]
- 46. Maievskyi O, Slіeptsova I. 2023. Histological changes in the rat’s jejunum wall under the conditions of action of common European adder (Vipera berus berus) venom. Agroecol. J. 84–91. ( 10.33730/2077-4893.1.2023.276732) [DOI] [Google Scholar]
- 47. Adamude FA, Dingwoke EJ, Abubakar MS, Mohamed G, Klein A, Sallau AB. 2023. Comparative venom toxin analyses of Nigerian Viperidae and Elapidae snakes. Sci. Afr. 20 , e01622. ( 10.1016/j.sciaf.2023.e01622) [DOI] [Google Scholar]
- 48. Avella I, Calvete JJ, Sanz L, Wüster W, Licata F, Quesada-Bernat S, Rodríguez Y, Martínez-Freiría F. 2022. Interpopulational variation and ontogenetic shift in the venom composition of Lataste’s viper (Vipera latastei, Boscá 1878) from northern Portugal. J. Proteomics 263 , 104613. ( 10.1016/j.jprot.2022.104613) [DOI] [PubMed] [Google Scholar]
- 49. Casewell NR, Harrison RA, Wüster W, Wagstaff SC. 2009. Comparative venom gland transcriptome surveys of the saw-scaled vipers (Viperidae: Echis) reveal substantial intra-family gene diversity and novel venom transcripts. BMC Genomics 10 , 564. ( 10.1186/1471-2164-10-564) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. op den Brouw B, Coimbra FCP, Casewell NR, Ali SA, Vonk FJ, Fry BG. 2021. A genus-wide bioactivity analysis of Daboia (Viperinae: Viperidae) viper venoms reveals widespread variation in haemotoxic properties. Int. J. Mol. Sci. 22 , 13486. ( 10.3390/ijms222413486) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Dryad . 2024. Data from: Comparative venom analysis between melanistic and normally-colored phenotypes of the common adder (Vipera berus). Dryad. ( 10.5061/dryad.v6wwpzh3x) [DOI]
- 52. Schulte L, Uhrig L, Eichberg J, Schwartze M, Auth I, Schulz Met al. Data from: Comparative venom analysis between melanistic and normally-colored phenotypes of the common adder (Vipera berus). Figshare. ( 10.6084/m9.figshare.c.7428278) [DOI]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data have been deposited on Dryad [51].
Supplementary material is available online [52].