

Abstract
The intestinal mucosa is lined by a single layer of epithelial cells that forms a dynamic barrier allowing paracellular transport of nutrients and water while preventing passage of luminal bacteria and exogenous substances. A breach of this layer results in increased permeability to luminal contents and recruitment of immune cells, both of which are hallmarks of pathologic states in the gut including inflammatory bowel disease (IBD).
Mechanisms regulating epithelial barrier function and transepithelial migration (TEpM) of polymorphonuclear neutrophils (PMN) are incompletely understood due to the lack of experimental in vivo methods allowing quantitative analyses. Here, we describe a robust murine experimental model that employs an exteriorized intestinal segment of either ileum or proximal colon. The exteriorized intestinal loop (iLoop) is fully vascularized and offers physiological advantages over ex vivo chamber-based approaches commonly used to study permeability and PMN migration across epithelial cell monolayers.
We demonstrate two applications of this model in detail: (1) quantitative measurement of intestinal permeability through detection of fluorescence-labeled dextrans in serum after intraluminal injection, (2) quantitative assessment of migrated PMN across the intestinal epithelium into the gut lumen after intraluminal introduction of chemoattractants. We demonstrate feasibility of this model and provide results utilizing the iLoop in mice lacking the epithelial tight junction-associated protein JAM-A compared to controls. JAM-A has been shown to regulate epithelial barrier function as well as PMN TEpM during inflammatory responses. Our results using the iLoop confirm previous studies and highlight the importance of JAM-A in regulation of intestinal permeability and PMN TEpM in vivo during homeostasis and disease.
The iLoop model provides a highly standardized method for reproducible in vivo studies of intestinal homeostasis and inflammation and will significantly enhance understanding of intestinal barrier function and mucosal inflammation in diseases such as IBD.
Introduction
The intestinal mucosa encompasses a single layer of columnar intestinal epithelial cells (IECs), underlying lamina propria immune cells and the muscularis mucosae. Besides its role in the absorption of nutrients, the intestinal epithelium is a physical barrier that protects the body interior from luminal commensal bacteria, pathogens, and dietary antigens. In addition, lECs and lamina propria immune cells coordinate the immune response inducing either tolerance or response depending on the context and stimuli. It has been reported that the disruption of the epithelial barrier can precede the onset of pathologic mucosal inflammation and contribute to inflammatory bowel disease (IBD) encompassing both ulcerative colitis and Crohn's disease1 , 2 , 3 , 4 , 5 , 6 , 7 . Individuals with ulcerative colitis present excessive transepithelial migration (TEpM) of polymorphonuclear neutrophils (PMN) forming crypt abscesses, a finding that has been associated with severity of disease8 , 9 . Although compromised epithelial barrier function and excessive immune responses are hallmarks of IBD, there is a lack of experimental in vivo assays to perform quantitative assessments of intestinal permeability and immune cell recruitment into the intestinal mucosa.
The most common methods used to study intestinal epithelial permeability and PMN TEpM employ ex vivo chamber-based approaches using IEC monolayers cultured on semipermeable porous membrane inserts10, 11 , 12 . The epithelial barrier integrity is monitored either by measurements of transepithelial electrical resistance (TEER) or the paracellular flux of the Fluorescein isothiocyanate (FITC)-labeled dextran from apical to basal compartment13 , 14 , 15 . Similarly, PMN TEpM is typically studied in response to a chemoattractant that is added in the lower chamber16 . PMN are placed in the upper chamber and after an incubation period, PMN that have migrated into the basal compartment are collected and quantified. While these methods are useful, easy to perform and very reproducible, they are obviously reductionist approaches and do not necessarily represent an accurate reflection of in vivo conditions.
In mice, a common assay to study intestinal paracellular permeability is by oral gavage of FITC-dextran and subsequent measurement of FITC-dextran appearance in the blood serum13 , 17 . The disadvantage of this assay is that it represents an assessment of overall barrier integrity of the gastrointestinal tract rather than that of regional intestinal contributions. In addition, Evans blue is commonly used to evaluate vascular leakage in vivo18 and has also been employed to evaluate intestinal mucosal permeability in mouse and rat19 , 20 , 21 . The quantification of Evans blue in the intestinal mucosa requires extraction from tissue employing incubation in formamide overnight. Therefore, the same tissue cannot be used to study intestinal epithelial permeability and neutrophil infiltration.
Here we highlight a simple protocol that reduces the number of animals needed to collect reproducible data on colonic mucosal permeability and leukocyte transepithelial migration in vivo. We, therefore, recommend the use of FITC-dextrans that are easily detectable in blood serum without compromising the integrity of intestinal loops which can be harvested for further analysis. Of note, the intestinal ligated loops have been used in various species (including mouse, rat, rabbit, calf) to study bacterial infection (such as Salmonella, Listeria monocytogenes and Escherichia coli)22 , 23 , 24 , 25 as well as intestinal permeability26 ; however, to the best of our knowledge there are no studies investigating mechanisms of PMN TEpM in specific regions in the intestine such as ileum or colon that are commonly involved in IBD.
Here we describe the mouse intestinal loop (iLoop) model that is a robust and reliable microsurgical in vivo method that employs a well-vascularized and exteriorized intestinal segment of either the ileum or proximal colon. The iLoop model is physiologically relevant and allows the assessment of intestinal barrier integrity and PMN TEpM on living mice under anesthesia. We demonstrate two applications: 1) quantification of serum levels of 4 kDa FITC-dextran after intraluminal administration in the iLoop 2) quantification of transmigrated PMN in the iLoop lumen after intraluminal injection of the potent chemottractant Leukotriene B4 (LTB4)27 . Moreover, utilizing the iLoop model with Jam-a-null mice or mice harboring selective loss of JAM-A on IECs (Villin-cre;Jam-a fl/fl) compared to control mice, we are able to corroborate previous studies that have reported a major contribution for tight junction-associated protein JAM-A to intestinal permeability and neutrophil transmigration15 , 28 , 29 , 30 , 31 .
The iLoop model is a highly functional and physiological method that can be used to corroborate in vitro assays. Furthermore, this is a versatile experimental model that allows the study of various reagents that can be injected into the loop lumen, including chemokines, cytokines, bacterial pathogens, toxins, antibodies and therapeutics.
Protocol
All animal experiments were conducted in accordance with the guidelines and policies of the National Institutes of Health and approved by the Institutional Animal Care & Use Committee at the University of Michigan.
1. Preoperative preparation
NOTE: This method was generated employing adult mice from C57BL/6 genetic background, aged 8 - 12 weeks. All mice were kept under strict specific pathogen free conditions with ad libitum access to normal chow and water. Results were obtained using C57BL/6, Jam-a - null mice (Jam-a−/−) or mice harboring selective loss of JAM-A on lECs (Villin-cre;Jam-afl/fl) and littermate Jam-afl/fl controls as previously described30.
- Area preparation
- Perform surgery in clean area. The intestinal Loop model, however, is a non-survival surgery that does not require aseptic/sterile technique. Observe veterinary sanitation practices and use cleaned surgical instruments (i.e., scrubbed with soap, rinsed with water followed by 70% ethanol).
- Turn on a temperature-controlled surgical board (or heating pads) and adapted light source to keep animal from hypothermia during the anesthesia and surgery.
- Prepare ligatures by cutting 6 cm segments of nonabsorbable 4-0 silk surgical sutures.
- Prepare cotton gauzes (5 cm x 5 cm) that are cut in the center following an ellipsoid shape. These will be used to cover the midline laparotomy and prevent direct contact between the exteriorized iLoop and animal fur. Soak the cut gauzes in warm Hanks' Balanced Salt Solution (HBSS) in a petri dish container.
- Prepare wet cotton swabs soaked in warm HBSS that will be used to handle organs and exteriorized iLoop.
- Prepare 10 mL syringe filled with warm HBSS and attach to a yellow feeding tube. This syringe will be used for moisturizing exposed tissues during surgery and to gently flush the iLoop of fecal content.
- Animal preparation
- Anesthetize the animal in accordance with the approved animal protocol. In this protocol a mix of isoflurane and oxygen is administered through an anesthesia vaporizer. According to manufacturer’s instructions, adjust oxygen flow rate to 1 L/min. Set the vaporizer to 5% and pre-charge the induction chamber. After 5 min, reduce isoflurane vaporizer to 2% - 2.5%.
-
Place the animal in the induction chamber for 3 min - 5 min, then transfer animal to a heated surgery board and connect an anesthesia nosecone plug. Restrain the animal in supine position by the four limbs using adhesive tape.NOTE: As an anesthesia alternative, a mixture of ketamine (80 mg/kg - 100 mg/kg) and xylazine (5 mg/kg - 10 mg/kg) diluted in saline solution (0.9% NaCl) can be administrated by intraperitoneal injection. Anesthesia should be maintained throughout the surgery by intramuscular administration of ketamine/xylazine (at 0.1 - 0.25 times of initial doses) to ensure anesthetic depth. If available, an isoflurane anesthesia vaporizer is highly recommended to assure better reproducibility, survivability and prevent animal pain.
- Apply ophthalmic ointment to both eyes to prevent corneal desiccation.
- Perform a physical exam that includes heart rate (around 500 beats/min) and rhythm, mucous membrane color (pink), capillary refill time (< 2 s), respiratory rate (not lower than 40 - 60 breaths/min), and temperature (36.5 °C)32.
-
Before proceeding to the next steps, assess anesthetic depth by pedal withdrawal reflex. Employ a painful stimulus (pinch) of the skin between the toes and/or toe pads. Mouse will respond by contracting and removing its leg. This pedal reflex disappears when animal is anesthetized deeply.NOTE: Monitoring of vitals and pedal reflex are recommended throughout anesthesia at minimum every 15 min. Institutional Animal Care and Use Committee (IACUC) guidelines for evaluation of anesthetic depth recommend monitoring of the following: (a) color of tail, foot and mucous membrane (such as tongue). Color pink as normal and pale or blue as indicative of decreased blood perfusion or respiratory distress; (b) evaluation of the breathing pattern as regular versus irregular breaths. A rectal temperature probe, rodent oximeter and heart rate monitors can be used for the assessment of body temperature, heart and respiratory rates, respectively.
2. Generation of the ileal loop
Skin preparation: Scrub fur of the abdominal midline with alcohol swabs or gauze sponge soaked with 70% Ethanol. Do not wet a wide area of fur with alcohol to prevent hypothermia.
Using scissors, perform a midline laparotomy. Make a vertical incision in the middle of the abdomen (about 2 cm in length) and expose the peritoneum. Be careful to not injure intra-abdominal organs.
Place pre-cut wet cotton gauze over the exposed intraabdominal cavity.
-
Use wet cotton swabs to mobilize and exteriorize the caecum. Carefully place the caecum on the wet cotton gauze.
NOTE: Caecum is localized in the left caudal quadrant of the abdominal cavity in a majority of mice independent of the sex of the animal.
Use wet cotton swabs to mobilize and gently exteriorize the ileum of which the terminal section (distal end) is attached to the caecum (Figure 1B).
-
Deploy at least 6 cm of terminal ileum on the wet cotton gauze without disruption of the mesenteric vessels and blood supply. Blood supply is maintained if there is no bleeding and the tissue maintains its pink color (Figure 1B).
NOTE: Avoid drying of exposed tissues by maintaining tissues moist at all times with warm HBSS (every 2 - 3 min) using 10 mL syringe attached to a yellow feeding tube (step 1.1.6).
Close to the caecum, identify the major artery supplying the ileum in the mesentery. Then locate two ligation sites in the mesentery that are free of critical blood vessels.
Using blunt tissue forceps, firmly grab the terminal ileum (closest to the caecum) and using fine tip forceps, fenestrate the mesentery avoiding blood vessels. Place silk suture across the perforation and tie a surgical knot to create the first ligation (distal end of the loop).
Use the ruler to measure 4 cm away from the first ligature and create the second ligature (proximal end of the loop) as mentioned in step 2.8 (Figure 1C).
-
With fine scissors carefully cut next to each ligation to isolate the 4 cm ileal loop, keeping intact blood supply and mesenteric membrane.
NOTE: Cut off both ends of the exteriorized segment of the iLoop, then flush gently as a necessary step that prevents interference with luminal contents (fecal matter), thus facilitating even dispersion of FITC-dextrans or chemotactic stimuli across the entire length of the isolated segment as well as allowing for more accurate quantification of leukocytes by flow cytometry. This procedure also allows uniform distension of the mucosa after injection of specified volumes of reagent and better reproducibility between animals.
Gently flush the content of the ileal loop segment with warm HBSS using a flexible yellow feeding tube attached to a 10 mL syringe (see step 1.1.6).
Ligate the two cut ends of the flushed ileal loop using silk suture.
-
Use a 1 mL syringe with 30 G needle to slowly inject 250 μL of reagent such as FITC-dextrans (step 4.2) or chemokine (step 5.3) into the intestinal lumen. The ileal loop will inflate causing a moderate distension of the mucosa (Figure 1D).
NOTE: Inject reagent into the loop lumen on the opposite side of the mesenteric artery. Be careful not to pull out the ileal loop from the animal while injecting to avoid tearing blood vessels and induce bleeding.
Using wet cotton swabs, gently put back the ileal loop, proximal ileum and caecum.
Use a needle holder, anatomical forceps and 3.0 nonabsorbable silk sutures with reverse cutting needle to close the abdominal wall.
Place the animal in a temperature-regulated anesthesia chamber for the incubation period.
Figure 1: The ileal loop model.
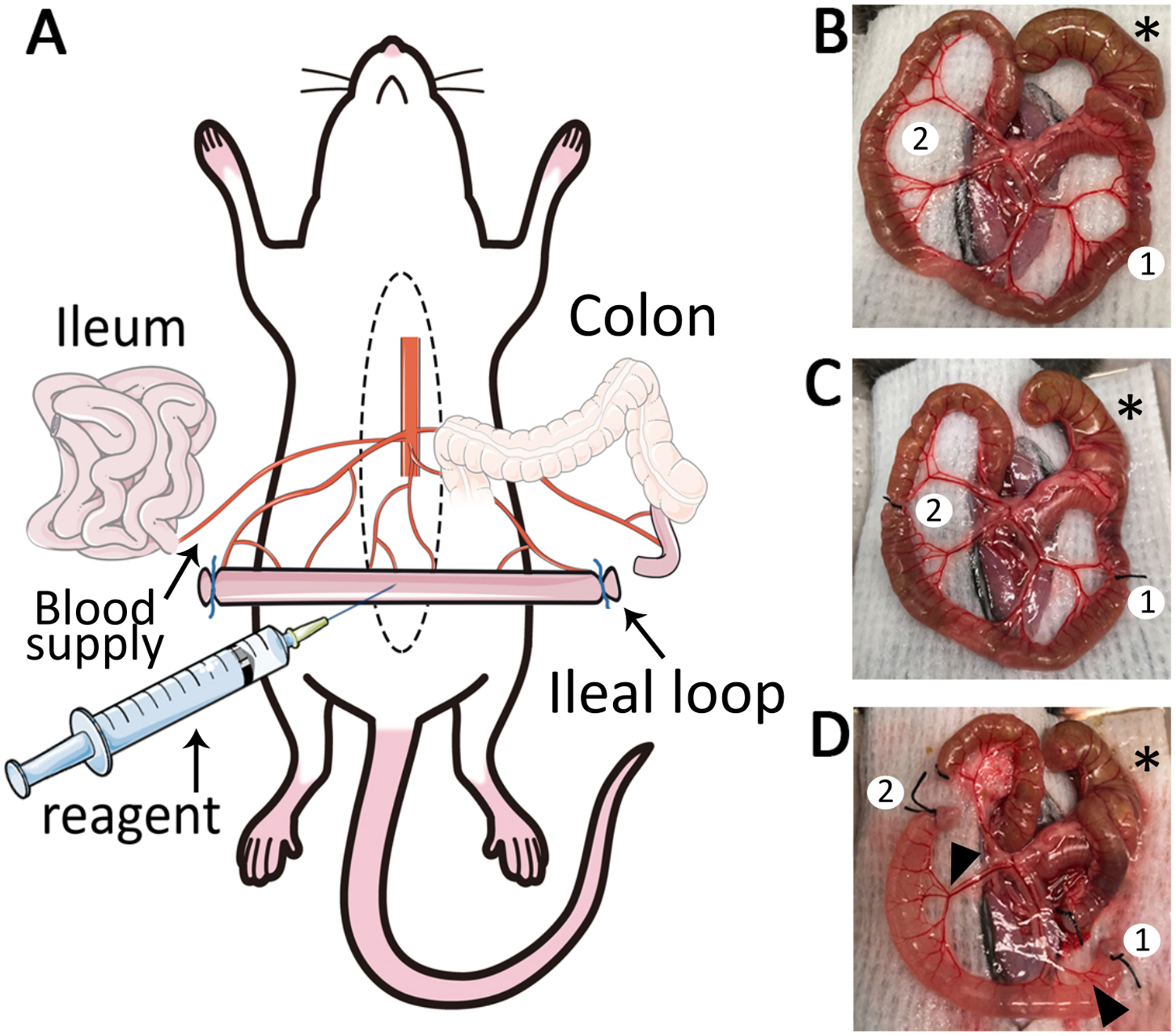
(A) Schematic overview of the ileal loop model. Median laparotomy is performed on mice under anesthesia and placed on a temperature-controlled surgery board. (B) Exteriorization of the caecum (*), ileum and mesentery. Two adequate sites for ligation are identified (1,2). (C) Isolate a segment of 4 cm length: the first ligature (1) is placed close to the ileo-caecal junction and a second ligature (2) is placed 4 cm away from the first ligature. (D) Two small incisions are made in the mesentery (1, 2) to create a 4 cm length ileal loop. After removal of luminal content and ligation of cut-ends, reagents such as fluorescent markers and chemoattractants can be injected into the lumen. The ileal loop is well vascularized (black arrowheads).
3. Generation of the proximal colon loop (pcLoop)
NOTE: For details about mice that were used for the generation of the pcLoop, see the information provided at the beginning of the protocol section.
Perform steps 2.1. - 2.4. as described above for the ileal loop.
Using wet cotton swabs, exteriorize the entire ileum and place it on the top of a wet cotton gauze. Identify the proximal colon and the blood supply located in the mesocolon. Mobilize the proximal colon and by using fine tip forceps create the first ligature in an area free of vessels in the mesocolon at about 0.5 cm distal from the caecum (Figure 2B).
Measure 2 cm from the first ligature and create a second ligature at an area free of blood supply in the mesocolon (Figure 2C).
-
Using fine scissors carefully cut next to each ligation to isolate a 2 cm long pcLoop.
NOTE: As mentioned in the note under step 2.10, it is important to cut off both ends to isolate a pcLoop that is gently cleaned of luminal contents. Carefully cut through the colonic tissue and mesocolon to prevent small vessels from bleeding into the intestinal lumen. If necessary, use thermal cautery to limit bleeding at the incision site.
Gently flush the pcLoop with warm HBSS to remove feces using a flexible yellow feeding tube attached to 10 ml_ syringe (see step 1.1.6).
Ligate the two cut ends of the flushed pcLoop using silk suture.
-
Use a 1 mL syringe with 30G needle to slowly inject 200 μL of reagent such as FITC-dextrans (step 4.2) or chemokine (step 5.3) into the intestinal lumen. The pcLoop will inflate causing a moderate distension of the mucosa (Figure 2D).
NOTE: Inject the reagent into the pcLoop lumen on the opposite side of the mesenteric artery. Ensure consistency between animals and create a 2 cm long pcLoop to ensure equal distension of the mucosa.
Utilize wet cotton swabs to gently place back the ligated pcLoop, ileum and caecum into the abdominal cavity.
Use a needle holder, anatomical forceps and 3.0 nonabsorbable silk sutures with reverse cutting needle to close the abdominal wall.
Place the animal in a temperature-regulated anesthesia chamber for the incubation period. 4
Figure 2: The proximal colon loop model.
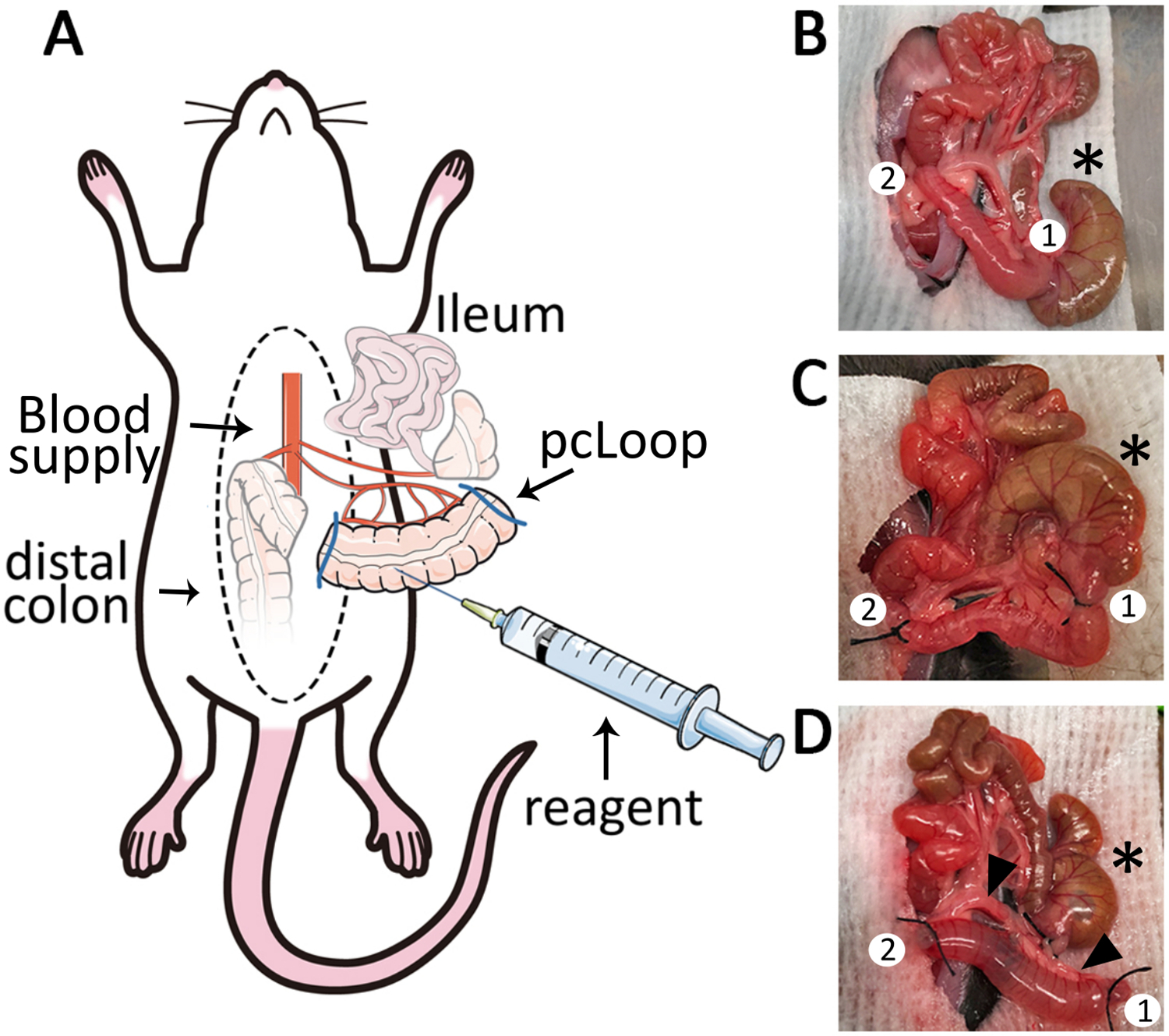
(A) Schematic overview of the pcLoop model. Median laparotomy is performed on mice under anesthesia placed on a temperature-controlled surgery board. (B) Exteriorization of the caecum (*), proximal colon, mesocolon and ileum. Two adequate sites for ligation are identified (1,2). (C) The first ligature (1) is placed close to the caecum and a second ligature (2) is placed 2 cm more distal from the first ligature. (D) The pcLoop is exteriorized, cleaned of luminal content and inflated with reagents such as fluorescent markers and chemoattractants. The pcLoop is a well-vascularized 2 cm segment of proximal colon (black arrowheads indicate blood supply).
4. Quantitative assessment of intestinal permeability: 4 kDa FITC-dextran assay
Perform an ileal loop or pcLoop (as described above).
Using a 1 mL syringe with 30 G needle, inject into the intestinal lumen either 250 μL (ileum - step 2.13) or 200 μL (colon - step 3.7) of 4 kDa FITC-dextran solution (1 mg / mL in HBSS). Keep the unused FITC-dextran solution protected from light to prepare the standard curve after serum collection.
For the ileal loop follow steps 2.14 to 2.16, for the pcLoop steps 3.8 to 3.10. Briefly, put organs and iLoop back in place into the abdominal cavity, close the abdominal wall.
Place the animal for 120 min in a heated anesthesia chamber.
-
After the incubation period open the abdominal wall, gain access to the heart and perform cardiac puncture using a 1 mL syringe with 25G needle to collect the blood. Transfer blood into a 1.3 mL serum clot activator tube, mix gently, and keep on ice protected from light. Collect at least 500 μL of blood per mouse.
NOTE: Animals are euthanized when being under anesthesia by using a physical method such as decapitation or cervical dislocation, and in accordance with the approved animal protocol.
Centrifuge serum clot activator tube for 5 min at 10,000 x g at room temperature according to manufacturer's recommendations. Collect the serum (supernatant) and transfer into a 1.7 mL centrifuge tube. Keep the tube on ice and protected from light.
- Quantification of fluorescence in blood serum
- Prepare a standard curve of FITC-dextran 4 kDa in serum of control mice (saline or HBSS is a valid alternative). Create a two-fold serial dilution with a starting concentration of 1 mg/mL FITC-dextran. FITC-dextran 4 kDa concentrations that are measured range between 0.25 mg/mL and 2 μg/mL.
- Transfer equal volume of the sample and standards to a black 96-well plate (flat bottom) and measure FITC in a fluorescence plate reader (Excitation 490 nm, Emission 520 nm), according to manufacturer's instructions and published protocols33 . Calculate the FITC concentration based on the standard curve or present permeability values as fold-change normalized to the experimental control group.
5. Quantitative assessment of migrated PMN in the intestinal lumen after intraluminal stimulation with chemokines
NOTE: Very few PMN reside in the intestinal mucosa at the baseline level. Pretreatment of animals with pro-inflammatory cytokines results in an inflammatory environment that facilitates PMN recruitment from bloodstream into the intestinal mucosa.
Using a 1 mL syringe with 30 G needle, perform intraperitoneal (i.p.) injection of a sterile solution of 100 ng of Tumor Necrosis Factor-α (TNFα) and 100 ng of Interferon-γ (INFγ) in 200 μL of Phosphate Buffered Saline (PBS).
After 4 - 24 h of pretreatment with pro-inflammatory cytokines, perform an ileal loop or pcLoop (as described above).
-
Using a 1 mL syringe with 30 G needle, inject into the intestinal lumen either 250 μL (ileum - step 2.13) or 200 μL (colon - step 3.7) of chemoattractant solution Leukotriene B4 (LTB4) 1 nM in HBSS.
NOTE: Leukotriene B4 (LTB4) is used in this protocol as a potent chemoattractant for PMN. Other chemoattractants such as N-Formylmethionyl-leucyl-phenylalanine (fMLF) or chemokine (C-X-C motif) ligand 1 (CXCL1/KC) can also be used to induce significant recruitment of PMN into the colonic lumen30 .
For the ileal loop follow steps 2.14 to 2.16. For the pcLoop follow steps 3.8 to 3.10. Briefly, put organs and iLoop back in place into the abdominal cavity, close the abdominal wall.
Place the animal for 60 min in a heated anesthesia chamber.
- Collection of the intestinal loop content
- Prepare solutions and store on ice: For the ileal loop and pcLoop, prepare a wash buffer containing 2 mM Ethylenediaminetetraacetic acid (EDTA), 5 mM Dithiothreitol (DTT) and 2% FBS in sterile PBS without calcium and magnesium.
- After the incubation period and under anesthesia maintenance, open the abdominal wall and pull out the iLoop (ileal loop or pcLoop). Euthanize animals when being under anesthesia by using a physical method such as decapitation or cervical dislocation, and in accordance with the approved animal protocol.
-
Rinse loop with cold PBS to remove any residual blood contaminant and absorb excess of PBS with tissue wipes. Carefully collect the loop content into a 1.7 mL centrifuge tube (about 250 μL for ileal loop and 200 μL for pcLoop). Flush the loop with 500 μL cold wash buffer and, immediately after collection, place the tube on ice.NOTE: DTT helps to dissolve mucus. If iLoop luminal content is very viscous (depending on the mouse genetic background), dilute it 1:2 or 1:3 with wash buffer containing DTT.
- Pass the luminal content solution through a 35 μm nylon mesh filter using a 5 mL round-bottom tube with cell strainer cap. This step helps to remove tissue fragments and cell aggregates. Rinse the cell strainer with 1 mL of wash buffer.
- Centrifuge tube at 400 x g for 5 min at 4 °C. Discard supernatant, rinse pellet with 500 μL - 1 mL wash buffer, then centrifuge at 400 x g for 5 min, 4 °C.
- Resuspend iLoop luminal cell pellet in 200 μL of flow cytometry buffer (FCB) containing 2% FBS in sterile PBS without calcium and magnesium. Cells can be kept in a tube or transferred to a 96-well round bottom plate for flow cytometry staining and analysis.
- Flow Cytometry staining and analysis
- Compensation controls: white blood cells
- Prepare a 1 mL syringe with 25 G needle prefilled with sterile 0.5 M EDTA (pH 8.0). 10% EDTA per expected blood volume (100 μL of EDTA for 1 mL blood).
-
Collect blood under anesthesia by cardiac puncture. Transfer blood into a 1.7 mL tube, then centrifuge at 400 x g for 10 min, 4 °C.NOTE: While the mouse is under anesthesia use a physical method to confirm death in accordance with the approved animal protocol (such as cervical dislocation).
- Aspirate the supernatant. Resuspend the pellet in 1 mL of Ammonium-Chloride-Potassium (ACK) lysis buffer for the lysis of red blood cells. Incubate for 3 min - 5 min on ice. Centrifuge 400 x g for 5 min, 4 °C. If the pellet is still red, repeat this ACK lysis buffer step until pellet turns white.
-
Resuspend the pellet in 1 mL FCB and plate 0.5 x 106- 1 x 106 of cells per well. Prepare five wells of a 96-well round-bottom plate. Place the plate on ice.NOTE: Use the same 96-well plate that contains the loop luminal contents (see step 5.6.6).
- Flow cytometry staining
- Centrifuge the 96-well plate for 5 min at 400 x g, 4 °C. Discard the supernatant and resuspend the pellets with 50 μL of Purified Rat Anti-Mouse CD16/CD32 as an Fc-Block (1 μg per 100 μL of FCB). Incubate for 5 min - 10 min on ice.
- Immunostaining of the iLoop luminal content: Prepare a mixture containing all fluorochrome-conjugated antibodies (1:50 dilution in FCB): anti-CD45-PerCP, anti-CD11b-PE and anti-Ly-6G-Alexa Fluor 647. Add 50 μL of the combination per well, for a final volume of 100 μL.
- Immunostaining of the white blood cells for compensations (1:50 dilution in FCB): Use 50 μL of FCB alone (unstained sample, well 1), 50 μL of each individual fluorochrome-conjugated antibody (wells 2 - 4), 50 μL of the combination of all fluorochrome-conjugated antibodies (well 5). Final volume of 100 μL.
- Incubate the plate for 30 min on ice protected from light.
- Centrifuge the plate for 5 min at 400 x g, 4 °C. Discard the supernatant and wash with 200 μL of FCB. Repeat this washing step twice.
- Add FCB 150 μL/well to the blood samples.
- Add 100 μL/well FCB to the iLoop luminal content sample. Then 50 μL/well of fluorescent counting beads.
- Flow cytometry analysis
- Gate for CD45 positive events and for the expression of Ly-6G-/Gr-1 and CD11b30 .
- Use 100 μL of the sample volume as a stop condition.
-
Calculate the absolute number of PMN that has migrated into the iLoop lumen following the information provided by the manufacturer of the fluorescent counting beads.
Representative Results
A schematic representation of the ileal loop and pcLoop models is depicted in Figure 1 and Figure 2, respectively. The anatomical pictures display the critical steps of the procedure including exteriorization of the intestinal segment (Figure 1B and Figure 2B), identification of an appropriate location for ligations that allows minimal disturbance of blood supply (Figure 1C and Figure 2C) and cleaning followed by ligation of cut ends of the iLoop that can be filled with reagent solution (Figure 1D and Figure 2D). Importantly, the iLoop model preserves vital blood supply and allows physiological absorption of applied reagents such as FITC-dextrans or the potent PMN chemoattractant LTB4. At the end of the assay, the iLoop should be inflated (as seen in Figure 1D and Figure 2D) and display normal mucosal perfusion with brightred mesenteric vessels. Depending on the assay, blood is collected to measure FITC-dextran in serum or iLoop luminal contents are processed for quantification of PMN TEpM prior to euthanizing the animal.
In order to verify the accuracy of the iLoop model for the assessment of intestinal permeability, a FITC-dextran pcLoop assay was performed to evaluate the role of TJ-associated protein JAM-A in the regulation of intestinal barrier function in vivo. Of note, it has been reported that JAM-A deficiency lead to increased epithelial intestinal permeability in vitro28 and after oral gavage in vivo29 . Herein, using the pcLoop model, a 2.5-fold increase in 4 kDa FITC-dextran serum levels was quantified in Jam-a-null mice (Jam-a−/−) compared to controls (Jam-a+/+) (Figure 3A)30 . Furthermore, similar results were obtained with mice harboring selective loss of JAM-A on IECs (Villin-cre;Jam-a fl/fl) compared to littermate controls (Jam-a fl/fl) (Figure 3B)30 . Therefore, the pcLoop model was able to corroborate previous studies that have reported a positive contribution for JAM-A to the intestinal barrier function.
Figure 3: JAM-A regulates intestinal permeability in vivo.
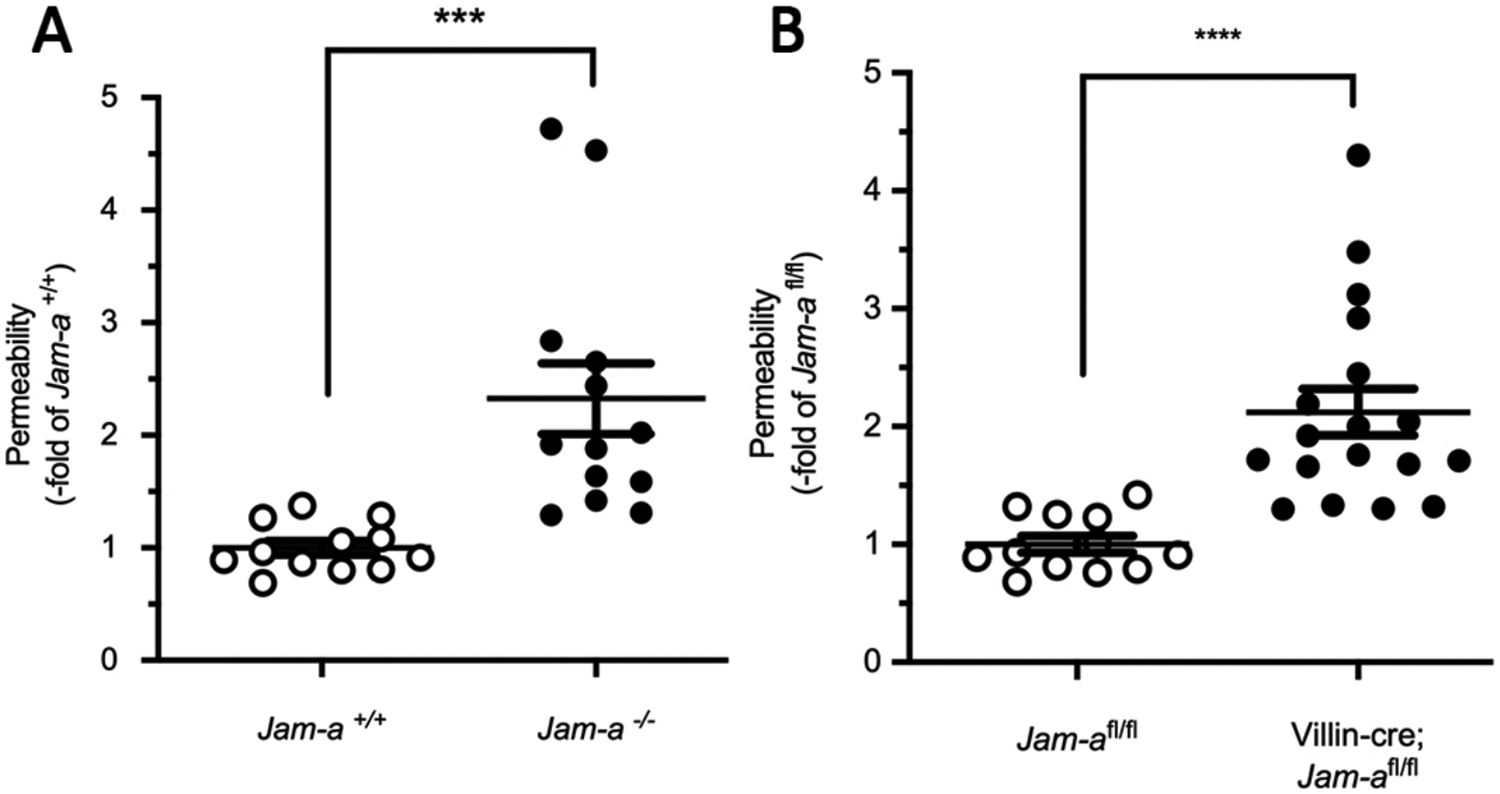
(A) JAM-A deficiency (Jam-a−/−) led to increased colonic permeability to 4 kDa FITC-dextran. Jam-a−/− (13x animals; black dots) were compared with Jam-a+/+ controls (12x animals; white dots). 4 kDa FITC-dextran (1 mg/mL) in HBSS was injected into the pcLoop lumen. Fluorescence was measured in blood serum after a 120 min incubation period. Data are expressed as means ± SEM; n = 3 independent experiments. ****P < 0.0001; Mann-Whitney U test. (B) Increased colonic permeability to 4 kDa FITC-dextran in Villin-cre; Jam-afl/fl (18x animals, black dots) compared to controls (Jam-afl/fl, 12x animals, white dots). Data are means ± SEM; n = 4 independent experiments. ****P < 0.0001; Mann-Whitney U test. This figure has been modified from Flemming S, Luissint AC et al.30.
Then pcLoop model was employed to study PMN recruitment into the intestinal mucosa and subsequent TEpM in vivo. As shown in Figure 4A, the number of PMN in the luminal content of the pcLoop was quantified by flow cytometry analysis. PMN were defined as cells positive for each of the cell-surface makers CD45, CD11b and Ly6G36 . Circulating white blood cells were used as positive control for gating strategy. As expected, the number of PMN present in a segment of proximal colon similar to the pcLoop was low under physiological conditions (Figure 4B). Pretreatment with pro-inflammatory cytokines TNFα and IFNγ prior to surgery resulted in augmented numbers of PMN recruited in the pcLoop lumen. The administration of the PMN chemoattractant LTB4 led to a dramatic increase in PMN counts supporting a LTB4-dependent PMN recruitment (Figure 4B). Immunohistochemical staining of PMN in the colonic mucosa corroborate the elevated recruitment of PMN following stimulation with cytokines and LTB4 when compared with cytokine treatment without LTB4 (Figure 4C)30 . The pcLoop model was employed to study the contribution of JAM-A to PMN TEpM by using Villin-cre;Jam-a fl/fl mice. Loss of epithelial JAM-A led to a reduced number of transmigrated PMN in the colonic lumen compared to littermate controls (Figure 4D)30 . These findings strongly support a role for JAM-A in facilitating PMN migration across the intestinal epithelium and provide complementary insights to studies that have reported the involvement of JAM-A in PMN migration across vascular endothelium in various models of inflammation31 , 37 , 38 .
Figure 4: JAM-A promotes LTB4-dependent recruitment of PMN into the lumen of the pcLoop.
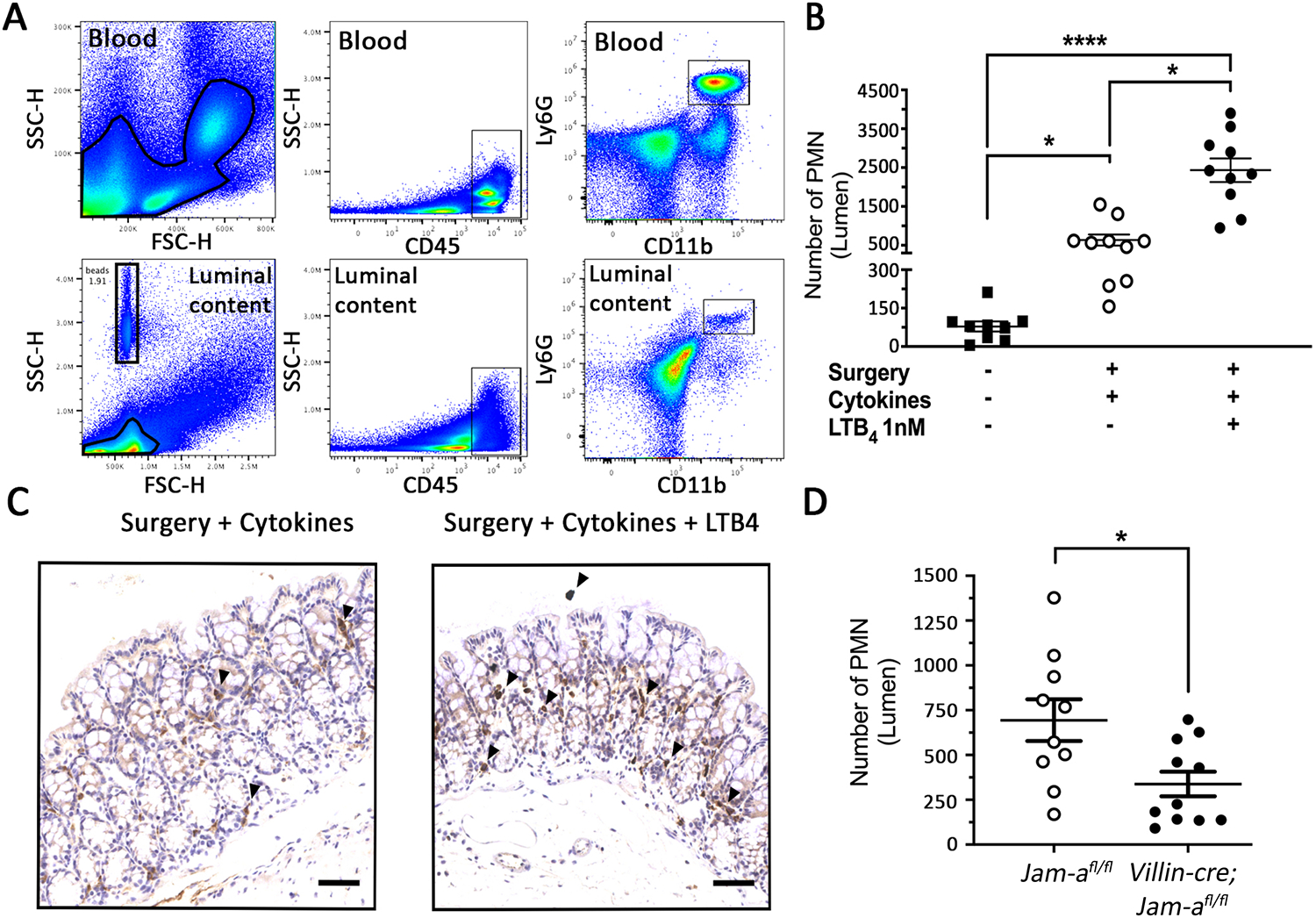
(A) Gating strategy to quantify PMN (CD45+, CD11b+, and Ly-6G/Gr1+ cells) in luminal content by flow cytometry with fluorescent counting beads. Leukocytes from blood samples were used as a positive control for the gating strategy. (B) Number of PMN recruited into the pcLoop lumen after cytokine (TNFα+IFNγ, 100ng each) treatment (10x animals; white dots) or after a combination of cytokines and 1 nM LTB4 (10x animals; black dots). Black squares represent the number of PMN at baseline as assessed in an intact colonic segment identical in length to the pcLoop that was not subjected to any surgery or treatment with proinflammatory cytokines and LTB4 (9x animals). Data are the mean ± SEM (n = 3 independent experiments), Kruskal-Wallis test with Dunn's multiple comparison test. *P < 0.05, ****P < 0.0001. (C) Immunohistochemical staining of PMN (anti-Ly6G/Gr1 antibody) in the epithelium of the pcLoop after treatment with cytokines alone (left panel, TNFα+IFNγ) or a combination of cytokines and LTB4 (right panel). The number of PMN recruited in the pcLoop is increased in the presence of LTB4 (black arrowheads). Scale bar: 100 μm. (D) Number of PMN recruited in the pcLoop lumen in Villin-cre; Jam-afl/fl mice (11x animals; black dots) compared to Jam-afl/fl mice (10x animals; white dots) in response to 1 nM LTB4. Data are means ± SEM; n = 3 independent experiments. *P < 0.05; 2-tailed Student's t test. This figure has been modified from Flemming S, Luissint AC et al.30.
Discussion
The mechanisms responsible for dysregulation of intestinal barrier function and immune cell recruitment under pathologic conditions such as IBD are incompletely understood due to a lack of experimental in vivo methods. Here, we detail a robust in vivo murine model that employs a well-vascularized exteriorized intestinal segment of either ileum or proximal colon and allows for assessment of intestinal permeability, neutrophil migration studies as well as other applications.
The iLoop is a non-recovery surgery that is performed on live animals. Anesthesia must be continuously monitored over the course of the experiment and evaluation of depth of sedation is mandatory. The most critical steps include (1) the isolation of the iLoop, (2) ligation of cut ends and, (3) the inflation of the iLoop by intraluminal injection of reagent solution. In each of these steps, bleeding can occur, compromising the blood supply of the iLoop and affecting accuracy of the results. Of note, in rare instances of intraluminal bleeding during the PMN TEpM assay, the flow cytometry gating strategy presented here will help to distinguish transmigrated PMN from PMN originating directly from the bloodstream (non-migrated PMN). Transmigrated PMN collected in the iLoop lumen express high levels of surface marker CD11b10 compared to circulating PMN (Figure 4D).
Given that the iLoop allows quantitative analyses of intestinal permeability and migration of blood PMN into the intestinal lumen, it is important to standardize the size of the absorbing mucosal area and the blood supply. In order to ensure consistency between animals, it is essential that a correct length of intestinal segment is exteriorized. The iLoop should be 4 cm for the ileal loop and 2 cm for the pcLoop and be perfused by comparable blood supply. Inconsistency in these parameters will also result in unequal distension of the iLoop after intraluminal injection of reagents and augment variability inter and between experimental groups. Furthermore, to avoid over-distension of the iLoop, we recommend that no more than 250 μL of reagent solution be injected in the lumen for the ileal loop and 200 μL for the pcLoop, respectively.
There are some limitations inherent with the nature of the procedure. The iLoop is a non-recovery surgery that is performed on live animals. This is a technically challenging microsurgical method; however, personnel can acquire surgical skills through practice. The average duration of the surgery should be short (maximum 15 min). We recommend 120 min as an ideal incubation time for measuring intestinal permeability and 60 min for the PMN TEpM. Incubation times can be reduced, but extended timepoints might affect the overall inflammatory state of the animal under anesthesia. In addition, the protocol from the start of the surgical procedure to sample collection / analysis cannot be paused.
This iLoop model presents key advantages with respect to existing methods: (1) the iLoop is fully vascularized and is more physiologically relevant, (2) in contrast to the oral gavage method that assesses the overall gastrointestinal tract integrity and depends on gastrointestinal motility13 , the iLoop allows to study the properties of specific localized areas in the intestine (terminal ileum or proximal colon) that are commonly involved in IBD, (3) the iLoop is the first in vivo model that allows the quantitative study of PMN TEpM into the gut lumen as well as other parts of the intestinal mucosa, including lamina propria and epithelial factions30 , 35 . It is possible to employ high versus low molecular weight FITC-labeled dextrans (4 to 150 kDa) to evaluate both size selectivity and/or severity of epithelial barrier defects in knockout/knock-in mice or various experimental models including, but not limited to, intestinal inflammation. In addition FITC-labeled dextrans can be quantified in other organs such as the liver39 or as a novel approach for studies of the blood brain barrier providing insights into the role of intestinal permeability in gut-liver and gut-brain axes40 , 41 , 42 . Furthermore, this method offers the possibility to perform two loops in parallel (ileal loop and pcLoop in the same animal) and instill two different fluorescent labeled probes for analyses of barrier properties in distinct areas in the intestine. Along similar lines, generation of two loops in parallel can be employed to specifically evaluate ileum versus colon for differences or similarities in recruitment of immune cells in response to the same reagent.
Here, by using the pcLoop with Jam-a-null mice or mice harboring selective loss of JAM-A on IECs (Villin-cre;Jam-a fl/fl), we corroborate findings from previous studies that have reported a positive role for the TJ-associated protein JAM-A in intestinal permeability and PMN TEpM. The applications of the iLoop can be expanded to various reagents including antibodies, microbial pathogens and therapeutic drugs30 , 34 , 35 . Of note, we used LTB4 (336.5 Da) to model PMN TEpM given that it is a well-accepted potent and physiologic PMN chemoattractant and its ability to induce TEpM at low concentrations (1 nM) in the physiologic range. However, our loop model is adaptable to other relevant chemoattractants. We have reported the use of the bacterial peptide N-formyl-methionyl-leucyl-phenylalanine (fMLF) to induce significant recruitment of PMN into the colonic lumen30 . fMLF (437.5 Da) is a lower affinity chemoattractant in mice which requires much higher concentrations to be effective (1μM). This model is adaptable for use of CXCL1/KC, another potent physiologic chemoattractant that we have successfully used, yet CXCL1/KC is expensive and a relatively large molecule (11 kDa) that is less efficient in crossing the epithelial barrier. We have also demonstrated that neutralizing antibodies against leukocytespecific integrin CD11b/CD18 (αMβ2) that were injected into the loop lumen prior administration of chemoattractant LTB4 resulted in reduced PMN TEpM corroborating results from in vitro studies10 , 30 , 35 . Furthermore, the pcLoop was recently employed to study the effect of PMN versus epithelial glycans in controlling the rate of PMN TEpM43 . Reagents were injected into the pcLoop lumen prior administration of chemoattractant LTB4. Therefore, with its broad spectrum of applications, the iLoop can complement and confirm findings obtained via in vitro assays. Ligated intestinal loops have also been used by others to study bacterial infection (such as Salmonella, L. monocytogenes and E. coli), therefore we believe that the ease in adaptability of this iLoop model can be used for these studies as well.
Following treatment with pro-inflammatory immune mediators, the iLoop can be used as an acute model of intestinal inflammation. Furthermore, the iLoop may enable studies elucidating the link between increased intestinal permeability and immune cell recruitment after exposure to intraluminal pathogens or in chronic inflammatory experimental models. Of note, we have recently observed by employing the pcLoop model that in response to high dose of proinflammatory cytokines TNFα and IFNγ (1 mg of each) intestinal paracellular permeability to 4 kDa FITC-dextran resulted in enhanced PMN recruitment into the pcLoop lumen in response to LTB4 in comparison to low dose cytokines (100ng of each)30 . Interestingly, here we show that increased epithelial permeability secondary to Jam-a deficiency did not lead to enhanced PMN TEpM but diminished it. All together these results suggest that intestinal paracellular permeability affects the rate of PMN TEpM but the correlation is not direct and depends on factors such as the expression of adhesion molecules (similar to JAM-A) that play an important role in both epithelial barrier function and leukocyte migration16 . Future studies are needed to investigate the fine tuning of immune cell responses by the intestinal epithelium, and contributions to pathologic mucosal inflammation such as inflammatory bowel disease.
In conclusion, the iLoop model provides a major improvement to existing approaches for the assessment of intestinal permeability and PMN TEpM in vivo that will significantly aid in understanding mechanisms underlying the regulation of intestinal inflammation and IBD.
Acknowledgments
The authors thank Dr. Sven Flemming of the University of Wuerzburg for his contributions to the establishment of the proximal colon loop model, Sean Watson for the management of the mouse colonies and Chithra K. Muraleedharan for helping with the acquisition of the pictures of the iLoop model. This work was supported by the German Research Foundation/DFG (BO 5776/2-1) to KB, R01DK079392, R01DK072564, and R01DK061379 to C.A.P.
Footnotes
A complete version of this article that includes the video component is available at http://dx.doi.org/10.3791/62093.
Disclosures
The authors have nothing to disclose.
References
- 1.Olson TS et al. The primary defect in experimental ileitis originates from a nonhematopoietic source. Journal of Experimental Medicine. 203 (3), 541–552 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jump RL, Levine AD Mechanisms of natural tolerance in the intestine: implications for inflammatory bowel disease. Inflammatory Bowel Diseases. 10 (4), 462–478 (2004). [DOI] [PubMed] [Google Scholar]
- 3.Peeters M. et al. Clustering of increased small intestinal permeability in families with Crohn's disease. Gastroenterology. 113 (3), 802–807 (1997). [DOI] [PubMed] [Google Scholar]
- 4.Michielan A, D'lnca R Intestinal permeability in inflammatory bowel disease: Pathogenesis, clinical evaluation, and therapy of leaky gut. Mediators of Inflammation. 2015 628157 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chin AC, Parkos CA Neutrophil transepithelial migration and epithelial barrier function in IBD: potential targets for inhibiting neutrophil trafficking. Annals of the New York Academy of Sciences. 1072, 276–287 (2006). [DOI] [PubMed] [Google Scholar]
- 6.Baumgart DC, Sandborn WJ Crohn's disease. Lancet. 380 (9853), 1590–1605 (2012). [DOI] [PubMed] [Google Scholar]
- 7.Ordás I, Eckmann L, Talamini M, Baumgart DC, Sandborn WJ Ulcerative colitis. Lancet. 380 (9853), 1606–1619 (2012). [DOI] [PubMed] [Google Scholar]
- 8.Muthas D. et al. Neutrophils in ulcerative colitis: A review of selected biomarkers and their potential therapeutic implications. Scandanavian Journal of Gastroenterology. 52 (2), 125–135 (2017). [DOI] [PubMed] [Google Scholar]
- 9.Pai RK et al. The emerging role of histologic disease activity assessment in ulcerative colitis. Gastrointestinal Endoscopy. 88 (6), 887–898 (2018). [DOI] [PubMed] [Google Scholar]
- 10.Parkos CA, Delp C, Arnaout MA, Madara JL Neutrophil migration across a cultured intestinal epithelium. Dependence on a CD11b/CD18-mediated event and enhanced efficiency in physiological direction. The Journal of Clinical Investigation. 88 (5), 1605–1612 (1991). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Brazil JC, Parkos CA Pathobiology of neutrophil-epithelial interactions. Immunological Reviews. 273 (1), 94–111 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Thomson A. et al. The Ussing chamber system for measuring intestinal permeability in health and disease. BMC Gastroenterology. 19 (1), 98 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Li BR et al. In vitro and in vivo approaches to determine intestinal epithelial cell permeability. Journal of Visualized Experiments. (140), e57032 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Srinivasan B. et al. TEER measurement techniques for in vitro barrier model systems. Journal of Laboratory Automation. 20 (2), 107–126 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fan S. et al. Role of JAM-A tyrosine phosphorylation in epithelial barrier dysfunction during intestinal inflammation. Molecular Biology of the Cell. 30 (5), 566–578 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Parkos CA Neutrophil-epithelial interactions: A doubleedged sword. American Journal of Pathology. 186 (6), 1404–1416 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Volynets V. et al. Assessment of the intestinal barrier with five different permeability tests in healthy C57BL/6J and BALB/cJ mice. Digital Diseases and Sciences. 61 (3), 737–746 (2016). [DOI] [PubMed] [Google Scholar]
- 18.Wick MJ, Harral JW, Loomis ZL, & Dempsey EC An optimized evans blue protocol to assess vascular leak in the mouse. Journal of Visualized Experiments. (139), e57037 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tateishi H, Mitsuyama K, Toyonaga A, Tomoyose M, Tanikawa K Role of cytokines in experimental colitis: relation to intestinal permeability. Digestion. 58 (3), 271–281 (1997). [DOI] [PubMed] [Google Scholar]
- 20.Mei Q, Diao L, Xu JM, Liu XC, Jin J A protective effect of melatonin on intestinal permeability is induced by diclofenac via regulation of mitochondrial function in mice. Acta Pharmacologica Sinica. 32 (4), 495–502 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vargas Robles H, Castro Ochoa KF, Nava P, Silva Olivares A, Shibayama M, Schnoor M Analyzing Beneficial Effects of Nutritional Supplements on Intestinal Epithelial Barrier Functions During Experimental Colitis. J. Vis. Exp (119), e55095 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Arques JL et al. Salmonella induces flagellin- and MyD88-dependent migration of bacteria-capturing dendritic cells into the gut lumen. Gastroenterology. 137 (2), 579–587 (2009). [DOI] [PubMed] [Google Scholar]
- 23.Coombes BK et al. Analysis of the contribution of Salmonella pathogenicity islands 1 and 2 to enteric disease progression using a novel bovine ileal loop model and a murine model of infectious enterocolitis. Infection and Immunity. 73 (11), 7161–7169 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Everest P. et al. Evaluation of Salmonella typhimurium mutants in a model of experimental gastroenteritis. Infection and Immunity. 67 (6), 2815–2821 (1999). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pron B. et al. Comprehensive study of the intestinal stage of listeriosis in a rat ligated ileal loop system. Infection and Immunity. 66 (2), 747–755 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Clayburgh DR et al. Epithelial myosin light chain kinase-dependent barrier dysfunction mediates T cell activation-induced diarrhea in vivo. The Journal of Clinical Investigation. 115 (10), 2702–2715 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Palmblad J. et al. Leukotriene B4 is a potent and stereospecific stimulator of neutrophil chemotaxis and adherence. Blood. 58 (3), 658–661 (1981). [PubMed] [Google Scholar]
- 28.Mandell KJ, Babbin BA, Nusrat A, Parkos CA Junctional adhesion molecule 1 regulates epithelial cell morphology through effects on beta1 integrins and Rap1 activity. The Journal of Biological Chemistry. 280 (12), 11665–11674 (2005). [DOI] [PubMed] [Google Scholar]
- 29.Laukoetter MG et al. JAM-A regulates permeability and inflammation in the intestine in vivo. Journal of Experimental Medicine. 204 (13), 3067–3076 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Flemming S, Luissint AC, Nusrat A, Parkos CA Analysis of leukocyte transepithelial migration using an in vivo murine colonic loop model. Journal of Clinical Investigation Insight. 3 (20), (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Luissint AC, Nusrat A, Parkos CA JAM-related proteins in mucosal homeostasis and inflammation. Seminars in Immunopathology. 36 (2), 211–226 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cesarovic N. et al. Isoflurane and sevoflurane provide equally effective anaesthesia in laboratory mice. Lab Animal. 44 (4), 329–336 (2010). [DOI] [PubMed] [Google Scholar]
- 33.JoVE Science Education Database. General Laboratory Techniques. Introduction to the Microplate Reader. JoVE, Cambridge, MA, (2020). [Google Scholar]
- 34.Kelm M. et al. Targeting epithelium-expressed sialyl Lewis glycans improves colonic mucosal wound healing and protects against colitis. Journal of Clinical Investigation Insight. 5 (12) (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Azcutia V. et al. Neutrophil expressed CD47 regulates CD11b/CD18-dependent neutrophil transepithelial migration in the intestine in vivo. Mucosal Immunology. 10.1038/s41385-020-0316-4 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yu YR et al. A protocol for the comprehensive flow cytometric analysis of immune cells in normal and inflamed murine non-lymphoid tissues. PloS One. 11 (3), e0150606 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bradfield PF, Nourshargh S, Aurrand-Lions M, Imhof BA JAM family and related proteins in leukocyte migration (Vestweber series). Arteriosclerosis, Thrombosis, and Vascular Biology. 27 (10), 2104–2112 (2007). [DOI] [PubMed] [Google Scholar]
- 38.Ebnet K. Junctional Adhesion Molecules (JAMs): Cell adhesion receptors with pleiotropic functions in cell physiology and development. Physiological Reviews. 97 (4), 1529–1554 (2017). [DOI] [PubMed] [Google Scholar]
- 39.Sorribas M. et al. FXR modulates the gut-vascular barrier by regulating the entry sites for bacterial translocation in experimental cirrhosis. Journal of Hepatology. 71 (6), 1126–1140 (2019). [DOI] [PubMed] [Google Scholar]
- 40.Mazzucco MR, Vartanian T, Linden JR In vivo Blood-brain Barrier Permeability Assays Using Clostridium perfringens Epsilon Toxin. Bio-Protocol. 10 (15), e3709 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kelly JR et al. Breaking down the barriers: the gut microbiome, intestinal permeability and stress-related psychiatric disorders. Frontiers in Cellular Neuroscience. 9, 392 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fiorentino M. et al. Blood-brain barrier and intestinal epithelial barrier alterations in autism spectrum disorders. Molecular Autism. 7 (1), 49 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kelm M. et al. Regulation of neutrophil function by selective targeting of glycan epitopes expressed on the integrin CD11b/CD18. FASEB Journal : An Official Publication of the Federation of American Societies for Experimental Biology. 34 (2), 2326–2343 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]