

ABSTRACT
Many bacteria co-exist and produce antibiotics, yet we know little about how they cope and occupy the same niche. The purpose of the present study was to determine if and how two potent antibiotic-producing marine bacteria influence the secondary metabolome of each other. We established an agar- and broth-based system allowing co-existence of a Phaeobacter species and Pseudoalteromonas piscicida that, respectively, produce tropodithietic acid (TDA) and bromoalterochromides (BACs). Co-culturing of Phaeobacter sp. strain A36a-5a on Marine Agar with P. piscicida strain B39bio caused a reduction of TDA production in the Phaeobacter colony. We constructed a transcriptional gene reporter fusion in the tdaC gene in the TDA biosynthetic pathway in Phaeobacter and demonstrated that the reduction of TDA by P. piscicida was due to the suppression of the TDA biosynthesis. A stable liquid co-cultivation system was developed, and the expression of tdaC in Phaeobacter was reduced eightfold lower (per cell) in the co-culture compared to the monoculture. Mass spectrometry imaging of co-cultured colonies revealed a reduction of TDA and indicated that BACs diffused into the Phaeobacter colony. BACs were purified from Pseudoalteromonas; however, when added as pure compounds or a mixture they did not influence TDA production. In co-culture, the metabolome was dominated by Pseudoalteromonas features indicating that production of other Phaeobacter compounds besides TDA was reduced. In conclusion, co-existence of two antibiotic-producing bacteria may be allowed by one causing reduction in the antagonistic potential of the other. The reduction (here of TDA) was not caused by degradation but by a yet uncharacterized mechanism allowing Pseudoalteromonas to reduce expression of the TDA biosynthetic pathway.
IMPORTANCE
The drug potential of antimicrobial secondary metabolites has been the main driver of research into these compounds. However, in recent years, their natural role in microbial systems and microbiomes has become important to determine the assembly and development of microbiomes. Herein, we demonstrate that two potent antibiotic-producing bacteria can co-exist, and one mechanism allowing the co-existence is the specific reduction of antibiotic production in one bacterium by the other. Understanding the molecular mechanisms in complex interactions provides insights for applied uses, such as when developing TDA-producing bacteria for use as biocontrol in aquaculture.
KEYWORDS: bromoalterochromides, mass spectrometry imaging, metabolomics, Phaeobacter, Pseudoalteromonas, tropodithietic acid
INTRODUCTION
For almost a century, microbial secondary metabolites have been exploited as antibiotics or other drugs, and this vast clinical perspective has been driving science in this area. Despite this, much less is known about their ecological role in natural microbial communities. It has widely been assumed that they are produced to give one species a competitive edge over other species (1) and that their primary function is to kill competitors, ensuring survival of the fittest (2). However, it is increasingly being acknowledged that these metabolites also have other functions, e.g., communication signals, metal-scavenging compounds, or quorum-sensing molecules (1, 2).
While most studies of microbial secondary metabolites, especially with antibiotic potential, have focused on terrestrial microbes, notably Streptomycetes and filamentous fungi (3), recent sequencing endeavors have demonstrated that marine microorganisms harbor a large genetic potential for production of these compounds (4). This is of interest in the current antimicrobial resistance crisis as this may point to environmental niches from which microorganisms that produce novel antibiotics can be bioprospected. However, as in the terrestrial environments, we lack an understanding of the natural role, function, and influence of microbial secondary metabolites with antibiotic activity in marine environments. Insights would be important to understand the dynamics of microbial communities and could point to novel strategies for bioprospecting. Several studies have demonstrated that the potential for secondary metabolite (antibiotic) production is high in marine biofilms (5, 6). Furthermore, some bacteria produce more antibiotics when grown in biofilms than when grown as planktonic cultures, as cell density and a structured environment can affect production (7). Two potent antibiotic-producing genera of marine bacteria are Phaeobacter and Pseudoalteromonas, which often co-exist in marine biofilms (8–10), thus providing a model system for studies of microbial co-existence and the possible roles of antibiotics in co-existence.
Phaeobacter belongs to the paraphyletic Roseobacter group (11) that is one of the most abundant and ubiquitous groups of marine bacteria (12). Within this group, a subset of species produces the potent tropone-derived antibiotic, tropodithietic acid (TDA) (13, 14). Species producing TDA include several species from the Phaeobacter genus, such as Phaeobacter gallaeciensis, Phaeobacter piscinae, and Phaeobacter inhibens (15–17), as well as species within the Tritonibacter and Pseudovibrio genera (14). TDA is broadly antimicrobial against both gram-negative and gram-positive bacteria (13, 18, 19), and resistance development toward TDA appears rare and is a fleeting phenotypic trait (19, 20). Adding TDA or TDA-producing bacteria to a microbial community can alter the relative abundance of bacteria belonging to the Alteromonadales order, including the genus Pseudoalteromonas (21, 22). TDA has been proposed to facilitate the invasion of Phaeobacter spp. on macroalgae surfaces by defeating other potent antimicrobial secondary metabolite producers, such as Pseudoalteromonas spp. (9, 10). In fact, it has been suggested that the ability to produce TDA is maintained by interspecies competition with Pseudoalteromonas species (9, 10).
On the other hand, Pseudoalteromonas is also a highly abundant marine genus, constituting on average up to 3% of the bacterial abundance in the upper surface of the ocean (12). Members of the Pseudoalteromonas genus are excellent biofilm producers and are often found associated with marine eukaryotes, such as algae (23). Pseudoalteromonas species can be categorized into two groups based on their ability to produce pigments (24). The pigmented strains can produce a broad range of bioactive secondary metabolites, such as the non-ribosomal peptide hybrids entitled bromoalterochromides (BACs) produced by Pseudoalteromonas piscicida (25).
One could assume that Phaeobacter and Pseudoalteromonas species would exclude one another so only one would be present at a time as it has been demonstrated that TDA-producing Phaeobacter can invade and take over a Pseudoalteromonas biofilm (26). Nevertheless, antibiotic-producing bacteria from these genera co-exist in marine environments (8). Considering the notion that antimicrobial secondary metabolites are inhibitory for competitors, it begs the questions: how do these two antimicrobial-producing bacteria interact, how do they exist in similar ecological niches, and how do they cope with the antimicrobial secondary metabolites produced by the other?
The purpose of this study was to determine if and how the antibiotic secondary metabolites of two species from these genera affect their interaction and the metabolome of the other. We used the two bacteria Phaeobacter sp. A36a-5a and Pseudoalteromonas piscicida B39bio, both isolated from biofilms in Danish harbors (8).
MATERIALS AND METHODS
Bacterial strains and culture conditions
Phaeobacter sp. A36a-5a (8) and Pseudoalteromonas piscicida B39bio (8) were cultured in Marine Broth (MB) (Difco 2216 Marine Broth; 37.4 g/L) or on Marine Agar (MA) (Difco 2216; 55.1 g/L). The tdaC transcriptional reporter fusion strain Phaeobacter sp. A36-5A-PtdaCGFP was cultured in MB with 100 µg/mL kanamycin. All strains were grown at 25°C and with 200 rpm when grown in liquid cultures unless otherwise stated. Escherichia coli strain Top10, used in cloning, was grown in Lysogeny Broth (LB; Difco 244620 LB Broth Miller; 25 g/L) or on Lysogeny Agar (LA; Difco 244620 LB Agar Miller; 40 g/L) , and E. coli strain WM3064, used in conjugation, was grown in LB and LA supplemented with 0.03 mM 2,6-diaminopimelic acid. Stock cultures were kept in 20% glycerol at −80°C.
Co-cultivation of Pseudoalteromonas and Phaeobacter on solid medium
Overnight (O/N) cultures were diluted 10-fold in MB. Six inocula (10 µL each) of Phaeobacter were spotted on MA in a vertical line and left to dry. Six inocula (10 µL each) of Pseudoalteromonas were then spotted next to Phaeobacter with increasing distance, with the closest spot being a co-inoculum of the two cultures at the same spot. Plates were incubated at 25°C for 3 days followed by visual inspection for growth inhibition and colony morphology changes.
Construction of tdaC transcriptional reporter fusion in Phaeobacter
The vector backbone carrying a promoter-less GFPmut3* gene was amplified from pBBR1-MCS2-PtdaCDE-gfp (6) using primer pairs Fw-gfp-backbone (5′-CGGTTGCGGTCTCAGCTGGAccagcttttgttccctttag-3′) and RV-gfp-backbone (5′-atacgcttggccctaccggaatgcgtaaaggagaagaacttttc-3′). The promoter region and 99 bp 5′ of tdaC gene were amplified from gDNA of Phaeobacter sp. A36-5a using primer sets tdaC_FW (5′-gaaaagttcttctcctttacgcattccggtagggccaagcgtat-3′) and tdaC_RW (5′-agcggaagagacgagtgtccagctgagaccgcaaccg-3′) and cloned onto the vector backbone via direct cloning (27). The recombined plasmid pBBR1-MCS2-PtdaCA36a-5aGFP was conjugated into Pheaobacter sp. A36-5a using protocol described by Lindqvist et al. (28) to generate the tdaC transcriptional reporter fusion strain Phaeobacter sp. A36-5A-PtdaCGFP.
A36-5A-PtdaCGFP was grown as described above on solid medium, and pictures of colony fluorescence were taken with a fluorescence stereo zoom microscope (ZEISS AXIO Zoom.V.16) with modes of GFP (excitation 485/20 nm; emission 520/20 nm) and processed using ZEN v. 2.3 pro.
Mass spectrometry imaging
O/N cultures were 10-fold diluted in 1.5% sterile Instant Ocean (Aquarium Systems Inc., Sarrebourg, France), and 20 µL of Phaeobacter and Pseudoalteromonas was spotted with a distance of 0.5 cm on MA and incubated at 25°C for 6 days. The regions of interest were excised from the plates using a scalpel and spatula, transferring the agar with colonies to conductive glass slides (Bruker IntelliSlides) that had previously had a thin layer of adhesive applied to them (2 Way Glue pen from Kuretake Co. Ltd.) to ensure the attachment of the agar to the slides. The samples were then dried at 35°C in an incubator until the agar was completely dry (2–4 h). A visual light image of the dried samples was then taken (Reflecta MF5000), followed by matrix application.
Depending on the target compound (TDA or BACs), two different matrix application methods were used. When targeting TDA, the matrix consisted of a mixture of 40 mg/mL 2,5-dihydroxybenzoic acid (DHB) and 0.1% trifluoroacetic acid (TFA) dissolved in 100% methanol (MeOH). When targeting BACs, the matrix was a mixture of 40 mg/mL DHB and 0.1% TFA dissolved in 50% MeOH in water. For both target compounds, the matrix was applied to the samples using a HTX Technologies TM-Sprayer with an N2 flow rate of approximately 8 psi, a path pattern of CC, a track spacing of 3 mm, a velocity of 1,200 mm/min, a matrix flow rate of 0.1 mL/min, and with a total of 10 passes. For targeting TDA, the matrix was applied with a nozzle temperature of 40°C, and for targeting BACs, the nozzle temperature was set to 80°C. Acquisition of the MALDI mass spectrometry image (MSI) was achieved using a Bruker TIMS TOF Flex in positive ionization, a pixel size of 30 µm with 600 shots per pixel, and a m/z range of 100–1,300. The mass spectrometer was calibrated using red phosphorus.
Genome sequencing, assembly, and mining
Genomic DNA was purified from both Phaeobacter and Pseudoalteromonas using the Wizard Genomic DNA Purification Kit (Promega) and eluted in 10 mM Tris-HCl (pH 8.0) buffer. DNA concentrations and quality of DNA were determined by fluorescence spectroscopy (Quibit dsDNA HS Assay: Invitrogen by Thermo Fisher Scientific Inc., Eugene, OR, USA) and absorption (DeNovix 439 DS-11+, DeNovix Inc., Wilmington, DE, USA), respectively. Samples were labeled using Rapid Barcoding Kit SQK-RBK004 (Oxford nanopore technologies) following the manufacturer’s instructions. Sequencing was done using the MinION (Oxford nanopore technologies) flow cell with data being analyzed using MinKNOW (Oxford nanopore technologies) and base calling performed using Guppy v. 4.2.2. All nanopore reads were trimmed using porechop v. 0.2.4 (29) to remove adapters. Genomic DNA from Phaeobacter and Pseudoalteromonas was also purified using the protocol described by Wilson (30). The quality was checked as above, and samples were sent for Illumina sequencing (Novogene).
A hybrid assembly from the nanopore reads together with Illumina reads was constructed using the Unicycler assembly pipeline v0.4.9 (31). Before assembly, all Illumina reads were pre-processed using fastp v. 0.23.0, and bandage v. 0.8.1 was used to assess assembly quality. Multilocus sequence analysis was performed using the autoMLST (32). Genomes were mined for biosynthetic gene clusters (BGCs) using antiSMASH v. 6.0.1 using default parameters (33).
Minimum inhibitory concentration assays
To test cross-inhibition (or tolerance) of Phaeobacter and Pseudoalteromonas, liquid monocultures were grown for 48 h, and cells harvested by centrifugation (8,000 × g, 5 min). Sterile-filtered (0.2 µm) supernatant was twofold diluted in sterile-filtered MB in a transparent 96-well plate (u shaped). Approximately 103 cells (of the reciprocal strain) were added to each well giving a total volume of 100 µL. Growth was measured as absorbance at 600 nm in a SpectraMax i3 (Molecular Devices) at 25°C, with shaking. The minimum inhibitory concentration (MIC) of purified bromoalterochromides (BACs) against Phaeobacter was tested in a similar setup.
Purification of bromoalterochromides
All glassware used during purification was amber due to the light sensitivity of the bromoalterochromides. Four liters of liquid fermentation were partitioned thrice with equal volumes of 1:3 isopropanol:ethyl acetate containing 0.1% formic acid. The resultant solution was dried in vacuo using a rotary evaporator (Buchi). Subsequently, liquid-liquid partitioning on the dried extract was undertaken using equal parts water:dichloromethane. The resultant aqueous extract was further separated via an Agilent Infinity II 1260 preparative HPLC. The UV cell was activated during method development and during final purification, and isolation was completed by time-based fractionation to avoid UV degradation. The extract was redissolved in MeOH and fractionated in five 100 mg portions on a Luna C18 preparative column (250 mm by 21.20 mm, 5 µm, 100 Å, Phenomenex). The mobile phase was composed of HPLC-grade water (solvent A) and acetonitrile (solvent B), a gradient of 40%–60% solvent B at 4 mL/min flow rate was used. NMR data for BAC A/A′, A′′, and three unknown structures related to BACs were deposited into NP-MRD (34).
Co-cultivation of Pseudoalteromonas and Phaeobacter in liquid medium
Phaeobacter and Pseudoalteromonas were inoculated in 20 mL MB in 100 mL Erlenmeyer flasks in different starting cell ratios from O/N cultures that had reached an approximate CFU of 109 cells/mL. Mono- and co-cultures were grown at 25°C with and without shaking at 135 rpm. Serial dilutions and plating on MA were used to determine CFU/mL, and the CFU/mL of each culture was based on their different colony morphologies.
Metabolomic analyses of mono- and co-culture
O/N cultures of Pseudoalteromonas and Phaeobacter were inoculated in MB at an approximate ratio of 1:1,000 cells of each strain, respectively, targeting a starting concentration of 5 and 5,000 CFU/mL of Pseudoalteromonas and Phaeobacter, respectively. Cultures were incubated at 25°C with 135 rpm. Samples were taken after 24 h, and cells harvested by centrifugation (8,500 × g, 2 min). The supernatant was mixed 1:1 with isopropanol:ethyl acetate 1:3 (vol/vol) with 0.1% formic acid. The pellets were resuspended in the same volume of isopropanol:ethyl acetate 1:3 (vol/vol) with 0.1% formic acid as used for the supernatant. The resuspended pellets were sonicated for 15 min at 50/60 Hz (Delta 220T Aerosec Industries). Supernatant samples were vortexed for 30 s, and the surface phase was collected. The sonicated samples were centrifuged (8,500 × g, 1 min), and the supernatant was collected. All samples were evaporated under N2 flow. All samples were resuspended in 500 µL methanol and stored at −20°C until further processing.
HR-LC-MS/MS was performed on an Agilent Infinity 1290 UHPLC coupled to an Agilent 6545 QTOF. Liquid chromatography of 1 or 5 µL extract was performed using an Agilent Poroshell 120 phenyl-C6 column (2.1 × 150 mm, 1.9 µm) at 60°C using CH3CN and H2O, both containing 20 mM formic acid. Initially, a linear gradient of 10% CH3CN/H2O to 100% CH3CN over 10 min was employed, followed by isocratic elution of 100% CH3CN for 2 min. Then, the gradient was returned to 10% CH3CN/H2O in 0.1 min, and finally, isocratic condition of 10% CH3CN/H2O for 1.9 min, all at a flow rate of 0.35 min−1 mL. HRMS data were recorded in positive ionization on an Agilent 6545 QTOF MS equipped with an Agilent Dual Jet Stream electrospray ion source with a drying gas temperature of 250°C, drying gas flow of 8 min−1 L, sheath gas temperature of 300°C, and sheath gas flow of 12 min−1 L. Capillary voltage was 4,000 V, and nozzle voltage was set to 500 V. The HRMS data were processed and analyzed using Agilent MassHunter Qualitative Analysis B.07.00. HPLC grade solvents (VWR Chemicals) were used for extractions, while LC-MS grade solvents (VWR Chemicals) were used for LC-MS.
Untargeted and targeted metabolomic analyses were performed using Agilent MassHunter Qualitative Analysis B.07.00, MS-DIAL v. 4.70, and statistical analysis was performed using MetaboAnalyst v. 5.0 (35). Feature-based molecular networking was performed using GNPS (36) and Cytoscape v. 3.9.0.
LC-MS detection limit of TDA
Phaeobacter was grown as monoculture in MB with a starting concentration of 104 CFU/mL. Samples were taken after 4, 12, 14, 16, and 20 h, and cell density determined by serial dilution and plating on MA. Supernatant samples were extracted and processed as described in “Metabolomic analyses of mono- and co-culture,” above, and analyzed by targeted LC-MS for TDA ([M + H]+: m/z 212.9675).
Quantification of tdaC gene expression with RT-qPCR
The Phaeobacter genome from the abovementioned assembly was used to design primers targeting tdaC. tdaC is part of the TDA biosynthetic gene cluster and resides in the same operon as tdaD-E, all of which are essential for TDA production (15). The tdaC sequence was used as template for Primer design in CLC workbench 8.0 (Qiagen, Aarhus, Denmark) using default settings, with a target melting temperature of 60°C. The primers used were tdaC_A36a-5a_forward (5´GGAACGAATTGTGAGCGAG´3) and tdaC_A36a-5a_reverse (5´GAGTGGAGATGGGGGTAG´3). Specificity was tested using DNA from outgrown monocultures of Phaeobacter sp. A36a-5a and Pseudoalteromonas piscicida B39bio. The optimal annealing temperature was 63°C, and the PCR product was verified using sequencing. O/N cultures were grown for the analysis of expression of the tdaC gene using RT-qPCR. RNA was extracted from 18 mL of culture and 2 mL of stop solution (ethanol with 5% phenol), after which lysis was performed using RNeasy Mini Kit (QIAGEN). Samples were DNase treated using TURBO DNA-Free Kit (Invitrogen). RNA was reverse transcribed using SuperScript IV Reverse Transcriptase (Invitrogen), using random hexamer. The qPCR was performed on cDNA using a CFX Opus 96 (Bio-Rad) instrument, with a master mix containing nuclease-free water, Luna Universal qPCR Master Mix (New England BioLabs), and tdaC primers (0.25 µM, final conc.). Thermal cycling conditions were as follows: 95°C for 1 min followed by 40 cycles of 95°C for 15 s, 63°C for 30 s, and finished with a melting curve going for 60°C–95°C with 0.5°C increments. Finally, results were analyzed using CFX Maestro v. 2.3 (Bio-Rad). Statistical analysis was performed using a Student’s t-tests on gene expression normalized to log10(CFU) in R.
RESULTS
Co-cultivation of Pseudoalteromonas and Phaeobacter on solid medium
Phaeobacter sp. strain A36a-5a and Pseudoalteromonas piscicida strain B39bio were co-cultured on Marine Agar. P. piscicida appeared as distinct yellow-pigmented colonies, and the Phaeobacter colonies appeared with a brown color [caused by Fe-TDA precipitation (37)]. Both colony types had regularly shaped edges, and the pigmentation was evenly distributed throughout the colony. When colonies were inoculated in close vicinity to each other (0.5 cm), the distinct brown color in the Phaeobacter colony was reduced in intensity in the area of the colony close to the Pseudoalteromonas colony; however, the growth of Phaeobacter as judged visually by the amount of cells in the colony appeared not affected (Fig. 1). The growth of the Pseudoalteromonas colony was slightly impeded in the interface with the Phaeobacter colony, but the intensity of the yellow pigmentation was only marginally affected. To determine if the presumed reduction in TDA concentration was due to a degradation of the molecule, potentially by Pseudoalteromonas, or a change in production, we fused a promoterless GFP gene to the promoter of tdaC gene to generate the reporter strain Phaeobacter sp. A36-5A-PtdaCGFP. When co-colony cultivations were in close vicinity, the tdaC gene expression, as monitored by green fluorescence, was lower in the area of the Phaeobacter colony close to the Pseudoalteromonas colony as compared to the areas of the Phaeobacter colony furthest from the Pseudoalteromonas (Fig. 2).
Fig 1.

Co-cultivation of Phaeobacter and Pseudoalteromonas on Marine Agar. From left to right: Phaeobacter monoculture, Pseudoalteromonas and Phaeobacter co-cultured, and Pseudoalteromonas monoculture.
Fig 2.

tdaC gene expression, monitored by green fluorescence in tagged Phaeobacter during co-colony cultivations of Phaeobacter and Pseudoalteromonas on Marine Agar. (A) Phaeobacter monoculture. (B) Co-colony cultivation of Phaeobacter (left) and Pseudoalteromonas (right, in dashed white line). tdaC gene expression is lower in the vicinity of the Pseudoalteromonas colony.
Mass spectrometry imaging of co-cultivated Pseudoalteromonas and Phaeobacter
Preparing MSI from Marine Agar-grown colonies was challenging as the drying process typically resulted in salt crystallization; however, TDA was clearly detectable in the monoculture colony of Phaeobacter (Fig. 3). In the co-culture with Pseudoalteromonas, the amount of TDA was higher at the Phaeobacter colony edges furthest from the Pseudoalteromonas colony, thus confirming the observations of the lower amount of the Fe-TDA complex and the reduction in tdaC gene expression. Bromoalterochromides (BAC-A/A′) were detectable in the monoculture of Pseudoalteromonas and in the coculture and diffused from the Pseudoalteromonas colony into the Phaeobacter colony in the area where the level of TDA was lower (Fig. 3). This led us to hypothesize that the BAC compounds were mediating the reduction of TDA production in Phaeobacter, and we, therefore, subsequently purified BACs from Pseudoalteromonas for further analysis (see “The effect of bromoalterochromides on the global metabolome and TDA production in Phaeobacter sp.,” below).
Fig 3.
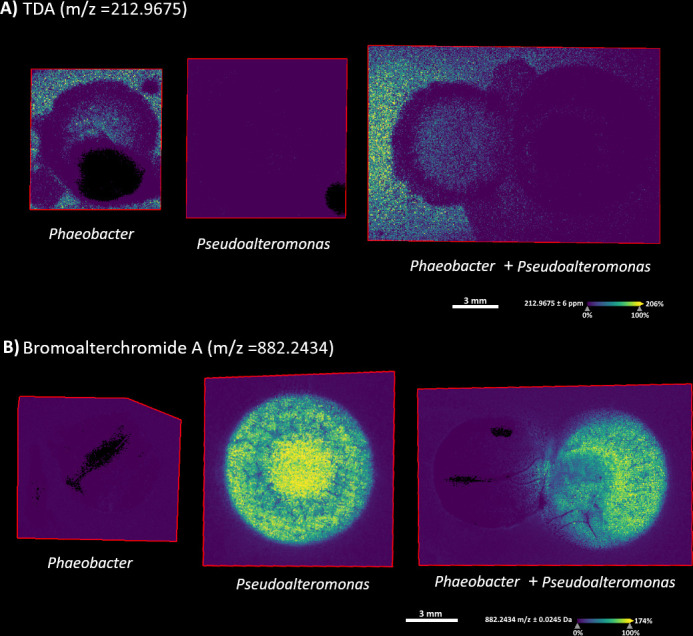
Mass spectrometry imaging of co-cultivated Pseudoalteromonas and Phaeobacter on Marine Agar. (A) MSI with an m/z 212.96 indicating tropodithietic acid. (B) MSI with an m/z of 882.2434 indicating the presence of bromoalterochromides A′.
Prediction of biosynthetic potential of genome-sequenced Phaeobacter and Pseudoalteromonas
To substantiate that the compounds involved in the interaction as detected by chemical analyses could be TDA and BACs, the genetic potential for secondary metabolite production of the two strains was analyzed using antiSMASH (6.0.1) (33). Phaeobacter harbored two small plasmids and Pseudoalteromonas one small plasmid. The total genome size was 4.05 and 5.42 Mbp, respectively. antiSMASH predicted eight biosynthetic gene clusters in the genome of Phaeobacter with a 100% match for a BGC encoding TDA in Phaeobacter inhibens DSM 17395. The number of predicted BGCs in the Pseudoalteromonas genome was 12, of which two could be annotated and encoded BACs and a rhizomide.
Minimum inhibitory concentration assay of Pseudoalteromonas vs Phaeobacter and of bromoalterochromides against Phaeobacter
To enable complete quantification and normalization of gene expression, we transferred the co-cultivation from agar to Marine Broth. Sterile-filtered supernatant of the two strains was inhibitory to each other: the MIC of crude Pseudoalteromonas supernatant against Phaeobacter was a 25% dilution of the supernatant, and the MIC of Phaeobacter supernatant against Pseudoalteromonas was a 50% dilution of the supernatant. Given the reported bioactivity of BACs (38, 39) and the results from the MSI analysis, we suspected that these compounds could course the alterations in the TDA production on Marine Agar. We purified and tested BAC activity against Phaeobacter using fractions containing BAC-A/A′, BAC-D/D′′, di-BAC-B, and a mixed fraction of all three. All three fractions exhibited poor antimicrobial activity: 316, 158, and 175 µM, respectively, against Phaeobacter. The MIC of the mixture of all the BAC fractions was 300 µg/mL. For comparison, BAC-A/A′ and BAC-D/D′′ have been reported to have an MIC of 59.3 and 58.3 µM against Vibrio fischeri and against Bacillus algicola an MIC of 7.4 and 7.3 µM (38). This, therefore, indicates that the BAC compounds are not the main inhibitory Pseudoalteromonas compounds against Phaeobacter sp.
Establishment of a liquid co-cultivation model system
In order to investigate the gene expression and metabolome further, we set up a liquid co-cultivation model system. The Pseudoalteromonas strain was the fastest growing and the most inhibitory of the two, and to allow for co-existence and parallel proliferation, the inoculum levels were adjusted. Multiple inoculum ratios were tested, and Phaeobacter was only able to grow and co-establish itself, when inoculated at a 1,000–10,000-fold higher cell density than Pseudoalteromonas. Monocultures of both strains grew to 108 CFU/mL in 24 h. When co-cultured, Pseudoalteromonas reached log10(CFU/mL) of 6.9 ± 0.26, and Phaeobacter reached log10(CFU/mL) of 6.1 ± 0.45. For the metabolomics analyses (see “Untargeted metabolomic analysis of Phaeobacter and Pseudoalteromonas in liquid mono- and co-culture,” below), samples from monocultures were taken at the same cell densities as were reached in the co-culture system.
Untargeted metabolomics analysis of Phaeobacter and Pseudoalteromonas in liquid mono- and co-culture
Untargeted metabolomics of the mono- and co-cultures detected a total of 1,499 features after LC-MS/MS analysis. The MS features of the supernatant samples were analyzed by principal component analysis (PCA) using Euclidean distances. A tight clustering was seen between features found in the supernatant from Pseudoalteromonas and features found in the supernatant from the co-culture (Fig. 4A), suggesting that the metabolites produced by Phaeobacter were reduced by co-culturing with Pseudoalteromonas. Principal components 1 and 2 were responsible for 72.3% of the sample variance and all data points fitted within the 95% CI. A Spearman correlation clearly demonstrated that the metabolite profile of the co-cultures was dominated by Pseudoalteromonas metabolites (Fig. S1).
Fig 4.

(A) Principal component analysis of metabolites in supernatant extraction from monoculture and co-culture of Phaeobacter and Pseudoalteromonas. Filled dots indicate samples, and translucent areas indicate the 95% CI for a given sample group. (B) Heatmap of the 200 most expressed features from Phaeobacter and Pseudoalteromonas in monoculture and co-culture. Y-axis not shown due to graphical constraints. Each cell on the y-axis represents a unique feature. The value is the normalized relative abundance of each feature and has been pareto scaled via MetaboAnalyst using “sum normalization.” The heatmap shows clustering between Pseudoalteromonas monoculture and co-culture samples.
A heatmap of the 200 most expressed features in all supernatant samples was constructed (Fig. 4B). Phaeobacter had a distinct metabolite profile at 106 CFU/mL in monoculture. However, despite reaching the same cell density in the co-culture, very few of these features were detectable. As expected from the PCA, Pseudoalteromonas metabolites were detectable in both mono- and co-cultures. We specifically searched for features indicative of TDA production by Phaeobacter; however, these were not detected, neither in mono- nor in co-culture. Subsequent metabolomic analyses of Phaeobacter monocultures at different cell densities revealed that TDA was not detectable by our methodology at cell densities below 1 × 107 CFU/mL (data not shown). Overall, the global metabolome of Phaeobacter was affected by the presence of Pseudoalteromonas; however, due to the detection limit, we could not, in the liquid system, quantify the presumed reduction in TDA production.
Gene expression of tdaC in Phaeobacter
Since the Phaeobacter cell densities reached did not allow the quantification of TDA in the liquid co-culture, we developed an RT-qPCR protocol for measuring expression of the tdaC gene, a key gene in the TDA biosynthetic pathway (40). When normalized to cell density, the tdaC gene expression was decreased eightfold (P-value = 0.006) when Phaeobacter was grown in co-culture with Pseudoalteromonas as compared to when grown in monoculture (Fig. 5).
Fig 5.
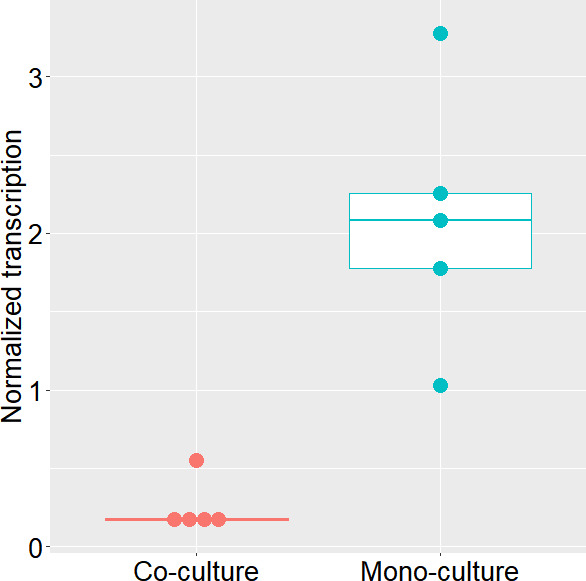
Phaeobacter sp. tdaC expression in mono- and in co-culture with Pseudoalteromonas piscicida in Marine Broth. Transcription is normalized to log10(CFU) levels in the cultures, showing clustering between Pseudoalteromonas monoculture and co-culture samples.
The effect of bromoalterochromides on the global metabolome and TDA production in Phaeobacter sp.
Based on the MSI results, we suspected that BACs could be mediating the reduced production of TDA in Phaeobacter sp. Untargeted metabolomics was therefore used to study changes in metabolite production when a sub-MIC mixture of BAC was added to a monoculture of Phaeobacter. After 24 h, Phaeobacter sp. reached log10(CFU/mL) 9.1 ± 0.16 and 8.7 ± 0.11, when grown with and without the addition of BACs, respectively.
A total of 809 features were detected in the combined samples of monocultures with and without the addition of BAC after LC-MS/MS analysis. Using a fold change of >2 and a non-parametric test, 46 features were detected at significantly (P-value 0.1) higher level, and 26 features were detected at a significantly lower level when the BAC mixture was added to a culture of Phaeobacter as compared to Phaeobacter grown without BACs. In total, 8.9% of the detectable features were produced in different concentrations as a result of the presence of the BAC mixture. However, LC-MS/MS analysis revealed no detectable change in TDA production with the addition of BAC to a monoculture of Phaeobacter.
DISCUSSION
Many bacteria produce secondary molecules with antibiotic activity and have been researched as part of bioprospecting for novel drugs. While the clinical interests have focused on terrestrial bacteria, specifically from Actinobacteria, it has become clear from sequence analyses that marine bacteria harbor a tremendous potential for the production of bioactive secondary metabolites (4, 41). Most bacteria do not exist as monocultures but in complex communities, in microbiomes. This raises the question of how antibiotic bacteria co-exist and whether the (antibiotic) metabolome is affected by co-existence. Indeed, several studies have demonstrated that co-cultivation can induce the production of antibiotics in several bacteria (42, 43). Here, we, in contrast, demonstrate that one antimicrobial-producing marine bacterium reduces the biosynthesis of another antimicrobial in a co-existing marine bacterium.
It is well known that (gut) bacteria may metabolize a range of drugs (44) and that several bacteria may actually live on (degrade) antibiotics as the sole source of carbon (45). Also, recent microbiome studies indicate that many bacteria metabolize (antibiotic) secondary metabolites produced by other members of the community (46, 47). Several studies have, in contrast, described that co-cultivation enhances antibiotic production (43), and this approach has in fact been used as a bioprospecting strategy (42, 48). The enhanced production of antibiotics in co-culture may be caused by the inducing effect of sub-inhibitory concentrations of antibiotics (49) as has also been demonstrated for marine bacteria (50). Our findings, in contrast, indicate that a bacterium may modulate the antibiotic production in another way not just by degradation but by affecting (reducing) the expression of the biosynthetic pathway, shown by the relative reduction of tdaC expression in co-culture.
Reduction of secondary metabolite levels has also been seen in a three-member synthetic communities where the level of koreenceine was reduced in a three-member rhizosphere community as compared to production in monoculture (51). Also, knocking out koreenceine production in Pseudomonas koreensis can lead to the upregulation of expression of BGCs in co-cultured bacteria, showing that secondary metabolites can play a role in gene expression in interspecies interactions (52).
We initially suspected that the bromoalterochromides were mediating the reduced tdaC expression, and hence, TDA concentration, in Phaeobacter based on MSI analysis of co-cultures where BACs were diffusing into the Phaeobacter colony. BACs from Pseudoalteromonas have been described as being antibacterial against both gram-positive bacteria (Bacillus agricola, Bacillus subtilis, and Enterococcus faecium) and gram-negative bacteria (Vibrio fischeri and Vibrio anguillarum) (24, 38); however, some studies have demonstrated a cytotoxic rather than an antibacterial effect (53). In our study, BACs purified from P. piscicida were not antimicrobial toward Phaeobacter sp. BACs have been suspected to play a role in the symbiosis between Pseudoalteromonas and larger microorganisms, by serving as defense against pathogens and fouling (38). This may explain why Phaebacter spp. are tolerant to BACs as many Phaeobacter species are often found associated with eukaryotes as great surface colonizers and will often colonize living surfaces of algae, bryozoan, and crustaceans (16, 54–56). Therefore, Phaeobacter sp. may have developed a tolerance toward BACs in order to occupy the same niche. By purifying BACs, we tested whether they were responsible for decreasing TDA production, which was not the case, and the exact mechanism by which Pseudoalteromonas affects tdaC expression was not uncovered.
Cultivating Phaeobacter sp. and P. piscicida in co-cultures allowing growth and co-existence of both proved to be highly sensitive to the initial inoculation ratios. When P. piscicida was introduced at CFU/mL ratios lower than 1:1,000 (e.g., 1:100, 1:10, 1:1, P. piscicida:Phaeobacter sp.), it consistently outcompeted Phaeobacter sp. Such imbalances are very common in co-cultivations as one strain typically grows faster than the other (57–59), and balancing inocula is required for co-existence. TDA-producing Phaeobacter may inhibit bacteria from the Pseudoalteromonas genus (22) and may even invade and take over already established Pseudoalteromonas biofilms (26); however, the faster growth of Pseudoalteromonas provides it with a growth fitness, allowing it to co-exist with Phaeobacter sp. in liquid culture. We here uncovered another mechanism increasing the fitness of P. piscicida, namely its ability to reduce TDA production. We suggest that the ability of P. piscicida to reduce tdaC expression is important for co-existence of these two bacteria.
Besides specifically reducing the tdaC expression, analysis of the global metabolome in both mono- and co-cultures of Phaeobacter sp. and P. piscicida revealed significant alterations in detectable metabolites. This is in-line with Chevrette et al. that demonstrated that bacterial co-cultures have a distinct metabolite profile as compared to monocultures (52). Notably, a lack of features from Phaeobacter sp. was observed when comparing the features in Phaeobacter sp. monocultures with those in co-culture, suggesting that not only TDA biosynthesis but also other Phaeobacter compounds is reduced by the presence of P. piscicida. This could indicate that the underlying mechanism by which P. piscicida modulates the expression of tdaC is affecting more than just the tdaC gene. However, Lindqvist et al. showed that a TDA-deficient mutant of Phaeobacter piscinae had a significantly altered metabolome compared to the wild type (28), and the change in metabolome of the co-culture could therefore also be a consequence of reduced tdaC transcription and TDA production in Phaeobacter.
The most intense metabolites produced by Phaeobacter sp. in monoculture became undetectable in the co-culture. This could be caused by a reduction in the production by Phaeobacter sp., for instance, by antimicrobial compounds from P. piscicida or P. piscicida could be metabolizing the compounds originating from Phaeobacter sp. These two possibilities are not mutually exclusive, and while it is conceivable that the compounds may degrade independently due to instability, this should occur in monoculture as well and does not account for the large change in detectability.
As mentioned previously, a TDA-producing Phaeobacter is capable of invading a Pseudoalteromonas tunicata biofilm (26) and having the capability of sensing and reducing TDA biosynthesis gene expression, therefore aligns well with the “competition sensing” hypothesis proposed by Cornforth and Foster (60). Competition sensing suggests that bacteria have evolved mechanisms to detect harmful effects caused by other bacteria and respond in a targeted manner to these effects. This encompasses physiological responses that a cell might exhibit when it detects harm or stress induced by other cells, which have evolved to some extent for that specific purpose (60). Thus, competition sensing could offer an evolutionary explanation for the ability of Pseudoalteromonas to reduce TDA production in Phaeobacter but likewise, and in contrast, could also explain that Phaeobacter refrains from producing TDA to prevent a detrimental response from P. piscicida, thus facilitating coexistence.
ACKNOWLEDGMENTS
This study was supported by a grant from the Danish National Research Foundation (DNRF137) and an infrastructure grant from the Novo Nordisk Foundation (NNF 19OC0055625).
P.B.S., N.N.S.E.H., and L.G. designed and planned the experiments. P.B.S. carried out microbiological assays, cultivations, genome sequencing, and analyzed data. A.J.C.A. performed mass spectroscopy imaging and data analysis. Quantification of tdaC gene expression with RT-qPCR, including primer design performed by M.D.S. and P.B.S. LC-MS data acquisition, and analysis was performed by P.B.S. and S.A.J. K.S. and S.A.J. purified bromoalterochromides. S.D.Z. designed and constructed tdaC transcriptional reporter fusion in Phaeobacter, and M.W.S. contributed with lab assistance in the construction. P.B.S., N.N.S.E.H., and L.G. wrote the original manuscript, and S.A.J., A.J.C.A., K.S., S.D.Z., M.D.S., N.N.S.E.H., and L.G. reviewed and edited.
Contributor Information
Lone Gram, Email: gram@bio.dtu.dk.
Isaac Cann, University of Illinois Urbana-Champaign, Urbana, Illinois, USA.
DATA AVAILABILITY
The genomes of Phaeobacter sp. A36a-5a and Pseudoalteromonas piscicida B39bio have been submitted to NCBI with the BioProject ID PRJNA1091865 and BioSample accession no. SAMN40615094 and SAMN40615095. All metabolomics data were deposited in GNPS-MassIVE under accession no. MSV000093286.
REFERENCES
- 1. Pishchany G, Kolter R. 2020. On the possible ecological roles of antimicrobials. Mol Microbiol 113:580–587. doi: 10.1111/mmi.14471 [DOI] [PubMed] [Google Scholar]
- 2. Davies J. 2013. Specialized microbial metabolites: functions and origins. J Antibiot (Tokyo) 66:361–364. doi: 10.1038/ja.2013.61 [DOI] [PubMed] [Google Scholar]
- 3. Traxler MF, Kolter R. 2015. Natural products in soil microbe interactions and evolution. Nat Prod Rep 32:956–970. doi: 10.1039/c5np00013k [DOI] [PubMed] [Google Scholar]
- 4. Paoli L, Ruscheweyh HJ, Forneris CC, Hubrich F, Kautsar S, Bhushan A, Lotti A, Clayssen Q, Salazar G, Milanese A, et al. 2022. Biosynthetic potential of the global ocean microbiome. Nature 607:111–118. doi: 10.1038/s41586-022-04862-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Gram L, Melchiorsen J, Bruhn JB. 2010. Antibacterial activity of marine culturable bacteria collected from a global sampling of ocean surface waters and surface swabs of marine organisms. Mar Biotechnol 12:439–451. doi: 10.1007/s10126-009-9233-y [DOI] [PubMed] [Google Scholar]
- 6. Bech PK, Zhang S-D, Henriksen NNSE, Bentzon-Tilia M, Strube ML, Gram L. 2023. The potential to produce tropodithietic acid by Phaeobacter inhibens affects the assembly of microbial biofilm communities in natural seawater. NPJ Biofilms Microbiomes 9:12. doi: 10.1038/s41522-023-00379-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Junkins EN, McWhirter JB, McCall L-I, Stevenson BS. 2022. Environmental structure impacts microbial composition and secondary metabolism. ISME Commun 2:15. doi: 10.1038/s43705-022-00097-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Bernbom N, Ng YY, Kjelleberg S, Harder T, Gram L. 2011. Marine bacteria from Danish coastal waters show antifouling activity against the marine fouling bacterium Pseudoalteromonas sp. strain S91 and zoospores of the green alga Ulva australis independent of bacteriocidal activity. Appl Environ Microbiol 77:8557–8567. doi: 10.1128/AEM.06038-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Majzoub ME, McElroy K, Maczka M, Thomas T, Egan S. 2018. Causes and consequences of a variant strain of Phaeobacter inhibens with reduced competition. Front Microbiol 9:2601. doi: 10.3389/fmicb.2018.02601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Lutz C, Thomas T, Steinberg P, Kjelleberg S, Egan S. 2016. Effect of interspecific competition on trait variation in Phaeobacter inhibens biofilms. Environ Microbiol 18:1635–1645. doi: 10.1111/1462-2920.13253 [DOI] [PubMed] [Google Scholar]
- 11. Wirth JS, Whitman WB. 2018. Phylogenomic analyses of a clade within the roseobacter group suggest taxonomic reassignments of species of the genera Aestuariivita, Citreicella, Loktanella, Nautella, Pelagibaca, Ruegeria, Thalassobius, Thiobacimonas and Tropicibacter, and the proposal of six novel genera. Int J Syst Evol Microbiol 68:2393–2411. doi: 10.1099/ijsem.0.002833 [DOI] [PubMed] [Google Scholar]
- 12. Wietz M, Gram L, Jørgensen B, Schramm A. 2010. Latitudinal patterns in the abundance of major marine bacterioplankton groups. Aquat Microb Ecol 61:179–189. doi: 10.3354/ame01443 [DOI] [Google Scholar]
- 13. Brinkhoff T, Bach G, Heidorn T, Liang L, Schlingloff A, Simon M. 2004. Antibiotic production by a Roseobacter clade-affiliated species from the German Wadden Sea and its antagonistic effects on indigenous isolates. Appl Environ Microbiol 70:2560–2565. doi: 10.1128/AEM.70.4.2560-2565.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Henriksen N, Lindqvist LL, Wibowo M, Sonnenschein EC, Bentzon-Tilia M, Gram L. 2022. Role is in the eye of the beholder-the multiple functions of the antibacterial compound tropodithietic acid produced by marine Rhodobacteraceae. FEMS Microbiol Rev 46:1–15. doi: 10.1093/femsre/fuac007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Brock NL, Nikolay A, Dickschat JS. 2014. Biosynthesis of the antibiotic tropodithietic acid by the marine bacterium Phaeobacter inhibens. Chem Commun 50:5487. doi: 10.1039/c4cc01924e [DOI] [PubMed] [Google Scholar]
- 16. Thole S, Kalhoefer D, Voget S, Berger M, Engelhardt T, Liesegang H, Wollherr A, Kjelleberg S, Daniel R, Simon M, Thomas T, Brinkhoff T. 2012. Phaeobacter gallaeciensis genomes from globally opposite locations reveal high similarity of adaptation to surface life. ISME J 6:2229–2244. doi: 10.1038/ismej.2012.62 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Sonnenschein EC, Phippen CBW, Bentzon-Tilia M, Rasmussen SA, Nielsen KF, Gram L. 2018. Phylogenetic distribution of roseobacticides in the Roseobacter group and their effect on microalgae. Environ Microbiol Rep 10:383–393. doi: 10.1111/1758-2229.12649 [DOI] [PubMed] [Google Scholar]
- 18. Bruhn JB, Nielsen KF, Hjelm M, Hansen M, Bresciani J, Schulz S, Gram L. 2005. Ecology, inhibitory activity, and morphogenesis of a marine antagonistic bacterium belonging to the Roseobacter clade. Appl Environ Microbiol 71:7263–7270. doi: 10.1128/AEM.71.11.7263-7270.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Porsby CH, Webber MA, Nielsen KF, Piddock LJV, Gram L. 2011. Resistance and tolerance to tropodithietic acid, an antimicrobial in aquaculture, is hard to select. Antimicrob Agents Chemother 55:1332–1337. doi: 10.1128/AAC.01222-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Rasmussen BB, Grotkjær T, D’Alvise PW, Yin G, Zhang F, Bunk B, Spröer C, Bentzon-Tilia M, Gram L. 2016. Vibrio anguillarum is genetically and phenotypically unaffected by long-term continuous exposure to the antibacterial compound tropodithietic acid. Appl Environ Microbiol 82:4802–4810. doi: 10.1128/AEM.01047-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Geng H, Tran-Gyamfi MB, Lane TW, Sale KL, Yu ET. 2016. Changes in the structure of the microbial community associated with Nannochloropsis salina following treatments with antibiotics and bioactive compounds. Front Microbiol 7:1155. doi: 10.3389/fmicb.2016.01155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Dittmann KK, Sonnenschein EC, Egan S, Gram L, Bentzon-Tilia M. 2019. Impact of Phaeobacter inhibens on marine eukaryote-associated microbial communities. Environ Microbiol Rep 11:401–413. doi: 10.1111/1758-2229.12698 [DOI] [PubMed] [Google Scholar]
- 23. Bowman JP. 2007. Bioactive compound synthetic capacity and ecological significance of marine bacterial genus Pseudoalteromonas. Mar Drugs 5:220–241. doi: 10.3390/md504220 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Offret C, Desriac F, Le Chevalier P, Mounier J, Jégou C, Fleury Y. 2016. Spotlight on antimicrobial metabolites from the marine bacteria Pseudoalteromonas: chemodiversity and ecological significance. Mar Drugs 14:129. doi: 10.3390/md14070129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Ross AC, Gulland LES, Dorrestein PC, Moore BS. 2015. Targeted capture and heterologous expression of the Pseudoalteromonas alterochromide gene cluster in Escherichia coli represents a promising natural product exploratory platform. ACS Synth Biol 4:414–420. doi: 10.1021/sb500280q [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Rao D, Skovhus T, Tujula N, Holmström C, Dahllöf I, Webb JS, Kjelleberg S. 2010. Ability of Pseudoalteromonas tunicata to colonize natural biofilms and its effect on microbial community structure. FEMS Microbiol Ecol 73:450–457. doi: 10.1111/j.1574-6941.2010.00917.x [DOI] [PubMed] [Google Scholar]
- 27. Wang H, Li Z, Jia R, Hou Y, Yin J, Bian X, Li A, Müller R, Stewart AF, Fu J, Zhang Y. 2016. RecET direct cloning and Redαβ recombineering of biosynthetic gene clusters, large operons or single genes for heterologous expression. Nat Protoc 11:1175–1190. doi: 10.1038/nprot.2016.054 [DOI] [PubMed] [Google Scholar]
- 28. Lindqvist LL, Jarmusch SA, Sonnenschein EC, Strube ML, Kim J, Nielsen MW, Kempen PJ, Schoof EM, Zhang S-D, Gram L. 2023. Tropodithietic acid, a multifunctional antimicrobial, facilitates adaption and colonization of the producer, Phaeobacter piscinae. mSphere 8:e0051722. doi: 10.1128/msphere.00517-22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Wick R, Volkening J. Porechop. Available from: https://github.com/rrwick/Porechop. Retrieved 29 Sep 2021.
- 30. Wilson K. 2001. Preparation of genomic DNA from bacteria. Curr Protoc Mol Biol Chapter 2:Unit. doi: 10.1002/0471142727.mb0204s56 [DOI] [PubMed] [Google Scholar]
- 31. Wick RR, Judd LM, Gorrie CL, Holt KE. 2017. Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput Biol 13:e1005595. doi: 10.1371/journal.pcbi.1005595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Alanjary M, Steinke K, Ziemert N. 2019. AutoMLST: an automated web server for generating multi-locus species trees highlighting natural product potential. Nucleic Acids Res 47:W276–W282. doi: 10.1093/nar/gkz282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Blin K, Shaw S, Kloosterman AM, Charlop-Powers Z, van Wezel GP, Medema MH, Weber T. 2021. antiSMASH 6.0: improving cluster detection and comparison capabilities. Nucleic Acids Res 49:W29–W35. doi: 10.1093/nar/gkab335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Wishart DS, Sayeeda Z, Budinski Z, Guo A, Lee BL, Berjanskii M, Rout M, Peters H, Dizon R, Mah R, et al. 2022. NP-MRD: the natural products magnetic resonance database. Nucleic Acids Res 50:D665–D677. doi: 10.1093/nar/gkab1052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Pang Z, Chong J, Zhou G, de Lima Morais DA, Chang L, Barrette M, Gauthier C, Jacques P-É, Li S, Xia J. 2021. MetaboAnalyst 5.0: narrowing the gap between raw spectra and functional insights. Nucleic Acids Res 49:W388–W396. doi: 10.1093/nar/gkab382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Wang M, Carver JJ, Phelan VV, Sanchez LM, Garg N, Peng Y, Nguyen DD, Watrous J, Kapono CA, Luzzatto-Knaan T, et al. 2016. Sharing and community curation of mass spectrometry data with global natural products social molecular networking. Nat Biotechnol 34:828–837. doi: 10.1038/nbt.3597 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. D’Alvise PW, Phippen CBW, Nielsen KF, Gram L. 2016. Influence of iron on production of the antibacterial compound tropodithietic acid and its noninhibitory analog in Phaeobacter inhibens. Appl Environ Microbiol 82:502–509. doi: 10.1128/AEM.02992-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Suria AM, Tan KC, Kerwin AH, Gitzel L, Abini-Agbomson L, Bertenshaw JM, Sewell J, Nyholm SV, Balunas MJ. 2020. Hawaiian bobtail squid symbionts inhibit marine bacteria via production of specialized metabolites, including new bromoalterochromides BAC-D/D′. mSphere 5:1–17. doi: 10.1128/mSphere.00166-20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Sobolevskaya MP, Smetanina OF, Speitling M, Shevchenko LS, Dmitrenok PS, Laatsch H, Kuznetsova TA, Ivanova EP, Elyakov GB. 2005. Controlling production of brominated cyclic depsipeptides by Pseudoalteromonas maricaloris KMM 636T. Lett Appl Microbiol 40:243–248. doi: 10.1111/j.1472-765X.2005.01635.x [DOI] [PubMed] [Google Scholar]
- 40. Geng H, Bruhn JB, Nielsen KF, Gram L, Belas R. 2008. Genetic dissection of tropodithietic acid biosynthesis by marine roseobacters. Appl Environ Microbiol 74:1535–1545. doi: 10.1128/AEM.02339-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Kaari M, Manikkam R, Baskaran A. 2022. Exploring newer biosynthetic gene clusters in marine microbial prospecting. Mar Biotechnol 24:448–467. doi: 10.1007/s10126-022-10118-y [DOI] [PubMed] [Google Scholar]
- 42. Zhuang L, Zhang H. 2021. Utilizing cross-species co-cultures for discovery of novel natural products. Curr Opin Biotechnol 69:252–262. doi: 10.1016/j.copbio.2021.01.023 [DOI] [PubMed] [Google Scholar]
- 43. Buijs Y, Zhang S-D, Jørgensen KM, Isbrandt T, Larsen TO, Gram L. 2021. Enhancement of antibiotic production by co-cultivation of two antibiotic producing marine Vibrionaceae strains. FEMS Microbiol Ecol 97:fiab041. doi: 10.1093/femsec/fiab041 [DOI] [PubMed] [Google Scholar]
- 44. Zimmermann M, Zimmermann-Kogadeeva M, Wegmann R, Goodman AL. 2019. Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature 570:462–467. doi: 10.1038/s41586-019-1291-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Dantas G, Sommer MOA, Oluwasegun RD, Church GM. 2008. Bacteria subsisting on antibiotics. Science 320:100–103. doi: 10.1126/science.1155157 [DOI] [PubMed] [Google Scholar]
- 46. Zhang S, Mukherji R, Chowdhury S, Reimer L, Stallforth P. 2021. Lipopeptide-mediated bacterial interaction enables cooperative predator defense. Proc Natl Acad Sci U S A 118:1–6. doi: 10.1073/pnas.2013759118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Hansen ML, Dénes Z, Jarmusch SA, Wibowo M, Lozano-Andrade CN, Kovács ÁT, Strube ML, Andersen AJC, Jelsbak L. 2024. Resistance towards and biotransformation of a Pseudomonas-produced secondary metabolite during community invasion. ISME J 18:wrae105. doi: 10.1093/ismejo/wrae105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Selegato DM, Castro-Gamboa I. 2023. Enhancing chemical and biological diversity by co-cultivation. Front Microbiol 14:1117559. doi: 10.3389/fmicb.2023.1117559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Seyedsayamdost MR. 2014. High-throughput platform for the discovery of elicitors of silent bacterial gene clusters. Proc Natl Acad Sci U S A 111:7266–7271. doi: 10.1073/pnas.1400019111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Buijs Y, Isbrandt T, Zhang S-D, Larsen TO, Gram L. 2020. The antibiotic andrimid produced by Vibrio coralliilyticus increases expression of biosynthetic gene clusters and antibiotic production in Photobacterium galatheae. Front Microbiol 11:622055. doi: 10.3389/fmicb.2020.622055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Lozano GL, Bravo JI, Garavito Diago MF, Park HB, Hurley A, Peterson SB, Stabb EV, Crawford JM, Broderick NA, Handelsman J. 2019. Introducing THOR, a model microbiome for genetic dissection of community behavior. mBio 10:e02846-18. doi: 10.1128/mBio.02846-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Chevrette MG, Thomas CS, Hurley A, Rosario-Meléndez N, Sankaran K, Tu Y, Hall A, Magesh S, Handelsman J. 2022. Microbiome composition modulates secondary metabolism in a multispecies bacterial community. Proc Natl Acad Sci U S A 119:e2212930119. doi: 10.1073/pnas.2212930119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Speitling M, Smetanina OF, Kuznetsova TA, Laatsch H. 2007. Bromoalterochromides A and A′, unprecedented chromopeptides from a marine Pseudoalteromonas maricaloris strain KMM 636T. J Antibiot (Tokyo) 60:36–42. doi: 10.1038/ja.2007.5 [DOI] [PubMed] [Google Scholar]
- 54. Freese HM, Sikorski J, Bunk B, Scheuner C, Meier-Kolthoff JP, Spröer C, Gram L, Overmann J. 2017. Trajectories and drivers of genome evolution in surface-associated marine Phaeobacter. Genome Biol Evol 9:3297–3311. doi: 10.1093/gbe/evx249 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Porsby CH, Nielsen KF, Gram L. 2008. Phaeobacter and Ruegeria species of the Roseobacter clade colonize separate niches in a Danish turbot (Scophthalmus maximus)-rearing farm and antagonize Vibrio anguillarum under different growth conditions. Appl Environ Microbiol 74:7356–7364. doi: 10.1128/AEM.01738-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Prado S, Montes J, Romalde JL, Barja JL. 2009. Inhibitory activity of Phaeobacter strains against aquaculture pathogenic bacteria. Int Microbiol 12:107–114. [PubMed] [Google Scholar]
- 57. Hibbing ME, Fuqua C, Parsek MR, Peterson SB. 2010. Bacterial competition: surviving and thriving in the microbial jungle. Nat Rev Microbiol 8:15–25. doi: 10.1038/nrmicro2259 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Chignell JF, Park S, Lacerda CMR, De Long SK, Reardon KF. 2018. Label-free proteomics of a defined, binary co-culture reveals diversity of competitive responses between members of a model soil microbial system. Microb Ecol 75:701–719. doi: 10.1007/s00248-017-1072-1 [DOI] [PubMed] [Google Scholar]
- 59. Khan N, Maezato Y, McClure RS, Brislawn CJ, Mobberley JM, Isern N, Chrisler WB, Markillie LM, Barney BM, Song HS, Nelson WC, Bernstein HC. 2018. Phenotypic responses to interspecies competition and commensalism in a naturally-derived microbial co-culture. Sci Rep 8:297. doi: 10.1038/s41598-017-18630-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Cornforth DM, Foster KR. 2013. Competition sensing: the social side of bacterial stress responses. Nat Rev Microbiol 11:285–293. doi: 10.1038/nrmicro2977 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The genomes of Phaeobacter sp. A36a-5a and Pseudoalteromonas piscicida B39bio have been submitted to NCBI with the BioProject ID PRJNA1091865 and BioSample accession no. SAMN40615094 and SAMN40615095. All metabolomics data were deposited in GNPS-MassIVE under accession no. MSV000093286.