

Abstract
Background
This study aimed to investigate the antiparasitic and anti-inflammatory potential of pumpkin seed oil in mice infected with Trichinella spiralis by demonstrating its impact on MMP-9 expression and pathogenesis during the intestinal and muscular phases.
Results
In this study, 100 mice were divided into five groups: an infected group, a pumpkin seed oil-treated group (1.5 mg/kg BW, administered three times per week), an albendazole-treated group, a native control group, and a pumpkin oil control group. Gas chromatography–mass spectrometry analysis of the pumpkin seed oil revealed a broad spectrum of biologically active compounds. The pumpkin seed oil treatment led to a significant reduction in the parasite burden, with a 75% decrease in adult worms and a 66% decrease in encysted larvae. Additionally, the infected animals treated with pumpkin oil exhibited a marked reduction in intestinal inflammation, characterized by a progressive increase in goblet cells. The number of encysted larvae in the diaphragm and muscle tissues was also significantly decreased. Furthermore, pumpkin seed oil treatment significantly reduced MMP-9 levels in both intestinal and muscular tissues, highlighting its potential to attenuate inflammation.
Conclusion
These findings underscore the effectiveness of pumpkin seed oil as anti-inflammatory and antiparasitic agent.
Supplementary Information
The online version contains supplementary material available at 10.1186/s12917-024-04241-2.
Keywords: Trichinella, Pumpkin seed oil, MMP-9, Anti-inflammatory, Antiparasitic
Introduction
Trichinella spiralis is a tiny nematode parasite found in rodents, pigs, and humans. It is a highly prevalent and significant parasite that causes trichinellosis globally [1]. This parasite is commonly used to test the effectiveness of different anthelmintic drugs [2]. Albendazole, a broad-spectrum anthelmintic drug, is the primary medication used to treat trichinellosis [3]. Unfortunately, its effectiveness is limited due to its low water solubility and the high level of resistance exhibited by parasitic stages [4]. Additionally, it has many side effects including neurological symptoms such as headaches, nausea, abdominal pain, fever [5]. Other side effects such as leukopenia, anemia, thrombocytopenia, and pancytopenia, and elevation in liver enzymes were also reported [6]. In neuro-helminth infection, albendazoles may cause focal neurologic deficits due to the destruction of helminthic larvae and cysts in the brain, in addition to meningeal signs and increased intracranial pressure [7]. Besides, it is restricted for pregnant women and children under the age of three years [2], and some are suspected to be carcinogenic [8]. Therefore, it is crucial to find safe and effective anti-Trichinellosis drugs, particularly natural agents with low cost, ecofriendly, and with no adverse effects [9]. Throughout history, natural plant extracts have emerged as powerful alternatives or complementary therapies against parasitic diseases. Pumpkin with oily seeds belong to the Cucurbitaceae family and are used to relieve several conditions, such as fever, bronchitis, and sore chests, as well as a diuretic and tonic. Previous literature demonstrated the therapeutic application of pumpkin oil against benign prostatic hyperplasia [10], cardiovascular health [11], antimicrobial properties [12], diabetes management [13], and against parasitic diseases [14–19]. The detailed physiochemical properties of pumpkin seed oil have been evaluated by our colleagues in the Faculty of Agriculture at Assiut University [20]. Pumpkin seed oil contains bioactive compounds that possess anthelmintic properties and is predominantly composed of four fatty acids: linoleic, oleic, stearic, and palmitic with a relative distribution of 33.1%, 43.8%, 7.8%, and 13.4%, respectively [21, 22].
It contains a specific amino acid that plays a major role in eliminating worms and is mainly concentrated in the seeds of Cucurbita species [23, 24].
Several studies have explored the diverse pharmacological effects and parasitological impacts of pumpkin on nematodes [17–19, 25]. This has been found to decrease the number of parasite eggs [26]. The in vitro studies revealed that the alcoholic extract of pumpkin (Cucurbita pepo) completely inhibits the mobility of T. spiralis and T. britovi larvae [27]. Furthermore, Abd Elsadek et al. [28] demonstrated the impact of pumpkin seed oil on the hepatic pathogenicity and inflammatory reactions resulting from T. spiralis infection.
Matrix metalloproteinases (MMPs) are endogenous regulators involved in tissue regeneration and inflammation. They play an essential role in granuloma formation during infection by promoting the infiltration of inflammatory cells and degrading extracellular matrix proteins [29]. In terms of parasitic infections, researchers have extensively studied the functions of these proteins in diseases such as malaria, neurocysticercosis, and angiostrongyliasis [30, 31]. In addition, MMP-9 is also known to have the ability to both generate and resolve fibrosis in the liver [28]. Many studies have reported a significant increase in the serum levels of MMPs in mice infected with T. spiralis and, to a lesser extent, in mice infected with T. pseudospiralis [32]. These results suggested that MMPs, specifically gelatinases, may serve as inflammatory markers [30]. To the best of our knowledge, little information is available regarding the role of pumpkin against T. spiralis infection. Therefore, the current study aimed to explore the effect of pumpkin seed oil on the enteral and parenteral phases of murine trichinellosis by reducing the pathogenesis and production of the inflammatory mediator MMP-9.
Materials and methods
Ethical standards
This study adhered to both national and international ethical guidelines. The study was approved by the Research Ethical Committee of the Faculty of Veterinary Medicine, Assiut University, Egypt, according to the guidelines of The OIE standards for the use of animals in research (protocol code: 06/2023/0140).
Materials
Pumpkin seed oil (purity 100%) was purchased from IMTENAN brand for Natural Oils and Herbs, Egypt. The oil was extracted using a cold-pressed method that preserves its natural benefits. It has a dark brown to green to dark red color with a strong nutty aroma. The oil is 100% pure and natural, and it has been certified by ISO, COA, and MSDS.3.5. Albendazole was provided as a suspension (Alzentale) by Egyptian International Pharmaceutical Industries [33].
Gas chromatography–mass spectrometry (GC–MS) analysis of the pumpkin seed oil
The phytochemical constituents of the pumpkin seed oil were analyzed with GC/MS Agilent 6890 gas chromatograph equipped with an Agilent mass spectrometric detector at Egyptian Pharmacopoeia, using a direct capillary interface and fused silica capillary column PAS-5ms (30m × 0.32 mm × 0.25μm film thickness). The oil under investigation was injected under the following conditions: helium was used as carrier gas at approximately 1.0 ml/min, pulsed split less mode, the solvent delay was 3 min, and the injection size was 1.0 μl. The mass spectrometric detector was operated in electron impact ionization mode with ionization energy of 70 e.v. scanning from m/z 50 to 500. The ion source temperature was 230 multiplier voltage (EM 5 voltage) maintained 1250 v above auto tune. The identification of compounds was achieved by library search on a Wiley 275 L GC/MS database (Thermo Fisher Technology, Waltham, Massachusetts, United States) and using AMDIS software (www.amdis.net), identified by its retention indices (relative to n-alkanes C8.0 – C24.0) and mass spectrum matching to available authentic standards. Wiley spectral library collection and the National Institute of Standards and Technology (NIST) library database curves generated by running GC analysis of representative authentic compounds.
Parasite strain
The strain of T. spiralis isolated from naturally infected pig obtained from El-Bassatine Abattoir, Cairo. In brief, the infected carcasses were skinned, minced, digested in digestive fluid, and then incubated at 37 °C overnight. Larvae were filtered using thieve to remove bones and hair then washed in PBS. The precipitated larvae were washed several times in PBS, the larvae number per ml was counted using light microscope (× 40). BALB/c mice were administered orally with 350 larvae under appropriate conditions and free of pathogens. The infection was maintained via regular passage in the Animal House, Faculty of Veterinary Medicine, Assiut University, Egypt. The larvae were recovered from the carcasses 30 days post-infection [34].
Experimental design
One hundred BALB/c mice (age 8–12 weeks, weighed 25-30 g) were purchased from the Animal House, Theodore Bilharz Research Institute, Cairo, Egypt. Animals were divided into the main five groups Table 1: the infected untreated group was administered orally with 350 T. spirallis larvae, the pumpkin-treated group was administered orally with 1.5 ml/kg BW pumpkin seed oil three times/ week [28, 35], albendazole-treated group was administered orally with 50 mg/kg BW [36]. The negative control and pumpkin control groups were also represented. The animals of all groups were euthanized at the end of the 7 day post infection (dpi) and 54 dpi for enteral and parenteral phase, respectively and anesthetized with intraperitoneal injection of sodium thiopental (100 mg/kg) [37, 38].
Table 1.
Experimental design showing the experimental groups and subgroups, dose of infection, and treatment
| Animal group | Subgroups | |
|---|---|---|
| GI (10 mice) | Control non-infected, non-treated group | –– |
| GII (15 mice) | Pumpkin non-infected group | –– |
| GIII (15 mice) | Infected non-treated group (orally administrated with 350 T. spirallis larvae) | –– |
|
GIV (30 mice) Pumpkin- treated group |
Infected mice were administered orally with 1.5 ml/kg BW pumpkin seed oil three times/ week according to Abd ELsadek et al., 2023 and Elhamalawy, 2018. | • IVa (enteral phase): 15 infected mice were treated with pumpkin seed oil from 2nd till 7th dpi |
| • IVc (parenteral phase): 15 infected mice were treated with pumpkin seed oil from 7th till 54th dpi | ||
|
GV (30 mice) Albendazole-treated group |
Each infected mouse was administered orally with Albendazole, as a reference drug (50 mg/kg BW) according to Abdel-Hakeem et al., 2024 and El-Hamed et al., 2022. | • Va (enteral phase): 15 infected mice were treated with albendazole from 3rd for three successive days |
| • Vb (parenteral phase): 15 infected mice were treated with albendazole from 31th for seven successive days | ||
Parasitological parameters
Evaluation of the adult worm burden in intestine
Mice in each group were euthanized, and the small intestines were removed, washed multiple times, and then gently scraped to expose the mucosa. The washed intestines were placed in phosphate buffered saline (PBS) and incubated for 4 h at 37°C to allow the worms to migrate out of the tissue and gather in a petri dish. Adult worms were examined, and the mean number was estimated under a stereomicroscope at a magnification of × 10 [33].
Evaluation of the larval burden in muscle
Briefly, 54 dpi mice were sacrificed, the skin of each mouse was removed, rectus abdominis muscles were minced, and digested using artificial digested fluid containing 1% pepsin (1:10,000) and 1% HCl in 200 mL distilled water. The mean number of encysted larvae was determined according to Goettstein et al. [3]. The larvae were counted using a stereomicroscope at a magnification of 40 × . The total number of larvae in the carcass was estimated by multiplying the number of larvae in 50 µl by 8000 (dilution factor).
The efficacy of treatment was counted according to Attia et al. [33]
Histopathology
The intestinal and muscle tissues were separated and preserved in a fixative (formalin-alcohol solution). After 48 h, the specimens were washed several times with 70% ethanol and serially dehydrated in increasing alcohol concentrations. The specimens were cleared in xylene three times/45 min each, impregnated, and embedded in paraplast. Transverse sections were cut at 4–5 µm and stained with hematoxylin and eosin [39, 40].
Immunohistochemistry and scoring of MMP-9
For immunohistochemical staining, paraffin-embedded tissues were cut at 3–4 µm on coated slides. All reagents used in the study were from the Dako EnVision™ FLEX system. Citric buffer (50 ×) at a low pH 6.1 (Code DM829) was used as the Antigen Retrieval technique. Peroxidase Blocking Reagent (Code SM801) was applied and incubated at room temperature for 5–10 min. A ready-to-use Tinto prediluted rabbit monoclonal antibody against MMP-9 (Catalog No. BSB 2538; Bio SB clone EP127, USA) was used according to the manufacturer's protocol with a tonsil tissue as a control (Supplementary 1). Horseradish peroxidase secondary antibody (Code SM802) was applied for 20 min at room temperature. DAB solution (Code DM827) was applied for 5–10 min to visualize the bound antibody, and the sections were stained with counterstain with haematoxylin and mounted using Dibutyl Phthalate Polystyrene Xylene. For scoring the immunostaining, three slides from three different animals in each group were examined under a light microscope (OPTICA, Italy) at a fixed high-power field of × 400 magnification. The number of positive cells was counted using ImageJ software, and the mean number of immunopositive cells was expressed [39, 40].
Serum level of MMP-9 using enzyme-linked immunosorbent assay (ELISA)
MMP-9 levels were assessed in all groups during the enteral phase on the 7th dpi and the parenteral phase on the 54th dpi using a commercially available quantitative sandwich immunoassay kit (Cat. No. E0321Ra, BT LAB, Shanghai, China). Briefly, 40 µL of serum sample (three replicated/each tested group) was added to a 96-well plate that had been coated with the Rat MMP-9 antibody. Then, 50 µL of streptavidin-HRP was added and mixed thoroughly. The mixture was incubated at 37°C for 60 min and washed with buffer five times/ per minute. Subsequently, equal volumes (50 µL) of substrate solutions A and B were added, and the plate was further incubated for 10 min at 37°C in dark. The stop solution (50 µL) was added, and MMP-9 was determined at an optical density of 450 nm. The standard values ranged from 0.375 to 6 ng/mL. The intra-assay coefficient of variation was 4% and the inter-assay coefficient of variation was 6.1%.
Data analysis
SPSS software (version 20) and Microsoft Excel sheet (version 2016) were used in analysis, and the data were expressed as mean ± standard deviation. The two-tailed unpaired data was used to evaluate the difference in MMP-9 level. We considered differences significant when P was less than or equal to 0.05.
Results
GC–MS analysis
GC–MS analysis of pumpkin seed oil indicated the presence of active sixteen phytochemical compounds (Fig. 1) with highly biological activities as shown in Table 2. The phytochemical constituents of the oil are mainly mono- and poly- saturated and unsaturated fatty acids, esters, carboxylic acid, and alkenes (Fig. 1). Molecular weight, chemical formula, chemical structure, retention time (RT), concentration (peak area %), and detailed biological activities of the detected bioactive compounds are listed in Table 2.
Fig. 1.

GC-Mass histogram shows the main sixteen phytochemical constituents in the pumpkin seed oil used in the current evaluation
Table 2.
GC–MS spectral analysis of phytochemical compounds identified in pumpkin seed oil
| N | Compound | Molecular weight | Nature | Chemical formula | Chemical structure | Area % | RT | Biological activities |
|---|---|---|---|---|---|---|---|---|
| 1 | Caprylic acid | 144.211 | Ester | C8H16O2 |
|
6.766 | 7.711 | Antioxidant and antibacterial [41] |
| 2 | Capric | 172.268 | Saturated fatty acid | C10H20O2 |
|
2.461 | 8.682 | Antibacterial [42] |
| 3 | Lauric | 200.322 | Saturated fatty acid | C12H24O2 |
|
0.361 | 10.003 | Antibacterial [43] and anti-inflammatory [44] |
| 4 | Myristic | 228.376 | Saturated fatty acid | C14H28O2 |
|
13.421 | 10.983 | Antiviral, antifungal, and anticancer [45] |
| 5 | Palmitic | 256.430 | Saturated fatty acid | C16H32O2 |
|
2.154 | 11.920 | Anti-inflammatory and antioxidant [46] |
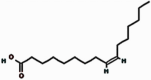
| 6 | Palmitoleic | 254.41 |
Mono unsaturated fatty acid |
C16H30O2 |
|
2.602 | 13.223 | Anti-inflammatory [47] and antibacterial [48] |
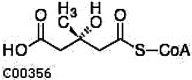
| 7 | Stearic | 284.484 | Saturated fatty acid | C18H36O2 |
|
3.141 | 14.781 | Anti-inflammatory and antioxidant [49] |
| 8 | Oleic | 282.468 |
Mono unsaturated omega-9 fatty acid |
C18H34O2 |
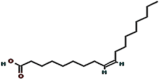
|
25.273 | 14.822 | Antioxidant [50], anti-inflammatory and anticancer [51], antiprotozoal, molluscicidal, and insecticidal [52] |
| 9 | Linoleic | 280.452 | alkane | C18H32O2 |

|
14.078 | 15.389 | Anti-inflammatory and antioxidant [53] |
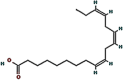
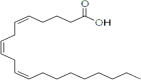
| 10 | Linolenic | 278.4 | triglyceride esters | C18H30O2 |
|
8.413 | 16.008 | Anti-inflammatory, anticancer, and immunomodulatory [54] |
| 11 | Eicosenoic | 310.5 |
Mono unsaturated omega-9 fatty acid |
C20H38O2 |
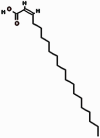
|
2.807 | 17.002 | Anti-inflammatory, anticancer, and antibacterial [55] |
| 12 | Arachidic | 304.474 | Poly-unsaturated omega-6 fatty acid | C20H32O2 |
|
6.011 | 17.540 | Anti-inflammatory, immunomodulatory, and anticancer [56], antibacterial [57] |
| 13 | (24R)-24-Ethyl cholesta-7, 25 (27)-dien-3β-ol | 386.7 | alkaloid | C27H46O |
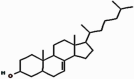
|
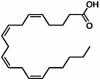
2.388 | 19.883 | Antioxidant, anti-inflammatory, antimicrobial, antidiabetic, and anticancer activity [58] |
| 14 | Eicosatrienoic | 306.4828 | carboxylic acid | C20H34O2 |
|
3.010 | 20.403 | Anti-inflammatory, immunostimulant, and anticancer [59] |
| 15 | Behenic | 340.592 | carboxylic acid | C22H44O2 |
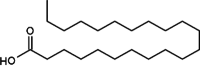
|
3.242 | 22.731 | Antibacterial and immunostimulant [60] |
| 16 | Lignoceric | 368.63 | Saturated fatty acid | C24H48O2 |
|
3.871 | 26.025 | Antitumor, antivirus, antioxidative, antibacterial, anti-UV activities [61] |
Effectiveness of pumpkin seed oil on reduction the mean number of the adult worms and encysted larvae
The pumpkin-treated group showed a significant reduction in the mean number of adult worms (39.1 ± 28.9) and encysted larvae (102,000 ± 27,332.3) with a reduction percentage 78.75% and 64.18%, respectively compared to the infected untreated group (Fig. 2). The albendazole-treated group showed a highly significant reduction (P < 0.0001) in the percentage of adult worms and encysted larvae 99.8% and 98.2%, respectively compared to the infected untreated group.
Fig. 2.

Dot plot shows a marked reduction in mean number of the adult worms (a) and encysted larvae (b) in the pumpkin and albendazole-treated groups
Histopathological findings
Histopathological alteration of the intestine, diaphragm, and muscle were evaluated in all groups. Compared to the normal intestine (Figs. 3a and 4a), the uninfected pumpkin-treated group showed infiltration of mononuclear cells in the intestine (Figs. 3b and 4b). In the enteral phase, the infected-untreated group showed signs of acute catarrhal inflammation, including desquamated epithelium and necrobiotic changes in the enterocytes (Fig. 3c). Furthermore, there were mucosal inflammatory cells, predominantly eosinophils and lymphocytes (Fig. 3c). In the pumpkin-treated group, the intestine showed normal structure with a significant reduction in the enteritis, mild inflammatory reaction in the core of villi, and desquamation of the villar epithelium (Fig. 3d). The intestine in the albendazole-treated group showed normal architecture with inflammatory cell reactions in the core of the villi (Fig. 3e). As shown in Fig. (3c–e), there was a progressively increased number of goblet cells during infection and treatment. In the parenteral phase, the intestine in the infected-untreated group showed milder lesions than in the enteral phase. The intestine showed sloughing of the epithelium in the tips of the villi with a normal number of goblet cells (Fig. 4c). The pumpkin and albendazole-treated groups showed normal intestine structure with a slight inflammatory reaction (Fig. 4d and e).
Fig. 3.
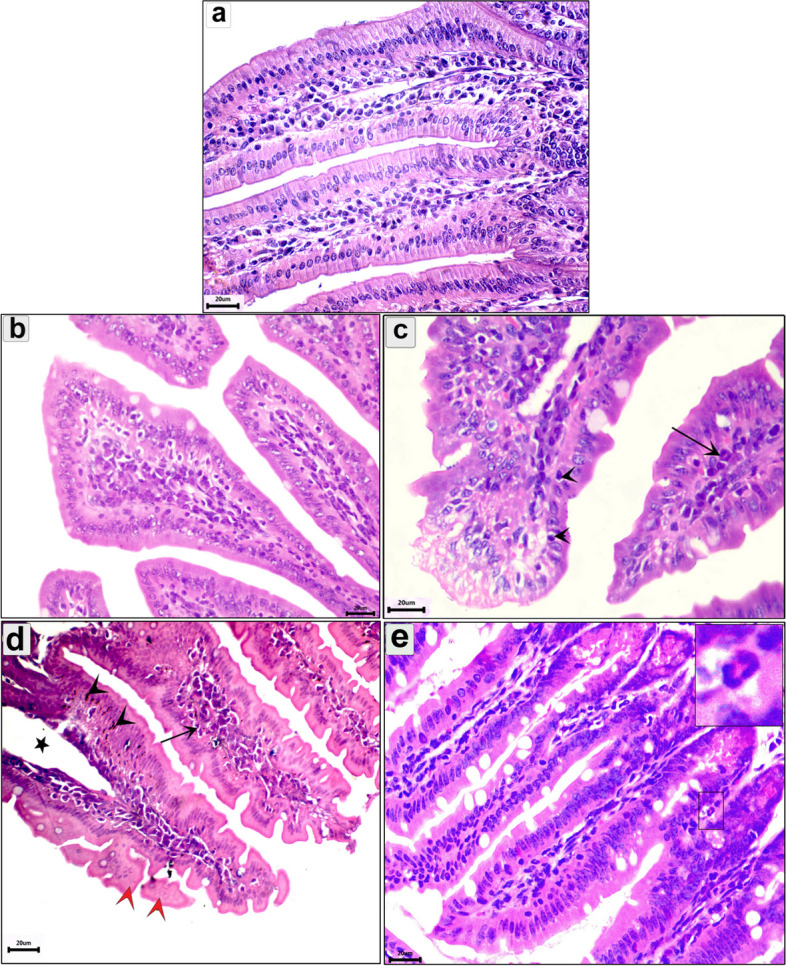
Photomicrograph of the intestine in the different groups in the enteral phase showing: a Normal structure of the intestinal mucosa; b uninfected pumpkin-treated intestine showing normal villi with healthy enterocytes; c Transverse section (T.s) in the intestine of the infected untreated group showing the parasitic enteritis characterized by shedding of the epithelium high inflammatory reaction in the core of the villi (arrow) and mononuclear cells between enterocytes (arrowhead); d T.s in the intestine of the pumpkin-treated group showing less normal intestinal mucosa, hyperplasia in some enterocytes (arrowhead), desquamation of the epithelium in tips of villi (red arrowhead), and mild inflammatory reaction in the core of villi (arrow); e T.s in the intestine of the albendazole-treated group showing normal intestinal villi with high number of goblet cells and eosinophils infiltration (small box). Stain H&E (400 ×)
Fig. 4.
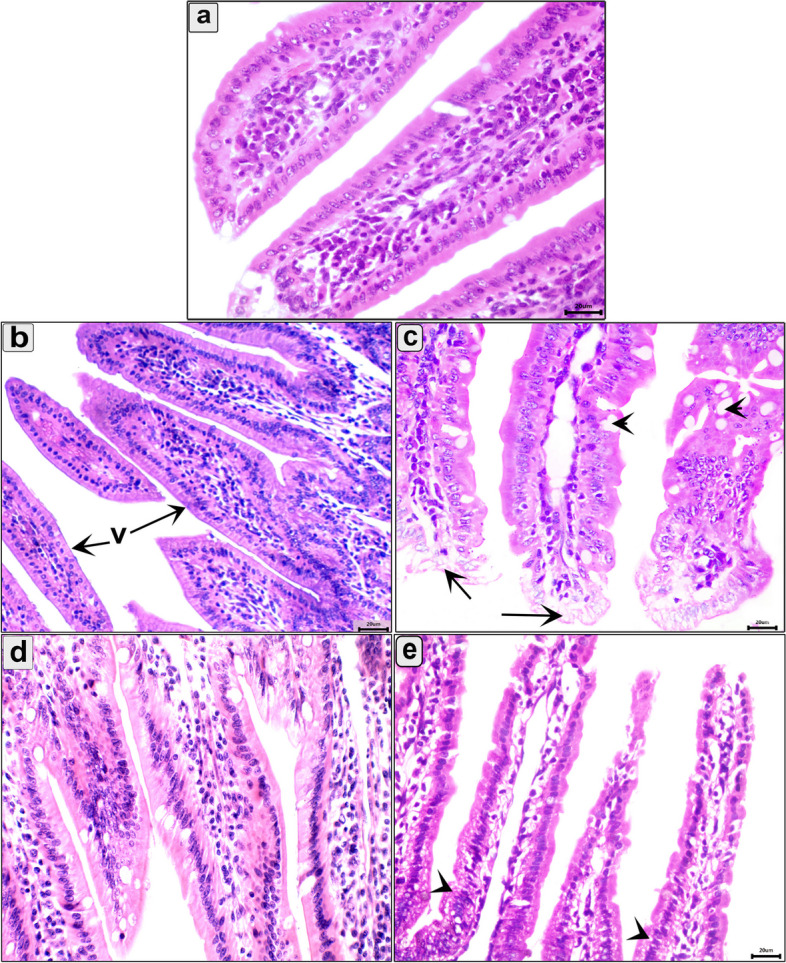
Photomicrograph of the intestine in the different groups in the parenteral phase showing: a Normal structure of the intestinal mucosa; b T.s in the intestine of uninfected pumpkin-treated showing normal villi (v) with healthy enterocytes; c T.s in the intestine of infected untreated group showing vacuolar degeneration in the enterocytes at the tips of villi (arroe) and increase the number of goblet cell (arrowhead); d T.s in the intestine of pumpkin-treated group showing normal architecture of intestinal mucosa; e T.s in the intestine of albendazole-treated group showing necrobiotic changes and vacuolar degeneration of enterocytes (arrowhead). Stain H&E (400 ×)
In the diaphragm and muscle, the control (Figs. 5a and 6a) and the uninfected pumpkin-treated group (Figs. 5b and 6b) showed healthy muscle bundles with flat nucleuses. Multiple cysts containing developed and viable larvae were observed in the infected-untreated group (Figs. 5c and 6c). These cysts were surrounded by a connective tissue capsule and had an inflammatory cell reaction from fibroblast and eosinophil cells (Figs. 5c and 6c). This caused severe parasitic myositis, which was represented by coagulative necrosis, vacuolation, and degeneration of muscle fibers. The pumpkin and albendazole-treated groups showed a significant reduction in the number of encysted larvae in the diaphragm (Fig. 5d and e) and muscle (Fig. 6d and e), which were completely degenerated. An extensive inflammatory cell reaction was observed between muscle fiber and the surrounding encysted larvae.
Fig. 5.
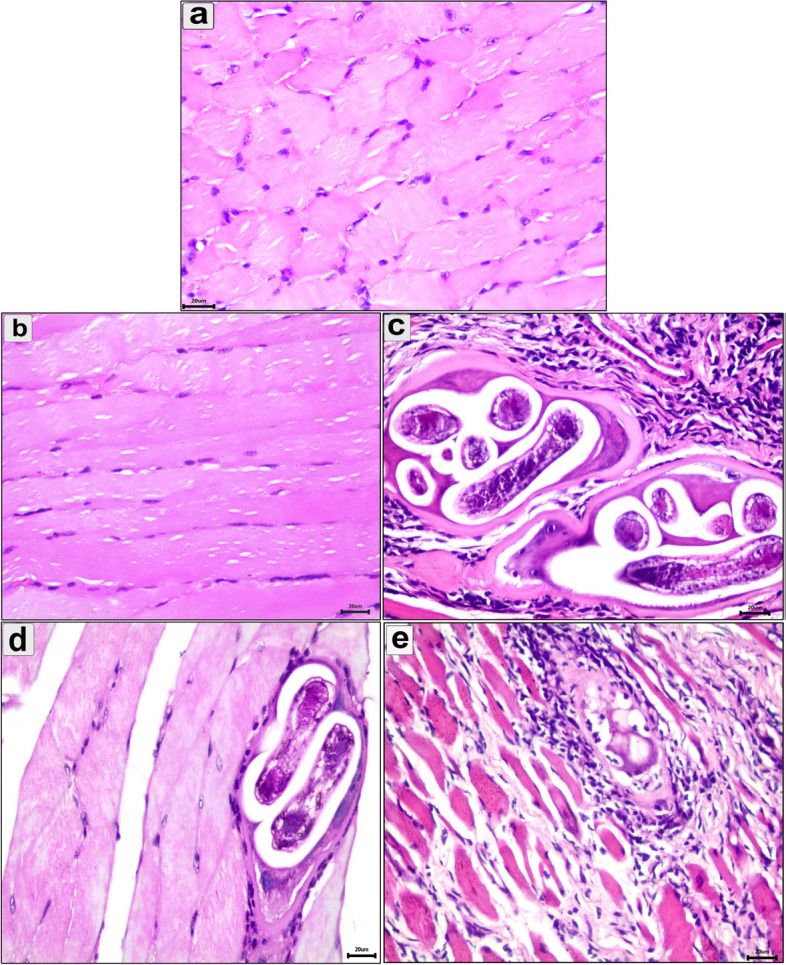
Photomicrograph of the diaphragm in the different groups showing: a Normal diaphragm consists of groups of striated muscle fibers; b The diaphragm of uninfected pumpkin-treated group showing normal muscle fibers; c The diaphragm of infected untreated group showing multiple well-developed cysts containing many larvae surrounded by connective tissue capsule; d The diaphragm of pumpkin-treated group showing marked reduction in the number and size of encysted larvae with normal architecture of the muscle fibers; e The diaphragm of albendazole-treated group showing degeneration larvae and muscle fibers with marked inflammatory reaction. Stain H&E (400 ×)
Fig. 6.
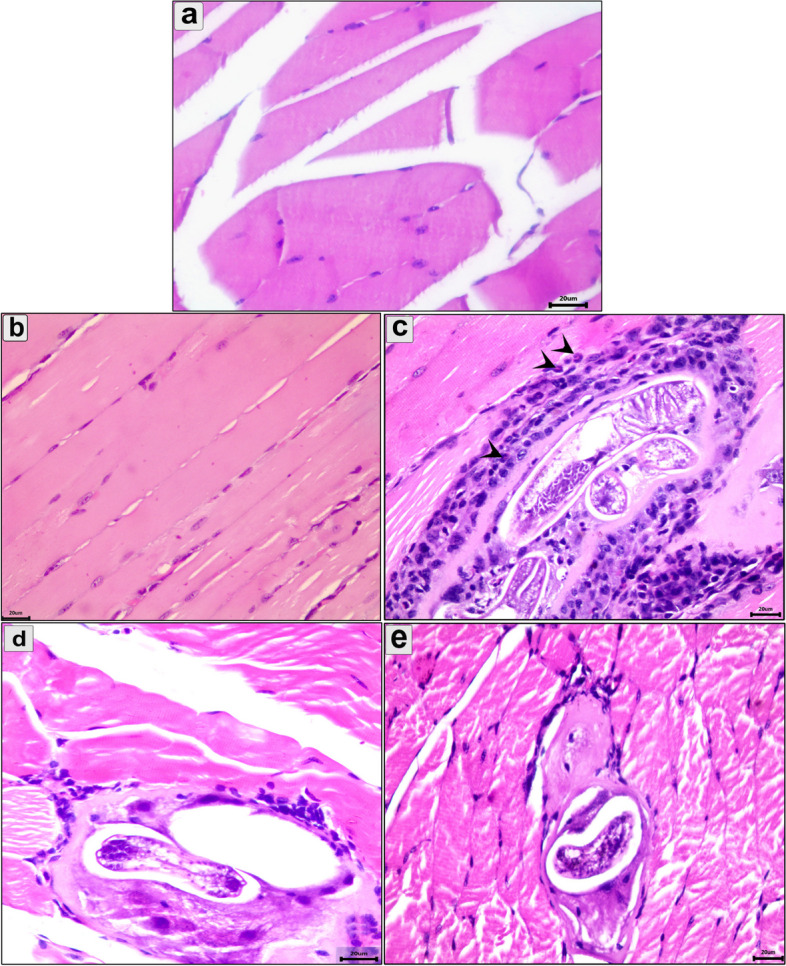
Photomicrograph of the muscle in the different groups showing: a Normal structure of muscle consists of bands of muscle fibers; b Muscle section of the uninfected pumpkin-treated group showing health muscle fibers with mild inflammatory reaction; c Muscle section of the infected untreated group showing multiple larval cysts surrounded by severe inflammatory cells, particularly eosinophils (arrowhead); d Muscle section of the pumpkin-treated group showing a degenerative encysted larvae with mild inflammatory reaction; e Muscle section of the albendazole-treated group showing minute encysted larvae with mild degeneration in the muscle fibers. Stain H&E (400 ×)
Downregulation in the expression of MMP-9 in the intestine, diaphragm, and muscle and accompanied with reduction of MMP-9 level
The immunohistochemical analysis showed cytoplasmic reaction of MMP-9 in the intestine, diaphragm, and muscles (Figs. 7, 8 and 9). The mean number of immunopositively cells in the intestine, diaphragm, and muscle was assessed (Fig. 10). The pumpkin and albendazole-treated groups exhibited a marked significant reduction (P = 0.0007) in the expression of MMP-9 in the intestine (Fig. 7c and d) compared to the infected-untreated group (Fig. 7b). Additionally, the pumpkin-treated group (Fig. 8c) showed a highly significant reduction in the expression of MMP-9 in the diaphragm (P < 0.05), whereas a non-significant expression in the albendazole-treated group (Fig. 8d). Contrarily, both the pumpkin and albendazole-treated groups showed a nonsignificant reduction (P = 0.262) in the expression of MMP-9 in the muscle (Fig. 9c and d) compared to the infected-untreated group (Fig. 9b).
Fig. 7.
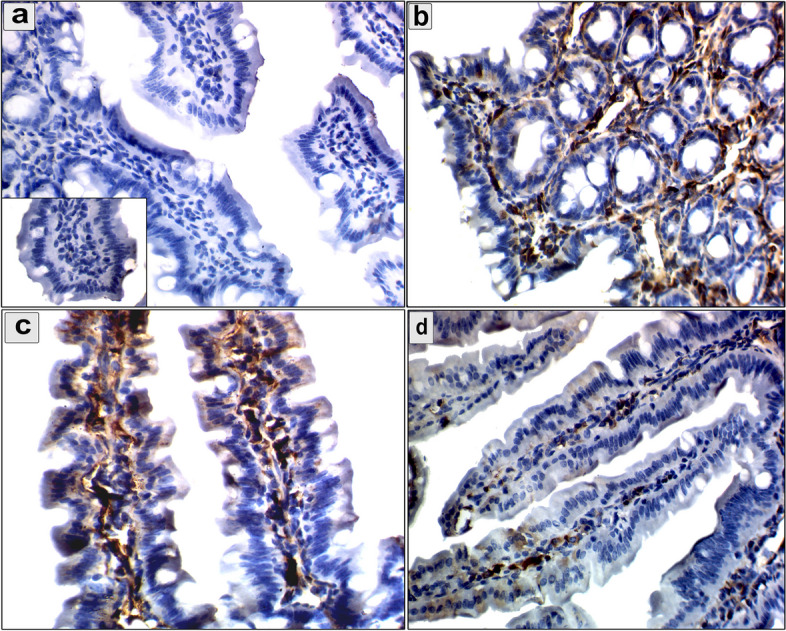
An immunohistochemical staining of MMP-9 the intestine demonstrates the anti-inflammatory effects of pumpkin oil and albendazole (a) The negative reaction in the uninfected pumpkin-treated group (negative control in the small box); b Severe reaction of MMP-9 positive cells in the infected untreated group were distributed in the mucosa and extending to the submucosa; c Marked downregulation in the MMP-9 expression in the pumpkin-treated group and the albendazole-treated group (d)
Fig. 8.
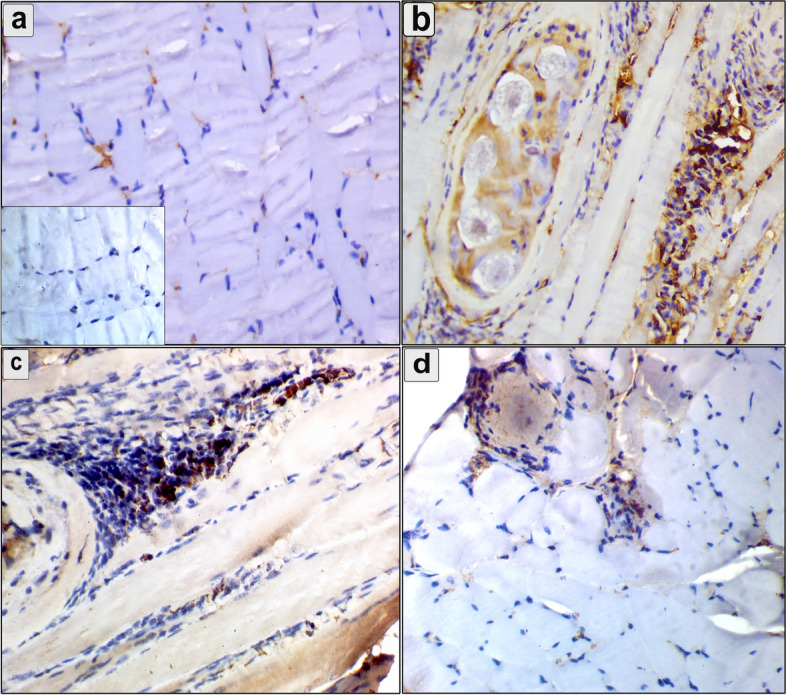
An immunohistochemical staining of MMP-9 in the diaphragm demonstrates the anti-inflammatory effects of pumpkin seed oil and albendazole (a) The negative reaction in the uninfected pumpkin-treated group (negative control in the small box); b Cytoplasmic reaction of MMP-9 positive cells were distributed around encysted larvae and between muscle bands in the infected untreated group; c Marked downregulation in the expression of MMP-9 around the encysted larvae in the pumpkin-treated group and the albendazole-treated group (d)
Fig. 9.
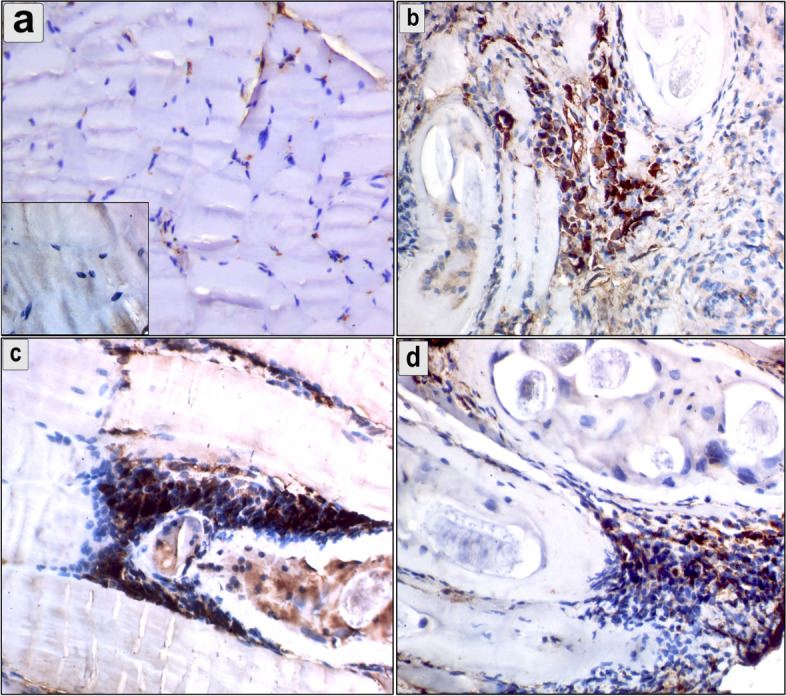
An immunohistochemical staining of MMP-9 in the muscle demonstrates the anti-inflammatory effects of pumpkin and albendazole. a The negative reaction in the uninfected pumpkin-treated group (negative control in the small box); b Cytoplasmic reaction of MMP-9 positive cells in the infected untreated group were distributed between muscle fibers and around the encysted larvae; c non-significant reduction in the MMP-9 expression was observed around the encysted larvae and between muscle bands in the pumpkin-treated group and the albendazole-treated group (d)
Fig. 10.

Histogram showing the mean number of MMP-9 positive cells in the intestine (a), diaphragm (b), and muscle (c) of the different groups. Differences were highly significant at ***P ≤ 0.0001, ns, non-significant
The level of MMP-9 in the different groups was determined using the ELISA technique in the enteral (7dpi) and parenteral phase (54dpi) of infection. Our results revealed a significant decrease (P = 0.0001) in the level of MMP-9 between the infected and treated groups in the enteral (Fig. 11) and parenteral (Fig. 12) phase. The pumpkin and albendazole-treated groups demonstrated a statistically significant reduction in the level of MMP-9 compared to the infected-untreated group.
Fig. 11.

Histogram showing the quantitative level of MMP-9 in the enteral phase of the different groups. A highly significant increase in the level of MMP-9 was observed between uninfected and infected groups. A significant reduction in the treated groups compared to the infected untreated group. Differences were highly significant at P ≤ 0.0001
Fig. 12.

Histogram showing the quantitative level of MMP-9 in the parenteral phase of the different groups. A highly significant increase in the level of MMP-9 was observed between uninfected and infected groups. A significant reduction in the treated groups compared to the infected untreated group. Differences were highly significant at P ≤ 0.0001
Discussion
Pumpkin seeds, as a natural supplement, can influence the hosts’ ability to cope with the negative impact resulting from parasitism and improve the host immunity. In this study, we explore the anthelminthic impact of pumpkin seed oil on the enteral and parenteral phases of T. spiralis infection, demonstrating its role in diminishing the inflammatory process. Our GC-Mass analysis shows that the pumpkin seed oil contains mono and poly saturated and unsaturated fatty acids which have a marked biological activity as antibacterial, antiparasitic, anti-inflammatory, and antioxidant. As antiparasitic agent, our results show successfully reduced the adult worm burden and muscle larval load of T. spiralis by 75% and 66%, respectively in the pumpkin-treated group. Grzybek et al. [19] previously studied the nematocidal activity of pumpkin extract, reporting that it reduced the egg hatching, larval development, and motility of adult worms of Heligmosoides bakeri in vitro. The in vitro impact of pumpkin seed oil against nematodes was also reported against larval stages of Haemonchus contortus [62], and Ascaridia galli [17]. In an vivo study, the extract of pumpkin showed the vigorous antiparasitic effect at high doses [19]. Maciel et al.[63] demonstrated that active substances such as nitrogen-containing chemicals in pumpkins have larvicidal and ovicidal effects, which may have inhibited larval development. Furthermore, anthelminthic impact of pumpkin seeds might be attributed to the secondary metabolites which correlated with get rid of the parasites from the host and weakening the attachment between the parasite and the intestinal mucosa [14]. In our experiment, the uninfected mice orally administered with the pumpkin seed oil exhibited a negative effect on the histological architecture of the intestine and muscle tissues with mild inflammatory reaction in the intestinal mucosa and around encysted larvae. This could be attributed to the activation of innate immune reaction. However, the pumpkin seeds possess a proteolytic effect and can damage the tegument, including the basal membrane [64]. Previous histological investigations showed a negative effect of pumpkin seeds on the general health of animal models such as rat and swine [65]. Our published data showed that pumpkin seed oil could diminish the liver damage and reduce the oxidative stress, which occasionally accompanies T. spiralis infection, potentially aiding in the recovery mechanism [28]. This could be attributed to the trace minerals such as zinc in pumpkin seeds, acting as an antioxidant having the ability to neutralize free radical generation and/or directly bind to the iron or copper binding sites of lipids, proteins, and DNA molecules [66].
The intestine, diaphragm, and muscle were the main organs affected by T. spiralis infection. In the current study, the intestine of the infected-untreated group showed marked parasite enteritis and inflammatory reactions, extending to the submucosa. These results were consistent with Gazzinelli-Guimaraes and Nutman [67], and Sorobetea et al. [68]. Furthermore, we reported mechanical damage in both intestinal and skeletal muscle cells with the accumulation of inflammatory cells. This could be due to the direct mechanical action of the parasite, causing pathological reactions. In the intestine, the administration of pumpkin seed oil to the infected mice restored the normal architecture of the intestinal tissue and reduced the inflammatory reaction. On the other hand, we noted the increase in the number of goblet cells in all infected groups which are usually associated with the invasion of T. spiralis [69, 70].
Bruschi and Chiumiento [71] reported that the inflammation of skeletal muscles correlated with high levels of oxygen reactive species and other free radicals. Interestingly, we reported a statistically significant decrease in the mean number of encysted larvae in the diaphragm and muscle at 54 dpi, suggesting that pumpkin seed oil contains active metabolites that might have a nematocidal effect. Additionally, it might be due to the degenerative effect of pumpkin seed oil on the reproductive organs of the parasites [72] and control of parasite fertility [73]. The anthelminthic effect of pumpkin seed extract in vivo studies was dependent on time and dose [17]. Although there was a non-significant difference between the efficiency of pumpkin seed oil in the present study compared to in vitro studies. This might be due to the effect of gastrointestinal factors such as PH that may change the response of parasite worms to treatment [74].
The anti-inflammatory effects of pumpkin seed oil could be attributed to the putative long-term health effects of isocaloric switch by partial replacement of saturated fats by unsaturated fats [75]. MMP-9 is recommended in the recent literature as an indirect marker to assess myositis severity in infected hosts [32, 76]. Our results clearly demonstrated a highly intracellular staining of MMP-9 in mononuclear cells infiltrating the lamina propria in the intestine and muscle tissues starting one week post infection in the infected-untreated group. This could be attributed cell-mediated reaction of macrophages that can induce large amounts of MMP production [77]. This was correlated with the fibrotic process in the intestinal and muscular diseases, which agreed with the results of Von Lampe et al. [78]. To the best of our knowledge, little is known about the effectiveness of pumpkin oil in the activation of MMP-9 during infection. The PI3K/Akt/NF-κB signaling pathway is indeed well-documented as a crucial regulator of MMP9 expression, particularly in inflammatory processes. Studies have shown that the activation of PI3K/Akt leads to the activation of NF-κB, which in turn upregulates MMP9 transcription [79, 80]. This pathway plays a critical role in mediating the inflammatory response, contributing to various pathological conditions, including those influenced by dietary components such as oils. For instance, omega-3 fatty acids, commonly found in fish oil, have been shown to inhibit this pathway, thereby reducing MMP9 expression and attenuating inflammation [81]. In our study primarily, we focus on the direct measurement of MMP9 as an inflammatory mediator in response to pumpkin oil treatments against T. spiralis infection. As pumpkin seed oil rich with carotenoids, phenolic compounds, and flavonoids, it modulates COX-2 gene which contain consensus sequences for NF-κB and play key role in inflammatory process. Pro-inflammatory stimuli activate the complex containing the NF-κB essential modulator (NEMO) and IκB kinase (IKK)1/2. IKK1/2 phosphorylates IκBs by IKK signalosome complex [82]. In this study, the pumpkin oil showed a statistically significant reduction in the expression of MMP-9 in the infected mice, suggesting its impact as anti-inflammatory agents and stimulation immune responses. The obtained results are promising for the putative anti-inflammatory effect of pumpkin seeds and an integrated approach to control inflammatory diseases.
Conclusion
Considering the outcomes of our results, pumpkin seed oil could be used as an effective alternative drug, characterized by costless and high clinical safety. Our results reported the therapeutic impact of pumpkin seed oil against parasitic infection and inflammatory diseases. More investigations are needed to elucidate the active components of Cucurbita species responsible for their anti-inflammatory agent.
Supplementary Information
Acknowledgements
The authors thank the Science, Technology, and Innovation Funding Authority (STDF) and the Egyptian Knowledge Bank (EKB) who provide open access funding.
Authors’ contributions
Author Contributions: S.S. Abdel-Hakeem, M.A.Z. Abdel-Samiee, M.S.E. Youssef, S.H.A. Abd Elbaset, and S. M. Abd-Elrahman involved in the conception of the research idea, methodology design, supervision, performed data analysis, and interpretation. S.S. Abdel-Hakeem, S.M. Alnasser, A. Meshal, M.A.Z. Abdel-Samiee, and S. M. Abd-Elrahman participated in data analysis, drafted, prepared the manuscript for publication, and revision. All authors read and approved of the final manuscript.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Availability of data and materials
No/Not applicable (this manuscript does not report data generation or analysis).
Data availability
No datasets were generated or analysed during the current study.
Declarations
Ethics approval and consent to participate
This study met requirements of the Research Ethical Committee of the Faculty of Veterinary Medicine, Assiut University, Egypt under approval number (06/2023/0140). The study participants provided their informed consent. All procedures were carried out in conformity with the applicable rules and regulations. The study was conducted in accordance with the ARRIVE (Animals in Research: Reporting In Vivo Experiments) criteria [83].
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Liu X, Song Y, Jiang N, Wang J, Tang B, Lu H, Peng S, Chang Z, Tang Y, Yin J. Global gene expression analysis of the zoonotic parasite Trichinella spiralis revealed novel genes in host parasite interaction. 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Yadav AK. Temjenmongla: Efficacy of Lasia spinosa leaf extract in treating mice infected with Trichinella spiralis. Parasitol Res. 2012;110:493–8. [DOI] [PubMed] [Google Scholar]
- 3.Gottstein B, Pozio E, Nöckler K. Epidemiology, diagnosis, treatment, and control of trichinellosis. Clin Microbiol Rev. 2009;22(1):127–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nassef NE, Moharm IM, Atia AF, Brakat RM, Abou Hussien NM, Shamseldeen A. Therapeutic efficacy of chitosan nanoparticles loaded with albendazole on parenteral phase of experimental Trichinellosis. J Egypt Soc Parasitol. 2019;49(2):301–11. [Google Scholar]
- 5.Speich B, Ali SM, Ame SM, Bogoch II, Alles R, Huwyler J, Albonico M, Hattendorf J, Utzinger J, Keiser J. Efficacy and safety of albendazole plus ivermectin, albendazole plus mebendazole, albendazole plus oxantel pamoate, and mebendazole alone against Trichuris trichiura and concomitant soil-transmitted helminth infections: a four-arm, randomised controlled trial. Lancet Infect Dis. 2015;15(3):277–84. [DOI] [PubMed] [Google Scholar]
- 6.Xing F, Ye H, Yang J. Chan JF-W, Seto W-K, Pai PM-C, Yuen K-Y, Hung DL-L: Fatal pancytopenia due to albendazole treatment for strongyloidiasis. IDCases. 2018;12:112–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Malik K, Dua A. Albendazole. In: StatPearls. edn. Treasure Island (FL): StatPearls Publishing. Florida, United States; 2023. https://www.ncbi.nlm.nih.gov/books/NBK553082/.
- 8.Shalaby MA, Moghazy FM, Shalaby HA, Nasr SM. Effect of methanolic extract of Balanites aegyptiaca fruits on enteral and parenteral stages of Trichinella spiralis in rats. Parasitol Res. 2010;107:17–25. [DOI] [PubMed] [Google Scholar]
- 9.Basyoni MM. El-Sabaa A-AA: Therapeutic potential of myrrh and ivermectin against experimental Trichinella spiralis infection in mice. Korean J Parasitol. 2013;51(3):297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hong H, Kim C-S, Maeng S. Effects of pumpkin seed oil and saw palmetto oil in Korean men with symptomatic benign prostatic hyperplasia. Nurs Res Pract. 2009;3(4):323–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Al-Okbi SY, Mohamed DA, Kandil E, Ahmed E, Mohammed S. Functional ingredients and cardiovascular protective effect of pumpkin seed oils. Grasas Aceites. 2014;65(1):e007. [Google Scholar]
- 12.Leichtweis MG, Molina AK, Pires TC, Dias MI, Calhelha R, Bachari K, Ziani BE, Oliveira MBP, Pereira C, Barros L. Biological activity of pumpkin byproducts: Antimicrobial and antioxidant properties. Molecules. 2022;27(23):8366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Makni M, Fetoui H, Gargouri NK, Garoui EM, Zeghal N. Antidiabetic effect of flax and pumpkin seed mixture powder: effect on hyperlipidemia and antioxidant status in alloxan diabetic rats. J Diabetes Complications. 2011;25(5):339–45. [DOI] [PubMed] [Google Scholar]
- 14.Acorda JA, Mangubat IYEC, Divina BP. Evaluation of the in vivo efficacy of pumpkin (Cucurbita pepo) seeds against gastrointestinal helminths of chickens. Turkish J Vet Anim Sci. 2019;43(2):206–11. [Google Scholar]
- 15.Alhawiti AO, Toulah FH, Wakid MH. Anthelmintic potential of Cucurbita pepo Seeds on Hymenolepis nana. Acta Parasitol. 2019;64:276–81. [DOI] [PubMed] [Google Scholar]
- 16.Ayaz E, Gökbulut C, Coşkun H, Uçar Türker A, Özsoy Ş, Ceylan K. Evaluation of the anthelmintic activity of pumpkin seeds (Cucurbita maxima) in mice naturally infected with Aspiculuris tetraptera. 2015. [Google Scholar]
- 17.Aziz ARA, AbouLaila MR, Aziz M, Omar MA, Sultan K. In vitro and in vivo anthelmintic activity of pumpkin seeds and pomegranate peels extracts against Ascaridia galli. Beni-Suef Univ J Basic Appl Sci. 2018;7(2):231–4. [Google Scholar]
- 18.Feitosa TF, Vilela VLR, Athayde ACR, Braga FR, Dantas ES, Vieira VD, de Melo LRB. Anthelmintic efficacy of pumpkin seed (Cucurbita pepo Linnaeus, 1753) on ostrich gastrointestinal nematodes in a semiarid region of Paraíba State Brazil. Trop Animal Health Prod. 2012;45:123–7. [DOI] [PubMed] [Google Scholar]
- 19.Grzybek M, Kukula-Koch W, Strachecka A, Jaworska A, Phiri AM, Paleolog J, Tomczuk K. Evaluation of anthelmintic activity and composition of pumpkin (Cucurbita pepo L.) seed extracts—in vitro and in vivo studies. Int J Mol Sci. 2016;17(9):1456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Khalifa A, Abdel-Rahman E, Mohamed SA, Al-Barbari F. Characteristics of Seeds And Seed Oils of Some Egyptian Pumpkin Cultivars. J Food Dairy Sci. 2012;3(12):681–97. [Google Scholar]
- 21.Bialek A, Bialek M, Jelinska M, Tokarz A. Fatty acid composition and oxidative characteristics of novel edible oils in Poland. CyTA-J Food. 2017;15(1):1–8. [Google Scholar]
- 22.Siano F, Straccia MC, Paolucci M, Fasulo G, Boscaino F, Volpe MG. Physico-chemical properties and fatty acid composition of pomegranate, cherry and pumpkin seed oils. J Sci Food Agric. 2016;96(5):1730–5. [DOI] [PubMed] [Google Scholar]
- 23.Abdel-Rahman MK. Effect of pumpkin seed (Cucurbita pepo L.) diets on benign prostatic hyperplasia (BPH): chemical and morphometric evaluation in rats. World J Chem. 2006;1(1):33–40. [Google Scholar]
- 24.François G, Nathalie B, Jean-Pierre V, Daniel P, Didier M. Effect of roasting on tocopherols of gourd seeds (Cucurbita pepo). Grasas Aceites. 2006;57(4):409–14. [Google Scholar]
- 25.Matthews K, O’Brien D, Whitley N, Burke J, Miller J, Barczewski R. Investigation of possible pumpkin seeds and ginger effects on gastrointestinal nematode infection indicators in meat goat kids and lambs. Small Rumin Res. 2016;136:1–6. [Google Scholar]
- 26.Obregón DD, Lozano LL, Zúñiga VC. Preclinical studies of Cucurbita maxima (pumpkin seeds) a traditional intestinal antiparasitic in rural urban areas. Revista de gastroenterologia del Peru: organo oficial de la Sociedad de Gastroenterologia del Peru. 2004;24(4):323–7. [PubMed] [Google Scholar]
- 27.Boros Z, Baies M-H, Gherman C, Cozma V. The effects of Artemisia absinthium (wormwood), Allium sativum (garlic), Cucurbita pepo (pumpkin), and Coriandrum sativum (coriander) on Trichinella spiralis and Trichinella britovi larvae, in vitro study. Sci Parasitol. 2021;22:70–8. [Google Scholar]
- 28.Abd ELsadek SH,. Abdel-Samiee MA-Z, Youssef MSE, Abdel-Hakeem SS: Hepatoprotective role of Nanourcumin and Pumpkin seed oil in Trichinella spiralis infection: pathogenesis and modulation of matrix metaloproteinasis (MMP9). Assiut Vet Med J. 2023;69(179):100–14. [Google Scholar]
- 29.Hu J, Van den Steen PE. Sang Q-XA, Opdenakker G: Matrix metalloproteinase inhibitors as therapy for inflammatory and vascular diseases. Nat Rev Drug Discovery. 2007;6(6):480–98. [DOI] [PubMed] [Google Scholar]
- 30.Bruschi F, Pinto B. The significance of matrix metalloproteinases in parasitic infections involving the central nervous system. Pathogens. 2013;2(1):105–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chiu P-S, Lai S-C. Matrix metalloproteinase-9 leads to blood–brain barrier leakage in mice with eosinophilic meningoencephalitis caused by Angiostrongylus cantonensis. Acta Trop. 2014;140:141–50. [DOI] [PubMed] [Google Scholar]
- 32.Bruschi F, D’amato C, Piaggi S, Bianchi C, Castagna B, Paolicchi A, Pinto B. Matrix metalloproteinase (MMP)-9: a realiable marker for inflammation in early human trichinellosis. Vet Parasitol. 2016;231:132–6. [DOI] [PubMed] [Google Scholar]
- 33.Attia RA, Mahmoud AE, Farrag HMM, Makboul R, Mohamed ME, Ibraheim Z. Effect of myrrh and thyme on Trichinella spiralis enteral and parenteral phases with inducible nitric oxide expression in mice. Mem Inst Oswaldo Cruz. 2015;110:1035–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Abd-ELrahman SM, Dyab AK, Mahmoud AE-S, Elossily NA, Alsharif FM, Mohamed SM, Abomughaid MM. Influence of chemically and biosynthesized silver nanoparticles on in vitro viability and infectivity of Trichinella spiralis muscle larvae. bioRxiv 2021:2020.2012. 2017.423206. [DOI] [PubMed]
- 35.Elhamalawy O. Protective effect of pumpkin seed oil against lead acetate toxicity in male mice. Al-Azhar Journal of Pharmaceutical Sciences. 2018;58(2):115–29. [Google Scholar]
- 36.El-Hamed A, Abd-Rabou A, Faramawy A. Therapeutic efficacy of curcuma and pomelo loaded chitosan nanoparticles in intestinal murine trichinellosis. Egypt J Chem. 2022;65(2):551–64. [Google Scholar]
- 37.Abd-Eldayem AM, Alnasser SM, Abd-Elhafeez HH, Soliman SA, Abdel-Emam RA. Therapeutic versus preventative use of Ginkgo biloba Extract (EGb 761) against indomethacin-induced gastric ulcer in mice. Molecules. 2022;27(17):5598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tavares NC, Mourão MM. Parasitemia evaluation in mice infected with Schistosoma mansoni. Bio-Protoc. 2021;11(10):e4017–e4017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Abdel-Hakeem SS, Abdel-Samiee MA, Abed GH. An insight into the potential parasitological effect of Schistosoma mansoni antigens in infected mice: Prophylactic role of cercarial antigen. Microsc Microanal. 2020;26(4):708–16. [DOI] [PubMed] [Google Scholar]
- 40.Alnasser SM, Alotaibi M, Ramadan NK, Abd ElHafeez HH, Abdel-Hakeem SS. The Efficiency of Schistosoma mansoni Crude Antigens in Inhibition of Heat Shock Protein, Apoptosis, and Lysosomal Activity: An Immunohistochemical Study. Microsc Microanal. 2023;29(2):739–53. [DOI] [PubMed] [Google Scholar]
- 41.Kim JJ, Kim HK. Antioxidant and antibacterial activity of caprylic acid vanillyl ester produced by lipase-mediated transesterification. J Microbiol Biotechnol. 2021;31(2):317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Murzyn A, Krasowska A, Stefanowicz P, Dziadkowiec D, Łukaszewicz M. Capric acid secreted by S. boulardii inhibits C. albicans filamentous growth, adhesion, and biofilm formation. Plos one. 2010;5(8):e12050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Anzaku AA, Akyala JI, Juliet A, Obianuju EC. Antibacterial activity of lauric acid on some selected clinical isolates. Ann Clin Lab Res. 2017;5(2):1–5. [Google Scholar]
- 44.Nakatsuji T, Kao MC, Fang JY, Zouboulis CC, Zhang L, Gallo RL, Huang CM. Antimicrobial property of lauric acid against Propionibacterium acnes: its therapeutic potential for inflammatory acne vulgaris. J Investig Dermatol. 2009;129(10):2480–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Javid S, Purohit MN, Yogish Kumar H, Ramya K, Mithuna NFA, Salahuddin MD, Prashantha Kumar BR. Semisynthesis of Myristic Acid Derivatives and Their Biological Activities: A Critical Insight. J Biol Active Prod Nat. 2020;10(6):455–72. [Google Scholar]
- 46.Jin XL, Wang K, Li QQ, Tian WL, Xue XF, Wu LM, Hu FL. Antioxidant and anti-inflammatory effects of Chinese propolis during palmitic acid-induced lipotoxicity in cultured hepatocytes. J Funct Foods. 2017;34:216–23. [Google Scholar]
- 47.Astudillo AM, Meana C, Bermúdez MA, Pérez-Encabo A, Balboa MA, Balsinde J. Release of anti-inflammatory palmitoleic acid and its positional isomers by mouse peritoneal macrophages. Biomedicines. 2020;8(11):480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Watanabe T, Yano S, Kawai T, Jinbo Y, Nonomura Y. Selective antibacterial activity of palmitoleic acid in emulsions and other formulations. J Surfactants Deterg. 2021;24(6):973–9. [Google Scholar]
- 49.Fratianni F, d’Acierno A, Ombra MN, Amato G, De Feo V, Ayala-Zavala JF, Nazzaro F. Fatty acid composition, antioxidant, and in vitro anti-inflammatory activity of five cold-pressed Prunus seed oils, and their anti-biofilm effect against pathogenic bacteria. Front Nutr. 2021;8:775751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wei CC, Yen PL, Chang ST, Cheng PL, Lo YC, Liao VHC. Antioxidative activities of both oleic acid and Camellia tenuifolia seed oil are regulated by the transcription factor DAF-16/FOXO in Caenorhabditis elegans. PLoS ONE. 2016;11(6):e0157195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Farag MA, Gad MZ. Omega-9 fatty acids: Potential roles in inflammation and cancer management. J Genet Eng Biotechnol. 2022;20(1):1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Al-Snafi AE. Antiparasitic, antiprotozoal, molluscicidal and insecticidal activity of medicinal plants (part 2)–plant-based review. Sch Acad J Pharm. 2016;5(6):194–207. [Google Scholar]
- 53.Kaseke T, Opara UL, Fawole OA. Fatty acid composition, bioactive phytochemicals, antioxidant properties and oxidative stability of edible fruit seed oil: Effect of preharvest and processing factors. Heliyon. 2020;6(9):e04962. 10.1016/j.heliyon.2020.e04962. [DOI] [PMC free article] [PubMed]
- 54.Pariza MW, Park Y, Cook M. Mechanisms of action of conjugated linoleic acid: Evidence and speculation (44457). Proc Soc Exp Biol Med. 2000;223(1):8–13. [DOI] [PubMed] [Google Scholar]
- 55.Sheppe AE, Edelmann MJ. Roles of eicosanoids in regulating inflammation and neutrophil migration as an innate host response to bacterial infections. Infect Immun. 2021;89(8):e0009521. 10.1128/IAI.00095-21. [DOI] [PMC free article] [PubMed]
- 56.Zhang Y, Chen H, Zhang W, Cai Y, Shan P, Wu D, Liang G. Arachidonic acid inhibits inflammatory responses by binding to myeloid differentiation factor-2 (MD2) and preventing MD2/toll-like receptor 4 signaling activation. Biochimica et Biophysica Acta (BBA)-Mol Basis Dis. 2020;1866(5):165683. [DOI] [PubMed] [Google Scholar]
- 57.Beavers WN, Monteith AJ, Amarnath V, Mernaugh RL, Roberts LJ, Chazin WJ, Skaar EP. Arachidonic acid kills Staphylococcus aureus through a lipid peroxidation mechanism. MBio. 2019;10(5):10–1128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Das N, Mishra SK, Bishayee A, Ali ES, Bishayee A. The phytochemical, biological, and medicinal attributes of phytoecdysteroids: An updated review. Acta Pharmaceutica Sinica B. 2021;11(7):1740–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Deng H, Li W. Monoacylglycerol lipase inhibitors: modulators for lipid metabolism in cancer malignancy, neurological and metabolic disorders. Acta Pharmaceutica Sinica B. 2020;10(4):582–602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Natnan ME, Low CF, Chong CM, Bunawan H, Baharum SN. Oleic acid as potential immunostimulant in metabolism pathways of hybrid grouper fingerlings (Epinephelus fuscoguttatus× Epinephelus lanceolatus) infected with Vibrio vulnificus. Sci Rep. 2023;13(1):12830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Shu F, Jiang B, Yuan Y, Li M, Wu W, Jin Y, Xiao H. Biological activities and emerging roles of lignin and lignin-based products─ A review. Biomacromol. 2021;22(12):4905–18. [DOI] [PubMed] [Google Scholar]
- 62.Marie-Magdeleine C, Hoste H, Mahieu M, Varo H, Archimède H. In vitro effects of Cucurbita moschata seed extracts on Haemonchus contortus. Vet Parasitol. 2009;161(1–2):99–105. [DOI] [PubMed] [Google Scholar]
- 63.Maciel M, Morais SM, Bevilaqua C, Camurça-Vasconcelos A, Costa C, Castro C. Ovicidal and larvicidal activity of Melia azedarach extracts on Haemonchus contortus. Vet Parasitol. 2006;140(1–2):98–104. [DOI] [PubMed] [Google Scholar]
- 64.Dotto JM, Chacha JS. The potential of pumpkin seeds as a functional food ingredient: A review. Sci Afr. 2020;10:e00575. [Google Scholar]
- 65.de Queiroz-Neto A, Mataqueiro MI, Santana AE, Alessi AC. Toxicologic evaluation of acute and subacute oral administration of Cucurbita maxima seed extracts to rats and swine. J Ethnopharmacol. 1994;43(1):45–51. [DOI] [PubMed] [Google Scholar]
- 66.Amara S, Abdelmelek H, Garrel C, Guiraud P, Douki T, Ravanat J-L, Favier A, Sakly M, Rhouma KB. Preventive effect of zinc against cadmium-induced oxidative stress in the rat testis. J Reprod Dev. 2008;54(2):129–34. [DOI] [PubMed] [Google Scholar]
- 67.Gazzinelli-Guimaraes PH, Nutman TB. Helminth parasites and immune regulation. F1000Research. 2018;7:F1000 Faculty Rev-1685. 10.12688/f1000research.15596.1. [DOI] [PMC free article] [PubMed]
- 68.Sorobetea D, Svensson-Frej M, Grencis R. Immunity to gastrointestinal nematode infections. Mucosal Immunol. 2018;11(2):304–15. [DOI] [PubMed] [Google Scholar]
- 69.Chen Y, Huang B, Huang S, Yu X, Li Y, Song W, Li Y, Lu F. Coinfection with Clonorchis sinensis modulates murine host response against Trichinella spiralis infection. Parasitol Res. 2013;112:3167–79. [DOI] [PubMed] [Google Scholar]
- 70.Khan W, Blennerhasset P, Ma C, Matthaei K, Collins S. Stat6 dependent goblet cell hyperplasia during intestinal nematode infection. Parasite Immunol. 2001;23(1):39–42. [DOI] [PubMed] [Google Scholar]
- 71.Bruschi F, Chiumiento L. Trichinella inflammatory myopathy: host or parasite strategy? Parasit Vectors. 2011;4(1):1–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Bauri R, Tigga MN, Kullu SS. A review on use of medicinal plants to control parasites. Indian J Nat Prod Resources (IJNPR) [Formerly Natural Product Radiance (NPR)]. 2015;6(4):268–77. [Google Scholar]
- 73.Strickland V, Krebs G, Potts W. Pumpkin kernel and garlic as alternative treatments for the control of Haemonchus contortus in sheep. Animal Prod Sci. 2009;49(2):139–44. [Google Scholar]
- 74.Ademola IO, Eloff J. Anthelminthic activity of acetone extract and fractions of Vernonia amygdalina against Haemonchus contortus eggs and larvae. Trop Anim Health Prod. 2011;43:521–7. [DOI] [PubMed] [Google Scholar]
- 75.Shaban A, Sahu RP. Pumpkin seed oil: an alternative medicine. Int J Pharm Phytochem Res. 2017;9(2):11. 10.25258/phyto.v9i2.8066. [DOI] [PMC free article] [PubMed]
- 76.Bruschi F, Pinto B, Fallahi P, Ferrari SM, Antonelli A. Increased neutrophil derived chemokines (CXCL10 and CCL2) in human trichinellosis as possible serological markers of the polarization of the immune response against the parasite. Cytokine. 2023;166:156205. [DOI] [PubMed] [Google Scholar]
- 77.Lacraz S, Isler P, Vey E, Welgus HG, Dayer J-M. Direct contact between T lymphocytes and monocytes is a major pathway for induction of metalloproteinase expression. J Biol Chem. 1994;269(35):22027–33. [PubMed] [Google Scholar]
- 78.Von Lampe B, Barthel B, Coupland S, Riecken E, Rosewicz S. Differential expression of matrix metalloproteinases and their tissue inhibitors in colon mucosa of patients with inflammatory bowel disease. Gut. 2000;47(1):63–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Shim Y-J, Kang B-H, Jeon H-S, Park I-S, Lee K-U, Lee I-K, Park G-H, Lee K-M, Schedin P, Min B-H. Clusterin induces matrix metalloproteinase-9 expression via ERK1/2 and PI3K/Akt/NF-κB pathways in monocytes/macrophages. J Leukoc Biol. 2011;90(4):761–9. [DOI] [PubMed] [Google Scholar]
- 80.Zhang W, Tu G, Lv C, Long J, Cong L, Han Y. Matrix metalloproteinase-9 is up-regulated by CCL19/CCR7 interaction via PI3K/Akt pathway and is involved in CCL19-driven BMSCs migration. Biochem Biophys Res Commun. 2014;451(2):222–8. [DOI] [PubMed] [Google Scholar]
- 81.Calder PC. Omega-3 polyunsaturated fatty acids and inflammatory processes: nutrition or pharmacology? Br J Clin Pharmacol. 2013;75(3):645–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Desai SJ, Prickril B, Rasooly A. Mechanisms of phytonutrient modulation of cyclooxygenase-2 (COX-2) and inflammation related to cancer. Nutr Cancer. 2018;70(3):350–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Du Sert NP, Ahluwalia A, Alam S, Avey MT, Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl U, Emerson M. Reporting animal research: Explanation and elaboration for the ARRIVE guidelines 2.0. PLoS biology. 2020;18(7):e3000411. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
No/Not applicable (this manuscript does not report data generation or analysis).
No datasets were generated or analysed during the current study.