

Abstract
The type III secretion system (T3SS) encoded by Salmonella Pathogenicity Island 2 (SPI2) is essential for virulence and intracellular proliferation of Salmonella enterica. We have previously identified SPI2-encoded proteins that are secreted and function as a translocon for the injection of effector proteins. Here, we describe the formation of a novel SPI2-dependent appendage structure in vitro as well as on the surface of bacteria that reside inside a vacuole of infected host cells. In contrast to the T3SS of other pathogens, the translocon encoded by SPI2 is only present singly or in few copies at one pole of the bacterial cell. Under in vitro conditions, appendages are composed of a filamentous needle-like structure with a diameter of 10 nm that was sheathed with secreted protein. The formation of the appendage in vitro is dependent on acidic media conditions. We analyzed SPI2-encoded appendages in infected cells and observed that acidic vacuolar pH was not required for induction of SPI2 gene expression, but was essential for the assembly of these structures and their function as translocon for delivery of effector proteins.
Keywords: intracellular pathogen, protein translocation, Salmonella-containing vacuole, surface appendage, type III secretion
Introduction
Gram-negative bacteria have evolved a variety of systems to solve the problem of protein transport across the two membranes of the cell envelope. One class, referred to as type III secretion systems (T3SS), consists of complex molecular machines that are functionally and structurally related to the flagella assembly systems. The main biological function of T3SS is not the secretion of proteins into the media, but rather the vectorial translocation of the so-called effector proteins over a third membrane system, namely the cytoplasm membrane or vacuolar membrane of a eukaryotic host cell (for a review, see Hueck, 1998). For many pathogens, the contact-dependent translocation of sets of effector proteins into a eukaryotic host cell has a central role in modulation of host cell functions by the pathogens. In addition, T3SS-dependent protein translocation is also important for the establishment of symbiotic relationships between bacteria and eukaryotic hosts.
The complex assembly of T3SS has made the understanding of the entire secretion apparatus, which consists of more than 20 gene products, elusive. However, it has been possible to visualize parts of T3SS of Salmonella enterica (Kubori et al, 1998), Shigella flexneri (Blocker et al, 1999) and other pathogens. The formation of short, needle-like structures extending from the outer membrane has been observed. The T3SS of certain pathogens are also involved in the formation of long pilus-like structures, as first observed for the ‘Esp pilus' of enteropathogenic Escherichia coli (EPEC) (Knutton et al, 1998). More detailed analyses in EPEC showed that this appendages consists of a needle-like structure composed of EscF subunits (Sekiya et al, 2001; Wilson et al, 2001) on which the filament consisting of EspA subunits (Daniell et al, 2001) is assembled.
S. enterica is a gastrointestinal pathogen of man and animal with a complex pathogenesis involving two different T3SS. S. enterica is able to invade eukaryotic cells such as enterocytes of the intestinal mucosa. The invasion phenotype is linked to the function of a T3SS encoded by genes within Salmonella Pathogenicity Island 1 (SPI1), which has been investigated in great detail (Galan, 2001). S. enterica is also a facultative intracellular pathogen that is able to survive phagocytosis and can proliferate inside infected host cells. A large number of gene functions are required for the intracellular phenotype of S. enterica. Among these is Salmonella Pathogenicity Island 2 (SPI2), a further large gene cluster encoding a second T3SS for translocation of virulence proteins. The structural characteristics of the SPI2-encoded T3SS have not been revealed so far. This system is required for the translocation of a set of effector proteins that modulate host cell functions in order to avoid antimicrobial activities of the host cell and to promote intracellular proliferation. SPI2 genes are induced if Salmonella resides in the phagosome of a host cell. SPI2 expression can also be induced in vitro by nutritional limitation (Deiwick et al, 1999). Under in vitro conditions, secretion of substrate proteins of the SPI2-encoded T3SS (SPI2-T3SS) can be induced by growth in minimal media of acidic pH (Beuzon et al, 1999). Interestingly, in vivo acidic vacuolar pH is important for the intracellular survival of Salmonella (Rathman et al, 1996). Previous studies indicated that after secretion in vitro, several SPI2-encoded substrate proteins are predominantly located on the outer surface of the bacterial cell and can be recovered by extraction with organic solvents or by mechanical shearing (Beuzon et al, 1999; Klein and Jones, 2001; Nikolaus et al, 2001). Use of antibodies labeled with a fluorescence marker indicated that the secreted protein is not uniformly distributed over the bacterial surface, but predominantly localized to one pole of the bacterial cell (Nikolaus et al, 2001). We further observed that SPI2 effectors of the ‘Salmonella-translocated effector' (STE) family of proteins are also localized on the bacterial surface after secretion in vitro (Hansen-Wester et al, 2002). Based on these observations, we set out to characterize the secretion of SPI2 substrate protein in more detail. Here, we used high-resolution field emission scanning and transmission electron microscopy to investigate structures that were formed by substrate proteins of the SPI2-T3SS. We further followed the fate of SPI2 translocon proteins during the intracellular growth of Salmonella enterica serotype Typhimurium (S. Typhimurium) in eukaryotic host cells and investigated the role of vacuolar pH for SPI2 function. Our data provide the first detailed insight into the structure and function of the SPI2-T3SS.
Results
Formation of surface structure by the SPI2-encoded T3SS
Based on previous observations on the secretion by the SPI2-T3SS, we set out to investigate the localization of secreted SPI2 proteins in more detail using high-resolution field emission scanning electron microscopy (FESEM) and immunofluorescence microscopy. S. Typhimurium wild type (WT) and an SPI2 null mutant strain (ssaV) were grown under conditions inducing expression of the SPI2-T3SS (PCN-P, pH 7.4), or both expression and secretion (PCN-P, pH 5.8, or F media, pH 5.0), and bacteria were analyzed by FESEM (Figure 1). After growth in minimal media at neutral pH, long polar structures with a rigid appearance were frequently observed on the surface of Salmonella WT and ssaV strains. These structures resembled long polar fimbriae (Bäumler and Heffron, 1995). In addition, thin, long and flexible structures were detected that represent flagella. Both of these long surface structures were absent if bacteria were grown in minimal media at acidic pH under conditions that induced T3SS secretion.
Figure 1.
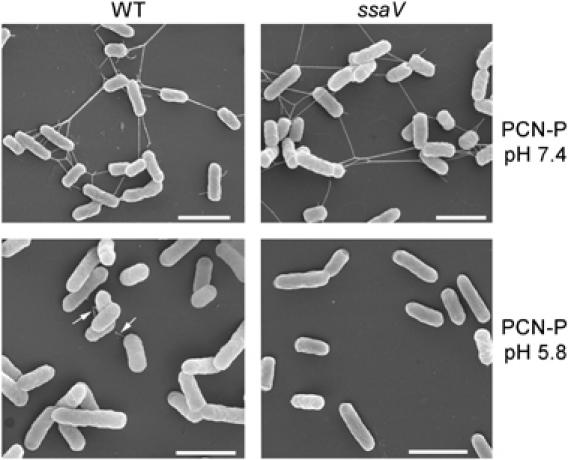
SPI2-dependent formation of surface structures in vitro. S. Typhimurium WT and a mutant strain deficient in the SPI2-encoded T3SS (ssaV) were grown in minimal media that induced expression of SPI2 (PCN-P, pH 7.4) or in minimal media inducing SPI2 expression and secretion of substrate proteins (PCN-P, pH 5.8). The bacteria were grown for 16 h with aeration, and aliquots of the cultures were taken and prepared for FESEM analyzes. The appearance of SPI2-dependent polar appendages is indicated by arrows. Similar appendages were observed for S. Typhimurium WT grown in low magnesium minimal media at pH 5.0 (data not shown). Scale bars represent 2 μM.
In contrast, surface structures with irregular appearance were detected (arrows) on the surface of WT S. Typhimurium grown at acidic pH (PCN-P, pH 5.8). These structures were never observed on bacteria grown at neutral pH, or in acidic minimal media that did not induce SPI2 expression. The SPI2 null mutant strain also did not express such surface structures. The SPI2-dependent appendages were mainly observed at one pole of the cell and most bacteria possessed only a single appendage.
We next analyzed the effect of mutations in genes encoding various T3SS components on the formation of these appendages (Figure 2). Strains with null mutations in the SPI1-encoded T3SS developed appendages that were indistinguishable from those of the WT strain (data not shown). In contrast, appendages were not detected on the surface of mutant strains deficient in the SPI2-encoded regulatory system SsrAB or in ssaC, encoding another structural component of the SPI2-T3SS (not shown).
Figure 2.

Effect of mutations in genes encoding T3SS components and secreted proteins on the formation of appendages. (A) Genetic organization of SPI2 genes investigated in this study. Genes encoding translocon subunits (SseB, C, D) and translocated effectors (SseF and G) are indicated by black and gray symbols, respectively. (B) Strains deficient in a structural component of the T3SS (ssaV) or in various translocon or effector proteins were grown in PCN-P media at pH 5.8 and analyzed by FESEM. Appendages were missing on the ssaV strain. Note the appearance of regular needle-like structures for the sseB strain (inset shows magnified view) and of irregular appendages for the other mutant strains. Plasmid complementation of the sseB strain was performed (sseB com). Scale bars represent 1 μm, 100 nm (inset sseB) and 250 nm (inset sseB com). (C) Mutant strains deficient in ssaG or ssaI, and mutant strains harboring a plasmid expressing ssaGHI for complementation (com) were analyzed. Appendage formation was also absent in an ssaH mutant strain (not shown), but was not restored in the complemented strains. Scale bars represent 5 μm for overview images and for the complemented strains 1 μm and 250 nm in the inset.
SseB, SseC and SseD have been identified as secreted proteins with translocon functions for the SPI2-T3SS (Nikolaus et al, 2001). SseB shares similarity with EspA, the structural component of the EPEC filament, SseD has limited sequence similarity to EspB and SseC is a member of the YopB family of translocon proteins involved in pore formation in the target membrane (Hensel et al, 1998). SseF and SseG are SPI2-encoded translocated effector proteins (Kuhle and Hensel, 2002). Mutations in sseC, sseD, sseF or sseG had no detectable effect on the morphology or frequency of surface structures. Surface structures were also formed by the sseB strain, but these had a rather regular appearance. Complementation of the sseB strain by plasmid-borne sseB restored the formation of appendages with an irregular appearance. SPI2 genes ssaG, ssaH and ssaI encode small proteins of 7.9, 8.1 and 9.0 kDa, respectively, with unknown functions. SsaG has low sequence similarity to EscF of EPEC and Citrobacter rodentium. Strains deleted in ssaG, ssaH or ssaI had a strong defect in intracellular replication and were unable to translocate effector proteins (Supplementary Figure 1). FESEM analyses indicated the absence of surface structures for the ssaG, ssaH and ssaI strain (Figure 2C). The ssaG and ssaI strains, but not the ssaH strain, could be complemented by a plasmid expressing ssaGHI. These observations indicate that translocon proteins SseB, SseC or SseD are not required for appendage formation but suggest a function for SsaG, SsaH and SsaI in the formation of a functional T3SS and of surface structures.
To investigate the kinetics of appendage formation, the appearance of SPI2-dependent appendages was analyzed at various time points after growth of the bacteria in PCN-P media at pH 5.8 (Figure 3). Surface structures were detectable at 5 h after inoculation, but not visible at earlier time points. The frequency of bacterial cells positive for appendages did not increase over time, and the number of appendages per bacterial cell remained similar. We also observed that aggregates of appendages were present in cultures grown for more than 10 h under inducing conditions. The aggregates were partially present on the surface, and also detached from the bacteria and formed massive complexes (data not shown). Negative staining of bacteria indicated that appendages were composed of an inner cylindrical structure (arrowheads in Figure 3B), which was sheathed by irregularly distributed, most likely proteinaceous material. The diameter of the inner structure was 10±0.7 nm. The length of about 160 nm for the cylindrical inner structures of sseB (Figure 2) and WT strains (Figure 3B, lower panel) was determined. For the sheath structures, highly variable diameters of 30–70 nm were estimated. Also, the length of the sheathed structures was highly variable at a given time point (see Supplementary Figure 2) and appeared to increase with longer incubation times.
Figure 3.
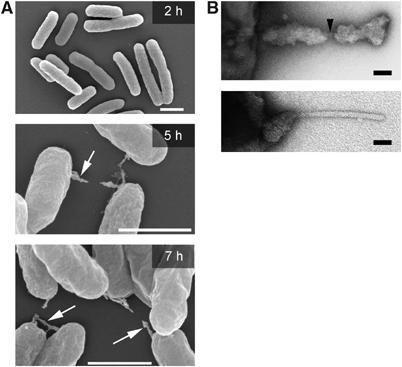
(A) Kinetics of formation of SPI2-dependent surface structures. S. Typhimurium wild type was cultured overnight in PCN media at pH 7.4. Bacteria were harvested by centrifugation and used for inoculation of PCN-P media at pH 5.8. Incubation at 37°C was continued with aeration and aliquots of the culture were taken at various time points for FESEM analysis. Appendages are indicated by arrows. Scale bars represent 2 μm. (B) Higher magnification view of negatively-stained single appendages revealing the inner cylindrical needle-like structure (arrow) and different extent of proteinaceous sheaths. Bacteria were cultured for 9 h. Scale bars correspond to 50 nm (upper panel) and 25 nm (lower panel).
Composition of SPI2-encoded appendages
We previously observed that immunostaining of bacteria grown under conditions inducing SPI2-dependent secretion with anti-SseB antisera resulted in a punctate staining on one pole of the bacterial surface (Nikolaus et al, 2001). We used polyclonal antibodies raised against recombinant SseB and SseC to analyze if these proteins are present in the structures described above. Immunogold labeling indicated that SseB (Figure 4A) and SseC (Figure 4C) were present on SPI2-dependent surface structures. Both proteins were distributed on the surface of the appendages, but there was no polarity detectable for either of the two proteins. Also, double labeling experiments (Figure 4D) indicated a random distribution on the surface of the appendages for these two proteins. Interestingly, the appearance of inner cylindrical structures was observed that were not labeled by immunogold staining for SseB or SseC.
Figure 4.
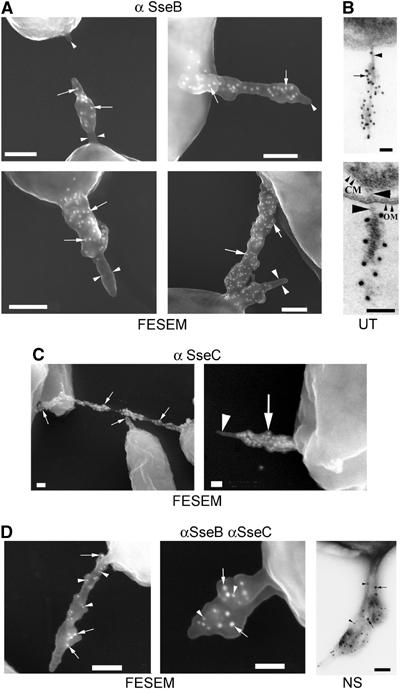
Detection of SPI-2-encoded proteins by immunoelectron microscopy. S. Typhimurium wild type was grown overnight in PCN-P media at pH 5.8 and cells were processed for immuno-EM. Fixed bacteria were incubated with antisera against SseB (A) or SseC (C) and protein A-coated gold-particles with a diameter of 15 nm. The distribution of the gold-particles (white dots) is detectable over the sheathed parts of the appendage (arrows). Both antibodies did not label the inner cylindrical structures of the appendages (arrowheads). (B) Immunogold labeling of SseB in ultrathin sections (UT). Note that the unlabeled inner structure (arrowhead) spans the entire cell envelope (CM, cytoplasmic membrane; OM, outer membrane). Since the bacteria were pre-embedded, immunogold labeling of SseB (dark dots) is only detectable on the bacterial exterior. (D) For double labeling, antibodies against SseB (arrowheads) and SseC (arrows) were detected with protein A-coated gold-particles of 10 nm for SseB and 15 nm for SseC. A negative-stained (NS) sample of a double-labeled bacterium is shown. Scale bars represent 100 nm.
To gain further insight into the SPI2-dependent surface structures, immunogold labeling of ultrathin sections of slightly plasmolyzed bacteria was performed (Figure 4B). These analyses revealed that the inner cylindrical structure of the appendages was inserted in the cytoplasmic membrane and spanned the periplasmic space and the outer membrane (arrowheads in Figure 4B). The sheathed material containing SseB, as indicated by immunogold labeling, was only present on the part exposed to the medium phase. Although secreted translocon proteins appeared randomly distributed, crosslinking experiments revealed the formation of dimers of SseB that was dependent on secretion (Supplementary Figure 3).
Location of SseB, SseC and SseD on intracellular Salmonella
We analyzed the localization of SseB, SseC and SseD on Salmonella residing within Salmonella-containing vacuoles (SCV) of infected host cells. Previous experiments demonstrated that SseB, SseC and SseD function as translocon for the translocation of effector proteins into host cells (Nikolaus et al, 2001). Antisera raised against SseB, SseC and SseD were suitable to detect surface appendages formed during growth in vitro, and direct labeling of one of the antibodies allowed double immunostaining of SseC and SseD in individual appendages (Figure 5A).
Figure 5.
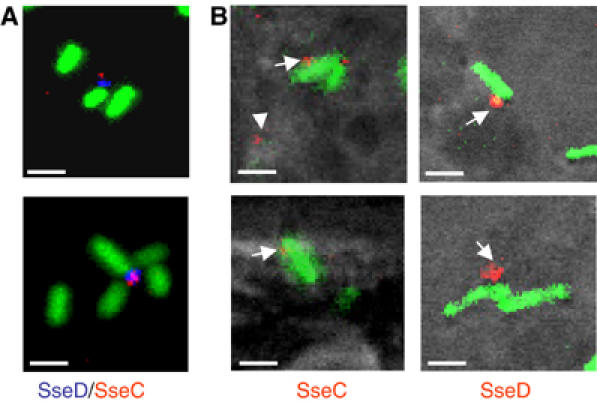
Detection of SPI2-encoded translocon proteins in vitro and in vivo using immunofluorescence. (A) S. Typhimurium WT harboring pFPV25.1 for the constitutive expression of GFP was grown in PCN-P at pH 5.8 for 7 h. Bacteria were harvested and processed for immunostaining. SseD was detected after incubation with rabbit antiserum against recombinant SseD followed by incubation with a Cy5-labeled secondary antibody (blue). SseC was detected by incubation with rabbit antiserum directly labeled with TRITC (red). (B) RAW macrophages were infected with Salmonella WT harboring pFPV25.1 12 h after infection, cells were fixed and immunostained for SseC and SseD (red). Note the appearance of SseC and SseD on the surface of intracellular bacteria (arrows), and also distant to the bacteria (arrowheads). Scale bars represent 1 μm.
To investigate the location of translocon proteins in vivo, the macrophage-like cell line RAW264.7 was infected with S. Typhimurium WT and immunostaining was performed. We detected SseC and SseD on the surface of intracellular Salmonella. As observed under in vitro conditions, a punctate appearance of SseC or SseD on one pole of the cell was also found for intracellular bacteria (Figure 5B). This observation indicates that only single or a low number of translocon structures were assembled by S. Typhimurium within the SCV.
The effect of mutations in various SPI2 genes encoding the T3SS, translocon proteins or STE gene sifA on the secretion of SseC and SseD by intracellular Salmonella are summarized in Table I. SseC and SseD were not detectable in host cells infected with strains deficient in regulator SsrB or the structural T3SS component SsaV, but were also absent in cells infected with spiC or sseB strains. In line with observations on the secretion in vitro (Nikolaus et al, 2001), an sseC mutant strain was able to secrete low amounts of SseD and vice versa. Lack of SifA, an effector required for induction of tubular aggregates of endosomes and maintenance of the SCV in Salmonella-infected cells (Beuzon et al, 2000), had no effect on the secretion of SseC and SseD.
Table 1.
Detection of translocon proteins SseC and SseD in RAW macrophages after infection with S. Typhimurium wild-type and various mutant strains
| Genotype | SseC | SseD |
|---|---|---|
| Wild type | + | + |
| sseC | − | (+) |
| sseD | (+) | – |
| ssaV | − | − |
| ssrB | − | − |
| sseB | − | − |
| sifA | + | + |
|
spiC |
− |
− |
| +, protein detectable; (±), weak signal; −, no signal. | ||
The distribution of translocon proteins and translocated effector proteins in Salmonella-infected macrophages was compared. We previously used an HA epitope-tagged derivative of effector protein SseJ to follow SPI2-dependent translocation in macrophages (Jantsch et al, 2003). Immunostaining for SseJ-HA and SseC or SseD revealed that the translocon proteins were predominantly located on the surface of S. Typhimurium, while the translocated effector protein appeared distant from the bacteria (data not shown).
We also investigated the appearance of SPI2-dependent surface structures in vivo by EM of ultrathin sections (Figure 6). On a few occasions, where the intracellular bacteria were not entirely enclosed by the membrane of the SCV, appendages on the surface of wild-type bacteria were detectable (Figure 6A, B, C and E). Such structures were never observed on the surface of SPI2 null mutant strains. The shaft of the appendages (Figure 6B, indicated by arrowhead) had a regular appearance and a diameter similar to that of the unsheathed inner structures shown in Figure 2B. The tips of the appendages had slightly larger diameters and the sheathed part of the appendage is indicated by an arrow in Figure 6B. Intracellular Salmonella are often in tight contact with the SCV membrane (Figure 6C). Occasionally, projections of the bacterial envelope were observed that were enclosed by the SCV membrane (Figure 6C, arrows). Immunogold-labeling indicated the presence of SseB in the bacterial cytoplasm and the distal part or the appendage (Figure 6D and E).
Figure 6.
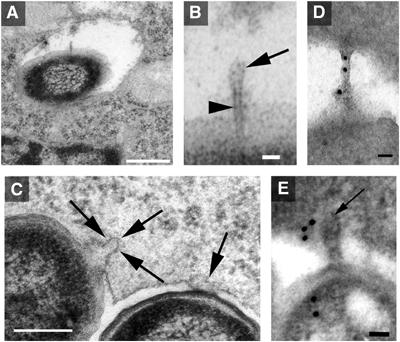
Ultrathin section analysis of intracellular Salmonella. RAW264.7 macrophages were infectedz with S. Typhimurium WT, and 7 h after infection, cells were fixed, dehydrated and embedded in epoxy resin. Analysis of ultrathin sections revealed the expression of appendages that were distant to the phagosomal membrane (A, B) or connecting the bacterial envelope to the phagosomal membrane (C–E). Depicted in (B) is a structure that exhibits a needle-like inner core (arrowhead) and a sheathed tip of the appendage (arrow). Immunogold labeling for SseB was performed (D, E). Scale bars represent 500 μm (A), 250 nm (C), 50 nm (D, E) and 25 nm (B).
In vivo function of the SPI2-encoded T3SS
The secretion of substrate proteins of the SPI2-encoded T3SS can be induced in vitro by growth in media with acidic pH (Beuzon et al, 1999; Klein and Jones, 2001; Nikolaus et al, 2001). We have recently reported that acidic pH is required for the assembly of a functional T3SS, especially of the secretion pore in the outer membrane composed of an oligomer of SsaC (Rappl et al, 2003). We reasoned that these media conditions mimic the acidic pH of the SCV in vivo and that acidic pH is required for the assembly of a functional T3SS. It has also been proposed that acidic pH is a signal that triggers the expression of SPI2 genes (Cirillo et al, 1998; Garmendia et al, 2003). To test these hypotheses, we analyzed the secretion of SseC and SseD by intracellular bacteria in the presence or absence of bafilomycin A1 (BAF), an inhibitor of the V-ATPase that is used to block acidification of phagosomes. In addition, BAF also affects the formation of vesicles, alters the intraphagosomal environment and induces apoptosis (Tapper and Sundler, 1995; Yamamoto et al, 1998). In order to separate the effects of this inhibitor, we first determined the minimal concentration required to inhibit phagosomal acidification under our assay conditions. We found that concentrations of 10–50 nM BAF were sufficient to fully prevent phagosomal acidification as indicated by acridine orange (AO) staining of living cells (Figure 7A). Higher concentrations of BAF resulted in altered morphology and detachment of cells, while a low proportion of vacuoles remained acidified in the presence of 5 nM BAF (Supplementary Figure 4). The addition of BAF caused reduced intracellular proliferation of wild-type S. Typhimurium at concentrations of 5 nM and full inhibition at 10 nM or higher (Supplementary Figure 5).
Figure 7.
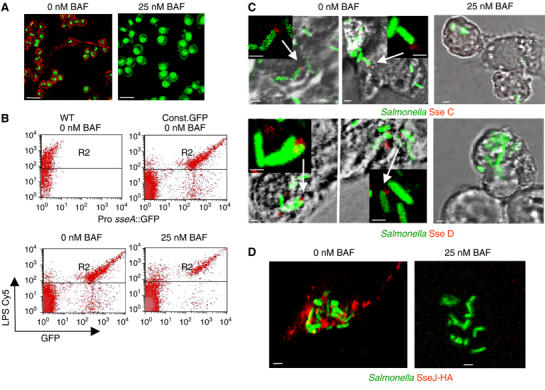
Secretion and translocation of SPI2 proteins requires acidic phagosomal pH. (A) Phagosome acidification is inhibited by low concentrations of BAF. Various concentrations of BAF (5–100 nM), or an equal amount of the solvent DMSO were added to RAW macrophages. After incubation for 1 h, AO was added to a final concentration of 1 μM and the fluorescence of living cells was analyzed by confocal microscopy. Red fluorescence indicating acidic vesicles was absent at BAF concentrations of 10 nM or higher (only shown for 25 nM BAF, for further details, see Supplementary Figure 4). Scale bars represent 16 μm. (B) Effect of BAF on expression of an SPI2 reporter gene by intracellular S. Typhimurium. RAW macrophages were infected with S. Typhimurium WT without plasmid or harboring plasmid pLS824 (Pro sseA::GFP fusion) or pFVP25.1 (constitutive GFP expression). After infection, 25 nM BAF was added and maintained throughout the experiment. Host cells were lysed 16 h after infection and released intracellular bacteria were labeled with antibody against LPS as primary antibody and a Cy5-conjugated secondary antibody. The GFP fluorescence of about 2000 bacteria was detected in gate R2. As the number of recovered bacteria was highly reduced in host cells that were exposed to BAF, the number of particles analyzed by FACS was increased about 10-fold. (C) Effect of BAF on secretion of translocon proteins by intracellular Salmonella WT. RAW macrophages fixed 16 h after infection and processed for immunostaining. The localization of SseC and SseD was analyzed as described in the legend of Figure 5. (D) Effect of BAF on translocation of the SPI2 effector protein SseJ. RAW macrophages were infected with S. Typhimurium WT harboring p2777 for expression of SseJ-HA and constitutive expression of GFP. After infection, 25 nM BAF or equal amounts of solvent were added. At 12 h after infection, cells were fixed and processed for immunostaining using antibodies against the HA-epitope tag and a Cy3-labeled secondary antibody (red). Representative intracellular bacteria are shown. Scale bars represent 16 μm (A) and 1 μM (C, D).
We next analyzed the effect of BAF on the expression of SPI2 genes by intracellular Salmonella (Figure 7B). The low copy number reporter plasmid pLS824 harbors a fusion of promoter sseA to GFP that is induced in an SsrAB-dependent manner in PCN-P minimal media and by intracellular Salmonella (see Supplementary Figure 6). Salmonella WT harboring plasmid pLS824 were used to infect cells and GFP fluorescence of individual bacteria was quantified after release from the host cells 16 h after infection. For a correct determination of the levels of reporter expression of individual bacteria by flow cytometry, the recovered bacteria were labeled with an antibody against LPS, and the GFP fluorescence of the LPS-positive population was measured. In contrast to analysis of GFP levels in ‘bacteria-sized particles', this procedure is not biased by different extents of intracellular replication of Salmonella under various assay conditions. We observed that expression of the SPI2 reporter gene fusion was only slightly reduced in the presence of various concentrations of bafilomycin (Figure 7B). Although the addition of 10 nM BAF or higher concentrations resulted in reduced numbers of intracellular bacteria, the GFP fluorescence of individual intracellular bacteria was identical to the GFP levels of bacteria recovered from nonbafilomycin-treated host cells. We did not observe the appearance of a bacterial population that had reduced levels of GFP expression after treatment with bafilomycin. This observation indicates that inhibition of phagosome acidification did not affect the expression of SPI2 genes by intracellular Salmonella.
Finally, the effect of BAF on translocon formation and translocation of SPI2 effectors by intracellular Salmonella was investigated. Immunofluorescence analyses indicated that the secretion of SseC and SseD was not detectable in the presence of 25 nM BAF (Figure 7C). The effector SseJ is translocated by the SPI2-T3SS by intracellular Salmonella (Kuhle and Hensel, 2002). In line with the absence of translocon proteins, the effector SseJ was not translocated in infected host cells exposed to BAF (Figure 7D).
Taken together, these data support our previous observation that acidic pH is a stimulus for the assembly of a functional T3SS (Rappl et al, 2003) and indicate that acidic phagosomal pH is required for the assembly of the SPI2 translocon and the translocation of SPI2 effectors into the host cells.
Discussion
We have identified a novel appendage structure on the surface of S. enterica serotype Typhimurium that is assembled dependent on the function of the SPI2-T3SS. Based on our observations, we propose that an ordered, filamentous structure is formed. The SPI2-dependent appendages consist of an inner cylindrical structure with a regular diameter of about 10 nm. The diameters of the needle-like structures of the other T3SS are in a similar range, that is, the SPI1-T3SS with 8 nm (Kubori et al, 2000) and the Shigella secreton (Tamano et al, 2000) with 8.4 nm. The EPEC T3SS appendages consist of a needle structure formed by EscF (Sekiya et al, 2001) and the EspA filament (Daniell et al, 2003), which have diameter of 8.8 or 12 nm, respectively.
When the secretion of substrate proteins of the SPI2-encoded T3SS was induced in vitro by specific growth conditions, the needle-like structures were sheathed with an irregular layer of secreted protein. The molecular appearance of the SPI2-dependent surface structures was distinct from the filamentous structures of EPEC- or the SPI1-dependent ‘needle complexes' of S. Typhimurium. The observation of a sheathed structure for the SPI2-encoded T3SS is reminiscent of the hrp pilus assembled by the T3SS of the plant pathogen Pseudomonas syringae (Jin and He, 2001), and also to a surface structure assembled by a T4SS encoded by the cag pathogenicity island of Helicobacter pylori (Rohde et al, 2003; Tanaka et al, 2003). The irregular appearance of surface structures indicates that secreted proteins are distributed along an ordered basal structure. Sheathed structures observed in vitro are likely a consequence of prolonged secretion and the lack of contact to a target membrane. Based on sequence similarity to T3SS substrate proteins of EPEC, it has been proposed that SseB, SseC and SseD are translocon subunits that are involved in the pore formation in the target membrane, that is, the phagosomal membrane. In the absence of such a membrane, these proteins will not adopt their proper conformation and appear randomly distributed in the form of a sheath on the needle-like structure.
SseB is neither required for the formation of the needle-like structure nor the formation of an ordered filament. Thus, SseB is, despite sequence similarity, functionally different to EspA of EPEC. EspA is the monomer of the ‘Esp pilus', a cylindrical tube with a regular diameter of 12 nm (Sekiya et al, 2001; Daniell et al, 2003). The phenotypes of sseB, sseC and sseD mutant strains match previous observations on the secretion and surface location of the translocon proteins. In the background of an sseB mutation, SseC and SseD were secreted, but no longer associated with the bacterial cell surface (Nikolaus et al, 2001). In the absence of SseB, in vitro-grown bacteria exhibit needle-like structures that are not sheathed with secreted protein. A similar situation appears to apply to intracellular bacteria. The sseB mutant secreted SseC and SseD, but the proteins had a rather diffuse location (data not shown). This may indicate that the proteins are secreted into the phagosomal lumen but cannot assemble a translocator in the SCV membrane in the absence of SseB.
The phenotypes of mutant strains lacking SsaG, SsaH or SsaI suggest that these proteins are involved in the formation of the filament and might represent monomers of the cylindrical inner structure. The inner structure might be composed of functional homologs of EscF (Sekiya et al, 2001), YscF (Hoiczyk and Blobel, 2001) or PrgI and PrgJ (Kimbrough and Miller, 2000; Kubori et al, 2000) that form needle-like structures in the EPEC, Yersinia and Salmonella SPI1 T3SS, respectively. Such homologs have not been identified in the SPI2 system, but ssaG, ssaH and ssaI may be interesting candidates as these genes are clustered with other apparatus genes and encode small proteins. SsaG is similar to EscF of EPEC, a protein that was shown to link the T3SS components in the cell envelope to the EspA filament (Sekiya et al, 2001; Wilson et al, 2001). However, we have not been able to detect any of the three small proteins in secreted protein fractions or material recovered from the surface of secreting bacteria and further work has to reveal the role of each protein.
Filamentous surface structures have been observed for a variety of T3SS. However, the SPI2-encoded structure is distinct in the formation of appendages singly or with few copies at one pole of the cell. T3SS-dependent surface structures encoded by the SPI1 of S. enterica or the mxi/spa genes of Shigella spp are present in as many as 200 copies per cell that are distributed over the bacterial surface (Kubori et al, 1998; Blocker et al, 1999). The observation that the SPI2-encoded surface structure is present in single or low copy number might reflect the different functional requirements of the T3SS involved in invasion (e.g. SPI1) or intracellular pathogenesis (such as SPI2). The SPI1-T3SS complexes are formed by extracellular Salmonella and are required to trigger invasion. A large number of injectisomes likely increases the probability of establishing contact between extracellular bacteria and host cells for the injection of a set of effector proteins. There is evidence that translocation of effector proteins only takes place by the injectisomes that have established contact with the host cell membrane to avoid shedding of effector proteins (Rosqvist et al, 1994). In contrast, the SPI2-T3SS is activated when Salmonella are inside the SCV. Here, the bacteria are enclosed by membranes and the likelihood of establishing contact between the injectisome and the target membrane is high (see Figure 6C). Consequently, a single injectisome could be sufficient to translocate SPI2 effectors. This difference may explain why different numbers of the SPI1- and SPI2-encoded injectisomes are assembled.
A further unique feature of the SPI2-T3SS is the requirement for acidic pH to trigger formation of the surface structure and translocation of effector proteins. For the majority of pathogens, the molecular signals triggering expression of T3SS genes are unknown. Furthermore, the identification of the stimuli for secretion or translocation of substrate proteins has been elusive for most T3SS. Previous studies (Beuzon et al, 1999; Rappl et al, 2003) as well as the present report strongly suggest a role of acidic pH as a stimulus for the SPI2-encoded T3SS. This stimulus, however, is required for the function of the T3SS rather than for inducing gene expression.
The role of the intraphagosomal pH in intracellular survival of Salmonella and regulation of gene expression is controversial (Rathman et al, 1996; Steele-Mortimer et al, 2000). To address this aspect, bafilomycin (BAF) is frequently used to inhibit phagosomal acidification. However, secondary effects of bafilomycin also involve altered maturation of phagosomes. Careful investigation indicated that BAF at a concentration of 25 nM, which was sufficient to inhibit phagosomal acidification, also inhibited translocation of SPI2 effector proteins and secretion of translocon subunits by intracellular Salmonella. However, we found that a concentration of 25 nM BAF had no significant effect on expression of a representative SPI2 gene. Higher concentrations of BAF resulted in reduced SPI2 expression. At such concentrations, morphology of the macrophages was also altered, indicating that normal cellular processes were significantly affected.
Our observations on the effect of phagosomal pH are in line with our previous finding that acidic pH is required for functional assembly of the SPI2-T3SS, but not for the expression of SPI2 genes (Rappl et al, 2003). The intraphagosomal signals that are sensed by Salmonella and required for the induction of SPI2 gene expression have not been identified. Work of several groups have indicated that nutritional limitation of the growth media is sufficient for induction (Beuzon et al, 1999; Deiwick et al, 1999; Miao et al, 2002). Whether these limitations are also present in the SVC has to be studied in greater detail.
We have previously shown that three secreted substrate proteins, SseB, SseC and SseD, are absolutely required for the translocation of effector proteins of the SPI2 system. Analyses of the T3SS of other pathogens also indicated that a subset of secreted proteins is required for translocation of effector proteins into the cytoplasm of the host cell. Future work has to reveal the interactions of these components of the SPI2-T3SS with host cell structures, for example, the molecular recognition of the endosomal membrane. It will also be of interest to understand the macromolecular organization of the SPI2 surface structures and their function as translocon in more detail.
In conclusion, we have shown the molecular analyses of the SPI2-T3SS in vitro and in vivo and observed that this structure differs from other T3SS structures. These structural characteristics might reflect the adaptation of Salmonella to the intracellular niche.
Materials and methods
Bacterial strains and growth conditions
S. Typhimurium strain NCTC 12023 was used as a wild-type and various mutant strains derived from S. Typhimurium 12023 used in this study are listed in Table II. Bacteria were routinely cultured in Luria broth (LB). For the induction of the expression of SPI2 genes, cultures were grown in minimal medium containing 25 mM phosphate (PCN) or 360 μM phosphate (PCN-P) as described previously (Deiwick et al, 1999). To induce expression and secretion, PCN-P media were adjusted to pH 5.8. Alternatively, N-salts minimal media containing 30 μM MgSO4 (F media) adjusted to pH 7.0 or 5.0 were used as described before (Deiwick et al, 1999). Bacterial cultures were grown at 37°C in glass test tubes with aeration in a ‘roller drum' (New Brunswick Scientific).
Table 2.
Bacterial strains used in this study
| Designation | Relevant characteristics | Reference |
|---|---|---|
| S. enterica serovar Typhimurium strains | ||
| NCTC 12023 | Wild type | Lab stock |
| P2D6 | ssaV::mTn5 | Shea et al (1996) |
| P8G12 | ssrB::mTn5 | Shea et al (1996) |
| EG10128 | spiC::aph | Uchiya et al (1999) |
| MvP101 | sseD::aphT | Medina et al (1999) |
| HH102 | sseB::aphT | Hensel et al (1998) |
| HH103 | sseB::aphT, psseB | Hensel et al (1998) |
| HH104 | sseC::aphT | Hensel et al (1998) |
| HH107 | sseF::aphT | Hensel et al (1998) |
| HH108 | sseG::aphT | Hensel et al (1998) |
| P3H6 | sifA::mTn5 | Beuzon et al (2000) |
| MvP514 | Δ ssaG | This study |
| MvP515 | Δ ssaH | This study |
| MvP520 | Δ ssaI | This study |
The generation of mutant strains and plasmids for complementation is described in Supplementary Material.
Cell culture and infection studies
The murine monocyte cell line RAW264.7 was cultured in DMEM containing 10% fetal calf serum at 37°C in an atmosphere containing 5% CO2. Prior to infection, RAW264.7 cells were seeded at a density of 5 × 104 cells/well into 24-well plates containing glass coverslips and allowed to adhere.
For infection studies, bacteria were grown in LB for 16 h with aeration, harvested by centrifugation and washed in DMEM. Bacteria were added to RAW264.7 cells at a multiplicity of infection of 10. Infection was synchronized by centrifugation at 1200 g for 5 min. Cultures were incubated for 30 min to allow infection, and noninternalized bacteria were removed by washing three times with PBS. DMEM containing 10 μg/ml gentamicin was added to kill remaining extracellular bacteria and incubation was continued for various periods of time.
Antibodies and immunofluorescence
Antisera raised in rabbits against recombinant SseB, SseC and SseD have been described before (Beuzon et al, 1999; Nikolaus et al, 2001). For some experiments, antibodies against SseD and SseC were directly labeled with TRITC using a ‘Pierce' labeling kit (Perbio, Bonn) according to the manufacturer's instruction. HA epitope-tagged effector proteins were detected using rat monoclonal antibody (Roche, Mannheim).
For detection of SseC and SseD in vitro, bacteria were cultured in LB overnight and 100 μl of the cultures were transferred to PCN-P media at pH 5.8, and incubated at 37°C for various periods of time. Bacteria were recovered by centrifugation at 6000 g for 10 min and fixed for 20 min in PBS containing 3% PFA. Subsequently, bacteria were washed twice with PBS. Care was taken to resuspend the bacteria very gently with the pipette and not by mixing. Antibodies were diluted 1:100 in PBS containing 1% BSA and 5% goat serum and added to 100 μl of the fixed bacteria suspension. The antibody staining was performed with shaking for 2 h followed by Cy3-conjugated anti-rabbit IgG for 1 h. After each antibody incubation, washes were performed six times with PBS. The final pellet was resuspended in 50% glycerol in PBS. A measure of 10 μl of the stained bacteria were mounted on glass slides, covered and sealed and analyzed by confocal laser microscopy (Leica TCS).
For detection of SseC and SseD in Salmonella-infected host cells, RAW cells were seeded onto glass coverslips and infected as described before. The antibodies were diluted in PBS containing 5% goat serum, 2% BSA and 0.1% saponin. For immunofluorescence, cells were fixed for 10 min with 3% PFA in PBS at RT, washed and incubated with 1:100 dilutions of antisera against SseD or SseC overnight at 4°C followed by washing with PBS. The cells were incubated with Cy5-conjugated goat anti-rabbit IgG antibody or directly labeled antibody against SseC and SseD for 1 h. After washing, the cells were analyzed using confocal microscopy and data were processed by using Adobe Photoshop.
Analysis of intracellular expression of SPI2 genes
For the analysis of the effect of bafilomycin on the expression of SPI2 genes by intracellular Salmonella, the procedure described by Jantsch et al (2003) was applied. Briefly, RAW264.7 cells were infected with S. Typhimurium WT harboring plasmid pLS824 for the expression of GFP under control of the promoter of sseA. To inhibit the vacuole acidification, various concentrations of BAF (Sigma, Munich, Germany) or an equal volume of the solvent DMSO were added immediately after infection and maintained throughout the experiment. After completion of the infection period, gentamicin was added to a final concentration of 20 μg/ml gentamicin to kill extracellular bacteria. At 16 h after infection, intracellular bacteria were released by lysis of host cells in PBS containing 0.1% Triton X-100 and incubated for 10 min with agitation to support lysis. Released bacteria were labeled with rabbit test sera against Salmonella LPS (Difco) and a Cy5-conjugated secondary antibody against rabbit IgG. Flow cytometry was performed using a FACScalibur (BD) to determine the GFP fluorescence of the bacterial population.
AO staining as indicator of vacuolar pH
AO vital staining produces a green color at neutral pH. When it accumulates within acidic vesicles, it produces a bright orange to red color. RAW macrophages were treated with various concentration of BAF for 1 h. AO was added at a final concentration of 1 μM to the cells and incubated for 5 min at 37°C. Cells were washed with prewarmed PBS and immediately visualized using confocal microscopy.
Field emission scanning electron microscopy
For morphological studies, bacteria were fixed in 1% formaldehyde in PBS for 1 h on ice and then washed with PBS. Coverslips with a diameter of 12 mm were coated with poly-L-lysine solution (Sigma, Munich, Germany) for 10 min, washed in distilled water and air-dried. A measure of 30 μl of a suspension of fixed bacteria were placed on a coverslip and allowed to settle for 10 min. Coverslips were then fixed in 2% glutaraldehyde in PBS for 5 min at RT and subsequently washed with TE-buffer (20 mM TRIS, 1 mM EDTA, pH 6.9) before dehydrating in a graded series of acetone (10, 30, 50, 70, 90, 100%) on ice for 15 min for each step. Samples in the 100% acetone step were allowed to reach RT before another change in 100% acetone. Samples were then subjected to critical-point drying with liquid CO2 (CPD 30, Balzers, Liechtenstein). Dried samples were covered with a gold film by sputter coating (SCD 40, Balzers) before examination in a field emission scanning electron microscope Zeiss DSM 982 Gemini using the Everhart Thornley SE detector and the inlens detector in a 50:50 ratio at an acceleration voltage of 5 kV.
Immuno-FESEM
Bacteria were fixed with 1% formaldehyde in PBS for 1 h on ice followed by a washing step with PBS containing 10 mM glycine for quenching free aldehyde groups. Samples were then incubated with the affinity-purified (protein A column) specific SseB or SseC polyclonal antibodies for 2 h at 30°C (1:20 dilution of the stock solution of 1.9 mg IgG protein/ml), followed by several washing steps in TE-buffer before incubation with protein A-coated colloidal gold-particles (1:75 dilution of the stock solution for 15 nm gold-particles, 1:200 dilution for the 10 nm gold-particles; British BioCell, Cardiff, UK) for 30 min at 30°C. After washing with PBS, samples were fixed in 2% glutaraldehyde for 15 min at RT, washed in TE-buffer and treated as described above.
For double labeling experiments, samples were incubated first with the specific SseB antibodies followed by protein A gold-particles with a diameter of 10 nm, followed by incubation with 0.2 mg/ml protein A for 15 min at RT and several washing steps in PBS before incubation of the samples with the specific SseC antibodies followed by 15 nm protein A gold-particles. After several washing steps in PBS, samples were placed on poly-L-lysine-coated coverslips, dehydrated and critical-point dried as described above. Samples were then coated with a thin carbon layer (one carbon string) using a Balzers MED 020 (Liechtenstein). The carbon layer method allows for detection of colloidal gold-particles on the bacterial surfaces without charging problems. Samples were examined in the Zeiss DSM 982 Gemini as described above at an acceleration voltage between 3 and 7 kV.
Further details on the procedure for electron microscopy are given in the supplement.
Supplementary Material
Supplementary Material
Supplementary Figure 1
Supplementary Figure 2
Supplementary Figure 3
Supplementary Figure 4
Supplementary Figure 5
Supplementary Figure 6
Acknowledgments
We are grateful to Daniela Jäckel for excellent technical assistance, to Volker Kuhle for support in confocal microscopy and to Garth Abrahams for critical comments on the manuscript. This work was supported by Grants HE 1964/8-1 and 8-2 of the priority program ‘Life inside cells' of the DFG. DC was recipient of a research fellowship of the Humboldt foundation.
References
- Bäumler AJ, Heffron F (1995) Identification and sequence analysis of lpfABCDE, a putative fimbrial operon of Salmonella typhimurium. J Bacteriol 177: 2087–2097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beuzon CR, Banks G, Deiwick J, Hensel M, Holden DW (1999) pH-dependent secretion of SseB, a product of the SPI-2 type III secretion system of Salmonella typhimurium. Mol Microbiol 33: 806–816 [DOI] [PubMed] [Google Scholar]
- Beuzon CR, Meresse S, Unsworth KE, Ruiz-Albert J, Garvis S, Waterman SR, Ryder TA, Boucrot E, Holden DW (2000) Salmonella maintains the integrity of its intracellular vacuole through the action of sifA. EMBO J 19: 3235–3249 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blocker A, Gounon P, Larquet E, Niebuhr K, Cabiaux V, Parsot C, Sansonetti P (1999) The tripartite type III secreton of Shigella flexneri inserts IpaB and IpaC into host membranes. J Cell Biol 147: 683–693 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cirillo DM, Valdivia RH, Monack DM, Falkow S (1998) Macrophage-dependent induction of the Salmonella Pathogenicity Island 2 type III secretion system and its role in intracellular survival. Mol Microbiol 30: 175–188 [DOI] [PubMed] [Google Scholar]
- Daniell SJ, Kocsis E, Morris E, Knutton S, Booy FP, Frankel G (2003) 3D structure of EspA filaments from enteropathogenic Escherichia coli. Mol Microbiol 49: 301–308 [DOI] [PubMed] [Google Scholar]
- Daniell SJ, Takahashi N, Wilson R, Friedberg D, Rosenshine I, Booy FP, Shaw RK, Knutton S, Frankel G, Aizawa S (2001) The filamentous type III secretion translocon of enteropathogenic Escherichia coli. Cell Microbiol 3: 865–871 [DOI] [PubMed] [Google Scholar]
- Deiwick J, Nikolaus T, Erdogan S, Hensel M (1999) Environmental regulation of Salmonella Pathogenicity Island 2 gene expression. Mol Microbiol 31: 1759–1773 [DOI] [PubMed] [Google Scholar]
- Galan JE (2001) Salmonella interactions with host cells: type III secretion at work. Annu Rev Cell Dev Biol 17: 53–86 [DOI] [PubMed] [Google Scholar]
- Garmendia J, Beuzon CR, Ruiz-Albert J, Holden DW (2003) The roles of SsrA–SsrB and OmpR–EnvZ in the regulation of genes encoding the Salmonella typhimurium SPI-2 type III secretion system. Microbiology 149: 2385–2396 [DOI] [PubMed] [Google Scholar]
- Hansen-Wester I, Stecher B, Hensel M (2002) Type III secretion of Salmonella enterica serovar Typhimurium translocated effectors and SseFG. Infect Immun 70: 1403–1409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hensel M, Shea JE, Waterman SR, Mundy R, Nikolaus T, Banks G, Vazquez-Torres A, Gleeson C, Fang F, Holden DW (1998) Genes encoding putative effector proteins of the type III secretion system of Salmonella Pathogenicity Island 2 are required for bacterial virulence and proliferation in macrophages. Mol Microbiol 30: 163–174 [DOI] [PubMed] [Google Scholar]
- Hoiczyk E, Blobel G (2001) Polymerization of a single protein of the pathogen Yersinia enterocolitica into needles punctures eukaryotic cells. Proc Natl Acad Sci USA 98: 4669–4674 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hueck CJ (1998) Type III protein secretion systems in bacterial pathogens of animals and plants. Microbiol Mol Biol Rev 62: 379–433 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jantsch J, Cheminay C, Chakravortty D, Lindig T, Hein J, Hensel M (2003) Intracellular activities of Salmonella enterica in murine dendritic cells. Cell Microbiol 5: 933–945 [DOI] [PubMed] [Google Scholar]
- Jin Q, He SY (2001) Role of the Hrp pilus in type III protein secretion in Pseudomonas syringae. Science 294: 2556–2558 [DOI] [PubMed] [Google Scholar]
- Kimbrough TG, Miller SI (2000) Contribution of Salmonella typhimurium type III secretion components to needle complex formation. Proc Natl Acad Sci USA 97: 11008–11013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein JR, Jones BD (2001) Salmonella Pathogenicity Island 2-encoded proteins SseC and SseD are essential for virulence and are substrates of the type III secretion system. Infect Immun 69: 737–743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knutton S, Rosenshine I, Pallen MJ, Nisan I, Neves BC, Bain C, Wolff C, Dougan G, Frankel G (1998) A novel EspA-associated surface organelle of enteropathogenic Escherichia coli involved in protein translocation into epithelial cells. EMBO J 17: 2166–2176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubori T, Matsushima Y, Nakamura D, Uralil J, Lara TM, Sukhan A, Galan JE, Aizawa SI (1998) Supramolecular structure of the Salmonella typhimurium type III protein secretion system. Science 280: 602–605 [DOI] [PubMed] [Google Scholar]
- Kubori T, Sukhan A, Aizawa SI, Galan JE (2000) Molecular characterization and assembly of the needle complex of the Salmonella typhimurium type III protein secretion system. Proc Natl Acad Sci USA 97: 10225–10230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuhle V, Hensel M (2002) SseF and SseG are translocated effectors of the type III secretion system of Salmonella pathogenicity island 2 that modulate aggregation of endosomal compartments. Cell Microbiol 4: 813–824 [DOI] [PubMed] [Google Scholar]
- Medina E, Paglia P, Nikolaus T, Müller A, Hensel M, Guzman CA (1999) Pathogenicity Island 2 mutants of Salmonella typhimurium are efficient carriers for heterologous antigens and enable modulation of immune responses. Infect Immun 67: 1093–1099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miao EA, Freeman JA, Miller SI (2002) Transcription of the SsrAB regulon is repressed by alkaline pH and is independent of PhoPQ and magnesium concentration. J Bacteriol 184: 1493–1497 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nikolaus T, Deiwick J, Rappl C, Freeman JA, Schröder W, Miller SI, Hensel M (2001) SseBCD proteins are secreted by the type III secretion system of Salmonella pathogenicity island 2 and function as a translocon. J Bacteriol 183: 6036–6045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rappl C, Deiwick J, Hensel M (2003) Acidic pH is required for the functional assembly of the type III secretion system encoded by Salmonella pathogenicity island 2. FEMS Microbiol Lett 226: 363–372 [DOI] [PubMed] [Google Scholar]
- Rathman M, Sjaastad MD, Falkow S (1996) Acidification of phagosomes containing Salmonella typhimurium in murine macrophages. Infect Immun 64: 2765–2773 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rohde M, Puls J, Buhrdorf R, Fischer W, Haas R (2003) A novel sheathed surface organelle of the Helicobacter pylori cag type IV secretion system. Mol Microbiol 49: 219–234 [DOI] [PubMed] [Google Scholar]
- Rosqvist R, Magnusson KE, Wolf-Watz H (1994) Target cell contact triggers expression and polarized transfer of Yersinia YopE cytotoxin into mammalian cells. EMBO J 13: 964–972 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sekiya K, Ohishi M, Ogino T, Tamano K, Sasakawa C, Abe A (2001) Supermolecular structure of the enteropathogenic Escherichia coli type III secretion system and its direct interaction with the EspA-sheath-like structure. Proc Natl Acad Sci USA 98: 11638–11643 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shea JE, Hensel M, Gleeson C, Holden DW (1996) Identification of a virulence locus encoding a second type III secretion system in Salmonella typhimurium. Proc Natl Acad Sci USA 93: 2593–2597 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steele-Mortimer O, St-Louis M, Olivier M, Finlay BB (2000) Vacuole acidification is not required for survival of Salmonella enterica serovar typhimurium within cultured macrophages and epithelial cells. Infect Immun 68: 5401–5404 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamano K, Aizawa SI, Katayama E, Nonaka T, Imajoh-Ohmi S, Kuwae A, Nagai S, Sasakawa C (2000) Supramolecular structure of the Shigella type III secretion machinery: the needle part is changeable in length and essential for delivery of effectors. EMBO J 19: 3876–3887 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka J, Suzuki T, Mimuro H, Sasakawa C (2003) Structural definition on the surface of Helicobacter pylori type IV secretion apparatus. Cell Microbiol 5: 395–404 [DOI] [PubMed] [Google Scholar]
- Tapper H, Sundler R (1995) Bafilomycin A1 inhibits lysosomal, phagosomal, and plasma membrane H(+)-ATPase and induces lysosomal enzyme secretion in macrophages. J Cell Physiol 163: 137–144 [DOI] [PubMed] [Google Scholar]
- Uchiya K, Barbieri MA, Funato K, Shah AH, Stahl PD, Groisman EA (1999) A Salmonella virulence protein that inhibits cellular trafficking. EMBO J 18: 3924–3933 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson RK, Shaw RK, Daniell S, Knutton S, Frankel G (2001) Role of EscF, a putative needle complex protein, in the type III protein translocation system of enteropathogenic Escherichia coli. Cell Microbiol 3: 753–762 [DOI] [PubMed] [Google Scholar]
- Yamamoto A, Tagawa Y, Yoshimori T, Moriyama Y, Masaki R, Tashiro Y (1998) Bafilomycin A1 prevents maturation of autophagic vacuoles by inhibiting fusion between autophagosomes and lysosomes in rat hepatoma cell line, H-4-II-E cells. Cell Struct Funct 23: 33–42 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Material
Supplementary Figure 1
Supplementary Figure 2
Supplementary Figure 3
Supplementary Figure 4
Supplementary Figure 5
Supplementary Figure 6