

Abstract
Bartonella spp. are opportunistic, vectorborne bacteria that can cause disease in both animals and humans. We investigated the molecular occurrence of Bartonella spp. in 634 phlebotomine sand fly specimens, belonging to 44 different sand fly species, sampled during 2017–2021 in north and northeastern Brazil. We detected Bartonella sp. DNA in 8.7% (55/634) of the specimens by using a quantitative real-time PCR targeting the 16S-23S internal transcribed spacer intergenic region. Phylogenetic analysis positioned the Lutzomyia longipalpis sand fly–associated Bartonella gltA gene sequence in the same subclade as Bartonella ancashensis sequences and revealed a Bartonella sp. sequence in a Dampfomyia beltrani sand fly from Mexico. We amplified a bat-associated Bartonella nuoG sequence from a specimen of Nyssomyia antunesi sand fly. Our findings document the presence of Bartonella DNA in sand flies from Brazil, suggesting possible involvement of these insects in the epidemiologic cycle of Bartonella species.
Keywords: Bartonella, phlebotomine, sand flies, Bartonellaceae, Phlebotominae, Carrion’s disease, Oroya fever, verruga peruana, vector, infections, vector-borne infections, Brazil
The genus Bartonella (Alphaproteobacteria: Bartonellaceae) comprises emergent and re-emergent opportunistic bacteria classified in 39 validated species (https://lpsn.dsmz.de/genus/bartonella), some of them capable of causing disease in both animals and humans (1). Mammals (e.g., rodents, bats, cats, dogs, ruminants), including humans, are the main reservoirs for bartonellae. The Bartonella species most often associated with disease in humans are B. henselae (the causative agent of cat scratch disease), B. quintana (the causative agent of trench fever), and B. bacilliformis and B. ancashensis (the causative agents of Carrion’s disease and verruga peruana) (2–4). Other species, including B. clarridgeiae, B. koehlerae, B. vinsonii subspecies berkhoffii, B. elizabethae, and Candidatus Bartonella mayotimonensis, also have been associated with disease in humans, especially in fever of unknown origin and culture-negative endocarditis cases (5,6). Bartonella spp. infect a variety of cells, including erythrocytes, pericytes, endothelial, dendritic, and macrophage cells and are associated with persistent intraerythrocytic bacteremia, suggesting a possible coevolution between these bacteria and their hosts, which may explain their remarkable adaptability to >1 mammal species (2,7,8). The ability of those bacterial species to maintain a persistent bacteremia over time dovetails with their main route of transmission, via bloodsucking arthropods (9). On the basis of molecular epidemiologic surveys and clinical observations, researchers have implicated many hematophagous arthropods in the transmission cycles of Bartonella spp.—mosquitoes (9), biting midges (10), triatomine bugs (11), mites (12,13), and flies (14)—adding to the list of those already identified as competent vectors (fleas, lice, phlebotomine sand flies, ticks) (15,16).
Phlebotomine sand flies (Diptera: Psychodidae: Phlebotominae) comprise >1,060 species, distributed worldwide, especially in tropical and subtropical regions (17). Given their hematophagous feeding habit, female sand flies are insects of considerable public health concern, because they act as vectors in the transmission of different pathogenic agents (bacteria, protozoa, virus), such as Bartonella sp., Leishmania sp., and Phleboviruses (18). Within the Bartonellaceae family, B. bacilliformis is notably the most important agent transmitted by phlebotomine sand flies. This Bartonella species is the causative agent of Carrion’s disease, which can manifest as 2 different syndromes (that can occur sequentially or independently): Oroya fever, characterized by an acute hemolytic anemia with an untreated fatality rate of up to 90%, and verruga peruana (also called Peruvian warts), characterized by a widespread formation of hemangiomas (verrugas) on the skin, along with a persistent bacteremia (3,7). The primary vectors of B. bacilliformis are Pintomyia verrucarum and Lutzomyia peruensis sand flies, which can be found in the Inter-Andean valleys of Peru, at altitudes ranging from 500 to 3,200 meters (7).
Carrion’s disease is a neglected disease because of its focal occurrence (Andean valleys of Peru and, to a lesser extent, in Colombia and Ecuador) and challenges in establishing diagnosis (lack of resources and difficult access to endemic areas). The occurrence of the disease in nonendemic areas and the detection of B. bacilliformis DNA in a growing range of sand fly species suggests that the epidemiologic cycle of Carrion’s disease might involve more sand fly species than first suspected (19). Researchers have detected B. bacilliformis DNA in wild-captured Pintomyia robusta sand flies in the border region between Ecuador and Peru (unpub. data, A.R. Carrazco-Montalvo, https://doi.org/10.13140/RG.2.2.17645.00481) and in Pintomyia maranonensis sand flies in northern Peru (20), but data have yet to confirm their role as vectors. Other possible vectors of Carrion’s disease were noted in Colombia, including Lutzomyia gomezi, Psychodopygus panamensis, Pintomyia serrana, and most notably Pintomyia columbiana sand flies, because of their presence in areas of disease outbreaks (21,22); however, those observations lacked molecular confirmation of the presence of Bartonella DNA in those sand fly specimens. Other suggested vectors for transmission of B. bacilliformis include Lutzomyia pescei, L. noguchii, and L. ayacuchensis sand flies (19,22,23).
Reports have identified Bartonella ancashensis, a species closely related to B. bacilliformis, from blood samples of patients undergoing treatment for verruga peruana in the rural region of Ancash, Peru (24,25). Although that species has not been isolated from blood samples of patients with Oroya fever and seems to be less pathogenic than B. bacilliformis, co-infections can occur, given that the geographic distribution of B. ancashensis overlaps with B. bacilliformis (4,25). Still, no reports have elucidated the involvement of sand flies in the transmission cycle of B. ancashensis.
Brazil has a rich diversity of 304 phlebotomine sand fly species (89 endemic), classified within 19 genera, distributed across all 5 federative regions of Brazil: 218 species in the north, 155 in the midwest, 132 in the southeast, 129 in the northeast, and 49 in the south (26). Despite the diverse phlebotomine sand fly fauna present in Brazil and the proximity to regions endemic for or reporting cases of Carrion’s disease, previous studies have not investigated the occurrence of Bartonella spp. in those dipterans. However, studies from other countries have detected the presence of Bartonella sp. DNA in sand fly species that inhabit Brazil. In Peru, individual female Pintomyia nevesi and Lutzomyia sherlocki sand flies and pooled female Nyssomyia whitmani and Psychodopygus hirsutus sand fly tested positive for Bartonella sp. DNA, phylogenetically associated with B. bacilliformis and Candidatus Bartonella rondoniensis (27). Researchers in Mexico detected Bartonella gltA genotypes, which have been associated with a putative new lineage of Bartonella in sand flies, in females Lutzomyia cruciata and Psathyromyia shannoni sand fly (28). In this study, we investigated the occurrence and molecular identity of Bartonella spp. in sand flies collected in 7 states across the north and northeast regions of Brazil.
Material and Methods
Sand Fly Specimens and Studied Areas
We analyzed sand fly specimens collected during November 2017–December 2021, captured by using Shannon traps or traps designed by the Centers for Disease Control and Prevention set up in ecologic reserves and parks throughout Brazil. Locations included preserved forest areas in the cities of Xapuri and Rio Branco (Acre); Murici Ecologic Station (Alagoas); Pau Brasil National Park (Bahia); Ubajara National Park (Ceará); Tapajós National Forest (Pará); Dois Irmãos State Park (Pernambuco); and Viruá National Park (Roraima). We extracted DNA from dissected sand flies by using Invitrogen TRIzol Reagent (Thermo Fisher Scientific, https://www.thermofisher.com); specimens were without heads and 3 last abdominal segments, which were used for morphologic identification according to previously described taxonomic keys (29). We evaluated DNA concentration and quality (260/280 ratio) with the use of a spectrophotometer (Nanodrop; Thermo Fisher Scientific). We assessed the presence of potential PCR inhibitors by using a conventional PCR based on cytochrome c oxidase subunit 1 (cox1), an endogenous gene among invertebrates. We investigated the occurrence of Bartonella sp. DNA in a total of 634 individual sand fly DNA samples, which we classified into 44 species belonging to 14 genera, obtained from 7 different states across north and northeast Brazil (Table).
Table. Species and number of sand flies, including regions they were collected, after PCR screening for amplification of the endogenous gene cox1 for investigation of Bartonella spp. in phlebotomine sand flies, Brazil*.
| Genera, no. | Species, no. | State of sampling |
|---|---|---|
|
Bichromomyia, 4
|
flaviscutellata, 4 |
Acre |
|
Brumptomyia, 12
|
sp., 12 |
Acre |
|
Evandromyia, 60
|
begonae, 1 |
Acre |
|
infraspinosa, 1 |
Acre |
|
|
saulensis, 14 |
Acre |
|
|
termitophila, 1 |
Acre |
|
|
walkeri, 43 |
Acre |
|
|
Lutzomyia, 46
|
longipalpis, 27 |
Ceará |
|
sherlocki, 19 |
Acre |
|
|
Micropygomyia, 2
|
trinidanensis, 1 |
Acre |
| sp., 1 |
Pará |
|
|
Nyssomyia, 132
|
antunesi, 76 |
Acre |
|
shawi, 15 |
Acre |
|
|
umbratilis, 28 |
Pará, n = 14; Pernambuco, n = 14 |
|
|
whitmani, 12 |
Acre |
|
| sp., 1 |
Acre |
|
|
Pintomyia, 13
|
nevesi, 5 |
Acre |
|
serrana, 6 |
Acre |
|
| sp., 2 |
Bahia |
|
|
Pressatia, 28
|
choti, 15 |
Bahia |
| sp., 13 |
Acre, n = 5; Bahia, n = 8 |
|
|
Psathyromia, 3
|
elizabethdorvalae, 2 |
Acre |
| sp., 1 |
Acre |
|
|
Psychodopygus, 163
|
amazonensis, 3 |
Acre |
|
ayrozai, 40 |
Alagoas, n = 2; Bahia, n = 8; Roraima, n = 30 |
|
|
carreirai, 25 |
Acre, n = 22; Roraima, n = 3 |
|
|
chagasi, 26 |
Alagoas, n = 2; Pará, n = 6; Roraima, n = 18 |
|
|
complexus, 3 |
Alagoas, n = 2; Pará, n = 1 |
|
|
davisi, 30 |
Acre, n = 24; Pará, n = 6 |
|
|
guyanensis, 1 |
Pará |
|
|
hirsutus, 2 |
Alagoas, n = 1; Bahia, n = 1 |
|
|
lainsoni, 2 |
Acre |
|
|
llanosmartinsi, 11 |
Acre |
|
|
paraensis, 17 |
Pará, n = 5; Roraima, n = 12 |
|
|
squamiventris, 1 |
Roraima |
|
| sp., 2 |
Acre, n = 1; Roraima, n = 1 |
|
|
Sciopemyia, 2
|
sordelli, 2 |
Acre |
|
Trichophoromyia, 106
|
ubiquitalis, 1 |
Pará |
|
viannamartins, 65 |
Alagoas |
|
| sp., 40 |
Acre, n = 24; Pará, n = 16 |
|
|
Trichopygomyia, 61
|
dasypodogeton, 2 |
Acre |
|
longispina, 55 |
Bahia |
|
| sp., 4 |
Bahia, n = 2; Roraima, n = 2 |
|
| Viannamyia, 2 | furcata, 2 | Acre |
*All sand fly samples were used for PCR amplification and phylogenetic characterization of Bartonella spp.
Molecular Assays
We conducted molecular screening for Bartonella spp. by using a quantitative real-time PCR (qPCR) based on a 243-bp fragment of the 16S-23S ribosomal DNA internal transcribed spacer (ITS). We performed all reactions in a final volume of 10 µL containing 2× qPCRBIO Probe Master Mix Buffer (PCR Biosystems, https://pcrbio.com), 1.2 µM of each primer and probe, 1 µL of DNA sample, and ultrapurified, sterilized water qsp (Appendix Table 1). For the construction of the standard curve of each reaction, we performed serial dilutions at different concentrations (107–101 copies) of a gBlock gene fragment encoding a 243-bp fragment of the ITS genic region of Bartonella henselae (GenBank accession no. L35101) (Integrated DNA Technologies, https://www.idtdna.com). We also used the gBlocksas positive controls.
We determined the number of gene copies by the formula (XG/μL DNA/[gene block length, bp × 660]) × 6.22 × 1023 × gene copies/μL. We calculated the amplification efficiency (E) according to the slope of the standard curve by using the formula E = 10−1/slope. We evalutated each DNA sample in duplicate and retested in triplicate those samples that presented differences in Cq values >0.5. We considered a Cq value cutoff of 42 for negative results. We carried out reactions in a C1000-CFX96 thermocycler (Bio-Rad Laboratories, https://www.bio-rad.com), using ultrapurified, sterilized water as a negative control.
We noted samples revealed to be positive in the screening qPCR and characterized them by using conventional PCRs based on 8 different molecular markers: gltA (380–400 bp), (767 bp), ftsZ (515 bp), groEL (752 bp), nuoG (346 bp), pap31 (564 bp), rpoB (825 bp), ribC (585–588 bp), and 16S-23S ITS (453–717 bp) (Appendix Table 1).
Purification and Phylogenetic Analyzes
We purified the amplicons obtained in the PCRs by using Wizard SV Gel and PCR Clean-Up System (Promega Corporation, https://www.promega.com). We submitted purified amplicons for Sanger sequencing in both directions (forward and reverse) at the Centro de Estudos do Genoma Humano e Células Tronco (University of São Paulo, São Paulo, Brazil) by using the BigDye Terminator v3.1 Cycle Sequencing Kit (Thermo Fisher Scientific). We assembled a consensus sequence for each sample by using Geneious Prime 2023.2 (Geneious, https://www.geneious.com) and BioEdit 7.2 (30) software programs.
We conducted BLASTn analyzes (https://blast.ncbi.nlm.nih.gov) to produce an alignment for each genetic region, by using the obtained sequences, closely related sequences, and reference sequences previously deposited in GenBank. We created alignments by using the MAFFT version 7 software (https://mafft.cbrc.jp/alignment/server/index.html) and trimmed by using BioEdit 7.2 software (30). For phylogenetic inferences, we performed a maximum-likelihood analysis, with 103 ultraFast bootstrap replicates for each alignment, by using IQTREE2 1.6.12 software (http://www.iqtree.org). We chose the best-fitting evolutionary model for each alignment by using MrModeltest2 2.4 (MrModeltest 2.4; https://github.com/nylander/MrModeltest2) through the PAUP4* Version 4c software (https://paup.phylosolutions.com). We rooted (via outgroups) and edited the resulting phylogenetic trees by using FigTree 1.4.4 (https://tree.bio.ed.ac.uk/software/figtree) and iTOL version 5 (https://itol.embl.de) software programs.
Results
The DNA extraction of individual specimens of sand flies by using TRIzol was satisfactory, yielding DNA concentrations ranging from 1 to 15 ng/µL. We were able to obtain positive samples in the cox1 conventional PCR for all 634 (100%) specimens.
Using the qPCR assay targeting the 16S-23S ITS region, we found that 55 (8.67%) of 634 sand flies tested positive for Bartonella spp. in the molecular screening: 48 from Acre (Nyssomyia antunesi [n = 18], Evandromyia walkeri [n = 7], Trichophoromyia sp. [n = 5], L. sherlocki [n = 4], Nyssomyia shawi [n = 3]; Psychodopygus llanosmartinsi [n = 2]; Psychodopygus davisi [n = 2]; Bichromomyia flaviscutellata [n = 1]; Evandromyia saulensis [n = 1]; Nyssomyia sp. [n = 1]; N. whitmani [n = 1]; P. nevesi [n = 1]; Pintomyia serrana [n = 1]; Viannamyia furcata [n = 1]); 2 from Alagoas (Trichophoromyia viannamartinsi); 2 from Roraima (Psychodopygus squamiventris [n = 1]; Psychodopygus ayrozai [n = 1]); 1 from Bahia (Trichopygomyia longispina); 1 from Ceará (Lutzomyia longipalpis); and 1 from Pará (Psychodopygus paraensis) (Figure 1). The range of Cq values of positive samples was 30.1–41.8. We selected 16 of those samples (based on the lowest PCR Cq values) and obtained 7 readable sequences. On the basis of BLASTn analysis, we confirmed that all 7 sequences corresponded to a Bartonella sp. (Appendix Table 2). However, the sequences were too short (179–222 bp) to be used for phylogenetic inferences. The value of the qPCR efficiency fell in the range of 98.7%–104.8% (mean 102.3, SD 2.29). The R2 value was 0.834–0.986 (mean 0.978, SD 0.05), the Y-intercept range was 34.429–42.318 (mean 37.97, SD, 2.75), and the slope was −3.22 to −3.35 (mean −3.27; SD, 0.05). We were unable to measure the DNA load of positive samples because the Cq difference between replicates was >0.5, possibly because of the Monte Carlo effect (31).
Figure 1.
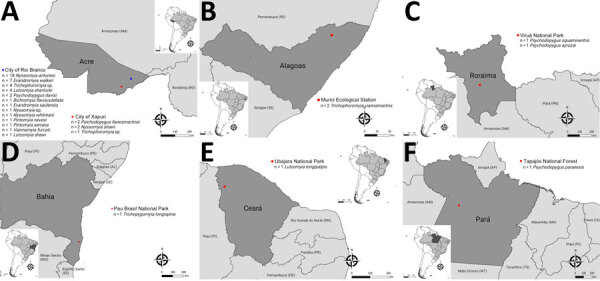
Sampling locations for sand flies that were qPCR positive in the screening for Bartonella spp. DNA from specimens collected in Brazil. A) State of Acre, northern Brazil; B) State of Alagoas, northeastern Brazil; C) State of Roraima, northern Brazil; D) State of Bahia, northeastern Brazil; E) State of Ceará, northeastern Brazil; F) State of Pará, northern Brazil. Dark gray indicates states with positive specimens, and red and blue dots representing the geographic location or city of sampling site. Inset maps show locations of each state in South America.
We performed further molecular characterization (by conventional PCR) of samples that tested positive in the ITS screening qPCR assay and generated amplicons for the following genes: 4 for the gltA, 4 for the ITS, 2 for the ftsZ, 2 for the pap31, 1 for the rpoB, and 1 for the nuoG. Of those, we obtained 2 readable sequences: one 377-bp gltA sequence (GenBank accession no. PP421218) from a L. longipalpis sand fly captured in the state of Ceará, and one 345-bp nuoG sequence from a N. antunesi sand fly from Acre.
The BLASTn analysis demonstrated that the gltA sequence obtained from L. longipalpis sand fly demonstrated >96% identity with B. ancashensis sequences previously obtained from infected humans (GenBank accession nos. CP010401.1, KC886736.1, and KC178618.1). Phylogenetic analyses positioned this sequence in the same subclade as B. ancashensis sequences and with a Bartonella sp. sequence detected in a Dampfomyia beltrani sand fly from Mexico (GenBank accession no. OQ343492.1), with a bootstrap clade support value of 95 (Figure 2).
Figure 2.
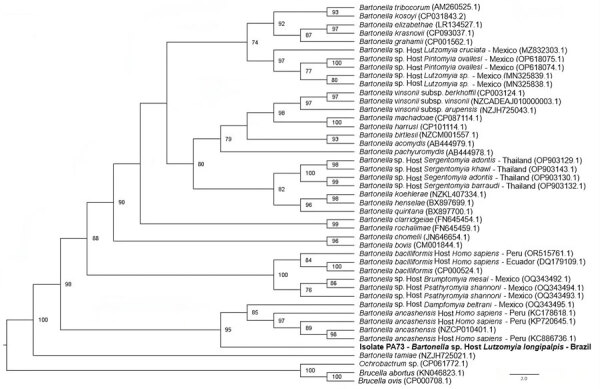
Phylogenetic tree based on an alignment of 380 bp-length of the gltA sequences obtained from phlebotomine sand flies collected in Brazil (bold) and reference sequences. Tree was created using the maximum-likelihood method and generalized time reversible plus invariate sites plus gamma as the evolutionary model. Ochrobactrum sp., Brucella ovis, and Brucella abortus were used as outgroups. Only bootstrap values >70 are shown. GenBank accession numbers are provided in parentheses.
The BLASTn analysis of the nuoG sequence from N. antunesi sand flies indicated a 94.04%–94.47% identity with 2 Bartonella sp. sequences obtained from Pteronotus davyi bats from Guatemala (GenBank accession nos. MN270091.1 and MN270098.1). The few Bartonella nuoG sequences in GenBank and low values of bootstrap clades hampered robust phylogenetic inference by using this molecular marker.
Discussion
We documented the presence of Bartonella spp. DNA in phlebotomine sand flies from Brazil. The occurrence rate observed in this study (55/634 specimens; 8.67%) is similar that reported in southern Mexico, where 2 (8.69%) of 23 specimens were positive (32). Other investigations have reported a range of rates; 2 studies in Peru found positive results in 17 (6.02%) of 228 pools (27) and 2 (2.63%) of 76 pools (20), whereas 2 other studies in Mexico found positive results in 27 (5.08%) of 531 specimens (33) and 11 (2.06%) of 532 specimens (28). Differences in lower occurrence rates can be explained by the wide diversity of sand fly species present in different countries, the method of molecular analysis employed for DNA amplification, and, as illustrated in this study, technical limitations in obtaining phylogenetically relevant Bartonella DNA sequences from these small insects. Although the phlebotomine vectors of Bartonella spp. are very restricted to defined geographic areas, there have been minimal efforts to investigate the prevalence of this bacterial genus in sand flies from regions other than Peru. In our study, the selection of a broad diversity of sand fly species for Bartonella detection can be misleading, since most of the species are not confirmed to be carriers of these bacteria. In this context, we can assume that sand flies that were negative for the Bartonella sp. detection are either unable to host the bacteria or can be considered infrequent vectors. Further studies are necessary to elucidate the role of different sand fly species in the Bartonella epidemiologic cycles.
Although pooling specimens for analysis might have yielded a higher quantity of DNA (ng/µL), we would not have been able to accurately quantify the number of specimens that contained Bartonella sp. DNA, potentially leading to an underrepresentation of PCR-positive sand flies. Therefore, we opted to individually extract the DNA from the specimens by using the TRIzol reagent (Thermo Fisher Scientific), which resulted in satisfactory DNA quality, with concentrations of 1–15 ng/µL, and provided enough volume to perform the molecular detection and characterization. We confirmed the absence of PCR inhibitors by successfully amplifying the invertebrate cox1 gene in all samples.
Our detection of Bartonella sp. in L. longipalpis sand flies from Ceará state in northeastern Brazil corroborates previous findings. Our obtained 377-bp Bartonella gltA sequence clustered in the same subclade as B. ancashensis sequences obtained from humans with verruga peruana and a genotype recently detected in pools of Dampfomyia beltrani sand flies from Mexico (34). Of interest, genotypes closely related to B. bacilliformis were previously detected in Psathyromyia sand flies from Mexico, a nonendemic country for Carrion’s disease (28). Collectively, findings to date highlight the occurrence of putative novel genotypes belonging to ancient Bartonella lineages in sand flies from Brazil and Mexico, whose zoonotic potential remains unknown.
Although natural Bartonella sp. infections have not been previously reported in L. longipalpis sand flies, experimental studies of this species demonstrated infection with B. ancashensis, which remained viable in the anterior midgut for up to 7 days (4). A subsequent report describing the experimental infection of L. longipalpis sand flies with B. bacilliformis noted similar bacterial viability results (35). Although the L. longipalpis species has been used as a model for sand fly infection with B. bacilliformis, there are no reports of this species in Peru, where Carrion’s disease is endemic (36). Prior investigators have suggested L. longipalpis sand flies might play a short-term role in the maintenance of Bartonella and potentially serve as a vector during that time (4,35). Our data further reinforce the need for additional investigation into the potential role of various sand flies for transmission of Bartonella spp. to human patients and sick animals. Future research specifically focusing on L. longipalpis sand flies is of particular importance because the species is the main vector of Leishmania infantum and is widely distributed in Brazil and throughout Central and South America (37). Although absent from Peru, the L. longipalpis sand fly belongs to the same genus, albeit from different subgenus, as the primary vector of Bartonella bacilliformis in Peru, namely the Lutzomyia (Helcocyrtomyia) peruensis sand fly. Furthermore, the L. longipalpis sand fly is related to sand fly species in which Bartonella DNA have already been detected, namely Lutzomyia (Tricholateralis) gomezi, Lutzomyia (Tricholateralis) cruciata, Lutzomyia (Tricholateralis) sherlocki, or to species that have been incriminated as additional putative vectors for B. bacilliformis, namely Lutzomyia (Helcocyrtomyia) pescei, Lutzomyia (Helcocyrtomyia) noguchii, and Lutzomyia (Helcocyrtomyia) ayacuchensis (7,19,22,23,29; A.R. Carrazco-Montalvo, unpub. data). Those findings highlight the importance of the sand fly genus Lutzomyia sensu stricto in the transmission cycles of Bartonella in South America.
Our investigation also revealed the amplification of a Bartonella sp. nuoG sequence with ≈94% identity to sequences previously detected in insectivorous P. davyi bats from Guatemala. The obtained genotype (detected in a Nyssomyia antunesi specimen captured in the state of Acre) shared 88%–91% identity with other Bartonella sp. sequences previously detected in bats and their associated ectoparasites from Brazil, including sequences amplified from Diphylla ecaudata and Desmodus rotundus vampire bats (38) and Trichobius dugesii flies (39). Despite the diverse phlebotomine sand fly fauna found across many Brazil biomes (≈370 species) and the country’s proximity to regions reporting cases of Carrion’s disease and Bartonella sp. in sand flies, the occurrence of Bartonella in those dipterans in Brazil has been unconfirmed. However, based on phlebotomine sand fly feeding habits (40), many studies have reported the occurrence of Bartonella sp. in vertebrates that act as hosts for sand fly blood meals, including rodents (41,42), marsupials 43), bats (38,39,42), and xenarthrans (44). Although Streblidae and Nycteribiidae flies act as the main putative vectors of Bartonella species transmission among bats (39,45), many sand fly species that feed on bats (and other hosts) can acquire Bartonella spp. infections during blood-feeding. We believe that sand fly feeding habits and the high prevalence of Bartonella infection in many reservoir mammal hosts indicates a potential relationship and involvement of sand flies in the epidemiologic cycles of these bacteria.
In conclusion, we amplified Bartonella spp. DNA and successfully sequenced from L. longipalpis and Ny. antunesi sand flies, indicating possible involvement of these phlebotomine species in the maintenance or transmission cycle of Bartonella spp. The Bartonella gltA genotype was closely related to B. ancashensis, and the nuoG genotype was most closely related to a bat-associated Bartonella sp. Determining the epidemiologic cycle of these agents in Brazil will require elucidating the species and lineages of Bartonella spp. circulating among sand flies and determining whether sand flies in Brazil are capable of Bartonella spp. transmission to animals, including humans.
More information for Bartonella spp. in phlebotomine sand flies, Brazil.
Acknowledgments
The dataset for this study is publicly available in the Sistema de Informação sobre a Biodiversidade Brasileira (SiBBr) and the Global Biodiversity Facility Information (GBIF) (https://doi.org/10.15468/3cnmuw).
This study was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP process nos. 2022/07008-6 and 2022/08543-2), and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico–Productivity Grant for MRA, CNPq process no. 303701/2021-8). P.H.F.S. recieved financial support from Fundação de Amparo à Pesquisa do Estado de Minas Gerais (PPM-00676-18).
Biography
Dr. Lee is a veterinarian and currently a PhD student at São Paulo State University in Brazil. His research is focused on the molecular detection of bloodborne agents in ectoparasites and wild and domestic animals.
Footnotes
Suggested citation for this article: Lee DAB, Shimabukuro PHF, Brilhante AF, Arantes PVC, Sanches GS, Franco EO, et al. Bartonella spp. in phlebotominae sand flies, Brazil. Emerg Infect Dis. 2024 Oct [date cited]. https://doi.org/10.3201/eid3010.240397
References
- 1.Okaro U, Addisu A, Casanas B, Anderson B. Bartonella species, an emerging cause of blood-culture-negative endocarditis. Clin Microbiol Rev. 2017;30:709–46. 10.1128/CMR.00013-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Breitschwerdt EB. Bartonellosis: one health perspectives for an emerging infectious disease. ILAR J. 2014;55:46–58. 10.1093/ilar/ilu015 [DOI] [PubMed] [Google Scholar]
- 3.Garcia-Quintanilla M, Dichter AA, Guerra H, Kempf VAJ. Carrion’s disease: more than a neglected disease. Parasit Vectors. 2019;12:141. 10.1186/s13071-019-3390-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Minnick MF, Robinson AJ, Powell RD, Rowland TE. Experimental colonization of sand flies (Lutzomyia longipalpis; Diptera: Psychodidae) by Bartonella ancashensis. Vector Borne Zoonotic Dis. 2023;23:324–30. 10.1089/vbz.2022.0087 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Regier Y, O Rourke F, Kempf VA. Bartonella spp. - a chance to establish One Health concepts in veterinary and human medicine. Parasit Vectors. 2016;9:261. 10.1186/s13071-016-1546-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lin EY, Tsigrelis C, Baddour LM, Lepidi H, Rolain JM, Patel R, et al. Candidatus Bartonella mayotimonensis and endocarditis. Emerg Infect Dis. 2010;16:500–3. 10.3201/eid1603.081673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gomes C, Ruiz J. Carrion’s disease: the sound of silence. Clin Microbiol Rev. 2017;31:e00056–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chomel BB, Boulouis HJ, Breitschwerdt EB, Kasten RW, Vayssier-Taussat M, Birtles RJ, et al. Ecological fitness and strategies of adaptation of Bartonella species to their hosts and vectors. Vet Res. 2009;40:29. 10.1051/vetres/2009011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rudolf I, Blažejová H, Mendel J, Straková P, Šebesta O, Rettich F, et al. Bartonella species in medically important mosquitoes, Central Europe. Parasitol Res. 2020;119:2713–7. 10.1007/s00436-020-06732-1 [DOI] [PubMed] [Google Scholar]
- 10.Sacristán C. das Neves CG, Suhel F, Sacristán I, Tengs T, Hamnes IS, et al. Bartonella spp. detection in ticks, Culicoides biting midges and wild cervids from Norway. Transbound Emerg Dis. 2021;68:941–51. 10.1111/tbed.13762 [DOI] [PubMed]
- 11.Laroche M, Berenger J-M, Mediannikov O, Raoult D, Parola P. Detection of a potential new Bartonella species “Candidatus Bartonella rondoniensis” in human biting kissing bugs (Reduviidae; Triatominae). PLoS Negl Trop Dis. 2017;11:e0005297. 10.1371/journal.pntd.0005297 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Melter O, Arvand M, Votýpka J, Hulínská D. Bartonella quintana transmission from mite to family with high socioeconomic status. Emerg Infect Dis. 2012;18:163–5. 10.3201/eid1801.110186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Loan HK, Cuong NV, Takhampunya R, Klangthong K, Osikowicz L, Kiet BT, et al. Bartonella species and trombiculid mites of rats from the Mekong Delta of Vietnam. Vector Borne Zoonotic Dis. 2015;15:40–7. 10.1089/vbz.2014.1604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Han HJ, Li ZM, Li X, Liu JX, Peng QM, Wang R, et al. Bats and their ectoparasites (Nycteribiidae and Spinturnicidae) carry diverse novel Bartonella genotypes, China. Transbound Emerg Dis. 2022;69:e845–58. 10.1111/tbed.14357 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Billeter SA, Levy MG, Chomel BB, Breitschwerdt EB. Vector transmission of Bartonella species with emphasis on the potential for tick transmission. Med Vet Entomol. 2008;22:1–15. 10.1111/j.1365-2915.2008.00713.x [DOI] [PubMed] [Google Scholar]
- 16.Król N, Militzer N, Stöbe E, Nijhof AM, Pfeffer M, Kempf VAJ, et al. Evaluating transmission paths for three different Bartonella spp. in Ixodes ricinus ticks using artificial feeding. Microorganisms. 2021;9:5:901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shimabukuro PHF, de Andrade AJ, Galati EAB. Checklist of American sand flies (Diptera, Psychodidae, Phlebotominae): genera, species, and their distribution. Zookeys. 2017;660:67–106. 10.3897/zookeys.660.10508 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jancarova M, Polanska N, Volf P, Dvorak V. The role of sand flies as vectors of viruses other than phleboviruses. J Gen Virol. 2023;104:001837. 10.1099/jgv.0.001837 [DOI] [PubMed] [Google Scholar]
- 19.Lydy SL, Lascano MS, Garcia-Perez JE, Williams-Newkirk AJ, Grijalva MJ. Seroprevalence and risk factors for infection with Bartonella bacilliformis in Loja province, Ecuador. Emerg Microbes Infect. 2018;7:115. 10.1038/s41426-018-0110-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ulloa GM, Vásquez-Achaya F, Gomes C, Del Valle LJ, Ruiz J, Pons MJ, et al. Molecular detection of Bartonella bacilliformis in Lutzomyia maranonensis in Cajamarca, Peru: a new potential vector of carrion’s disease in Peru? Am J Trop Med Hyg. 2018;99:1229–33. 10.4269/ajtmh.18-0520 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Alexander B. A review of bartonellosis in Ecuador and Colombia. Am J Trop Med Hyg. 1995;52:354–9. 10.4269/ajtmh.1995.52.354 [DOI] [PubMed] [Google Scholar]
- 22.Minnick MF, Anderson BE, Lima A, Battisti JM, Lawyer PG, Birtles RJ. Oroya fever and verruga peruana: bartonelloses unique to South America. PLoS Negl Trop Dis. 2014;8:e2919. 10.1371/journal.pntd.0002919 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Noguchi H, Shannon RC, Tilden EB, Tyler JR. Etiology of Oroya fever: XIV. The insect vectors of Carrion’s disease. J Exp Med. 1929;49:993–1008. 10.1084/jem.49.6.993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mullins KE, Hang J, Jiang J, Leguia M, Kasper MR, Ventosilla P, et al. Description of Bartonella ancashensis sp. nov., isolated from the blood of two patients with verruga peruana. Int J Syst Evol Microbiol. 2015;65:3339–43. 10.1099/ijsem.0.000416 [DOI] [PubMed] [Google Scholar]
- 25.Mullins KE, Hang J, Clifford RJ, Onmus-Leone F, Yang Y, Jiang J, et al. Whole-genome analysis of Bartonella ancashensis, a novel pathogen causing verruga peruana, rural Ancash region, Peru. Emerg Infect Dis. 2017;23:430–8. 10.3201/eid2303.161476 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Shimabukuro PHF, Andrade AJ, Galati EAB. Phlebotominae in Catálogo Taxonômico da Fauna do Brasil. PNUD. 2024. [cited 2024 Feb 14]. http://fauna.jbrj.gov.br/fauna/faunadobrasil/3297
- 27.Zorrilla VO, Lozano ME, Espada LJ, Kosoy M, McKee C, Valdivia HO, et al. Comparison of sand fly trapping approaches for vector surveillance of Leishmania and Bartonella species in ecologically distinct, endemic regions of Peru. PLoS Negl Trop Dis. 2021;15:e0009517. 10.1371/journal.pntd.0009517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lozano-Sardaneta YN, Soto-Olguín NJ, Rodríguez-Rojas JJ, Sánchez-Montes S, Rebollar-Téllez EA, Becker I. Molecular detection of Bartonella sp. in Psathyromyia shannoni and Lutzomyia cruciata from northeastern Mexico. Front Trop Dis. 2021;2:780808. 10.3389/fitd.2021.780808 [DOI] [Google Scholar]
- 29.Galati EAB. Phlebotominae (Diptera, Psychodidae) classificação, morfologia, terminologia e identificação de adultos. Apostila. Bioecologia e Identificação de Phlebotominae. 2016;1:131. [Google Scholar]
- 30.Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. 1999;41:95–8. [Google Scholar]
- 31.Bustin SA, Nolan T. Pitfalls of quantitative real-time reverse-transcription polymerase chain reaction. J Biomol Tech. 2004;15:155–66. [PMC free article] [PubMed] [Google Scholar]
- 32.Lozano-Sardaneta YN, Colunga-Salas P, Sánchez-Montes S, Cáceres AG, Becker I. First report of Bartonella sp. in sand flies (Diptera: Psychodidae: Phlebotominae) from southern Mexico. J Am Mosq Control Assoc. 2019;35:224–7. 10.2987/19-6854.1 [DOI] [PubMed] [Google Scholar]
- 33.Lozano-Sardaneta YN, Marina CF, Torres-Monzón JA, Sánchez-Cordero V, Becker I. Molecular detection of Wolbachia and Bartonella as part of the microbiome of phlebotomine sand flies from Chiapas, Mexico. Parasitol Res. 2023;122:1293–301. 10.1007/s00436-023-07829-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Martínez-Burgos M, Lozano-Sardaneta YN, Rodríguez-Rojas JJ, Gómez-Rivera AS, Canto-Mis KL, Flores-Escobar E, et al. Species diversity and detection of pathogens in phlebotomine sand flies collected from forest management areas of Quintana Roo, Mexico. Med Vet Entomol. 2023;37:845–58. 10.1111/mve.12691 [DOI] [PubMed] [Google Scholar]
- 35.Battisti JM, Lawyer PG, Minnick MF. Colonization of Lutzomyia verrucarum and Lutzomyia longipalpis sand flies (Diptera: Psychodidae) by Bartonella bacilliformis, the etiologic agent of Carrión’s disease. PLoS Negl Trop Dis. 2015;9:e0004128. 10.1371/journal.pntd.0004128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sousa-Paula LC, Otranto D, Dantas-Torres F. Lutzomyia longipalpis (sand fly). Trends Parasitol. 2020;36:796–7. 10.1016/j.pt.2020.05.007 [DOI] [PubMed] [Google Scholar]
- 37.Lainson R, Rangel EF. Lutzomyia longipalpis and the eco-epidemiology of American visceral leishmaniasis, with particular reference to Brazil: a review. Mem Inst Oswaldo Cruz. 2005;100:811–27. 10.1590/S0074-02762005000800001 [DOI] [PubMed] [Google Scholar]
- 38.André MR, Gutiérrez R, Ikeda P, do Amaral RB, de Sousa KCM, Nachum-Biala Y, et al. Genetic diversity of Bartonella spp. in vampire bats from Brazil. Transbound Emerg Dis. 2019;66:2329–41. 10.1111/tbed.13290 [DOI] [PubMed] [Google Scholar]
- 39.Ikeda P, Marinho Torres J, Perles L, Lourenço EC, Herrera HM, de Oliveira CE, et al. Intra- and inter-host assessment of Bartonella diversity with focus on non-hematophagous bats and associated ectoparasites from Brazil. Microorganisms. 2020;8:1822. 10.3390/microorganisms8111822 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sousa RLT, Vasconcelos SA, Santos-Mallet JR, Nascimento EF, Teixeira CR, Silva CLM, et al. Padrões de fonte alimentar dos Flebotomíneos (Diptera: Psychodidae) vetores das Leishmanioses: uma revisão bibliográfica. Revista Eletrônica Acervo Saúde. 2021;13:e8567. 10.25248/reas.e8567.2021 [DOI]
- 41.do Amaral RB, Cardozo MV, Varani AM, Gonçalves LR, Furquim MEC, Dias CM, et al. Bartonella machadoae sp. nov. isolated from wild rodents in the Pantanal wetland. Acta Trop. 2022;229:106368. 10.1016/j.actatropica.2022.106368 [DOI] [PubMed] [Google Scholar]
- 42.Pacheco TDA, Amaral RBD, Ikeda P, Maia MO, Lee DAB, Semedo TBF, et al. Molecular detection and characterization of Bartonella spp. in small mammals in the Amazonia and Cerrado biomes, midwestern Brazil. Acta Trop. 2024;251:107129. 10.1016/j.actatropica.2024.107129 [DOI] [PubMed] [Google Scholar]
- 43.Braga MDSCO, Costa FB, Calchi AC, de Mello VVC, Mongruel ACB, Dias CM, et al. Molecular detection and characterization of vector-borne agents in common opossums (Didelphis marsupialis) from northeastern Brazil. Acta Trop. 2023;244:106955. 10.1016/j.actatropica.2023.106955 [DOI] [PubMed] [Google Scholar]
- 44.Calchi AC, Vultão JG, Alves MH, Yogui DR, Desbiez ALJ, do Amaral RB, et al. Multi‐locus sequencing reveals a novel Bartonella in mammals from the Superorder Xenarthra. Transbound Emerg Dis. 2020;67:tbed.13545. 10.1111/tbed.13545 [DOI] [PubMed]
- 45.do Amaral RB, Lourenço EC, Famadas KM, Garcia AB, Machado RZ, André MR. Molecular detection of Bartonella spp. and Rickettsia spp. in bat ectoparasites in Brazil. PLoS One. 2018;13:e0198629. 10.1371/journal.pone.0198629 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
More information for Bartonella spp. in phlebotomine sand flies, Brazil.