

Abstract
The cultivation of grapevines has spanned millennia, leading to thousands of varieties through exchanges, mutations, and crosses between genotypes, as well probably as gene flow from wild populations. These varieties are typically categorized by regional origin and primary use, either for wine production or fruit consumption. France, within the Western European group, hosts many of the world’s renowned wine grape varieties. However, the historical development of cultivated grapevines in France and in the world remains poorly understood. This study applies morphometry on 19,377 charred and waterlogged archaeological grape pips to investigate the evolutionary history of grapevine in France over the last 10,000 years. The study compares seed outlines and lengths, corrected for taphonomic distortions, with a reference collection of 80 wild and 466 modern domestic grapevine accessions. Findings reveal a shift from wild grapevine exploitation to the expansion of domestic varieties around 600–500 BCE, coinciding with Mediterranean cultural influences and the introduction of eastern grape types. The identification of the East-Table group, a group of varieties of eastern origin for fruit consumption, indicates that grapes were also grown for food, especially in Mediterranean regions and near urban areas, alongside wine production. Early French viticulture featured a notable presence of Western European wine-type grapevines. The abundance of pips with wild-like morphology suggests early cultivation involved plants at an initial domestication stage and gene flow between introduced and wild grapevines. As viticulture spread northward, wild and Eastern morphotypes declined, leading to the dominance of Western European wine types in inner France during the Middle Ages.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-024-72692-6.
Keywords: Vitis vinifera, Domestication, Morphometrics, Archaeobotany, Taphonomy, Diffusion, Perennial plant
Subject terms: Palaeoecology, Plant domestication, Archaeology, Cultural evolution
Introduction
The grapevine (Vitis vinifera L.) stands as a cornerstone of cultural and economic significance in the history of the Mediterranean basin and Europe, serving as a source of edible fresh and dried fruits and wine1. Its domestication, an intricate historical process, originated from the Eurasian wild grapevine (Vitis vinifera subsp. sylvestris, hereafter V. sylvestris) in South-West Asia, potentially in the Caucasus and the Levant2. This process, initiated between 8,000 and 6,000 years ago3,4, set the stage for the gradual spread of vinegrowing and cultivated grapevines (Vitis vinifera subsp. vinifera, hereafter V. vinifera) westwards across the Mediterranean and Europe, driven by trade, cultural exchanges, and population movements1,5. Millennia of sexual crosses and clonal propagation, combined with gene flows between V. vinifera and V. sylvestris and somatic genetic mutations resulted in a diversity of 5,000–10,000 cultivated varieties (cultivars), nowadays categorized into distinct groups according to their main use (table vs. wine) and geographical origin6.
Genomic analyses of over 1,600 V. vinifera cultivars and 800 V. sylvestris accessions reveal six main groups of domestic grapevines related to their main use and regional origins, including “Western Asian table grapes”, “Caucasian wine varieties”, “Muscat varieties”, “Balkan wine varieties”, “Iberian wine varieties” and “Western European wine varieties”2. Notably, three major clusters consistently emerge throughout previous studies7–10, supporting the classification into three main groups or Proles previously proposed by Negrul based on morphology and geography: Proles orientalis, grouping table grape varieties from the Near and the Middle East, Proles pontica, with wine and multipurpose varieties from the Balkans and the Black Sea area, and Proles occidentalis, composed of Central and Western European small berried wine varieties11. The latter encompasses some of the most renowned and widely grown varieties in the world, like Cabernet-Sauvignon, Merlot, Tempranillo, Chardonnay and Syrah12,13.
However, the chronological and geographical origins of grapevine varieties and variety groups remain elusive. It is still challenging to date the spread and origins of winegrowing throughout the Mediterranean. In Greece, Neolithic evidence points to the initiation of winemaking and vine cultivation around 4,500-4,000 BCE, but this appears to have been based on wild grapevines only14,15. Current data indicates that domestic V. vinifera appeared in the first half of the 2nd millennium BCE, potentially introduced from the East14. In France, there exists no evidence of vine cultivation before the 6th century BCE and the first contacts with Mediterranean peoples, especially Greeks and Etruscans, and ensuing colonial settlements16,17. Viticulture was rapidly adopted by local Mediterranean populations, raising questions about the introduction of varieties from the East or the use of autochthonous V. sylvestris.
The evolution of cultivated grapevines and their adaptation to climatic and stational variations, pathogen infestations and socio-economic changes, has probably been crucial for the resilience and sustainability of wine production over the centuries. Today, the effects of climate change on viticulture are a major concern. Adopting new grape varieties is considered as one of the most effective ways to ensure the sustainability of viticulture13,18. This study endeavours to quantify the morphological changes of grape seeds and relate them to fluctuations in agrobiodiversity and to the evolution of cultivated grapevines since the beginnings of viticulture in France.
Following the Roman colonization, vinegrowing and wine making in Mediterranean France underwent profound transformations. From the end of the 1st century BCE onwards, it changed from localized cultivation to a large-scale specialized activity, based on the Italian colonial model, leading to the production of vast quantities of wine exported across the expanse of the Roman Empire19,20. As the 1st century CE unfolded, viticulture began to extend its reach beyond its Mediterranean origins, venturing into territories characterized by oceanic and continental climates. This westward and northward expansion into climates different from its traditional Mediterranean habitat illustrates viticulture’s adaptability to different environmental conditions21. Warmer temperatures during the Roman climate optimum probably had a positive impact on the spread of viticulture to the north22. Indeed, more recent historical records suggest that viticulture reached its zenith in north-western France during the 13th century, coinciding with the warming trends of the Medieval climate optimum23.
Archaeological excavations yield invaluable insights into the evolutionary trajectory of fruit domestication, particularly based on the analysis of changes in fruit size and shape over time24. Seed morphometrics are a cornerstone of archaeobotany in studying processes of domestication and changes in agrobiodiversity25–29. They have been instrumental in discerning the domestic state of grapevines and elucidating morphological diversity within V. vinifera14,30–37. V. sylvestris seeds are globular in shape, with a short beak, whereas V. vinifera seeds are more elongated, pyriform and have a long beak38. In this study, we present a comprehensive analysis of seed outlines derived from a dataset comprising 231 samples corresponding to 19,377 grape seeds in total, spanning archaeological sites across France, from approximately 10,000 BCE to the 19th century CE.
We track the temporal and spatial evolution of seed size and shape, taking into account deformations due to taphonomic conditions (preservation by charring or waterlogging) and correlate these changes with the geographic and bioclimatic contexts of the archaeological sites. By implementing a comparative approach, we juxtapose ancient seed morphologies with a reference database encompassing 80 modern V. sylvestris accessions and 466 V. vinifera varieties. This methodological framework allows us to delineate the emergence and dissemination of the domestic morphotype from its advent around 500 BCE. Furthermore, we refine our characterization of domestic archaeological seeds by comparing them with contemporary grapevine groups associated with specific utilization and geographical origins39.
Results
Morphological diversity of archaeological seeds
We explored the morphological diversity of 3,007 charred and 16,370 waterlogged archaeological grape seeds originating from 176 sites across France (Fig. 1 and Supplementary Table 1), using Principal Component Analysis (PCA) on elliptical Fourier transforms (EFT) coefficients (Supplementary Fig. 1). Throughout the chronology, the morphology of charred seeds is consistently shifted towards more globular seed shapes compared to waterlogged specimens (Fig. 2a). However, the PCA unveils on the first principal component a discernible and similar temporal trajectory in pip morphology for the two preservation modes, from globular to pyriform seed shapes, with more elongated beaks. Changes in shape are particularly strong between the Bronze Age (approximately 2,200–750 BCE) and the Iron Age (approximately 750–27 BCE) but continue until the Middle Ages and beyond. Variations in seed shape across different regions manifest themselves distinctly during several chronological periods (Fig. 2). In the Mediterranean region, a pronounced transformation in pip morphology is observed in the Iron Age, followed by a comparatively mild morphological change. Conversely, in the inland regions of France (WEST, CENO, EAMC), the main morphological shifts happen later, during the Roman period. However, the Iron Age is represented by few samples.
Fig. 1.
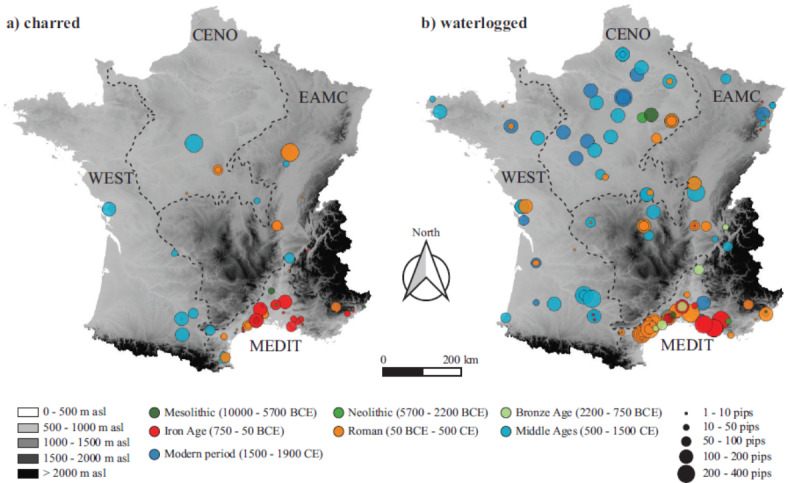
Geographical distribution of the studied archaeobotanical samples of grape seeds according to the mode of preservation, (a) charred, (b) waterlogged. The colour of the circles indicates sample age and the size is proportional to the number of pips analized. The dotted lines represent the limits of the 4 large ecological regions: Mediterranean (MEDIT), West (WEST), Centre-North (CENO) and East-Massif Central (EAMC).
Fig. 2.

Representation of the results on PC1 of the Principal Component Analysis on elliptical Fourier transforms (EFT) coefficients of the archaeological grape seeds. PC1 registers a transition from globular seeds with short beaks to more elongated and slender seeds with long beaks. (a) Representation according to preservation mode and archaeological periods (Mesolithic + Neolithic, Bronze Age, Iron Age, Roman, Middle Ages, Modern period). (b) Representation according to ecological regions (Mediterranean, West, Centre-North, East-Massif-Central) and archaeological periods. The number of studied pips is indicated for each combination preservation x period x region. Differences between consecutive periods and differences between preservation modes within periods were tested using pairwise Wilcoxon tests: *. Differences between regions within periods were tested using Kruskal-Wallis tests: *. Only p-values < 0.001 are shown.
Correcting distortions of ancient seeds and identifying grapevine groups
Linear Discriminant Analyses (LDA) performed on Fourier coefficients alone enables the discrimination between wild and domestic seed types (status)14,30,32,40, here with a remarkable accuracy of 95.86%, for wild and 95.97% for domestic seeds. Additionally, it allows the classification of domestic seeds into four Use and Regional Groups (URG) of varieties, albeit with a slightly lower global precision of 68.66% (Fig. 3). Here we use the 4 main URG identified from 10,000 genetic SNP markers in the study of Laucou et al.10: East-Table, Balkans-Wine, Iberian-Wine, West-Wine. Considering the length of the seeds in addition to EFT coefficients improves marginally the accuracy of status identification and more significantly the accuracy of URG identification (73.18% globally). It particularly enhances the identification of the East-Table and Iberian-Wine groups.
Fig. 3.

General results of the classification of modern and archaeological grape pips by comparson to a reference collection and using Linear Discriminant Analysis (LDA). Identification of (a) status and (b) Use and Regional Groups of varieties using 100 balanced LDA and 3 different methods: shape (EFT), form (EFT + Length), form corrected (EFT*+L*; see M&M section). Left: training accuracies on the modern reference collection of 466 cultivars and 80 wild accessions. Classification accuracies are represented by the percentage of seeds correctly assigned to the group after the 100 LDA. The inferred proportions on the archaeological material are represented by distinguishing results on charred (middle) and waterlogged (right) material.
Classifying archaeological seeds shows the impact of the preservation mode and the morphometric method used on the results. Using exclusively EFT coefficients reveals a higher global proportion of the domestic type for waterlogged seeds (65.33%) than for charred seeds (49.98%). The 4 URG of varieties are recognised within domestic seeds with a similar dominance of West-Wine (57.41% for charred, 53.08% for waterlogged) and East-Table (35.23% for charred, 33.94% for waterlogged) groups.
Considering seed form, combining seed length with EFT coefficients (EFT + L) has the most impact on the classification of charred seeds. The proportion of those classified as domestic is then considerably lower (36.78%) while that of waterlogged seeds is similar (62.1%). This underlines the strong effect of charring on seed length (Supplementary Fig. 2). The effect on URG identification is also strong, with a reduced proportion of seeds allocated to the East-Table group, particularly for charred material. In return, the proportion of the West-Wine group increases.
Building upon experimental charring and waterlogging research41,42, we implemented an average reverse vector of shape alteration, computed separately for charred and waterlogged specimens, in order to address taphonomic distortions and to mitigate the consequences of different preservation conditions. We also accounted for the effect of charring by correcting seed length of charred seeds. Here the results also differ for charred and waterlogged seeds. The proportion of domestic morphology seeds is similar to the one obtained with EFT only for charred material (51.58%) but reaches its highest value for waterlogged remains (74.54%). Regarding the identification of URG, the results are close to the ones obtained with EFT for charred seeds and differ more for waterlogged material, with a better representation of the Balkan-Wine and Iberian-Wine groups, at the expenseof the East-Table group, in particular.
Evolution of pip and berry size
The temporal dynamics of seed size variation, as evidenced by the examination of charred and waterlogged material, exhibit a consistent trend (Supplementary Fig. 2). Domestication of grapevine led to an increase in berry size and consequently of seed size, as it has been established that a relationship exists between seed size and berry size43–45. A regression analysis, predicated upon measurements of 1470 modern pips and as much berries, allows inferring berry volume from pip length (Fig. 4). The final model 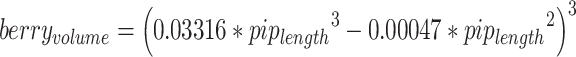 was significant (Pearson correlation test: P < 10−15; adjusted R2 = 0.95). After correcting for taphonomic influences on charred seed dimensions, the inferred berry volumes are close to those inferred from waterlogged seeds, although still slightly lower, suggesting that our correction for charred seeds length is somewhat insufficient. In addition, while the inferred berry volumes globally approximate those documented in modern grapes, they are still slightly lower, even for waterlogged remains for which seed length is less affected by taphonomic processes; this suggests that all inferred berry volumes are still slightly underestimated.
was significant (Pearson correlation test: P < 10−15; adjusted R2 = 0.95). After correcting for taphonomic influences on charred seed dimensions, the inferred berry volumes are close to those inferred from waterlogged seeds, although still slightly lower, suggesting that our correction for charred seeds length is somewhat insufficient. In addition, while the inferred berry volumes globally approximate those documented in modern grapes, they are still slightly lower, even for waterlogged remains for which seed length is less affected by taphonomic processes; this suggests that all inferred berry volumes are still slightly underestimated.
Fig. 4.

Approximation of berry size evolution through time compared to modern wild and domestic grapes. Berry size and its temporal changes are inferred from the length of the archaeological pips (Supplementary Fig. 2) using length correction for charred pips and a linear model (Supplementary Fig. 3). The dots represent the median value and the ends of the bars the first and third quartiles. Temporal trends are represented using (a) mean dates BCE/CE and (b) archaeological periods, distinguishing the results from charred and waterlogged material. The values inferred for archaeological seeds are juxtaposed with (c) the measured berry volumes for 49 modern wild individuals, cultivated wild grapevines and wine and table varieties. Differences between consecutive periods were tested for waterlogged material using pairwise Wilcoxon tests: *. Only p-values < 0.001 are shown.
Considering the two preservation modes together, the estimated berry volumes decrease slightly from the Mesolithic to the Bronze Age. However, throughout this long period of time, they stay close to the sizes characteristic of berries from modern wild grapevine populations. Estimated berry volumes increase sharply in the Iron Age to reach a maximum between circa 500 BCE and 500 CE, throughout the Iron Age and Roman period. Following this peak, there is a discernible decline in berry volume between Roman times and the Middle Ages, although it remains notably elevated compared to pre-500 BCE samples. From the Iron Age onwards, and in spite of a notable variability in the predicted volumes, these largely encompass the dimensions of contemporary cultivated V. sylvestris and wine cultivars. According to these predictions, the most voluminous berries reported in present-day grapes (1.5–1.8 cm3), corresponding to table varieties, have few analogues in archaeological material.
Wild and domestic Vitis vinifera types
The observed pattern of seed morphological change is largely explained by a chronological evolution of proportions between wild and domestic types, as revealed by the comparison of archaeological seeds to the reference collection of modern material. Until approximately 500 BCE (Iron Age), the majority of archaeological seeds are attributed to the wild morphotype, whether using only EFT coefficients or EFT*+L* (Fig. 5, a, Supplementary Tables 2, 3 and 4). From the Mesolithic to the Bronze Age, a minor proportion of pips is identified as having a domestic morphology in most of the waterlogged samples. This proportion is at most 19% in the Late Bronze Age site of La Motte (ca. 900 BCE), with the method EFT*+L*. In this case, it can be hypothesized that the shape correction is excessive, resulting in an overestimation of the domestic type. However, using only EFT + L the proportion of the domestic type reach 4.8% in Mesolithic Noyen-sur-Seine (ca. 6,900 BCE), 7.4% in Early Bronze Age Nîmes-Vistres (ca. 2,150 BCE) and finally 11.9% in Late Bronze Age La Motte.
Fig. 5.

Evolution of wild pips size and shape according to archaeological periods, seed preservation and ecological regions. (a) Proportion of wild pips identified by 100 balanced LDA in the assemblages using the EFT (shape) method. Only groups > 2 samples per period are shown. The dots represent the median proportion and the ends of the bars the first and third quartiles, (b) Changes in the length of the pips identified as wild (mm), (c) Changes in the shape of the pips identified as wild. The shape is represented by the coordinates of the pips on the first component of the PCA performed on EFT coefficients. The number of studied pips is indicated for each combination preservation x period x region. Differences between consecutive periods were tested using pairwise Wilcoxon tests: *. Differences between regions within periods were tested using Kruskal-Wallis tests: *. Only p-values < 0.001 are shown.
After 500 BCE, the proportion of domestic-type seeds increases progressively, with a first threshold in the Iron Age, followed by a second one during the Middle Ages (Fig. 5). Considerable variability in the proportion of domestic seeds exists among samples. In the Iron Age, domestic-type seeds are often predominant in the MEDIT region, but not in all samples. The few Iron Age samples available from other regions only contain a minority of domestic-type pips. It is during the Roman period that the proportion of the wild type sharply decreases in the WEST, CENO, and EAMC regions. The subsequent medieval decrease is noticeable everywhere.
While remaining well represented after the Iron Age and the emergence of the domestic morphotype, the wild morphotype undergoes morphological evolution. The average length of wild type seeds increases, especially when we consider waterlogged seeds (Fig. 5b). Seed shape changes at the same time, following a similar trend and converging towards the domestic morphotype (Fig. 5c). After the Roman period, the size and shape of the wild morphotype seem to shift back towards values closer to those found in V. sylvestris today. Differences in seed size and shape between regions are noticeable at different periods, in particular during Roman times.
Domestic vine groups and origins
Despite the variation in the identification of the 4 URGs according to the preservation mode and the morphometric method (EFT, EFT + L or EFT*+L*), a consistent pattern of spatio-temporal variations is recorded (Fig. 6, Supplementary Fig. 4). From the Iron Age to the Modern period, the main groups recorded are consistently East-Table and West-Wine. The Balkan-Wine group is better represented, mostly at the expense of the East-Table group, if we use the EFT*+L* method. The East-Table group shows high and increasing proportions during the Iron Age, before decreasing in the Roman period. The West-Wine group has the opposite dynamic. It already accounted for a large proportion during the Iron Age and gradually increased from the beginning of the Roman period, reaching its height in the Middle Ages. From the second half of the Middle Ages onwards, this Western group gradually decreased, although it remained in the majority. The mix of grape types cultivated in the Late Middle Ages and Modern period seems therefore to be more composite. The East-Table group regains importance, while the Balkan-Wine and the Iberian-Wine groups reach their highest percentages, although they are still always in the minority whatever the method and preservation mode. There are also recurring differences between regions (Fig. 7, Supplementary Fig. 5). The East-Table group is consistently the most strongly represented in the MEDIT region, in relatively constant figures from the Iron Age to the Modern period. While fairly well represented in the more northerly regions during the Roman period, it declined in the Middle Ages, as the West-Wine group became even more prominent. The Iberian-Wine group is always the best represented in the West region.
Fig. 6.

Changes through time in the proportions of pips allocated to the different Use and Regional Groups of varieties in the archaeological assemblages, according to seed preservation and ecological regions. Only the assemblages with > 20 pips are considered. The graph represents a combination of the proportions according to the 3 different methods: EFT (shape), EFT + L (form), EFT*+L* (form corrected). The dots represent the median proportion and the ends of the bars the proportions for Q10 and Q90. The number of studied samples is indicated for each combination preservation x time period. The graphs for each individual method are given in Supplementary Fig. 4.
Fig. 7.

Changes in the proportions of pips allocated to the different Use and Regional Groups of varieties in the archaeological assemblages according to archaeological periods, seed preservation and ecological regions. Only the assemblages with > 20 pips are considered. The graph represents a combination of the proportions according to the 3 different methods: EFT (shape), EFT + L (form), EFT*+L* (form corrected). The dots represent the median proportion and the ends of the bars the proportions for Q10 and Q90. The number of studied samples is indicated for each combination preservation x period x region. The graphs for each individual method are given in Supplementary Fig. 5.
Discussion
The comprehensive study of over 19,000 archaeological grape seeds confirms the impact of preservation modes on seed form as suggested by previous experimental studies41,42, by recognizing consistent differences in size and shape between charred and waterlogged seeds across various periods. As indicated by experiments, charred seeds become notably smaller and more spherical, getting closer to the morphology of wild grape seeds33,41,46,47. This probably explains the higher proportion of wild morphotypes in charred seeds compared to waterlogged specimens. Waterlogging experiments indicate that the resultant deformations are less pronounced than those from charring and do not prevent the distinction between wild and domestic morphotypes42. These contrasting taphonomic effects may partially also account for variations in the representation of Usage and Regional Groups (URG) of grape varieties between charred and waterlogged seeds. Corrections made to the shape and size of archaeological seeds partially compensate for taphonomic deformations. Berry volumes estimated from corrected charred seed lengths do not completely match those from waterlogged seeds. All inferred volumes nearly equate those of contemporary berries, but remain slightly lower. In addition, the proportion of seeds allocated to the wild morphotype remains slightly higher in charred material despite the corrections. Ultimately, these findings suggest that current methods for correcting seed lengths and shape may still be insufficient. Inferred berry volumes are probably slightly underestimated.
Despite some discrepancies in identifying wild and domestic morphotypes as well as URGs within the samples, the diverse methodologies applied in this study show broadly consistent patterns of spatio-temporal variability. Additionally, the findings are congruent across charred and waterlogged seeds. This convergence supports the reliability and robustness of the reconstructed evolutionary history. The study reveals ongoing morphological changes over time, with significant shifts towards the domestic morphotype during the Iron Age (circa 600–500 BCE) in the Mediterranean region and during the Roman era in inland regions. These findings are consistent with established timelines for the emergence of domesticated grapevines and the onset of viticulture in these areas, supporting the theory that cultivated grapevines were introduced to inland France from the south21,48,49. Prior to these periods, fruits were likely collected from naturally occurring wild grapevines widespread across France. From the Mesolithic to the Iron Age, archaeological seeds consistently match with berry volumes of contemporary wild grapes. A slight, gradual reduction in berry volume from the Mesolithic to the Bronze Age may be indicative of environmental shifts, potentially involving a decrease in water availability during the initial phases of berry development50,51.
Could grapevine nevertheless have already been cultivated before the Iron Age in the south and the Roman period in the north? Previous research suggest wine production in Iron Age northwestern France52 and grapevine cultivation in northern Italy during the Bronze Age53–55. Our study identified small percentages of domestic seeds from the Mesolithic period, but likely due mainly to model error (less than 4% for EFT and EFT + L). Percentages of 7.4% and 11.9% of domestic seeds at two Bronze Age sites (ca. 2,150–900 BCE) in the Mediterranean region suggest possible early domestication, but the evidence is weak. The higher frequency of Vitis remains in Neolithic and Bronze Age archaeological sites suggests that grapes were a more common food resource for people in the Mediterranean region compared to inland France but not necessarily cultivated17. The wild plant may have been more widespread in the Mediterranean region due to favourable ecological conditions. Nevertheless, small-scale cultivation prior to the Iron Age cannot be entirely ruled out, especially since the cultivation of wild plants may have occurred without inducing seed morphological change14,15,56.
After the Phocaeans established Marseille in 600 BCE and began cultivating grapevine and selling wine, grapevine seeds and charcoal became more prevalent at sites across the French Mediterranean region17,48,49. Our data show that domestic seed morphotypes were commonly found at many sites, indicating the widespread adoption of viticulture in Mediterranean France. The timing suggests influences from Greek settlers in Marseille, with potential contributions from other Mediterranean cultures such as the Etruscans and Phoenicians57–60. This is consistent with our finding that a significant proportion of domestic seeds is comparable with current varieties characteristic of the eastern Mediterranean and southwest Asia, mainly eastern table types. However, when applying the size and shape correction (EFT*+L*) to the waterlogged pips we obtain more balanced proportions with the wine Balkan and Caucasian group. This supports the model of westward spread of domesticated grapevines and viticulture1,61,62. Furthermore, paleogenomic evidence corroborates eastern genetic lineage in Nîmes (eastern Languedoc, south-eastern France), by the 2nd -1st century BCE63, while Italian variety names referenced by Latin authors point to Greek origins for several varieties64.
Our study also reveals the presence of the group of western European wine varieties in southern France from the early days of viticulture, with proportions comparable to those of the eastern Mediterranean types. Western European wine cultivars include late-blooming types with short vegetative periods, small to medium-sized compact bunches, small spherical berries with thick skin, and acidic juice11,38,65. This group is now characteristic of Germany, Alpine regions, France and the Iberian Peninsula. In Mediterranean France, ancient DNA provided evidence of close kinship between Roman grape seeds and modern western European wine varieties63. Our results suggest the regional emergence of the western European wine group from the Iron Age onwards, thought this type may have originated earlier in other western European wine regions and been introduced to southern France in the same way as eastern cultivated grapevines. They could have come from northern Italy, where grapevines have been grown since the 16th -10th centuries BCE, the date of emergence still a matter of debate66,67. The weak isolation-by-distance correlation in current cultivars suggests extensive exchange and interbreeding of varieties across regions throughout history2.
Wild-type morphology seeds are a significant component of Iron Age assemblages in the Mediterranean region, sometimes representing most of the sample. Although their proportion decreases over time, wild-type pips remain common in Roman sites in southern France. They are however less frequent in inland France during the roman period. Wild-type pips, or intermediate morphologies between wild and domestic, have been reported at early vinegrowing sites in various regions14,30,35,36,43,68. We show here that Iron Age and Roman pips classified as wild differ in size and shape from present-day wild pips, indicating the impact of cultivation and an incipient selection process. Middle Bronze Age ancient DNA and morphometric evidence from Pertosa cave (Campania, southern Italy) suggest a mix of eastern domestic varieties, wild type vines and, remarkably, a hybrid between both groups69. Combined cultivation of introduced domesticated grapevines and wild individuals likely promoted gene flow between the two groups. Gene flow from V. sylvestris is considered to have played a key role in adapting domestic grapevines to new environments and in developing wine varieties, with wild grape traits different from eastern table grapes (berry size, berry skin thickness)2,8,70. The western wine group formed from eastern Mediterranean domestic germplasm with crosses and introgressions from western V. sylvestris38. Genetic data show the contribution of local wild grapevines to regional grape varieties in several parts of western Europe8,71–74. The importance of the wild morphotype declined during the Middle Ages, and seeds classified in this wild group reverted to shapes resembling present-day V. sylvestris. This implies that, from the Middle Ages onwards, the wild morphotype more often represents spontaneous wild grapevines rather than lowly-selected cultivated plants, suggesting a growing separation between wild and cultivated grape populations.
The slight decline in the importance of grapevines of eastern Mediterranean origin during the Roman period is probably linked to the expansion of large-scale viticulture in south-eastern France from the 1st to the 3rd centuries CE20,75–77. The focus on wine production and export is likely to have favoured specialised wine varieties. As viticulture spread to inland regions the importance of eastern grape types declined, and their role further diminished after the Roman period. Eastern varieties, typically early-blooming plants with a long vegetative period and poor cold tolerance11,38,65, are better suited to the Mediterranean climate than to more northerly climates with oceanic or continental influences. Introduced from the south, eastern varieties may have been maintained outside the Mediterranean region during the Roman period, before Medieval and Modern winegrowing focused on western European types, well adapted to local ecological conditions. Written records from the 19th century onwards confirm that major inland winegrowing regions in France relied on western European varieties78–81. The availability of varieties suited to regional climates probably contributed to the significant expansion of winegrowing in inland France during the Middle Ages, as shown by archaeobotanical records17.
The presence of seeds assigned to the East-Table modern group, particularly in the Mediterranean region, suggests that viticulture was partially oriented towards table grape production, more particularly in its early Iron Age phase, although these varieties can also produce wine. Genetics suggest that East-Table grapevines date back to the initial domestication of grapevine in southwest Asia and played a significant role in the development of major European grape clusters2. Latin authors such as Columella (De re rustica, 3,2) and Pliny the Elder (Naturalis Historia, 14, 42) referenced specialized table varieties in the 1st c. CE. Table grape records in archaeobotanical assemblages may have been favoured by the fact that a large number of samples originated from urban rubbish deposits and latrines, contexts that mainly register fruit consumption82–84. Columella (De re rustica, 3, 2) advised planting table grape vineyards near towns for easier transport to market without damage. Thus, the cultivation of table grapes close to urban areas may have contributed to the level of the table seed morphotype in our samples. But, in ancient Rome, grapes were also preserved by drying or by immersion in wine or grape juice85, enabling trade over long distances. Consequently, some seeds found at French sites could have been imported from other Mediterranean regions86. The same situation could have occurred in the medieval period, when grapevines were often grown in gardens, orchards, and vineyards near or within towns, while dried raisins were traded throughout France84,87. Medieval recipe books indicate grapes were more frequently used in southern French cuisine88, which is consistent with the higher frequency of the East-Table morphotype in the Mediterranean region. Limited increase in the East-Table and Iberian types after 1,500 CE may result from urban expansion, trade development and a new preference for exotic varieties89.
The speed of the process of grapevine domestication remains a debated topic90,91. Over the past two decades, archaeobotany and genetics have shifted views on plant domestication towards protracted processes27,92,93. However, because grapevine and other perennial plants are generally cross-pollinated, highly heterozygous, and have long juvenile periods, their domestication and the fixation and multiplication of cultivars primarily depended on clonal propagation. This considerably reduced the number of sexual generations since the onset of domestication compared to annual plants, while domestication syndrome traits are more limited and new mutations may arise somatically rather than sexually70,94,95. Changes associated with domestication could have occurred rapidly through mutations and clonal propagation91 or taken much longer, through gradual accumulation of sexual crosses95.
Some seeds of the early French cultivated grapevines bear morphological signs of advanced domestication. These vines may include imported varieties propagated vegetatively, possibly only a few generations away from the first domesticates, several millennia before in southwest Asia96–98. On the other hand, the high proportions of domesticated western grapevines and cultivated vines resembling wild types suggest a dynamic domestication process, likely involving multiple sexual crosses. Despite thousands of registered varieties, current cultivated grape diversity may represent only a fraction of past diversity, that gradually concentrated around elite cultivars with close parental relationships6,99,100. Surveys in historic wine regions may reveal the extent of ancient diversity in traditional agrosystems, including well-known regional and foreign varieties, as well as previously unknown varieties grown from seeds or cultivated V. sylvestris plants101. Temporal and spatial variations in grape morphotypes reflect adaptation to environmental conditions and usage, notably marked by the expansion of the western wine type at the time of the extension of wine-growing in Roman Mediterranean France and during the Middle Ages in inland regions. Seed morphometric data reveal a complex evolutionary history, characterized by both rapid and gradual processes, encompassing long-term fixation and spread of selected traits alongside dynamic mechanisms, probably involving sexual crosses and the integration of less productive genotypes into the viticultural system. This dynamism was probably stronger during periods of significant viticultural changes but slowed down after the Middle Ages. This trajectory is in line with a landscape-level domestication process over long periods of time, involving long-distance germplasm exchange and continuous gene flow with wild populations102.
Materials and methods
Archaeological material
Collaborative efforts among 25 scientists and 17 laboratories throughout France enabled the assembly of a comprehensive dataset of 231 samples of archaeological seeds, coming from 176 sites. These samples, drawn from the repositories of participating laboratories, encompass a diverse array of archaeological contexts and represent a crucial resource for studying ancient grapevine domestication.
The collected samples have been subject to meticulous curation, some remaining housed at the Institute of Evolutionary Sciences of Montpellier (ISEM), while others were returned to their respective laboratories and institutions or entrusted to the custodianship of the French Ministry of Culture for long-term preservation. Our selection criteria prioritized seeds preserved through charring and waterlogging. Mineralized seeds were not considered due to their strong and highly variable deformation depending on the taphonomic history of the assemblages.
To establish temporal frameworks for our analyses, samples were dated based on their stratigraphic context, with the stratigraphic levels themselves being dated either by calibrated radiocarbon dates or by archaeological material, occasionally by dendrochronology. The chronology of 17 samples was ascertained by direct radiocarbon dating of V. vinifera seeds. Subsequent data analysis involved the use of either mean dates of stratigraphic phases or the categorizing of samples into broader chrono-cultural periods including the Epipaleolithic/Mesolithic (10,000–5,700 BCE), the Neolithic (5,700-2,200 BCE), the Bronze Age (2,200–750 BCE), the Iron Age (750–50 BCE), the Roman period (50 BCE-500 CE), the Middle Ages (500–1,500 CE) and the Modern period (1,500–1,900 CE).
Geographical distribution of sites was classified into four distinct ecological regions, in accordance with the classification delineated by the National Forest Inventory103: Mediterranean (MEDIT), characterized by its hot and dry climate; West (WEST), with an oceanic climate; Centre-North (CENO), typified by extensive plains and semi-oceanic conditions; and East-Massif Central (EAMC), encompassing mountainous terrain and a mountain or semi-continental climate.
Modern material
This investigation builds upon the modern dataset analyzed by Bonhomme et al.39, comprising 466 V. vinifera cultivars and 80 V. sylvestris accessions originating from diverse regions across West Asia, Europe, and the Mediterranean basin. The majority of the cultivars were procured from the INRAE Grape Germplasm Repository located at Domaine de Vassal, Marseillan-Plage, France (https://vassal.montpellier.hub.inrae.fr), and the Saguramo Grape Repository situated in Jighaura, Georgia. V. sylvestris accessions were obtained from collections belonging to the Institute of Evolutionary Sciences of Montpellier (ISEM), France.
This modern dataset serves as a comparative reference for the analysis and classification of archaeological samples, allowing categorization based on key parameters including domestication status (wild vs. domestic), primary cultivar use (table vs. wine), supposed geographical origin, and genetic clustering, as delineated by Bonhomme et al.39. Our analysis utilizes the four principal Use and Regional Groups (URG) identified by Laucou et al.10, derived from a comprehensive examination of 18,000 single nucleotide polymorphisms (SNPs) across a dataset comprising 783 modern accessions. These groups include Eastern table cultivars, wine cultivars originating from the Balkans, wine cultivars from Western Europe, and a distinct category comprising wine-table grapes from the Iberian Peninsula.
Morphometrics
Pip shape was studied by quantitative seed outline description, using elliptical Fourier transforms (EFT), which proved powerful in differentiating between wild and domestic grape seeds, as well as different groups of cultivars within the domestic type14,30,32,39. We applied the morphometric procedure employed in these studies. To eliminate the risk of inter-operator variability, modern and archaeological grape pips were photographed, by one single operator, in dorsal and lateral views, using a stereomicroscope and digital camera. The images were converted into black shapes. The (x; y) coordinates of 360 equidistant points along the curvilinear abscissa were extracted and two landmarks positioned (one at each tip of the pips) on each image. Landmarks were used to normalize the position, size, rotation and first point of the outlines. After EFT computations we kept only the five first harmonics, which represent 95% of the total harmonic power. For each seed, 40 shape descriptors, often named “Fourier coefficients” (2 views x 5 harmonics x 4 coefficients per harmonic) were used as quantitative variables. Pip size is approximated based on their length, averaged between the two views, and numerically obtained from outline coordinates39.
Correcting the deformation of archaeological seeds
Our archaeological material was preserved either by charring or waterlogging, with each mode inducing differential alterations in the size and shape of the seeds. Several experimental studies have been dedicated to the impact of charring on grape pip morphology33,41,46,47, which document and quantify the characteristic reduction in size and rounding of charred pips. As a result of this process seed shape of cultivated grape varieties moves towards the morphology of their wild counterparts. Recent investigations also shed light on the effects of waterlogging on seed shape42, revealing a more constrained deformation pattern, primarily affecting the beak of the seed.
Despite these alterations, shape differences between V. sylvestris and V. vinifera pips, and between groups of interest within the latter, remain detectable by EFT outline analysis. However, the discriminatory effectiveness of the method diminishes, particularly for charred specimens and when attempting to distinguish between different groups of varieties41,42.
In this study, we conducted comparative analyses between archaeological seeds and a modern reference collection, employing raw EFT coefficients and length measurements for identifications. Concurrently, we undertook similar comparisons and identifications after previously correcting the effects of charring and waterlogging on the archaeological seeds. To mitigate deformation, an average “reverse” vector of shape changes was applied independently to charred and waterlogged seeds, calibrated by observations on experimentally charred and waterlogged seeds. For charring, we used the results obtained for pips charred at 250 °C under reducing conditions41. Pip length, which is independent of shape, was also corrected using the same experimental data. For waterlogging, we used the average deformation among pips soaked in HNO3 and NaOH for 360 min42, only for shape correction as pip length is less affected by waterlogging than charring. Finally, three different datasets were studied: EFT (shape), EFT + L (form), EFT*+L* (form corrected).
Status and variety groups inference on archaeological material
Linear discriminant analyses (LDA) were used to infer the status (wild vs. domestic) and variety groups using the grouping into 4 Use and Regional Groups (URG) obtained from 18,000 genetic SNP markers (SNP4): East-Table, Balkans-Wine, Iberian-Wine, West-Wine10,39. For each group of interest, we trained 100 different balanced LDA on the modern reference collection, then inferred the status/URG for each archaeological pip. Such permutational-based LDA is described elsewhere37,39,104. Each LDA is trained on a different set of modern pips, targeting 50% of the smallest group, which both balances sample size and allows more robust training dataset in terms of e.g. varietal sampling. Each pip having 100 predictions, we considered the majority result (≥ 51). We then calculated proportion for status/URG prediction in each archaeobotanical sample. These proportions allowed us to test differences between successive periods (using Wilcoxon rank sum tests) and within periods, among regions or URG (using Kruskal-Wallis rank sum tests). Analyses were done separately on charred and waterlogged material.
Pip size and approximation of berry size evolution
We utilized the length measurements of archaeological grape pips to infer the volume of ancient berries and track its temporal evolution. Berry size stands as a primary target of domestication, with the fruits of domestic grapevines exhibiting substantial enlargement (ranging from 8 to over 30 mm) compared to their wild counterparts (< 10 mm)78. Previous research established a strong correlation between seed size and berry size43,45, paving the way for the inference of berry dimensions from archaeobotanical remains based on pip dimensions44.
In this study, we used the modern reference collection of Bonhomme et al.44. Because we only had pip length and no information on the status, we constructed a new linear model aiming at predicting berry volume from pip length. To linearize the relationship, we used the cubic-root of the berry volume, a degree 3 model with no intercept. Backward stepwise selection was used using Akaike’s criterion. This ended up with a predictive model of berry volume (back-transformed) from pip length. The predicted volumes of ancient berries were juxtaposed with the measured volumes of berries from a diverse collection of 49 modern accessions, encompassing wild grapevines, wild vines cultivated in INRAE ampelographic collection, as well as cultivated wine and table varieties. This comparative analysis offers insights into the evolutionary trajectory of grape berry size and the domestication process.
Supplementary Information
Acknowledgements
This research was funded by French National Agency, ANR-16-CE27-0013 “Vignes et vins en France du Néolithique au Moyen Age. Approche intégrée en archéosciences” and ANR-22-CE27-0026 “Modelling interactions between climate change and agriculture in the ancient west”. AE has received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation program (grant agreement No. 852573).We are deeply grateful to all the directors of the archaeological excavations and their teams who made it possible to collect the samples studied in this work. We are grateful to the Centre de Ressources biologiques de la Vigne, INRAE, Domaine de Vassal-Montpellier (France) and to the Saguramo Scientific Research Center of Agriculture, Jighaura (Georgia) which allowed us to collect the pips from modern cultivated varieties used in this study. All relevant permits or permissions have been obtained to obtain the plants and the studies conducted comply with local and national regulations or guidelines.
Author contributions
Conceptualization: L.B., J.F.T., V.B., R.B., T.L.; Data curation: all authors; Formal analysis: V.B., L.B., A.E.; Funding acquisition: L.B., A.E.; Investigation: L.B., V.B., S.I., T.P.; Methodology: V.B., L.B.; Project administration: L.B., J.F.T.; Resources: all authors; Software: V.B.; Visualization: V.B., L.B.; Writing—original draft: L.B., V.B.; Writing—review and editing: all authors.
Data availability
Full datasets are available at this address. https://doi.org/10.6084/m9.figshare.25907659.v2.
Declarations
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.McGovern, P. E. Ancient Wine: The Search for the Origins of Viniculture (Princeton University Press, 2019). [Google Scholar]
- 2.Dong, Y. et al. Dual domestications and origin of traits in grapevine evolution. Science379, 892–901 (2023). [DOI] [PubMed] [Google Scholar]
- 3.Fuller, D. Q. & Stevens, C. J. Between domestication and civilization: The role of agriculture and arboriculture in the emergence of the first urban societies. Veget. Hist. Archaeobot.28, 263–282 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.McGovern, P. et al. Early Neolithic wine of Georgia in the South Caucasus. Proc. Natl. Acad. Sci. U.S.A.144 (2017). [DOI] [PMC free article] [PubMed]
- 5.Zohary, D., Hopf, M. & Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin (Oxford University Press, 2012). [Google Scholar]
- 6.Lacombe, T. et al. Large-scale parentage analysis in an extended set of grapevine cultivars (Vitis vinifera L). Theor. Appl. Genet.126, 401–414 (2013). [DOI] [PubMed] [Google Scholar]
- 7.Bacilieri, R. et al. Genetic structure in cultivated grapevines is linked to geography and human selection. BMC Plant. Biol.13, 25 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Magris, G. et al. The genomes of 204 Vitis vinifera accessions reveal the origin of European wine grapes. Nat. Commun.12, 7240 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Emanuelli, F. et al. Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC Plant. Biol.13, 39 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Laucou, V. et al. Extended diversity analysis of cultivated grapevine Vitis vinifera with 10K genome-wide SNPs. PLoS ONE13, e0192540 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Negrul, A. M. Origin and classification of cultivated grape. In The Ampelography of the USSR Vol. 1 (eds Baranov, A. et al.) 159–216 (Pischepromizdat, 1946). [Google Scholar]
- 12.OIV. Distribution of the World’s Grapevine Varieties (International Organisation of Vine and Wine, 2017).
- 13.Morales-Castilla, I. et al. Diversity buffers winegrowing regions from climate change losses. Proc. Natl. Acad. Sci. U.S.A.117, 2864–2869 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pagnoux, C. et al. Local domestication or diffusion? Insights into viticulture in Greece from Neolithic to Archaic times, using geometric morphometric analyses of archaeological grape seeds. J. Archaeol. Sci.125, 105263 (2021). [Google Scholar]
- 15.Valamoti, S. M. Harvesting the ‘wild’? Exploring the context of fruit and nut exploitation at Neolithic Dikili Tash, with special reference to wine. Veget. Hist. Archaeobot.24, 35–46 (2015). [Google Scholar]
- 16.Brun, J. P. Archéologie Du Vin et de l’huile de La Préhistoire à l’époque Hellénistique (Errance, 2004). [Google Scholar]
- 17.Bouby, L. et al. The Holocene history of grapevine (Vitis vinifera) and viticulture in France retraced from a large-scale archaeobotanical dataset. Palaeogeogr. Palaeoclimatol. Palaeoecol.625, 111655 (2023). [Google Scholar]
- 18.Santos, J. A. et al. A review of the potential climate change impacts and adaptation options for European viticulture. Appl. Sci.10, 3092 (2020). [Google Scholar]
- 19.Brun, J. P. Viticulture et oléiculture en Gaule. In Comment les Gaules devinrent romaines 231–253 (La Découverte, Paris, (2010).
- 20.Rice, C. M. Comparative advantage, specialized viticulture, and the economic development of Gallia Narbonensis. J. Roman Archaeol.36, 261–299 (2023). [Google Scholar]
- 21.Brun, J. P. La viticulture en gaule tempérée. Gallia68, 1–12 (2011). [Google Scholar]
- 22.Bernigaud, N., Bondeau, A. & Guiot, J. Understanding the development of viticulture in Roman Gaul during and after the Roman climate optimum: The contribution of spatial analysis and agro-ecosystem modeling. J. Archaeol. Sci. Rep.38, 103099 (2021). [Google Scholar]
- 23.Lachiver, M. Vins, vignes et vignerons: histoire du vignoble français (le Grand livre du mois, Paris, 2006). [Google Scholar]
- 24.Fuller, D. Q. Long and attenuated: Comparative trends in the domestication of tree fruits. Veget. Hist. Archaeobot.27, 165–176 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gros-Balthazard, M. et al. The discovery of wild date palms in Oman reveals a complex domestication history involving centers in the Middle East and Africa. Curr. Biol.27, 2211-2218e8 (2017). [DOI] [PubMed] [Google Scholar]
- 26.Jesus, A. et al. A morphometric approach to track opium poppy domestication. Sci. Rep.11, 9778 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Q Fuller, D. et al. Convergent evolution and parallelism in plant domestication revealed by an expanding archaeological record. Proc. Natl. Acad. Sci. U.S.A.111, 6147–6152 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Roushannafas, T., Bogaard, A. & Charles, M. Geometric morphometrics sheds new light on the identification and domestication status of ‘new Glume Wheat’ at Neolithic Çatalhöyük. J. Archaeol. Sci.142, 105599 (2022). [Google Scholar]
- 29.Terral, J. F. et al. The shape diversity of olive stones resulting from domestication and diversification unveils traits of the oldest known 6500-years-old table Olives from Hishuley Carmel Site (Israel). Agronomy11, 2187 (2021). [Google Scholar]
- 30.Bouby, L. et al. Tracking the history of grapevine cultivation in georgia by combining geometric morphometrics and ancient DNA. Veget. Hist. Archaeobot.30, 63–76 (2021). [Google Scholar]
- 31.Karasik, A., Rahimi, O., David, M., Weiss, E. & Drori, E. Development of a 3D seed morphological tool for grapevine variety identification, and its comparison with SSR analysis. Sci. Rep.8, 6545 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Terral, J. F. et al. Evolution and history of grapevine (Vitis vinifera) under domestication: New morphometric perspectives to understand seed domestication syndrome and reveal origins of ancient European cultivars. Ann. Bot.105, 443–455 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mangafa, M. & Kotsakis, K. A new method for the identification of wild and cultivated charred grape seeds. J. Archaeol. Sci.23, 409–418 (1996). [Google Scholar]
- 34.Ucchesu, M. et al. Earliest evidence of a primitive cultivar of Vitis vinifera L. during the Bronze age in Sardinia (Italy). Veget. Hist. Archaeobot.24, 587–600 (2015). [Google Scholar]
- 35.Chen, G. et al. Morphotype broadening of the grapevine (Vitis vinifera L.) from oxus civilization 4000 BP, Central Asia. Sci. Rep.12, 16331 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Valera, J., Matilla-Seiquer, G., Obón, C., Alcaraz, F. & Rivera, D. Grapevine in the ancient upper Euphrates: Horticultural implications of a Bayesian morphometric study of archaeological seeds. Horticulturae9, 803 (2023). [Google Scholar]
- 37.Bonhomme, V. et al. Seed morphology uncovers 1500 years of vine agrobiodiversity before the advent of the champagne wine. Sci. Rep.11, 2305 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Levadoux, L. Les Populations Sauvages et cultivées des Vitis vinifera L. Ann. Amélioration Des. Plantes1, 59–117 (1956). [Google Scholar]
- 39.Bonhomme, V. et al. Pip shape echoes grapevine domestication history. Sci. Rep.11, 21381 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Pagnoux, C. et al. Inferring the agrobiodiversity of Vitis vinifera L. (Grapevine) in ancient Greece by comparative shape analysis of archaeological and modern seeds. Veget. Hist. Archaeobot.24, 75–84 (2015). [Google Scholar]
- 41.Bouby, L. et al. Back from burn out: Are experimentally charred grapevine pips too distorted to be characterized using morphometrics?. Archaeol. Anthropol. Sci.10, 943–954 (2018). [Google Scholar]
- 42.Bouby, L., Bonhomme, V., Ivorra, S. & Pastor, T. Experimental waterlogging of grape seeds, impact on seed shape and geometrical reversing for morphometric inference. J. Archaeol. Sci. Rep.51, 104204 (2023). [Google Scholar]
- 43.Bouby, L. et al. Bioarchaeological insights into the process of domestication of grapevine (Vitis vinifera L.) during Roman Times in Southern France. PLoS ONE8, e63195 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bonhomme, V. et al. Eco-evo-devo implications and archaeobiological perspectives of trait covariance in fruits of wild and domesticated grapevines. PLoS ONE15, e0239863 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Terpo, A. The carpological examination of wild-growing vine species of Hungry. II, Qualitative and quantitative chraracteristics of vine seeds. Acta Bot. Acad. Sci. Hung. 23.
- 46.Smith, H. & Jones, G. Experiments on the effects of charring on cultivated grape seeds. J. Archaeol. Sci. 317–327 (1990).
- 47.Ucchesu, M. et al. Predictive method for correct identification of archaeological charred grape seeds: Support for advances in knowledge of grape domestication process. PLoS ONE11, e0149814 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bouby, L., Marinval, P. & Terral, J. F. From secondary to speculative production? The protohistorical history of viticulture in Southern France. In Plants and People: Choices and Diversity through Time 175–181 (London & Philadelphia, London & Philadelphia, 2014).
- 49.Py, M. & Buxo Capdevila, R. La Viticulture en Gaule à l’Âge Du Fer. Gallia58, 29–43 (2001). [Google Scholar]
- 50.Ferrer, M., Echeverria, G. & Carbonneau, A. Effect of berry weight and its components on the contents of sugars and anthocyanins of three varieties of Vitis vinifera L. under different water supply conditions. S. Afr. J. Enol. Vitic.35, 103–113 (2014). [Google Scholar]
- 51.Ojeda, H., Deloire, A. & Carbonneau, A. Influence of water deficits on grape berry growth. VITIS J. Grapevine Res.3, 141–145 (2001). [Google Scholar]
- 52.Cherel, A. F. & Frère, D. Du vin en Bretagne dès le premier âge du Fer ? Fabrication et consommation locales. Archeopages 12–21, 10.4000/archeopages.5018 (2020).
- 53.Dodd, E. The archaeology of wine production in Roman and Pre-roman Italy. Am. J. Archaeol.126, 443–480 (2022). [Google Scholar]
- 54.Mercuri, A. M. et al. Sharing the agrarian knowledge with archaeology: First evidence of the dimorphism of Vitis pollen from the Middle Bronze age of N Italy (Terramara Santa Rosa Di Poviglio). Sustainability13, 2287 (2021). [Google Scholar]
- 55.Pecci, A. et al. Wine consumption in bronze age Italy: Combining organic residue analysis, botanical data and ceramic variability. J. Archaeol. Sci.123, 105256 (2020). [Google Scholar]
- 56.Allaby, R. G., Stevens, C., Lucas, L., Maeda, O. & Fuller, D. Q. Geographic mosaics and changing rates of cereal domestication. Philos. Trans. R Soc. B372, 20160429 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Dietler, M. Archaeologies of Colonialism: Consumption, Entanglement, and Violence in Ancient Mediterranean France (University of California Press, 2010). 10.1525/9780520947948. [Google Scholar]
- 58.Garcia, D. La Celtique Méditerranéenne. Habitats et Sociétés En Languedoc et En Provence VIIIe-IIe Siècles Av. J.C (Errance & Picard, 2014). [Google Scholar]
- 59.Py, M. Les Gaulois Du Midi. De La Fin de l’âge Du Bronze à La Conquête Romaine (Errance, 2012). [Google Scholar]
- 60.Sourisseau, J. C. Documents archéologiques et réseaux d’échanges en Méditerranée centrale. In Mobilités grecques. Mouvements, réseaux, contacts en Méditerranée, de l’époque archaïque à l’époque hellénistique (eds Capdetrey, L. & Zurbach, J.) 179–197 (Ausonius Editions, Pessac, 2012). [Google Scholar]
- 61.Forni, G. P. The Origin of ‘Old World’ Viticulture. In Caucasus and Northern Black Sea Region Ampelography (eds Maghradze, D. et al.) 27–38 (Vitis, Quedlinburg, 2012). [Google Scholar]
- 62.Grassi, F. & De Lorenzis, G. Back to the origins: Background and perspectives of grapevine domestication. IJMS22, 4518 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ramos-Madrigal, J. et al. Palaeogenomic insights into the origins of french grapevine diversity. Nat. Plants5, 595–603 (2019). [DOI] [PubMed] [Google Scholar]
- 64.André, J. Contribution Au Vocabulaire De La Viticulture: les noms de cépages. Revue Des. études Latines30, 126–156 (1952). [Google Scholar]
- 65.Migicovsky, Z. et al. Patterns of genomic and phenomic diversity in wine and table grapes. Hortic. Res.4, 17035 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Bellini, C., Mariotti-Lippi, M., Mori Secci, M., Aranguren, B. & Perazzi, P. Plant gathering and cultivation in prehistoric Tuscany (Italy). Veget. Hist. Archaeobot.17, 103–112 (2008). [Google Scholar]
- 67.Rottoli, M. Before the Empire: Prehistoric fruit gathering and cultivation in northern Italy. In Plants and People Choices and Diversity through Time (eds Chevalier, A. et al.) 163–169 (Oxbow Books, Oxford & Philadelphia, 2014). [Google Scholar]
- 68.Jacquat, C. & Martinoli, D. Wild or cultivated? Study of the grape pips found at Petra, Jordan; 150 B.C.–A.D. 40. Veget. Hist. Archaebot.8, 25–30 (1999). [Google Scholar]
- 69.Breglia, F., Bouby, L., Wales, N., Ivorra, S. & Fiorentino, G. Disentangling the origins of viticulture in the western mediterranean. Sci. Rep.13, 17284 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Gaut, B. S., Díez, C. M. & Morrell, P. L. Genomics and the contrasting dynamics of annual and perennial domestication. Trends Genet.31, 709–719 (2015). [DOI] [PubMed] [Google Scholar]
- 71.Cunha, J. et al. Genetic relationships among Portuguese cultivated and Wild Vitis vinifera L. germplasm. Front. Plant. Sci.11, 127 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.De Michele, R. et al. Phylogenetic relationship among wild and cultivated grapevine in sicily: A hotspot in the middle of the mediterranean basin. Front. Plant. Sci.10, 1506 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Grassi, F. et al. Evidence of a secondary grapevine domestication centre detected by SSR analysis. Theor. Appl. Genet.107, 1315–1320 (2003). [DOI] [PubMed] [Google Scholar]
- 74.Freitas, S. et al. Pervasive hybridization with local wild relatives in Western European grapevine varieties. Sci. Adv.7, eabi8584 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Brun, J. P. Archéologie Du Vin et de l’huile En Gaule Romaine (Errance, 2005). [Google Scholar]
- 76.Carrato, C. Dynamique De La Production vinicole en Gaule Narbonnaise à l’époque Romaine à partir de l’étude des capacités de stockage (Ier s. Av.–Ve s. ap. J.-C.). In A. Making Wine in Western-Mediterranean B. Production and the Trade of Amphorae: Some New Data from Italy: Panel 3.5 (eds Brun, J. P. et al.) 159–166 (Propylaeum, Heidelberg, 2020). [Google Scholar]
- 77.Buffat, L., Pellecuer, C., Mauné, S. & Pomarèdes, H. La Viticulture Antique en Languedoc-Roussillon: La viticulture en Gaule. Galia58, 91–111 (2001). [Google Scholar]
- 78.Galet, P. Dictionnaire Encyclopédique Des Cépages (Hachette, 2000). [Google Scholar]
- 79.Ampélographie: Traité Général de Viticulture. Vol. 1 (Masson et Cie, Paris, 1910).
- 80.Bisson, J. Essai de classement des cépages français en écogéogroupes phénotypiques. J. Int. Des. Sci. de la. Vigne et du Vin33, 105–110 (1999). [Google Scholar]
- 81.Bisson, J. Classification Des Vignes Françaises (éditions Féret, 2009). [Google Scholar]
- 82.Greig, J. The Investigation of a Medieval barrel-latrine from Worcester. J. Archaeol. Sci.8, 265–282 (1981). [Google Scholar]
- 83.Ruas, M. P. Les Plantes consommées Au Moyen Âge en France méridionale d’après les semences archéologiques. Archéologie Du Midi Médiéval15–16, 179–204 (1998). [Google Scholar]
- 84.Ruas, M. P., Bouby, L., Mane, P., Puig, C. & Pradat, B. Les fruits de l’alimentation médiévale en France du Sud, entre marchés, recettes et dépotoirs. Amime23, 195–206 (2005). [Google Scholar]
- 85.André, J. L’alimentation et La Cuisine à Rome (Les Belles Lettres, 2018). [Google Scholar]
- 86.Tillier, M. Economie Végétale Des Espaces Portuaires En Méditerranée Romaine: Alimentation, Productions Locales, Échanges et Paysages (éditions Mergoil, 2023). [Google Scholar]
- 87.Higounet-Nadal, A. Presses universitaires du Midi,. Les jardins urbains dans la France médiévale. In Jardins et vergers (ed. Higounet, C.) 115–144 (1989). 10.4000/books.pumi.22517
- 88.Mane, P. Les Fruits dans les traités culinaires français (XIIIe-XVe siècles). Amime23, 129–144 (2005). [Google Scholar]
- 89.Braudel, F. Civilisation Matérielle, Économie et Capitalisme - Tome 2, Ls Jeux de l’échange (Armand Colin, 2022). [Google Scholar]
- 90.Zhou, Y., Massonnet, M., Sanjak, J. S., Cantu, D. & Gaut, B. S. Evolutionary genomics of grape (Vitis vinifera ssp. vinifera) domestication. Proc. Natl. Acad. Sci. U.S.A.114, 11715–11720 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.This, P., Lacombe, T. & Thomas, M. Historical origins and genetic diversity of wine grapes. Trends Genet.22, 511–519 (2006). [DOI] [PubMed] [Google Scholar]
- 92.Fuller, D. Q., Denham, T. & Allaby, R. Plant domestication and agricultural ecologies. Curr. Biol.33, R636–R649 (2023). [DOI] [PubMed] [Google Scholar]
- 93.Purugganan, M. D. Evolutionary insights into the nature of plant domestication. Curr. Biol.29, R705–R714 (2019). [DOI] [PubMed] [Google Scholar]
- 94.Miller, A. J. & Gross, B. L. From forest to field: Perennial fruit crop domestication. Am. J. Bot.98, 1389–1414 (2011). [DOI] [PubMed] [Google Scholar]
- 95.Meyer, R. S., DuVal, A. E. & Jensen, H. R. Patterns and processes in crop domestication: An historical review and quantitative analysis of 203 global food crops. New Phytol.196, 29–48 (2012). [DOI] [PubMed] [Google Scholar]
- 96.Abbo, S., Gopher, A. & Lev-Yadun, S. Fruit domestication in the near east. Plant. Breed. Rev.39, 325–377 (2015). [Google Scholar]
- 97.Zohary, D. & Spiegel-Roy, P. Beginnings of fruit growing in the old world. Science187, 319–327 (1975). [DOI] [PubMed] [Google Scholar]
- 98.Weiss, E. Beginnings of fruit growing in the old world two generations later. Isr. J. Plant. Sci.62, 75–85 (2015). [Google Scholar]
- 99.D’Onofrio, C. et al. Parentage Atlas of Italian grapevine varieties as inferred from SNP genotyping. Front. Plant. Sci.11, 605934 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Zdunić, G. et al. Genetic structure and relationships among wild and cultivated grapevines from Central Europe and part of the Western Balkan Peninsula. Genes11, 962 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Maraš, V. et al. Population genetic analysis in old Montenegrin vineyards reveals ancient ways currently active to generate diversity in Vitis vinifera. Sci. Rep.10, 15000 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Allaby, R. G., Stevens, C. J., Kistler, L. & Fuller, D. Q. Emerging evidence of plant domestication as a landscape-level process. Trends Ecol. Evol.37, 268–279 (2022). [DOI] [PubMed] [Google Scholar]
- 103.Cavaignac, S. Les sylvoécorégions (SER) de France métropolitaine: Étude de définition (2009).
- 104.Evin, A. et al. The long and winding road: Identifying pig domestication through molar size and shape. J. Archaeol. Sci.40, 735–743 (2013). [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Full datasets are available at this address. https://doi.org/10.6084/m9.figshare.25907659.v2.