

Abstract
The amount of excess polymerase and RNase H activity in human immunodeficiency virus type 1 virions was measured by using vectors that undergo a single round of replication. Vectors containing wild-type reverse transcriptase (RT), vectors encoding the D110E mutation to inactivate polymerase, and vectors encoding mutations D443A and E478Q to inactivate RNase H were constructed. 293 cells were cotransfected with different proportions of plasmids encoding these vectors to generate phenotypically mixed virions. The resulting viruses were used to infect human osteosarcoma cells, and the relative infectivity of the viruses was determined by measuring transduction of the murine cell surface marker CD24, which is encoded by the vectors. The results indicated that there is an excess of both polymerase and RNase H activities in virions. Viral replication was reduced to 42% of wild-type levels in virions with where half of the RT molecules were predicted to be catalytically active but dropped to 3% of wild-type levels when 25% of the RT molecules were active. However, reducing RNase H activity had a lesser effect on viral replication. As expected, based on previous work with murine leukemia virus, there was relatively inefficient virus replication when the RNase H and polymerase activities were encoded on separate vectors (D110E plus E478Q and D110E plus D443A). To determine how virus replication failed when polymerase and RNase H activities were reduced, reverse transcription intermediates were measured in vector-infected cells by using quantitative real-time PCR. The results indicated that using the D11OE mutation to reduce the amount of active polymerase reduced the number of reverse transcripts that were initiated and also reduced the amounts of products from the late stages of reverse transcription. If the E478Q mutation was used to reduce RNase H activity, the number of reverse transcripts that were initiated was reduced; there was also a strong effect on minus-strand transfer.
The viral enzyme reverse transcriptase (RT) converts the single-stranded genomic RNA found in retroviruses into double-stranded DNA. RT contains two enzymatic activities that collaborate in this conversion: a DNA polymerase that can copy either an RNA or a DNA template, and RNase H, which cleaves RNA if (and only if) it is part of an RNA-DNA hybrid (see references 6, 25, and 66 for reviews). Both the polymerase and RNase H activities are necessary for retroviral replication; mutations that block RNase H activity also block virus replication (47, 49, 56, 61). RNase H activity is required during several steps of reverse transcription. RNase H degradation is required for the first-strand transfer reaction and generates the RNA primer for plus-strand DNA synthesis. RNase H also removes the RNA primers used to initiate plus- and minus-strand DNA synthesis (15, 18, 37, 38, 44–46, 51, 58, 62).
Human immunodeficiency virus type 1 (HIV-1) RT is a heterodimeric protein; the two subunits are p66 and p51 (10). The p66 subunit contains 560 amino acids, and the p51 subunit is composed of the same sequence as the first 440 amino acids of the p66 subunit. Based on the crystal structure, the polymerase domain of HIV-1 RT has been likened to a right hand composed of fingers, palm, thumb, and connection subdomains (31). Although the folding within each of these subdomains is similar in the two subunits, the overall spatial arrangement of the subdomains differs between the subunits (27, 31). The polymerase and RNase H activities of RT reside on the p66 subunit. A triad of aspartic acids residues (positions 110, 185, and 186) forms the polymerase active site that is located in the p66 palm subdomain (27, 31). Although the p51 subunit contains the same three aspartic acid residues, the p51 subunit does not directly contribute to the polymerase activity of RT (33). The RNase H domain is primarily derived from the 15-kDa segment that is present only in the p66 subunit; p51 lacks an RNase H domain (6). Crystallographic data show that the RNase H active site is positioned approximately 17 nucleotides from the polymerase active site; this distance is supported by biochemical experiments (16, 19, 27, 43, 67).
The HIV-1 RT RNase H domain and the Escherichia coli RNase H are related in structure and activity. The polypeptides chains share 24% sequence identity and fold into similar structures (29, 68). There is a strict conservation of the amino acids that comprise the RNase H active site. The active-site amino acid residues D443, E478, D498, and D549 of HIV-1 RT correspond to amino acids D10, E48, D70, and D134 of E. coli RNase H (8, 25). Mutations at any of these amino acids profoundly affect RNase H activity and block virus replication. The degradation of the viral RNA by RNase H is not sequence specific, and a range of different-size RNase H cleavage products are generated. However, the RNase H cleavages that generate the polypurine tract (ppt) primer and the cleavages that remove the tRNA and ppt primers are site specific. In HIV-1 replication, the ppt primer is completely removed and the tRNA primer is cleaved one nucleotide from the RNA/DNA junction (44, 45, 52, 65). The generation (and removal) of the ppt primer must be very specific to ensure the proper synthesis of the left end of unintegrated viral DNA; removal of the tRNA primer defines the right end of viral DNA.
Although RT can cleave an RNA template during polymerization, the enzyme's two activities (polymerization and RNase H cleavage) can function independently (9). However, both the structure of RT and the biochemical behavior of wild-type and mutant enzymes show that while the two enzymatic activities of RT are not strictly coupled, the two domains (polymerase and RNase H) are interdependent in their interactions with nucleic acid substrates. The degree of interdependence of polymerase and RNase H domains is somewhat different for different RTs. In the case of murine leukemia virus (MLV) RT, the separately expressed RNase H and polymerase domains retain high levels of enzymatic activity (32, 34, 57). However, with HIV-1 RT, the separately expressed domains have little activity. The lack of activity of the separate RNase H domain of HIV-1 RT appears to be due to weak binding of the substrate. When combined with a purified HIV-1 polymerase domain, the isolated HIV-1 RNase H domain regains activity (24, 53). In addition, introducing a substrate-binding element from E. coli RNase H into the isolated RNase H domain of HIV-1 RT also restores enzymatic activity (30, 54).
The polymerase and RNase H activities of MLV RT can complement inefficiently in vivo when present on separate RT molecules, demonstrating that the two enzymatic activities can act independently to successfully complete the synthesis of viral DNA (60). Various MLV RNase H mutants were tested for the ability to complement RTs lacking polymerase activity. Some RNase H mutants failed to complement, suggesting that these RNase H mutations may also affect the polymerase activity of RT (60). In support of this idea, some RNase H mutations of HIV-1 RT appear to affect the polymerase activity; mutations at position 478 have been reported to affect the in vitro polymerase activity of RT (12, 13, 40, 63). This mutation did not affect the initiation of reverse transcription in vitro, suggesting that DNA synthesis was impaired at the level of elongation (26).
Retroviral virions each contain approximately 50 to 100 molecules of RT. There are approximately 2,000 molecules of Gag in a virion (55; see also reference 6 for a review). The ratio of Gag to Gag-Pol is about 20 to 1 (21, 28, 41). This means that there are about 100 Gag-Pol molecules per virion. For retroviruses, like HIV-1, in which the RT is a dimer, the number of RT molecules in a virion should be approximately 50. This is a relatively large number, and it is possible that there is an excess of polymerase activity, of RNase H activity, or of both. In support of this idea, HIV-2 RT has, in an in vitro assay, approximately 10% of the RNase H activity of HIV-1 RT (50). HIV-2 replicates reasonably well, which suggests the possibility that HIV-1 virions contain excess RNase H activity. In vitro, HIV-1 RT is not a strongly processive polymerase. HIV-1 RT dissociates from its primer-template frequently, suggesting that viral DNA is synthesized in relatively short segments. The relatively low processivity of RT might be the reason that there is a large number of RT molecules in a virion; a relatively large number of RT molecules may be required to complete viral DNA synthesis.
We performed phenotypic mixing experiments in which the virions contained different amounts of catalytically active and catalytically inactive RTs to determine whether there is an excess of either polymerase or RNase H activity in the virion. The D110E mutation was used to inactivate the polymerase; in parallel experiments, RNase H was inactivated using either the D443A or E478Q mutation. The ability of viruses containing different mixtures of active and inactive RTs to replicate was determined. In HIV-1 virions, there is a modest excess of polymerase activity; there is a greater excess of RNase H activity. The process of reverse transcription in vivo was monitored by real-time PCR. The analysis suggests that some RNase H mutations can interfere with polymerase activity in vivo.
MATERIALS AND METHODS
Vector construction.
pNLNgoMIVR−E−.HSA was derived from pNL4-3HSAR−E− (obtained from the AIDS Reagent Program). A vector containing a unique SmaI site and a unique Asp718 site in the pol coding region of the pNL4-3-based vector was constructed. The Asp718 site at position 6343 was removed by digesting pNL4-3HSAR−E− with SalI and BamHI. Then the restriction fragment containing the Asp718 site at position 6343 was ligated into the cloning vector pKSΔAsp718 (which was created by digesting pKS+ [obtained from Stratagene] with Asp718 and then using T4 DNA polymerase and deoxynucleoside triphosphates [dNTPs] to fill in the overhang and religating the vector). pKS containing the SalI-to-BamHI fragment of pNL4-3HSAR−E− was digested with Asp718, then T4 DNA polymerase and dNTPs were used to fill in the overhangs, and the vector was ligated using T4 DNA ligase. The pKS cloning vector containing the SalI-to-BamHI fragment of pNL4-3HSAR−E− that lacked the Asp718 site was digested with SalI and BamHI, and the fragment containing the pNL-derived sequence missing the Asp718 site was cloned into pNL4-3HSAR−E− digested with SalI and BamHI, generating the vector pNL4-3ΔAsp718#3. This vector retains three of the Asp718 sites originally present in pNL4-3, but the site at position 6343 is destroyed. The Asp718 site at position 9006 was removed by digesting pNL4-3HSAR−E− with NsiI (removing bases 1247 to 6738) and religating the vector. This vector, called pNL-NsiI, was digested with Asp718, then treated with T4 DNA polymerase and dNTPs to fill in the overhangs, and religated, generating the vector pNL-NsiIΔAsp718#4. pNL-NsiIΔAsp718#4 was digested with BamHI and NcoI. The fragment containing the filled-in Asp718 site (position 9006) was ligated to pNL pNL4-3ΔAsp718#3 digested with BamHI and NcoI. This generated the vector pNLΔAsp718#3,4. A unique SmaI site was introduced into the pol coding region at the 14th amino acid of RT. A PCR primer 5′ of the ApaI site at position 2005 and a PCR primer that generates the SmaI site in RT were used to amplify pNL4-3HSAR−E−. The PCR product was digested with ApaI and SmaI, and the fragment was cloned into PKS+ΔAsp718 digested with ApaI and SmaI. Next, the Asp718 site at position 4154 was changed to an NgoMIV site by using a PCR primer that anneals to the 5′ end of the RT coding region, a downstream PCR primer that changes the Asp718 site at 4154 to an NgoMIV site, and a previously described RT expression plasmid as the PCR template (3). pNL4-3HSAR−E− was used as a PCR template with a primer that changes the Asp718 site at 4154 to an NgoMIV site plus a primer that lies downstream of the SalI site. These fragments were cloned as a SmaI-to-NgoMIV fragment and an NgoMIV-to-SalI fragment into pKS containing the ApaI-to-SmaI fragment as a three-way ligation. Next, the ApaI-to-SalI fragments were moved into pNLΔ3,4 digested with ApaI and SalI, creating the final vector, pNLNgoMIVR−E−.HSA.
The plasmid encoding the vector with the D110E mutation in RT that inactivates polymerase was created by digesting a previously described RT expression plasmid containing the D110E mutation (3) with SmaI and Asp718, gel eluting the 1.3-kb fragment, and ligating the fragment into pNLNgoMIVR−E−.HSA digested with SmaI and Asp718 by using standard cloning procedures.
The E478Q and D443A mutations in RT were created by subcloning the Asp718-to-SalI fragment of pNLNgoMIVR−E−.HSA into pKS+ and using a QuickChange site-directed mutagenesis kit (Stratagene) to introduce the desired mutations into the RNase H coding region. DNA sequence analysis was performed to ensure that the clones contained only the desired mutation, and then the mutated sequences were cloned into pNLNgoMIVR−E−.HSA as an Asp718-to-SalI fragment.
Cells, transfection, and infection.
The human embryonal kidney cell line 293 was obtained from American Type Culture Collection. The human osteosarcoma cell line HOS was obtained from Richard Schwartz (Michigan State University, Lansing). 293 and HOS cells were maintained in Dulbecco's modified Eagle's medium (Life Technologies) supplemented with 5% fetal bovine serum, 5% newborn calf serum, and penicillin (50 U/ml) plus streptomycin (50 μg/ml) (Quality Biological).
Transfection, infection, and phenotyping protocol.
293 cells were transfected with 2 μg of pNLNgoMIVR−E−.HSA and 2 μg of pHCMV-g (obtained from Jane Burns, University of California, San Diego) by the calcium phosphate method. 293 cells were plated in 100-mm-diameter dishes at a density of 1.8 × 106 cells per plate on the day prior to transfection such that they would be approximately 30% confluent on the day of transfection The precipitate was added to 293 cells dropwise. Eight hours after transfection, the cells were washed once with 8 ml of Dulbecco's phosphate-buffered saline (PBS; Gibco BRL/Life Technologies), and fresh medium was added. The medium was changed again 24 h after transfection. The 48-h supernatants were harvested, clarified by low-speed centrifugation, filtered through a Millex-GS 0.22-μm-pore-size filter (Millipore), and used to infected HOS cells. The amount of p24 in the samples was determined using an HIV-1 p24 antigen capture assay kit to control for the amounts of virus in the samples (AIDS Vaccine Program, SAIC, Frederick, Md.). The virus was allowed to adsorb to the cells for 6 h, and then fresh medium was added. Forty-eight hours after infection, the cells were harvested from the plate by treating with 1.5 ml of EDTA (Gibco BRL/Life Technologies), an additional 2.5 ml of PBS was added; then the cells were collected by centrifugation, washed, resuspended in 200 μl of PBS, labeled with phycoerythrin-conjugated rat anti-mouse CD24 monoclonal antibody (PharMingen) by using standard procedures, fixed with glutaraldehyde, and subjected to fluorescence-activated cell sorting (FACS) to determine virus titer.
Transfections, infections, and nucleic acid extraction for analysis of DNA synthesis using real-time PCR.
293 cells were transfected with 5 μg of pNLNgoMIVR−E−.HSA and 3 μg of pHCMV-g by the calcium phosphate method. The transfections for the real-time PCR experiments were performed with 5 μg of plasmid to maximize the amount of virus in the supernatant and obtain acceptable levels of residual plasmid carryover. After 7 h, the cells were rinsed two times with 8 ml of PBS, and fresh medium was added. The plates were inspected for the presence of precipitate by using a light microscope. Twenty hours after transfection, the cells were washed with 8 ml of PBS, and fresh medium was added. Thirty hours after transfection, the medium was changed again. Forty-eight hours after transfection, the plates were inspected with a light microscope to ensure that the precipitate had been removed. Virus-containing supernatants were harvested and clarified by centrifugation, and 5 ml of the virus-containing supernatant was filtered through a 0.22-μm-pore-size filter; 4 ml of virus was used to infect HOS cells, plated the day prior to infection at 5 × 105 cells per 60-mm-diameter plate. The virus was allowed to adsorb to the cells for 4 h, the cells were washed three times with 4 ml of PBS, and fresh medium was added. Twenty hours after infection, the total DNA was isolated from the cells by the Blood DNA kit protocol (Qiagen) and eluted with 200 μl of elution buffer.
Determination of DNA copy number by real-time PCR.
Real-time PCR can be used to measure the amount of a specific nucleic acid present in a solution. The technique has been described in the literature (23, 35, 36); only a brief description is given here. Real-time PCR monitors the amplification of a specific PCR product during each amplification cycle. Taqman technology uses a nucleic acid probe that contains a fluorescent group and a quencher. This probe is degraded by the exonucleolytic activity of Taq polymerase if it is part of a DNA duplex; degradation separates the fluorescent group from the quencher, increasing the fluorescence in the PCR sample. The fluorescence is monitored during each PCR cycle. When the amount of fluorescence in the sample reaches a certain assigned level, the PCR cycle number needed to achieve this threshold is recorded. By comparing the threshold cycle in a sample to those of standards containing known amounts of the target sequence, the number of copies of the sequence in the sample is determined. Real-time PCR was performed with an ABI7700 apparatus (Applied Biosystems) in a final volume of 50 μl. The PCR mix contained 2× Universal PCR Master Mix (Applied Biosystems) (25 μl), 10 μl of nucleic acid sample, and 5 μl of primers, probe, and water. The PCR primers and Taqman probes were present at a final concentration of 200 nM. The unknown samples were compared to duplicate samples containing 107 to 102 copies of plasmid DNA. The cycling conditions were 2 min at 50°C and 10 min at 95°C, then 40 cycles of 95°C for 15 s and 60°C for 1 min. The Taqman primer sets used to monitor different stages of reverse transcription are given in Table 1.
TABLE 1.
Taqman primers and probes
| Amplicon | Forward primer | Reverse primer | Taqman probea |
|---|---|---|---|
| RU5 | 5′-TCTGGCTAACTAGGGAACCCA-3′ | 5′-CTGACTAAAAGGGTCTGAGG-3′ | 5′-R-TTAAGCCTCAATAAAGCTTGCCTTGAGTGC-Q-3′ |
| U3 | 5′-CTAATTCACTCCCAAAGAAGACAAGA-3′ | 5′-GTACTAGCTTGTAGCACCATCCAAAG-3′ | 5′-R-CACACAAGGCTACTTCCCTGATTGGCAG-Q-3′ |
| Gag | 5′-TTCGCAGTTAATCCTGGCCTT-3′ | 5′-GCACACAATAGAGGACTGCTATTGTA-3′ | 5′-R-TAGAGACATCAGAAGGCTGTAGACAAA-Q-3′ |
| 2nd-strand transfer | 5′-TTTTAGTCAGTGTGGAAAATCTGTAGC-3′ | 5′-TACTCACCAGTCGCCGCC-3′ | 5′-R-TCGACGCAGGACTCGGCTTGCT-Q-3′ |
| 2-LTR circle junction | 5′-GCCTGGGAGCTCTCTGGCTAA-3′ | 5′-GCCTTGTGTGTGGTAGATCCA-3′ | 5′-R-AAGTAGTGTGTGCCCGTCTGTTGTGTGACTC-Q-3′ |
6-Carboxyfluorescin is the reporter dye (R); 6-carboxytetramethylrhodamine is the quencher (Q).
The HIV-1 2-LTR circle junction was cloned into the pKS plasmid as an Asp718-to-Sal fragment by PCR amplification from the 2-LTR circles, using total DNA extracted from infected HOS cells as the template for the reaction. Serial dilutions of this plasmid were used as a standard to determine the copy number of 2-LTR circles in the samples as described for the determination of other DNA copy numbers. The Taqman set used to monitor the presence of 2-LTR circles in the samples is given in Table 1.
RESULTS
Vectors and replication assay.
The pNLNgoMIVR−E−.HSA vectors (Fig. 1A) are based on previously described vectors derived from the NL4-3 isolate of HIV-1 (7, 22). In the experiments described here, the portion of RT located between amino acids 14 and 427 of RT was derived from the BH10 isolate of HIV-1. The env and vpr genes and are inactivated in the vectors (7, 22); this limits the vectors to a single cycle of replication. Cotransfecting 293 cells (Fig. 1B) with vector DNA and the plasmid, pHCMV-g, that expresses the vesicular stomatitis virus G envelope glycoprotein produced infectious virions (1, 69). Forty-eight-hour supernatants were harvested and used to infect HOS cells. The murine heat-stable antigen gene (hsa) (64) is expressed from the nef reading frame of the vector. Infected cells were identified and enumerated by being labeled with anti-HSA and subjected to FACS. Viral titers were determined 48 h after infection.
FIG. 1.

HIV-1 based vector and biological assay for viral infectivity. (A) The HIV-1 vector pNLNgoMIVR−E−.HSA was created from the vector pNL4-3HSAR−E− (obtained from the AIDS Reagent Program) by introducing a unique SmaI restriction enzyme site and making the Asp718 restriction enzyme site in the pol coding region unique (see Materials and Methods for details). The sequences encoding RT from the SmaI site to the Asp718 site were derived from the BH10 isolate of HIV and were subcloned from previously described HIV-1 RT expression plasmids (3, 4). All other viral sequences in the vector are derived from the NL4-3 strain of HIV-1. The vector expresses the murine cell surface marker CD24 (HSA) from the nef open reading frame. The env gene and the vpr gene have been inactivated in this vector. The drawing is not to scale. Infectious virus was produced by pseudotyping with the vesicular stomatitis virus G envelope glycoprotein (expressed from pHCMV-g). The vector undergoes only a single cycle of replication. pbs, primer binding site. (B) 293 cells were cotransfected with pNLNgoMIVR−E−.HSA vectors and pHCMV-g to generate virus stocks. Forty-eight hours after transfection, virus-containing supernatants were used to infect the HOS cell line. Forty-eight hours following infection, the HOS cells were labeled with antibody against the HSA. The cells were then fixed with formaldehyde and subjected to FACS to determine the virus titer. ELISA, enzyme-linked immunosorbent assay.
Phenotypic mixing experiments.
To determine if there is an excess of polymerase or RNase H activity in HIV-1 virions, we performed phenotypic mixing experiments in which virions were produced that contain different amounts of catalytically active and inactive RT. 293 cells were cotransfected with different proportions of plasmids encoding the wild-type vector (pNLNgoMIVR−E−.HSA) and vectors that contained mutations that inactivate polymerase (D110E) or RNase H (D443A or E478Q). The relative amount of active RT was inferred from the mixture of plasmids used to generate the virus.
293 cells were cotransfected with mixtures of plasmids encoding vectors containing wild-type RT and the D110E mutation to produce virions containing 100% or approximately 50, 25, 10, or 0% of the polymerase activity present in wild-type virions. The effect of decreasing the percentage of RT having polymerase activity on relative virus titer is shown in the Fig. 2A. When 50% of the plasmids encoded wild-type RT and the other 50% encoded the D110E mutant, the relative infectivity of the virus stock was 42%. When the percentage of plasmids encoding wild-type RT was decreased to 25%, the relative infectivity was only 3% of the wild-type level. When less than 25% of the plasmid encoded wild-type RT, there was no measurable infection of HOS cells.
FIG. 2.

Limiting polymerase or RNase H activity affects virus infectivity. (A) 293 cells were cotransfected with different proportions of plasmid containing vector wild-type RT and vector containing the D110E mutation in RT. This generated virions containing different percentages of active polymerase. (B) 293 cells were cotransfected with different proportions of plasmid containing vector encoding wild-type RT and vector containing the E478Q mutation in RT. This generated virions containing different percentages of active RNase H. (C) 293 cells were cotransfected with different proportions of plasmid containing vector encoding wild-type RT and vector containing the D443A mutation in RT. This generated virions containing different percentages of active RNase H. In each case, the predicted percentage of polymerase or RNase H activity is shown on the x axis, and the relative infectivity, expressed as the percentage of the wild-type control, is shown on the y axis. The relative amounts of polymerase or RNase H activity are inferred from the mixtures of plasmids used to generate the virions.
To measure the amount of excess RNase H activity in virions, we prepared phenotypically mixed virions that contained 100, 50, 25, 10, and 0% of the RNase H activity of wild-type virions. These virions were prepared by cotransfecting 293 cells with plasmid encoding the wild-type vector and a vector encoding the E478Q mutation in RT. The effect of decreasing the amount of RNase H activity on virus replication was not as dramatic as the effect of decreasing the polymerase activity (Fig. 2B). When virions contained 50 or 25% of the wild-type level of RNase H, the specific infectivity was 70 or 42%, respectively, of that of the wild-type vector. When the amount of RNase H activity was decreased to 10%, a greater effect was seen; the relative infectivity was still 8% of the wild-type level. When all RT in the virions had the E478Q mutation, there was no measurable infection.
When the D443A mutation was used to inactivate the RNase H activity, the results were similar to the results of the E478Q mixing experiments, although the virions seemed to tolerate low levels of the D443A mutation better than low levels of the E478Q mutation. When 50% of the RT contained the D443A mutation there was no measurable effect on infectivity. When the percentage of RT containing the D443A mutation was increased to 75 or 90% the relative infectivity was about 45 or 35%, respectively, of the wild-type level. When the virion contained only RT carrying the D443A mutation, no infected HOS cells were found.
Monitoring reverse transcription in vivo using real-time PCR.
Real-time PCR can be used to determine the copy number of specific nucleic acids. The strategy used to monitor specific steps of reverse transcription is shown in Fig. 3. As shown in Fig. 3A, minus-strand DNA synthesis is initiated from a tRNA primer bound to the RNA genome at the primer binding site. When the U5 and R regions of the viral RNA are copied into DNA, the resulting product is called minus-strand strong-stop DNA (−sssDNA). The synthesis of −sssDNA can be measured using a set of PCR primers and a Taqman probe that anneals to the R and U5 regions of the DNA (Table 1). Once −sssDNA is synthesized, it is transferred to the 3′ end of the RNA in a process called first strand (or minus-strand) transfer. Since the U3 region of the RNA is copied into DNA immediately after the strand transfer step, measuring viral DNA specific for U3 and comparing it to the amount of −sssDNA monitors first-strand transfer. RT then copies the viral structural genes; comparison of the amounts of DNA specific for structural genes such as env, pol, and gag can be used to monitor the progress of minus-strand synthesis. Plus-strand DNA transfer can be measured if the PCR amplicon spans the primer binding site. Plus-strand elongation and completion of reverse transcription cannot be monitored directly because PCR-based methods do not readily distinguish complementary DNA sequences. However, monitoring the formation of the 2-LTR circles, which arise from the ligation of the ends of the full-length linear viral DNA, can be used as a surrogate for the completion of reverse transcription. Viral DNA and locations of the Taqman primer sets used to monitor individual steps in reverse transcription are depicted in the Fig. 3B.
FIG. 3.

Use of real-time PCR to monitor viral DNA synthesis. (A) The process of reverse transcription. The thin line represents viral RNA, and the thick line represents viral DNA synthesized by RT. RNase H cleavage of RNA is shown by a dashed line. Measuring the amount of RU5 DNA synthesized monitors minus-strand DNA synthesis; measuring the amount of U3 DNA synthesized monitors minus-strand DNA transfer; measuring DNA synthesis in the gag gene monitors minus-strand elongation. PCR primers spanning the primer binding site (pbs) determine the amount of plus-strand transfer. Monitoring the formation of 2-LTR circles using primers spanning the 2-LTR circle junctions serves as a surrogate for the completion of reverse transcription. The drawing is not to scale. (B) Structures of a DNA provirus and of PCR primer and probe sets. Boxes on the viral DNA indicate the positions of Taqman probe and primer sets during each successive step of reverse transcription.
Limiting polymerase activity affects DNA synthesis.
Phenotypically mixed virions were generated by cotransfecting 293 cells with different proportions of plasmids carrying a vector encoding wild-type RT and vector encoding the D110E mutation in RT. The 48-h supernatants were harvested, clarified by centrifugation, filtered, and used to infect HOS cells for 4 h (see Materials and Methods for details). The HOS cells were washed three times with 4 ml of PBS and then maintained in medium for 20 h. Total DNA was extracted from the infected cells, and the synthesis of viral DNA was analyzed by real-time PCR.
In samples of virus that contain only RT carrying the D110E mutation, 2-LTR circles were not detected, and 1.0 × 102 copies of RU5 and U3 were detected. We also detected 2.1 × 102 copies of gag and 8.0 × 101 copies of strand transfer products, which we believe represents residual plasmid carried over from the transfection. Samples derived entirely from wild-type vector contained 2.4 × 106 copies of DNA corresponding to the RU5 region of HIV-1 (Table 2). Since 4 orders of magnitude more products were detected in cells infected with vectors carrying wild-type RT, this small amount of plasmid carryover is not critical to the experiments. There were 7.9 × 105 copies of U3, indicating that about 30% of the −sssDNA undergoes minus-strand DNA transfer; the detection of 6.9 × 105 copies of gag indicated that minus-strand DNA synthesis was also quite efficient. The presence of 2.0 × 105 copies of the product spanning the primer binding site indicated that plus-strand transfer occurred approximately 10% of the time −sssDNA was made. There was approximately 0.2% the amount of the 2-LTR circles as for RU5 (5.5 × 103 versus 2.4 × 106).
TABLE 2.
Quantitative analysis of viral DNA synthesis in cells infected with phenotypically mixed viruses containing wild-type RT and RT lacking polymerase activity
| Virus stocka | DNA synthesis (no. of copies)
|
||||
|---|---|---|---|---|---|
| RU5 | U3 | Gag | Plus-strand transfer | Circle junction | |
| 100% wt | 2.4 × 106 | 7.9 × 105 | 6.9 × 105 | 2.0 × 105 | 5.5 × 103 |
| 20% wt + 80% D110E | 2.9 × 104 | 1.2 × 104 | 1.2 × 104 | 6.2 × 102 | Not detected |
| 10% wt + 90% D110E | 1.3 × 104 | 7.2 × 103 | 4.5 × 103 | 2.0 × 102 | Not detected |
| 100% D110E | 1.0 × 102 | 1.0 × 102 | 2.1 × 102 | 8.0 × 101 | Not detected |
Virus stocks were derived by transfecting 293 cells with different proportions of plasmids carrying vectors encoding wild-type (wt) RT or the D110E mutation in RT that inactivates the polymerase activity.
When the amount of RT containing polymerase activity was decreased, the profile of DNA synthesis changed. There were significant decreases in the synthesis of −sssDNA in samples containing 20 and 10% polymerase compared to the wild type (2.9 × 104 and 1.3 × 104 copies versus 2.4 × 106 copies). Minus-strand DNA transfer and synthesis through the gag gene were relatively efficient in the samples containing 10 and 20% active RT. However, the percentage of DNA undergoing plus-strand DNA transfer decreased relative to virions containing only wild-type RT; no 2-LTR circles were detected. These results are consistent with the results of the biological assay that indicated that retroviral replication is severely impaired when the percentage of polymerase activity is less than 25% of the wild-type level. These results do not support the suggestion that the low processivity of HIV-1 RT plays a role in the requirement for a large number of active RTs in a virion but suggest instead that it is the initiation of minus-strand synthesis and some step or steps in plus-strand synthesis that are limiting (see Discussion).
Effects of limiting RNase H levels on DNA synthesis.
When the virions contained only RT carrying the E478Q mutation, 1.6 × 104 copies of −sssDNA were produced (Table 3). Approximately 2 × 102 copies of U3, gag, and strand transfer products were detected. As has already been discussed, approximately 102 copies presumably represent residual plasmid DNA from the transfection of the 293 cells. No 2-LTR circles were detected. This result suggests that there is a defect in the initiation of viral DNA synthesis as well as a block to first-strand transfer.
TABLE 3.
Quantitative analysis of viral DNA synthesis in cells infected with phenotypically mixed viruses containing wild-type RT and RT lacking RNase H activity
| Virus stocka | DNA synthesis (no. of copies)
|
||||
|---|---|---|---|---|---|
| RU5 | U3 | Gag | Plus-strand transfer | Circle junction | |
| 100% wt | 2.4 × 106 | 7.9 × 105 | 6.9 × 105 | 2.0 × 105 | 5.5 × 103 |
| 10% wt + 90% E478Q | 4.6 × 105 | 1.5 × 105 | 1.1 × 105 | 2.8 × 104 | 2.2 × 102 |
| 5% wt + 95% E478Q | 1.1 × 105 | 4.1 × 104 | 3.3 × 104 | 4.9 × 103 | 5.4 × 101 |
| 100% E478Q | 1.6 × 104 | 1.8 × 102 | 2.2 × 102 | 1.9 × 102 | Not detected |
Virus stocks were derived by transfecting 293 cells with different proportions of plasmids carrying vectors encoding wild-type (wt) RT or the E478Q mutation in RT that inactivates the RNase H.
When virions containing only 10 or 5% of the wild-type level of RNase H activity were used to infect HOS cells, DNA synthesis was moderately affected. Initiation of reverse transcription was reduced by approximately 90% (4.5 × 105 and 1.1 × 105 copies, for 10 and 5% activity respectively, versus 2.4 × 106 for the wild type). About 30% of the −sssDNAs were able to complete minus-strand transfer. DNA synthesis through the structural genes was relatively efficient for those samples that completed strand transfer. In samples with limiting RNase H activity, plus-strand DNA transfer occurred about 5% of the time −sssDNA was synthesized. Circle junctions were detected at about 0.05% of the level of −sssDNA when 10 or 5% RNase H activity was present.
Virions containing defective RTs carrying the D110E mutation to inactivate polymerase and D443A or E478Q to inactivate RNase H infect cells inefficiently.
To determine if virions containing the polymerase and RNase H activities on separate RTs can undergo reverse transcription and infect cells, we performed phenotypic mixing experiments in which approximately half of the RTs in the virion possess the D110E mutation and half possess either the E478Q or D443A mutation. The resulting virions are solely dependent on the RTs containing the D110E mutation for RNase H activity and on the RTs containing either E478Q or D443A for polymerase activity.
293 cells were cotransfected with a mixture of plasmids encoding wild-type RT or a 50-50 mixture of plasmids encoding the D110E mutation to inactivate the polymerase activity of RT and plasmid encoding either the E478Q or D443A mutation to inactivate RNase H activity of RT. Results for the viruses containing the two RT activities on separate RT molecules are shown in Fig. 4. The infection of HOS cells was reduced to approximately 5 or 8%, respectively, for viruses containing the D443A-D110E mixture of RTs or the E478Q-D110E mixture of RTs compared to virions containing wild-type RT. This result indicates that the two defective RTs complement inefficiently. A similar result has been reported for MLV (60).
FIG. 4.

RTs with inactive polymerase and inactive RNase H complement inefficiently. In control experiments, 293 cells were transfected with plasmids containing vectors encoding wild-type RT or the E478Q or D443A mutant. In parallel, cells were cotransfected with equal proportions of plasmids carrying the D110E mutant and the E478Q mutant or the D110E mutant and the D443A mutant. This generated virions containing only wild-type RT, only the two RNase H mutations, or equal mixtures of RTs defective in polymerase and RNase H. The RT composition of the virions is shown on the x axis, and the relative infectivity is shown on the y axis.
DISCUSSION
Phenotypically mixed HIV-1 vectors that undergo only a single cycle of replication were used to measure the amount of excess polymerase and RNase H activity in virions. When the D11OE mutation was used to reduce the amount of active RT in the virion 50%, infectivity was 42% of the wild-type level. While this still allows a substantial amount of viral replication, it would be expected to cause a reduction in viral fitness, and we would expect viruses whose RTs had approximately half the normal level of polymerase to be selected against during passage relative to the wild-type virus. Replication was severely impaired when less than 25% of the RT in the virions contained active polymerase. When the D443A mutation was used to inactivate RNase H by 90%, the virions retained 35% of their infectivity. The analysis of mixed virions sets the stage for the analysis of the replication of vectors encoding different RT mutants whose biochemical properties have been previously characterized in vitro. The characterization of reverse transcription in a phenotypically mixed virion containing 25% wild-type RT and 75% inactive RT establishes a baseline for the comparison of different RT mutants that contain approximately 25% of a given RT activity (polymerase or RNase H) in in vitro assays.
The ability to monitor reverse transcription accurately by measuring specific DNA intermediates allows us to determine where reverse transcription fails for virions containing limiting amounts of polymerase or RNase H activity. Determining where reverse transcription fails defines the limiting activity (or activities) for a mixed virion or a particular RT mutant.
In the phenotypically mixed virions, a substantial decrease in virus infectivity was observed when the virions contained 25% or less catalytically active polymerase. Since the virions must package enough polymerase activity to ensure that that viral RNA genome is completely reverse transcribed, this result shows that multiple RTs are needed to complete reverse transcription. It has been estimated that virions contain approximately 50 copies of RT; this suggests more than 12 RTs are needed to efficiently complete viral DNA synthesis. RT has a relatively low affinity for its template-primer, and DNA polymerization in vitro results in the synthesis of relatively short segments of DNA. These properties of RT could contribute to the requirement for multiple RTs to convert the RNA into double-stranded DNA. When the polymerase activity of RT mutants was measured in vitro, the lowest amount of polymerase activity for a mutant RT (G190E) found in HIV-1-infected patients was about 20% the wild-type level [using poly(rC)-oligo(dG) as the substrate] (5). This is similar to the amount of active polymerase in the phenotypically mixed virions that were still able to infect cells in this study. The G190E mutant also had reduced processivity compared to wild-type RT (5). The fact that this mutant displayed poor processivity and decreased polymerase activity but can be selected in HIV-1-infected individuals supports the notion that there is excess polymerase activity in the virion and that multiple RTs are involved in reverse transcription.
The dramatic reduction in the amount of −sssDNA synthesized when the amount of active polymerase was systematically reduced in the virions indicates a failure at the level of initiation. Although the virions with limited polymerase activity still contained enough polymerase activity to extend most of the minus-strand DNAs that were initiated, there was little, if any, plus-strand transfer and no 2-LTR circles were found, suggesting a problem with plus-strand DNA synthesis. Taken together, these data suggest that when the amount of active RT is limiting, the critical stages are the initiation of minus-strand DNA synthesis and the synthesis of plus-strand DNA. Processivity, at least in terms of the synthesis of minus-strand DNA, does not appear to be limiting. This is a somewhat surprising result; it suggests that the processivity of minus-strand DNA synthesis may be better in vivo than in vitro. The failure at the plus-strand transfer step implies failure at some step in plus-strand synthesis. Because the initiation of minus-strand DNA synthesis appears to be a limiting step in viral DNA synthesis, it is tempting to speculate that the initiation of plus-strand DNA synthesis is also a limiting step. However, PCR-based technology does not readily distinguish between plus-strand and minus-strand DNA, and we cannot, with the data now available, determine what step in plus-strand DNA synthesis and/or plus-strand transfer is limiting.
The replication of phenotypically mixed virions containing reduced amounts of RNase H activity indicated that there is a greater excess of RNase H activity in virions than there is of polymerase activity. Viruses containing relatively small amounts of the RT with the D443A mutation replicated better than viruses containing the equivalent amount of the RT with the E478Q mutation. Virions that contain RT with the E478Q mutation are defective in the initiation of viral DNA synthesis; it is possible that the effect of the E478Q mutation on polymerization contributes to the replication defect. This excess of RNase H activity in HIV-1 virions does not cause premature degradation of the RNA template or result in premature cleavage of the ppt. We would estimate, based on the data obtained with the D443A mutant, as few as five RTs produce sufficient RNase H cleavage for HIV-1 replication. In support of the estimate that there is approximately a 10-fold excess of RNase H activity in HIV-1 virions, HIV-2 RT has only about 10% the RNase H activity of HIV-1 RT when the specific activities of the two enzymes are compared in vitro (50). The fact that there is a large excess of RNase H activity in HIV-1 virions suggests that the specificity of RNase H cleavage is tightly controlled. Cleavage of the RNA by RNase H must be precise in order to create the proper ends of the linear viral DNA that serves as the substrate for integration into the host genome. We have suggested that this specificity is based not on nucleic acid sequence but on the structure of the nucleic acid in the context of the enzyme (4, 17, 48). The polymerase domain has a substantial role in binding the nucleic acid. The observation that mutations in the polymerase domain of RT affect the specificity of cleavage of the RNA template indicates that the proper positioning of nucleic acid within RT is important for specific RNase H cleavage (4, 17, 42). It would appear that the control of cleavage specificity is sufficient to allow the virion to contain a considerable excess of RNase H activity.
Virions that contained significantly reduced levels of RNase H activity were deficient in minus-strand transfer. This observation is expected based on the requirement for RNase H cleavage for the minus-strand transfer reaction (16, 56, 59). However, we also found that virions containing the E478Q mutation did not efficiently synthesize −sssDNA. The D443N mutation was reported to have minimal effects on polymerase activity when homopolymeric templates were used as substrates (39); however, when heteropolymeric templates were used, this RNase H mutant appeared to interfere with DNA synthesis (12, 40). There are MLV RNase H mutants that have similar properties (2, 20, 57, 60). The E478Q mutation has been reported to interfere with DNA synthesis in vitro; however, initiation was not affected (26). If the RNase H mutations caused a reduction in processivity in vivo, we would have seen greater effects on the synthesis of minus-strand DNA. Severely limiting the RNase H activity appears to have a minimal effect on the late stages of viral DNA synthesis. The ratio of the gag DNA to the plus-strand transfer product and the ratio of the circle junction to either the gag DNA or the plus-strand transfer product are approximately the same in the virions that contain 100% wild-type RT and those that contain 5% wild-type RT, despite the fact that infectivity is reduced. The two steps that appear to be most affected are −sssDNA synthesis and first-strand transfer.
There are two things that are surprising about the results. First, it seems unlikely that the RNase H mutations are affecting general DNA synthesis (processivity); the deficient virions extend minus-strand DNA from U3 through gag reasonably efficiently. The problem seems to be specific for initiation of DNA synthesis, a step that appears to be limiting when the polymerase activity is reduced. This suggests that the initiation of minus-strand DNA synthesis is quite difficult in vivo and is sensitive to alterations in the levels of both polymerase and RNase H activity. Second, it appears that the first-strand transfer reaction is more sensitive to limiting RNase H activity than is the generation or removal of the plus-strand (ppt) primer. The data also suggest that removal of the tRNA primer is not limiting in vivo. Although it might seem at first that the steps of viral DNA synthesis at which RNase H must act with considerable precision (generation and removal of the plus-strand primer and removal of the minus-strand primers) would be the steps that would be most sensitive if the level of RNase H was limiting, this does not appear to be the case. One possible explanation is that the minus-strand transfer step, which is sensitive to limiting the level of RNase H activity, is actually a carefully regulated process. Because the end of the HIV-1 genome is a hairpin (transactivation response region), −sssDNA hairpin formation can be seen in vitro; hairpin formation does not appear to occur in vivo. The available data suggest that RNase H cleavage is carefully controlled. In the presence of nucleocapsid, a specific RNA segment from the 5′ end of the genome blocks hairpin formation and promotes strand transfer (11, 14). It would appear that the requirements are equally specific in vivo, which would explain why strand transfer is sensitive to limiting amounts of RNase H activity.
ACKNOWLEDGMENTS
pNL4-3HSAR−E− from Nathaniel Landau, was obtained through the AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH.
We thank Marilyn Powers for DNA sequencing, Louise Finch for FACS analysis, and Hilda Marusiodis for help in preparing the manuscript. Research in Stephen H. Hughes' laboratory was supported by the National Cancer Institute and by the National Institute for General Medical Sciences.
REFERENCES
- 1.Bartz S R, Vodicka M A. Production of high-titer human immunodeficiency virus type 1 pseudotyped with vesicular stomatitis virus glycoprotein. Methods. 1997;12:337–342. doi: 10.1006/meth.1997.0487. [DOI] [PubMed] [Google Scholar]
- 2.Blain S W, Goff S P. Effects on DNA synthesis and translocation caused by mutations in the RNase H domain of Moloney murine leukemia virus reverse transcriptase. J Virol. 1995;69:4440–4452. doi: 10.1128/jvi.69.7.4440-4452.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Boyer P L, Ferris A L, Hughes S H. Cassette mutagenesis of the reverse transcriptase of human immunodeficiency virus type 1. J Virol. 1992;66:1031–1039. doi: 10.1128/jvi.66.2.1031-1039.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Boyer P L, Ferris A L, Hughes S H. Mutational analysis of the fingers domain of human immunodeficiency virus type 1 reverse transcriptase. J Virol. 1992;66:7533–7537. doi: 10.1128/jvi.66.12.7533-7537.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Boyer P L, Gao H-Q, Hughes S H. A mutation at position 190 of human immunodeficiency virus type 1 reverse transcriptase interacts with mutations at positions 74 and 75 via the template primer. Antimicrob Agents Chemother. 1998;42:447–452. doi: 10.1128/aac.42.2.447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Coffin J M, Hughes S H, Varmus H E, editors. Retroviruses. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1997. [PubMed] [Google Scholar]
- 7.Connor R I, Chen B K, Choe S, Landau N R. Vpr is required for efficient replication of human immunodeficiency virus type-1 in mononuclear phagocytes. Virology. 1995;206:935–944. doi: 10.1006/viro.1995.1016. [DOI] [PubMed] [Google Scholar]
- 8.Davies J F, II, Hostomska Z, Hostomsky Z, Jordan S R, Matthews D A. Crystal structure of the ribonuclease H domain of HIV-1 reverse transcriptase. Science. 1991;252:88–95. doi: 10.1126/science.1707186. [DOI] [PubMed] [Google Scholar]
- 9.DeStefano J J, Buiser R G, Mallaber L M, Myers T W, Bambara R A, Fay P J. Polymerization and RNase H activities of the reverse transcriptases from avian myeloblastosis, human immunodeficiency and Moloney murine leukemia viruses are functionally uncoupled. J Biol Chem. 1991;266:7423–7431. [PubMed] [Google Scholar]
- 10.Di Marzo Veronese F, Copeland T D, DeVico A L, Rahman R, Oroszlan S, Gallo R C, Sarngadharan M G. Characterization of highly immunogenic p66/p55 as the reverse transcriptase of HTL VIII/LAV. Science. 1986;231:1289–1291. doi: 10.1126/science.2418504. [DOI] [PubMed] [Google Scholar]
- 11.Driscoll M D, Golinelli M-P, Hughes S H. In vitro analysis of human immunodeficiency virus type 1 minus-strand strong-stop DNA synthesis and genomic RNA processing. J Virol. 2001;75:672–686. doi: 10.1128/JVI.75.2.672-686.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dudding L R, Nkabinde N C, Mizrahi V. Analysis of the RNA- and DNA-dependent DNA polymerase activities of point mutants of HIV-1 reverse transcriptase lacking ribonuclease H activity. Biochemistry. 1991;30:10498–10506. doi: 10.1021/bi00107a019. [DOI] [PubMed] [Google Scholar]
- 13.Dudding L R, Mizrahi V. Rapid kinetic analysis of a point mutant of HIV-1 reverse transcriptase lacking ribonuclease H activity. Biochemistry. 1993;32:6116–6120. doi: 10.1021/bi00074a025. [DOI] [PubMed] [Google Scholar]
- 14.Fu T B, Taylor J. When retroviral reverse transcriptases reach the ends of their RNA templates. J Virol. 1992;66:4271–4278. doi: 10.1128/jvi.66.7.4271-4278.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Furfine E S, Reardon J E. Human immunodeficiency virus reverse transcriptase ribonuclease H: specificity of tRNA(Lys3)-primer excision. Biochemistry. 1991;30:7041–7046. doi: 10.1021/bi00243a001. [DOI] [PubMed] [Google Scholar]
- 16.Furfine E S, Reardon J E. Reverse-transcriptase-RNaseH from human immunodeficiency virus: relationship of the DNA polymerase and RNA hydrolysis activities. J Biol Chem. 1991;266:406–412. [PubMed] [Google Scholar]
- 17.Gao H-Q, Boyer P L, Arnold E, Hughes S H. Effects of mutations in the polymerase domain on the polymerase, RNase H and strand transfer activities of human immunodeficiency virus type 1 reverse transcriptase. J Mol Biol. 1998;277:559–572. doi: 10.1006/jmbi.1998.1624. [DOI] [PubMed] [Google Scholar]
- 18.Gilboa E, Mitra S W, Goff S, Baltimore D. A detailed model of reverse transcription and tests of crucial aspects. Cell. 1979;18:93–100. doi: 10.1016/0092-8674(79)90357-x. [DOI] [PubMed] [Google Scholar]
- 19.Gopalakrishnan V, Peliska J A, Benkovic S J. Human immunodeficiency virus type 1 reverse transcriptase: spatial and temporal relationship between the polymerase and RNase H activities. Proc Natl Acad Sci USA. 1992;89:10763–10767. doi: 10.1073/pnas.89.22.10763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Guo J, Wu W, Yuan Z Y, Post K, Crouch R J, Levin J G. Defects in primer-template binding, processive DNA synthesis, and RNase H activity associated with the chimeric reverse transcriptases having the murine leukemia virus polymerase domain joined to Escherichia coli RNase H. Biochemistry. 1995;34:5018–5029. doi: 10.1021/bi00015a013. [DOI] [PubMed] [Google Scholar]
- 21.Hayman M J. Viral polyproteins in chick embryo fibroblasts infected with avian sarcoma leukosis viruses. Virology. 1978;85:241–252. doi: 10.1016/0042-6822(78)90428-2. [DOI] [PubMed] [Google Scholar]
- 22.He J, Choe S, Walker R, Di Marzio P, Morgan D O, Landau N R. Human immunodeficiency virus type 1 viral protein R (Vpr) arrests cells in the G2 phase of the cell cycle by inhibiting p34cdc2 activity. J Virol. 1995;69:6705–6711. doi: 10.1128/jvi.69.11.6705-6711.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Heid C A, Stevens J, Livak K J, Williams P M. Real time quantitative PCR. Genome Res. 1996;6:986–994. doi: 10.1101/gr.6.10.986. [DOI] [PubMed] [Google Scholar]
- 24.Hostomsky Z, Homstomska Z, Hudson G O, Moomaw E W, Nodes B R. Reconstitution in vitro of RNase H activity by using purified N-terminal and C-terminal domains of HIV-1 reverse transcriptase. Proc Natl Acad Sci USA. 1991;88:1148–1152. doi: 10.1073/pnas.88.4.1148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hughes S H, Arnold E, Hostomsky Z. RNase H of retroviral reverse transcriptases. 1998. pp. 195–224. . Chapter In R. J. Crouch and J. J. Toulmé (ed.), Ribonucleases H. Les Editions INSERM, Paris, France. [Google Scholar]
- 26.Isel C, Lanchy J-M, Le Grice S F, Ehresmann C, Ehresmann B, Marquet R. Specific initiation and switch to elongation of human immunodeficiency virus type 1 reverse transcriptase require the post-transcriptional modifications of primer tRNA3lys. EMBO J. 1996;15:917–924. [PMC free article] [PubMed] [Google Scholar]
- 27.Jacobo-Molina A, Ding J, Nanni R G, Clark A D, Jr, Lu X, Tantillo C, Williams R L, Kamer G, Ferris A L, Clark P, Hizi A, Hughes S H, Arnold E. Crystal structure of human immunodeficiency virus type 1 reverse transcriptase complexed with double-stranded DNA at 3.0 Å resolution shows bent DNA. Proc Natl Acad Sci USA. 1993;90:6320–6324. doi: 10.1073/pnas.90.13.6320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jamjoon G A, Naso R B, Arlinghaus R B. Further characterization of intracellular precursor polyproteins of Rauscher leukemia virus. Virology. 1977;78:11–34. doi: 10.1016/0042-6822(77)90075-7. [DOI] [PubMed] [Google Scholar]
- 29.Katayanagi K, Miyagawa M, Matsushima M, Ishikawa M, Kanaya S, Ikehara M, Mastsuzaki T, Morikawa K. Three-dimensional structure of ribonuclease H from E. coli. Nature. 1990;347:306–309. doi: 10.1038/347306a0. [DOI] [PubMed] [Google Scholar]
- 30.Keck J L, Marqusee S. Substitution of a highly basic helix/loop sequence into the RNase H domain of human immunodeficiency virus reverse transcriptase restores its Mn2+ dependent RNase H activity. Proc Natl Acad Sci USA. 1995;92:2740–2744. doi: 10.1073/pnas.92.7.2740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kohlstaedt L A, Wang J, Friedman J M, Rice P A, Steitz T A. Crystal structure at 3.5 Å resolution of HIV-1 reverse transcriptase complexed with an inhibitor. Science. 1992;256:1783–1790. doi: 10.1126/science.1377403. [DOI] [PubMed] [Google Scholar]
- 32.Kotewicz M L, Samson C M, D' Alessio J M, Gerard G F. Isolation of cloned Moloney murine leukemia virus reverse transcriptase lacking ribonuclease H activity. Nucleic Acids Res. 1988;16:265–277. doi: 10.1093/nar/16.1.265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Le Grice S F J, Naas T, Wohlgensinger B, Schatz O. Subunit selective mutagenesis indicates minimal polymerase activity in heterodimer-associated p51 HIV-1 reverse transcriptase. EMBO J. 1991;10:3905–3911. doi: 10.1002/j.1460-2075.1991.tb04960.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Levin J G, Crouch R J, Post K, Hu S C, McKelvin D, Zweig M, Court D L, Gerwin B I. Functional organization of the murine leukemia virus reverse transcriptase: characterization of a bacterially expressed AKR DNA polymerase deficient in RNase H activity. J Virol. 1988;62:4376–4380. doi: 10.1128/jvi.62.11.4376-4380.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lie Y S, Petropoulos C J. Advances in quantitative PCR technology; 5′ nuclease assays. Curr Opin Biotechnol. 1998;9:43–48. doi: 10.1016/s0958-1669(98)80082-7. [DOI] [PubMed] [Google Scholar]
- 36.Livak K J, Flood S J A, Marmaro J, Giusti W, Deetz K. Oligonucleotides with fluorescent dyes at opposite ends provide a quenched probe system useful for detecting PCR product and nucleic acid hybridization. PCR Methods Appl. 1995;4:357–362. doi: 10.1101/gr.4.6.357. [DOI] [PubMed] [Google Scholar]
- 37.Luo G X, Taylor J. Template switching by reverse transcriptase during DNA synthesis. J Virol. 1990;64:4321–4328. doi: 10.1128/jvi.64.9.4321-4328.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mitra S W, Goff S, Gilboa E, Baltimore D. Synthesis of a 600-nucleotide-long plus-strand DNA by virions Moloney murine leukemia virus. Proc Natl Acad Sci USA. 1979;76:4355–4359. doi: 10.1073/pnas.76.9.4355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mizrahi V, Usdin M T, Harington A, Dudding L R. Site-directed mutagenesis of the conserved Asp-443 and Asp-498 carboxy-terminal residues of HIV-1 reverse transcriptase. Nucleic Acids Res. 1990;18:5359–5363. doi: 10.1093/nar/18.18.5359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mizrahi V, Brooksbanks R L, Nkabinde N C. Mutagenesis of the conserved aspartic acid 443, glutamic acid 478, aspargine 494, and aspartic acid 498 residues in the ribonuclease H domain of p66/p51 human immunodeficiency virus type I reverse transcriptase. J Biol Chem. 1994;269:19245–19249. [PubMed] [Google Scholar]
- 41.Oppermann H, Bishop J M, Varmus H E, Levintow L. A joint product of the genes gag and pol of avian sarcoma virus: a possible precursor of reverse transcriptase. Cell. 1977;12:993–1005. doi: 10.1016/0092-8674(77)90164-7. [DOI] [PubMed] [Google Scholar]
- 42.Palaniappan C, Wisniewski M, Jacques P S, Le Grice S F, Fay P J, Bambara R A. Mutations within the primer grip region of reverse transcriptase result in loss of RNase H function. J Biol Chem. 1997;272:11157–11164. doi: 10.1074/jbc.272.17.11157. [DOI] [PubMed] [Google Scholar]
- 43.Post K, Guo J, Kalman E, Uchida T, Crouch R J, Levin J G. A large deletion in the connection subdomain of murine leukemia virus reverse transcriptase or replacement of the RNase H domain with Escherichia coli RNase H results in altered polymerase and RNase H activities. Biochemistry. 1993;32:5508–5517. doi: 10.1021/bi00072a004. [DOI] [PubMed] [Google Scholar]
- 44.Pullen K A, Champoux J J. Plus-strand origin for human immunodeficiency virus type 1: implications for integration. J Virol. 1990;64:6274–6277. doi: 10.1128/jvi.64.12.6274-6277.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Pullen K A, Ishimoto L K, Champoux J J. Incomplete removal of the RNA primer for minus-strand DNA synthesis by human immunodeficiency virus type 1 reverse transcriptase. J Virol. 1992;66:367–373. doi: 10.1128/jvi.66.1.367-373.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rattray A J, Champoux J J. Plus-strand priming by Moloney murine leukemia virus. The sequence features important for cleavage by RNase H. J Mol Biol. 1989;208:445–456. doi: 10.1016/0022-2836(89)90508-1. [DOI] [PubMed] [Google Scholar]
- 47.Repaske R, Hartley J W, Kavlick M F, O'Neill R R, Austin J B. Inhibition of RNase H activity and viral replication by single mutations in the 3′ region of Moloney murine leukemia virus reverse transcriptase. J Virol. 1989;63:1460–1464. doi: 10.1128/jvi.63.3.1460-1464.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sarafianos S G, Das K, Tantillo C, Clark A D, Jr, Ding J, Whitcomb J, Gait M, Boyer P L, Hughes S H, Arnold E. Crystal structure of HIV-1 reverse transcriptase in complex with a polypurine tract RNA:DNA. EMBO J. 2001;20:1449–1461. doi: 10.1093/emboj/20.6.1449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Schatz O, Cromme F V, Naas T, Lindemann D, Mous J, Le Grice S F J. Inactivation of the RNase H domain of HIV-1 reverse transcriptase blocks viral infectivity. In: Papas T S, editor. Gene regulation and AIDS. Houston, Tex: Portfolio; 1990. pp. 293–404. [Google Scholar]
- 50.Shaharabany M, Hizi A. The catalytic functions of chimeric reverse transcriptases of human immunodeficiency viruses type 1 and type 2. J Biol Chem. 1992;267:3674–3678. [PubMed] [Google Scholar]
- 51.Smith J K, Cywinski A, Taylor J M. Specificity of initiation of plus-strand DNA by Rous sarcoma virus. J Virol. 1984;52:314–319. doi: 10.1128/jvi.52.2.314-319.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
-
52.Smith J S, Roth M. Specificity of human immunodeficiency virus-1 reverse transcriptase-associated ribonuclease H in removal of the minus-strand primer, tRNA
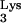 J Biol Chem. 1992;267:15071–15079. [PubMed] [Google Scholar]
J Biol Chem. 1992;267:15071–15079. [PubMed] [Google Scholar] - 53.Smith J S, Gritsman K, Roth M J. Contributions of DNA polymerase subdomains to the RNase H activity of human immunodeficiency virus type 1 reverse transcriptase. J Virol. 1994;68:5721–5729. doi: 10.1128/jvi.68.9.5721-5729.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Stahl S J, Kaufman J D, Vikic-Topic S, Crouch R J, Wingfield P T. Construction of an enzymatically active ribonuclease H domain of human immunodeficiency virus type 1 reverse transcriptase. Protein Eng. 1994;7:1103–1108. doi: 10.1093/protein/7.9.1103. [DOI] [PubMed] [Google Scholar]
- 55.Stromberg K, Hurley N E, Davis N L, Rueckert R R, Fleisnner E. Structural studies of avian myeloblastosis virus: Comparison of polypeptides in virion and core component by dodecyl sulfate-polyacrylamide gel electrophoresis. J Virol. 1974;13:513–528. doi: 10.1128/jvi.13.2.513-528.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Tanese N T, Telesntitsky A, Goff S P. Abortive reverse transcription by mutants of Moloney murine leukemia virus deficient in the reverse transcriptase-associated RNase H function. J Virol. 1991;65:4387–4397. doi: 10.1128/jvi.65.8.4387-4397.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Tanese N, Goff S P. Domain structure of the Moloney murine leukemia virus reverse transcriptase: mutational analysis and separate expression of the polymerase and RNase H activities. Proc Natl Acad Sci USA. 1988;85:1777–1781. doi: 10.1073/pnas.85.6.1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Taylor J M, Hsu T W. Reverse transcription of avian sarcoma virus RNA into DNA might involve copying of the tRNA primer. J Virol. 1980;33:531–534. doi: 10.1128/jvi.33.1.531-534.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Telesnitsky A, Blain S W, Goff S P. Defects in Moloney murine leukemia virus replication caused by a reverse transcriptase mutation modeled on the structure of Escherichia coli ribonuclease H. J Virol. 1992;66:615–622. doi: 10.1128/jvi.66.2.615-622.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Telesnitsky A, Goff S P. Two defective forms of reverse transcriptase can complement to restore retroviral infectivity. EMBO J. 1993;12:4433–4438. doi: 10.1002/j.1460-2075.1993.tb06128.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Tisdale M, Schultze T, Larder B A, Moelling K. Mutations within the RNase H domain of HIV-1 reverse transcriptase abolish viral infectivity. J Gen Virol. 1991;72:59–66. doi: 10.1099/0022-1317-72-1-59. [DOI] [PubMed] [Google Scholar]
- 62.Varmus H E, Heasley S, Kung H-J, Opperman H, Smith V C, Bishop J M, Shank P R. Kinetics, of synthesis, structure, and purification of avian sarcoma virus-specific DNA made in the cytoplasm of acutely infected cells. J Mol Biol. 1978;120:55–82. doi: 10.1016/0022-2836(78)90295-4. [DOI] [PubMed] [Google Scholar]
- 63.Volkmann S, Wöhrl B M, Tisdale M, Moelling K. Enzymatic analysis of two HIV-1 reverse transcriptase mutants with mutations in the carboxyl-terminal amino acid residues conserved among retroviral ribonucleases H. J Biol Chem. 1993;268:2674–2683. [PubMed] [Google Scholar]
- 64.Wegner R H, Rochelle J M, Seldin M F, Kohler G, Nielsen P J. The heat stable antigen (mouse CD24) gene is differentially regulated but has a housekeeping promoter. J Biol Chem. 1993;268:23345–23352. [PubMed] [Google Scholar]
- 65.Whitcomb J M, Kumar R, Hughes S H. Sequence of the circle junction of human immunodeficiency virus type 1: implications for reverse transcription and integration. J Virol. 1990;64:4903–4906. doi: 10.1128/jvi.64.10.4903-4906.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Whitcomb J M, Hughes S H. Retroviral reverse transcription and integration: progress and problems. Annu Rev Cell Biol. 1992;8:275–306. doi: 10.1146/annurev.cb.08.110192.001423. [DOI] [PubMed] [Google Scholar]
- 67.Wohrl B M, Moelling K. Interaction of HIV-1 ribonuclease H with polypurine tract containing RNA-DNA hybrids. Biochemistry. 1990;29:10141–10147. doi: 10.1021/bi00496a001. [DOI] [PubMed] [Google Scholar]
- 68.Yang W, Hendrickson W A, Crouch R J, Satow Y. Structure of ribonuclease H phased at 2 Å resolution by MAD analysis of the selenomethionyl protein. Science. 1990;249:1398–1405. doi: 10.1126/science.2169648. [DOI] [PubMed] [Google Scholar]
- 69.Yee J K, Friedmann T, Burns J C. Generation of high-titer pseudotyped retroviral vectors with very broad host range. Methods Cell Biol. 1994;43:99–112. doi: 10.1016/s0091-679x(08)60600-7. [DOI] [PubMed] [Google Scholar]