

ABSTRACT
Grasslands represent a major biome on Earth and play a vital role in ecosystem functioning and dynamics. However, owing to the variations among grassland types, the impact of grazing on plant community diversity and stability remains unclear. This study is based on the typical steppe of the Mongolian Plateau. Field sampling and data analysis were combined to qualitatively and quantitatively investigate the structural characteristics, species diversity, and stability of plant communities under varying grazing intensities, that is, four‐season nomadic, two‐season rotational, and sedentary grazing (FSNG, TSRG, and SG, respectively). The results indicated that FSNG pastures exhibited the largest number of plant species while FSNG and TSRG pastures exhibited relatively high importance values for the primary dominant species. Carex duriuscula, Chenopodium glaucum, and Cleistogenes squarrosa were prominent in SG pastures, with C. duriuscula having the largest importance value. The mean height, cover, and aboveground biomass of plant communities in FSNG were significantly higher than those in SG (p < 0.05), with no significant difference observed between FSNG and TSRG. FSNG also demonstrated the highest Shannon–Wiener, Simpson, and Pielou indexes. The Shannon–Wiener and Simpson indexes between the FSNG, TSRG, and SG pastures showed significant differences (p < 0.05). Nomadic plant communities displayed positive loosely interspecific traits, suggesting independence and positive succession. Conversely, communities in TSRG and SG exhibited negative correlations and higher instability. The stability analysis ranked community stability as FSNG > TSRG > SG, suggesting that judicious grazing practices could enhance grassland stability. The findings reveal that grazing patterns influence plant community composition and function and that FSNG pastures promote higher species diversity, perennial dominance, and overall stability compared with TSRG and SG pastures.
Keywords: community stability, community structure, different grazing patterns, species diversity, typical steppe of the Mongolian plateau
This paper focuses on the Wuzhumuqin–Wendurhan Sample Strip, a typical grassland on the Mongolian Plateau, and explores the structural characteristics, species diversity, and stability of plant communities in response to different grazing practices, which is relevant to the scope of the journal's theme.

1. Introduction
Plant community diversity is a vital aspect of biodiversity and plays a key role in supplying material and energy to ecosystems, maintaining ecosystem stability, and regulating global climate (Zhao et al. 2023). Community stability encompasses the system's capacity to withstand external disturbances and includes factors such as resilience, resistance, and persistence (Yao et al. 2016; Ishii et al. 2021). Grazing exerts a widespread influence on the Mongolian Plateau, impacting not only species composition, community diversity, and ecosystem stability but also material cycles and energy flows. Consequently, it stands as a pressing concern within the international ecological community (Briske et al. 2008; Conte and Tilt 2014; Zhang et al. 2022). In recent years, there has been a surge in research focusing on precise grazing management systems. Controlled experiments have scrutinized nomadic, rotational, and continuous grazing to ascertain their respective advantages and disadvantages (Jacobo et al. 2006; Na, Bao, et al. 2018). Despite these efforts, the effects of varied grazing practices on community diversity and stability within typical Mongolian Plateau grasslands remain inadequately elucidated (De Boeck et al. 2018; Valerio et al. 2022).
The Mongolian Plateau is located in Central Asia and has a typical continental arid and semiarid climate that is sensitive to climate change and exhibits fragile ecosystems (Zhang et al. 2009). Typical grasslands of the Mongolian Plateau are present in Sukhbaatar, Kent, and Dongfang Provinces, as well as in the eastern part of the Xilin Gol League and western part of Hulunbeier City in Inner Mongolia, and represent the core components of the temperate grasslands (Wulan 2021). The Inner Mongolian Steppe represents the southern part of this steppe belt, serving as one of China's major animal husbandry bases and an important ecological barrier in northern China (Wulan 2021). However, owing to the wide distribution area of grasslands and the intensity of their utilization, the issues of grassland desertification and vegetation degradation are becoming increasingly serious (Li and Qu 2002; Liu et al. 2009; Zhang et al. 2018). Such changes are threatening the ecological security, social stability, and economic development of China's northern border (Bao, Liu, et al. 2013; Zhang et al. 2018).
Grazing is the primary method of grassland utilization on the Mongolian Plateau, constituting a significant ecological disturbance. This disturbance can profoundly affect entire ecosystems by modifying soil physicochemical properties and increasing habitat heterogeneity, thereby influencing the distribution of species as well as the structure and diversity patterns of grassland communities. Notably, the pasture contracting system introduced in Inner Mongolia during the early 1980s has led to a gradual transition in grassland utilization from nomadic pastoralism to sedentary pastoralism (Na, Li, et al. 2018). This shift encompasses diverse approaches, such as settled grazing, restricted grazing, fallow pastoralism, and rotational pastoralism, thereby deviating from the traditional nomadic pastoralism practiced in Mongolia. This change exposed the grassland zone to different pressures and influencing factors, which decreased grassland productivity, escalated grass–animal conflict, and reduced vegetation diversity and stability (Liu et al. 2009; Na, Li, et al. 2018), posing a serious threat to the sustainable development of grassland animal husbandry. Therefore, in recent years, Mongolian Plateau vegetation has become a focus of many researchers, both at home and abroad, who have studied the impact of grazing on plant community diversity and stability. Studies have suggested that short‐term grazing exclusion, rotational grazing, deferred grazing, and moderate disturbance levels are beneficial for maintaining species diversity (Zhao et al. 2023). However, other research indicates that long‐term grazing exclusion can lead to an excessive increase in the dominance of one or a few species, thereby reducing grassland species richness and diversity (Wang et al. 2019). Additionally, continuous grazing degrades grassland habitats, thus causing significant shifts in the dominance of key functional plant groups and reducing community species diversity and structural complexity. Grazing disturbance is one of the factors that influences grassland ecosystem stability, and it has a direct impact on community stability (Zhao et al. 2023). Studies have shown that as grazing intensity increases, community functional diversity and stability significantly decrease (Shen, Wang, and Han 2022). Under varying degrees of grazing disturbance, the stability of desert grassland ecosystems differs, with excessive grazing pressure negatively affecting grassland productivity and ecological functions. However, due to limitations imposed by international borders, communication barriers, and adverse weather conditions, research is often confined to small areas within a single country (Yin et al. 2013). Changes in typical steppe ecosystems of the Mongolian Plateau are primarily studied using remote sensing images to extract the normalized difference vegetation index (NDVI) of grassland vegetation and analyze spatial and temporal variations (Bao, Bao, et al. 2013; Batunacun et al. 2015; Tao et al. 2015). However, comprehensive research on the effects of different grazing practices on species diversity and ecosystem stability across the transboundary typical steppe of China and Mongolia is still lacking.
Plant community structure, species diversity, and stability constitute fundamental components of community ecology, thereby serving as crucial indicators for assessing and managing degraded grassland resources (Miao et al. 2014). Therefore, studying the community structure and stability is important for understanding and protecting grassland ecosystems, and further investigations are warranted on the effects of grazing methods on plant community species diversity and stability in typical grasslands on the Mongolian Plateau.
This study aimed to determine whether unwarranted grazing practices (1) degrade the structural characteristics of the plant community in the direction of succession, (2) reduce the species diversity of plant communities, or (3) decrease plant community stability. Determining the most sensible grazing practices will help achieve the sustainable use of typical grasslands on the Mongolian Plateau. Furthermore, our findings will provide a reference for the restoration and stabilization of grassland ecosystems.
2. Materials and Methods
2.1. Sample Zone Delineation
The study describes the setup of plant sample plots and the recording of geographic information at the sampling site (Figure 1). The vegetation type within the sample plot is herbaceous, with specific geographic coordinates and heights noted on a whiteboard. Based on the unified context of a typical steppe–chestnut–calcium soil operating area, the delineation of sample zones prioritized the homogeneity of subsurface natural geographic conditions, particularly focusing on selecting high plains with flat topography. Additionally, the sample zones were strategically positioned to span across territories of different countries, ensuring a comprehensive representation of the changing characteristics of the same grassland under varied country borders, utilization modes, and intensities (Wulan 2021). Specifically, the sample zone encompasses the core area of the typical steppe on the Mongolian Plateau, stretching from the city of Wendurhan, Kent Province, in northwestern Mongolia, to Bayanhua Town, Xiwuzhumuqin Banner, in Inner Mongolia, China. The sample encompasses four control points: (111°8′51.25″ E, 47°56′20.13″ N) (northwest), (119°13′25.31″ E, 45°13′9.35″ N) (northeast), (118°14′32.64″ E, 44°32′30.80″ N) (southeast), (110°13′34.08″ E, 47°13′58.02″ N) (southwest). These points span diagonally across the Kent, Sukhbaatar, and Dongfang Provinces of Mongolia and the Xilinguole League of Inner Mongolia, with a total area of 69,850,000 km2 (Figure 2). Within the sample zone, 66.73% of the area is in Mongolia, and 33.27% is in Inner Mongolia (Wulan 2021).
FIGURE 1.

Setup of plant sample plots at the sampling site.
FIGURE 2.

Study area location. FSNG, four‐season nomadic grazing; SG, settled grazing; TSRG, two‐season rotational grazing.
2.2. Experimental Design and Field Sampling
While the sample plots in this study are situated in various regions of China and Mongolia, they share similar topographies, soil compositions, plant diversity, grazing levels, and livestock demographics. To mitigate the impact of environmental variables, this paper performed a significance analysis on the ecological factors of the study area samples, as shown in Table 1. Interviews with herders and field surveys revealed that three types of grazing practices are common in the study area: four‐season nomadic grazing (FSNG), two‐season rotational grazing (TSRG), and settled grazing (SG). In FSNG areas, herders migrate according to seasonal changes and typically utilize four pastures for spring, summer, autumn, and winter. In TSRG areas, herders alternate pastures between the spring and autumn seasons or between the summer and winter seasons. In SG areas, herders grazing in a fixed location, with the pasture position remaining largely unchanged.
TABLE 1.
Basic information of the sample plots.
| Sample plot | Altitude (m) | Average stocking rate (sheep unit/km2) | Annual mean temperature (°C) | Annual total precipitation (mm) | Soil type | Sample size | Main dominant plants |
|---|---|---|---|---|---|---|---|
| FSNG | 1020a | 0.14a | 1.92a | 244.41a | Castanoems | 8 | Leymus chinensis, Stipa grandis, Cleistogenes squarrosa |
| TSRG | 939.5a | 0.16a | 1.90a | 234a | Castanoems | 8 | C. squarrosa, L. chinensis, Chenopodium glaucum |
| SG | 962.2a | 0.15a | 1.90a | 234a | Castanoems | 8 | Carex duriuscula, C. squarrosa, C. glaucum |
Note: Lowercase letters indicate significant differences at the 5% level. Source: Meteorological Bureau of Xilingol League and National Agency for Meteorology and Environmental Monitoring of Mongolia. Stocking rate—total number of livestock (sheep) owned by herders in the sample plot/total area of grassland owned by herders. All types of livestock were converted to sheep units according to China's sheep unit conversion standard as follows: one camel—seven sheep, one horse—six sheep, one cow—five sheep, and one goat—one sheep.
During July and August of 2016 and 2017, we surveyed plant communities in the study area, specifically in the herdsmen's pastures, and established a total of 24 sampling points, with eight points randomly selected within each of the three grazing types. At each sampling point, which covered a 1 m2 plot. In sedentary and two‐season rotational grazing systems, the distances between adjacent sampling points were generally consistent, typically within 200 m. In contrast, the distance between adjacent sampling points was increased to effectively capture the spatial variability present within these pastures owing to the high heterogeneity of the four‐season nomadic pastures. In these plots, we assessed various parameters, including plant height, density, cover, aboveground biomass, and species composition. Plant height was measured from the base to the canopy using a tape, with average heights calculated from 3 to 5 specimens per species. Plant density was determined by counting the total number of plants within the sample area. To estimate vegetation cover, we captured photographs of the samples from a perpendicular angle and used the Photoshop 6 software to calculate the pixels occupied by plants based on the color contrast between plants and soil. The ratio of plant pixels to the total pixels of the sample provided the cover value. Aboveground biomass was obtained using a harvesting method for each sample plot. The fresh weight of each plant by species was measured, and samples were transported to the laboratory. The plants were subsequently dried to a constant weight at 65°C, and their dry weight was measured (Jian et al. 2018).
2.3. Data Processing and Methods
2.3.1. Community Species Importance Values and Plant Diversity
In this study, species occurring in the plant community were categorized into four functional groups according to differences in life type: perennial grasses, perennial forbs, annual and biennial plants, shrubs, and semishrubs (Jian et al. 2018; Sasaki et al. 2019; Zhang et al. 2020; Suriguga et al. 2022). Diversity indexes are typically used to represent plant community diversity (Sun, Zhu, and Zhang 2014). Thus, the Shannon–Wiener, Margalef, Pielou, and Simpson indexes and the dominance of species in a plant community were determined based on the importance value (pi) (Jian et al. 2018).
The associated formulae are provided below.
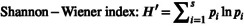 |
(1) |
| (2) |
| (3) |
| (4) |
where S denotes the number of species in the plant community, N denotes the sum of the number of individuals of all species, and p i denotes the species importance value.
Next, we measured the β‐diversity indexes of plant communities among different grazing patterns in typical grasslands in China and Mongolia. A simpler measure of plant community similarity coefficients was used, that is, the Jaccard coefficient (Jian et al. 2018).
| (5) |
where a and b are the number of species in the two plant communities in different habitats, and c is the number of species common to the two plant communities. Additionally, β ranges from (0, 1), where β values in the range of (0, 0.25) indicate that the two communities are extremely dissimilar; (0.25, 0.50), the two communities are moderately dissimilar; (0.5, 0.75), the two communities are moderately similar; and (0.75, 1.00), the two communities are extremely similar (Bátori et al. 2012).
2.3.2. Overall Community Correlation Analysis
The correlation among plant species illustrates their adaptation to various grazing patterns in China and Mongolia's typical grasslands, effectively depicting plant community structures. In this study, the variance ratio method derived from the null‐linkage model (Roxburgh, Shea, and Wilson 2004) was used to quantitatively verify the overall correlation of typical steppe plant communities in China and Mongolia.
| (6) |
| (7) |
where S is the total number of species, p i = n i /N, n i is the number of samples where species i occurs, and N is the total number of samples; T j is the total number of species occurring in sample plot j; t is the average number of species within the sample; VR is the overall association index between species within the community; VR > 1 indicates that the species shows an overall positive association, VR = 1 indicates no association between all species, and VR < 1 indicates an overall negative association between species; and W is used to test the significance of the VR offset 1, W = VR × N. If the interspecific association is not significant, there is a 90% probability that W falls into χ 20.05(N) < W < χ 20.95(N) (Jian et al. 2018).
2.3.3. Community Stability Analysis
We employed the Godron contribution law method to rank plant species surveyed under different grazing practices by frequency and convert them to relative frequency for stepwise accumulation. This involved calculating the cumulative sum of the plant species in their sorted order and expressing it as percentages. Subsequently, we generated a fuzzy scatter smoothing curve, plotting the cumulative inverse percentage of plant species on the x‐axis against the cumulative relative frequency on the y‐axis. This visualization method effectively illustrates the relationship between grazing practices and the distribution of plant species across the study area. Finally, coordinates of the intersection point with the x + y = 100 line were found, which represent the stabilization point of the desired plant community. The closer the cumulative inverse percentage of plant species and the cumulative relative frequency percentage of the plant community is to 20/80, the more stable the plant community will be (Huang et al. 2015; Linders et al. 2019).
| (8) |
| (9) |
The coordinates of the intersection point (x, y) were derived from Equation (9) and compared with 20/80 to determine the degree of stabilization of the plant community for different grazing practices.
Excel (2007) and SPSS (22.0) were used for data processing and statistical analyses, respectively. The t‐test was used for identifying significant differences between the two variables (α = 0.05), and Origin 9.1 software was used for plotting.
3. Results
3.1. Changes in Plant Community Characteristics Under Different Grazing Methods
3.1.1. Plant Community Composition and Structure
Grassland plant community composition determines the community structure, appearance, function, and succession direction and thus is one of the most intuitive characteristics of vegetation (Gao et al. 2019; Chang et al. 2020). The choice of grazing method influences the composition of plant communities, as evidenced by the total numbers of species observed in the FSNG, TSRG, and SG areas, at 37, 27, and 23, respectively (Table 2). In terms of species importance value, dominant species differed across the grazing areas. In the FSNG area, Leymus chinensis, Stipa grandis, and Carex duriuscula held the highest importance value at 49.16%. Conversely, the TSRG area was characterized by brown Cleistogenes squarrosa, L. chinensis, and Chenopodium glaucum, with an importance value of 35.19%. Finally, the SG area was dominated by C. duriuscula, C. squarrosa, and C. glaucum, with an importance value of 47.73%. The results reveal differences among the dominant species of typical steppe of FSNG, TSRG, and SG. The dominant species in FSNG pastures remained relatively stable, whereas, in TSRG, L. chinensis and C. squarrosa dominated, with a shift in the relative importance of the species. However, the presence of degradation indicator plants such as C. duriuscula and C. glaucum in the SG area indicated severe degradation.
TABLE 2.
Composition and importance values of typical steppe communities in China and Mongolia.
| Functional group | Species | Importance value % | ||
|---|---|---|---|---|
| FSNG | TSRG | SG | ||
| Perennial grasses | Leymus chinensis | 19.97 | 11.28 | 10.66 |
| Stipa grandis | 13.85 | 3.45 | 8.63 | |
| Cleistogenes squarrosa | 11.59 | 14.22 | 15.43 | |
| Stipa breviflora | 2.34 | — | — | |
| Stipa mongolorum Tzvel. | 4.64 | — | — | |
| Agropyron cristatum | 1.45 | 1.09 | 1.93 | |
| Achnatherum splendens | 1.71 | — | — | |
| Stipa krylovii | — | 9.58 | 9.53 | |
| Perennial forbs | Stellera chamaejasme Linn | — | 3.58 | — |
| Allium tenuissimum | 1.19 | 2.03 | 2.29 | |
| Allium ramosum | — | 1.45 | — | |
| Iris tenuifolia Pall. | — | — | 1.00 | |
| Carex duriuscula | 15.34 | 9.36 | 18.07 | |
| Allium polyrhizum | 3.45 | 6.91 | 1.33 | |
| Saussurea mongolica | 1.08 | — | — | |
| Asparagus dauricus Link | 0.33 | — | 1.21 | |
| Convolvulus ammannii | 1.12 | 1.69 | 1.62 | |
| Serratula komarovii | 1.22 | 1.96 | 1.43 | |
| Anemarrhena asphodeloides Bunge | — | 3.91 | — | |
| Allium ramosum | — | — | 2.11 | |
| Bromus inermis Leyss. | — | — | — | |
| Thalictrum petaloideum L. | 1.18 | — | — | |
| Potentilla acaulis | — | — | 1.1 | |
| Astragalus melilotoides Pall | — | 1.05 | 1.83 | |
| Allium mongolicum Regel | 1.06 | — | — | |
| Gymnocarpos przewalskii | 3.53 | — | — | |
| Shrubs and semi‐shrubs | Caragana microphylla Lam | 3.76 | — | — |
| Artemisia frigida | 1.64 | 4.05 | 2.96 | |
| Artemisia halodendron | — | 2.88 | 1.83 | |
| Artemisia adamsii Bess. | — | 3.38 | — | |
| Ptilotricum canescens | — | 2.17 | 1.35 | |
| Annual and biennial plants | Artemisia annua | 5.67 | — | — |
| Chenopodium glaucum | — | 9.69 | 14.23 | |
| Artemisia palustris Linn. | — | — | — | |
| Salsola collina | — | 3.38 | 1.32 | |
| Lappula myosotis Moench | — | 0.59 | 2.17 | |
| Atriplex tatarica (L.) Bung | 1.19 | — | 1.20 | |
Note: “—” indicates a plant community where the species does not occur or has an importance value of 1.
Abbreviations: FSNG, four‐season nomadic grazing; SG, settled grazing; TSRG, two‐season rotational grazing.
In terms of functional groups of species, FSNG, TSRG, and SG pastures were dominated by perennial grasses and perennial forbs (Figure 3). Among them, the average percentage of perennial grasses in FSNG, TSRG, and SG areas were 54.7%, 44.68%, and 38.43%, respectively.
FIGURE 3.

Quantitative characteristics of plant communities in each sample area.
The mean perennial forbs ratio was 31.84%, 30.80%, and 37.42%, respectively. Shrub and semishrub plants showed importance values of 4.99%, 10.24%, and 10.30%, respectively. The mean values of the proportion of annual and biennial plants were 8.48%, 14.27%, and 23.23%. Furthermore, the order of taxa proportion of the functional groups of grassland communities with different grazing modes was perennial grasses > perennial forbs > annual and biennial plant > shrub and semishrub. However, there was a difference in the mean value of the percentage. Moreover, the structure of sedentary grazing plant communities progressively degraded.
3.1.2. Quantitative Characteristics of the Plant Communities
Community characterization is the basis for ecological restoration and reestablishment and the integration of vegetation adaptations to ecosystems (Chang et al. 2020). Different grazing methods affect the quantitative characteristics of plant communities (Cumming et al. 2016). The average heights of the FSNG, TSRG, and SG plant communities were 25.30, 10.80, and 10.16 cm, respectively, with a significant difference between FSNG, TSRG, and SG (p < 0.05) (Figure 4). This result indicates that the heights of FSNG were higher than that of the other grazing modes and that the heights of the communities were significantly lower, especially in the case of SG conditions (p < 0.05). The plant community densities of FSNG, TSRG, and SG were recorded as 228, 317.36, and 303 plants/m2, respectively, with no significant difference observed among the three grazing methods (Figure 4). Despite this, TSRG demonstrated the highest density, suggesting its density surpassed that of the other grazing methods. Additionally, the plant community cover for FSNG, TSRG, and SG was reported as 79.37%, 35.62%, and 28.11%, respectively, with significant differences noted between the FSNG community and TSRG and SG communities, while no significant differences were observed between TSRG and SG communities (p < 0.05). This outcome implies that the community cover of the FSNG plot exceeded that of the TSRG and SG plots. The aboveground biomass of plant communities in Mongolia was 262.87, 170.58, and 131.74 for FSNG, TSRG, and SG, respectively, and with a significant difference between FSNG, TSRG, and SG (p < 0.05) (Figure 3). This result indicates that the aboveground biomass of the FSNG pasture was higher than that of the TSRG and SG Mongolian pastures.
FIGURE 4.
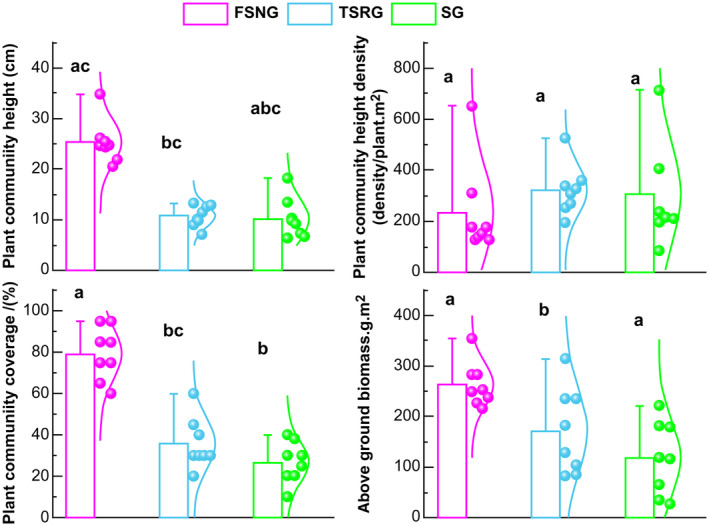
Comparison of the quantitative characteristics of typical steppe communities in China and Mongolia. The small cubes in each box represent mean values. Different lowercase letters indicate significant differences between grazing practices (p < 0.05). FSNG, four‐season nomadic grazing; SG, settled grazing; TSRG, two‐season rotational grazing.
3.2. Characterization of Plant Community Diversity
The Shannon–Wiener, Margalef, Pielou, and Simpson community diversity indices were used to characterize the changes in community diversity (Bao et al. 2009; Liu et al. 2022). The Shannon–Wiener, Simpson, and Pielou indexes were the highest for TSRG at 2.14, 0.90, and 0.85, respectively, and the Margalef index was the highest for FSNG at 1.72 (Table 3). The Margalef index was not significantly different across grazing practices; the Shannon–Wiener, Simpson, and Pielou indexes were significantly higher (p < 0.05) for FSNG than for TSRG and SG.
TABLE 3.
Species diversity in typical steppe communities in China and Mongolia.
| Test sample | Shannon–Wiener index | Simpson index | Pielou index | Margalef index |
|---|---|---|---|---|
| FSNG | 2.14 ± 0.06a | 0.90 ± 0.11a | 0.85 ± 0.11a | 1.06 ± 0.21a |
| TSRG | 1.95 ± 0.09a | 0.83 ± 0.03b | 0.83 ± 0.14a | 1.72 ± 0.12a |
| SG | 1.70 ± 0.08b | 0.87 ± 0.21ab | 0.78 ± 0.02ab | 1.13 ± 0.10a |
Note: Lowercase letters indicate significant differences between grazing practices (p < 0.05).
Abbreviations: FSNG, four‐season nomadic grazing; SG, settled grazing; TSRG, two‐season rotational grazing.
Jaccard's similarity coefficient was used as a measure of the similarity of typical steppe communities with different grazing practices, and the similarity coefficients were analyzed (Jian et al. 2018). The Jaccard coefficients of FSNG and TSRG pastures were 0.25, indicating moderate dissimilarity; those of FSNG and sedentary grazing were 0.23 (ranging from 0.01 to 0.25), indicating that the communities were at a very dissimilar level, whereas those of TSRG and SG were 0.61 (ranging from 0.5 to 0.75), indicating that the communities were at a moderately similar level (Table 4). Therefore, large habitat variability was observed between the plant communities under different grazing patterns in typical grasslands in China and Mongolia. This result suggests that different grazing practices affected population dispersal and settlement, which in turn changed the composition of plant communities and formed different community structures.
TABLE 4.
Similarity coefficients of typical steppe communities in China and Mongolia.
| Test sample | FSNG | TSRG | SG |
|---|---|---|---|
| FSNG | 1 | 0.25 | 0.23 |
| TSRG | 0.25 | 1 | 0.61 |
| SG | 0.23 | 0.61 | 1 |
Abbreviations: FSNG, four‐season nomadic grazing; SG, settled grazing; TSRG, two‐season rotational grazing.
3.3. Stability Analysis of Plant Community Stability
Under different grazing modes, the plant communities in the FSNG pastures exhibited a VR > 1, indicating that the community populations were generally positively correlated (Table 5). The test value was W = 9.52, within the interval of (X20.95, X20.05), indicating that it did not reach the significance level. The community plant populations were in the unstable stage of overall positive succession. TSRG and SG pastures had VR < 1, indicating that the community had overall negative associations. The test values W = 5.46 and W = 6.42, respectively, fell within the (X20.95, X20.05) interval, indicating that they did not reach the significance level. This result suggests that the community is in the unstable stage of negative succession. Compared with the FSNG areas, the TSRG and SG areas were characterized by looser interspecific linkages, greater independence between species, and plant community degradation risk.
TABLE 5.
Overall correlation between typical steppe plant communities in China and Mongolia.
| Test sample | Variance ratio | Test statistic | X2 threshold | Test results |
|---|---|---|---|---|
| FSNG | 1.19 | 9.52 | (2.73, 15.51) | Positive correlation |
| TSRG | 0.683 | 5.46 | Negative correlation | |
| SG | 0.802 | 6.42 | Negative correlation |
Abbreviations: FSNG, four‐season nomadic grazing; SG, settled grazing; TSRG, two‐season rotational grazing.
The Godron plant community stability calculation method was employed to simulate the community using a smoothing curve (Figure 5). Furthermore, the coordinates of the intersection point of the smoothed curve and a straight line were determined. By integrating the principle of stability from Godron, the proximity of these coordinates (x, y) to the equilibrium point (20, 80) indicates the community's stability level. Conversely, a greater distance from these coordinates to the equilibrium point suggests lower stability. For visual representation, the Euclidean distance between the intersection point coordinates and those of the equilibrium point is utilized. The Godron stability determination results for communities with different grazing methods indicated that the Euclidean distance of Godron stability determination was 7.16 for FSNG, 13.22 for TSRG, and 16.45 for SG (Table 6). According to the principle, the closer to the equilibrium point (20, 80), the smaller the Euclidean distance, and the higher the stability of the community. The FSNG area was closer to the coordinates of the stabilization point, followed by TSRG, and the SG pastures were the farthest from the coordinates of the stabilization point. Overall, vegetation communities in FSNG pastures were more stable.
FIGURE 5.

Godron stability index plots for typical steppe plant communities in Inner Mongolia and Mongolia. FSNG, four‐season nomadic grazing; SG, settled grazing; TSRG, two‐season rotational grazing.
TABLE 6.
Stability analysis of typical steppe communities under different grazing treatments.
| Test sample | Fitting curve | R 2 | Intersection coordinates | Euclidean distance |
|---|---|---|---|---|
| FSNG | y = −0.011x 2 + 1.65x + 41.89 | 0.88 | (23.35, 73.67) | 7.16 |
| TSRG | y = −0.011 ± 1.647 + 41.89 | 0.87 | (29.38, 70.68) | 13.22 |
| SG | y = −0.013x 2 + 1.91x + 35.00 | 0.89 | (30.77, 67.56) | 16.45 |
Abbreviations: FSNG, four‐season nomadic grazing; SG, settled grazing; TSRG, two‐season rotational grazing.
4. Discussion
4.1. Different Grazing Patterns and Plant Community Structures
Community structure is an important indicator of the health of grassland plant communities, particularly changes in the dominant species that play a significant role in the direction of grassland ecosystem succession (Chang et al. 2020). First, different grazing practices have a discernible impact on grassland community composition and structural characteristics. In this study, the FSNG grassland had the highest number of species, followed by TSRG and SG had the lowest number of species. This indicates that FSNG has a high species composition diversity and is significantly different from the TSRG and SG communities. Higher grazing rates and consistently higher levels of disturbance resulted in rapid degradation of the pasture, whereas FSNG provided time for pasture restoration and community vegetation recovery due to the degree of mobility of the livestock (Bao, Yi, et al. 2013). Second, in the FSNG area, perennial dominant species, such as L. chinensis, S. grandis, and C. duriuscula, had higher importance values, whereas annuals species had a lower proportion and importance; in addition, the dominance of inchworm moss among the perennial miscellaneous grasses decreased (Wang et al. 2017). In contrast, under SG conditions, the importance value of annual and biennial plants and shrubs and semishrubs increased while the importance value of inchworm moss among the perennial forbs was relatively high. This finding indicated that the plant community is undergoing degradation and succession as a result of the reduction of branches and leaves of the more palatable perennial grasses and the reduction of the effective photosynthetic area due to overfeeding by livestock, resulting in the replacement of perennial grasses by perennial forbs, annual and biennial plants, and shrubs. Overall, FSNG reduces the intensity of livestock disturbance, improves soil physicochemical properties, and promotes vegetation growth (Bao et al. 2014), while SG degrades grassland ecosystems due to long‐term livestock disturbance, which can be restored through the implementation of grazing methods, such as nomadic grazing.
4.2. Different Grazing Patterns and Plant Community Characteristics
Differences in grazing patterns affect the degree of grassland vegetation utilization, causing changes in the quantitative features of plant communities (Bat‐Oyun et al. 2016). The density, height, and cover of community vegetation, as the most basic indicators for grassland monitoring are the most intuitive parameters for describing the basic conditions of grasslands as well as auxiliary evaluation indicators in grassland community studies (Schönbach et al. 2011).
In this study, both community cover and community height were higher in the FSNG pasture than the TSRG and SG pastures, suggesting that moderate grazing disturbance and sufficient recreational time positively influence the increase in vegetation cover while rainfall mainly affects vegetation height (Sun, Zhu, and Zhang 2014). However, plant communities under SG conditions exhibited the highest density due to overgrazing, which led to a notable decline in perennial grasses, which are preferred by livestock, and a simultaneous increase in the number of annual and biennial plants, which are less favored by livestock. Additionally, significant differences were observed in the total aboveground biomass of plant communities across different grazing practices, with FSNG pastures displaying notably higher biomass compared to SG pastures. This disparity in biomass could be attributed to grazing pressure reducing plant abundance and subsequently diminishing the overall photosynthetic area, thereby impacting the efficiency of plant organic matter accumulation. Overall, the production performance of FSNG and TSRG was higher than that of SG, indicating that grassland degradation was more pronounced under SG.
4.3. Different Grazing Practices and Plant Community Species Diversity
The species diversity of plant communities is a measurable indicator of community structure and closely related to ecosystem functioning. Community diversity changes because of different grazing practices (McIntyre, Heard, and Martin 2003; Grenke et al. 2022). This study demonstrated that the FSNG pastures have the highest values for the three alpha diversity indices, except for the Margalef index, indicating that the communities contained the most information, had the highest number of vegetation species, and were more evenly distributed. Moreover, the FSNG communities were more complex and stable compared with the TSRG and sedentary grazing. The population differences between TSRG and SG were similar, which might be related to the differences in the zonal habitats of the sample sites, the experimental sampling, and their community features. Additionally, the FSNG pastures have lower grazing pressure in summer and are less disturbed, and some of the less palatable plants may affect livestock foraging at a certain stage of growth, which increases their dominance. However, their effects on the community require investigation in fixed‐point continuous experiments.
4.4. Current Status of Interspecific Connectivity and Stability of Plants Under Different Grazing Patterns
The overall correlation of plant communities with different grazing practices is a general pattern of interspecific relationships that can reflect the stability and direction of succession of plant communities under different grazing practices (Jian et al. 2018). The stronger the overall association strength, the more robust the mutualism among typical steppe plant species, leading to a more stable community (Qi et al. 2010). Conversely, heightened interspecific competition tends to unfavorably impact plant survival. Upon analyzing the overall interspecific correlation of plant communities, we observed that FSNG pasture communities displayed a positive correlation overall, although it was not statistically significant. This finding indicates that FSNG interspecific associations are loose and somewhat independent and in the stage of continuous improvement; moreover, the plant community structure is still unstable (Jian et al. 2018). Additionally, the Godron plant community stability determination method revealed that FSNG was closer to the coordinates of the stabilization point, followed by TSRG, whereas SG was the farthest away. This disparity arises from the high‐intensity, sustained utilization of grasslands in SG. Overgrazing by domestic animals during the pasture growing season not only curtails the time for compensatory growth of pasture grasses but also prompts further harvesting of deadfalls at the end of the season. Consequently, this leads to a significant reduction in grassland vegetation cover and biomass, resulting in an unstable structure of the plant community. TSRG pastures end grazing earlier in the year compared with SG pastures, thereby allowing more time for compensatory growth after grazing ends and increasing the biomass of pregrazed grasslands more quickly.
FSNG contributes to the stability of the community based on the Godron analysis. SG leads to the disappearance of community species, and species disappearance tends to lead to the loss of a certain plant function from the community. Furthermore, invasive species change the original composition of the community, which has an impact on the stability of the community (Lei et al. 2023).
Our study conclusions are summarized as follows:
Grazing mode affects the composition and function of plant communities, and there are differences in the dominant species under different grazing modes. The number of species was higher under FSNG, in which perennial dominant species such as L. chinensis, S. grandis, and C. duriuscula had larger importance values and smaller percentages of annuals and biennials. The number of species in the TSRG and SG pastures was lower, and the perennial forbs C. squarrosa and the typical steppe degradation indicator plants C. duriuscula and C. glaucum appeared in the SG pastures at the same time. Perennials dominated under FSNG, while annuals and biennials were more pronounced under SG pastures.
Community cover, community height, and aboveground biomass were higher in FSNG pastures than in TSRG and SG pastures; community densities were the highest in TSRG pastures but not significantly different.
Diversity characterization revealed that the Shannon–Wiener, Simpson, and Pielou indexes were higher in the FSNG pastures compared with TSRG and SG pastures, while the Margalef index was highest for TSRG. However, none of these indices showed significant differences. FSNG appeared to support the maintenance of diversity and stability in typical steppe communities, with no significant difference observed between the effects of TSRG and SG on community diversity. Additionally, FSNG, TSRG, and SG plant communities exhibited considerable dissimilarity, showcasing a wide range of structural differentiation and a high degree of habitat variability among the communities.
Analysis of interspecific correlation revealed that the FSNG pasture plant communities were positively correlated, with loose interspecific linkages and a certain degree of independence, and in the stage of continuous improvement. The TSRG and SG pastures were negatively correlated and were in the unstable stage of dynamic succession. Stability analysis was used to further explain that FSNG is more stable than TSRG and SG.
Author Contributions
Suriguga Gao: conceptualization (equal), data curation (equal), formal analysis (equal), investigation (equal), writing – original draft (equal). Yu Hong: funding acquisition (equal), project administration (equal), resources (equal). Wulan Tuya: conceptualization (equal), project administration (equal), writing – review and editing (equal). Weiqing Zhang: conceptualization (equal), project administration (equal), supervision (equal). Chang An: methodology (equal), project administration (equal), resources (equal). Siqin Chaoketu: conceptualization (equal), software (equal), validation (equal). Xu Sha: data curation (equal), resources (equal). Bu He: data curation (equal), software (equal). Wu Yinga: data curation (equal), resources (equal).
Conflicts of Interest
The authors declare no conflicts of interest.
Acknowledgments
This research was funded by the National Nature Science Foundation of China (Nos 41861024, 42161023, and 42061031), Natural Nature Science Foundation of Inner Mongolia (No. 2024MS04014), Inner Mongolia Autonomous Region Science and Technology Program (No. 2020GG0039), and Inner Mongolia Normal University Basic Research Operating Expenses Special Project (Nos 2022JBZD017 and 2022JBTD010), and Inner Mongolia Autonomous Region Key R&D and Achievement Transformation Programme Project (No. 2023KJHZ0027).
Funding: This research was funded by the National Nature Science Foundation of China (Nos 41861024, 42161023, and 42061031), Natural Nature Science Foundation of Inner Mongolia (No. 2024MS04014), Inner Mongolia Autonomous Region Science and Technology Program (No. 2020GG0039), and supported by the Fundamental Reasearch Funds for the Inner Mongolia Normal University (No. 2022JBTD010 and 2022JBZD017), and Inner Mongolia Autonomous Region Key R&D and Achievement Transformation Programme Project (No. 2023KJHZ0027).
Data Availability Statement
The data that support the findings of this study are available from the corresponding author (Wulan Tuya) upon reasonable request.
References
- Bao, G. , Bao Y., Qin Z., Zhou Y., and Shiirev A.. 2013. “Vegetation Cover Changes in Mongolian Plateau and Its Response to Seasonal Climate Changes in Recent 10 Years.” Scientia Geographica Sinica 33, no. 5: 613–621. [Google Scholar]
- Bao, G. , Liu Y., Liu N., and Wang W. P.. 2013. “Characteristics of Climate Changes During the Instrumental Period in the Eastern and Southern Mongolian Plateau and Their Ecological‐Environmental Effects.” Journal of Earth & Environment 4, no. 5: 1444–1460. [Google Scholar]
- Bao, X. , Yi J., Lian Y., Liu S., and Jimuse W.. 2009. “Study on Communities Diversity of Different Grazing in Typical Steppe.” Acta Agriculturae Boreali‐Sinica 24, no. 5: 229–233. [Google Scholar]
- Bao, X. , Yi J., Lian Y., Xiao X., and Bao X.. 2013. “Effect of Grazing on Plant Functional Group Characteristics in Stipa Steppe Between China and Mongolia.” Bulletin of Botanical Research 33, no. 2: 159–165. [Google Scholar]
- Bao, X. X. , Lian Y., Yi J., and Zhang R. X.. 2014. “Effects of Grazing on Plant Functional Group Characteristics in Stipa klemenzii Desert Steppe in China and Mongolia.” Chinese Journal of Ecology 33, no. 11: 2966–2972. http://www.cje.net.cn/EN/Y2014/V33/I11/2966. [Google Scholar]
- Bátori, Z. , Körmöczi L., Erdös L., Zalatnai M., and Csiky J.. 2012. “Importance of Karst Sinkholes in Preserving Relict, Mountain, and Wet‐Woodland Plant Species Under Sub‐Mediterranean Climate: A Case Study From Southern Hungary.” Journal of Cave & Karst Studies 74, no. 1: 127–134. [Google Scholar]
- Bat‐Oyun, T. , Shinoda M., Cheng Y., and Purevdorj Y.. 2016. “Effects of Grazing and Precipitation Variability on Vegetation Dynamics in a Mongolian Dry Steppe.” Journal of Plant Ecology 9, no. 5: 508–519. [Google Scholar]
- Batunacun, H. , Biligejifu Y., Liu J., and Zhen L.. 2015. “Spatial Distribution and Variety of Grass Species on the Ulan Bator‐Xilinhot Transect of Mongolian Plateau.” Journal of Natural Resources 30, no. 1: 24–36. [Google Scholar]
- Briske, D. D. , Derner J. D., Brown J. R., et al. 2008. “Rotational Grazing on Rangelands: Reconciliation of Perception and Experimental Evidence.” Rangeland Ecology & Management 61, no. 1: 3–17. [Google Scholar]
- Chang, H. , Sun H. L., Liu Y. H., Qiu X., Shi L., and Wen C.. 2020. “Community Structure and Diversity Under Different Degrees of Degraded Grassland in East Wuzhumuqin Meadow.” Acta Agrestia Sinica 28, no. 1: 184–192. [Google Scholar]
- Conte, T. J. , and Tilt B.. 2014. “The Effects of China's Grassland Contract Policy on Pastoralists' Attitudes Towards Cooperation in an Inner Mongolian Banner.” Human Ecology 42, no. 6: 837–846. [Google Scholar]
- Cumming, W. F. P. , Newman R. F., Bradfield G. E., Krzic M., and Thompson D. J.. 2016. “Grazing and Solar Irradiance Drive Patterns of Change in Two Grassland Plant Communities in Southern British Columbia.” Journal of Arid Environments 133: 94–101. [Google Scholar]
- De Boeck, H. J. , Bloor J. M. G., Kreyling J., et al. 2018. “Patterns and Drivers of Biodiversity‐Stability Relationships Under Climate Extremes.” Journal of Ecology 106, no. 3: 890–902. [Google Scholar]
- Gao, L. , Zhang S., Zhu Z., Xu R., and Zhang P.. 2019. “Effects of Grazing on Plant Communities and Ecological Functions in Typical Steppe.” Research of Soil & Water Conservation 26, no. 6: 205–211. [Google Scholar]
- Grenke, J. S. J. , Bork E. W., Carlyle C. N., Boyce M. S., and Cahill J. F.. 2022. “Limited Impacts of Adaptive Multi‐Paddock Grazing Systems on Plant Diversity in the Northern Great Plains.” Journal of Applied Ecology 59, no. 7: 1734–1744. [Google Scholar]
- Huang, X. , Wang S., Huang B., et al. 2015. “Analyses of Community Stability and Interspecific Associations Between the Rare Plant Phyllitis scolopendrium and Its Associated Species.” Acta Ecologica Sinica 35, no. 1: 80–90. [Google Scholar]
- Ishii, N. I. , Takahashi M., Yoshihara Y., et al. 2021. “Genetic Diversity, Population Size, and Population Stability of Common Plant Species in a Mongolian Grassland.” Journal of Arid Environments 194: 104607. [Google Scholar]
- Jacobo, E. J. , Rodríguez A. M., Bartoloni N., and Deregibus V. A.. 2006. “Rotational Grazing Effects on Rangeland Vegetation at a Farm Scale.” Rangeland Ecology & Management 59, no. 3: 249–257. [Google Scholar]
- Jian, X. , Shui W., Wang Y., et al. 2018. “Species Diversity and Stability of Grassland Plant Community in Heavily‐Degraded Karst Tiankeng: A Case Study of Zhanyi Tiankeng in Yunnan, China.” Acta Ecologica Sinica 38, no. 13: 4704–4714. [Google Scholar]
- Lei, S. L. , Liao L. R., Wang J., et al. 2023. “The Diversity‐Godron Stability Relationship of Alpine Grassland and Its Environmental Drivers.” Acta Prataculturae Sinica 32, no. 3: 1–12. [Google Scholar]
- Li, X. , and Qu Q.. 2002. “Soil Microbial Biomass Carbon and Nitrogen in Mongolian Grassland.” Acta Pedologica Sinica 39, no. 1: 97–104. [Google Scholar]
- Linders, T. E. W. , Schaffner U., Eschen R., et al. 2019. “Direct and Indirect Effects of Invasive Species: Biodiversity Loss is a Major Mechanism by Which an Invasive Tree Affects Ecosystem Functioning.” Journal of Ecology 107, no. 6: 2660–2672. [Google Scholar]
- Liu, K. , Liu Z., Zhou N., et al. 2022. “Diversity‐Stability Relationships in Temperate Grasslands as a Function of Soil pH.” Land Degradation & Development 33, no. 10: 1704–1717. [Google Scholar]
- Liu, S. , Yu G., Jun A., et al. 2009. “The Thawing‐Freezing Processes and Soil Moisture Distribution of the Steppe in Central Mongolian Plateau.” Acta Pedologica Sinica 46, no. 1: 46–51. [Google Scholar]
- McIntyre, S. , Heard K. M., and Martin T. G.. 2003. “The Relative Importance of Cattle Grazing in Subtropical Grasslands: Does It Reduce or Enhance Plant Biodiversity?” Journal of Applied Ecology 40, no. 3: 445–457. [Google Scholar]
- Miao, L. , Jiang C., He B., Liu Q., Zhu F., and Cui X.. 2014. “Response of Vegetation Coverage to Climate Change in Mongolian Plateau During Recent 10 Years.” Acta Ecologica Sinica 34, no. 5: 1295–1301. [Google Scholar]
- Na, Y. , Bao S., Hashimoto K., McCarthy C., and Hoshino B.. 2018. “The Effects of Grazing Systems on Plant Communities in Steppe Lands‐A Case Study From Mongolia's Pastoralists and Inner Mongolian Settlement Areas.” Land 7, no. 1: 10. [Google Scholar]
- Na, Y. , Li J., Hoshino B., Bao S., Qin F., and Myagmartseren P.. 2018. “Effects of Different Grazing Systems on Aboveground Biomass and Plant Species Dominance in Typical Chinese and Mongolian Steppes.” Sustainability 10, no. 12: 4753. [Google Scholar]
- Qi, K. , Zhang C. Y., Hou J. H., and Zhao X. H.. 2010. “Dynamics of Species Diversity and Interspecific Associations of Herbaceous Plants in a Pinus tabulaeformis Forest on a Sandy Site in Chifeng, China.” Acta Ecologica Sinica 30, no. 18: 5106–5112. [Google Scholar]
- Roxburgh, S. H. , Shea K., and Wilson J. B.. 2004. “The Intermediate Disturbance Hypothesis: Patch Dynamics and Mechanisms of Species Coexistence.” Ecology 85, no. 2: 359–371. [Google Scholar]
- Sasaki, T. , Lu X., Hirota M., and Bai Y.. 2019. “Species Asynchrony and Response Diversity Determine Multifunctional Stability of Natural Grasslands.” Journal of Ecology 107, no. 4: 1862–1875. [Google Scholar]
- Schönbach, P. , Wan H., Gierus M., et al. 2011. “Grassland Responses to Grazing: Effects of Grazing Intensity and Management System in an Inner Mongolian Steppe Ecosystem.” Plant and Soil 340, no. 1–2: 103–115. [Google Scholar]
- Shen, T. , Wang Y., and Han G.. 2022. “Response of Stipa Breviflora Desert Steppe Ecosystem Stability to Grazing.” Grassland & Turf 42, no. 6: 88–94. [Google Scholar]
- Sun, Z. , Zhu J., and Zhang X.. 2014. “Influence of Short‐Period Grazing Intensity on Vegetation Characteristic and Diversity of Meadow Steppe in Zhaosu.” Journal of Xinjiang Agricultural University 37, no. 1: 35–39. [Google Scholar]
- Suriguga, G. , Wulantuya S., Zhang S., and Meili W.. 2022. “Relationship Between Species Diversity and Biomass of Stipa krylovii Steppe Community and Its Responses to Grazing Intensity.” Acta Ecologica Sinica 42, no. 23: 9736–9746. [Google Scholar]
- Tao, S. L. , Fang J. Y., Zhao X., et al. 2015. “Rapid Loss of Lakes on the Mongolian Plateau.” Proceedings of the National Academy of Sciences of the United States of America 112, no. 7: 2281–2286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valerio, M. , Ibáñez R., Gazol A., and Götzenberger L.. 2022. “Long‐Term and Year‐to‐Year Stability and Its Drivers in a Mediterranean Grassland.” Journal of Ecology 110, no. 5: 1174–1188. [Google Scholar]
- Wang, X. , Ma H., Shen Y., et al. 2019. “Effects of Different Rotational Grazing Patterns on Plant Community Characteristics in Desert Steppe Grassland.” Acta Prataculturae Sinica 28, no. 4: 23–33. [Google Scholar]
- Wang, X. F. , Wang C., Hu J. P., Liu S. R., Zeng Z. H., and Hu Y. G.. 2017. “Plant Communities Diversity of Meadow Steppe Under Different Grazing Systems of Household Pasture.” Acta Agrestia Sinica 25, no. 3: 466–473. [Google Scholar]
- Wulan, T. 2021. “Characteristics of Grassland Utilization in Mongolian Plateau and Their Differences Among Countries.” Acta Geographica Sinica 76, no. 7: 1722–1731. [Google Scholar]
- Yao, T. , Zhu Z., Li Y., et al. 2016. “Effects of Functional Diversity and Functional Redundancy on the Community Stability of an Alpine Meadow.” Acta Ecologica Sinica 36, no. 6: 1547–1558. 10.5846/stxb201408301725. [DOI] [Google Scholar]
- Yin, Y. , Liu H. Y., Liu G., Hao Q., and Wang H.. 2013. “Vegetation Responses to Mid‐Holocene Extreme Drought Events and Subsequent Long‐Term Drought on the Southeastern Inner Mongolian Plateau, China.” Agricultural and Forest Meteorology 178–179: 3–9. [Google Scholar]
- Zhang, X. , Hu Y., Zhuang D., and Qi Y.. 2009. “The Spatial Pattern and Differentiation of NDVI in Mongolia Plateau.” Geographical Research 28, no. 1: 10–19. [Google Scholar]
- Zhang, X. , Zhang W., Sai X., et al. 2022. “Grazing Altered Soil Aggregates, Nutrients and Enzyme Activities in a Stipa kirschnii Steppe of Inner Mongolia.” Soil & Tillage Research 219: 105327. [Google Scholar]
- Zhang, Y. Z. , Wang Z. Q., Yang Y., Li J. L., Zhang Y., and Zhang C. B.. 2018. “Research on the Quantitative Evaluation of Grassland Degradation and Spatial and Temporal Distribution on the Mongolia Plateau.” Pratacultural Science 35, no. 2: 233–243. [Google Scholar]
- Zhang, Z. , Li Y., Ding Y., Li F., Sun J., and Li X.. 2020. “Responses of Plant Community Diversity and Functional Community Characteristics of Typical Grassland to Grazing Factors.” Chinese Journal of Grassland 42, no. 5: 48–54. [Google Scholar]
- Zhao, M. , Zhao K., Wang Y., et al. 2023. “Long‐Term Grazing Disturbance Reduced Plant Diversity in Stipa breviflora Desert Steppe.” Acta Prataculturae Sinica 32, no. 9: 39–49. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author (Wulan Tuya) upon reasonable request.