

Abstract
Background
False negative rapid diagnostic tests (RDTs) accruing to the non-detection of Plasmodium falciparum histidine-rich protein 2/3 (Pfhrp2/3) is threatening the diagnosis and management of malaria. Although regular monitoring is necessary to gauge the level of efficacy of the tool, studies in Cameroon remain limited. This study assessed Plasmodium spp. prevalence and Pfhrp2/3 gene deletions across ecological and transmission zones in Cameroon.
Methods
This is a cross-sectional, multi-site, community- and hospital- based study, in 21 health facilities and 14 communities covering all five ecological settings in low seasonal (LS) and intense perennial (IPT) malaria transmission zones between 2019 and 2021. Participants were screened for malaria parasite using Pfhrp2 RDT and light microscopic examination of thick peripheral blood smears. DNA was extracted from dried blood spot using chelex®-100 and P. falciparum confirmed using varATS real-time quantitative Polymerase Chain Reaction (qPCR), P. malariae and P. ovale by real-time qPCR of Plasmepsin gene, and P. vivax using a commercial kit. Isolates with amplified Pfcsp and Pfama-1 genes were assayed for Pfhrp 2/3 gene deletions by conventional PCR.
Results
A total of 3,373 participants enrolled, 1,786 Plasmodium spp. infected, with 77.4% P. falciparum. Discordant RDT and qPCR results (False negatives) were reported in 191 (15.7%) P. falciparum mono-infected samples from LS (29%, 42) and IPT (13.9%, 149). The Pfhrp2+/Pfhrp3 + genotype was most frequent, similar between LS (5.5%, 8/145) and IPT (6.0%, 65/1,076). Single Pfhrp2 and Pfhrp3 gene deletions occurred in LS (0.7%, 1/145 each) and IPT (3.6%, 39/1,076 vs. 2.9%, 31/1,076), respectively. Whilst a single sample harboured Pfhrp2-/Pfhrp3- genotype in LS, 2.4% (26/1,076) were double deleted at IPT. Pfhrp2+/Pfhrp3- (0.3%, 3/1,076) and Pfhrp2-/Pfhrp3+ (1.2%, 13/1,076) genotypes were only observed in IPT. Pfhrp2, Pfhrp3 deletions and Pfhrp2-/Pfhrp3- genotype accounted for 78.8% (26), 69.7% (23) and 63.6% (21) RDT false negatives, respectively.
Conclusion
Plasmodium falciparum remains the most dominant and widely distributed Plasmodium species across transmission and ecological zones in Cameroon. Although the low prevalence of Pfhrp2/3 gene deletions supports the continued use of HRP2-based RDTs for routine malaria diagnosis, the high proportion of false-negatives due to gene deleted parasites necessitates continued surveillance to inform control and elimination efforts.
Supplementary Information
The online version contains supplementary material available at 10.1186/s12879-024-09935-4.
Keywords: Malaria diagnosis, Plasmodium falciparum, Histidine-rich protein, Gene deletion
Background
Malaria remains a socio-economic burden and global public health concern, despite the intensification of control interventions and overall decline in the disease incidence [1]. Sub-Saharan Africa (sSA) continues to bear the brunt of the disease [2], with children below five years of age, pregnant women, and naïve travellers from malaria free to the disease endemic areas most vulnerable [1]. The high incidence of malaria in Africa, particularly in sSA has long been attributed to several factors, including poverty, poor implementation of intervention measures, and insufficient medical infrastructure [3]. In addition, hasty, uncontrolled and haphazard urbanization, dilapidated semi-urban and rural constructions, population mobility, discrepancy and inconsistency in implementing control measures are increasing transmission in cities and rural areas across Africa [4, 5]. In addition, most of sSA remain malaria endemic, with a significant reservoir for seeding resurgence and spread of the parasite due partly to the increasing parasite and vector resistance to drugs and insecticides, two important malaria control interventions [6].
The incidence of malaria has been on the rise in Cameroon [6], with an estimated 2 million cases in 2018, representing 24% medical consultations, 45% hospitalizations, and 3 thousand malaria deaths in health facilities, equivalent to 13% of all cases of death. Malaria parasite transmission in Cameroon is heterogeneous and the epidemiology categorised into five ecological zones with varying transmission intensities; intense and perennial in the Humid Forest (HF) with monomodal (HF-M) or bimodal (HF-B) rainfall, and Humid Savannah (HS), low in the High Guinea Savannah (HGS) and seasonal in the Sudano-Sahelian (SS) zone [7]. The majority (71%) of the population of approximately 25 million reside in intense and perennial transmission settings in the country, where there are several circulating primate-infecting Plasmodium species but human infections are mainly by Plasmodium falciparum, although minor non-falciparum species (P. malariae, P. ovale, and P. vivax) are also being transmitted [7, 8].
In Cameroon, measures to combat malaria initiated since 2002 by the government and put in place by the National Malaria Control Program (NMCP) include, amongst others, early diagnosis of disease cases [9]. Current malaria management guidelines in the country, recommend prompt parasitological confirmation of all suspected malaria cases by light microscopy and/or rapid diagnostic tests (RDTs) prior to antimalarial treatment [10]. Although laboratory diagnosis of malaria traditionally involves the microscopic examination of Giemsa-stained blood smears and remains the gold standard tool [11, 12], its limitations, including the need for skilled technicians and electricity supply [13] necessitated the development of RDTs [11].
Rapid diagnostic tests are fast, affordable, easy to perform and interpret, reliable and effective diagnostic complement to light microscopy for the point of care malaria case management especially during emergencies and in endemic rural settings and areas with limited laboratory facilities [13–15]. These RDTs rely on P. falciparum histidine-rich protein 2 (Pfhrp2) encoded by the Pfhrp2 gene, as well as its paralogue, Pfhrp3, lactate dehydrogenase (LDH), or aldolase antigens, which bind to antibodies on the test strip in parasite infected samples [16]. These antibodies are designed to recognise the hrp2 antigen but may cross-react with hrp3, a structurally homologous [17] protein of the hrp2 family [18]. Nevertheless, RDTs are limited by their inability to detect infections at low parasitaemia, the possibility of false positive or negative results due to the persistence of target antigen even after successful treatment or deletions in the hrp2/3 genes, respectively [11, 19] or detection of non-Plasmodium infection [20, 21].
Variation in the sensitivity of Pfhrp RDTs have been reported and attributed to several factors, including poor quality products from some brands or operator proficiency. The heterogeneity of false negative RDTs has been attributed to variation in malaria prevalence, with low transmission regions and the resulting low parasitaemia being an important determinant [22]. However, there are also reports of extensive genetic variations worldwide in the Exon 1 and 2 regions of Pfhrp2/3 genes, including complete or partial deletion of parts or all of Pfhrp2 and Pfhrp3 genes [23–26]. These result in the absence or truncation of the hrp2/3 antigens, with the resultant false-negative diagnosis and poor management of the malaria [27, 28]. The Pfhrp-deleted mutant parasite strains present a threat to malaria elimination as they cannot be identified for interventions. Their prevalence across endemic populations should be assessed and reported to support local diagnostic policies for malaria.
Although there are numerous reports of false negative RDTs and Pfhrp2/3 deletions in malaria endemic settings in sSA, studies in Cameroon have been limited to a few localities on the slope of mount Cameroon [29], Douala, Maroua, Mayo-Oulo and Pette [30], Mfou and Yaounde [31]. Regular monitoring is necessary to determine whether local Pfhrp2/3 deletion prevalence is increasing towards the WHO threshold of 5% of false negatives among symptomatic individuals [6] requiring local or national change in diagnostic strategy. This is particularly important as deletion in Pfhrp2/3 genes have been reported in neighbouring countries (Nigeria, Equatorial Guinea, Ghana) with high and unrestricted trans-border mobility [6, 32, 33]. This study assessed the Plasmodium spp. prevalence, false negative malaria RDTs and Pfhrp2/3 gene deletions across ecological and malaria transmission zones in Cameroon to determine whether the prevalence has reached a threshold requiring local or national change in diagnostic strategy. Detection of low level parasitaemia undetected by RDTs, and low frequency of Pfhrp2/3 deletions have implications for RDT surveillance, control and management of malaria in Cameroon.
Methods
Study area
This study was undertaken across all five ecological zones in Cameroon (Table 1, Fig. 1), located on the Gulf of Guinea in the Central Africa region, between latitude 7° 22’ 11” North and longitude 12° 20’ 41” East (Fig. 1). The distinct geo-climatic features in these zones mimic most of sSA allowing for relatively high biodiversity in the country. This includes hot, humid, and dry tropical climate [34, 35], vegetation, coastal lowlands, wetlands and highlands, deep equatorial evergreen forest, and humid savannah [36].
Table 1.
Major characteristics of the study sites
| Characteristics | Malaria parasite transmission | ||||
|---|---|---|---|---|---|
| Intense and Perennial | Low and seasonal | ||||
| Ecological zone | HF-M | HS | HF-B | HGS | SS |
| Region(s) | Southwest | Northwest | Centre, East | Adamawa | Far North |
| Town(s) | Buea, Limbe, Mamfe, Mutengene, Tiko | Bamenda | Bertoua, Yaounde | Meiganga | Mora |
| Number of Health facilities | 16 | 5 | |||
| Enrolment* | 2,849a | 108 | 149 | 181 | 86 |
| Temperature (°C) | 17–34 | 15–28 | 17–33 | 12–34 | 15–40 |
| Season | Long rainy & short dry | Two rainy & two dry | Long rainy & short dry | Short rainy & long dry | |
| Humidity (%) | 82–87 | 56–86 | 48–86 | 78–83 | 49–80 |
| Precipitation (mm) | 3,033 | 2,400–3,000 | 1,357–1,540 | 1,500–1,533 | 610–760 |
| Annual Rainfall (mm) | 1,600–4,000 | 2,000–3,000 | 1,600–2,000 | 1,200–1,600 | 500–1,200 |
| EIR | 100 | 10 | |||
*Samples were enrolled from Health facilities (clinical samples) in all ecological zones; aIncludes 747 clinical and 2,102 community samples. EIR = Entomological Inoculation Rate, SS = Sudano-Sahelian, HGS = High Guinea Savannah, HF-B = Humid Forest with bimodal rainfall, HF-M = Humid Forest with monomodal rainfall, HS = Humid Savannah
Fig. 1.

Sampling sites across the geo-climatic zones in Cameroon
Plasmodium spp. transmission in Cameroon is highly heterogenous (Table 1): intense perennial lasting 7–12 months in the HF and HS and 1–3 months in the HGS and SS zones with low and seasonal transmission [7]. The SS zone is characterised by a high population migration from neighbouring Nigeria, Chad, and Central African Republic, while the HGS zone harbours several camps of refugees or displaced persons from the aforementioned countries. The outbreak of socio-political unrest in the HS and HF-M since 2017 has led to the displacement of persons to other zones.
Study design, population and sampling
This study applied both cross-sectional community- and hospital- based malaria parasite surveys between April 2019 and October 2021 during the dry and rainy seasons. Sites were located across the three malaria transmission settings and five ecological zones of Cameroon (Fig. 1). Participants included individuals reporting for medical consultation or screening for febrile illness at selected health facilities (HFs) or mobilisation points in the different target localities. Each individual was tested for malaria by RDT and parasite infection confirmed by PCR. A total of 21 HFs were selected randomly from ten towns (Table 1) as well as fourteen communities from Buea, Tiko, and Limbe health districts in the HS-M of the Southwest region.
For the clinical samples, health facilities were randomly selected in each ecological zone among the main public health facilities serving the town and all febrile individuals reporting to the Outpatient Department for consultation or parturient women at Antenatal Clinic eligible for enrolment. For the community surveys, communities were first identified as rural and semi-urban and then randomly selected across altitudinal zones. Prior to participant enrolment, each local population was then sensitized on the project objectives, methods, and possible benefits/risks through health personnel, elites and/or community leaders. Participants were then invited to a central enrollment point and resident adults interviewed in either English/French (national languages) or pidgin-English (common local creole).
Due to the absence of conclusive preliminary evidence on false-negative RDT prevalence or Pfhrp2/3 gene deletions in the country, an initial minimum of 370 symptomatic P. falciparum patients per sampling domain (ecological/transmission zone) was targeted in line with WHO guidelines [16]. This sample size would be adequate to demonstrate that a Pfhrp2/3 gene deletion prevalence is below the 5% threshold for an expected population prevalence of 3.2% with 95% confidence.
A pre-tested semi-structured questionnaire was used to document participant’s socio-demographic and clinical parameters (Supplementary File 1). Approximately 2 ml of whole blood was collected from each participant by venepuncture into vacutainers coated with EDTA (Microtainer, Becton Dickinson) for malaria parasite detection by HRP2-based RDT and microscopy. About 50 μl of blood was then spotted on Whatman™ 3 mm filter paper (Sigma-Aldrich), dried for 4 h at ambient temperature and stored sealed in a zip lock bag containing a desiccant at -20 °C for subsequent PCR analysis.
Malaria rapid diagnostic testing
The presence of Plasmodium spp. in blood was assessed using the PfHRP2-based malaria rapid diagnostic test kit (SD Bioline™, Alere, South Korea) according to the manufacturer’s guide. Briefly, 5 μl of blood was placed in the sample window of the RDT cassette and three drops of diluent added. The results were then read after 15 min, with the presence of two (or three), one or no distinct band indicative of a positive, negative or invalid result, respectively.
Light microscopy
Thick blood smears were prepared on clean grease-free glass slides and allowed to air-dry at ambient temperature. The smears were then stained with 10% Giemsa (Sigma-Aldrich) for 15 min as described [37], air-dried and viewed under the 100x oil immersion objective and 10x eyepiece of an optical binocular microscope (Olympus Optical Co. Ltd., Japan). Each slide was examined by two independent microscopists and considered negative if no parasite was seen after counting 500 leucocytes. With each positive slide, the parasite densities were estimated by counting the parasites against a minimum of 200 white blood cells and assuming an average leucocyte count of 8,000 per μl of blood [38].
DNA extraction and diagnostic PCR
DNA was extracted from dried blood spots (DBS) on filter paper using the Chelex® – Tween® 20 method as described [39], with some modifications. Briefly, three 3 mm DBS discs were punched into 1 ml of 0.5% Tween®-20 and incubated at 4 °C overnight to lyse the red blood cells. The DBS were then washed in 1 ml of 1 x PBS and allowed to incubate at 4 °C for 30 min and incubated in 150 μl of hot 5% of chelex®-100 resins in the water bath at 95 °C for 15 min. The supernatant of each well (DNA extract) was then transferred into a new tube, its concentration checked using a Qubit® 3.0 fluorometer and the DNA stored at -20 °C.
Plasmodium spp. were detected by quantitative real-time PCR (qPCR) amplification targeting the var gene acidic terminal sequence (varATS) for P. falciparum [40], Plasmepsin 4 for P. malariae, P. ovale [41] and P. vivax (Genesig kit, PrimerDesign UK). 3D7 reference strain DNA and nuclease-free water were used as positive and negative controls, respectively, in all amplifications.
All P. falciparum amplifications were carried out on a CFX96™ real-time PCR system, in a 20 μl reaction volume. The varATS assay contained 0.8 μM each of the forward and reverse primers (Table S1), 0.4 μM of probe, 1x TaqMan™ Universal PCR Master-mix (Applied Biosystems), and 5 μl of genomic DNA. Thermocycling conditions involved one cycle of pre-incubation at 50 °C for 2 min, one cycle initial denaturation at 95 °C for 10 min, followed by 45 cycles of a two-step denaturation and annealing/elongation at 95 °C for 15 s and 55 °C for 1 min [40].
Plasmodium malariae and P. ovale identification was undertaken by multiplex semi-nested and real-time quantitative PCR. A pre-amplification reaction was undertaken on a C1000 Touch™ thermal cycler, in a total volume of 15 μl, comprising 0.1 μM of each forward and reverse primers (Table S1) for P. malariae and P. ovale, 1x Qiagen Multiplex PCR Master-mix (Qiagen), and 5 μl of genomic DNA. Thermocycling conditions were set at initial denaturation of 95 °C for 5 min, and 12-cycle reaction of denaturation, annealing and elongation at 95 °C, 60 °C and 72 °C for 30, 60 and 90 s, respectively.
The speciation qPCR was done on a Bio-Rad CFX96™ real-time system, in a total volume of 15 μl containing 0.4 μM each of forward and reverse primers for either P. malariae or P. ovale, 0.2 μM of the species-specific probe (Table S1), 1x TaqMan™ Universal PCR Master-mix (Applied Biosystems), and 3.0 μl of the pre-amplification product. Thermocycling conditions were set at 95 °C for 10 min, followed by 39 cycles of denaturation, annealing and elongation at 95 °C and 60 °C for 10 s and 60 s, respectively, with fluorescence acquisition.
Plasmodium vivax was identified using a commercial kit (Genesig®, PrimerDesign, UK) according to the manufacturers’ protocol.
Amplifications of Pfcsp and Pfama-1 genes
The circumsporozoite protein (csp) and apical membrane antigen 1 (ama-1) genes were amplified in all P. falciparum positive isolates prior to the detection of deletion in Pfhrp 2 and 3 genes. Only samples with amplified Pfcsp and Pfama-1 gene targets were assayed for hrp2/3 gene deletions. 3D7 reference DNA and nuclease-free water were used as positive and negative controls, respectively, in all amplifications.
The Pfcsp and Pfama-1 genes were amplified by nested PCR using specific predesigned primers as described [42]. Briefly, the Pfcsp primary reaction was done on a C1000 Touch™ thermal cycler in a 25 μl reaction volume containing 0.25 μM each of forward and reverse primers, 0.2 mM dNTPs (Qiagen), 0.02 U/μl Q5 Hot Start High-Fidelity DNA Taq (Applied Biosystems), 1.5 mM MgCl2, 1x Q5 buffer (New England Biolabs) and 5 μl of genomic DNA. Thermocycling conditions were set at initial denaturation of 98 °C for 30 s, followed by 34 cycles of denaturation at 98 °C for 30 s, annealing at 56 °C for 30 s, elongation at 72 °C for 2 min and a final extension at 72 °C for 2 min.
The nested reaction was similar to the primary reaction using the specific nested primer set and 1 μl of primary PCR amplicon. Thermocycling conditions were slightly modified with an initial denaturation at 98 °C for 1 min, followed by 34 cycles of denaturation at 98 °C for 45 s, annealing at 56 °C for 45 s, elongation at 72 °C for 1 min, and a final elongation at 72 °C for 5 min.
Pfama-1 gene amplification conditions were similar to the Pfcsp amplification, except the use of 0.5 μM of the forward and reverse primers for the primary PCR reaction and an annealing temperature of 63 °C for 46 s. The nested PCR reaction also contained 0.5 μM each of forward and reverse primers and 1 μl of primary PCR amplicon as template. Thermocycling conditions were set at initial denaturation of 98 °C for 1 min, followed by 34 cycles of denaturation at 98 °C for 45 s, annealing at 63 °C for 45 s, elongation at 72 °C for 1 min and a final elongation at 72 °C for 5 min.
Detection of deletion of Pfhrp2 and 3 genes
This was undertaken as a one-step PCR on the C1000 Touch™ thermal cycler using specific predesigned primers (Table S2) as described [29]. Both Pfhrp2 and Pfhrp3 genes were amplified separately in a 25 μl reaction containing 0.2 μM of forward and reverse primers, 0.1 U/μl Taq DNA polymerase (New England Biolabs), 0.2 mM dNTPs (Qiagen), 1x Thermopol buffer (New England Biolabs), and 5 μl of genomic DNA. Thermocycling conditions were set at initial denaturation of 95 °C for 5 min, followed by 39 cycles of denaturation at 95 °C for 45 s, annealing at 50 °C for 1 min, elongation at 68 °C for 1 min, and a final elongation at 70 °C for 5 min.
Gel electrophoresis
The nested PCR products were separated on a 1.5% agarose gel (Sigma, USA) stained with 5 μl Redsafe dye (Chembio, UK) in 1x TBE at 100 volts for 40 min. The bands were visualised on the Gel Doc™ EZ System (Biorad).
Data analysis
All data were double entered in Microsoft Excel version 2016 and analysed using SPSS Statistics 20.0 (IBM Corp, Atlanta, GA, USA). Associations between proportions were explored using the Pearson’s Chi square (x2) test while differences in group means were assessed using the student’s t test or analysis of variance (ANOVA) test.
Plasmodium falciparum infection status was classified as reported previously [38] as negative if all tests (RDT, microscopy and PCR, where applicable) were negative, submicroscopic if microscopy (and RDT, where applicable) was negative but PCR positive, microscopic if microscopy or RDT (where applicable) was positive whether the PCR was positive or negative. False positives or negatives denoted samples that were positive or negative by routine tests (RDT or microscopy) but negative or positive by PCR respectively.
Parasitaemia was defined as presence of parasite or parasite antigen/DNA in blood using light microscopy, RDT or PCR. In order to identify “true Pfhrp2/3 deletions”, parasitaemia was corrected for Ct value below 37 using qPCR to minimize the risk of finding false deletion results associated with a single copy of the target gene (Pfhrp2) and low-density infection [43]. Symptomatic infection was defined as fever or history of fever with parasitaemia while asymptomatic parasitaemia was defined as parasite infection in the absence of fever or history of fever as previously reported [35]. All analyses were performed at 95% confidence intervals and the level of significance (p) set at ≤ 0.05 for all statistical tests.
Results
Malaria parasite infection
A total of 3,373 participants were pre-screened from Antenatal Care (12,5%, 420), clinical (25.2%, 851) and community (62.3%, 2,102) surveys across all five ecological zones (Table 2). The prevalence of malaria parasite infection by RDT was 60.7% (255/420) and 32.6% (684/2,099) from ANC and community surveys, respectively. In all, 37.8% (1,237/3,269) and 25.5% (832/3,269) malaria parasite infections were symptomatic and asymptomatic respectively. One thousand three hundred and ninety-eight (78.3%) of the 1,786 RDT positives and clinical samples were confirmed positive by qPCR, with 7% (125/1,784) false positive RDT prevalence overall. PCR identified 20 (1.1%) and 42 (2.4%) of false positive and false negative light microscopic results respectively.
Table 2.
Socio-demographic and clinical characteristics of P. falciparum infected participants
| Characteristics | Subgroup | N | Overall [% (n)] |
Transmission setting [% (n)] | P value | |
|---|---|---|---|---|---|---|
| Low and seasonal | Intense and perennial | |||||
| Age group (years) | < 5 | 1,354 | 13.6 (184) | 7.4 (13) | 14.4 (171) | < 0.001 |
| 5–9 | 14.4 (195) | 7.9 (13) | 15.3 (182) | |||
| 10–14 | 11.6 (157) | 4.8 (8) | 12.5 (149) | |||
| ≥ 15 | 60.4 (818) | 79.4 (131) | 57.8 (687) | |||
| Gender | Male | 1,376 | 35.8 (493) | 32.5 (54) | 36.3 (439) | 0.343 |
| Female | 64.2 (883) | 67.5 (112) | 63.7 (771) | |||
| Ethnic group | Bantu | 1,273 | 27.2 (346) | 1.1 (1) | 29.2 (345) | < 0.001 |
| Semi-Bantu | 63.9 (813) | 6.5 (6) | 68.4 (807) | |||
| Sudanese | 7.8 (99) | 92.5 (86) | 1.1 (13) | |||
| Non-Cameroonian | 1.2 (15) | 0 (0) | 1.3 (15) | |||
| Clinical phenotype | Asymptomatic | 1,285 | 32.2 (414) | 3.2 (5) | 36.3 (409) | < 0.001 |
| Symptomatic | 67.8 (871) | 96.8 (153) | 63.7 (718) | |||
| Infection status | Microscopic | 1,383 | 98.4 (1,361) | 92.8 (154) | 99.2 (1,207) | < 0.001 |
| Submicroscopic | 1.6 (22) | 7.2 (12) | 0.8 (10) | |||
| Non falciparum species | 1,383 | 11.7 (162) | 12.7 (21) | 11.6 (141) | 0.689 | |
| Anaemia | 1,088 | 46.6 (507) | 34.5 (20) | 47.3 (487) | 0.057 | |
| Bednet usage | 1,283 | 46.5 (596) | 75 (69) | 44.2 (527) | < 0.001 | |
| Hrp2-based RDT positives | 1,381 | 84.9 (1,173) | 69.9 (116) | 87 (1,057) | < 0.001 | |
| RDT False negatives | 15.1 (208) | 30.1 (50) | 13.0 (158) | |||
| GMPD (parasites/μl of blood) | 1,335 | 9,566 | 4,991 | 10,406 | 0.013 | |
GMPD = Geometric Mean Parasite Density; Masl = Metres above Sea Level
Distribution of Plasmodium spp.
Plasmodium falciparum was the predominant human malaria parasite species across all ecological and transmission zones in Cameroon, infecting 77.4% (1,383/1,786), 76.2% (1,361/1,786) and 75.1% (1,342/1,786) of participants overall following qPCR, light microscopy and RDT respectively (Fig. 2).
Fig. 2.
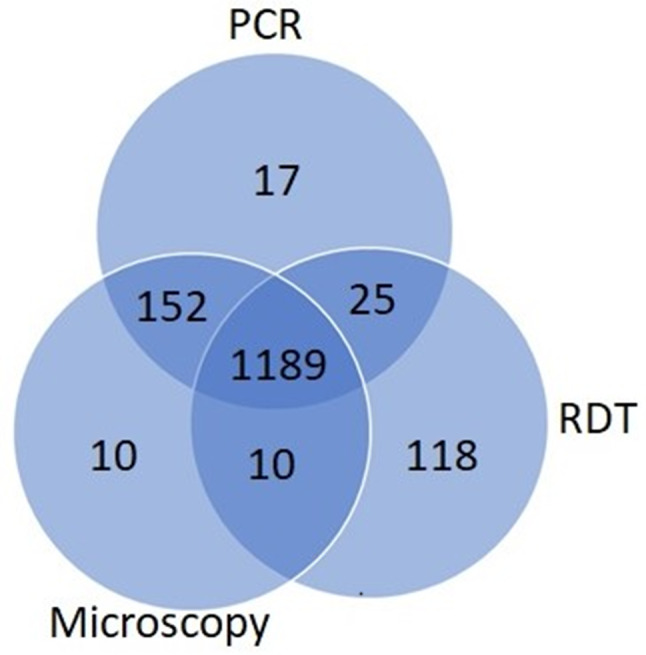
Number of P. falciparum infected individuals detected by light microscopy, RDT and qPCR across ecological and transmission zones in Cameroon (n = 1,383)
The proportion of P. falciparum infections was higher (p < 0.001) in areas with intense and perennial transmission (80.1%, 1,217/1,519) compared to low and seasonal transmission zones (62.2%, 166/267) (Fig. 3). One hundred and seventy-seven (9.9%) of the participants harboured non-falciparum infections, with 6.4% (115/1,786) and 4.1% (73/1,785) of the population containing P. malariae and P. ovale, respectively, while P. vivax was undetected in all samples analysed. Mixed infections of P. falciparum with P. malariae and P. ovale were detected in 5.9% (106/1,786) and 3.7% (66/1,786) participants, respectively, while 0.6% (11) samples were co-infected with P. malariae and P. ovale, and 0.6% (10) with P. falciparum, P. malariae and P. ovale.
Fig. 3.

Distribution of infecting Plasmodium species in individuals across ecological (a) and transmission (b) zones in Cameroon (n = 1,786). Only Plasmodium spp. mono-infections were considered. SS = Sudano-Sahelian, HGS = High Guinea Savannah, HF-B = Humid Forest with bimodal rainfall, HF-M = Humid Forest with monomodal rainfall, HS = Humid Savannah, LS = Low and seasonal, IPT = Intense and Perennial transmission
Socio-demographic and clinical characteristics of P. falciparum infected participants
The characteristics of the 1,383 P. falciparum positive participants by qPCR is shown (Table 2). Most participants were females (64.2%, 883), aged 15 years and above (60.4%, 818), enrolled from clinical surveys (44.4%, 614), in Urban settings (58.9%, 814), the Semi-Bantu ethnic group (63.9%, 813), and low altitude areas (51.6%, 713) with high and intense transmission (88.8%, 1,217). Plasmodium falciparum hrp2 antigen was detected in 1,173 participants, resulting in an overall RDT false negative prevalence of 15.1% (208/1,383). The prevalence of asymptomatic and submicroscopic infections were 32.2% (414) and 1.6% (22) respectively. Almost half of the population were anaemic (46.6%, 507/1,088) and reportedly frequent bednet users (46.5%, 596/1,283), with 11.7% (162) harbouring non-falciparum infections. Overall, study participants were infected with 9,566 (range: 80–438, 225) asexual parasites/μl of blood, with higher (p = 0.013) parasite loads in areas with intense and perennial transmission compared to their low and seasonal counterparts (Table 2).
Pfhrp2-based RDT false negatives
A total of 1,221 P. falciparum mono-infected samples from the SS (5.5%, 67) and HGS (6.4%, 78) with low and seasonal transmission (11.9%, 145) and HS (6.4%, 78), HF-B (10.2%, 124) and HF-M (71.6%, 874) intense and perennial transmission zones (88.1%, 1,076) were retained for the analysis of Pfhrp2/3 gene deletion (Fig. 4). Of the 191 (15.7%) samples with discordant RDT and qPCR results (False negatives) from LS (29%, 42) and IPT (13.9%, 149) settings (and 10% qPCR positives as controls), 134 were deemed appropriate for Pfhrp2/Pfhrp3 genotyping following both Pfcsp and Pfama-1 gene amplifications to control DNA quantity and quality (Table 3). False negative RDT samples (36.11 ± 5.51) had higher (p < 0.001) qPCR Ct values (i.e., lower parasite DNA concentration) compared to RDT positive cases (24.13 ± 3.41) (Fig. 4). Similarly, the parasite load by light microscopy was lower (p = 0.001), on average, in false negatives compared to their positive counterparts (Table 3).
Fig. 4.

Sample processing and results for Pfhrp2 and Pfhrp3 genotyping in ecological and transmission zones in Cameroon. Terminal boxes show the presence (+) or absence (−) of the Pfhrp2 and Pfhrp3 genes from samples appropriate for genotyping. All percentages shown are out of the total of 145 and 1,076 DBS samples with P. falciparum mono-infections analysed in the zones with low/seasonal and intense/perennial malaria parasite transmission, respectively. SS = Sudano-Sahelian, HGS = High Guinea Savannah, HF-B = Humid Forest with bimodal rainfall, HF-M = Humid Forest with monomodal rainfall, HS = Humid Savannah
Table 3.
Characteristics of the P. falciparum mono-infected Pfcsp and Pfama-1 amplified isolates genotyped for Pfhrp2/Pfhrp3
| Parameter | All subjects | Pfhrp2-based RDT Status | P value | |
|---|---|---|---|---|
| Positive | False Negative | |||
| DBS Processed | 134 | 97 | 37 | |
| Transmission zone [% (n)] | ||||
| IPT | 93.3 (125) | 91.8 (89) | 97.3 (36) | 0.183 |
| LS | 6.7 (9) | 8.2 (8) | 2.7 (1) | |
| Hrp2/3 gene deletions [% (n)] | ||||
| Pfhrp2 | 34.5 (40) | 16.9 (14) | 78.8 (26) | < 0.001 |
| Pfhrp3 | 23.9 (32) | 8.2 (8) | 64.9 (24) | < 0.001 |
| Pfhrp2-/Pfhrp3- | 23.3 (27) | 7.2 (6) | 63.6 (21) | < 0.001 |
| Pfhrp2-/Pfhrp3+ | 11.2 (13) | 9.6 (8) | 15.2 (5) | 0.292 |
| Pfhrp2+/Pfhrp3- | 2.6 (3) | 1.2 (1) | 6.1 (2) | 0.194 |
| Pfhrp2+/Pfhrp3+ | 62.9 (73) | 81.9 (68) | 15.2 (5) | < 0.001 |
| Ct (Pf), Mean ± SD (range) |
27.3 ± 6.6 (16.3–43.9) |
24.1 ± 3.5 (16.3–31.8) |
35.5 ± 5.9 (24.4–43.9) |
< 0.001 |
| GMPD [range] (parasites/μl blood) |
15,967 [160–405,000] |
20,029 [160–405,000] |
8,523 [400–120,000] |
0.001 |
GMPD = Geometric Mean Parasite Density; LS = Low/Seasonal Transmission; IPT = Intense and Perennial Transmission
Pfhrp2/Pfhrp3 gene deletions
The wild-type Pfhrp2+/Pfhrp3 + genotype was the most frequent and similar in individuals with asymptomatic and symptomatic infections as well as across both LS (5.5%, 8/145) and IPT (6.0%, 65/1,076) transmission zones. Single Pfhrp2 and Pfhrp3 gene deletions were observed in both the LS (0.7%, 1/145 each) and IPT (3.6%, 39/1,076 vs. 2.9%, 31/1,076) zones, respectively. Whilst only a single sample harboured the Pfhrp2-/Pfhrp3- double deletion genotype in the LS setting, 2.4% (26/1,076) were double deleted at IPT (Fig. 4). Parasites with the Pfhrp2+/Pfhrp3- (0.3%, 3/1,076) and Pfhrp2-/Pfhrp3+ (1.2%, 13/1,076) genotypes were only observed in areas with intense and perennial transmission.
In comparison to wild-type Pfhrp2+/Pfhrp3 + parasites (24.04 ± 3.74, range: 16.25–34.44), significantly higher qPCR Ct values (i.e., lower parasite densities) were observed in infections with P. falciparum lacking the Pfhrp2 gene alone, Pfhrp2-/Pfhrp3+ (28.43 ± 5.04, range: 21.60–39.67, p = 0.010) (Fig. 5). Infections with P. falciparum double deletion isolates, Pfhrp2-/Pfhrp3- (36.17 ± 6.20, range: 23.23–43.91) had significantly higher Ct values compared to wild-type Pfhrp2+/Pfhrp3+ parasites (p < 0.001) and those parasites lacking the Pfhrp2 gene alone, Pfhrp2-/Pfhrp3+ (p < 0.001).
Fig. 5.

Variation in (a) all Ct values (n = 116), (b) Ct values < 37 (n = 101), (c) all parasite density (n = 114) and (d) parasite density > 1,000 parasites/μl (n = 109) with Pfhrp2/Pfhrp3 gene deletion status in Pfhrp2-based RDT positive and false negative patients in Cameroon
Overall, the prevalence of false-negative HRP2-RDT caused by confirmed Pfhrp2, Pfhrp3 and Pfhrp2-/Pfhrp3-, Pfhrp2-/Pfhrp3+, and Pfhrp2+/Pfhrp3- deletions among symptomatic P. falciparum cases were 78.8% (26/33), 69.7% (23/33), 63.6% (21/33), 15.2% (5/33), and 6.1% (2/33), respectively, with 22,2% (7/33) and 30.3% (10/33) of the RDT false negatives were found to be Pfhrp2 and Pfhrp3 positive by PCR. Eighteen isolates were considered ‘true deletions’ harbouring 61.1% (11/18), 50.0% (9/18), 38.9% (7/18), 22.2% (4/18), 11.1% (2/18) and 27.8% (5/18) hrp2 and hrp3 deleted genes as well as the Pfhrp2-/Pfhrp3-, Pfhrp2-/Pfhrp3+, Pfhrp2+/Pfhrp3- and Pfhrp2+/Pfhrp3 + genotypes, respectively, when corrected for Ct values < 37.0. Conversely, 45.5% (15) samples with Pfhrp2/3 deleted genes were found to be submicroscopic in this study (Ct value above 37 using qPCR), 100% (15) and 93.3% (14) of which harboured hrp2 and hrp3 deleted genes as well as 93.3% (14), 6.7% (1), 0% (0) and 0% (0) with the Pfhrp2-/Pfhrp3-, Pfhrp2-/Pfhrp3+, Pfhrp2+/Pfhrp3- and Pfhrp2+/Pfhrp3 + genotypes, respectively.
Discussion
Histidine-rich protein 2-based RDTs are reliable, affordable, accessible, and easy to use tools in remote settings and remain critical in the diagnosis of malaria especially in sub-Saharan Africa where P. falciparum is most prevalent. In fact, most countries in Africa have adopted the HRP2-based RDT as a pragmatic, sensitive and the only P. falciparum-specific diagnostic test in many settings. Although this tool has been primarily limited by the inability to detect low parasitaemia in blood, the recurrent evidence of false negatives has also been attributed to the non-detection of the protein [44–46]. The presence of P. falciparum lacking either or both hrp2 and hrp3 genes has been documented in numerous countries [47, 48] and recognized as a substantial additional threat to malaria case management, control, and elimination because of false-negative diagnostic results [49]. Loss of this tool would be a considerable setback to accurate monitoring of malaria case incidence within a country and to achieving the goal of universal confirmation of malaria infection before antimalarial chemotherapy [50]. This study enrolled participants across all ecological/transmission zones in Cameroon to assess the presence and patterns of Plasmodium spp. and Pfhrp2/3 gene deletions throughout the country where information on the extent of deletions remains inadequate [29–31].
Plasmodium falciparum was the most abundant circulating human infecting Plasmodium species across all malaria transmission and ecological zones in Cameroon, reflecting the outcome of the relative species abundance and co-infection with non-falciparum species in previous studies in the country [51, 52]. Such varying distribution of Plasmodium species definitely would trigger varying competitiveness in the host, adaptations, and diversity in disease pathogenesis, manifestations, and response to control and elimination efforts. In line with previous reports in the southern part of Cameroon [53] and elsewhere, P. vivax was not detected in this study, in spite of the very wide geographical spread of the sampling undertaken and highly sensitive nature of the detection assay. This is suggestive of a more restrictive / limited or localised distribution of the species in the country to a few foci in Dschang, Santchou, and Kyé-ossi [54], Bolifamba [8], Douala, Ebolowa, Yaounde, and Bertoua [55]. A countrywide mapping of species and rigorous and targeted implementation of efficient measures would be necessary to effectively eliminate the disease.
This current study found evidence of deletions in hrp2 and hrp3 genes in P. falciparum isolates from different disease spectrum (parturient, symptomatic, and asymptomatic individuals) across zones with low/seasonal and intense/perennial transmission in Cameroon, obtained in 2019–2021. However, minimal circulation of parasites with single and double Pfhrp2/3 gene deletions were detected, consistent with previous reports in the country [29, 31], elsewhere in DRC, Central Africa [56] and even South Sudan [57] near the Horn of Africa with documented high prevalence of deletions [43, 58]. Although the reported prevalence of hrp2/3 gene deletions is below the 5% WHO threshold and support the continued use of HRP2-based RDTs, regular and sustained surveillance / monitoring of Pfhrp2/3 gene deletions, with careful laboratory workflows are necessary to support malaria control and elimination efforts [56].
Pfhrp2/3 double-deleted parasites genes were detected across both transmission zones, indicating the potential challenge of PfHRP2 RDT-based malaria diagnosis in the country [43], since these do not produce the HRP2/HRP3 antigen targets of the RDT and their assessment with HRP-based RDTs will produce a high false-negative rate [59]. False-negative RDT results not only prevent appropriate treatment when needed but also impede malaria control and elimination efforts by systematically under-reporting the true prevalence of the infection [56, 60]. Further multi-site and more extensive studies, conducted according to WHO’s protocol on Pfhrp2/3 gene deletion surveillance will help facilitate decisions regarding the appropriateness and effectiveness of Hrp2-based RDTs currently in use in Cameroon [61] since the implementation of the tool is likely exerting selective pressure on the parasite population and could lead to the spread of Pfhrp2/3 gene-deleted parasites [62].
The WHO recommends surveillance of a combination of HRP2-based RDT with an LDH-based RDT or smear microscopy on malaria cases with discordant/HRP2 false negative RDT result as an initial trigger for widespread hrp2/hrp3 gene deletion surveillance [16]. The results presented here only reflect the presence and geographical spread, not a precise prevalence estimate of Pfhrp2/3 gene deletions in Cameroon [56] and may have been influenced by a number of factors. First, false positives were screened using the more sensitive qPCR in this analysis, irrespective of symptoms, 10% of which had positive RDT results, with submicroscopic infection potentially mitigating the prevalence of Pfhrp2/3 gene deletion [63]. In addition, the timing of the surveillance during the transmission season is quite relevant to the survey since HRP2 may persist in the plasma up to 28 days after treatment particularly in high parasitaemia infections [64] and a new infection with P. falciparum parasites lacking Pfhrp2/3 genes may have circulating HRP2 from the previous infection in areas with high malaria transmission. Furthermore, circulating HRP3 protein alone can give a positive HRP2-RDT signal in the absence of the Pfhrp2 locus when parasitaemia is more than 1,000 parasite/μl [48], since high sequence and structural homology between epitopes of the two isoform proteins [65] have been shown to mask the deletion effect on either of the genes on the performance of RDT, allowing cross-reaction of the monoclonal antibodies of PfHRP2 with PfHRP3 [66] at moderate to high parasite density. Pfhrp2/3 gene deletions have been reported in asymptomatic infections in previous studies [60] and their inclusion here highlights the importance of surveying individuals with different disease spectrum and outcomes [67].
Although molecular assays targeting exon 1/2 have been shown to yield spurious amplification of the paralogous gene [59], lower parasite densities (high Ct values), common among asymptomatic individuals, could have influenced the prevalence of deletions in RDT false-negative (lower plasma HRP2 level) infections [68] since they may not provide a high-quality sample for detection of gene deletions by PCR [61]. In fact, 21.2% (7/33) and 35.1% (13/37) of the RDT false negatives were Pfhrp2 and Pfhrp3 positive, respectively, by PCR (Fifteen samples with the Pfhrp2/3 gene deletion overall) in this study. These probably reflect samples with low levels of parasitaemia below the detection threshold of Pfhrp2 RDT, submicroscopic infections, and the absence of Pfhrp2 antigen due to host immune response in the high transmission areas [69, 70]. This hypothesis is supported by higher parasitaemia in this study as detected by light microscopy in RDT positives compared to false positives [68]. For this reason, the WHO recommends assessing Pfhrp2/3 gene deletions among symptomatic persons [16]. Only eighteen samples with a Ct value below 37 using qPCR (and hence had > 9 parasite density/μl) were considered true deletions to minimize the risk of finding false deletion results associated with a single copy of the target gene (Pfhrp2) and low-density infection [43]. Plasmodium falciparum positive isolates with Pfhrp2/3 gene deletion in Pfhrp2 RDT negative submicroscopic infections are potentially infectious to mosquitoes and contribute to ongoing malaria transmission, selectively increasing to challenge malaria control and prevention if not continuously monitored [43].
Limitations
This study did not enrol the WHO recommended number of health centres across all ecological/transmission zones and so does not provide a precise prevalence estimate of Pfhrp2/3 gene deletions in Cameroon. Secondly, the surveys were largely conducted in the middle of peak transmission season (April – September) and in some areas previously reported to have high polyclonal P. falciparum infections, possibly with both hrp2/3-deleted and hrp2/3-wild type clones, possibly underestimating gene deletions since the multiplicity of infection was not ascertained. Additionally, it was not possible to assess the contribution of causes other than gene deletions to false-negative RDT, such as adherence to manufacturer’s instructions, transportation or storage conditions. Measurement of PfHRP2 antigen in the plasma was also not performed.
Conclusion
Plasmodium falciparum remains the most widely distributed and abundant human infecting Plasmodium species across all transmission and ecological zones in Cameroon. This study demonstrates less than 5% of Pfhrp2/3 gene deletions in P. falciparum isolates, supporting the continued use of HRP2-based RDTs for routine malaria diagnosis in the country. The high proportion of false-negatives due to gene deleted parasites necessitates continued nationwide surveillance to continuously inform malaria diagnosis, case management, control, and elimination efforts.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Acknowledgements
We appreciate participants from the communities and health facilities that made this study possible, and the administrative authorities, medical personnel, and community leaders who assisted with this work.
Abbreviations
- GMPD
Geometric mean parasite density
- HF-B
Humid forest with bimodal rainfall
- HF-M
Humid forest with monomodal rainfall
- HGS
High Guinea Savannah
- HS
Humid Savannah
- IPT
Intense perennial transmission
- LS
Low and seasonal
- Pfhrp
Plasmodium falciparum histidine-rich protein
- qPCR
Quantitative polymerase chain reaction
- RDT
Rapid diagnostic test
- SS
Sudano-Sahelian
- sSA
Sub-Saharan Africa
- varATS
Variable acidic terminal sequence
Author contributions
TOA: conceptualized, designed, coordinated the study, and drafted the manuscript. LNT: coordinated the study, collected samples, performed experiments, and drafted the manuscript. ECO: coordinated laboratory activities and improved on the manuscript. BNE and BN: performed experiments. SD: processed samples and performed experiments. MAO: supervised experiments and improved on the manuscript. VNN and PNK: collected and processed samples and improved on the manuscript. HFC: provided logistics and supervised sample collection. EAA: provided critical review of the manuscript. AAN: provided logistics, coordinated and substantially improved on the manuscript. All authors reviewed the manuscript.
Funding
This study received financial support in the form of a postdoctoral grant number 107740/Z/15/Z from the Developing Excellence in Leadership and Genetics Training for Malaria Elimination in sub-Saharan Africa (DELGEME) program, sponsored by the Developing Excellence in Leadership, Training and Science (DELTAS) Africa initiative and doctoral research assistantship number H3AFull/17/008 from the Pan African Malaria Genetic Epidemiology Network (PAMGEN) - Human Heredity and Health in Africa (H3Africa). The funders have no role in designing the study, collecting and analysing the data, deciding to publish, and preparing the manuscript.
Data availability
All data generated or analysed during this study are included in this published article [and its supplementary information files].
Declarations
Ethics approval and consent to participate
Ethical clearance was obtained from the Cameroon National Ethics Committee (Ref. N0 2019/09/1188/CE/CNERSH/SP and 2020/12/1315/CE/CNERSH/SP). Administrative authorisations were sought from the Regional Delegations of Public Health for the Adamawa, Centre, East, Far North, Northwest and Southwest regions, and their respective District Medical Services and the Directorate of Human Resources of the Cameroon Development Corporation. At community level, authorization was obtained from Chiefs or Quarter heads where appropriate. Only prospective participants or their guardians who volunteered to participate by signing a written informed consent, after sensitization were enrolled. Assent was also obtained from all children above 12 years of age and pregnant women below 16 years of age in line with national gynaecological guidelines.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Tobias Obejum Apinjoh, Email: apinjoh.tobias@ubuea.cm.
Alfred Amambua-Ngwa, Email: alfngwa@lshtm.ac.uk.
References
- 1.World Health Organization. World malaria report 2019. Geneva: World Health Organization. 2019 [cited 2024 Mar 21]. https://iris.who.int/handle/10665/330011
- 2.By-Nc-Sa C. World malaria report 2023.
- 3.Gallup J, Sachs J. The economic burden of malaria. Am J Trop Med Hyg. 2001;64:85–96. [DOI] [PubMed] [Google Scholar]
- 4.Wang S-J, Lengeler C, Smith TA, Vounatsou P, Cissé G, Diallo DA, et al. Rapid urban malaria appraisal (RUMA) in sub-Saharan Africa. Malar J. 2005;4:40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ndo C, Menze-Djantio B, Antonio-Nkondjio C. Awareness, attitudes and prevention of malaria in the cities of Douala and Yaoundé (Cameroon). Parasites Vectors. 2011;4:181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.World malaria. Report 2020 20 years of global progress and challenges. Geneva: World Health Organization; 2020. [Google Scholar]
- 7.Antonio-Nkondjio C, Ndo C, Njiokou F, Bigoga JD, Awono-Ambene P, Etang J, et al. Review of malaria situation in Cameroon: technical viewpoint on challenges and prospects for disease elimination. Parasites Vectors. 2019;12:501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fru-Cho J, Bumah VV, Safeukui I, Nkuo-Akenji T, Titanji VP, Haldar K. Molecular typing reveals substantial Plasmodium vivax infection in asymptomatic adults in a rural area of Cameroon. Malar J. 2014;13:170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.The world health report. 2000: Health systems: improving performance. 2000.
- 10.World Health Organization. Global malaria programme. Guidelines for the treatment of malaria. 2015.
- 11.Haditsch M. Quality and reliability of current malaria diagnostic methods. Travel Med Infect Dis. 2004;2(3–4):149–60. [DOI] [PubMed] [Google Scholar]
- 12.World Health Organization. Malaria microscopy quality assurance manual. Version 2. Geneva: World Health Organization; 2016. [Google Scholar]
- 13.Wilson ML. Laboratory diagnosis of malaria: conventional and rapid diagnostic methods. Arch Pathol Lab Med. 2013;137(6):805–11. [DOI] [PubMed] [Google Scholar]
- 14.World Health Organization. World Health Organization, Global Malaria Programme. Universal access to malaria diagnostic testing: an operational manual. Geneva: World Health Organization; 2011. [Google Scholar]
- 15.Cheng Q, Gatton ML, Barnwell J, Chiodini P, McCarthy J, Bell D, et al. Plasmodium falciparum parasites lacking histidine-rich protein 2 and 3: a review and recommendations for accurate reporting. Malar J. 2014;13:283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.World Health Organization. False-negative RDT results and P. Falciparum histidine-rich protein 2/3 gene deletions. WHO. 2019;10(1):12. [Google Scholar]
- 17.Akinyi S, Hayden T, Gamboa D, Torres K, Bendezu J, Abdallah JF, et al. Multiple genetic origins of histidine-rich protein 2 gene deletion in Plasmodium falciparum parasites from Peru. Sci Rep. 2013;3:2797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lee N, Baker J, Andrews KT, Gatton ML, Bell D, Cheng Q, et al. Effect of sequence variation in Plasmodium falciparum histidine-rich protein 2 on binding of specific monoclonal antibodies: implications for rapid diagnostic tests for malaria. J Clin Microbiol. 2006;44(8):2773–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gamboa D, Ho M-F, Bendezu J, Torres K, Chiodini PL, Barnwell JW, et al. A large proportion of P. Falciparum isolates in the Amazon Region of Peru Lack pfhrp2 and pfhrp3: implications for malaria rapid diagnostic tests. PLoS ONE. 2010;5(1):e8091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gatton ML, Ciketic S, Barnwell JW, Cheng Q, Chiodini PL, Incardona S et al. An assessment of false positive rates for malaria rapid diagnostic tests caused by non-Plasmodium infectious agents and immunological factors. PLoS One. 2018.13(5):e0197395. [DOI] [PMC free article] [PubMed]
- 21.Hosch S, Yoboue CA, Donfack OT, Guirou EA, Dangy JP, Mpina M, et al. Analysis of nucleic acids extracted from rapid diagnostic tests reveals a significant proportion of false positive test results associated with recent malaria treatment. Malar J. 2022;21:2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Watson OJ, Sumner KM, Janko M, Goel V, Winskill P, Slater HC, et al. False-negative malaria rapid diagnostic test results and their impact on community-based malaria surveys in sub-saharan Africa. BMJ Glob Health. 2019;4:e001582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Baker J, Ho M-F, Pelecanos A, Gatton M, Chen N, Abdullah S et al. Global sequence variation in the histidine-rich proteins 2 and 3 of Plasmodium falciparum: implications for the performance of malaria rapid diagnostic tests. Malar J. 2010;9:129. [DOI] [PMC free article] [PubMed]
- 24.Gupta H, Matambisso G, Galatas B, Cisteró P, Nhamussua L, Simone W, et al. Molecular surveillance of pfhrp2 and pfhrp3 deletions in Plasmodium falciparum isolates from Mozambique. Malar J. 2017;16:416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fontecha G, Mejía RE, Banegas E, Ade MP, Mendoza L, Ortiz B, et al. Deletions of pfhrp2 and pfhrp3 genes of Plasmodium falciparum from Honduras, Guatemala and Nicaragua. Malar J. 2018;17:320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Rachid Viana GM, Akinyi Okoth S, Silva-Flannery L, Lima Barbosa DR, Macedo De Oliveira A, Goldman IF et al. Histidine-rich protein 2 (pfhrp2) and pfhrp3 gene deletions in Plasmodium falciparum isolates from select sites in Brazil and Bolivia. Gatton ML, editor. PLoS ONE. 2017;12:e0171150. [DOI] [PMC free article] [PubMed]
- 27.Gendrot M, Fawaz R, Dormoi J, Madamet M, Pradines B. Genetic diversity and deletion of Plasmodium falciparum histidine-rich protein 2 and 3: a threat to diagnosis of P. Falciparum malaria. Clin Microbiol Infect. 2019;25:580–5. [DOI] [PubMed] [Google Scholar]
- 28.Berzosa P, González V, Taravillo L, Mayor A, Romay-Barja M, García L, et al. First evidence of the deletion in the pfhrp2 and pfhrp3 genes in Plasmodium falciparum from Equatorial Guinea. Malar J. 2020;19:99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Eyong EM, Etutu SJM, Jerome F-C, Nyasa RB, Kwenti TE, Moyeh MN. Plasmodium Falciparum histidine-rich protein 2 and 3 gene deletion in the Mount Cameroon region. IJID Reg. 2022;3:300–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Foko LPK, Moukoko CEE, Jakhan J, Narang G, Hawadak J, Motse FDK, et al. Deletions of histidine-rich protein 2/3 genes in natural Plasmodium falciparum populations from Cameroon and India: role of asymptomatic and submicroscopic infections. Am J Trop Med Hyg. 2024;110(6):1100–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Nana RRD, Ngum NL, Makoge V, Amvongo-Adja N, Hawadak J, Singh V. Rapid diagnostic tests for malaria diagnosis in Cameroon: impact of histidine rich protein 2/3 deletions and lactate dehydrogenase gene polymorphism. Diagn Microbiol Infect Dis. 2024;108:1116103. [DOI] [PubMed] [Google Scholar]
- 32.Kojom LP, Singh V. Prevalence of Plasmodium falciparum field isolates with deletions in histidine-rich protein 2 and 3 genes in context with sub-Saharan Africa and India: a systematic review and meta-analysis. Malar J. 2020;19:46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Prosser C, Gresty K, Ellis J, Meyer W, Anderson K, Lee R, et al. Plasmodium Falciparum histidine-rich protein 2 and 3 gene deletions in strains from Nigeria, Sudan, and South Sudan. Emerg Infect Dis. 2021;27:471–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Beck HE, Zimmermann NE, McVicar TR, Vergopolan N, Berg A, Wood EF. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci Data. 2018;5:180214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tangi LN, Ajonina MU, Moyeh MN, Chi HF, Ntui VN, Kwi PN, et al. Knowledge, attitude, and adherence to malaria control guidelines and the prevalence of Plasmodium species infection in localities across transmission and ecological zones in Cameroon. Front Public Health. 2023;11:1060479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mbenda HGN, Awasthi G, Singh PK, Gouado I, Das A. Does malaria epidemiology project Cameroon as ‘Africa in miniature’? J Biosci. 2014;39:727–38. [DOI] [PubMed] [Google Scholar]
- 37.Cheesbrough M. District laboratory practice in tropical countries. 2nd ed. Leiden: Cambridge University Press; 2006. [Google Scholar]
- 38.Apinjoh TO, Ntui VN, Chi HF, Moyeh MN, Toussi CT, Mayaba JM et al. Intermittent preventive treatment with sulphadoxine-pyrimethamine (IPTp-SP) is associated with protection against sub-microscopic P. falciparum infection in pregnant women during the low transmission dry season in southwestern Cameroon: a semi - longitudinal study. PLoS ONE. 2022;18. [DOI] [PMC free article] [PubMed]
- 39.Simon N, Shallat J, Williams Wietzikoski C, Harrington WE. Optimization of Chelex 100 resin-based extraction of genomic DNA from dried blood spots. Biology Methods Protocols. 2020;5:bpaa009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hofmann N, Mwingira F, Shekalaghe S, Robinson LJ, Mueller I, Felger I. Ultra-sensitive detection of Plasmodium falciparum by amplification of multi-copy subtelomeric targets. L Von Seidlein editor. PLoS Med. 2015;12:e1001788. [DOI] [PMC free article] [PubMed]
- 41.Daniels RF, Deme AB, Gomis JF, Dieye B, Durfee K, Thwing JI, et al. Evidence of non-plasmodium falciparum malaria infection in Kédougou, Sénégal. Malar J. 2017;16:9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Baker J, McCarthy J, Gatton M, Kyle DE, Belizario V, Luchavez J, et al. Genetic diversity of Plasmodium falciparum histidine-rich protein 2 (PfHRP2) and its effect on the performance of PfHRP2-based. Rapid Diagnostic Tests; 2005. [DOI] [PubMed]
- 43.Alemayehu GS, Blackburn K, Lopez K, Cambel Dieng C, Lo E, Janies D, et al. Detection of high prevalence of Plasmodium falciparum histidine-rich protein 2/3 gene deletions in Assosa Zone, Ethiopia: implication for malaria diagnosis. Malar J. 2021;20:109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ali IM, Bigoga JD, Forsah DA, Cho-Ngwa F, Tchinda V, Moor VA, et al. Field evaluation of the 22 rapid diagnostic tests for community management of malaria with artemisinin combination therapy in Cameroon. Malar J. 2016;15:31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bamou R, Nematchoua-Weyou Z, Lontsi-Demano M, Ningahi LG, Tchoumbou MA, Defo-Talom BA, et al. Performance assessment of a widely used rapid diagnostic test CareStart™ compared to microscopy for the detection of Plasmodium in asymptomatic patients in the western region of Cameroon. Heliyon. 2021;7:e06271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.By-Nc-Sa C. World malaria report 2022. 2022.
- 47.Berhane A, Anderson K, Mihreteab S, Gresty K, Rogier E, Mohamed S, et al. Major threat to malaria control programs by Plasmodium falciparum lacking histidine-rich protein 2, Eritrea. Emerg Infect Dis. 2018;24:462–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kong A, Wilson SA, Ah Y, Nace D, Rogier E, Aidoo M. HRP2 and HRP3 cross-reactivity and implications for HRP2-based RDT use in regions with Plasmodium falciparum hrp2 gene deletions. Malar J. 2021;20:207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Poti KE, Sullivan DJ, Dondorp AM, Woodrow CJ. HRP2: transforming malaria diagnosis, but with caveats. Trends Parasitol. 2020;36:112–26. [DOI] [PubMed] [Google Scholar]
- 50.Lee N, Gatton ML, Pelecanos A, Bubb M, Gonzalez I, Bell D, et al. Identification of optimal epitopes for Plasmodium falciparum rapid diagnostic tests that target histidine-rich proteins 2 and 3. J Clin Microbiol. 2012;50:1397–405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Feufack-Donfack LB, Sarah-Matio EM, Abate LM, Bouopda Tuedom AG, Ngano Bayibéki A, Maffo Ngou C, et al. Epidemiological and entomological studies of malaria transmission in Tibati, Adamawa region of Cameroon 6 years following the introduction of long-lasting insecticide nets. Parasites Vectors. 2021;14:247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Nguiffo-Nguete D, Nongley Nkemngo F, Ndo C, Agbor J-P, Boussougou-Sambe ST, Salako Djogbénou L, et al. Plasmodium malariae contributes to high levels of malaria transmission in a forest–savannah transition area in Cameroon. Parasites Vectors. 2023;16:31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sundararaman SA, Liu W, Keele BF, Learn GH, Bittinger K, Mouacha F, et al. Plasmodium falciparum -like parasites infecting wild apes in southern Cameroon do not represent a recurrent source of human malaria. Proc Natl Acad Sci USA. 2013;110:7020–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Djeunang Dongho GB, Gunalan K, L’Episcopia M, Paganotti GM, Menegon M, Sangong RE et al. Plasmodium vivax infections detected in a large number of febrile Duffy-Negative Africans in Dschang, Cameroon. The American Journal of Tropical Medicine and Hygiene. 2022 [cited 2024 Mar 21]; 10.4269/ajtmh.20-1255 [DOI] [PMC free article] [PubMed]
- 55.Ngassa Mbenda HG, Das A. Occurrence of multiple chloroquine-resistant pfcrt haplotypes and emergence of the S(agt)VMNT type in Cameroonian Plasmodium falciparum. J Antimicrob Chemother. 2014;69:400–3. [DOI] [PubMed] [Google Scholar]
- 56.McCaffery JN, Nace D, Herman C, Singh B, Sompwe EM, Nkoli PM, et al. Plasmodium Falciparum pfhrp2 and pfhrp3 gene deletions among patients in the DRC enrolled from 2017 to 2018. Sci Rep. 2021;11:22979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lynch E, Jensen TO, Assao B, Chihana M, Turuho T, Nyehangane D, et al. Evaluation of HRP2 and pLDH-based rapid diagnostic tests for malaria and prevalence of pfhrp2/3 deletions in Aweil, South Sudan. Malar J. 2022;21:261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Mihreteab S, Anderson K, Pasay C, Smith D, Gatton ML, Cunningham J, et al. Epidemiology of mutant Plasmodium falciparum parasites lacking histidine-rich protein 2/3 genes in Eritrea 2 years after switching from HRP2-based RDTs. Sci Rep. 2021;11:21082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Parr JB, Anderson O, Juliano JJ, Meshnick SR, Streamlined. PCR-based testing for pfhrp2- and pfhrp3-negative Plasmodium falciparum. Malar J. 2018;17:137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kaaya RD, Kavishe RA, Tenu FF, Matowo JJ, Mosha FW, Drakeley C, et al. Deletions of the Plasmodium falciparum histidine-rich protein 2/3 genes are common in field isolates from north-eastern Tanzania. Sci Rep. 2022;12:5802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Leonard CM, Assefa A, McCaffery JN, Herman C, Plucinski M, Sime H, et al. Investigation of Plasmodium falciparum pfhrp2 and pfhrp3 gene deletions and performance of a rapid diagnostic test for identifying asymptomatic malaria infection in northern Ethiopia, 2015. Malar J. 2022;21:70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Watson OJ, Slater HC, Verity R, Parr JB, Mwandagalirwa MK, Tshefu A, et al. Modelling the drivers of the spread of Plasmodium falciparum hrp2 gene deletions in sub-Saharan Africa. eLife. 2017;6:e25008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Rogier E, McCaffery JN, Nace D, Svigel SS, Assefa A, Hwang J, et al. Plasmodium Falciparum pfhrp2 and pfhrp3 gene deletions from persons with symptomatic malaria infection in Ethiopia, Kenya, Madagascar, and Rwanda. Emerg Infect Dis. 2022;28:608–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Houzé S, Boly MD, Le Bras J, Deloron P, Faucher J-F. Pf HRP2 and pf LDH antigen detection for monitoring the efficacy of artemisinin-based combination therapy (ACT) in the treatment of uncomplicated falciparum malaria. Malar J. 2009;8:211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Grignard L, Nolder D, Sepúlveda N, Berhane A, Mihreteab S, Kaaya R, et al. A novel multiplex qPCR assay for detection of Plasmodium falciparum with histidine-rich protein 2 and 3 (pfhrp2 and pfhrp3) deletions in polyclonal infections. EBioMedicine. 2020;55:102757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Apinjoh TO, Ouattara A, Titanji VPK, Djimde A, Amambua-Ngwa A. Genetic diversity and drug resistance surveillance of Plasmodium falciparum for malaria elimination: is there an ideal tool for resource-limited sub-saharan Africa? Malar J. 2019;18:217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Nolder D, Stewart L, Tucker J, Ibrahim A, Gray A, Corrah T, et al. Failure of rapid diagnostic tests in Plasmodium falciparum malaria cases among travelers to the UK and Ireland: identification and characterisation of the parasites. Int J Infect Dis. 2021;108:137–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Thang ND, Rovira-Vallbona E, Binh NTH, Dung DV, Ngoc NTH, Long TK, et al. Surveillance of pfhrp2 and pfhrp3 gene deletions among symptomatic Plasmodium falciparum malaria patients in Central Vietnam. Malar J. 2022;21:371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kaaya RD, Matowo J, Kajeguka D, Tenu F, Shirima B, Mosha F, et al. The impact of submicroscopic parasitemia on malaria rapid diagnosis in Northeastern Tanzania, an area with diverse transmission patterns. Infect Disease Rep. 2022;14:798–809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Markwalter CF, Mudenda L, Leelawong M, Kimmel DW, Nourani A, Mbambara S, et al. Evidence for histidine-rich protein 2 immune complex formation in symptomatic patients in Southern Zambia. Malar J. 2018;17:256. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data generated or analysed during this study are included in this published article [and its supplementary information files].