

Abstract
Respiratory syncytial virus (RSV) is a major global pathogen, causing lower respiratory tract disease in at-risk populations including young children. Antibodies form a crucial layer of protection from RSV disease, particularly in immunologically naïve infants. Such antibodies are derived from the mother via transplacental transfer and breast milk, but may be particularly low in high-risk infants such as those born preterm. Maternally derived antibodies can now be supplemented by the administration of anti-RSV monoclonal antibodies, while a rising wave of maternal and paediatric vaccine strategies are approaching. The implementation of these prophylactics may profoundly decrease the healthcare burden of RSV. In this article, we review the role of antibody-mediated immunity in protecting children from RSV. We focus on maternally derived antibodies as the main source of protection against RSV and study factors that influence the scale of this transfer. The role of passive and active prophylactic approaches in protecting infants against RSV are discussed and knowledge gaps in our understanding of antibody-mediated protection against RSV are identified.
Shareable abstract
Antibodies play a major role in protecting children against severe RSV infections. Here, we review the transfer of antibodies from mother to infant and the relative roles of different antibody isotypes and specificities. https://bit.ly/3AgS8Fa
Introduction
Human respiratory syncytial virus (RSV) is the leading cause of bronchiolitis in young infants, with those <6 months old at highest risk of severe lower respiratory tract infection (LRTI) [1–3]. At this young age, the developing immune system may mount ineffective responses following RSV exposure [4], resulting in an ongoing need for protective therapies against RSV [4–6]. RSV is an RNA virus, composed of 10 genes expressing 11 proteins including fusion (F), the attachment glycoprotein (G) and nucleoprotein (N) [7]. F and G are surface proteins involved in viral entry of host cells, and key targets of neutralising antibodies [8, 9]. High antibody titres against these proteins are associated with reduced RSV-LRTI severity [10]. The two subtypes of RSV (RSV-A and RSV-B) are serologically distinct due to G protein sequence diversity (Ga and Gb, respectively), while the F protein is highly conserved [7, 11]. Most children are infected with RSV in the first year of life and virtually all by 2 years of age [12]. Hospitalisation rates peak at 2 to 3.9 months after birth [13]; about 1.8% of healthy term infants in high-income settings require hospitalisation because of RSV and 5.5% of these lead to intensive care unit admissions [2]. RSV thus poses a major healthcare burden in young children.
Vaccine development for RSV has been slow [14–20]. Early trials involving formalin-inactivated RSV enhanced disease severity [21, 22], resulting in caution in further attempts at vaccination [23, 24]. Enhanced respiratory disease (ERD) has been related to low-affinity antibodies and T-helper 2 biased immune responses observed in some animal studies, with antibodies contributing to ERD by altering innate immune responses and facilitating RSV uptake by monocytes [23, 25]. Neutralising antibodies alone (e.g. immunoprophylaxis with anti-F monoclonal antibodies such as palivizumab (Synagis®) and nirsevimab (Beyfortus™)) are sufficient to protect infants against LRTI [26, 27], indicating that induction of neutralising antibodies is a vital function of vaccine candidates. Induction of neutralising antibodies has been aided by structural analyses that identified major conformational differences between pre- and post-fusion (“pre-F” and “post-F”) formations of the RSV-F protein. While pre-F and post-F share many antigenic sites, such as site II that is targeted by palivizumab, those that are unique to pre-F (such as site Ø that is targeted by nirsevimab) are superior targets for neutralisation [28, 29]. These advances in rational structure-based pre-F antigen design have enabled the RSV vaccines and monoclonal antibodies now in development and licensure.
This review summarises recent literature on the humoral immune response following a childs’ first RSV infection and the influence of maternal antibodies. Given recent advances, we also explore the effect of immunoprophylaxis and maternal vaccination on humoral immunity and protection.
Results
We followed a literature search strategy (see supplementary material) to identify manuscripts for inclusion. This identified the antibody isotypes IgG and IgA and their antigen specificities to be of fundamental importance to the subject, and these are presented first in this review. This is followed by a discussion of literature pertaining to maternal antibody transfer and the protection they provide against RSV. Finally, we present current data on prophylactic strategies aiming at improving protection for children, including vaccination and immunoprophylaxis.
Anti-RSV antibodies in infants and adults
In seropositive adults, RSV infection boosts antibody titres via the reactivation of memory B-cells, resulting in elevated anti-RSV immunoglobulin (Ig)M, IgG, and IgA titres [30–32]. Following infection, strong IgG and IgA responses are found to both F and G proteins, but also to N [8, 31, 33]. High titres of anti-RSV IgG and IgA during infection are associated with reduced severity [8, 31]. Anti-RSV IgE was once proposed to predispose individuals to atopy or the development of wheeze and asthma [34, 35], but the existence of anti-RSV IgE and this association have now been refuted [36]. The following sections summarise the literature on the contribution of anti-RSV IgG and IgA to immunity.
IgG
IgG is the main immunoglobulin circulating in adult plasma (∼75% of all antibodies) [37]. Variations in heavy chains and hinge regions define four IgG subclasses (IgG1–4), which all bind antigen but differ in their effector functions [37]. During infection, anti-RSV IgGs bind both F and G [8, 31, 33]; however, neutralising IgGs typically bind epitopes unique to the pre-fusion conformation of F (pre-F) which block virion fusion with target cells [38]. Serum neutralisation titres typically correlate closely with anti-F antibodies (particularly pre-F) [9, 38] with IgG making a dominant contribution due to its abundance in blood; however, diverse isotypes and antigen specificities may also contribute to neutralisation.
Analysing blood IgG titres between acute and convalescent samples revealed that the strongest responses are seen at 13–18 months of life [39]. While boosting of IgG following infection or reinfection was observed in all age groups, the subclass of IgG boosted differs markedly between age groups, where infants induce IgG3 responses absent in adults [39]. As IgG3 has the strongest capacity to induce Fc-effector functions, such as phagocytosis and complement deposition, this age-associated shift in IgG subclasses induced by infection may alter the functions of anti-RSV antibodies over the life course [40]. Indeed, serum from 24-month-old children has been reported to induce weaker natural killer cell activation than that of adults after accounting for differences in antibody titres [41]. At 2–3 weeks post-infection, increases in anti-F IgG are weaker in 0–3-month-old infants relative to 4–6-month-olds, which in turn are weaker than those aged 7–12 months and adults [33, 39, 42]. The plateauing of the blood IgG response to RSV infection from 7 months suggests that this immune response has achieved maturation at this age. IgG1 and IgG3 follow this pattern, whereas IgG2 levels stay low from birth until 19–36 months before increasing until adulthood [39].
IgG titres in blood gradually wane following birth or RSV infection, with one report demonstrating that anti-G and anti-F IgG in RSV-infected 2–4-month-old infants are detected in the serum for 4 months (figure 1) [31]. Together, these reports demonstrate that IgG titres are boosted throughout life due to repeated infections, giving rise to repeated cycles of antibody rising and waning [13, 31], though the IgG subclasses and Fc-mediated effector functions of these repeatedly induced IgGs may change over the life course.
FIGURE 1.
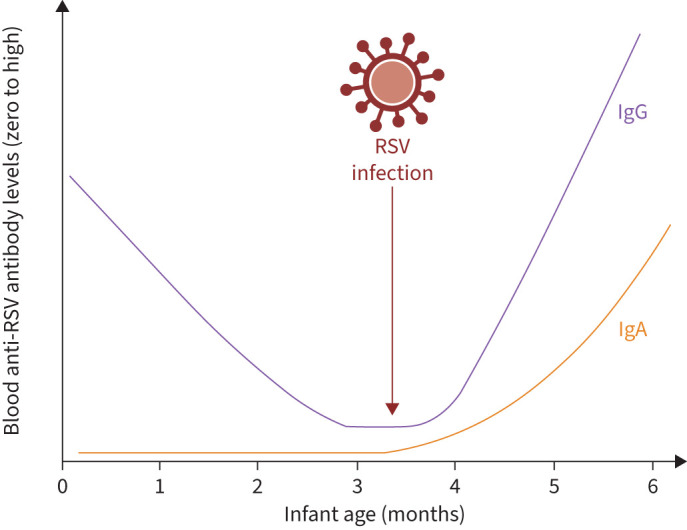
Infant blood anti-respiratory syncytial virus (RSV) IgG and IgA titres. Maternally derived IgG is evident in the infant's blood from birth but declines rapidly to undetectable levels around 4 months of age. Following RSV infection, de novo antibody responses occur in infants older than 1–2 months of life, resulting in IgG and IgA titre rises. Created with Biorender.com.
IgA
Monomeric IgA is present in circulation, but IgA is more abundant in mucosal surfaces in a dimeric or multimeric form where it blocks pathogen entry [37]. Variations in hinge regions give rise to two IgA subclasses, IgA1 and IgA2, with IgA2 more frequent in mucosal surfaces whereas IgA1 forms ∼90% of total circulating IgA [37].
IgA induction following RSV infection wanes within 6–12 months in adults and possibly within 2 months in children <2 years [32]. The kinetics of this response may differ from other isotypes [30, 43]. In infants and toddlers aged 0–16 months, anti-RSV IgA are present in nasopharyngeal secretions 2 weeks post-infection [44]. The timing of this nasal IgA response is similar in adults [30, 33], while IgG responses in adults may occur earlier [33]. While these reports support the principle of differing IgA and IgG response kinetics, these studies have not used consistent definitions of infection or methodologies, limiting cross-interpretation [32, 33, 43, 44]. Controlled human infection model studies determined that the robust mucosal IgA responses to infection seen in young adults were absent in older adults, despite similar blood IgG responses [45]. Given the prominent role of mucosal IgA in protection from infection in these models [30], these results indicate that older adults may gain limited protection from each RSV infection and so experience more frequent reinfections [45]. The mechanism behind this weakened IgA response in older adults is unclear, but indicates that the antibody isotypes and subclasses induced by infection and potentially vaccination may change with advancing age.
Differences in mucosal IgA and blood IgG responses to infection in some age groups may indicate differences in memory B-cells expressing different immunoglobulin isotypes [45]. F-specific IgA+ memory B-cells were present in blood samples from some adults at the end of the RSV season [31], but not after experimental infections despite increases in nasal IgA [30]. By contrast, F-specific IgA memory B-cells were not detected in blood from infants or children following RSV infection [31]. Like IgG, IgA may have a protective role, as RSV-infected adults with shorter infection duration (<2 weeks) have higher mucosal anti-G IgA titres [33].
Together, these studies show that IgA, particularly in the mucosa, can protect against RSV infections but that the induction of IgA after RSV infection may be dysregulated, particularly in older adults.
Maternal antibodies
Transplacental transfer of anti-RSV antibodies
To mediate protection, maternal antibodies must first reach the infant's circulation. IgG transfer from maternal to fetal blood depends on crossing both the multinucleated placental syncytiotrophoblasts and fetal capillary endothelial cells [46]. Maternal IgGs are endocytosed by syncytiotrophoblasts, bind neonatal Fc receptors lining the endosome and are transported basolaterally for release into the fetal bloodstream [46].
IgG1 is considered the most transplacentally transferred antibody relative to other isotypes and IgG subclasses, with cord:maternal blood ratios of ∼1.5 [39, 47]. RSV-specific antibodies are similarly transferred, with reports showing cord:maternal ratios between 1.03 and 1.22 [13, 48–50]. Variance in these ratios could be explained by the differing study populations or methodologies employed. Anti-RSV IgG titres decline after birth [39, 51]. The levels of maternally derived IgG required for protection against RSV LRTI is unclear, but this antibody transfer is believed to mediate protection of neonates [48].
The time of gestation when IgG transfer starts is unclear; some studies indicate it begins at 13 weeks’ gestation but only becomes sufficient at 36 weeks, whereas others suggest it begins at 28 weeks [52, 53]. This discrepancy could be explained twofold. Firstly, Fc receptors that mediate transplacental transfer are not abundantly expressed until 26 weeks’ gestation [53]. Secondly, IgG transfer is impeded before 26 weeks by cytotrophoblasts residing under the syncytiotrophoblast layer in the placenta [53]. The timing of mother-to-fetus IgG transfer implies that infants born preterm are vulnerable to RSV, partly due to deficient IgG transfer [54, 55]. In contrast, anti-pre-F IgG transfer does not differ between infants with low birth weight and those with normal birth weight for their gestational age [55].
Infants remain seropositive for maternal anti-RSV antibodies until ∼4–6 months after birth [13, 48, 56, 57]; however, maternal antibodies have a half-life of ∼1 month [13, 49, 53, 56]. Variations in maternal antibody duration between studies could reflect differences in population ethnicity, comorbidities or RSV seasonality. In fact, neutralising antibody half-life in high-income countries (HICs) is ∼1 month whereas in the low- and middle-income countries (LMICs) Kenya and Bangladesh it was ∼2 months, although such differences require formal testing [49, 58]. The decay from seropositivity at birth to seronegativity has been reported at 4.5–4.7 months in two Kenyan cross-sectional studies [58, 59]. However, we must consider that this could be an overestimation of maternal antibody duration, as infections in cross-sectional studies can give the impression of prolonged durability [51]. Birth cohort studies with active surveillance for RSV infection are needed to definitively answer this question. Overall, the evidence indicates that in term-born infants, RSV-specific maternal antibodies start to decline by 2 months of life, reaching seronegativity around 6 months. Far less is known about the presence and durability of these antibodies in preterm infants.
How protective are maternal antibodies?
High cord:maternal blood IgG ratios indicate successful maternal IgG transfer, but the actual amount transferred likely determines protection from RSV disease. One report demonstrated that infants with neutralising antibodies inhibitory concentrations >239 and >60 against RSV-A and RSV-B, respectively, were ∼fourfold more likely to be protected against RSV [60]. Maternal antibodies are associated with protection against RSV hospitalisation in infants <3 months old [10, 49]. Higher maternal serum neutralising antibody titres are associated with lower risk of RSV infection [49], while estimated serum IgG titres at birth were associated with delayed time to first RSV infection [61]. Maternally derived blood anti-pre-F IgG and neutralising antibodies are considered to protect infants from RSV hospitalisation, as lower serum pre-F IgG titres were observed in mothers with infants hospitalised with RSV than those with nonhospitalised infants (23.9 (range 1.4–273.7) µg·L−1 and 30.6 (3.4–220.0) µg·L−1, respectively, p=0.003) [10]. Infants with a serum neutralising antibody titre of 8.0 log2 were three times more likely to be protected from RSV hospitalisation than those with titres <8.0 log2. Together, such studies raise the possibility of establishing a serological correlate of protection for RSV (figure 2) [62, 63].
FIGURE 2.

Maternal antibody transfer in preterm, low- and high-titre scenarios. The scale of maternal antibody transfer profoundly influences the scale of antibody transfer to infants. In the scenario of preterm birth, transplacental antibody transfer may be low, resulting in low titres of anti-respiratory syncytial virus (RSV) antibodies in infants and high probability of susceptibility to severe RSV from birth. In maternal low-titre scenarios with term births, transplacental transfer does occur, but may quickly decay below a protective threshold. The amount of antibody needed to achieve this protective threshold is unknown and may be influenced by demographic and environmental factors. In high-titre scenarios, robust transplacental antibody transfer occurs, possibly owing to recent maternal RSV infection or vaccination. These high titres provide a longer window of protection against susceptibility to severe RSV infection. FcRN: neonatal Fc receptor. Created with Biorender.com.
However, some studies refute the possibility of defining a protective threshold. One study of 3646 mother–infant pairs in Nepal found no correlation between cord blood maternal antibody titres and the infant's age at primary RSV infection, suggesting that maternal antibodies did not delay primary RSV infection [50]. Another study reported no significant difference in antibody titres at birth between infants hospitalised with RSV and nonhospitalised, apparently uninfected, controls, though the sample size of 30 RSV cases and 60 controls was small [64]. These studies were conducted in LMIC settings and RSV severity and hospitalisation may be influenced by factors aside from maternal antibodies. An Australian birth cohort study reported that each 10-fold increase in cord blood anti-RSV neutralising antibodies was associated with a 37% decreased risk of RSV-LRTI at 12–24 months of age [65]. This study also reported that higher cord blood RSV neutralising antibody titres were associated with enhanced risk of non-RSV LRTI in 6–12-month-old infants [65]. This result suggested that maternal anti-RSV antibodies may be adversely associated with immunity to other pathogens, but our review found no studies replicating this result. A direct role of anti-RSV antibodies in this association may be unlikely as maternal antibodies have been consistently demonstrated to decay toward seronegativity by 6 months [33, 48, 49, 56–59]. Anti-pre-F IgG in breast milk has also been associated with protection, as mothers of RSV-infected infants aged <6 months had lower breast milk IgG levels than uninfected infants [66]. However, this study did not measure antibody levels in the infants; therefore, the extent of transfer of breast milk antibodies to the infant's plasma is uncertain. Some studies suggest that the presence of maternal antibodies against RSV in infants aged <6 months interferes with the infant's ability to develop a humoral response to RSV, explaining the infant's weak de novo response, though immunological immaturity in infants could also mediate this effect [6, 42, 53, 59].
Many studies highlight the importance of neutralisation as the main correlate of protection for maternally derived antibodies [10, 49, 61, 67]. While many studies report anti-RSV IgG titres, the antigenic targets of these antibodies are crucial. In accordance with the central role of anti-F antibodies in mediating neutralisation, one study reported that IgG anti-N titres in cord blood were not associated with protection from severe LRTI in infants <3 months old [67]. Live virus assays are the gold standard for determining neutralisation, but modern antigens such as pre-F may offer a higher-throughput surrogate [51]. Finally, mucosal antibodies that may be most likely to confer protection from infection [30] are not commonly studied, nor are the subclasses or Fc-dependent effector functions. Together, a majority of studies support a protective role for maternally derived antibodies against hospitalisation with RSV, but less evidence supports protection against acquiring RSV infection. Prevention of disease and prevention of infection may, therefore, be mediated by distinct immunological parameters.
Enhancing protection of infants
Immunoprophylaxis
Infants generate a poor immune response to RSV infection, including a weakened and short-lived memory response, leaving them at risk of severe RSV infection and hindering vaccine development [3, 5, 6]. Passive immunisation approaches have therefore been developed to protect these at-risk infants.
Passive immunoprophylaxis was first achieved with polyclonal RSV hyperimmune globulin (RSV-IGIV) (RespiGam®) [68]. RSV-IGIV prophylaxis trials in groups of at-risk infants demonstrated a 41% risk reduction of RSV-LRTI hospitalisation in infants with bronchopulmonary dysplasia (BPD) or preterm birth [69] and a 31% risk reduction in children with congenital heart disease [70]. The benefit of RSV-IVIG in this latter study was most pronounced in infants <6 months old, where a 58% risk reduction was observed [70]. Contrary to this success as a prophylactic, RSV-IVIG given to previously healthy children during hospitalisation with RSV-LRTI yielded little therapeutic benefit [71]. Together, these studies demonstrated that prophylaxis of at-risk infants could decrease the burden of RSV disease [68].
Coupling the success of RSV-IVIG with an increasing understanding of the importance of anti-F antibodies in neutralisation of RSV led to the development of palivizumab, an anti-F monoclonal IgG with potent in vitro and in vivo activity [72]. Trials of palivizumab in high-risk infants with BPD or preterm birth resulted in a 55% reduction in RSV-LRTI hospitalisation [73]. This was most pronounced in infants born at ≤35 weeks gestation, where a 78% reduction in RSV-LRTI hospitalisation was observed [73]. These studies confirmed the value of anti-RSV prophylaxis, and experimentally demonstrated the protection afforded by systemically delivered IgG anti-RSV-F [68]. The efficacy of palivizumab led to its use in HICs to protect at-risk infants, including preterm infants and young children (≤24 months of age) with haemodynamically significant congenital cardiac disease and/or chronic lung disease. However, meta-analysis of palivizumab cost-effectiveness studies reported that one 100 mg vial cost between USD 904 and 1866 [74], while the US Department of Veteran Affairs lists the maximum price per 100 mg palivizumab dose as USD 2487 (with five doses needed per RSV season) relative to USD 1691 for a 100 mg nirsevimab dose [75]. While considerable variation was noted between studies based on study population and methodology, the cost-effectiveness of palivizumab for preterm infants was estimated at between USD 5188 and 791 265 per quality-adjusted life-year (with 90% of estimates USD <50 000) [74]. These costs proved prohibitive for palivizumab use in many LMICs and limited use to high-risk infants in HICs [74].
With the aim of increasing efficacy and reducing costs, the next-generation monoclonal antibody for RSV immunoprophylaxis, nirsevimab, was developed [76]. Nirsevimab targets the highly conserved antigenic site Ø of RSV pre-F and has >50-fold-higher neutralisation activity than palivizumab [76]. Coupled with substitution of three amino acids in the antibody Fc region (M252Y/S254 T/T256E; “YTE”) that prolong the in vivo half-life to approximately 71 days, this aimed to result in a prophylactic that could be administered as a single-dose, compared to the 5 monthly doses required for palivizumab [74, 76].
Trials in preterm infants demonstrated 78% reduction in RSV-LRTI hospitalisation following a single nirsevimab dose [77]. This was followed by two large-scale multi-centre phase 3 studies; MELODY (1490 healthy infants born ≥35 weeks gestational age with a primary end-point of medically attended RSV associated LRTI) and HARMONIE (8058 infants born ≥29 weeks gestational age with a primary end-point of hospitalisation for RSV-LRTI). In healthy term-born infants, MELODY reported a 62% reduction in RSV-LRTI hospitalisation [78]. Similarly, HARMONIE reported an 83% reduction in RSV-LRTI hospitalisation in infants <12 months old [79]. Together, these studies support the efficacy of nirsevimab in preventing RSV-LRTI. In MELODY, nirsevimab administration increased serum RSV neutralising antibody titres >140-fold at day 31, relative to baseline [80], and titres remained >50-fold above baseline at day 151 [76, 80]. Nirsevimab administration was not associated with a decreased rate of IgG seroconversion to the post-F conformation of RSV-F [80], indicating that RSV infections still occur in nirsevimab-recipient infants but are less severe. Since approval for use in the EU, a real-world population cohort study of 1177 infants determined that nirsevimab was 88.7% effective at preventing RSV hospitalisation [81]. Together, these clinical trials and real-world studies support the effectiveness of immunoprophylaxis in preventing RSV disease in infants.
One theoretical risk of widespread prophylaxis using nirsevimab is selection pressure-led accumulation of binding site mutations that result in escape from neutralisation. To understand the impact of binding site mutations, a metanalysis of 5675 RSV sequences demonstrated that of the 25 amino acids in the nirsevimab binding site, 25 and 22 were highly conserved in RSV-A and RSV-B, respectively [82]. One prevalent RSV-B binding site polymorphism (Ile206Met:Gln209Arg) was still neutralised by nirsevimab, suggesting that while binding site mutations exist and should be monitored, these may have minimal impact on nirsevimab efficacy [82].
In anticipation of nirsevimab licensure, one modelling study estimated the purchasing price per dose of nirsevimab would need to be less than GBP 63 to be cost effective to immunise all infants ahead of the RSV season [83]. Nirsevimab was licensed in the UK and EU in November 2022 and approved by the US Food and Drug Administration in July 2023. Together, immunoprophylactic therapies experimentally confirmed that RSV disease could be prevented by conferring neutralising antibodies to infants in the first months of life, paving the way for next-generation vaccination strategies.
Maternal vaccination
Maternal vaccination confers passive immunity to infants through transplacental transfer of maternal IgG against F, N and G during the third trimester and via breast milk after birth [13, 48–50, 52, 60]. Maternal antibody transfer underpins the success of maternal vaccines, so the quantity of anti-RSV IgG and the time-point of gestation at which IgG is transferred to the fetus are important considerations for maternal vaccine development [84, 85]. As previously described, maternal anti-RSV IgG starts to decline by 2–4 months, with most infants becoming seronegative by 6 months [13, 50, 59]. An infant's immune system is continually maturing, so responses to pathogens and active immunisations in the first months of life may not elicit effective immune responses [19, 50]. This leaves infants vulnerable to severe RSV infection, highlighting the urgency of boosting immunity at birth [4, 5, 13]. Maternal vaccination aims to provide protection against RSV to the infant at birth and prevent severe RSV disease during the first months of life [5] (figure 2). This could also protect the mother, blocking one route of transmission to the infant [10].
Neutralising maternal IgG boosted by RSV vaccine candidates are successfully transplacentally transferred to infants [16–18, 20]; however, like natural infection-derived antibodies, antibody transfer ratios were variable among vaccinees. The Novavax recombinant RSV-F protein nanoparticle maternal vaccine candidate boosted maternal antibody concentrations and demonstrated a transplacental transfer ratio of 1.1–1.2 in infants born >30 days after maternal immunisation versus 0.6–0.8 in those born <30 days after immunisation [16]. This highlights the need to carefully plan maternal immunisation schedules to achieve maximal antibody transfer and benefit. The Pfizer RSV pre-F protein vaccine Abrysvo™ resulted in transfer ratios of 1.41–2.10 [18]. This ratio was higher than the transfer of natural infection induced antibodies, suggesting that transfer after vaccination may be particularly efficient.
One phase I/II study (NCT03674177) of the GSK pre-F3 vaccine demonstrated that a single dose in nonpregnant women increased anti-pre-F IgG neutralising titres 8–14-fold (depending on vaccine dose) 1 week after vaccination, which remained 5–6-fold higher 3 months post-vaccination [84]. The authors acknowledge that they did not account for RSV infections during the study, which may account for increases in antibody titres and confound determination of vaccine immunogenicity. Following this success, a maternal vaccination trial was performed (NCT04126213), showing similar increases in neutralising antibody titres [20]. Transplacental transfer ratios were high (1.62 in the 60 μg dose arm) and maternal antibody titres remained elevated for 6 months [20]. These remaining anti-RSV IgG maternal antibodies may continue to transfer to the infant via breast milk and provide additional protection [66]. The relative vaccine efficacy between breast-fed and nonbreast-fed infants was not reported in these studies but warrants investigation. A phase III trial (NCT04605159) of this vaccine stopped early after an increased rate of preterm birth was observed in vaccinated mothers, relative to placebo (relative risk 1.37) [86].
One Novavax vaccine candidate study demonstrated an efficacy of 44% in reducing RSV-LRTI, 35% in reducing RSV-LRTI hospitalisation and 47% in reducing severe hypoxaemia in the first 3 months of life, although these reported efficacies did not achieve significance in their primary end-point (LRTI in infants after 3 months of birth) [17].
Trials of the Pfizer RSV pre-F maternal vaccine (phase IIb NCT04032093 and phase III NCT04424316) demonstrated reduced risk of RSV-LRTI hospitalisation in the first 6 months of life [18, 19]. The phase IIb trial showed a vaccine efficacy of 91.5% against severe RSV-associated LRTI in the first 6 months of life and 81.8% (99.5% CI; 40.6–96.3) and 69.4% (97.58% CI 44.3–84.1) within 3 and 6 months of life, respectively, in phase III [18, 19]. The decline of vaccine efficacy over time likely reflects waning of maternal antibodies. While efficacy appears promising for Pfizer's RSV vaccine, which has received regulatory approval, some safety concerns have arisen around the potential for increasing rates of preterm birth in vaccinees relative to placebo [87]. While such an effect has not been recapitulated in most studies, the possibility of these risks requires vigilant monitoring and post-marketing surveillance if licenced [88]. Such risks must be balanced against the impact of RSV, with potential risks and benefits communicated to trial participants and patients.
These studies raise important considerations on monitoring vaccine effectiveness. Firstly, paediatric RSV disease will be influenced by the scale of transplacental antibody transfer, which may be lower in LMICs than HICs and in preterm births [17, 18]. Similarly, maternal antibody transfer is less efficient in infants who are small or large for their gestational age; thus, vaccines may be less effective in these high-risk infants [48]. Secondly, breastfeeding may enhance effectiveness through continual provision of maternal antibody, representing a low-cost strategy for RSV prevention that could have a high impact in areas where breastfeeding rates are low [66, 89].
Overall, maternal vaccines have shown promise in successfully transferring maternal antibodies, reducing RSV-LRTI and hospitalisation within 6 months of life. Modelling has demonstrated that maternal vaccination could prevent 62–75% and 76–87% of paediatric intensive care unit admissions with RSV in the UK and Netherlands, respectively, and 29–48% of RSV-related in-hospital deaths worldwide in infants <12 months [90].
Paediatric vaccination
Direct vaccination is considered unlikely to protect infants, owing to the immaturity of immune responses in the first months of life when risk of severe RSV disease is highest [3, 5, 6]. However, passive immunisation only provides transient protection for the first months of life, leaving older infants and toddlers at risk. To meet this gap in immunisation, direct vaccination with a variety of platform technologies approaches are being trialled, including a chimpanzee adenovirus expressing the RSV F, N and M2-1 proteins (e.g. NCT02927873), mRNA (e.g. NCT04528719) and live-attenuated vaccines (e.g. NCT04909021) [91].
While many studies are still ongoing, or yet to report all data, there are promising demonstrations of immunogenicity with these approaches. In one study of 21 seronegative 6–24-month-old infants and toddlers, the nasally administered live-attenuated RSV D46/NS2/N/ΔM2-2-HindIII vaccine induced serum neutralising antibodies in 95% of infants [91]. Immunogenicity was coupled with a high rate of mild upper respiratory tract (URT) symptoms (76% in vaccinees versus 18% in placebo controls), suggesting that a balance between immunogenicity and reactogenicity may be anticipated. A meta-analysis of five live-attenuated RSV vaccine candidates across seven trials indicated that a ≥4-fold rise in serum neutralising antibody titres in response to these vaccines was associated with protection against medically attended RSV infections (odds ratio 0.26) [92]. Interestingly, this analysis found no association between either the detection or scale of vaccine shedding and protection. Given the stronger association between protection and nasal antibodies, relative to blood antibodies [30], it would be of value to study mucosal humoral immune responses in these live-attenuated vaccination studies.
Identifying RSV cases in vaccine efficacy trials
To identify RSV infections in infants enrolled in vaccine efficacy trials, studies typically perform molecular testing for viral RNA during symptomatic episodes [16, 18, 19]. This design captures relative rates of symptomatic disease but does not inform our understanding of paucisymptomatic or asymptomatic infections. Regular viral swab testing outside of symptomatic episodes is burdensome and, depending on frequency and sensitivity, may still miss infections. Serological testing provides an alternative route to capturing RSV infections. Indeed, studies of antibody responses after nirsevimab administration indicate equivalent infection rates between nirsevimab recipients and placebo controls, observed as equal IgG seroconversion to RSV post-F [80]. As maternal vaccination would induce both IgG and IgA against epitopes unique to pre-F (such as site Ø) and common with post-F (such as site 2), but only IgG is transferred transplacentally, infants’ anti-RSV IgA levels have been proposed as a biomarker of RSV infection [51] (figure 1).
One complication is that IgA could be transferred to infants via breast milk [66]. Maternal anti-RSV IgA antibodies may be present in the breast milk, but infant seropositivity depends on if, and how much, IgA is absorbed from the gut to the bloodstream. It has also been proposed that infants might aspirate breast milk into the URT, thus instilling maternal IgA into the airway [93]. Further work is needed to confirm whether infant blood and airway IgA can be maternally derived. Additionally, infections in the first months of life may not trigger antibody responses in infants, so serum IgA-based definitions of RSV infection history may be inaccurate in this age group [42].
Discussion
Summary of main findings
In this review, we explored the humoral response to RSV in infants and the role of maternally derived antibodies in protecting infants. We also explored the effects of prophylactics against RSV and serological methods for identifying RSV infections. A systematic approach was implemented by identifying literature on PubMed, which was supplemented with papers known to the authors. The literature supports the transfer of maternal anti-RSV antibodies to infants via the placenta during gestation [13, 17, 48, 49]. These antibodies remain for ∼4 months [31]. Despite active transfer, infants are not wholly protected from RSV infection by maternal antibodies [39]. This leaves them vulnerable to severe RSV infection in these periods [4, 5, 13]. Immunoprophylaxis with IVIG and monoclonal anti-RSV antibodies experimentally demonstrated the benefit that can be achieved by antibodies [68]. Maternal vaccines could protect the infant in this vulnerable period by increasing transfer of anti-RSV antibodies during pregnancy [4, 5, 13]. Maternal vaccine studies have directly evaluated maternal antibody transfer by comparing anti-RSV IgG titres between maternal and cord blood [13, 49, 50, 52]. These support the observation that maternal antibody is transferred to cord and infant blood [46, 48, 53]. Maternal vaccines have shown promise in reducing RSV incidence and severity, with the MATISSE maternal vaccine trial paving the way for the licensure of the Pfizer RSV vaccine (Abrysvo™) [18, 19]. Infant prematurity, low or high birth weight, HIV infection, malaria and hypergammaglobulinaemia are factors that have been associated with decreased maternal anti-RSV antibody transfer [48, 53, 54]. The effect of these factors on transplacental transfer of maternal antibodies, and hence potentially vaccine efficacy, should be further studied [53].
Interpretation of findings
A broad limitation of the literature is the variety of methods used, including enzyme-linked immunosorbent assays against numerous antigens and plaque-reduction neutralisation assays or microneutralisation assays to measure neutralising antibody titres [31, 39, 49]. This could explain the variability between studies in IgG and IgA titres measured in infancy, the transferred maternal antibody ratio, the durability of antibodies in infant serum and the timing of maternal antibody transfer during gestation. This highlights the need for standardised assays and antigens across studies, allowing easier inter-study comparison and confirmation of key humoral immune responses. Standardised assays would also help to clarify when in gestation maternal vaccines should be delivered for optimal antibody transfer, thereby providing the infant with maximal protection [84].
Further work is needed in investigating whether maternal antibodies influence the nature of the infant's antibody response when they are inevitably infected with RSV for the first time, whether infants of RSV-vaccinated mothers have sterilising immunity at birth or whether infants from RSV-vaccinated mothers simply get milder infections. Seroconversion to post-F in nirsevimab recipient infants indicates that immunoprophylaxis may not prevent RSV infection but does prevent disease [80]; however, the same cannot yet be assumed with maternal or infant vaccination. Changes in the infants’ immune response to RSV could be studied by observing whether antibody isotypes, targeted epitopes and effector functions differ following infection with or without maternal or infant vaccination and whether any changes in antibody responses become fixed for life. In particular if would be valuable to understand whether infants receiving monoclonal antibodies still form de novo antibody responses against those monoclonal-targeted epitopes during infections. An understanding of protective antibody titres and the rate of maternal antibody waning may indicate the most appropriate age for infant vaccinations, should these become available in the future.
Literature regarding the efficacy of protection of maternal antibodies against RSV infection primarily measure serum neutralising antibody titres, with serum neutralisation predominantly mediated by IgG [10, 49, 61, 67]. As opposed to IgG, IgA is not transplacentally transferred, thus making the latter antibody a potential marker in infants of vaccinated mothers to indicate whether the infant has been RSV-infected [51]. This may support vaccine testing as active surveillance for RSV infections by viral PCR is logistically challenging. Indeed, cross-sectional serosurveillance studies have demonstrated the low anti-RSV IgA titres at birth steeply elevate through the first months of life, likely as a result of RSV infections [51]. Notably, nasal mucosal anti-RSV IgA has been demonstrated to be protective against RSV infection, is correlated with reduced viral replication and has been suggested as a superior correlate of protection than serum neutralising antibody titres [30, 94]. Nevertheless, further work in measuring serum and nasal IgA in infants is required to establish the development of IgA responses at different stages of infancy. This would determine the age at which infants mount an IgA response and identify a threshold indicative of infection. Greater understanding of anti-RSV mucosal antibody responses could also influence the development of mucosal or intramuscular vaccines for infants and toddlers and identify which vaccine platforms elicit the most effective and durable immune responses in children, thus presenting new opportunities for another tier of protection against RSV disease at different ages.
We must also consider that IgA has been detected in colostrum and studies have identified that breastfeeding is a protective factor against severe RSV [66]. Whether this protection is due to antibody transfer or aspiration of milk IgA into the airway is unclear. Maternal IgA may also be transferred via amniotic fluid, providing mucosal immunity against RSV for at least 1 week [95]. Nevertheless, clinical outcomes such as decreasing RSV hospitalisation rates must remain the key objectives, rather than solely aiming for increased antibody responses as a primary end-point in vaccine trials [62].
Conclusion
Antibodies are key effectors of RSV immunity which prevent severe disease in infants. Maternal anti-RSV IgG is transferred to the infant during gestation and can protect infants in their first months of life. Infants <6 months are particularly vulnerable to severe RSV disease and maternal vaccine and passive immunisation approaches have been developed. Direct vaccination of infants and toddlers with mucosal vaccines could further add to the repertoire of prophylactics against RSV and add a crucial layer of mucosal immunity. This growing toolbox of prophylactics against RSV is set to revolutionise the management of this burdensome disease. Standardisation of humoral immune assays and antigens for key endpoints would allow direct comparisons between studies. The effects of immunoprophylaxis and vaccination on infant's de novo humoral responses to RSV warrants further study. Overall, the ability to prevent RSV disease is undergoing a revolution, but questions on therapeutic deployment, antibody durability and long-term effects remain.
Questions for future research
What factors determine the scale of maternal antibody transfer to infants?
Do maternal vaccines protect infants from RSV infection or just disease?
Is a child's de novo immunity to RSV altered by the presence of antibodies derived from maternal vaccination or immunoprophylaxis?
Are mucosal antibodies important in protection and are they induced by RSV prophylactics?
Can standardised assays for humoral immunity improve comparability between studies?
Can a protective threshold of neutralising antibodies be determined?
Do non-neutralising features of antibodies (e.g. Fc-dependent effector properties, non-F antigens) confer protection beyond that mediated by neutralising antibodies?
Supplementary material
Please note: supplementary material is not edited by the Editorial Office, and is uploaded as it has been supplied by the author.
Supplementary material ERR-0106-2024.SUPPLEMENT (396.9KB, pdf)
Acknowledgements
The PROMISE investigators are: Jeroen Aerssens, Benoit Callendret, and Gabriela Ispas (Janssen, Beerse, Belgium); Bahar Ahani (AstraZeneca, Gaithersburg, MD, USA); Jessica Atwell, Elizabeth Begier, Monica Turiga, and Tin Tin Htar (Pfizer, Paris, France); Mathieu Bangert, Rolf Kramer, and Charlotte Vernhes (Sanofi Pasteur, Lyon, France); Philippe Beutels (University of Antwerp, Antwerpen, Belgium); Louis Bont (University Medical Centre Utrecht, Utrecht, the Netherlands); Harry Campbell, Harish Nair, You Li, Sebastien Kenmoe, Richard Osei-Yeboah, and Xin Wang (University of Edinburgh, Edinburgh, UK); Rachel Cohen, Gael Dos Santos, Philip Joosten, and Theo Last (GSK, Wavre, Belgium); Veena Kumar (Novavax, Gaithersburg, MD, USA); Nuria Machin (Teamit Research, Barcelona, Spain); Hanna Nohynek (Finnish National Institute for Health and Welfare, Helsinki, Finland); Peter Openshaw (Imperial College London, London, UK); John Paget (Netherlands Institute for Health Services Research, Utrecht, the Netherlands); Andrew Pollard (University of Oxford, Oxford, UK); Anne Teirlinck (National Institute for Public Health and the Environment, Bilthoven, the Netherlands); Arantxa Urchueguía-Fornes, Ainara Mira-Iglesias, Alejandro Orrico-Sánchez, and Javier Díez-Domingo (Vaccine Research Department, FISABIO-Public Health and CIBER de Epidemiología y Salud Pública, Instituto de Salud Carlos III, Valencia, Spain); Johannesen Caroline Klint (Nordsjællands Hospital, Denmark); Mark Miller (School of Public Health and Community Medicine, Institute of Medicine, University of Gothenburg, Gothenburg, Sweden); Rafael Mikolajczyk (Institute for Medical Epidemiology, Biometry, and Informatics, Medical Faculty, Martin Luther University of Halle-Wittenberg, Halle, Germany); and Terho Heikkinen (Department of Pediatrics, University of Turku and Turku University Hospital, Turku, Finland).
Provenance: Submitted article, peer reviewed.
Disclaimer: This article reflects only the authors’ view, and the Joint Undertaking is not responsible for any use that may be made of the information it contains herein.
C. Vernhes has changed affiliation since the work was performed and is now employed by Vaccines Europe, Brussels, Belgium.
Conflict of interest: A. Moureau is an employee of Sanofi and owns Sanofi stock as an employee. C. Vernhes was an employee of Sanofi at the time the work was performed, owns Sanofi stock as a former employee and is currently employed by the trade association Vaccines Europe. P.J.M. Openshaw reports consulting fees from GSK, Moderna, Janssen, Seqirus, Pfizer, Sanofi, AstraZeneca, and Icosavax; honoraria for lectures from GSK, Moderna, Seqirus, Sanofi, and AstraZeneca; participation on a Data Safety Monitoring Board for Sanofi; board membership for the Science Media Centre and a role as governor for Sidcot School. R.S. Thwaites reports consulting fees from AOBiome, Gossamer Bio, Indalo Therapeutics, and PrEP Biopharm; and honoraria for lectures from AstraZeneca and GSK. All other authors report no conflicts of interest.
Support statement: This work is supported by the Preparing for RSV Immunisation and Surveillance in Europe (PROMISE) project, which has received funding from the Innovative Medicines Initiative 2 Joint Undertaking (grant 101034339); this Joint Undertaking receives support from the European Union's Horizon 2020 research and innovation program and the European Federation of Pharmaceutical Industries (EFPIA). This work is also supported by the Imperial Biomedical Research Centre (NIHR Imperial BRC, grant IS-BRC-1215-20013), the Health Protection Research Unit in Respiratory Infections at Imperial College London. P.J.M. Openshaw is supported by a NIHR Senior Investigator Award (award 201385). Funding information for this article has been deposited with the Crossref Funder Registry.
References
- 1.Shi T, McAllister DA, O'Brien KL, et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: a systematic review and modelling study. Lancet 2017; 390: 946–958. doi: 10.1016/S0140-6736(17)30938-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wildenbeest JG, Billard MN, Zuurbier RP, et al. The burden of respiratory syncytial virus in healthy term-born infants in Europe: a prospective birth cohort study. Lancet Respir Med 2023; 11: 341–353. doi: 10.1016/S2213-2600(22)00414-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Li Y, Wang X, Blau DM, et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in children younger than 5 years in 2019: a systematic analysis. Lancet 2022; 399: 2047–2064. doi: 10.1016/S0140-6736(22)00478-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lambert L, Sagfors AM, Openshaw PJ, et al. Immunity to RSV in early-life. Front Immunol 2014; 5: 466. doi: 10.3389/fimmu.2014.00466 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Esposito S, Abu Raya B, Baraldi E, et al. RSV prevention in all infants: which is the most preferable strategy? Front Immunol 2022; 13: 880368. doi: 10.3389/fimmu.2022.880368 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Eichinger KM, Kosanovich JL, Lipp M, et al. Strategies for active and passive pediatric RSV immunization. Ther Adv Vaccines Immunother 2021; 9: 2515135520981516. doi: 10.1177/2515135520981516 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Collins PL, Fearns R, Graham BS. Respiratory syncytial virus: virology, reverse genetics, and pathogenesis of disease. Curr Top Microbiol Immunol 2013; 372: 3–38. doi: 10.1007/978-3-642-38919-1_1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Capella C, Chaiwatpongsakorn S, Gorrell E, et al. Prefusion F, postfusion F, G antibodies, and disease severity in infants and young children with acute respiratory syncytial virus infection. J Infect Dis 2017; 216: 1398–1406. doi: 10.1093/infdis/jix489 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ngwuta JO, Chen M, Modjarrad K, et al. Prefusion F-specific antibodies determine the magnitude of RSV neutralizing activity in human sera. Sci Transl Med 2015; 7: 309ra162. doi: 10.1126/scitranslmed.aac4241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Koivisto K, Nieminen T, Mejias A, et al. Respiratory syncytial virus (RSV)-specific antibodies in pregnant women and subsequent risk of RSV hospitalization in young infants. J Infect Dis 2022; 225: 1189–1196. doi: 10.1093/infdis/jiab315 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cantu-Flores K, Rivera-Alfaro G, Munoz-Escalante JC, et al. Global distribution of respiratory syncytial virus A and B infections: a systematic review. Pathog Glob Health 2022; 116: 398–409. doi: 10.1080/20477724.2022.2038053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Coultas JA, Smyth R, Openshaw PJ. Respiratory syncytial virus (RSV): a scourge from infancy to old age. Thorax 2019; 74: 986–993. doi: 10.1136/thoraxjnl-2018-212212 [DOI] [PubMed] [Google Scholar]
- 13.Pasittungkul S, Thongpan I, Vichaiwattana P, et al. High seroprevalence of antibodies against human respiratory syncytial virus and evidence of respiratory syncytial virus reinfection in young children in Thailand. Int J Infect Dis 2022; 125: 177–183. doi: 10.1016/j.ijid.2022.10.039 [DOI] [PubMed] [Google Scholar]
- 14.Munoz FM, Piedra PA, Glezen WP. Safety and immunogenicity of respiratory syncytial virus purified fusion protein-2 vaccine in pregnant women. Vaccine 2003; 21: 3465–3467. doi: 10.1016/S0264-410X(03)00352-9 [DOI] [PubMed] [Google Scholar]
- 15.August A, Glenn GM, Kpamegan E, et al. A phase 2 randomized, observer-blind, placebo-controlled, dose-ranging trial of aluminum-adjuvanted respiratory syncytial virus F particle vaccine formulations in healthy women of childbearing age. Vaccine 2017; 35: 3749–3759. doi: 10.1016/j.vaccine.2017.05.045 [DOI] [PubMed] [Google Scholar]
- 16.Munoz FM, Swamy GK, Hickman SP, et al. Safety and immunogenicity of a respiratory syncytial virus fusion (F) protein nanoparticle vaccine in healthy third-trimester pregnant women and their infants. J Infect Dis 2019; 220: 1802–1815. doi: 10.1093/infdis/jiz390 [DOI] [PubMed] [Google Scholar]
- 17.Madhi SA, Polack FP, Piedra PA, et al. Respiratory syncytial virus vaccination during pregnancy and effects in infants. N Engl J Med 2020; 383: 426–439. doi: 10.1056/NEJMoa1908380 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Simoes EAF, Center KJ, Tita ATN, et al. Prefusion F protein-based respiratory syncytial virus immunization in pregnancy. N Engl J Med 2022; 386: 1615–1626. doi: 10.1056/NEJMoa2106062 [DOI] [PubMed] [Google Scholar]
- 19.Kampmann B, Madhi SA, Munjal I, et al. Bivalent prefusion F vaccine in pregnancy to prevent RSV illness in infants. N Engl J Med 2023; 388: 1451–1464. doi: 10.1056/NEJMoa2216480 [DOI] [PubMed] [Google Scholar]
- 20.Bebia Z, Reyes O, Jeanfreau R, et al. Safety and immunogenicity of an investigational respiratory syncytial virus vaccine (RSVPreF3) in mothers and their infants: a phase 2 randomized trial. J Infect Dis 2023; 288: 299–310. doi: 10.1093/infdis/jiad024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kim HW, Canchola JG, Brandt CD, et al. Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am J Epidemiol 1969; 89: 422–434. doi: 10.1093/oxfordjournals.aje.a120955 [DOI] [PubMed] [Google Scholar]
- 22.Kapikian AZ, Mitchell RH, Chanock RM, et al. An epidemiologic study of altered clinical reactivity to respiratory syncytial (RS) virus infection in children previously vaccinated with an inactivated RS virus vaccine. Am J Epidemiol 1969; 89: 405–421. doi: 10.1093/oxfordjournals.aje.a120954 [DOI] [PubMed] [Google Scholar]
- 23.Muralidharan A, Li C, Wang L, et al. Immunopathogenesis associated with formaldehyde-inactivated RSV vaccine in preclinical and clinical studies. Expert Rev Vaccines 2017; 16: 351–360. doi: 10.1080/14760584.2017.1260452 [DOI] [PubMed] [Google Scholar]
- 24.Mazur NI, Higgins D, Nunes MC, et al. The respiratory syncytial virus vaccine landscape: lessons from the graveyard and promising candidates. Lancet Infect Dis 2018; 18: e295–e311. doi: 10.1016/S1473-3099(18)30292-5 [DOI] [PubMed] [Google Scholar]
- 25.Openshaw PJ, Culley FJ, Olszewska W. Immunopathogenesis of vaccine-enhanced RSV disease. Vaccine 2001; 20: Suppl. 1, S27–S31. doi: 10.1016/S0264-410X(01)00301-2 [DOI] [PubMed] [Google Scholar]
- 26.Garegnani L, Styrmisdottir L, Roson Rodriguez P, et al. Palivizumab for preventing severe respiratory syncytial virus (RSV) infection in children. Cochrane Database Syst Rev 2021; 11: CD013757. 10.1002/14651858.CD013757.pub2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bergeron HC, Tripp RA. Breakthrough therapy designation of nirsevimab for the prevention of lower respiratory tract illness caused by respiratory syncytial virus infections (RSV). Expert Opin Investig Drugs 2022; 31: 23–29. doi: 10.1080/13543784.2022.2020248 [DOI] [PubMed] [Google Scholar]
- 28.Graham BS. The journey to RSV vaccines – heralding an era of structure-based design. N Engl J Med 2023; 388: 579–581. doi: 10.1056/NEJMp2216358 [DOI] [PubMed] [Google Scholar]
- 29.McLellan JS, Chen M, Joyce MG, et al. Structure-based design of a fusion glycoprotein vaccine for respiratory syncytial virus. Science 2013; 342: 592–598. doi: 10.1126/science.1243283 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Habibi MS, Jozwik A, Makris S, et al. Impaired antibody-mediated protection and defective IgA B-cell memory in experimental infection of adults with respiratory syncytial virus. Am J Respir Crit Care Med 2015; 191: 1040–1049. doi: 10.1164/rccm.201412-2256OC [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Green CA, Sande CJ, de Lara C, et al. Humoral and cellular immunity to RSV in infants, children and adults. Vaccine 2018; 36: 6183–6190. doi: 10.1016/j.vaccine.2018.08.056 [DOI] [PubMed] [Google Scholar]
- 32.Stensballe LG, Kofoed PE, Nante EJ, et al. Duration of secretory IgM and IgA antibodies to respiratory syncytial virus in a community study in Guinea–Bissau. Acta Paediatr 2000; 89: 421–426. doi: 10.1080/080352500750028122 [DOI] [PubMed] [Google Scholar]
- 33.Ye X, Iwuchukwu OP, Avadhanula V, et al. Humoral and mucosal antibody response to RSV structural proteins in RSV-infected adult hematopoietic cell transplant (HCT) recipients. Viruses 2021; 13: 991. doi: 10.3390/v13060991 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Smith-Norowitz TA, Mandal M, Joks R, et al. IgE anti-respiratory syncytial virus antibodies detected in serum of pediatric patients with asthma. Hum Immunol 2015; 76: 519–524. doi: 10.1016/j.humimm.2015.06.002 [DOI] [PubMed] [Google Scholar]
- 35.Welliver RC, Wong DT, Sun M, et al. The development of respiratory syncytial virus-specific IgE and the release of histamine in nasopharyngeal secretions after infection. N Engl J Med 1981; 305: 841–846. doi: 10.1056/NEJM198110083051501 [DOI] [PubMed] [Google Scholar]
- 36.Holt PG, Sly PD. Interactions between RSV infection, asthma, and atopy: unraveling the complexities. J Exp Med 2002; 196: 1271–1275. doi: 10.1084/jem.20021572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schroeder HW Jr, Cavacini L. Structure and function of immunoglobulins. J Allergy Clin Immunol 2010; 125: S41–S52. doi: 10.1016/j.jaci.2009.09.046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Huang J, Diaz D, Mousa JJ. Antibody epitopes of pneumovirus fusion proteins. Front Immunol 2019; 10: 2778. doi: 10.3389/fimmu.2019.02778 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Jounai N, Yoshioka M, Tozuka M, et al. Age-specific profiles of antibody responses against respiratory syncytial virus infection. EBioMedicine 2017; 16: 124–135. doi: 10.1016/j.ebiom.2017.01.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Vidarsson G, Dekkers G, Rispens T. IgG subclasses and allotypes: from structure to effector functions. Front Immunol 2014; 5: 520. doi: 10.3389/fimmu.2014.00520 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lakerveld AJ, Gelderloos AT, Schepp RM, et al. Difference in respiratory syncytial virus-specific Fc-mediated antibody effector functions between children and adults. Clin Exp Immunol 2023; 214: 79–93. doi: 10.1093/cei/uxad101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sande CJ, Cane PA, Nokes DJ. The association between age and the development of respiratory syncytial virus neutralising antibody responses following natural infection in infants. Vaccine 2014; 32: 4726–4729. doi: 10.1016/j.vaccine.2014.05.038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tsutsumi H, Matsuda K, Yamazaki H, et al. Different kinetics of antibody responses between IgA and IgG classes in nasopharyngeal secretion in infants and children during primary respiratory syncytial virus infection. Acta Paediatr Jpn 1995; 37: 464–468. doi: 10.1111/j.1442-200X.1995.tb03356.x [DOI] [PubMed] [Google Scholar]
- 44.Yamazaki H, Tsutsumi H, Matsuda K, et al. Effect of maternal antibody on IgA antibody response in nasopharyngeal secretion in infants and children during primary respiratory syncytial virus infection. J Gen Virol 1994; 75: 2115–2119. doi: 10.1099/0022-1317-75-8-2115 [DOI] [PubMed] [Google Scholar]
- 45.Ascough S, Dayananda P, Kalyan M, et al. Divergent age-related humoral correlates of protection against respiratory syncytial virus infection in older and young adults: a pilot, controlled, human infection challenge model. Lancet Healthy Longev 2022; 3: e405–e416. doi: 10.1016/S2666-7568(22)00103-9 [DOI] [PubMed] [Google Scholar]
- 46.Palmeira P, Quinello C, Silveira-Lessa AL, et al. IgG placental transfer in healthy and pathological pregnancies. Clin Dev Immunol 2012; 2012: 985646. doi: 10.1155/2012/985646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Clements T, Rice TF, Vamvakas G, et al. Update on transplacental transfer of IgG subclasses: impact of maternal and fetal factors. Front Immunol 2020; 11: 1920. doi: 10.3389/fimmu.2020.01920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yildiz M, Kara M, Sutcu M, et al. Evaluation of respiratory syncytial virus IgG antibody dynamics in mother–infant pairs cohort. Eur J Clin Microbiol Infect Dis 2020; 39: 1279–1286. doi: 10.1007/s10096-020-03841-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chu HY, Steinhoff MC, Magaret A, et al. Respiratory syncytial virus transplacental antibody transfer and kinetics in mother-infant pairs in Bangladesh. J Infect Dis 2014; 210: 1582–1589. doi: 10.1093/infdis/jiu316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Chu HY, Tielsch J, Katz J, et al. Transplacental transfer of maternal respiratory syncytial virus (RSV) antibody and protection against RSV disease in infants in rural Nepal. J Clin Virol 2017; 95: 90–95. doi: 10.1016/j.jcv.2017.08.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Berbers G, Mollema L, van der Klis F, et al. Antibody responses to respiratory syncytial virus: a cross-sectional serosurveillance study in the Dutch population focusing on infants younger than 2 years. J Infect Dis 2021; 224: 269–278. doi: 10.1093/infdis/jiaa483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mashiyama F, Hashimoto K, Norito S, et al. Neutralizing and epitope-specific antibodies against respiratory syncytial virus in maternal and cord blood paired samples. Viruses 2022; 14: 2702. doi: 10.3390/v14122702 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Nyiro JU, Bukusi E, Mwaengo D, et al. Efficiency of transplacental transfer of respiratory syncytial virus (RSV) specific antibodies among pregnant women in Kenya. Wellcome Open Res 2022; 7: 43. doi: 10.12688/wellcomeopenres.17636.2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Atwell JE, Lutz CS, Sparrow EG, et al. Biological factors that may impair transplacental transfer of RSV antibodies: implications for maternal immunization policy and research priorities for low- and middle-income countries. Vaccine 2022; 40: 4361–4370. doi: 10.1016/j.vaccine.2022.06.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kachikis AB, Rumfelt K, Pike M, et al. Transfer of respiratory syncytial virus prefusion F protein antibody in low birthweight infants. Open Forum Infect Dis 2024; 11: ofae314. doi: 10.1093/ofid/ofae314 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Arankalle VA, Kulkarni R, Malshe N, et al. Seroepidemiology of respiratory syncytial virus in western India with special reference to appropriate age for infant vaccination. J Med Virol 2019; 91: 1566–1570. doi: 10.1002/jmv.25489 [DOI] [PubMed] [Google Scholar]
- 57.Suara RO, Piedra PA, Glezen WP, et al. Prevalence of neutralizing antibody to respiratory syncytial virus in sera from mothers and newborns residing in the Gambia and in the United States. Clin Diagn Lab Immunol 1996; 3: 477–479. doi: 10.1128/cdli.3.4.477-479.1996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ochola R, Sande C, Fegan G, et al. The level and duration of RSV-specific maternal IgG in infants in Kilifi Kenya. PLoS One 2009; 4: e8088. doi: 10.1371/journal.pone.0008088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Nyiro JU, Kombe IK, Sande CJ, et al. Defining the vaccination window for respiratory syncytial virus (RSV) using age-seroprevalence data for children in Kilifi, Kenya. PLoS One 2017; 12: e0177803. doi: 10.1371/journal.pone.0177803 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Buchwald AG, Graham BS, Traore A, et al. Respiratory syncytial virus (RSV) neutralizing antibodies at birth predict protection from RSV illness in infants in the first 3 months of life. Clin Infect Dis 2021; 73: e4421–e4427. doi: 10.1093/cid/ciaa648 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Walsh EE, Wang L, Falsey AR, et al. Virus-specific antibody, viral load, and disease severity in respiratory syncytial virus infection. J Infect Dis 2018; 218: 208–217. doi: 10.1093/infdis/jiy106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Openshaw PJM. Using correlates to accelerate vaccinology. Science 2022; 375: 22–23. doi: 10.1126/science.abn0007 [DOI] [PubMed] [Google Scholar]
- 63.Piedra PA, Jewell AM, Cron SG, et al. Correlates of immunity to respiratory syncytial virus (RSV) associated-hospitalization: establishment of minimum protective threshold levels of serum neutralizing antibodies. Vaccine 2003; 21: 3479–3482. doi: 10.1016/S0264-410X(03)00355-4 [DOI] [PubMed] [Google Scholar]
- 64.Nyiro JU, Sande CJ, Mutunga M, et al. Absence of association between cord specific antibody levels and severe respiratory syncytial virus (RSV) disease in early infants: a case control study from coastal Kenya. PLoS One 2016; 11: e0166706. doi: 10.1371/journal.pone.0166706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Takashima MD, Grimwood K, Sly PD, et al. Cord-blood respiratory syncytial virus antibodies and respiratory health in first 5 years of life. Pediatr Pulmonol 2021; 56: 3942–3951. doi: 10.1002/ppul.25688 [DOI] [PubMed] [Google Scholar]
- 66.Mazur NI, Horsley NM, Englund JA, et al. Breast milk prefusion F immunoglobulin G as a correlate of protection against respiratory syncytial virus acute respiratory illness. J Infect Dis 2019; 219: 59–67. doi: 10.1093/infdis/jiy477 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Receveur M, Ottmann M, Reynes JM, et al. Level of maternal antibodies against respiratory syncytial virus (RSV) nucleoprotein at birth and risk of RSV very severe lower respiratory tract infection. Influenza Other Respir Viruses 2023; 17: e13025. doi: 10.1111/irv.13025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Wu H, Pfarr DS, Losonsky GA, et al. Immunoprophylaxis of RSV infection: advancing from RSV-IGIV to palivizumab and motavizumab. Curr Top Microbiol Immunol 2008; 317: 103–123. doi: 10.1007/978-3-540-72146-8_4 [DOI] [PubMed] [Google Scholar]
- 69.The PREVENT Study Group . Reduction of respiratory syncytial virus hospitalization among premature infants and infants with bronchopulmonary dysplasia using respiratory syncytial virus immune globulin prophylaxis. Pediatrics 1997; 99: 93–99. doi: 10.1542/peds.99.1.93 [DOI] [PubMed] [Google Scholar]
- 70.Simoes EA, Sondheimer HM, Top FH,Jr, et al. Respiratory syncytial virus immune globulin for prophylaxis against respiratory syncytial virus disease in infants and children with congenital heart disease. The Cardiac Study Group. J Pediatr 1998; 133: 492–499. doi: 10.1016/S0022-3476(98)70056-3 [DOI] [PubMed] [Google Scholar]
- 71.Rodriguez WJ, Gruber WC, Groothuis JR, et al. Respiratory syncytial virus immune globulin treatment of RSV lower respiratory tract infection in previously healthy children. Pediatrics 1997; 100: 937–942. doi: 10.1542/peds.100.6.937 [DOI] [PubMed] [Google Scholar]
- 72.Johnson S, Oliver C, Prince GA, et al. Development of a humanized monoclonal antibody (MEDI-493) with potent in vitro and in vivo activity against respiratory syncytial virus. J Infect Dis 1997; 176: 1215–1224. doi: 10.1086/514115 [DOI] [PubMed] [Google Scholar]
- 73.The Impact-RSV Study Group . Palivizumab, a humanized respiratory syncytial virus monoclonal antibody, reduces hospitalization from respiratory syncytial virus infection in high-risk infants. Pediatrics 1998; 102: 531–537. doi: 10.1542/peds.102.3.531 [DOI] [PubMed] [Google Scholar]
- 74.Mac S, Sumner A, Duchesne-Belanger S, et al. Cost-effectiveness of palivizumab for respiratory syncytial virus: a systematic review. Pediatrics 2019; 143: e20184064. doi: 10.1542/peds.2018-4064 [DOI] [PubMed] [Google Scholar]
- 75.US Department of Veterans Affairs . Pharmaceutical catalog search. Date last accessed: 24 July 2024. www.vendorportal.ecms.va.gov/NAC/Pharma/List
- 76.Zhu Q, McLellan JS, Kallewaard NL, et al. A highly potent extended half-life antibody as a potential RSV vaccine surrogate for all infants. Sci Transl Med 2017; 9: eaaj1928. doi: 10.1126/scitranslmed.aaj1928 [DOI] [PubMed] [Google Scholar]
- 77.Griffin MP, Yuan Y, Takas T, et al. Single-dose nirsevimab for prevention of RSV in preterm infants. N Engl J Med 2020; 383: 415–425. doi: 10.1056/NEJMoa1913556 [DOI] [PubMed] [Google Scholar]
- 78.Hammitt LL, Dagan R, Yuan Y, et al. Nirsevimab for prevention of RSV in healthy late-preterm and term infants. N Engl J Med 2022; 386: 837–846. doi: 10.1056/NEJMoa2110275 [DOI] [PubMed] [Google Scholar]
- 79.Drysdale SB, Cathie K, Flamein F, et al. Nirsevimab for prevention of hospitalizations due to RSV in infants. N Engl J Med 2023; 389: 2425–2435. doi: 10.1056/NEJMoa2309189 [DOI] [PubMed] [Google Scholar]
- 80.Wilkins D, Yuan Y, Chang Y, et al. Durability of neutralizing RSV antibodies following nirsevimab administration and elicitation of the natural immune response to RSV infection in infants. Nat Med 2023; 29: 1172–1179. doi: 10.1038/s41591-023-02316-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Ezpeleta G, Navascues A, Viguria N, et al. Effectiveness of nirsevimab immunoprophylaxis administered at birth to prevent infant hospitalisation for respiratory syncytial virus infection: a population-based cohort study. Vaccines (Basel) 2024; 12: 383. doi: 10.3390/vaccines12040383 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wilkins D, Langedijk AC, Lebbink RJ, et al. Nirsevimab binding-site conservation in respiratory syncytial virus fusion glycoprotein worldwide between 1956 and 2021: an analysis of observational study sequencing data. Lancet Infect Dis 2023; 23: 856–866. doi: 10.1016/S1473-3099(23)00062-2 [DOI] [PubMed] [Google Scholar]
- 83.Hodgson D, Koltai M, Krauer F, et al. Optimal respiratory syncytial virus intervention programmes using nirsevimab in England and Wales. Vaccine 2022; 40: 7151–7157. doi: 10.1016/j.vaccine.2022.10.041 [DOI] [PubMed] [Google Scholar]
- 84.Schwarz TF, Johnson C, Grigat C, et al. Three dose levels of a maternal respiratory syncytial virus vaccine candidate are well tolerated and immunogenic in a randomized trial in nonpregnant women. J Infect Dis 2022; 225: 2067–2076. doi: 10.1093/infdis/jiab317 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Albrecht M, Arck PC. Vertically transferred immunity in neonates: mothers, mechanisms and mediators. Front Immunol 2020; 11: 555. doi: 10.3389/fimmu.2020.00555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Dieussaert I, Hyung Kim J, Luik S, et al. RSV prefusion F protein-based maternal vaccine – preterm birth and other outcomes. N Engl J Med 2024; 390: 1009–1021. doi: 10.1056/NEJMoa2305478 [DOI] [PubMed] [Google Scholar]
- 87.Wadman M. FDA advisers agree maternal RSV vaccine protects infants, but are divided on its safety. Date last accessed: 27 August 2024. Date last updated: 19 May 2023. www.science.org/content/article/fda-advisers-agree-maternal-rsv-vaccine-protects-infants-divided-safety [Google Scholar]
- 88.Bonhoeffer J, Black S, Izurieta H, et al. Current status and future directions of post-marketing vaccine safety monitoring with focus on USA and Europe. Biologicals 2012; 40: 393–397. doi: 10.1016/j.biologicals.2012.07.007 [DOI] [PubMed] [Google Scholar]
- 89.Mineva GM, Purtill H, Dunne CP, et al. Impact of breastfeeding on the incidence and severity of respiratory syncytial virus (RSV)-associated acute lower respiratory infections in infants: a systematic review highlighting the global relevance of primary prevention. BMJ Glob Health 2023; 8: e009693. doi: 10.1136/bmjgh-2022-009693 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Scheltema NM, Kavelaars XM, Thorburn K, et al. Potential impact of maternal vaccination on life-threatening respiratory syncytial virus infection during infancy. Vaccine 2018; 36: 4693–4700. doi: 10.1016/j.vaccine.2018.06.021 [DOI] [PubMed] [Google Scholar]
- 91.McFarland EJ, Karron RA, Muresan P, et al. Live-attenuated respiratory syncytial virus vaccine with M2-2 deletion and with small hydrophobic noncoding region is highly immunogenic in children. J Infect Dis 2020; 221: 2050–2059. doi: 10.1093/infdis/jiaa049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Karron RA, Atwell JE, McFarland EJ, et al. Live-attenuated vaccines prevent respiratory syncytial virus-associated illness in young children. Am J Respir Crit Care Med 2021; 203: 594–603. doi: 10.1164/rccm.202005-1660OC [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Downham MA, Scott R, Sims DG, et al. Breast-feeding protects against respiratory syncytial virus infections. Br Med J 1976; 2: 274–276. doi: 10.1136/bmj.2.6030.274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Russell CD, Unger SA, Walton M, et al. The human immune response to respiratory syncytial virus infection. Clin Microbiol Rev 2017; 30: 481–502. doi: 10.1128/CMR.00090-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Jacobino SR, Nederend M, Hennus M, et al. Human amniotic fluid antibodies protect the neonate against respiratory syncytial virus infection. J Allergy Clin Immunol 2016; 138: 1477–80.e5. doi: 10.1016/j.jaci.2016.06.001 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Please note: supplementary material is not edited by the Editorial Office, and is uploaded as it has been supplied by the author.
Supplementary material ERR-0106-2024.SUPPLEMENT (396.9KB, pdf)