


Abstract
DNA lesion bypass is an important cellular response to genomic damage during replication. Human DNA polymerase η (Polη), encoded by the Xeroderma pigmentosum variant (XPV) gene, is known for its activity of error-free translesion synthesis opposite a TT cis-syn cyclobutane dimer. Using purified human Polη, we have examined bypass activities of this polymerase opposite several other DNA lesions. Human Polη efficiently bypassed a template 8-oxoguanine, incorporating an A or a C opposite the lesion with similar efficiencies. Human Polη effectively bypassed a template abasic site, incorporating an A and less frequently a G opposite the lesion. Significant –1 deletion was also observed when the template base 5′ to the abasic site is a T. Human Polη partially bypassed a template (+)-trans-anti-benzo[a]pyrene-N2-dG and predominantly incorporated an A, less frequently a T, and least frequently a G or a C opposite the lesion. This specificity of nucleotide incorporation correlates well with the known mutation spectrum of (+)-trans-anti-benzo[a]pyrene-N2-dG lesion in mammalian cells. These results show that human Polη is capable of error-prone translesion DNA syntheses in vitro and suggest that Polη may bypass certain lesions with a mutagenic consequence in humans.
INTRODUCTION
Lesion bypass is an important cellular response to unrepaired DNA damage during replication. Two modes of lesion bypass are known, error-free bypass and error-prone bypass. Error-free lesion bypass results in the preferential incorporation of the correct nucleotide opposite the damage, whereas error-prone lesion bypass leads to the preferential incorporation of an incorrect nucleotide opposite the damage. Consequently, error-free lesion bypass is a mutation-avoiding mechanism, whereas error-prone lesion bypass is a mutation-generating mechanism. In eukaryotes, an important error-prone lesion bypass mechanism, also known as the damage-induced mutagenesis pathway, has been discovered originally in the yeast Saccharomyces cerevisiae (1–5). Later, this mechanism was also found in humans (5–9). In this mutagenesis pathway, DNA polymerase ζ (Polζ) (the REV3–REV7 protein complex) and the REV1 dCMP transferase are involved in the translesion DNA synthesis step (7,10,11).
More recently, it has been demonstrated that Polη is involved in error-free lesion bypass of a TT dimer (12–14), which apparently operates independently of the Polζ mutagenesis pathway (15). Polη is encoded by the RAD30 gene in the yeast S.cerevisiae and the Xeroderma pigmentosum variant (XPV) gene in humans (12,13). Polη plays an important role in response to UV radiation in humans, since a defect in this gene will lead to the hereditary XPV disease. XPV patients exhibit sensitivity to the sunlight and a predisposition to skin cancer (16). The molecular pathology of XPV could be attributed to the function of Polη in error-free lesion bypass of TT dimers and perhaps other cyclobutane pyrimidine dimers (CPD) as well. One molecular explanation for XP disease is as follows. During replication, some unrepaired CPDs would be bypassed by Polη without mutations. In the absence of Polη, CPDs that are normally bypassed by Polη would accumulate during replication. Some of those CPDs may lead to cell death, resulting in UV sensitivity; while other CPDs may be bypassed by the Polζ mutagenesis pathway, resulting in an elevated frequency of UV mutations. XPV cells show a high proportion of mutations at cytosine-containing photoproducts and a strand bias for mutation specificity (17). These observations remain unexplained.
Most recently, translesion syntheses opposite a cisplatin adduct and an acetylaminofluorene-adducted guanine (AAF-G) have been observed with purified human Polη in vitro (18,19) (our unpublished results). These observations, together with our studies on yeast Polη (20), raised the possibility that human Polη may be capable of bypassing other DNA lesions. Furthermore, it is not very clear whether human Polη is specifically an error-free lesion bypass polymerase. To address these questions, we have examined the response of purified human Polη to several kinds of DNA damage in vitro. In this report, we show that human Polη is capable of error-prone bypass opposite an 8-oxoguanine lesion, an apurinic/apyrimidinic (AP) site and a (+)-trans-anti-benzo[a]pyrene-N2-dG bulky adduct. Our results suggest that human Polη can bypass different types of DNA lesions during replication, but certain lesions are bypassed with mutagenic consequences.
MATERIALS AND METHODS
Materials
A mouse monoclonal antibody against the His6 tag was obtained from Qiagen (Valencia, CA). Alkaline phosphatase conjugated anti-mouse IgG was obtained from Sigma Chemicals (St Louis, MO). Platinum High Fidelity Tag DNA polymerase was purchased from BRL (Bethesda, MD). The human T cell ZAP Express cDNA library was purchased form Stratagene (La Jolla, CA). The yeast rad30 deletion mutant strain BY4741rad30Δ (MATa his3 leu2 met15 ura3 rad30Δ) was from Research Genetics (Huntsville, AL).
Damaged DNA templates
A 30mer DNA template containing a site-specific 8-oxoguanine was synthesized via automated DNA phosphoramidite methods by Operon (Alameda, CA). The sequence is 5′-GGATGGACTGCAGGATCCGGAGGCCGCGCG-3′, where the position of the 8-oxoguanine is underlined. The 36mer templates containing a site-specific tetrahydrofuran (AP site analog) were also synthesized by Operon. The sequences are 5′-GAAGGGATCCTTAAGACTXTAACCGGTCTTCGCGCG-3′, 5′-GAAGGGATCCTTAAGACAXTAACCGGTCTTCGCGCG-3′, 5′-GAAGGGATCCTTAAGACGXTAACCGGTCTTCGCGCG-3′, 5′-GAAGGGATCCTTAAGACCXTAACCGGTCTTCGCGCG-3′, where X designates the AP site. A 49mer DNA template containing a site-specific cis-syn TT dimer or a TT (6-4) photoproduct was prepared as previously described (21). Its sequence is 5′-AGCTACCATGCCTGCACGAATTAAGCAATTCGTAATCATGGTCATAGCT-3′, where the modified TT is underlined. A 33mer DNA template, 5′-CTCGATCGCTAACGCTACCATCCGAATTCGCCC-3′, was reacted with (+)-7R,8S-dihydrodiol-9S,10R-epoxy-7,8,9,10-tetrahydrobenzo[a]pyrene [(+)-trans-anti-BPDE] to generate the 10S (+)-trans-anti-BPDE-N2-dG adduct site at the underlined G and purified as previously described (22–24).
Overexpression plasmid of the human XPV gene
The human XPV cDNA was obtained by polymerase chain reaction (PCR) amplification from the human T cell ZAP Express cDNA library using Platinum High Fidelity Tag DNA polymerase and two primers, 5′-CGGGATCCATGGCTACTGGACAGGATCGAG-3′ and 5′-ACGCGTCGACCATTGTACCCGGCCGAG-3′. The resulting 2.6 kb PCR product was then cloned into the BamHI and SalI sites of the vector pECUh6, yielding pECUh6–XPV. The human XPV gene was verified by DNA sequencing. This expression construct contained the 2 µm origin for multicopy plasmid replication, the URA3 gene for plasmid selection, the CUP1 promoter for inducible XPV gene expression and six His codons preceding the ATG initiator codon of the XPV gene.
Purification of human DNA Polη
Yeast rad30 deletion mutant cells containing pECUh6–XPV were grown at 30°C for 2 days in minimum medium containing 2% dextrose. After 10-fold dilution in 16 l of YPD (2% Bacto-peptone, 1% yeast extract, 2% dextrose) medium, cells were grown for 6 h at 30°C. Expression of human Polη was induced by adding CuSO4 to 0.3 mM and grown for another 3 h. Cells were collected by centrifugation and washed in water. After resuspending in an extraction buffer containing 50 mM Tris–HCl pH 7.5, 1 M KCl, 10% sucrose, 20% glycerol, 5 mM β-mercaptoethanol and protease inhibitors (25), cells were homogenized by zirconium beads in a bead-beater for 15 pulses of 30 s each on ice. The clarified extract (∼130 ml) was loaded onto a HiTrap chelating column charged with NiSO4 (Amersham Pharmacia, 10 ml), followed by washing the column sequentially with 100 ml of Ni buffer A (20 mM phosphate buffer pH 7.4, 1 M KCl, 10% glycerol, 5 mM β-mercaptoethanol and protease inhibitors) containing 10 mM imidazole and 100 ml of Ni buffer A containing 35 mM imidazole. Bound proteins were eluted with a linear gradient of 35–108 mM imidazole. The His6-tagged human Polη was identified by western blot using a mouse monoclonal antibody specific to the His6 tag. The pooled nickel column sample was concentrated by PEG 10 000 and desalted through five 5-ml Sephadex G-25 columns in FPLC buffer A (50 mM Tris–HCl pH 7.5, 1 mM EDTA, 10% glycerol and 5 mM β-mercaptoethanol) containing 100 mM KCl. The resulting sample (∼40 ml) was loaded onto a Mono S HR5/5 column (Amersham Pharmacia) and eluted with a 30-ml linear gradient of 100–400 mM KCl in FPLC buffer A. Polη was eluted at ∼250 mM KCl. Fractions containing Polη were pooled and concentrated by PEG 10 000. Then, the sample was loaded onto an FPLC Superdex 200 gel filtration column equilibrated with FPLC buffer A containing 300 mM KCl and the column was developed in the same buffer.
DNA lesion bypass assays
Lesion bypass assays were performed in standard DNA polymerase reactions using various damaged DNA templates as indicated in the text. The standard DNA polymerase reaction (10 µl) contained 25 mM potassium phosphate pH 7.0, 5 mM MgCl2, 5 mM dithiothreitol, 100 µg/ml bovine serum albumin, 10% glycerol, 50 µM dNTPs (dATP, dCTP, dTTP and dGTP individually or together as indicated), 50 fmol of a DNA substrate containing a 32P-labeled primer and purified DNA Polη. After incubation at 30°C for 10 min, reactions were terminated with 7 µl of a stop solution (20 mM EDTA, 95% formamide, 0.05% bromophenol blue and 0.05% xylene cyanol). The reaction products were resolved on a 20% polyacrylamide gel containing 8 M urea and visualized by autoradiography. Primer extension was quantitated by scanning densitometry of the autoradiogram using the SigmaGel software (Sigma) for analysis.
RESULTS
Purification of human DNA Polη
To facilitate protein purification and detection, we tagged human Polη with six histidine residues at its N-terminus. The tagged protein was expressed in yeast cells of the rad30 deletion mutant strain to avoid potential contamination by the yeast Polη. Human Polη was then purified to near homogeneity (Fig. 1A). The identity of the tagged human Polη was confirmed by western blot analysis using a mouse monoclonal antibody specific to the His6 tag (Fig. 1B). The purified human Polη migrated as a 77 kDa protein on a 10% SDS–polyacrylamide gel (Fig. 1A), consistent with its calculated molecular weight of 78 kDa. Using a 49mer DNA template containing a site-specific cis-syn TT dimer or a TT (6-4) photoproduct (Fig. 2A), we performed lesion bypass assays with the purified human Polη. A 32P-labeled 15mer primer was annealed right before the UV lesion (Fig. 2A). As expected, the purified human Polη efficiently bypassed the TT dimer (Fig. 2B), but was unable to bypass the TT (6-4) photoproduct (Fig. 2C, lane 2). After incorporating one nucleotide opposite the 3′ T of the TT (6-4) photoproduct, DNA synthesis by human Polη was completely stopped (Fig. 2C, lane 2). To reveal the identity of this incorporated nucleotide, we performed DNA synthesis assays with only one deoxyribonucleoside triphosphate: dATP, dCTP, dTTP or dGTP individually. As shown in Figure 2C (lanes 3–6), human Polη predominantly incorporated a G opposite the 3′ T of the TT (6-4) photoproduct. Less frequently, an A was also incorporated (Fig. 2C, lane 3). These results show that our purified human Polη is a full-length protein and an active enzyme.
Figure 1.
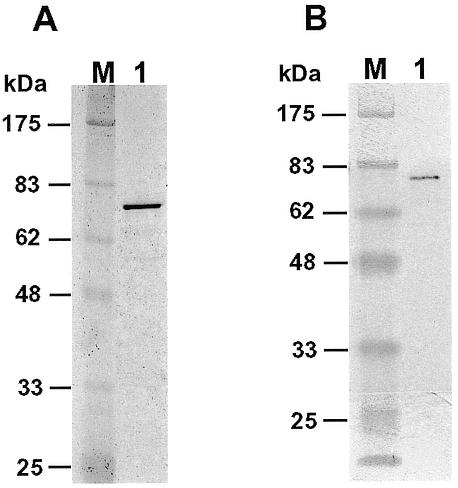
Analyses of purified human Polη. (A) Purified human Polη (80 ng) was analyzed by electrophoresis on a 10% SDS–polyacrylamide gel and visualized by silver staining. Protein size markers (lane M) are indicated on the left. (B) Purified human Polη (16 ng) was analyzed by a western blot using a mouse monoclonal antibody against the His6 tag. Protein size markers (lane M) are indicated on the left.
Figure 2.
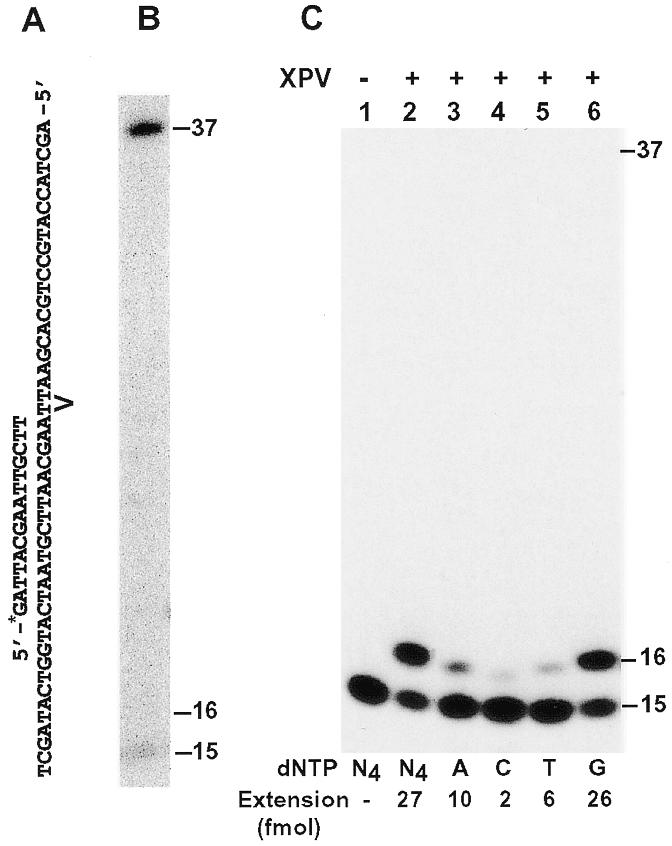
Response of human Polη to a TT dimer and a TT (6-4) photoproduct in template DNA. (A) Primed DNA template containing a TT dimer or a TT (6-4) photoproduct as indicated at the TT sequence. The 15mer primer was labeled with 32P at its 5′-end (*). (B) A lesion bypass assay was performed on the TT dimer template with 0.8 ng of purified human Polη. (C) DNA polymerase assays were performed with 0.8 ng (10 fmol, 1 nM) human Polη using the template (50 fmol) containing a TT (6-4) photoproduct. Polymerase reactions were carried out in the presence of a single deoxyribonucleoside triphosphate dATP (lane 3), dCTP (lane 4), dTTP (lane 5), dGTP (lane 6) or all four dNTPs (lane 2). Lane 1, control reaction without DNA polymerase. Quantitation of extended primers is shown at the bottom of the gel. DNA size markers in nucleotides are indicated on the right.
Error-prone bypass of template 8-oxoguanine by human Polη
8-Oxoguanine is a major form of oxidative damage in DNA. To examine whether human Polη can bypass this lesion, we synthesized a 30mer DNA template containing a site-specific 8-oxoguanine residue (Fig. 3). A 32P-labeled 17mer primer was annealed to the template, right before the 8-oxoguanine residue (Fig. 3A). As shown in Figure 3A (lane 6), human Polη efficiently bypassed the template 8-oxoguanine. To identify the base incorporated opposite 8-oxoguanine, we performed DNA synthesis assays with only one deoxyribonucleoside triphosphate: dATP, dCTP, dGTP or dTTP individually. As shown in Figure 3A (lanes 7–10), human Polη extended 79% of the primers using dATP, 75% of the primers using dCTP and 32% of the primers using dGTP opposite the template 8-oxoguanine. In comparison, human Polη predominantly incorporated the correct C opposite the undamaged template G (Fig. 3A, lanes 1–5). We consistently observed that copying the last template base by human Polη from undamaged DNA was not very efficient, often resulting in two bands visible after separation on a denaturing polyacrylamide gel (e.g. Fig. 3A, lane 1).
Figure 3.

Bypass of a template 8-oxoguanine (8-oxoG) by human Polη. (A) DNA polymerase assays were performed with purified human Polη (0.16 ng, 2 fmol) using the indicated DNA templates without (lanes 1–5) or with (lanes 6–10) a site-specific 8-oxoG. The 17mer primer was labeled with 32P at its 5′-end. Polymerase reactions were carried out in the presence of a single deoxyribonucleoside triphosphate dATP (A), dCTP (C), dTTP (T), dGTP (G) or all four dNTPs (N4) as indicated. (B) Two 32P-labeled primers were annealed separately to the 8-oxoG template as indicated, forming an 8-oxoG-A mismatch (lanes 1–5) and an 8-oxoG-C base pair (lanes 6–10), respectively, at the primer 3′-end. Then, DNA polymerase assays were performed with 0.16 ng of purified human Polη as in (A). DNA size markers in nucleotides are indicated on the right.
Misincorporation of A opposite template 8-oxoguanine would lead to G→T transversion. To examine whether the misincorporated A could be extended by human Polη, we separately annealed two 32P-labeled 18mer primers to the 8-oxoguanine DNA template, forming an 8-oxoguanine-A mismatch and an 8-oxoguanine-C base pair at the primer 3′-end, respectively (Fig. 3B). As shown in Figure 3B (lanes 1 and 6), both primers were efficiently extended by human Polη. In both cases, the correct nucleotide T was predominantly incorporated opposite the undamaged template base A 5′ to the lesion by human Polη (Fig. 3B, lanes 4 and 9). Human Polη extended 52 and 54% of the 8-oxoguanine-A and the 8-oxoguanine-C base pairs, respectively, using dTTP (Fig. 3B, lanes 4 and 9).
To obtain a more quantitative comparison between A and C incorporations opposite the template 8-oxoguanine and subsequent extensions to the 5′ undamaged template base, we performed steady-state kinetic analyses using a previously described method (26). As shown in Table 1, the template 8-oxoguanine slowed human Polη 5.3-fold with respect to C incorporation, as indicated by the finc (0.19). However, human Polη incorporated both C and A opposite the 8-oxoguanine with essentially the same efficiency (Table 1). Consequently, A misincorporation by human Polη increased 421-fold (3.8 × 10–4/1.6 × 10–1) from opposite undamaged template G to 8-oxoguanine (Table 1). Further extension of the misincorporated A from opposite the lesion to the next template base was only 1.7-fold (1/5.8 × 10–1) slower than extension from the correct 8-oxoguanine-C base pair (Table 1). Therefore, we conclude that, upon encountering 8-oxoguanine in DNA, human Polη will perform efficient error-prone lesion bypass, leading to frequent G→T transversions.
Table 1. Kinetic measurements of nucleotide incorporation opposite 8-oxoguanine and subsequent extension to the next template base by human Polη.
| Vmax (fmol/min) | Km (µM) | Vmax/Km | finc or fexta | |
|---|---|---|---|---|
| Incorporation (template-insertion)b | ||||
| G-dCTP | 3.0 ± 0.12 | 0.044 ± 0.001 | 68 | 1 |
| G-dATP | 2.7 ± 0.21 | 102 ± 28 | 0.026 | 3.8 × 10–4 |
| 8-oxoG-dCTP | 3.2 ± 0.14 | 0.24 ± 0.05 | 13 | 1.9 × 10–1 |
| 8-oxoG-dATP | 3.0 ± 0.09 | 0.27 ± 0.04 | 11 | 1.6 × 10–1 |
| Extension (template-primer)c | ||||
| 8-oxoG-C | 2.7 ± 0.18 | 0.18 ± 0.06 | 15 | 1 |
| 8-oxoG-A | 2.6 ± 0.11 | 0.31 ± 0.06 | 8.7 | 5.8 × 10–1 |
afinc = (Vmax/Km)incorrect/(Vmax/Km)correct; fext = (Vmax/Km)mismatched primer /(Vmax/Km)matched primer.
bNucleotide incorporation opposite undamaged or damaged template G.
cExtension from matched C or mismatched A opposite the template 8-oxoG to the next template base (A). The correct nucleotide (T) was incorporated opposite this 5′ undamaged base and dTTP was used for the kinetic measurements of extension.
Mutagenic translesion synthesis opposite a template AP site by human Polη
AP sites are significant DNA lesions, which can arise in the genome spontaneously or can be induced by many environmental agents. Using purified human Polη, we examined the response of this polymerase to an AP site in DNA. A 14mer primer was labeled at its 5′-end with 32P and annealed to a DNA template (template AP-T). The 3′-end of the primer was annealed three nucleotides before the template AP site (Fig. 4). As shown in Figure 4 (lane 3), purified human Polη was able to bypass this AP site. Human Polη efficiently incorporated one nucleotide opposite the template AP site, but its further extension was inhibited by the lesion (Fig. 4, lane 3), as evidenced by the accumulation of the 18mer synthesis products. In the absence of the AP site, the 18mer synthesis product was not accumulated by human Polη (Fig. 4, lane 2). By increasing the amount of human Polη from 21 to 103 fmol in the reaction, the majority of the 14mer primers were extended to the end of the template within 10 min at 30°C, bypassing the template AP site (data not shown).
Figure 4.
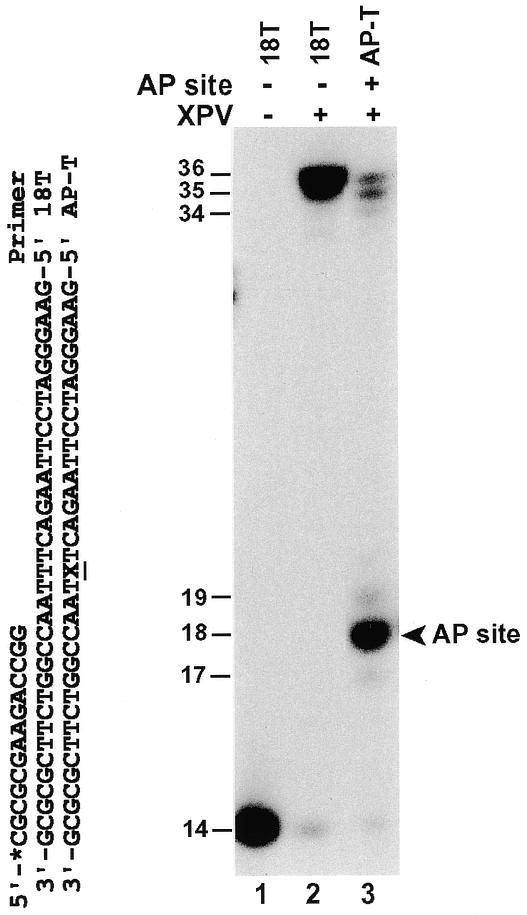
Bypass of a template AP site by human Polη. DNA templates without (template 18T) or with (template AP-T) an AP site were annealed to a 32P-labeled 14mer primer as shown. The AP site is indicated by X. Polymerase assays were performed with 1.6 ng (21 fmol) human Polη using undamaged template 18T (lane 2) or the AP site-containing template AP-T (lane 3). The 18mer DNA band extended opposite the AP site is indicated by an arrowhead. Lane 1, control reaction without DNA polymerase. DNA size markers in nucleotides are indicated on the left.
Bypass of AP sites by human Polκ was strongly influenced by the sequence context 5′ to the AP site (27,28). To examine whether AP site bypass by human Polη is also influenced by sequence context, we synthesized three more DNA templates that differed from the above AP site template (template AP-T) by one nucleotide 5′ to the AP site (Fig. 5A). A 17mer primer was labeled at its 5′-end with 32P and annealed to the four AP site templates, right before the template AP site (Fig. 5A). Then, the primed DNA templates were incubated with human Polη for lesion bypass assays. As shown in Figure 5B, the efficiency of AP site bypass by human Polη was similar among the four templates. Again, primer extension was inhibited after incorporating one nucleotide opposite the AP site (18mer DNA fragment) (Fig. 5B, lanes 2–5).
Figure 5.

Effect of sequence context on AP site bypass by human Polη. (A) DNA templates used for AP site bypass assays. The 32P-labeled 17mer primer was annealed right before the template AP site that is indicated by X. Different template bases 5′ to the AP site are underlined. The AflII restriction cleavage site on the 32P-labeled strand is shown, as well as the AflII recognition sequence. (B) Polymerase assays were performed with 1.6 ng (21 fmol) of purified human Polη using an undamaged DNA template (lane 1) or the various AP site-containing templates as indicated (lanes 2–5). DNA size markers in nucleotides are indicated on the right.
When the template base 5′ to the AP site is a T (template AP-T), the majority of the bypassed products appeared to be one nucleotide shorter (Fig. 5B, lane 4). This result raised the possibility that deletions might be involved during AP site bypass by human Polη in the AP-T template. To examine this possibility, we performed lesion bypass assays followed by digestion with the restriction endonuclease AflII. For normal primer extension and after AflII digestion of the extended products, a 32P-labeled 22mer band was expected to arise (Fig. 5A). Indeed, using the undamaged template (template 18T), DNA synthesis products of human Polη (Fig. 6, lane 1) were cleaved by AflII to a 22-nt fragment (Fig. 6, lane 2). However, using the AP site template AP-T, lesion bypass products of human Polη (Fig. 6, lane 7) were cleaved by AflII to three bands, 22mer, 21mer and 20mer fragments with relative intensities of 6.7:7.7:1, respectively (Fig. 6, lane 8). Thus, 44, 50 and 6% of the bypass products were derived from bypass without deletion, –1 deletion and –2 deletion mechanisms, respectively. Using AP site templates AP-A, AP-C and AP-G containing a template A, C or G, respectively, 5′ to the AP site (Fig. 5A), we performed similar AflII cleavage experiments following lesion bypass by human Polη. As shown in Figure 6 (lanes 4, 6 and 10), the major cleavage product was the 22mer DNA fragment, indicating bypass without deletion.
Figure 6.
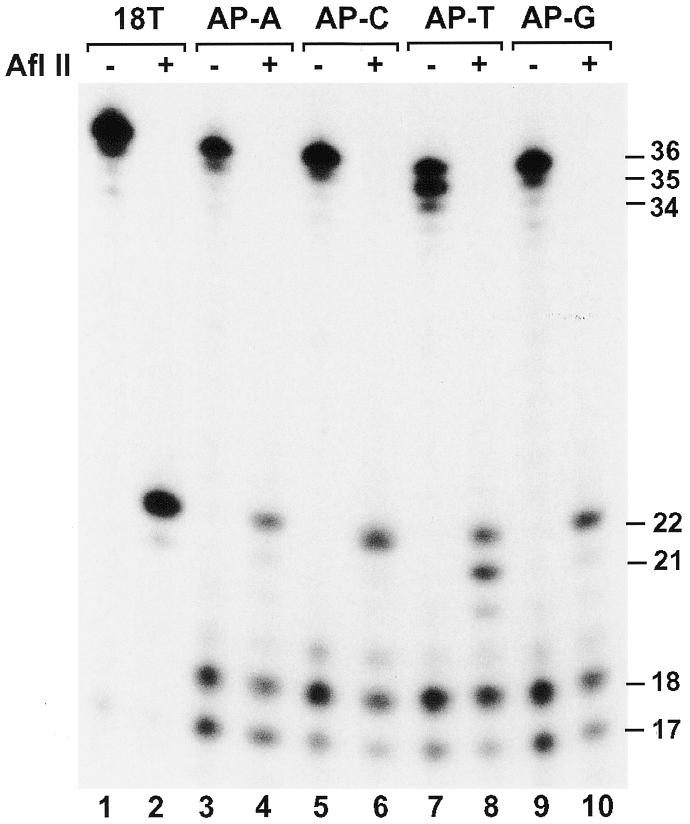
Analysis of AP site bypass products by AflII restriction digestion. DNA polymerase assays were performed with the undamaged DNA template (18T) or the four AP site-containing templates using 8 ng (103 fmol) of human Polη. After the polymerase reaction, 5 µl of the reaction products were mixed with 2 µl of H2O, 1 µl of the 10× AfIII buffer (500 mM Tris–HCl pH 8.0, 100 mM MgCl2) and 2 µl of AfIII (20 U). AfIII digestions were at 37°C for 4 h. The digested products were separated by electrophoresis on a 20% denaturing polyacrylamide gel and visualized by autoradiography. Samples without (AflII, –) or with (AflII, +) AflII treatment are indicated. DNA size markers in nucleotides are indicated on the right.
To identify the nucleotide incorporated by human Polη opposite the template AP site, we performed lesion bypass assays with only one deoxyribonucleoside triphosphate. As shown in Figure 7, nucleotide incorporation opposite the AP site by human Polη followed the order from most frequent to least frequent: A>G>C>T. Taken together, these results show that human Polη mainly incorporates A opposite a template AP site, whose further extension is the rate-limiting step. When the template base 5′ to the AP site is a T, 50% of the AP site bypass is mediated by a –1 deletion mechanism.
Figure 7.
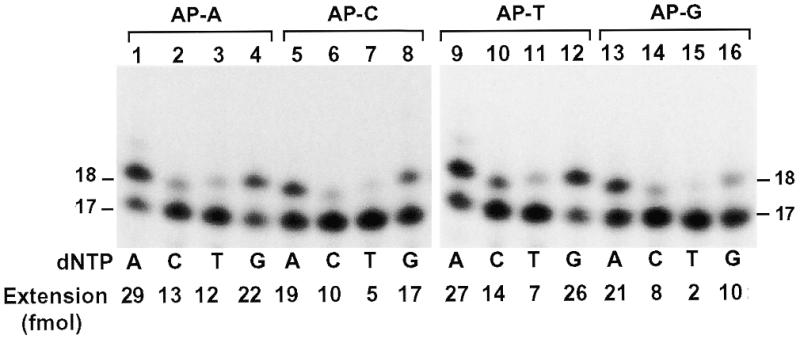
Nucleotide incorporation opposite the template AP site. Lesion bypass assays were performed with 0.8 ng (10 fmol, 1 nM) human Polη using various AP site-containing templates (50 fmol, 5 nM) in a standard DNA polymerase reaction buffer containing dATP (A), dCTP (C), dTTP (T) and dGTP (G) individually as indicated. Templates contained a 32P-labeled 17mer primer annealed right before the template AP site. DNA sequences of the various templates are shown in Figure 5A. Quantitation of extended primers is shown at the bottom of the gels. DNA size markers in nucleotides are indicated on the sides.
Error-prone translesion synthesis opposite a (+)-trans-anti-benzo[a]pyrene-N2-dG adduct by human Polη
Like a TT dimer, (+)-trans-anti-benzo[a]pyrene-N2-dG is a bulky lesion in DNA. This lesion represents the major DNA damage caused by the reaction of racemic anti-benzo[a]pyrene-7,8-dihydrodiol-9,10-epoxide (BPDE), a potent ultimate carcinogen of benzo[a]pyrene, with DNA (29,30). Using purified human Polη, we examined the response of this polymerase to a template (+)-trans-anti-BPDE-N2-dG bulky lesion. A 19mer primer was labeled with 32P at its 5′-end and annealed right before the template (+)-trans-anti-BPDE-N2-dG lesion (Fig. 8). As shown in Figure 8 (lane 1), human Polη effectively incorporated one nucleotide opposite the lesion and extended one more nucleotide downstream, generating a 21mer DNA fragment. However, further DNA synthesis was inhibited by the lesion. Nevertheless, some of the stalled 21mer DNA fragments were further extended by human Polη to near the end of the DNA template (Fig. 8, lane 1). To identify the nucleotide incorporated opposite the lesion, we performed lesion bypass assays with only one of the four deoxyribonucleoside triphosphates at a time. As shown in Figure 8 (lanes 2–5), human Polη predominantly incorporated an A opposite the template (+)-trans-anti-BPDE-N2-dG. Less frequently, a T was incorporated opposite this lesion. Least frequently, a C or a G was also incorporated opposite this lesion. These results demonstrate that human Polη is capable of error-prone translesion synthesis opposite a template (+)-trans-anti-BPDE-N2-dG.
Figure 8.
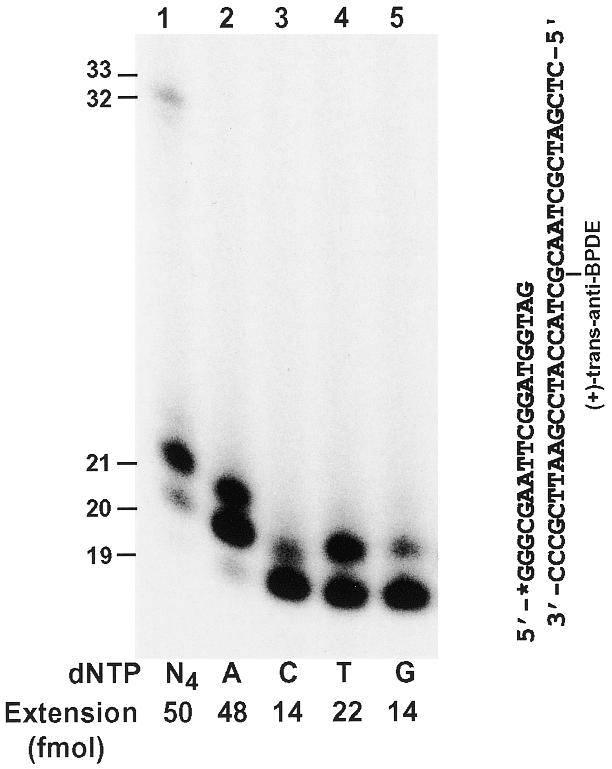
Bypass of a template (+)-trans-anti-BPDE-N2-dG lesion by human Polη. A 19mer primer was labeled with 32P at its 5′-end and annealed right before a template (+)-trans-anti-BPDE-N2-dG as shown on the right. Using 50 fmol (5 nM) DNA, polymerase reactions were performed with 2 ng (26 fmol, 2.6 nM) of human Polη in the presence of a single deoxyribonucleoside triphosphate dATP (lane 2), dCTP (lane 3), dTTP (lane 4), dGTP (lane 5) or all four dNTPs (lane 1). Quantitation of extended primers is shown at the bottom of the gel. DNA size markers in nucleotides are indicated on the left.
DISCUSSION
Polη was originally identified as an error-free lesion bypass polymerase in response to UV radiation (12–14). This activity of human Polη is critical in preventing the XPV disease (13,31). However, our recent biochemical studies on the yeast Polη suggest that this polymerase may also be involved in error-prone translesion synthesis (20). In this study, we show that human Polη is indeed able to bypass an 8-oxoguanine, an AP site and a (+)-trans-anti-benzo[a]pyrene-N2-dG bulky adduct in an error-prone manner in vitro. Hence, depending on the specific DNA lesion, human Polη is capable of both error-free and error-prone translesion syntheses, at least in vitro.
Error-free AA incorporations opposite a TT dimer by human Polη can be interpreted by two mechanisms. One is indiscriminating A incorporation opposite a damaged template. The other is that the TT dimer is recognized by Polη as a coding TT sequence. We found that human Polη predominantly incorporates a G opposite the 3′ T of a TT (6-4) photoproduct. This result supports the second mechanism of lesion bypass by Polη and is inconsistent with the first interpretation. An intriguing observation is that a site-specific TT (6-4) photoproduct induced primarily 3′ T→C substitution mutations in COS cells, resulting from G incorporation opposite the 3′ T of the lesion (32). Since human Polη did not bypass the 5′ T of the TT (6-4) photoproduct at the enzyme concentrations used, it is not clear at present whether G incorporation by human Polη opposite the 3′ T of the lesion is physiologically significant. However, it is possible that, following G incorporation, the 5′ T of the lesion may be bypassed by human Polζ. Supporting this model, cooperation between yeast Polη and yeast Polζ to bypass a template AP site has been observed in vitro (20). Furthermore, when a primer 3′-end is a G and annealed opposite the 3′ T of the TT (6-4) photoproduct, this primer is effectively extended to the end of the template by purified yeast Polζ (D.Guo, and Z.Wang, unpublished results).
Significant AP site bypass was observed at a low polymerase to DNA ratio: 21 fmol Polη versus 50 fmol DNA (Fig. 4). Thus, it is likely that human Polη plays a role in error-prone bypass of AP sites in vivo. The consequence of AP site bypass by human Polη is mainly A incorporation opposite the lesion, although less frequently, G is also incorporated. When the template base 5′ to the AP site is a T, a –1 deletion was observed among 50% of the bypass products. This –1 deletion bypass probably results from re-alignment of the incorporated A with the next template T. This property of human Polη is reminiscent of the AP site bypass by human Polκ (27,28). However, in contrast to human Polκ (27), the efficiency of AP site bypass by human Polη is not significantly affected by the sequence context 5′ to the AP site. AP site bypass by human Polη was also observed by Masutani et al. (19). However, the –1 deletion bypass and the sequence context effect on AP site bypass by human Polη were not examined in the study of Masutani et al. (19).
In Escherichia coli, translesion synthesis of an AP site results in preferential incorporation of an A opposite the lesion, leading to the ‘A rule’ hypothesis (33). In mammals, however, it seems that A incorporation opposite an AP site is not strongly biased. Similar incorporations of A, C and T opposite an AP site were observed (34–37). In one study, preferential A incorporation opposite an AP site was noticed (38). Yet in another study, preferential G incorporation was detected opposite an AP site (39). Relative to A incorporation, G is also significantly incorporated by purified human Polη opposite an AP site, especially when the template base 5′ to the lesion is a pyrimidine. Thus, Polη may have contributed in part to the previously reported A and G incorporations opposite a template AP site in mammalian cells.
The (+)-trans-anti-BPDE-N2-dG bulky adduct is highly mutagenic in COS cells (40,41). The mutagenic potential of (+)-trans-anti-BPDE-N2-dG lesions in different base sequence contexts has been investigated using site-specific mutagenesis methods in a variety of prokaryotic and eukaryotic cellular systems (40–45). In some or all of these studies, bypass polymerases may have played a role in the mutagenic bypass of this lesion. In this study we found that human Polη can insert incorrect bases opposite the (+)-trans-anti-BPDE-N2-dG lesion. Human Polη predominantly incorporates an A, less frequently a T and even less frequently a G or C opposite the lesion (Fig. 8). Therefore, if these single nucleotide insertion experiments are reflected in full-length primer extension, then these results predict that Polη causes predominantly G→T transversions, less frequent G→A transitions and even less frequent G→C transversions. Similar mutagenic specificities were observed in COS cells (40,41). This correlation between in vitro and in vivo results suggest a role for Polη in mutagenic bypass of the (+)-trans-anti-BPDE-N2-dG lesion in mammalian cells. An earlier study has attempted to examine the cytotoxicity and mutagenesis induced by BPDE in cultured XPV cells and concluded that neither was affected without functional XPV gene (46). In light of our new biochemical information on the response of human Polη to the (+)-trans-anti-BPDE-N2-dG adduct in vitro, more rigorously controlled in vivo studies are needed to definitively answer whether Polη is important for in vivo bypass and mutagenesis of benzo[a]pyrene adducts.
Comparing the yeast and the human Polη, some differences were noticed. While yeast Polη predominantly incorporates C opposite 8-oxoguanine (20), human Polη inserts C and A with similar efficiencies. Whereas yeast Polη predominantly incorporates G opposite the AP site (20), human Polη prefers A. Nevertheless, both yeast and human Polη were capable of translesion synthesis opposite multiple different DNA lesions. As suggested by two earlier reports, the ability of Polη to bypass DNA lesions may have derived from its relaxed requirement for correct base pairing geometry at the polymerase active site (47,48). As a result of such a specialized function in lesion bypass, Polη synthesizes DNA from undamaged templates with extraordinarily low fidelity (47,48).
Our results show that human Polη is capable of error-prone translesion DNA syntheses in vitro. These results raised the possibility that human Polη may be involved in mutagenesis induced by certain DNA lesions such as AP sites and (+)-trans-anti-BPDE-N2-dG adducts in vivo.
Acknowledgments
ACKNOWLEDGEMENTS
This work was supported by a THRI grant from the Tobacco and Health Research Institute of the University of Kentucky (Z.W.), a New Investigator Award in Toxicology from Burroughs Wellcome Fund (Z.W.), a NIH grant CA40463 (J.-S.T.) and a NIH grant CA20851 (N.E.G.).
REFERENCES
- 1.Lemontt J.F. (1971) Mutants of yeast defective in mutation induction by ultraviolet light. Genetics, 68, 21–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lemontt J.F. (1977) Pathways of ultraviolet mutability in Saccharomyces cerevisiae. III. Genetic analysis and properties of mutants resistant to ultraviolet-induced forward mutation. Mutat. Res., 43, 179–204. [DOI] [PubMed] [Google Scholar]
- 3.Lawrence C. (1994) The RAD6 DNA repair pathway in Saccharomyces cerevisiae: what does it do, and how does it do it? Bioessays, 16, 253–258. [DOI] [PubMed] [Google Scholar]
- 4.Lawrence C.W. and Hinkle,D.C. (1996) DNA polymerase ζ and the control of DNA damage induced mutagenesis in eukaryotes. Cancer Surv., 28, 21–31. [PubMed] [Google Scholar]
- 5.Gibbs P.E., McGregor,W.G., Maher,V.M., Nisson,P. and Lawrence,C.W. (1998) A human homolog of the Saccharomyces cerevisiae REV3 gene, which encodes the catalytic subunit of DNA polymerase ζ. Proc. Natl Acad. Sci. USA, 95, 6876–6880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lin W., Wu,X. and Wang,Z. (1999) A full-length cDNA of hREV3 is predicted to encode DNA polymerase ζ for damage-induced mutagenesis in humans. Mutat. Res., 433, 89–98. [DOI] [PubMed] [Google Scholar]
- 7.Lin W., Xin,H., Zhang,Y., Wu,X., Yuan,F. and Wang,Z. (1999) The human REV1 gene codes for a DNA template-dependent dCMP transferase. Nucleic Acids Res., 27, 4468–4475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gibbs P.E., Wang,X.D., Li,Z., McManus,T.P., McGregor,W.G., Lawrence,C.W. and Maher,V.M. (2000) The function of the human homolog of Saccharomyces cerevisiae REV1 is required for mutagenesis induced by UV light. Proc. Natl Acad. Sci. USA, 97, 4186–4191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Murakumo Y., Roth,T., Ishii,H., Rasio,D., Numata,S., Croce,C.M. and Fishel,R. (2000) A human REV7 homolog that interacts with the polymerase ζ catalytic subunit hREV3 and the spindle assembly checkpoint protein hMAD2. J. Biol. Chem., 275, 4391–4397. [DOI] [PubMed] [Google Scholar]
- 10.Nelson J.R., Lawrence,C.W. and Hinkle,D.C. (1996) Deoxycytidyl transferase activity of yeast REV1 protein. Nature, 382, 729–731. [DOI] [PubMed] [Google Scholar]
- 11.Nelson J.R., Lawrence,C.W. and Hinkle,D.C. (1996) Thymine-thymine dimer bypass by yeast DNA polymerase ζ. Science, 272, 1646–1649. [DOI] [PubMed] [Google Scholar]
- 12.Johnson R.E., Prakash,S. and Prakash,L. (1999) Efficient bypass of a thymine-thymine dimer by yeast DNA polymerase, Polη. Science, 283, 1001–1004. [DOI] [PubMed] [Google Scholar]
- 13.Masutani C., Kusumoto,R., Yamada,A., Dohmae,N., Yokoi,M., Yuasa,M., Araki,M., Iwai,S., Takio,K. and Hanaoka,F. (1999) The XPV (xeroderma pigmentosum variant) gene encodes human DNA polymerase η. Nature, 399, 700–704. [DOI] [PubMed] [Google Scholar]
- 14.Masutani C., Araki,M., Yamada,A., Kusumoto,R., Nogimori,T., Maekawa,T., Iwai,S. and Hanaoka,F. (1999) Xeroderma pigmentosum variant (XP-V) correcting protein from HeLa cells has a thymine dimer bypass DNA polymerase activity. EMBO J., 18, 3491–3501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.McDonald J.P., Levine,A.S. and Woodgate,R. (1997) The Saccharomyces cerevisiae RAD30 gene, a homologue of Escherichia coli dinB and umuC, is DNA damage inducible and functions in a novel error-free postreplication repair mechanism. Genetics, 147, 1557–1568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cleaver J.E. and Kraemer,K.H. (1989) Xeroderma pigmentosum. In Scriver,C.R., Beaudet,A.L., Sly,W.S. and Valle,D. (eds), The Metabolic Basis of Inherited Disease, 6th Edn. McGraw-Hill Book Co., New York, NY, pp. 2949–2971.
- 17.McGregor W.G., Wei,D., Maher,V.M. and McCormick,J.J. (1999) Abnormal, error-prone bypass of photoproducts by xeroderma pigmentosum variant cell extracts results in extreme strand bias for the kinds of mutations induced by UV light. Mol. Cell. Biol., 19, 147–154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vaisman A., Masutani,C., Hanaoka,F. and Chaney,S.G. (2000) Efficient translesion replication past oxaliplatin and cisplatin GpG adducts by human DNA polymerase η. Biochemistry, 39, 4575–4580. [DOI] [PubMed] [Google Scholar]
- 19.Masutani C., Kusumoto,R., Iwai,S. and Hanaoka,F. (2000) Mechanisms of accurate translesion synthesis by human DNA polymerase η. EMBO J., 19, 3100–3109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yuan F., Zhang,Y., Rajpal,D.K., Wu,X., Guo,D., Wang,M., Taylor,J.-S. and Wang,Z. (2000) Specificity of DNA lesion bypass by the yeast DNA polymerase η. J. Biol. Chem., 275, 8233–8239. [DOI] [PubMed] [Google Scholar]
- 21.Smith C.A. and Taylor,J.S. (1993) Preparation and characterization of a set of deoxyoligonucleotide 49-mers containing site-specific cis-syn, trans-syn-I, (6-4), and Dewar photoproducts of thymidylyl(3′→5′)-thymidine. J. Biol. Chem., 268, 11143–11151. [PubMed] [Google Scholar]
- 22.Cosman M., Ibanez,V., Geacintov,N.E. and Harvey,R.G. (1990) Preparation and isolation of adducts in high yield derived from the binding of two benzo[a]pyrene-7,8-dihydroxy-9,10-oxide stereoisomers to the oligonucleotide d(ATATGTATA). Carcinogenesis, 11, 1667–1672. [DOI] [PubMed] [Google Scholar]
- 23.Geacintov N.E., Cosman,M., Mao,B., Alfano,A., Ibanez,V. and Harvey,R.G. (1991) Spectroscopic characteristics and site I/site II classification of cis and trans benzo[a]pyrene diolepoxide enantiomer-guanosine adducts in oligonucleotides and polynucleotides. Carcinogenesis, 12, 2099–2108. [DOI] [PubMed] [Google Scholar]
- 24.Rechkoblit O., Amin,S. and Geacintov,N.E. (1999) Primer length dependence of binding of DNA polymerase I Klenow fragment to template-primer complexes containing site-specific bulky lesions. Biochemistry, 38, 11834–11843. [DOI] [PubMed] [Google Scholar]
- 25.Xin H., Lin,W., Sumanasekera,W., Zhang,Y., Wu,X. and Wang,Z. (2000) The human RAD18 gene product interacts with HHR6A and HHR6B. Nucleic Acids Res., 28, 2847–2854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Creighton S., Bloom,L.B. and Goodman,M.F. (1995) Gel fidelity assay measuring nucleotide misinsertion, exonucleolytic proofreading, and lesion bypass efficiencies. Methods Enzymol., 262, 232–256. [DOI] [PubMed] [Google Scholar]
- 27.Zhang Y., Yuan,F., Wu,X., Wang,M., Rechkoblit,O., Taylor,J.-S., Geacintov,N.E. and Wang,Z. (2000) Error-free and error-prone lesion bypass by human DNA polymerase κin vitro. Nucleic Acids Res., 28, 4138–4146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ohashi E., Ogi,T., Kusumoto,R., Iwai,S., Masutani,C., Hanaoka,F. and Ohmori,H. (2000) Error-prone bypass of certain DNA lesions by the human DNA polymerase κ. Genes Dev., 14, 1589–1594. [PMC free article] [PubMed] [Google Scholar]
- 29.Cheng S.C., Hilton,B.D., Roman,J.M. and Dipple,A. (1989) DNA adducts from carcinogenic and noncarcinogenic enantiomers of benzo[a]pyrene dihydrodiol epoxide. Chem. Res. Toxicol., 2, 334–340. [DOI] [PubMed] [Google Scholar]
- 30.Peltonen K. and Dipple,A. (1995) Polycyclic aromatic hydrocarbons: chemistry of DNA adduct formation. J. Occup. Environ. Med., 37, 52–58. [DOI] [PubMed] [Google Scholar]
- 31.Johnson R.E., Kondratick,C.M., Prakash,S. and Prakash,L. (1999) hRAD30 mutations in the variant form of xeroderma pigmentosum. Science, 285, 263–265. [DOI] [PubMed] [Google Scholar]
- 32.Kamiya H., Iwai,S. and Kasai,H. (1998) The (6-4) photoproduct of thymine-thymine induces targeted substitution mutations in mammalian cells. Nucleic Acids Res., 26, 2611–2617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Strauss B.S. (1991) The ‘A rule’ of mutagen specificity: a consequence of DNA polymerase bypass of non-instructional lesions? Bioessays, 13, 79–84. [DOI] [PubMed] [Google Scholar]
- 34.Gentil A., Renault,G., Madzak,C., Margot,A., Cabral-Neto,J.B., Vasseur,J.J., Rayner,B., Imbach,J.L. and Sarasin,A. (1990) Mutagenic properties of a unique abasic site in mammalian cells. Biochem. Biophys. Res. Commun., 173, 704–710. [DOI] [PubMed] [Google Scholar]
- 35.Gentil A., Cabral-Neto,J.B., Mariage-Samson,R., Margot,A., Imbach,J.L., Rayner,B. and Sarasin,A. (1992) Mutagenicity of a unique apurinic/apyrimidinic site in mammalian cells. J. Mol. Biol., 227, 981–984. [DOI] [PubMed] [Google Scholar]
- 36.Neto J.B., Gentil,A., Cabral,R.E. and Sarasin,A. (1992) Mutation spectrum of heat-induced abasic sites on a single-stranded shuttle vector replicated in mammalian cells. J. Biol. Chem., 267, 19718–19723. [PubMed] [Google Scholar]
- 37.Cabral Neto J.B., Cabral,R.E., Margot,A., Le Page,F., Sarasin,A. and Gentil,A. (1994) Coding properties of a unique apurinic/apyrimidinic site replicated in mammalian cells. J. Mol. Biol., 240, 416–420. [DOI] [PubMed] [Google Scholar]
- 38.Takeshita M. and Eisenberg,W. (1994) Mechanism of mutation on DNA templates containing synthetic abasic sites: study with a double strand vector. Nucleic Acids Res., 22, 1897–1902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Klinedinst D.K. and Drinkwater,N.R. (1992) Mutagenesis by apurinic sites in normal and ataxia telangiectasia human lymphoblastoid cells. Mol. Carcinog., 6, 32–42. [DOI] [PubMed] [Google Scholar]
- 40.Moriya M., Spiegel,S., Fernandes,A., Amin,S., Liu,T., Geacintov,N. and Grollman,A.P. (1996) Fidelity of translesional synthesis past benzo[a]pyrene diol epoxide-2′-deoxyguanosine DNA adducts: marked effects of host cell, sequence context, and chirality. Biochemistry, 35, 16646–16651. [DOI] [PubMed] [Google Scholar]
- 41.Fernandes A., Liu,T., Amin,S., Geacintov,N.E., Grollman,A.P. and Moriya,M. (1998) Mutagenic potential of stereoisomeric bay region (+)- and (–)-cis-anti-benzo[a]pyrene diol epoxide-N2-2′-deoxyguanosine adducts in Escherichia coli and simian kidney cells. Biochemistry, 37, 10164–10172. [DOI] [PubMed] [Google Scholar]
- 42.Mackay W., Benasutti,M., Drouin,E. and Loechler,E.L. (1992) Mutagenesis by (+)-anti-B[a]P-N2-Gua, the major adduct of activated benzo[a]pyrene, when studied in an Escherichia coli plasmid using site-directed methods. Carcinogenesis, 13, 1415–1425. [DOI] [PubMed] [Google Scholar]
- 43.Jelinsky S.A., Liu,T., Geacintov,N.E. and Loechler,E.L. (1995) The major, N2-Gua adduct of the (+)-anti-benzo[a]pyrene diol epoxide is capable of inducing G→A and G→C, in addition to G→T, mutations. Biochemistry, 34, 13545–13553. [DOI] [PubMed] [Google Scholar]
- 44.Shukla R., Liu,T., Geacintov,N.E. and Loechler,E.L. (1997) The major, N2-dG adduct of (+)-anti-B[a]PDE shows a dramatically different mutagenic specificity (predominantly, G→A) in a 5′-CGT-3′ sequence context. Biochemistry, 36, 10256–10261. [DOI] [PubMed] [Google Scholar]
- 45.Page J.E., Zajc,B., Oh-hara,T., Lakshman,M.K., Sayer,J.M., Jerina,D.M. and Dipple,A. (1998) Sequence context profoundly influences the mutagenic potency of trans-opened benzo[a]pyrene 7,8-diol 9,10-epoxide-purine nucleoside adducts in site-specific mutation studies. Biochemistry, 37, 9127–9137. [DOI] [PubMed] [Google Scholar]
- 46.Watanabe M., Maher,V.M. and McCormick,J.J. (1985) Excision repair of UV- or benzo[a]pyrene diol epoxide-induced lesions in xeroderma pigmentosum variant cells is ‘error free’. Mutat. Res., 146, 285–294. [DOI] [PubMed] [Google Scholar]
- 47.Matsuda T., Bebenek,K., Masutani,C., Hanaoka,F. and Kunkel,T.A. (2000) Low fidelity DNA synthesis by human DNA polymerase η. Nature, 404, 1011–1013. [DOI] [PubMed] [Google Scholar]
- 48.Washington M.T., Johnson,R.E., Prakash,S. and Prakash,L. (1999) Fidelity and processivity of Saccharomyces cerevisiae DNA polymerase η. J. Biol. Chem., 274, 36835–36838. [DOI] [PubMed] [Google Scholar]