

Abstract

The organization of chromatin is critical for gene expression, yet the underlying mechanisms responsible for this organization remain unclear. Recent work has suggested that phase separation might play an important role in chromatin organization, yet the molecular forces that drive chromatin phase separation are poorly understood. In this work we interrogate a molecular model of chromatin to quantify the driving forces and thermodynamics of chromatin phase separation. By leveraging a multiscale approach, our molecular model is able to reproduce chromatin’s chemical and structural details at the level of a few nanometers, yet remain efficient enough to simulate chromatin phase separation across 100 nm length scales. We first demonstrate that our model can reproduce key experiments of phase separating nucleosomal arrays, and then apply our model to quantify the interactions that drive their formation into chromatin condensates with either liquid- or solid-like material properties. We next use our model to characterize the molecular structure within chromatin condensates and find that this structure is irregularly ordered and is inconsistent with existing 30 nm fiber models. Lastly we examine how post-translational modifications can modulate chromatin phase separation and how the acetylation of chromatin can lead to chromatin decompaction while still preserving phase separation. Taken together, our work provides a molecular view into the structure and dynamics of phase-separated chromatin and provides new insights into how phase separation might manifest in the nucleus of living cells.
Introduction
Eukaryotic DNA must become highly compacted to fit within the confined space of the nucleus yet simultaneously organized to correctly facilitate gene expression. To meet these requirements, eukaryotic DNA complexes with histone octamers to form chromatin: a hierarchically compacted material which exhibits a heterogeneous and dynamic organization.1 Chromatin’s organization is driven by a diverse set of interactions between chromatin, RNAs, and proteins, resulting in several concurrent mechanisms which regulate chromatin’s organization.2
During interphase, chromatin is organized into distinct membraneless compartments like the nucleolus as well as heterochromatin and euchromatin domains. These compartments consist of different mixtures of DNA, RNAs, and proteins, and experiments have found that their constituents often interact to induce phase separation.3 Although the details of how phase separation might establish chromatin compartments is unclear,4 phase separation is now understood to play a role in the self-assembly and maintenance of membraneless bodies within the nucleus.5 For example, in the nucleolus liquid–liquid phase separation driven by weak, multivalent interactions is responsible for nucleolus formation as well as the management of nucleolar subcompartments.6 In heterochromatin, phase separation initiated by either HP1 bridging interactions or condensation explains how distal heterochromatic loci associate to form domains with regulated properties.7−10
Other recent work has focused on understanding the intrinsic phase separation of chromatin in vitro. In these studies, short chromatin segments consisting of reconstituted nucleosomal arrays have been observed to phase separate into dense chromatin condensates in the absence of chromatin-associated proteins and RNAs.11−14 These studies also provide clues into how chromatin phase separation may be regulated in the cell, such as the importance of linker histones and post-translational modifications in mediating chromatin phase separation.11
Nonetheless, many outstanding questions regarding the phase separation of chromatin remain. One open question relates to whether chromatin is liquid-like or solid-like and how this material state of chromatin might be modulated under different conditions. For example, recent experiments have observed that chromatin can be either liquid- or solid-like depending on the spacing of nucleosomes, the ionic conditions, and the presence of linker histones.11−14 Another open question relates to the mesoscale structure of chromatin condensates and whether there is any evidence for the solenoid and zigzag 30 nm fiber models.15,16 A final open question is how post-translational modifications, such as the acetylation of histone tails, can modulate the phase separation of chromatin across various length scales.
Simulations might be expected to provide insight into these questions, yet the many length and time scales that characterize chromatin phase separation make such simulations difficult. Recent efforts to understand chromatin can generally be classified as chemically specific models which are limited to low chromatin densities that do not phase separate17−19 or as highly coarse-grained models that lack the chemical details necessary to elucidate the subtle molecular forces that regulate chromatin phase separation.20−28 Multiscale models that systematically combine chemically specific and coarse-grained models are an attractive strategy to address these challenges,29,30 yet efforts to apply these models to chromatin phase separation have only recently been pursued.31 As a consequence, a detailed understanding of how molecular interactions give rise to the physical properties of phase-separated chromatin has yet to be established.
In this work, we investigate the structure and properties of phase-separated chromatin using a multiscale model of chromatin. Our results indicate that chromatin phase separation is driven by heterogeneous self-interactions which give rise to liquid-like behaviors on short length and time scales. However, we find that chromatin’s material state is sensitive to post-translational modifications and that solid-like chromatin can form when internucleosomal interactions are increased. Our results also show that chromatin condensates lack a regular mesoscale structure like the solenoid and zigzag 30 nm fiber models, and that instead chromatin assumes a more disordered and irregular structure. Lastly we examine how H4 tail acetylation disrupts chromatin condensates and how H4 acetylation can decompact chromatin while still preserving its ability to phase separate. Taken together, our results provide a molecular view into chromatin condensates and how chromatin’s self-interactions can establish its organization.
Methods
Simulating Chromatin Condensates
We use the 1CPN model29 to simulate nucleosomal arrays and their phase separation. 1CPN is a coarse-grained, multiscale model that models linker DNA as a negatively charged twistable worm-like chain and nucleosomes as anisotropic sites with implicit histone tail interactions. These histone tail interactions are incorporated into 1CPN through coarse-grained potentials whose parameters are obtained using detailed free energy calculations from the near-atomistic 3SPN-AICG nucleosome model32−37 (see ref. (29) and details below). This multiscale procedure permits the 1CPN model to be efficient enough to perform megabase-scale simulations of genomic DNA while retaining biochemical details such as post-translational modifications. 1CPN has been to shown to predict chromatin’s mesoscale structural features like tetranucleosomal folding motifs38,39 and is compatible with a recent model of the linker histone.40 Within 1CPN, electrostatics are treated using Debye–Hückel theory and the solvent is treated implicitly using Langevin dynamics.
Because 1CPN is parametrized using detailed 3SPN-AICG free energy calculations, the 1CPN model can be used to examine specific DNA sequences and post-translational modifications. In this work we use the strongly binding 601 DNA sequence and both unmodified nucleosomes and those with acetylated H4 tails. Specifically, the 601 DNA sequence is incorporated into 1CPN by matching its free energy profile for DNA’s rotation around the nucleosome to a profile calculated using 3SPN-AICG. Similarly, H4 tail acetylation is simulated by first computing anisotropic nucleosome pair potentials using the 3SPN-AICG model37 and then fitting 1CPN’s interaction parameters so that these potentials are reproduced.29 Additional details of this fitting process are provided in the Supporting Information.
We simulate the phase separation of nucleosomal arrays using the so-called direct coexistence method where both chromatin-dense and chromatin-dilute phases are allowed to equilibrate within a single simulation box. In order to mitigate the effects of the interface between phases, we adopt a slab-shaped simulation box which has been shown to reduce the finite-size effects present in direct coexistence simulations.41 To initialize our simulations, we first place nucleosomal arrays randomly in a large cubic simulation box and then compress the box until the x- and y-dimensions of the slab-shaped box are reached. The simulation box is then expanded along its z-dimension to form a slab-shaped box and the simulation is then run until the dense and dilute phase concentrations equilibrate. We confirmed that these concentrations were not influenced by any of the parameters used in our initialization procedure (see Supporting Information). For bulk simulations of the chromatin-dense phase (i.e., chromatin condensate), we use a cubic simulation box and a nucleosome concentration equal to that of the chromatin-dense phase.
Classifying Liquid- and Solid-Like Chromatin Condensates
Chromatin condensates are viscoelastic materials whose liquid- and solid-like properties depend on the length and time scales over which they are observed. In order to classify whether chromatin condensates are liquid- or solid-like we use two complementary metrics. For the first, we calculate the mean-squared displacement (MSD) of nucleosomes as a function of time:42
| 1 |
where  is the
position vector of the ith nucleosome at time t and N is
the total number of nucleosomes. By examining the slope of the MSD
for a specified lag time, chromatin’s material state at different
time scales can be obtained. For example, a MSD which scales linearly
with time (indicated by a slope of 1 on a log–log plot) denotes
freely diffusive nucleosomes characteristic of liquid-like chromatin.
Alternatively, MSDs with slopes less than one denote subdiffusive
nucleosome dynamics and more solid-like behavior.
is the
position vector of the ith nucleosome at time t and N is
the total number of nucleosomes. By examining the slope of the MSD
for a specified lag time, chromatin’s material state at different
time scales can be obtained. For example, a MSD which scales linearly
with time (indicated by a slope of 1 on a log–log plot) denotes
freely diffusive nucleosomes characteristic of liquid-like chromatin.
Alternatively, MSDs with slopes less than one denote subdiffusive
nucleosome dynamics and more solid-like behavior.
The second
metric we use to quantify the material state of chromatin examines
the lifetime of nucleosome–nucleosome contacts within a condensate.
To define this metric, we first specify that two nucleosomes with
positions ri and rj are in contact if their spatial separation  is less than the cutoff
is less than the cutoff  Å.
We then measure how these contacts
are disrupted over time. Specifically, we define the fraction of persistent
contacts P(t) as
Å.
We then measure how these contacts
are disrupted over time. Specifically, we define the fraction of persistent
contacts P(t) as
 |
2 |
where T0 is the set of nucleosomes in contact with another nucleosome at time t = 0 and T is the set of nucleosomes which have persisted in their contact since t = 0. The time scale over which P(t) decays can be used to quantify the lifetimes of nucleosome–nucleosome contacts and corresponds to a transition from solid-like to liquid-like behavior. For both of these metrics time is given in units of τ, the time scale over which a 12-mer nucleosomal array with a linker DNA length of 25 bp will diffuse its radius of gyration in dilute conditions. This calculation and a further interpretation of τ is given in the Supporting Information.
Characterizing Structures within Chromatin Condensates
Chromatin
structures such as the solenoid and zigzag 30 nm fiber
models can be characterized by how neighboring nucleosomes interact.
To quantify these interactions, we first define a pair of nucleosomes
to be kth neighbors if they exist on the same nucleosomal
array and have k – 1 nucleosomes between them.
We then define the orientation of two nucleosomes i and j using three angles: α which is the
angle between the unit vectors normal to each nucleosome’s
top face  and
and  , βi which
is the angle between
, βi which
is the angle between  and the unit vector connecting the nucleosomes’
centers
and the unit vector connecting the nucleosomes’
centers  , and βj which is the angle between
, and βj which is the angle between  and
and  . Using these definitions, we then classify
nucleosome interactions by their different types: “Face–face”,
“Face–side”, or “Side–side”
as described previously31 (see Table 1). Face–face
type nucleosome interactions between first and second neighbors are
characteristic of solenoid and zigzag 30 nm fiber models respectively,43 therefore their frequency quantifies the presence
of these 30 nm fiber models within our simulations.
. Using these definitions, we then classify
nucleosome interactions by their different types: “Face–face”,
“Face–side”, or “Side–side”
as described previously31 (see Table 1). Face–face
type nucleosome interactions between first and second neighbors are
characteristic of solenoid and zigzag 30 nm fiber models respectively,43 therefore their frequency quantifies the presence
of these 30 nm fiber models within our simulations.
Table 1. Definitions for Nucleosome Interaction Types.
| Nucleosome interaction type | Angles between nucleosomes |
|---|---|
| face–face |  |
 or or 
|
|
| face–side |  |
| side–side |  |
 and and 
|
We also quantify the structure of nucleosomal arrays by measuring their diameters and linear packing densities. To do this, we approximate each array’s shape with a cylinder by using the following procedure. First, we calculate a nucleosomal array’s radius of gyration tensor S:44
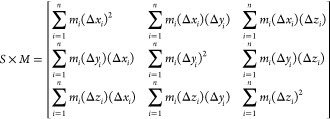 |
3 |
where M is
the total mass of the array, mi is the mass of an individual site i, n is the array’s number of sites, and Δxi is the distance between site i and the array’s center of mass along the x-axis, and similarly for the y- and z-axes. Once S is computed with eq 3, we then decompose S into eigenvectors v1, v2, and v3 with corresponding
eigenvalues λ1, λ2, and λ3 where  , and then define v1 as the central axis of the approximating cylinder.
Finally
the cylinder’s length is then determined by the minimum and
maximum positions of the array’s sites along its central axis,
and the cylinder’s diameter is set equal to 2 times the maximum
distance of the array’s sites from its central axis. When performing
this procedure, we found that only using the single nucleosome site
present in 1CPN underestimated the excluded volume of the nucleosome.
This issue was easily rectified by introducing additional ghost sites
which collectively represent the nucleosome’s excluded volume
more faithfully (see Supporting Information).
, and then define v1 as the central axis of the approximating cylinder.
Finally
the cylinder’s length is then determined by the minimum and
maximum positions of the array’s sites along its central axis,
and the cylinder’s diameter is set equal to 2 times the maximum
distance of the array’s sites from its central axis. When performing
this procedure, we found that only using the single nucleosome site
present in 1CPN underestimated the excluded volume of the nucleosome.
This issue was easily rectified by introducing additional ghost sites
which collectively represent the nucleosome’s excluded volume
more faithfully (see Supporting Information).
The last metric we use to quantify chromatin’s structure within condensates is the nucleosome–nucleosome structure factor S(k), which is obtained by performing a Fourier transform on the nucleosome–nucleosome radial distribution function g(r) (see ref. (45)):
| 4 |
where r is a given radial distance, k is the magnitude of a given scattering vector, and ρ is the bulk nucleosome density. The magnitude of a scattering vector corresponds to a radial distance of r = 2π/k, meaning that a structure factor peak at k = 2 indicates that nucleosome positions are correlated over distances of r = π.
Results and Discussion
Before interrogating the structure and thermodynamic properties of chromatin condensates, our first step is to validate our model and confirm that model predictions are consistent with existing experimental measurements. We validate our model using experimental data from Gibson et al. where dilute solutions of short nucleosomal arrays are observed to undergo phase separation. This seminal work has comprehensively mapped out the phase diagram of short nucleosomal arrays and has examined how phase boundaries can be modulated by numerous factors such as linker DNA length, salt concentration, the presence of linker histone H1, and histone acetylation. If our model can reproduce this wide range of experimental data, then we can have increased confidence in other model predictions for which experimental data is not available.
The first validation of our model focuses on experimental data from Gibson et al. which indicates that longer linker DNA lengths weaken chromatin phase separation, leading to lower concentrations of nucleosomes within the chromatin condensate. Gibson et al.11 also observe that the addition of linker histone H1 counteracts this effect and results in condensate nucleosome concentrations that are insensitive to linker DNA length. To examine if our model can reproduce these observations, we perform simulations that mirror the conditions used in the experiments: we simulate 12-mer nucleosomal arrays in solution with different linker lengths, in the presence and absence of linker histones, and examine whether these arrays undergo phase separation. If phase separation occurs, the nucleosome concentration within the condensate is measured. We find that our model predictions are in good agreement with experimental measurements (Figure 1). Our model accurately predicts that the nucleosome concentration within condensates decreases for larger linker lengths in the absence of H1 but is approximately constant if H1 is present.
Figure 1.
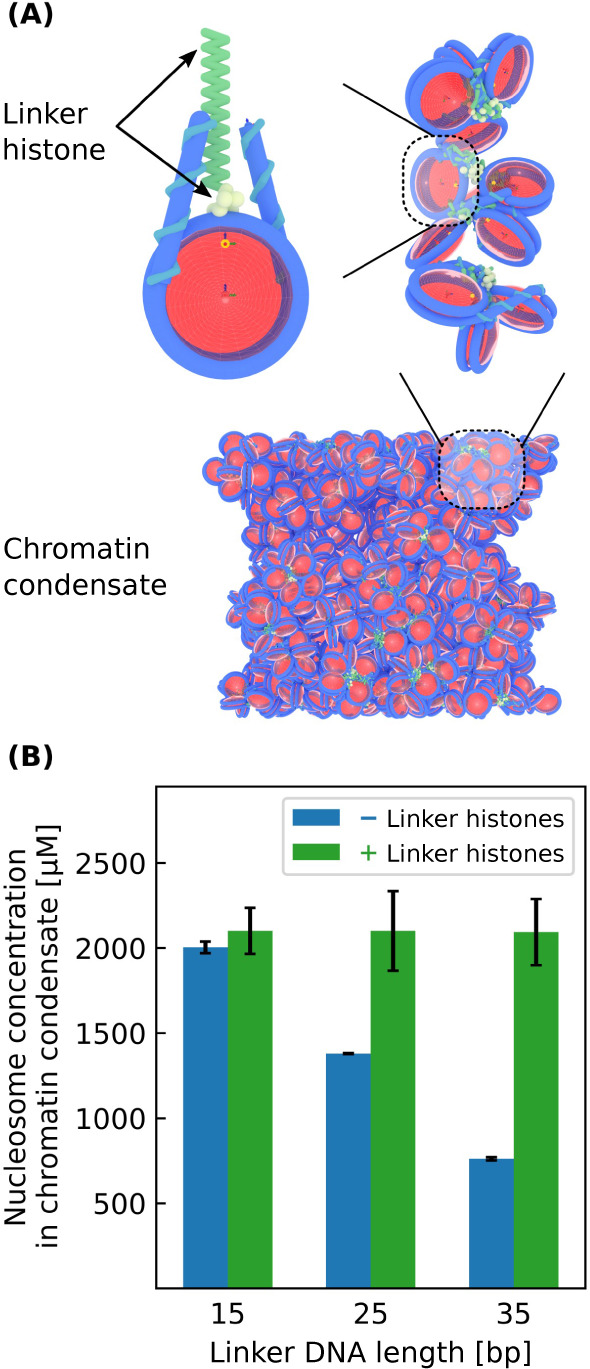
Phase separation of 12-mer nucleosomal arrays with varying linker DNA lengths with and without linker histones. (A) Model visualization of a chromatin condensate containing linker histones. (B) Nucleosome concentration within various chromatin condensates. Each simulation has a bulk nucleosome concentration of 200 μM and a box geometry of 100 × 100 × 1000 nm.
The next validation of our model focuses on other data from Gibson et al. where it was observed that lower salt concentrations and shorter nucleosomal arrays lengths (i.e., having fewer nucleosomes per array) weaken chromatin phase separation. To test if our model can reproduce these results, we use our model to calculate the phase diagram for nucleosomal arrays with varying salt concentrations and nucleosomal array lengths. In these simulations the linker DNA lengths are fixed at 25 bp and nucleosome–nucleosome interaction strengths are assumed to be independent of salt concentration. Our model predictions agree well with experiments: while 12-mer solutions at high salt concentrations readily phase separate, solutions with salt concentrations less than 70 mM are incapable of phase separation (Figure 2A). Similarly, while 12-mer nucleosomal arrays are capable of phase separation at a salt concentration of 150 mM, shorter 4-mer arrays are not (Figure 2B).
Figure 2.

Phase separation of nucleosomal arrays under varying salt concentrations and nucleosomal arrays lengths. (A) Phase diagram with respect to salt concentration for 12-mer nucleosomal arrays alongside experimental results. Simulations with 70 mM of salt or less do not phase separate. (B) Phase diagram with respect to nucleosomal array length for nucleosomal arrays with a salt concentration of 150 mM alongside experimental results. Simulations with 6 nucleosomes per array or less do not phase separate. Each simulation has a bulk nucleosome concentration of 200 μM and a box geometry of 100 × 100 × 1000 nm. Experimental data is from ref. (11).
We emphasize that these simulation results are purely predictive and that no modifications were made to the originally published 1CPN model.29 As a consequence, the good agreement between simulation and experiment is quite remarkable, especially when considering that the conditions within a chromatin condensate are very different from the dilute conditions under which 1CPN was originally parametrized.29 This parameter-free validation of our model gives us confidence that the 1CPN model can accurately predict the phase separation of chromatin condensates and that 1CPN can be used to provide molecular insights into their structural and material properties.
It is also noteworthy that the predictions of our 1CPN model are in good agreement with other recent simulations of chromatin phase separation.31 This recent work uses a model similar in spirit to 1CPN and also relies on a multiscale approach to systematically incorporate the molecular details of chromatin into increasingly coarse-grained models. Though this past work used a slightly shorter DNA linker length than we have used here (18 bp versus 25 bp), the critical salt concentration predicted by this model of 69 mM is remarkably close to the value of about 70 mM predicted by the 1CPN model (see Figure 2). Though the nucleosome concentrations predicted by the 1CPN model are in better agreement with Gibson et al., the general agreement between these two models suggests that nucleosomal breathing is adequately captured in the 1CPN model so as to permit the weak multivalent interactions that are necessary to drive chromatin phase separation.
Liquid- and Solid-Like Chromatin Condensates
Now that we have validated our model, we next turn to examine the conditions under which chromatin exhibits liquid- or solid-like material properties. Experiments have shown chromatin to be liquid-like,11,13,14 or solid-like,12,46,47 in various in vitro and in vivo systems, and these experiments collectively demonstrate that chromatin has a material state dependent on the length and time scales considered (i.e., chromatin is viscoelastic).48 Chromatin’s material state is likely important to nuclear processes like transcription, but its significance and molecular origins are still poorly understood.
In order to examine how molecular interactions establish chromatin’s material properties, we focus on the role of internucleosomal interactions which can be modulated by numerous factors such as solution conditions or post-translational modifications.37,49−52 While we will examine the importance of specific post-translational modifications in a later subsection, we begin here by examining the role of internucleosomal interactions at a coarse-grained level by varying the ϵ0 parameter in the 1CPN model (Figure 3A), which scales the overall strength of the anisotropic interactions between nucleosomes (see eq 5 in ref. (29)).
Figure 3.

Liquid- and solid-like chromatin condensates. (A) Model visualization of the nucleosome binding energy ϵ0 which scales all internucleosomal interactions. (B) Phase diagram with respect to ϵ0 for 12-mer nucleosomal arrays with linker DNA lengths of 25 bp. ϵ0 has a default value of 2.0 kBT in our model. (C) Mean-squared displacement (MSD) over time for nucleosomes within different condensates. Liquid-like condensates have diffusive nucleosomes which linearly scale their MSD with time. (D) Fraction of persistent nucleosome contacts over time within different condensates. When this fraction equals 0, all initial nucleosome–nucleosome contacts are broken. τ is the time it takes a nucleosomal array simulated here to diffuse its radius of gyration in dilute conditions and is equal to 40 μs. All simulations have a salt concentration of 150 mM. Simulations in (B) have bulk nucleosome concentrations of 300 μM and box geometries of 80 × 80 × 1000 nm. Simulations in (C) and (D) have bulk nucleosome concentrations equal to the concentrations within their corresponding chromatin-dense phase found in (B) and each contain 100 nucleosomal arrays.
To examine the effect of ϵ0 on chromatin’s material properties, we first compute the phase diagram for nucleosomal arrays as a function of ϵ0 (Figure 3B). As anticipated, the phase diagram is very sensitive to ϵ0, with increasing values of ϵ0 resulting in a wider phase diagram and more strongly segregated chromatin. Notably, the phase diagram has a critical point of about 1.7 kBT which indicates that relatively weak internucleosomal interactions are sufficient to drive chromatin phase separation.
We next observe that the
material state of chromatin is also very
sensitive to ϵ0. Using eq 1, we characterize chromatin’s material
state within different condensates by computing the mean-squared displacement
(MSD) over time of their nucleosomes (Figure 3C). Initially the MSD scales sublinearly
with time which denotes subdiffusive nucleosomes typical of a solid-like
chromatin state. As time progresses, the MSD eventually scales linearly
with time which indicates liquid-like behavior. Over the ∼40
ms time scale of our simulations, we find that condensates with  exhibit liquid-like behavior,
while condensates
with
exhibit liquid-like behavior,
while condensates
with  remain
solid-like. We also find that the
mobility of nucleosomes differs considerably between the two types
of condensates and that solid-like nucleosomes with
remain
solid-like. We also find that the
mobility of nucleosomes differs considerably between the two types
of condensates and that solid-like nucleosomes with  displace their size (∼11 nm) during
their simulation.
displace their size (∼11 nm) during
their simulation.
It is possible that diffusive nucleosomes have long-lived contacts characteristic of a solid-like material, and so we check for this possibility by measuring the lifetime of nucleosome–nucleosome contacts within the condensates (Figure 3D). We measure this lifetime by calculating the fraction of persistent contacts over time with eq 2, which equals zero when all initial contacts are broken (see Classifying Liquid- and Solid-Like Chromatin Condensates). Figure 3C,D shows that the transition from subdiffusive to diffusive nucleosomes coincides with the termination of initial nucleosome–nucleosome contacts, and so we conclude that diffusive nucleosomes here characterize a liquid-like chromatin state.
Overall we find that internucleosomal interactions play a major role in establishing chromatin’s material properties. For example, by increasing internucleosomal interactions by a mere 2 kBT, we observe that chromatin which was initially liquid-like will become solid-like (Figure 3). Experimentally, the precise strength of internucleosomal interactions is the subject of some debate.53−58 Some recent experiments have estimated internucleosomal attractions to be at most 2.7 kBT,57 which based on our results suggests that short segments of unmodified chromatin are liquid-like in agreement with some recent experiments,11,14,59 but not others.12 Since chromatin’s material state is sensitive to ϵ0, it is likely that changes to ϵ0 through post-translational modifications can modulate the time scales over which chromatin is liquid- or solid-like. Such regulation is likely important for transcription which occurs over similar time scales of 10–100 ms per nucleotide.60
Mesoscale Structure of Chromatin Condensates
We now turn to characterize chromatin’s mesoscale structure within chromatin condensates. Our first objective is to investigate whether solenoid or zigzag 30 nm fibers61 are present in our simulations. These structures can be identified by the way their neighboring nucleosomes interact43 and so we begin by analyzing these interactions. To carry out this analysis, we first perform a bulk simulation of a liquid-like chromatin condensate (Figure 4A) and then measure the frequency of different interaction types between first and second nucleosome neighbors (Figure 4B). Our results indicate that liquid-like chromatin contains mostly face–side interactions between first and second neighbors and that face–face interactions rarely occur. Since both solenoid and zigzag models are characterized by sustained face–face interactions, Figure 4B shows that neither solenoid nor zigzag fibers are present within liquid-like chromatin condensates.
Figure 4.

Interactions
between neighboring nucleosomes for liquid-like chromatin
( ). (A) Model visualization of a bulk chromatin
condensate simulation (top) and a cartoon depicting nucleosome neighbors
(bottom). (B) Relative frequencies of different interaction types
between neighboring nucleosomes. Visualizations of each interaction
type are shown above their respective values. Neighboring nucleosomes
more than 150 Å apart are negligibly interacting and have interaction
type “No contact”. The bulk condensate simulation contains
100 12-mer nucleosomal arrays having linker DNA lengths of 25 bp with
a salt concentration of 150 mM and bulk nucleosome concentration of
1400 μM.
). (A) Model visualization of a bulk chromatin
condensate simulation (top) and a cartoon depicting nucleosome neighbors
(bottom). (B) Relative frequencies of different interaction types
between neighboring nucleosomes. Visualizations of each interaction
type are shown above their respective values. Neighboring nucleosomes
more than 150 Å apart are negligibly interacting and have interaction
type “No contact”. The bulk condensate simulation contains
100 12-mer nucleosomal arrays having linker DNA lengths of 25 bp with
a salt concentration of 150 mM and bulk nucleosome concentration of
1400 μM.
We next characterize chromatin’s mesoscale structure directly by fitting each nucleosomal array to a cylinder of minimal volume (see Characterizing Structures within Chromatin Condensates). This fitting allows us to quantify the diameter of each array and the corresponding distribution of array diameters throughout a chromatin condensate (Figure 5A). Surprisingly, we observe that the average array diameter is around 30 nm despite the previous lack of evidence for 30 nm fiber models. However, the distribution of array diameters is quite broad which indicates that chromatin’s mesoscale structure is irregular throughout the liquid-like condensate. To better understand this broad distribution, we examine the linear packing densities of nucleosomal arrays which measures how tightly compacted each array is (Figure 5B). The arrays’ packing densities are also broadly distributed and the average packing density is roughly half that of either 30 nm fiber model. Together these distributions show that liquid-like chromatin has an irregular mesoscale organization different from the 30 nm fiber models. These results suggest that chromatin’s mesoscale structure is instead better described as a disordered polymer chain which is qualitatively consistent with chromatin electron microscopy tomography (ChromEMT) experiments.62
Figure 5.
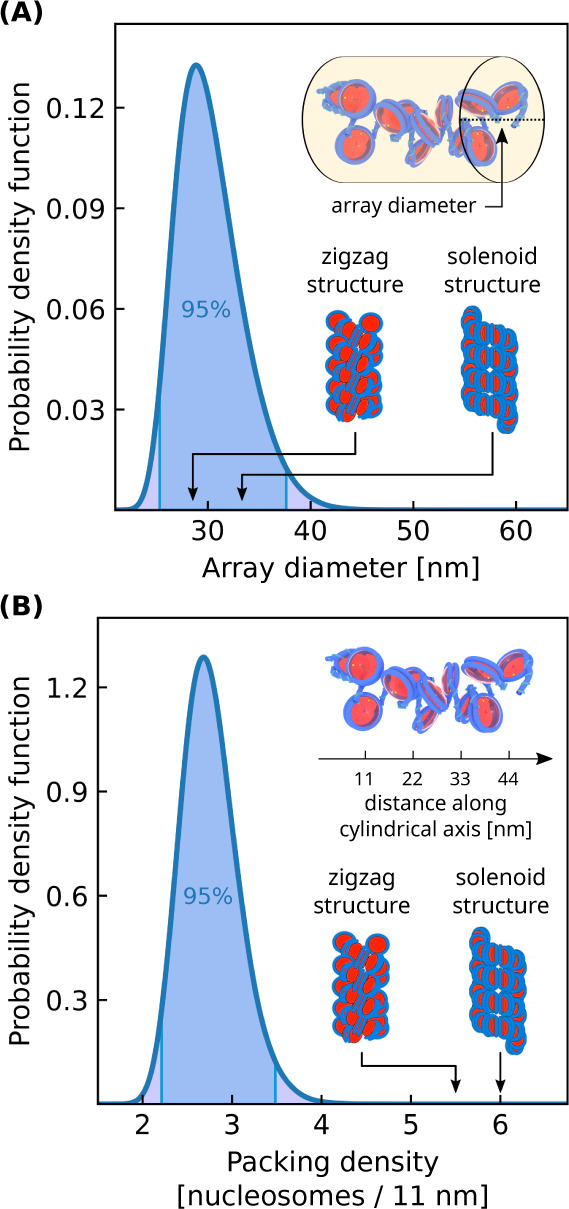
Nucleosomal array structures
within a liquid-like chromatin condensate
( ). (A) Distribution of array diameters.
(B) Distribution of array packing densities. The packing density is
defined as the number of nucleosomes per 11 nm along the array’s
central axis. Values for solenoid and zigzag 30 nm fiber models are
indicated using illustrations from ref. (63). The bulk condensate simulation contains 100
12-mer nucleosomal arrays having linker DNA lengths of 25 bp with
a salt concentration of 150 mM and bulk nucleosome concentration of
1400 μM. Reproduced or adapted with permission from ref (63). Copyright 2019 Elsevier.
). (A) Distribution of array diameters.
(B) Distribution of array packing densities. The packing density is
defined as the number of nucleosomes per 11 nm along the array’s
central axis. Values for solenoid and zigzag 30 nm fiber models are
indicated using illustrations from ref. (63). The bulk condensate simulation contains 100
12-mer nucleosomal arrays having linker DNA lengths of 25 bp with
a salt concentration of 150 mM and bulk nucleosome concentration of
1400 μM. Reproduced or adapted with permission from ref (63). Copyright 2019 Elsevier.
Finally, we examine how the mesoscale structure
of chromatin changes
as a condensate varies from liquid- to solid-like. To perform this
analysis, we first conduct bulk simulations of chromatin condensates
with different internucleosomal interaction strengths scaled by ϵ0, which we previously found to change chromatin’s material
state (see Figure 3). We then use eq 4 to
calculate the nucleosome–nucleosome structure factor of each
condensate (Figure 6). For liquid-like condensates ( ), the structure factor is peaked at 8.5
nm and shows no peak at 30 nm. The peak at 8.5 nm corresponds to frequent
face–side nucleosome interactions which occur over distances
of 7–11 nm and is consistent with our observations in Figure 4B. The absence of
a peak at 30 nm is also consistent with the broad distribution of
array diameters that we observed in Figure 5A. As ϵ0 is increased and
chromatin becomes more solid-like, the peak in the structure factor
shifts to smaller distances and is located at 7.1 nm for
), the structure factor is peaked at 8.5
nm and shows no peak at 30 nm. The peak at 8.5 nm corresponds to frequent
face–side nucleosome interactions which occur over distances
of 7–11 nm and is consistent with our observations in Figure 4B. The absence of
a peak at 30 nm is also consistent with the broad distribution of
array diameters that we observed in Figure 5A. As ϵ0 is increased and
chromatin becomes more solid-like, the peak in the structure factor
shifts to smaller distances and is located at 7.1 nm for  . This indicates that stronger internucleosomal
interactions result in more compact chromatin structures, consistent
with Figure 3B. Even
at larger values of ϵ0, we see no peak at 30 nm which
suggests that no regularly ordered 30 nm fibers are present within
our simulations. We also observe that larger values of ϵ0 decrease chromatin’s isothermal compressibility which
is equal to the structure factor S(k) as k → 0.64
. This indicates that stronger internucleosomal
interactions result in more compact chromatin structures, consistent
with Figure 3B. Even
at larger values of ϵ0, we see no peak at 30 nm which
suggests that no regularly ordered 30 nm fibers are present within
our simulations. We also observe that larger values of ϵ0 decrease chromatin’s isothermal compressibility which
is equal to the structure factor S(k) as k → 0.64
Figure 6.

Nucleosome–nucleosome
structure factor for liquid- and solid-like
chromatin condensates. Structure factor peaks indicate correlated
nucleosome positions over the given reciprocal scattering vector length
2π/k. Nucleosome binding energies  correspond to liquid-like condensates
while
correspond to liquid-like condensates
while  correspond
to solid-like condensates. Each
bulk condensate simulation contains 100 12-mer nucleosomal arrays
having linker DNA lengths of 25 bp with a salt concentration of 150
mM and bulk nucleosome concentration equal to that of a condensate
with nucleosome binding energy ϵ0.
correspond
to solid-like condensates. Each
bulk condensate simulation contains 100 12-mer nucleosomal arrays
having linker DNA lengths of 25 bp with a salt concentration of 150
mM and bulk nucleosome concentration equal to that of a condensate
with nucleosome binding energy ϵ0.
By further increasing internucleosomal interactions, our results suggest that the face–face interactions of 30 nm fiber models will eventually become favorable. Such highly attractive internucleosomal interactions have been explored in experiments of nucleosomal arrays with superphysiological concentrations of divalent salts.14,43 In agreement with our model predictions, these nucleosomal arrays were shown to phase separate into solid-like condensates14 and have frequent face–face interactions.43 Interestingly, these arrays did not regularly form into 30 nm fiber models despite their frequent face–face interactions.43 This may be explained by the 60 bp linker DNA lengths of the arrays which are much longer than the 25 bp linker lengths considered here. Longer linker DNA lengths drive chromatin’s structure to be more irregular and flexible38 and so altogether it is likely that the formation of 30 nm fibers is sensitive to both the nuclear environment and chromatin’s composition. This sensitivity can explain experimental results showing that 30 nm fibers are rare occurrences in human chromatin65 yet common in chicken erythrocytes.66
We have also simulated chromatin condensates with 20 bp linker lengths and have analyzed their mesoscale structure in the presence and absence of 2 mM of divalent salt (see Supporting Information). Though nucleosomal arrays with 20 bp linker lengths have been shown to form zigzag-like fibers in similar dilute conditions,67 our simulations show that in dense condensates these arrays have a mesoscale structure like a disordered polymer chain consistent with our 25 bp results presented in Figures 4–6. We also observe that removing 2 mM of divalent salt results in small changes to chromatin’s mesoscale structure (see Supporting Information). This finding agrees well with experiments showing that divalent salts weakly alter chromatin’s mesoscale structure when physiological concentrations of monovalent salts are present.68 Divalent salts may therefore predominately alter the material state of condensed chromatin, as shown experimentally14 and in Figure 3, rather than its mesoscale structure.
H4 Acetylated Chromatin Condensates
We now turn to examine how specific post-translational modifications modulate chromatin phase separation. We focus here on histone acetylation which has been shown to weaken chromatin’s self-interactions37,57 and inhibit the phase separation of 12-mer nucleosomal arrays.11 Since chromatin’s H4 tails are essential for its compaction,49 we focus on how the acetylation of H4 tails (here referred to as H4 acetylation) can modulate chromatin phase separation.
In order to simulate H4 acetylated chromatin, we first compute the orientation-dependent pair potential between two H4 acetylated nucleosomes using the near-atomistic 3SPN-AICG model (Figure 7A). This orientation-dependent pair potential is then used to reparametrize the 1CPN model so that it can incorporate the subtle effects of H4 acetylation (see Figure 7B,C and Simulating Chromatin Condensates). We note that this multiscale approach to incorporate post-translational modifications is parameter-free and provides a systematic route to include chemically specific information into the 1CPN model.
Figure 7.

H4 tail acetylation of chromatin condensates. (A) The near-atomistic 3SPN-AICG nucleosome model was used to obtain orientation-dependent pair potentials which parametrize the coarse-grained 1CPN model. The orientations shown are (I) side–side, (II) face–face, and (III) face–side. (B) Near-atomistic and coarse-grained 1CPN nucleosome pair potentials for unmodified chromatin and (C) H4 acetylated chromatin. (D) Simulations of unmodified 12-mer nucleosomal arrays and H4 acetylated 12-mer/60-mer arrays with linker DNA lengths of 25 bp. (E) Phase diagram with respect to nucleosomal array length for unmodified and H4 acetylated chromatin.
We first apply our model of H4 acetylation to solutions of 12-mer nucleosomal arrays. Recent experiments have observed that this system should undergo phase separation when chromatin is unmodified but should not when chromatin’s H4 tails are partially acetylated.11 Our model predictions are in agreement with these experiments and accurately predict that H4 acetylation is sufficient to disrupt the phase separation of 12-mer arrays (Figure 7D). Our model also predicts that the phase separation of H4 acetylated chromatin can be restored by increasing its length to 60-mer arrays (Figure 7D). This reemergence of phase separation for longer arrays can be explained by the longer arrays’ lower entropy of mixing which favors phase separation.69 The ability of longer stretches of acetylated chromatin to reinitiate phase separation suggests that chromatin phase separation in living cells is likely a multifaceted process, where both the type and density of post-translational modifications can be used to tune phase separation.
To quantify the effect of H4 acetylation on chromatin phase separation, we compute a phase diagram with respect to array length for H4 acetylated chromatin and compare it to unmodified chromatin (Figure 7E). We find that H4 acetylation shifts the phase diagram to longer array lengths, and that H4 acetylated chromatin requires 40-mer arrays to phase separate while unmodified chromatin only requires 7-mer arrays. We also find that H4 acetylation results in a lower concentration of nucleosomes in the chromatin-dense phase (650–850 μM) which is approximately half the concentration observed in unmodified chromatin (1050–1400 μM). Taken together, our results indicate that even though H4 acetylation can decompact chromatin,59 this decompaction does not exclude the possibility that acetylated chromatin can phase separate, especially for long stretches of chromatin. This result helps explain recent evidence suggesting that acetylated chromatin regions are liquid-like, phase separated domains47 and further shows how phase separation can play a major role in chromatin’s organization.
Conclusions
In this work we examine chromatin condensates composed of nucleosomal arrays by utilizing a multiscale, coarse-grained chromatin model. Our model can reproduce key experimental observations of these condensates including their dependence on linker histones, salt concentration, linker DNA length, and array length. Using this model, we have demonstrated that the material state of chromatin condensates can be modulated by the strength of their nucleosome–nucleosome interactions. Based on experimental estimates of chromatin’s nucleosome–nucleosome interactions,57 our model predicts that unmodified chromatin is liquid-like on short length scales in agreement with experiments.11,13,14 Like other viscoelastic materials, chromatin transitions from solid- to liquid-like behavior depending on the length and time scale considered47,48 and our results demonstrate that post-translational modifications can regulate this transition for chromatin.
Our work also provides a molecular view into chromatin’s mesoscale structure which regulates gene expression yet is poorly defined.16 Within chromatin condensates, our results show that chromatin lacks a regular secondary structure like the solenoid and zigzag 30 nm fiber models. Instead chromatin is structured irregularly and frequently self-interacts through heterogeneous nucleosome–nucleosome interactions as also observed experimentally.62 We additionally show that chromatin’s mesoscale structure becomes more regular as chromatin’s self-interactions are increased, which can occur under superphysiological concentrations of divalent cations43 or when nucleosomes are more closely spaced.38
Lastly our work investigates whether H4 tail acetylation can prevent chromatin phase separation. In agreement with experiments,11 we find that H4 acetylation dissolves chromatin condensates composed of short, 12-mer nucleosomal arrays. However, we also observe that condensates of acetylated 40-mer arrays can reinitiate phase separation, albeit at a lower degree of compaction relative to unmodified chromatin. This result reveals that acetylation’s decompaction of chromatin is compatible with chromatin phase separation and also exemplifies how phase separation may play a major role in chromatin’s organization. Collectively, our work demonstrates that multiscale, coarse-grained models are a powerful tool for probing the structural and dynamic properties of in vitro chromatin systems.
Acknowledgments
A.G. was supported by the Department of Education under Award No. P200A180026. This work was also partially supported by a New Investigator Grant from the Charles E. Kaufman Foundation. Simulations were run on hardware supported by Drexel’s University Research Computing Facility which is partially funded by NSF OAC Major Research Infrastructure grant #1919691.
Supporting Information Available
The Supporting Information is available free of charge at https://pubs.acs.org/doi/10.1021/acs.jpcb.4c04420.
Additional details for simulating chromatin condensates with 1CPN; validation of our simulation box geometry and Langevin thermostat; calculation of the characteristic time scale τ used in our analysis; structural analysis of chromatin condensates with 20 bp linker DNA lengths; fitting process for 1CPN’s nucleosome pair potentials; visualization of ghost sites used in our cylindrical approximation (PDF)
The authors declare no competing financial interest.
Supplementary Material
References
- Maeshima K.; Iida S.; Tamura S. Physical nature of chromatin in the nucleus. Cold Spring Harbor Perspect. Biol. 2021, 13, a040675. 10.1101/cshperspect.a040675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Misteli T. The self-organizing genome: principles of genome architecture and function. Cell 2020, 183, 28–45. 10.1016/j.cell.2020.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rippe K. Liquid–liquid phase separation in chromatin. Cold Spring Harbor Perspect. Biol. 2022, 14, a040683. 10.1101/cshperspect.a040683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hildebrand E. M.; Dekker J. Mechanisms and functions of chromosome compartmentalization. Trends Biochem. Sci. 2020, 45, 385–396. 10.1016/j.tibs.2020.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feric M.; Misteli T. Phase separation in genome organization across evolution. Trends Cell Biol. 2021, 31, 671–685. 10.1016/j.tcb.2021.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lafontaine D. L.; Riback J. A.; Bascetin R.; Brangwynne C. P. The nucleolus as a multiphase liquid condensate. Nat. Rev. Mol. Cell Biol. 2021, 22, 165–182. 10.1038/s41580-020-0272-6. [DOI] [PubMed] [Google Scholar]
- Larson A. G.; Elnatan D.; Keenen M. M.; Trnka M. J.; Johnston J. B.; Burlingame A. L.; Agard D. A.; Redding S.; Narlikar G. J. Liquid droplet formation by HP1α suggests a role for phase separation in heterochromatin. Nature 2017, 547, 236–240. 10.1038/nature22822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strom A. R.; Emelyanov A. V.; Mir M.; Fyodorov D. V.; Darzacq X.; Karpen G. H. Phase separation drives heterochromatin domain formation. Nature 2017, 547, 241–245. 10.1038/nature22989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L.; Gao Y.; Zheng X.; Liu C.; Dong S.; Li R.; Zhang G.; Wei Y.; Qu H.; Li Y. Histone modifications regulate chromatin compartmentalization by contributing to a phase separation mechanism. Mol. Cell 2019, 76, 646–659. 10.1016/j.molcel.2019.08.019. [DOI] [PubMed] [Google Scholar]
- Erdel F.; Rademacher A.; Vlijm R.; Tünnermann J.; Frank L.; Weinmann R.; Schweigert E.; Yserentant K.; Hummert J.; Bauer C. Mouse heterochromatin adopts digital compaction states without showing hallmarks of HP1-driven liquid-liquid phase separation. Mol. Cell 2020, 78, 236–249. 10.1016/j.molcel.2020.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson B. A.; Doolittle L. K.; Schneider M. W.; Jensen L. E.; Gamarra N.; Henry L.; Gerlich D. W.; Redding S.; Rosen M. K. Organization of chromatin by intrinsic and regulated phase separation. Cell 2019, 179, 470–484. 10.1016/j.cell.2019.08.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strickfaden H.; Tolsma T. O.; Sharma A.; Underhill D. A.; Hansen J. C.; Hendzel M. J. Condensed chromatin behaves like a solid on the mesoscale in vitro and in living cells. Cell 2020, 183, 1772–1784. 10.1016/j.cell.2020.11.027. [DOI] [PubMed] [Google Scholar]
- Chen Q.; Zhao L.; Soman A.; Arkhipova A. Y.; Li J.; Li H.; Chen Y.; Shi X.; Nordenskiöld L. Chromatin Liquid–Liquid Phase Separation (LLPS) Is Regulated by Ionic Conditions and Fiber Length. Cells 2022, 11, 3145. 10.3390/cells11193145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson B. A.; Blaukopf C.; Lou T.; Chen L.; Doolittle L. K.; Finkelstein I.; Narlikar G. J.; Gerlich D. W.; Rosen M. K. In diverse conditions, intrinsic chromatin condensates have liquid-like material properties. Proc. Natl. Acad. Sci. U. S. A. 2023, 120 (18), e2218085120 10.1073/pnas.2218085120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeshima K.; Tamura S.; Hansen J. C.; Itoh Y. Fluid-like chromatin: Toward understanding the real chromatin organization present in the cell. Curr. Opin. Cell Biol. 2020, 64, 77–89. 10.1016/j.ceb.2020.02.016. [DOI] [PubMed] [Google Scholar]
- Krietenstein N.; Rando O. J. Mesoscale organization of the chromatin fiber. Curr. Opin. Genet. Dev. 2020, 61, 32–36. 10.1016/j.gde.2020.02.022. [DOI] [PubMed] [Google Scholar]
- Kenzaki H.; Takada S. Linker DNA length is a key to tri-nucleosome folding. J. Mol. Biol. 2021, 433, 166792. 10.1016/j.jmb.2020.166792. [DOI] [PubMed] [Google Scholar]
- Ding X.; Lin X.; Zhang B. Stability and folding pathways of tetra-nucleosome from six-dimensional free energy surface. Nat. Commun. 2021, 12 (1), 1091. 10.1038/s41467-021-21377-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu S.; Lin X.; Zhang B. Chromatin fiber breaks into clutches under tension and crowding. Nucleic Acids Res. 2022, 50, 9738–9747. 10.1093/nar/gkac725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Pierro M.; Cheng R. R.; Lieberman Aiden E.; Wolynes P. G.; Onuchic J. N. De novo prediction of human chromosome structures: Epigenetic marking patterns encode genome architecture. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, 12126–12131. 10.1073/pnas.1714980114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacPherson Q.; Beltran B.; Spakowitz A. J. Bottom–up modeling of chromatin segregation due to epigenetic modifications. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, 12739–12744. 10.1073/pnas.1812268115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuebler J.; Fudenberg G.; Imakaev M.; Abdennur N.; Mirny L. Chromatin organization by an interplay of loop extrusion and compartmental segregation. Biophys. J. 2018, 114, 30a. 10.1016/j.bpj.2017.11.211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falk M.; Feodorova Y.; Naumova N.; Imakaev M.; Lajoie B. R.; Leonhardt H.; Joffe B.; Dekker J.; Fudenberg G.; Solovei I. Heterochromatin drives compartmentalization of inverted and conventional nuclei. Nature 2019, 570, 395–399. 10.1038/s41586-019-1275-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi Y.; Zhang B. Predicting three-dimensional genome organization with chromatin states. PLoS Comput. Biol. 2019, 15, e1007024 10.1371/journal.pcbi.1007024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agrawal A.; Ganai N.; Sengupta S.; Menon G. I. Nonequilibrium biophysical processes influence the large-scale architecture of the cell nucleus. Biophys. J. 2020, 118, 2229–2244. 10.1016/j.bpj.2019.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin X.; Qi Y.; Latham A. P.; Zhang B. Multiscale modeling of genome organization with maximum entropy optimization. J. Chem. Phys. 2021, 155 (1), 010901. 10.1063/5.0044150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sommer J.-U.; Merlitz H.; Schiessel H. Polymer-assisted condensation: A mechanism for hetero-chromatin formation and epigenetic memory. Macromolecules 2022, 55, 4841–4851. 10.1021/acs.macromol.2c00244. [DOI] [Google Scholar]
- Wakim J. G.; Spakowitz A. J. Physical modeling of nucleosome clustering in euchromatin resulting from interactions between epigenetic reader proteins. Proc. Natl. Acad. Sci. U. S. A. 2024, 121 (26), e2317911121 10.1073/pnas.2317911121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lequieu J.; Córdoba A.; Moller J.; de Pablo J. J. 1CPN: A coarse-grained multi-scale model of chromatin. J. Chem. Phys. 2019, 150 (21), 215102. 10.1063/1.5092976. [DOI] [PubMed] [Google Scholar]
- Huertas J.; Woods E. J.; Collepardo-Guevara R. Multiscale modelling of chromatin organisation: resolving nucleosomes at near-atomistic resolution inside genes. Curr. Opin. Cell Biol. 2022, 75, 102067. 10.1016/j.ceb.2022.02.001. [DOI] [PubMed] [Google Scholar]
- Farr S. E.; Woods E. J.; Joseph J. A.; Garaizar A.; Collepardo-Guevara R. Nucleosome plasticity is a critical element of chromatin liquid–liquid phase separation and multivalent nucleosome interactions. Nat. Commun. 2021, 12 (1), 2883. 10.1038/s41467-021-23090-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li W.; Wolynes P. G.; Takada S. Frustration, specific sequence dependence, and nonlinearity in large-amplitude fluctuations of allosteric proteins. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 3504–3509. 10.1073/pnas.1018983108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freeman G. S.; Lequieu J. P.; Hinckley D. M.; Whitmer J. K.; De Pablo J. J. DNA shape dominates sequence affinity in nucleosome formation. Phys. Rev. Lett. 2014, 113, 168101. 10.1103/PhysRevLett.113.168101. [DOI] [PubMed] [Google Scholar]
- Freeman G. S.; Hinckley D. M.; Lequieu J. P.; Whitmer J. K.; de Pablo J. J. Coarse-grained modeling of DNA curvature. J. Chem. Phys. 2014, 141 (16), 165103. 10.1063/1.4897649. [DOI] [PubMed] [Google Scholar]
- Lequieu J.; Córdoba A.; Schwartz D. C.; de Pablo J. J. Tension-dependent free energies of nucleosome unwrapping. ACS Cent. Sci. 2016, 2, 660–666. 10.1021/acscentsci.6b00201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lequieu J.; Schwartz D. C.; de Pablo J. J. In silico evidence for sequence-dependent nucleosome sliding. Proc. Natl. Acad. Sci. U. S. A. 2017, 114 (44), E9197–E9205 10.1073/pnas.1705685114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moller J.; Lequieu J.; de Pablo J. J. The free energy landscape of internucleosome interactions and its relation to chromatin fiber structure. ACS Cent. Sci. 2019, 5, 341–348. 10.1021/acscentsci.8b00836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarado W.; Moller J.; Ferguson A. L.; de Pablo J. J. Tetranucleosome interactions drive chromatin folding. ACS Cent. Sci. 2021, 7 (6), 1019–1027. 10.1021/acscentsci.1c00085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarado W.; Agrawal V.; Li W. S.; Dravid V. P.; Backman V.; de Pablo J. J.; Ferguson A. L. Denoising autoencoder trained on simulation-derived structures for noise reduction in chromatin scanning transmission electron microscopy. ACS Cent. Sci. 2023, 9, 1200–1212. 10.1021/acscentsci.3c00178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luque A.; Collepardo-Guevara R.; Grigoryev S.; Schlick T. Dynamic condensation of linker histone C-terminal domain regulates chromatin structure. Nucleic Acids Res. 2014, 42, 7553–7560. 10.1093/nar/gku491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dignon G. L.; Zheng W.; Best R. B.; Kim Y. C.; Mittal J. Relation between single-molecule properties and phase behavior of intrinsically disordered proteins. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, 9929–9934. 10.1073/pnas.1804177115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frenkel D.; Smit B.. Understanding molecular simulation: From algorithms to applications; Elsevier, 2023. [Google Scholar]
- Maeshima K.; Rogge R.; Tamura S.; Joti Y.; Hikima T.; Szerlong H.; Krause C.; Herman J.; Seidel E.; DeLuca J. Nucleosomal arrays self-assemble into supramolecular globular structures lacking 30-nm fibers. EMBO J. 2016, 35, 1115–1132. 10.15252/embj.201592660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arkın H.; Janke W. Gyration tensor based analysis of the shapes of polymer chains in an attractive spherical cage. J. Chem. Phys. 2013, 138 (5), 054904. 10.1063/1.4788616. [DOI] [PubMed] [Google Scholar]
- Allen M. P.; Tildesley D. J.. Computer simulation of liquids; Oxford university press, 2017. [Google Scholar]
- Marshall W.; Straight A.; Marko J.; Swedlow J.; Dernburg A.; Belmont A.; Murray A.; Agard D.; Sedat J. Interphase chromosomes undergo constrained diffusional motion in living cells. Curr. Biol. 1997, 7, 930–939. 10.1016/S0960-9822(06)00412-X. [DOI] [PubMed] [Google Scholar]
- Nozaki T.; Shinkai S.; Ide S.; Higashi K.; Tamura S.; Shimazoe M. A.; Nakagawa M.; Suzuki Y.; Okada Y.; Sasai M. Condensed but liquid-like domain organization of active chromatin regions in living human cells. Sci. Adv. 2023, 9 (14), eadf1488 10.1126/sciadv.adf1488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zidovska A. Chromatin: liquid or solid?. Cell 2020, 183, 1737–1739. 10.1016/j.cell.2020.11.044. [DOI] [PubMed] [Google Scholar]
- Dorigo B.; Schalch T.; Bystricky K.; Richmond T. J. Chromatin fiber folding: requirement for the histone H4 N-terminal tail. J. Mol. Biol. 2003, 327, 85–96. 10.1016/S0022-2836(03)00025-1. [DOI] [PubMed] [Google Scholar]
- Lu X.; Simon M. D.; Chodaparambil J. V.; Hansen J. C.; Shokat K. M.; Luger K. The effect of H3K79 dimethylation and H4K20 trimethylation on nucleosome and chromatin structure. Nat. Struct. Mol. Biol. 2008, 15, 1122–1124. 10.1038/nsmb.1489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pepenella S.; Murphy K. J.; Hayes J. J. Intra-and inter-nucleosome interactions of the core histone tail domains in higher-order chromatin structure. Chromosoma 2014, 123, 3–13. 10.1007/s00412-013-0435-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tolsma T. O.; Hansen J. C. Post-translational modifications and chromatin dynamics. Essays Biochem. 2019, 63, 89–96. 10.1042/EBC20180067. [DOI] [PubMed] [Google Scholar]
- Cui Y.; Bustamante C. Pulling a single chromatin fiber reveals the forces that maintain its higher-order structure. Proc. Natl. Acad. Sci. U. S. A. 2000, 97, 127–132. 10.1073/pnas.97.1.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kruithof M.; Chien F.-T.; Routh A.; Logie C.; Rhodes D.; Van Noort J. Single-molecule force spectroscopy reveals a highly compliant helical folding for the 30-nm chromatin fiber. Nat. Struct. Mol. Biol. 2009, 16, 534–540. 10.1038/nsmb.1590. [DOI] [PubMed] [Google Scholar]
- Chien F.-T.; Van Der Heijden T. Characterization of nucleosome unwrapping within chromatin fibers using magnetic tweezers. Biophys. J. 2014, 107, 373–383. 10.1016/j.bpj.2014.05.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meng H.; Andresen K.; Van Noort J. Quantitative analysis of single-molecule force spectroscopy on folded chromatin fibers. Nucleic Acids Res. 2015, 43, 3578–3590. 10.1093/nar/gkv215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Funke J. J.; Ketterer P.; Lieleg C.; Schunter S.; Korber P.; Dietz H. Uncovering the forces between nucleosomes using DNA origami. Sci. Adv. 2016, 2 (11), e1600974 10.1126/sciadv.1600974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin X.; Zhang B. Explicit ion modeling predicts physicochemical interactions for chromatin organization. eLife 2024, 12, RP90073. 10.7554/eLife.90073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y.-J. C.; Koutelou E.; Dent S. Y. Now open: evolving insights to the roles of lysine acetylation in chromatin organization and function. Mol. Cell 2022, 82, 716–727. 10.1016/j.molcel.2021.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shamir M.; Bar-On Y.; Phillips R.; Milo R. SnapShot: timescales in cell biology. Cell 2016, 164, 1302–1302. 10.1016/j.cell.2016.02.058. [DOI] [PubMed] [Google Scholar]
- Tremethick D. J. Higher-order structures of chromatin: the elusive 30 nm fiber. Cell 2007, 128, 651–654. 10.1016/j.cell.2007.02.008. [DOI] [PubMed] [Google Scholar]
- Ou H. D.; Phan S.; Deerinck T. J.; Thor A.; Ellisman M. H.; O’shea C. C. ChromEMT: Visualizing 3D chromatin structure and compaction in interphase and mitotic cells. Science 2017, 357 (6349), eaag0025 10.1126/science.aag0025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeshima K.; Ide S.; Babokhov M. Dynamic chromatin organization without the 30-nm fiber. Curr. Opin. Cell Biol. 2019, 58, 95–104. 10.1016/j.ceb.2019.02.003. [DOI] [PubMed] [Google Scholar]
- Salacuse J.; Denton A.; Egelstaff P. Finite-size effects in molecular dynamics simulations: Static structure factor and compressibility. I. Theoretical method. Phys. Rev. E 1996, 53, 2382. 10.1103/PhysRevE.53.2382. [DOI] [PubMed] [Google Scholar]
- Sanborn A. L.; Rao S. S. P.; Huang S.-C.; Durand N. C.; Huntley M. H.; Jewett A. I.; Bochkov I. D.; Chinnappan D.; Cutkosky A.; Li J. Chromatin extrusion explains key features of loop and domain formation in wild-type and engineered genomes. Proc. Natl. Acad. Sci. U. S. A. 2015, 112 (47), E6456–E6465 10.1073/pnas.1518552112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheffer M. P.; Eltsov M.; Frangakis A. S. Evidence for short-range helical order in the 30-nm chromatin fibers of erythrocyte nuclei. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 16992–16997. 10.1073/pnas.1108268108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Routh A.; Sandin S.; Rhodes D. Nucleosome repeat length and linker histone stoichiometry determine chromatin fiber structure. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 8872–8877. 10.1073/pnas.0802336105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Widom J. Physicochemical studies of the folding of the 100 Å nucleosome filament into the 300 Å filament: cation dependence. J. Mol. Biol. 1986, 190, 411–424. 10.1016/0022-2836(86)90012-4. [DOI] [PubMed] [Google Scholar]
- Shell M. S.Thermodynamics and statistical mechanics: an integrated approach; Cambridge University Press, 2015. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.