

Abstract
Background/Aim
Alzheimer’s disease (AD) is the most common form of dementia worldwide. Nattokinase is a serine protease extracellularly produced by natto, a fermented product of Bacillus subtilis var. natto. In this study, we investigated the therapeutic effects of nattokinase in a rat model of AD induced by aluminum and D-galactose.
Materials and Methods
Forty Wistar rats were randomly divided into four groups: normal, vehicle, and orally administered nattokinase (NK65 and NK130 groups). Except for the normal group, all groups were treated with AlCl3 and D-galactose for 10 weeks. The NK65 and NK130 groups additionally received 65 mg/kg/day and 130 mg/kg/day nattokinase, respectively. We analyzed β-amyloid levels in the cerebrospinal fluid (CSF), and the spatial reference test was evaluated using the Morris water maze test. After the Morris water maze test, rats of all groups were subjected to micro-computed tomography (μCT) to assess constructional changes in the brain. Aluminum concentration and β-amyloid levels were analyzed by histochemical staining in all brain tissues.
Results
Oral administration of nattokinase in the AD rat model increased free-form β-amyloid levels in the CSF and improved aluminum and amyloid plaque accumulation in the brain. Brain μCT images showed enhanced brain volume with fewer constructional changes after treatment with nattokinase. In the behavioral tests, both the escape latency time in the spatial reference test and the time taken to cross the platform area in the spatial probe test improved partially.
Conclusion
The results suggest that nattokinase has potential therapeutic applications in the treatment of AD.
Keywords: Alzheimer’s disease, nattokinase, natto, rat model
Alzheimer’s disease (AD) is the main cause of dementia and one of the greatest healthcare challenges worldwide in the 21st century. AD continues to be an issue of concern owing to the aging population and elevated incidence in many countries (1). AD is characterized by impaired learning, cognitive deficits, behavioral disturbances, progressive neuronal loss, and cerebral atrophy (2). The main pathogenesis of AD is the accumulation of extracellular amyloid β (Aβ) plaques (3). Aβ production initiates a cascade of pathological changes in the brain including inflammation, oxidative damage, and excitotoxicity, whereas tau protein hyperphosphorylation results in microtubule disassembly. Both these changes can lead to neuronal destruction and cognitive deficiencies (4). Until recently, only two classes of drugs were approved to treat AD: inhibitors of cholinesterase enzymes and antagonists of N-methyl d-aspartate (NMDA) (5). In January 2023, the FDA granted anti-Aβ protofibril mAb to reduce the progression of memory loss (6). Thus, the development of drugs or agents that potentially reduce plasma Aβ aggregates such as anti-oligomeric Aβ mAb or proteolytic enzymes is currently a priority.
Continuous exposure to heavy metals has several negative effects on human health. Aluminum, a widely used heavy metal, selectively accumulates in the brain leading to AD via different mechanisms, including oxidative and endoplasmic reticulum stress, mitochondrial dysfunction, neuroinflammation, cell death, interaction Aβ and α-synuclein, cytoskeletal abnormalities, as well as alteration of synaptic plasticity and signal transduction through interference with neurotransmitter system (7).
Nattokinase (NK) is an extracellular serine protease produced by B. subtilis var. natto found in natto (8). NK has potent fibrinolytic and antithrombotic activities. Additionally, NK has anticoagulant/anti-aggregant, anti-atherosclerotic, lipid-lowering, antihypertensive, and neuroprotective effects (9). NK intake leads to positive outcomes in the pathophysiology of AD in rat models (10). NK is a natural product with proven safety for human consumption following oral administration. In this study, we aimed to investigate the protective effects of NK in a rat model of AD induced by exposure to aluminum chloride and D-galactose. We evaluated the effects of NK on learning and memory. We also analyzed the levels of Aβ in cerebrospinal fluid, aluminum concentration, and β-amyloid levels by histochemical staining. Brain micro-computed tomography (CT) was used to assess constructional changes in the brain (Figure 1).
Figure 1.
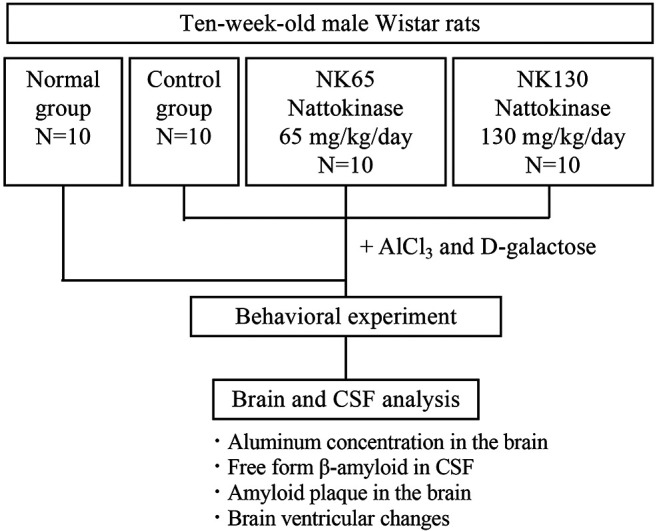
Flow chart of the study design.
Materials and Methods
Animals. Ten-week-old male Wistar rats weighing 410-575 g were used in this study. The animals were purchased from BioLASCO Taiwan Co., LTD and kept under standard housing conditions, which included a 12-hour light and dark cycle, room temperature, and relative humidity (50-70%). They were allowed water and food ad libitum throughout the experimental period.
Ethics approval. All experimental procedures were performed by Trineo Biotechnology Co. Ltd. The research facility was accredited by the Taiwan Accreditation Foundation and certified as a compliance-registered GLP facility for preclinical testing laboratories (Certificate No.: GLP-0031). All procedures were conducted in compliance with the Animal Experiment Regulations of Trineo Biotechnology Co., Ltd., Animal Experiment Committee Regulations, and Animal Experiment Approval Provisions (approval no. IACUC-2017-SH-007).
Experimental design. Forty male Wister rats were randomly allocated into four groups, each consisting of 10 rats: the normal group, the control group, and two dosing groups. Except for the normal group, all groups were treated with aluminum chloride (AlCl3; 200 mg/kg/day, intragastrically) and D-galactose (300 mg/kg/day, subcutaneously) for 10 weeks. In the NK-treated group, rats were orally administered deionized water (control) or two different doses of NK (65 or 130 mg/kg body weight) once per day. The fibrin decomposition units (FU) of 65 mg and 130 mg NK (Contek, Co., Ltd.) were 2,600 FU and 5,200 FU, respectively. All solutions were administered by oral gavage. The dose was 10 ml/kg. The administered volume was calculated based on the body weight of the rats recorded before the first dose.
Analysis of free-form β-amyloid in CSF. The CSF samples were diluted with ddH2O and mixed with 0.8 M urea. The supernatant was then directly subjected to LC-MS/MS analysis. LC analysis was performed using an Eksigent Ekspert ultraLC 110 UHPLC system (AB Sciex LLC, Framingham, MA, USA). The chromatographic separation was carried out on a C18 column (2.1×100 mm, 2.5 μm) with a 5.5 min linear gradient of mobile phase A, comprising 0.3% NH4OH in water, and mobile phase B, consisting of a mixture of 0.3% NH4OH in water/ACN at a ratio of 10/90. The gradient ranged from 10% to 45% B, at a flow rate of 300 μl/min. The injection volume was set to 10 μl.
MS/MS analysis was conducted using an AB Sciex QTRAP® 5500 LC/MS/MS system with IonDrive™ Turbo V ion source and electrospray ionization (ESI) probe. All compounds and source/gas parameters were optimized based on direct injection or split T injection of peptide standards. An MRM transition of 847.7→824 provided the best S/N ratio for the analysis of for 1-40 and was selected for quantitation. The data were processed using MultiQuant™ software.
Micro-CT scanning. CT images were acquired using Quantum FX micro-CT scanner (Perkin Elmer Inc., Waltham, MA, USA) with an FOV of 40, tube potential peaks of 90 kVp, tube current of 200 μA, and 4.5 min exposure time. Tomographic images were obtained using the Quantum FX micro-CT viewer. 3D reconstruction was performed using 512 images processed by the volume rendering method, and brain volume was measured using Analyze 11.0 (AnalyzeDirect, Inc., Overland Park, KS, USA). Ventricular dilatation, cerebral cortex atrophy, and hippocampal atrophy in the hippocampal images were monitored to assess changes in the brain ventricles. The changes were defined as not obvious (–), partial dilatation or atrophy (+/–), and obvious (+).
Determination of aluminum concentration and histological investigation. Tissues were digested in a microwave (MARS Xpress, CEM Corp., Matthews, NC, USA). The total aluminum content in each sample was measured using graphite furnace atomic absorption spectrophotometry with matrix-matched standards and an established analytical program. Brain tissues were fixed in 10% formalin buffer for 24 h and dehydrated. Specimens were cleared in xylene and embedded in paraffin for sectioning. The tissue sections obtained were deparaffinized and stained with hematoxylin and Congo red for histological examination under a light microscope (11). Three images were obtained from each rat for the amyloid plaque count.
Behavioral experiment. The Morris water maze test was used to assess neurocognitive function at the end of 9 weeks (12). A circular pool (210 cm in diameter and 50 cm in height) was filled with water, and the platform was submerged. Each rat was placed in water to search for the platform in four trials per day for three consecutive days. The probe test was performed on the third day after the four trials. In the probe test, the platform was removed, and each rat was forced to search for the platform for 90 s. The track was recorded, and the visit, path, and time of the platform quadrant were analyzed using a video tracking software.
Statistical analysis. Data are presented as mean and standard deviation (mean±S.D.). Data were compared between groups using two-tailed t-tests. A value of p<0.05 was considered statistically significant.
Results
Aluminum concentration in the brain. The aluminum concentration in the brain was determined in the 10th week after continuous treatment with AlCl3 (200 mg/kg/day, intragastrically) and D-galactose (300 mg/kg/day, subcutaneously). The aluminum content in the vehicle group treated with AlCl3 was higher than that in the normal group (Table I). In the NK65 and NK130 groups, the rats were orally administered two different doses of NK (65 and 130 mg/kg body weight, respectively) once daily simultaneous to continuous treatment with AlCl3. The aluminum content in the NK130 group (1.75±0.85 μg/g) was significantly reduced compared to that in the vehicle group (2.55±0.85 μg/g) and nearly similar level to that in the normal group (1.74±0.85 μg/g), whereas that in the NK65 group (2.63±1.02 μg/g) was not different from that in vehicle group. These results imply that NK reduces the accumulation or metabolism of aluminum in the brain.
Table I. Aluminum concentration in the brain.

*p<0.05, compared to the normal group. #p<0.05, compared to the vehicle group. S.D: Standard deviation.
Histological staining of free-form β-amyloid in CSF and amyloid plaque. The presence of free-form β-amyloid and amyloid plaques has been associated with AD. We first measured the levels of free-form β-amyloid in the CSF. Because low level of free-form β-amyloid in the CSF could be an indicator of AD with a high diagnostic sensitivity and specificity, free-form β-amyloid in CSF could serve as a crucial biomarker for early disease detection (13). As shown in Table II, the level of free-form β-amyloid in the vehicle group (11.20±10.97 ng/ml) was lower than that in the normal group (18.20±10.97 ng/ml). NK65 and NK130 treatment groups showed higher free-form β-amyloid levels (NK65; 17.50±13.79 ng/ml and NK130; 16.30±12.52 ng/ml) than that in vehicle group and an equivalent level to that in the normal group (Table II).
Table II. Free-form β-amyloid in CSF at 10 weeks.

0: Data below the limit of detection; the minimum detectable value of the assay is typically less than 0.01 ng/ml. CSF: Cerebrospinal fluid.
Next, we measured amyloid plaques using histological staining. The accumulation of β-amyloid plaques accompanies altered glucose metabolism and synaptic loss, which contribute to a gradual decline in cognitive function. Therefore, the hippocampus and cortex were stained with Congo red dye to detect the presence of these plaques. The number of amyloid plaques was significantly higher in the vehicle group than that in the normal group (Table III). Compared with that in the vehicle group, no significant reduction in amyloid plaques was observed in the NK (65 mg/kg) treatment group; however, the number of plaques in the NK (130 mg/kg) treatment group was significantly reduced from 18.5 to 11.6 (Table III). These results indicated that NK partially mitigates β-amyloid plaque formation.
Table III. Number of amyloid plaques in the brain.

*p<0.05, compared with the normal group. #p<0.05, compared with the vehicle group.
Brain ventricular changes. The changes in ventricular dilatation, cerebral cortex atrophy, and hippocampus atrophy were assessed using micro-CT scanning. Image data were extracted and quantified as brain volumes (mm3) (Table IV). The average brain volume of the vehicle group was drastically reduced because of the more intense disorder of ventricular dilatation, cerebral cortex atrophy, and hippocampal atrophy than those in the normal group. The brain volume in the NK65 and NK130 groups treated with NK restored and was significantly higher than that in the vehicle group, almost reaching a level similar to that in the normal group.
Table IV. Definition of ventricular dilatation, cerebral cortex atrophy, and hippocampus atrophy.

*p<0.05, compared with the normal group. #p<0.05, compared with the vehicle group.
Behavioral experiments (Morris water maze test and probe test). Morris water maze and probe tests were conducted to investigate spatial learning and memory. Animals were given cues and trained for three days. In the training phase shown in Table V, the vehicle group took longer to locate the platform than that taken by the normal group. In the groups treated with 65 and 130 mg/kg NK, performance improved (Table V). However, no statistically significant differences were observed between the groups. During the probe test, the vehicle group made fewer visits to the platform than did the normal group (Table VI). Furthermore, the number of visits in the NK treatment groups showed an increasing trend compared to that in the vehicle group, whereas the path length decreased in the NK treatment groups. Specifically, the NK65 group showed a statistically significant decrease compared with that in the vehicle group, suggesting that NK intake may contribute to memory improvement.
Table V. Morris water maze test results (s).

Table VI. Probe test results.

*p<0.05, significant difference.
Discussion
In this study, we evaluated the effects of NK in the AD rat model. Oral administration of NK increased free-form β-amyloid levels in CSF and improved aluminum and amyloid plaque accumulation in the brain at the 10th week. Brain micro-CT images showed enhanced brain volume with fewer constructional changes after treatment with NK. These results indicate that NK has therapeutic applications in the treatment of AD. Further studies are required to elucidate the underlying mechanisms. We also analyzed gene expression related to antioxidant enzymes including HO-01, NQO-1, GSTP-1, and SOD-3, and β-amyloid metabolism including ADAM9 and ADAM10 (Data not shown). However, we could not identify any obvious molecular factors underlying the improvement of the AD rat model phenotype after the oral administration of NK. Previous study indicated that NK had AchE inhibitory effects in the brain and decreases ADAM9 and ADAM10 expression in AD rat models (10). In our study, although we could not obtain the same results for gene expression, we evaluated the quantitative results of free-form β-amyloid in the CSF and amyloid plaque in the brain in addition to behavioral experiment tests. These findings indicate that NK has the potential to delay the onset or slow the progression of AD.
Conclusion
We evaluated the effects of the oral administration of NK in a rat model of AD induced by AlCl3 and D-galactose treatment. In the AD rat model, administering NK orally led to an increase in the level of free-form β-amyloid in the CSF and improved the accumulation of aluminum and amyloid plaques in the brain. Brain μCT images revealed enhanced brain volume following NK treatment. Moreover, in the behavioral tests, partial improvement was observed in both escape latency time during the spatial reference test and the frequency of crossing the platform area in the spatial probe test. Further investigation is required to understand how NK intake affects AD progression.
Conflicts of Interest
The Authors declare no conflicts of interest.
Authors’ Contributions
Conceptualization, T.T., J.Y. and M.K.; Methodology, J.Y., K.H. and S.C.; Formal analysis, T.T. and M.K.; Supervision, T.T. and M.K.; Writing–original draft preparation, T.T. and M.K. Writing–review and editing, T.T., J.Y., K.H., S.C., A.I. and M.K. All Authors have read and agreed to the published version of the article.
References
- 1.Reitz C, Mayeux R. Alzheimer disease: epidemiology, diagnostic criteria, risk factors and biomarkers. Biochem Pharmacol. 2014;88(4):640–651. doi: 10.1016/j.bcp.2013.12.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Scheltens P, De Strooper B, Kivipelto M, Holstege H, Chételat G, Teunissen CE, Cummings J, van der Flier WM. Alzheimer’s disease. Lancet. 2021;397(10284):1577–1590. doi: 10.1016/S0140-6736(20)32205-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.d’Errico P, Meyer-Luehmann M. Mechanisms of pathogenic Tau and Aβ protein spreading in Alzheimer’s disease. Front Aging Neurosci. 2020;12:265. doi: 10.3389/fnagi.2020.00265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Yiannopoulou KG, Papageorgiou SG. Current and future treatments in Alzheimer disease: an update. J Cent Nerv Syst Dis. 2020;12:1179573520907397. doi: 10.1177/1179573520907397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Breijyeh Z, Karaman R. Comprehensive review on Alzheimer’s disease: causes and treatment. Molecules. 2020;25(24):5789. doi: 10.3390/molecules25245789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hong M, Bitan G. Recent advances and future therapy development for Alzheimer’s disease and related disorders. Neural Regen Res. 2024;19(9):1877–1878. doi: 10.4103/1673-5374.391182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Skalny AV, Aschner M, Jiang Y, Gluhcheva YG, Tizabi Y, Lobinski R, Tinkov AA. Molecular mechanisms of aluminum neurotoxicity: Update on adverse effects and therapeutic strategies. Adv Neurotoxicol. 2021;5:1–34. doi: 10.1016/bs.ant.2020.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sumi H, Hamada H, Tsushima H, Mihara H, Muraki H. A novel fibrinolytic enzyme (nattokinase) in the vegetable cheese Natto; a typical and popular soybean food in the Japanese diet. Experientia. 1987;43(10):1110–1111. doi: 10.1007/BF01956052. [DOI] [PubMed] [Google Scholar]
- 9.Hristova M. Nattokinase – Uses and benefits. TJS. 2023;21(4):343–350. doi: 10.15547/tjs.2023.04.007. [DOI] [Google Scholar]
- 10.Fadl N, Ahmed H, Booles H, Sayed A. Serrapeptase and nattokinase intervention for relieving Alzheimer’s disease pathophysiology in rat model. Hum Exp Toxicol. 2013;32(7):721–735. doi: 10.1177/0960327112467040. [DOI] [PubMed] [Google Scholar]
- 11.Morikawa T, Uehara O, Paudel D, Yoshida K, Harada F, Hiraki D, Sato J, Matsuoka H, Kuramitsu Y, Michikawa M, Abiko Y. Systemic administration of lipopolysaccharide from Porphyromonas gingivalis decreases neprilysin expression in the mouse hippocampus. In Vivo. 2023;37(1):163–172. doi: 10.21873/invivo.13065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ren H, Zhu M, Yu H, Weng Y, Yu W. The effect of propofol on the hippocampus in chronic cerebral hypoxia in a rat model through Klotho regulation. In Vivo. 2024;38(3):1162–1169. doi: 10.21873/invivo.13551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Anoop A, Singh PK, Jacob RS, Maji SK. CSF Biomarkers for Alzheimer’s disease diagnosis. Int J Alzheimers Dis. 2010;2010:606802. doi: 10.4061/2010/606802. [DOI] [PMC free article] [PubMed] [Google Scholar]