

Abstract
Background
Educational attainment (EduA) is correlated with life outcomes, and EduA itself is influenced by both cognitive and non-cognitive factors. A recent study performed a ‘genome-wide association study (GWAS) by subtraction,’ subtracting genetic effects for cognitive performance from an educational attainment GWAS to create orthogonal ‘cognitive’ and ‘non-cognitive’ factors. These cognitive and non-cognitive factors showed associations with behavioral health outcomes in adults; however, whether these correlations are present during childhood is unclear.
Methods
Using data from up to 5517 youth (ages 9–11) of European ancestry from the ongoing Adolescent Brain Cognitive DevelopmentSM Study, we examined associations between polygenic scores (PGS) for cognitive and non-cognitive factors and cognition, risk tolerance, decision-making & personality, substance initiation, psychopathology, and brain structure (e.g. volume, fractional anisotropy [FA]). Within-sibling analyses estimated whether observed genetic associations may be consistent with direct genetic effects.
Results
Both PGSs were associated with greater cognition and lower impulsivity, drive, and severity of psychotic-like experiences. The cognitive PGS was also associated with greater risk tolerance, increased odds of choosing delayed reward, and decreased likelihood of ADHD and bipolar disorder; the non-cognitive PGS was associated with lack of perseverance and reward responsiveness. Cognitive PGS were more strongly associated with larger regional cortical volumes; non-cognitive PGS were more strongly associated with higher FA. All associations were characterized by small effects.
Conclusions
While the small sizes of these associations suggest that they are not effective for prediction within individuals, cognitive and non-cognitive PGS show unique associations with phenotypes in childhood at the population level.
Keywords: academic achievement, cognitive performance, educational attainment, genetics, late childhood, middle childhood, neuroimaging, polygenic scores
Introduction
As educational attainment (EduA) is among the strongest predictors of positive outcomes across the lifespan (e.g. income, health, well-being; Gutacker, Kinge, & Olsen, 2023; Raghupathi & Raghupathi, 2020; Zajacova & Lawrence, 2018), understanding its contributing factors is important. Despite often being thought of as primarily ‘environmental,’ EduA itself is moderately heritable (h2 = 0.41–0.47; Heath et al., 1985; Silventoinen et al., 2020). Evidence that EduA is strongly impacted by cognitive ability and a set of broadly defined ‘non-cognitive skills’ (e.g. emotion regulation and personality traits such as grit and curiosity; Chamorro-Premuzic & Furnham, 2003; Duckworth, Peterson, Matthews, & Kelly, 2007, 2019; Kovas et al., 2015; Malanchini, Engelhardt, Grotzinger, Harden, & Tucker-Drob, 2019; Noftle & Robins, 2007) has inspired recent approaches that have deconstructed the genetic architecture of EduA into cognitive and non-cognitive components that have shared and unique associations with complex EduA-related phenotypes (e.g. risk tolerance and psychopathology; Demange et al., 2021; Malanchini et al., 2024; Tucker-Drob & Harden, 2012; Tucker-Drob, Briley, Engelhardt, Mann, & Harden, 2016). However, the extent to which these differential associations are present in childhood, before education has been completed, remains poorly understood (Malanchini et al., 2024). Moreover, prior research in adolescents underscores the critical role of development in understanding factors such as substance use and sleep disturbances that can significantly impact educational outcomes and often are exacerbated during adolescence (de Zeeuw et al., 2014; Donnellan, Martin, & Senia, 2021; Hicks et al., 2021). By studying PGS of cognitive and non-cognitive components of EduA in a developmental sample, we can study the extent to which genetic liability for these cognitive and non-cognitive factors are associated with outcomes such as substance use and personality prior to the completion of education.
This study aims to characterize the associations between cognitive and non-cognitive PGS and psychosocial and neuroimaging phenotypes in a cohort of individuals aged 9–11 who have not yet been as impacted by socioenvironmental factors such as amount of schooling. We also controlled for potential confounders such as population stratification, assortative mating, and passive gene-environment correlations through within-sibling analyses.
Deconstructing the genetic architecture of educational attainment into cognitive and non-cognitive components
Demange et al. (2021) demonstrated that genetic effects on EduA can be parsed into cognitive and non-cognitive factors. To study genetic influences on ‘non-cognitive skills,’ Demange et al. (2021) performed a novel ‘genome-wide association study (GWAS)-by-subtraction’ by residualizing the genetic effects of EduA (N = 1 131 881; Lee et al., 2018) on the genetic effects of cognitive performance (Lee et al., 2018) in a Cholesky decomposition using genomic structural equation modeling (Grotzinger et al., 2019), leaving a residual ‘non-cognitive’ genetic factor (N = 510 715) and ‘cognitive’ genetic factor (N = 257 841; Lee et al., 2018). In this way, the authors generated two new GWASs, one of ‘non-cognitive skills’ that represents genetic influences on EduA that are not shared with cognitive performance, and an orthogonal ‘cognitive skills’ GWAS. Their study identified 157 independent loci associated with the non-cognitive factor and observed that the cognitive and non-cognitive factors showed distinct associations with other relevant phenotypes. For example, the non-cognitive factor showed positive genetic correlations with risk tolerance and some psychiatric disorders (e.g. bipolar disorder and schizophrenia), and was positively genetically correlated with personality traits including conscientiousness, extraversion, and agreeableness, while the cognitive factor showed negative or null correlations with these same phenotypes (Demange et al., 2021).
While the GWASs mentioned above consisted largely of adult participants, adolescence is a critical stage of both cognitive and non-cognitive development. Prior studies have shown that early individual differences in cognitive (e.g. cognitive performance; Lövdén, Fratiglioni, Glymour, Lindenberger, & Tucker-Drob, 2020) and non-cognitive factors (e.g. personality; Chamorro-Premuzic & Furnham, 2003; Noftle & Robins, 2007) may contribute to EduA, employment outcomes, and overall success later in life (Duckworth et al., 2019; Moffitt et al., 2011). A recent preprint reported that the association between a non-cognitive PGS and academic achievement nearly doubled between the ages of 7 and 16 (Malanchini et al., 2024), suggesting that genetic factors related to non-cognitive facets of EduA may become particularly influential during adolescence, a critical developmental period for factors related to EduA. To date, most examinations of associations between genetic propensities for cognitive and non-cognitive factors and brain imaging have been conducted in adult samples (Demange et al., 2021). Little is known of the brain mechanisms related to either cognitive or non-cognitive factors during middle/late childhood, a critical period for neural plasticity. In sum, despite adolescence being an important time for future academic achievement, little is known about the cognitive and non-cognitive influences on EduA and their correlations with relevant traits, behaviors, and brain structure during critical developmental periods such as middle/late childhood.
The current study
In the current study, we estimated the associations between PGS for cognitive and non-cognitive factors of EduA and outcome measures among children enrolled in the ongoing Adolescent Brain Cognitive Development (ABCD) Study (Volkow et al., 2018; see Fig. 1 for overview). We focused on behavioral phenotypes implicated in studies on adults (i.e. cognition, risk tolerance, personality, & decision-making, substance initiation, psychopathology; Demange et al., 2021) and examined associations with brain structure. Finally, as prior research has shown that confounds such as assortative mating (Horwitz, Balbona, Paulich, & Keller, 2023) can inflate GWAS test statistics (Yengo et al., 2018) and polygenic score associations (Okbay et al., 2022), we performed post hoc within-sibling analyses to assess whether any significant associations may be independent of assortative mating, passive gene-environment correlation, or other sociodemographic confounders.
Figure 1.

Overview of study design. Demange et al. (2021) performed a ‘genome-wide association study (GWAS) by subtraction,’ subtracting genetic effects for a cognitive performance GWAS from an educational attainment (EduA) GWAS to create orthogonal ‘cognitive’ and ‘non-cognitive’ factors. Here, we created cognitive and non-cognitive PGSs using summary statistics from this GWAS-by-subtraction and evaluated whether EduA cognitive and non-cognitive PGSs are associated with cognition, risk tolerance, personality, & decision-making, substance initiation, psychopathology, and neuroimaging phenotypes in the Adolescent Brain Cognitive Development (ABCD) Study.
Methods
Participants
The ongoing Adolescent Brain and Cognitive Development SM (ABCD) Study® is a longitudinal study following 11 879 children (ages 8.9–11 at baseline; born between 2005 and 2009) recruited from 21 research sites across the United States to study the development of complex behavior and biology from late childhood to late adolescence/young adulthood in the context of experience and genetic background (Volkow et al., 2018). It includes a family-based component in which twin (n = 2108), triplet (n = 30), non-twin siblings (n = 1589), and singletons (n = 8148) were recruited. Caregivers provided written informed consent and their children provided verbal assent. For the present study, we used data from the baseline visit (2016–2018; ages: 9–11). Analyses were only conducted in individuals with genetic ancestry most similar to those of European ancestry reference populations (see Polygenic Scores section below), due to the lack of relevant well-powered discovery GWAS in other ancestries and the low predictive utility of PGS when applied across ancestries (Martin et al., 2017, 2019). After excluding individuals with missing outcome or covariate data, described below, analytic Ns ranged from 4688 to 5517 (see online Supplemental Fig. S1 for analytic sample size flow chart).
Measures
Cognition, risk tolerance, personality & decision-making, substance initiation, psychopathology, and neuroimaging data were drawn from the baseline assessments from the National Institute of Mental Health Data Archive (NDA; https://nda.nih.gov/); data release 4.0 and 5.1, see online Supplemental Table S1 for details). Socioeconomic status and genomic data (release 3.0) were also derived from the baseline session.
Cognition
Crystallized and fluid intelligence as well as their total composite were estimated from the NIH Toolbox assessment (Luciana et al., 2018).
Risk tolerance, personality, & decision-making
Risk tolerance and impulsivity were measured at baseline using the 20-item UPPS-P Impulsive Behavior scale short form (Cyders, Littlefield, Coffey, & Karyadi, 2014; Watts, Smith, Barch, & Sher, 2020) which captures five dimensions related to impulsivity: (1) Negative Urgency (i.e. tendency to act rashly when in a heightened negative mood state), (2) Positive Urgency (i.e. tendency to act rashly when in a heightened positive mood state), (3) Lack of Perseverance (i.e. inability to keep focused or motivated to finish a task), (4) Lack of Premeditation (i.e. tendency to not plan ahead or to act without forethought of behavioral consequences), and (5) Sensation Seeking (i.e. tendency to pursue novel or exciting activities and experiences); see online Supplemental Table S1 for details. A modified version of the BIS/BAS scales (Carver & White, 1994; Pagliaccio et al., 2016) was used to broadly assess two motivational systems: the behavioral inhibition system (BIS) and the behavioral activation system (BAS). Measures included the BIS subscale, sensitivity to punishment resulting in avoidance and negative emotionality, and two BAS subscales: Drive, positive motivation toward goals, and Reward Responsiveness, sensitivity to reward resulting in approach behaviors and positive emotionality (see online Supplemental Table S1 for details). The BAS Fun-Seeking subscale was excluded from analyses as previous research has indicated it had poor loading in age ranges that match the ABCD sample at baseline (Pagliaccio et al., 2016). A single item from the Sensation Seeking scale ‘I enjoy taking risks’ was also included as a stand-alone measure of general risk tolerance. Delayed discounting, as an index of decision-making, was measured at baseline using the single-item cash choice task where youth decide whether they would ‘rather have $75 in three days or $115 in 3 months’ (Luciana et al., 2018; Wulfert, Block, Santa Ana, Rodriguez, & Colsman, 2002). This single-item measure was used, as opposed to behavioral data acquired (Kohler, Lichenstein, & Yip, 2022), due to quality control procedures of these data resulting in the exclusion of large amounts of baseline data.
Substance use initiation
Given the low endorsement for all other substances at the baseline assessment, only lifetime alcohol initiation, defined as ‘sips’ or ‘full drinks,’ from the baseline substance use interview (Volkow et al., 2018) was analyzed in the current study. Individuals endorsing alcohol use only in the context of religious ceremonies were excluded. Participants who had endorsed alcohol use were compared to substance naive participants who endorsed no substance initiation (e, g., individuals who endorsed alcohol use were compared to individuals who did not initiate alcohol use nor use of cannabis, tobacco, or any other substances; Miller et al., 2024; see online Supplemental Tables S1, S2 for additional information).
Psychopathology
We assessed six mental health-related measures. Given that the distribution of the scores of severity of youth psychotic-like experiences (PLEs) (Karcher & Barch, 2021) was skewed, we first winsorized to three standard deviations. However, as the variable still had a skew of ⩾ |1.96|, we then performed a rank-based inverse normal transformation of the variable (See online Supplemental Table S1 for details). We also included a caregiver-reported diagnosis screener for autism (Barch et al., 2018). Baseline data from the 5.1 release was used to generate KSADS-5 diagnoses (Kaufman et al., 1997). KSADS-5 diagnoses were created for: obsessive compulsive disorder (OCD), bipolar disorder, major depressive disorder (MDD), and attention deficit hyperactivity disorder (ADHD). The analytical N for these measures ranged from 5493 to 5506 and all definitions and items used to create the lifetime mental health diagnoses measures can be found in online Supplemental Table S1.
Neuroimaging
Indices of gray matter structure (i.e. cortical volumes) and white matter tracts (i.e. fractional anisotropy [FA], mean diffusivity [MD]) were derived using the Desikan-Killiany atlas (Desikan et al., 2006) and Atlas Tract (Basser, Mattiello, & Lebihan, 1994), respectively. No task-related functional magnetic resonance imaging (fMRI) data were examined due to test–retest reliability concerns (Elliott et al., 2020). Acquisition and preprocessing methodology (Casey et al., 2018; Hagler et al., 2019) as well as additional information can be found in both the online Supplemental Table 3 and Supplemental Methods.
Polygenic scores
Quality control was performed on the genomic data (ABCD data release 3.0) using the Rapid Imputation and COmputational PIpeLIne for Genome-Wide Association Studies (Lam et al., 2020). Briefly, after performing QC and using a combination of parent-reported demographic information and principal components analysis to identify a subset of n = 5556 individuals of genetic ancestry most similar to those of European ancestry reference populations, the genetic data were imputed to the TOPMed imputation reference panel (Taliun et al., 2021; online Supplemental Methods). Only individuals of European ancestry were analyzed, as the Demange et al., discovery GWAS only included individuals of European ancestry, and there is poor predictive utility across ancestries which may lead to erroneous conclusions (Martin et al., 2017, 2019). While there are GWAS of educational attainment and cognitive performance in other ancestries (such as those available for download from the Pan-UK Biobank), these GWAS are very small (N < 10 000) and would result in extremely under-powered PGS. Thus, we chose not to extend our analyses to other ancestries, given the low statistical power of the existing GWAS and the potentially sensitive nature of phenotypes like cognition and educational attainment. Single nucleotide polymorphisms (SNPs) with imputation R2 > 0.8 and minor allele frequency>0.01 were retained for PGS analyses.
We used PRS-CS (Ge, Chen, Ni, Feng, & Smoller, 2019) to calculate polygenic scores (PGS) in the European ancestry subset of the ABCD Study sample, using effect sizes from the Demange et al., GWAS of ‘cognitive skills’ (GWAS catalog accession GCST90011875; effective n = 257 700; Demange et al., 2021) and ‘non-cognitive skills’ (GWAS catalog accession GCST90011874; effective n = 510 795; Demange et al., 2021), derived by using ‘GWAS-by-subtraction’ to residualize the genetic effects of EduA on the genetic effects of cognitive performance. We used the 1000 Genomes Phase 3 European ancestry sample as a reference panel (1000 Genomes Project Consortium et al., 2015). We used the ‘auto’ function of PRS-CS, allowing the software to learn the global shrinkage parameter from the data (see online Supplemental Methods for details).
Statistical analyses
Analyses were preregistered on the Open Science Network (https://doi.org/10.17605/OSF.IO/EV4BH) and conducted using mixed effects models implemented using lmer (for continuous outcomes) and glmer (for dichotomous outcomes) from the lme4 package (Austin, 2010; Bates, Maechler, Bolker, & Walker, 2015) in R (v4.3; R Core Team). Both the cognitive and non-cognitive PGS were included as predictors in the same model (i.e. they were analyzed simultaneously). Age, sex, and the first 10 genetic principal components were included as fixed effect covariates, with family ID and recruitment site as random effects to account for data dependence. For imaging models, recruitment site was replaced by MRI serial number. Imaging models also included MRI manufacturer, global brain metrics representing the mean for each modality, and mean motion for DTI as fixed effects. We used false discovery rate (FDR; Benjamini and Hochberg, 1995) to account for multiple testing (pfdr < 0.05); FDR was applied separately to the non-imaging phenotypes and each respective imaging modality. R2 for the continuous outcomes was calculated using the MuMIn package (Bartóń, 2024). For the categorical outcomes, we used the same package to calculate Nagelkerke pseudo R2 (Nagelkerke, 1991).
Three additional non-pre-registered post-hoc tests were included: First, we tested whether the regression coefficients for the cognitive and non-cognitive PGS significantly differed from each other in each model (pdiff; online Supplemental Methods). Second, to assess whether associations with cognitive and non-cognitive PGS were influenced by socioeconomic status (SES), we performed supplementary analyses that covaried for two SES related variables at baseline: (1) caregiver-reported combined past-12-month family income and (2) neighborhood deprivation index, a composite of neighborhood socioeconomics (Fan et al., 2021). Third, we conducted post hoc within-sibling analyses to assess whether any significant associations arising from the primary analyses may plausibly represent direct genetic effects. Polygenic score associations can be confounded by gene-environment correlations (rGE), where an individual's genetic predisposition may be correlated with their environment. This phenomenon may occur via passive rGE, whereby parents shape a child's environment based on their own genetic predispositions (which are also passed down to their children), or evocative or active rGE, whereby children evoke responses or actively seek environments that correlate with their own genetic disposition. Within-sibling PGS analyses are one way to test whether PGS associations are inconsistent with passive rGE. Because alleles are randomly assigned at birth, any genetic differences between siblings are assumed to be free from environmental influences that are shared, such as passive rGE, and also free from other between-family confounding factors, such as assortative mating or population stratification. Thus, significant within-sibling effects indicate that these associations are unconfounded by population stratification, assortative mating, passive gene-environment correlations (passive rGE), and other potential population-level confounds, though it should be noted that active and evocative rGE will still influence within-sibling variation in PGS effects (Brumpton et al., 2020; Howe et al., 2022; Young et al., 2018). For these analyses, we included both the family mean PGS and each sibling's deviation from their family mean PGS as predictors in a mixed-effect model, as has been done previously (Selzam et al., 2019):
where Yijk represents the outcome for sibling i in family j at site k, PGSij represents the polygenic score for sibling i in family j, PGSj represents the mean PGS for family j, γj represents a random intercept for family j, γk represents a random intercept for site k, and 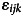 represents the independent random error for each individual i in family j at site k. We also included the same fixed-effect covariates (age, sex, and genetic principal components) as in our primary models described above (not shown in equation). In this way, we were able to partial the variance in the outcome explained by the PGS into between-family (βB) and within-family (βW) effects. We also included the same fixed-effect covariates (age, sex, and genetic principal components) as in our primary models described above (not shown in equation). Because the sample of siblings (N = 1702) was considerably smaller than the full analytic sample (N = 5517), we derived the population-level estimates (the ‘Total’ effects) between PGS and outcomes in a sample of equivalent size (and statistical power). See online Supplemental Methods for additional details.
represents the independent random error for each individual i in family j at site k. We also included the same fixed-effect covariates (age, sex, and genetic principal components) as in our primary models described above (not shown in equation). In this way, we were able to partial the variance in the outcome explained by the PGS into between-family (βB) and within-family (βW) effects. We also included the same fixed-effect covariates (age, sex, and genetic principal components) as in our primary models described above (not shown in equation). Because the sample of siblings (N = 1702) was considerably smaller than the full analytic sample (N = 5517), we derived the population-level estimates (the ‘Total’ effects) between PGS and outcomes in a sample of equivalent size (and statistical power). See online Supplemental Methods for additional details.
Results
Demographic descriptive statistics for the baseline analytic sample (max N = 5517) are available in Table 1. The cognitive and non-cognitive PGS were somewhat negatively correlated with one another (r = −0.12, p < 2 × 10−16) in this sample.
Table 1.
ABCD European ancestry baseline demographic table
| Variable | Mean (SD)/n (%) baseline |
|---|---|
| Sex (male) | 2612(47.0%) |
| Age (years) | 9.93(0.63) |
| Household income | |
| < $49 000 | 658 (12.42%) |
| $50 000–$74 999 | 717 (13.53%) |
| $75 000–$99 999 | 896(16.93%) |
| $100 000–$199 999 | 2178(41.12%) |
| > $200 000 | 849(16.00%) |
| Highest caregiver education | |
| Less than high school | 30(0.54%) |
| High school degree or equivalent | 183(3.30%) |
| Some college, associate degree | 1046(18.83%) |
| College degree | 1753(31.57%) |
| Master's degree | 1723(31.01%) |
| Doctorate/professional degree | 820(14.75%) |
| Parental marital status | |
| Married or co-habiting | 4721(85.1%) |
| Widowed | 41(0.7%) |
| Divorced/separated | 620(11.2%) |
| Never married | 164(3.0%) |
Behavioral and sociodemographic outcomes
Cognition (Fig. 2a; online Supplemental Table S4)
Figure 2.

Associations between cognitive and non-cognitive PGS and neurocognition, risk-tolerance, personality & decision making, substance initiation, and psychopathology. Blue and purple asterisks correspond to significant associations (pfdr < 0.05) between the outcome measures of (a) cognition, (b) risk tolerance, personality, & decision-making, (c) substance initiation, and (d) psychopathology and cognitive PGS or both PGS, respectively. Blue hashtags correspond to associations that are significantly different for the cognitive PGS compared to the non-cognitive PGS. ADHD, attention deficit hyperactivity disorder; ASD, autism spectrum disorder; MDD, major depressive disorder; OCD, obsessive compulsive disorder; PLE, psychotic-like experiences.
The cognitive PGS was positively associated with all three cognition measures (i.e. fluid, crystallized, total score); Bs > 0.136, pfdrs < 3.90 × 10−24). The non-cognitive PGS was associated with crystallized intelligence and total scores (Bs > 0.047, pfdrs < 3.83 × 10−9), but not fluid intelligence (B = 0.013, pfdr = 0.454). For all three measures, the cognition scores were more strongly associated with the cognitive PGS than non-cognitive PGS (pdiff < 0.05). Post-hoc analyses revealed that when we covaried for SES (family income and neighborhood deprivation index), associations were somewhat attenuated for both the cognitive and non-cognitive PGS (online Supplemental Table S5). For example, associations with crystallized intelligence were attenuated for both the cognitive PGS (original Beta = 0.244, s.e. = 0.013; Beta when covaried by SES = 0.220, s.e. = 0.013) and the non-cognitive PGS (original Beta = 0.082, s.e. = 0.014; Beta when covaried by SES = 0.061, s.e. = 0.013).
Risk tolerance, personality, & decision-making. (Fig. 2b; online Supplemental Table S4)
The cognitive PGS was associated with greater odds of choosing the delayed reward (i.e. preferring the $115 in 3 months rather than the $75 in 3 days; OR 1.076, pfdr = 0.024) and risk tolerance (B = 0.031, pfdr = 0.044). The non-cognitive PGS was associated with decreased UPPS-P Lack of Perseverance and BAS Reward Responsiveness (Bs > −0.040, pfdrs < 0.016). Both the cognitive and non-cognitive PGS were associated with decreased BAS Drive and UPPS-P Positive and Negative Urgency (Bs > −0.081, pfdrs < 0.008). Post-hoc analyses showed that adjusting for both SES measures resulted in attenuation of associations (online Supplemental Table S5; e.g. associations with UPPS-P Positive Urgency were attenuated for both the cognitive PGS [original Beta = −0.048, s.e. = 0.014; Beta when covaried by SES = −0.039, s.e. = 0.015] and the non-cognitive PGS [original Beta = −0.080, s.e. = 0.014; Beta when covaried by SES = −0.062, s.e. = 0.015]).
Substance initiation. (Fig. 2c; online Supplemental Table S4)
No associations between alcohol initiation and either the cognitive or non-cognitive PGS were significant after multiple testing correction (pfdr > 0.05), and these associations did not significantly differ (pdiff > 0.05) between the cognitive and non-cognitive PGS.
Psychopathology. (Fig. 2d; online Supplemental Table S4)
The cognitive PGS was associated with decreased odds of ADHD diagnosis (OR 0.888, pfdr = 0.008) and bipolar diagnosis (OR 0.799, pfdr = 0.036). Both the cognitive and non-cognitive PGS were associated with lower severity of PLEs (Bs < −0.075, pfdrs < 0.003). The cognitive and non-cognitive PGS did not significantly differ from one another in their associations with any of the assessed mental health outcomes (pdiffs > 0.05). When we covaried for measures of SES, associations were somewhat attenuated (online Supplemental Table S5, e.g. associations with PLE: original cognitive PGS Beta = −0.074, s.e. = 0.013; Beta when covaried by SES = −0.060, s.e. = 0.014); original non-cognitive PGS Beta = −0.047, s.e. = 0.014; Beta when covaried by SES = −0.036, s.e. = 0.014).
Within-sibling analyses. (Fig. 3; online Supplemental Table S6).
Figure 3.

Total, between, and within-family estimates for the associations between cognitive and non-cognitive pgs and psychosocial measures. total, within- and between-family associations between Cognitive and Noncognitive PGS (p < 0.05) and significant measures in the domains of cognition, substance initiation, risk tolerance, personality, & decision-making, and psychopathology (i.e. outcomes with pfdr < 0.05 in Fig. 3 and online Supplemental Table 4). For the cognitive PGS, black, dark blue, light blue, and purple asterisks correspond to significant total, between-, within-family, and all three associations, respectively. For the non-cognitive PGS, red, orange, yellow, and purple asterisks correspond to significant total, between-, within-family, and all three associations, respectively.
Post-hoc analyses revealed that within-family variation in the cognitive PGS was associated with all three cognitive outcomes after accounting for between-family variation (Bs > 0.043, ps < 0.020). Within-family variation in both PGS were associated with PLEs after accounting for between-family variation, but within-family variation in the cognitive PGS was associated with decreased PLE severity scores (B = −0.044, p = 0.031) while within-family variation in the non-cognitive PGS was associated with increased PLE severity scores (B = 0.053, p = 0.008).
Brain Structure
Global volume indices. (Fig. 4a; online Supplemental Table S7)
Figure 4.

Significant associations between cognitive and non-cognitive pgs and neural indices of interest. significant associations between cognitive and non-cognitive PGS and significant imaging modalities including: (a) global brain indices, (b) cortical volume, (c) fractional anisotropy, and (d) mean diffusivity. Blue, orange, and purple asterisks correspond to significant associations (pfdr < 0.05) between the outcome measure and cognitive, non-cognitive, or both PGS respectively. Blue hashtags correspond to associations with cognitive PGS that are of significantly greater magnitude than for the non-cognitive PGS, while orange hashtags represent the opposite.
Both the cognitive and non-cognitive PGS were positively associated with all (n = 9) global volumes of interest, including whole brain, whole brain cortical volume, total left and right hemisphere cortical volume, left and right cerebral white matter, subcortical, supratentorial, and intracranial volume (Bs > 0.037, pfdrs < 0.002). All associations were significantly larger for the cognitive PGS relative to the non-cognitive PGS (all pdiff < 0.05).
Global white matter tracts. (Fig. 4a; online Supplemental Table S7)
The non-cognitive PGS was positively associated with the global measure of mean diffusivity (B = 0.029, pfdr = 0.023) but not with the global measure of average FA. The cognitive PGS was not associated with any global atlas tract fiber measures, but the association for the non-cognitive PGS was not significantly stronger than that observed for the cognitive PGS.
Global models covarying for SES. (online Supplemental Table S8)
When we covaried for SES, associations with global volumes were somewhat attenuated for both the cognitive and non-cognitive PGS (e.g. associations with total whole brain cortical volume: original cognitive PGS Beta = 0.095, s.e. = 0.012; Beta when covaried by SES = 0.073, s.e. = 0.012; original non-cognitive PGS Beta = 0.046, s.e. = 0.012; Beta when covaried by SES = 0.036, s.e. = 0.012). After controlling for SES, neither global white matter tract remained significant (pfdrs > 0.05).
Regional volume. (Fig. 4b; online Supplemental Table S9)
The cognitive PGS was positively associated with the following 13 regional brain volumes: (seven lateral and three bilateral: (1–2) bilateral inferior temporal gyri, (3–4) bilateral precentral gyri, (5–6) bilateral superior temporal gyri, (7) right (R) banks of superior temporal sulcus, (8) R caudal anterior cingulate, (9) R inferior parietal gyrus, (10) R middle temporal gyrus, (11) R pars orbitalis, (12) R temporal pole, (13) left (L) rostral anterior cingulate (B = 0.032–0.0383, pfdr = 0.005–0.035). The following regional associations were significantly greater for the cognitive PGS relative to the non-cognitive PGS: rh-pars orbitalis, lh-superiortemporal, rh-middle temporal, rh-inferior parietal, and bilateral inferior temporal volumes. The non-cognitive PGS was not significantly associated with any regional cortical volume.
DTI-FA. (Fig. 4c; online Supplemental Table S9)
The cognitive and non-cognitive PGS were associated with one bilateral white matter tract and 10 white matter tracts (one unilateral, one interhemispheric, and four bilateral), respectively. The cognitive PGS was positively associated with the bilateral corticospinal/pyramidal tract (B = 0.035–0.041, pfdr = 0.001–0.005). Tracts associated with the non-cognitive PGS included: (1–2) bilateral corticospinal/pyramidal, (3–4) bilateral superior corticostriatal-frontal cortex, (5–6) bilateral superior corticostriatal-parietal cortex, (7–8) bilateral superior corticostriatal, bilateral corpus callosum, and bilateral forceps minor (Bs = −0.029–0.038, pfdrs = 0.001–0.046). Both the forceps minor and the left superior corticostriatal-frontal cortex showed associations with the non-cognitive PGS that were of significantly greater magnitude (pdiffs < 0.05) than with the cognitive PGS.
DTI-MD. (Fig. 4d; online Supplemental Table S9)
The cognitive PGS was negatively associated with the bilateral corticospinal/pyramidal tracts (Bs = −0.035 to −0.027, pfdr = 0.003–0.026). The non-cognitive PGS was not significantly associated with any tract MD, but its association with the bilateral corticospinal/pyramidal tracts did not significantly differ from that observed for the cognitive PGS.
No significant regions in any neuroimaging modality showed evidence for laterality, (i.e. estimates within one hemisphere resided within the 95% confidence interval of the other hemisphere).
Regional volume and DTI models covarying for SES. (online Supplemental Table S10)
Post-hoc analyses indicated that when controlling for both measures of SES, associations between the cognitive PGS and seven of the 13 regional brain volumes (bilateral inferior temporal, bilateral precentral gyri, right (R) inferior parietal gyrus, R middle temporal gyrus, left superior temporal gyrus) and one of the two MD & FA white matter tracts respectively (i.e. left corticospinal/pyramidal) were attenuated but remained significant, while associations with the other six regional volumes were no longer significantly associated. For the non-cognitive PGS, when we covaried for both measures of SES, four out of the 10 FA white matter tracts (i.e. corpus callosum, forceps minor, left (L) superior corticostriatal-frontal cortex, L superior corticostriatal-parietal cortex only) remained significant; however, these associations were all reduced.
Within-sibling analyses. (online Supplemental Fig. S2; Supplemental Table S11)
Post-hoc analyses revealed that within-family variation in the cognitive PGS contributed to associations with two regional volume measures (right hemisphere caudal anterior cingulate and pars orbitalis) after accounting for significant between-family variation (ps < 0.048). The non-cognitive PGS showed significant within-family effects for the FA measure right superior corticostriate-frontal cortex (p < 0.034).
Discussion
Our study of phenotypic correlates of genetic propensity for cognitive and non-cognitive factors during late childhood (ns = 4688–5517) revealed four broad findings: First, cognitive and non-cognitive polygenic propensity both showed associations with cognition scores, impulsivity, and psychopathology. As seen for other polygenic associations with complex phenotypes, all effect sizes for the PGS were small (non-imaging: R2 > 2.19 × 10−5; global imaging: R2 > 9.0 × 10−4; regional imaging R2 > 9.67 × 10−4); thus, these PGS are not informative at the level of individual prediction. However, they could be relevant etiologically by providing insights into the differing associations with polygenic propensity for cognitive v. non-cognitive factors at a population level. Second, as revealed by our within-family analyses, associations between the cognitive PGS and some non-imaging outcomes (i.e. cognitive performance and PLE severity score) were consistent with direct genetic effects and/or evocative/active rGE, while the non-cognitive PGS showed potential direct genetic effects on PLE severity score (although in the opposite direction from the population-level estimate). Third, the cognitive PGS was more strongly associated with greater regional cortical volumes than the non-cognitive PGS, while the non-cognitive PGS generally showed greater magnitude of associations with white matter tract FA. Fourth, within-family analyses revealed significant within-family associations between the cognitive PGS and right hemisphere caudal anterior cingulate and pars orbitalis cortical volumes, while the non-cognitive PGS had significant within-family associations with the FA tract right superior corticostriate-frontal cortex. Overall, these findings converge with other recent data to suggest that the phenotypic correlates of genetic liability to cognitive and non-cognitive factors underlying educational attainment are present during childhood, prior to the completion of most education, and largely mirror patterns in adults.
Polygenic scores for cognitive and non-cognitive factors related to educational attainment have shared and unique correlates
The cognitive PGS was associated with higher fluid intelligence, crystallized intelligence, and a total combined score. The largest association was with crystallized intelligence, mirroring a prior ABCD study where researchers found that an intelligent PGS was most predictive of the crystallized intelligence score (Loughnan et al., 2023). We observed that the cognitive PGS was associated with several behavioral traits: increased risk tolerance and greater odds of choosing the delayed reward, decreased UPPS-P Positive and Negative Urgency and BAS Drive. Prior research has shown that in children, BAS Drive is associated with decreased inhibitory control (Vervoort et al., 2015). Thus, the current finding of decreased BAS Drive may indicate an increased tendency toward reserved, calculated, or cautious decision-making, which could facilitate further educational advancement (Almy, Kuskowski, Malone, Myers, & Luciana, 2018). Similarly, the observed association between cognitive PGS and greater odds of choosing a delayed reward (e.g. preferring $115 in 3 months over $75 in 3 days) is consistent with other work linking the ability to delay gratification among adolescents to higher academic achievement (Lee et al., 2012; Mischel, Shoda, & Rodriguez, 1989). Increases in Negative and Positive Urgency, theorized to be components of a broader dimension of impulsive action related to emotion (Billieux et al., 2021), are known risk factors for various psychopathologies (Elliott, Johnson, Pearlstein, Muñoz Lopez, & Keren, 2023). Thus, the negative associations between the cognitive PGS and both Negative and Positive Urgency are consistent with the negative associations between the cognitive PGS and decreased risk of ADHD, bipolar disorder, and severity ofPLEs. Conversely, associations between cognitive PGS and greater risk tolerance may reflect adaptive or positive risk taking associated with better educational engagement and outcomes (Duell & Steinberg, 2019, 2020).
While the non-cognitive PGS was also associated with higher crystallized and total cognition scores (but not fluid intelligence), associations between these outcomes and the non-cognitive PGS were significantly smaller than with the cognitive PGS (pdiffs = 2.2 × 10−16–1.51 × 10−12). Similar to findings with the cognitive PGS, the non-cognitive PGS was associated with lower PLE severity scores as well as decreased self-reported Positive and Negative Urgency, and Drive. However, only the non-cognitive PGS was associated with decreased Reward Responsiveness and Lack of Perseverance. These findings may reflect a tendency for academically high-achieving individuals to be motivated to not be distracted (e.g. self-regulate) and focus on tasks and accomplishing goals (McClelland & Cameron, 2011). As such, it may be that the genetic propensity for non-cognitive skills that contribute to educational attainment may relate more strongly to personality, while cognitive skills may be more associated with overall decision-making (e.g. taking calculated risks and weighing the impact of rewards [immediate v. delayed]).
In contrast to phenotypic outcomes described above, neuroimaging measures showed notable differences in associations between cognitive and non-cognitive PGS. Globally, all volume measures (N = 9; Fig. 4) were associated with both the cognitive and non-cognitive PGSs, but all associations were significantly stronger for the cognitive PGS. Across the majority of assessed non-global metrics, there were significant differences in the strength of association between the two PGS. For example, bilateral inferior temporal volume, a region implicated in studies of academic achievement, showed a significantly stronger association with the cognitive PGS (Mackey et al., 2015). Conversely, the tract forceps minor, which has previously been implicated in executive functioning and achievement (Loe, Adams, & Feldman, 2019), showed a significantly stronger (negative) association with the non-cognitive PGS. Overall, these findings suggest that genetic propensity for cognitive skills that contribute to educational attainment may relate more strongly to cortical volumes, while non-cognitive factors of educational success may be better reflected in the microstructure of the white matter tracts connecting between brain regions.
Evidence for direct and indirect genetic effects on phenotypes
Using post hoc within-sibling analyses, we contrasted within- v. between-family estimates for phenotypic correlates to determine whether associations with the PGS might be affected by confounding by passive rGE or other mechanisms. The cognitive PGS displayed significant within-sibling associations with all three cognition measures (online Supplemental Table S6, Fig. S4), suggesting that the associations between the cognitive PGS and these outcomes reflect direct genetic effects and/or evocative/active rGE. The cognitive PGS also had significant within-sibling associations with the right hemisphere caudal anterior cingulate and pars orbitalis measures, both of which have been associated with the salience brain network and cognitive salience (Snyder, Uddin, & Nomi, 2021). Both the cognitive and non-cognitive PGS displayed significant within-sibling associations with severity of PLEs; however, within-sibling differences in the cognitive PGS were associated with decreased severity of PLE, while within-sibling differences in the non-cognitive PGS were associated with increased severity of PLE. This divergence in direction of the association may be due to the PGS reflecting different functional outcomes or features of individuals at clinically high risk for psychosis. The negative association between cognitive PGS and PLE severity may reflect direct genetic effects related to aspects of cognitive symptoms associated with psychosis (e.g. cognitive dysfunction; Chun, Cooper, and Ellman, 2020), which are correlated with lower EduA. The observed positive association between greater PLE severity and non-cognitive PGS may reflect direct genetic contributions to creativity and openness to experience, which have been positively genetically correlated with schizophrenia (Duncan et al., 2018).
The non-cognitive PGS showed a significant positive within-sibling association with right superior corticostriate-frontal cortex FA, which is associated with many relevant pathways, including learning and reward sensitivity (Shipp, 2017). The null within-sibling findings for all other phenotypes for the non-cognitive PGS suggest that associations between the non-cognitive PGS and cognition scores and other outcomes may be due to passive rGE (e.g. parents with greater genetic predisposition for motivation or curiosity might enroll their children in additional courses or training) or other confounding factors that vary between families, such as assortative mating. Our findings for the non-cognitive PGS contrast with a recent preprint from Malanchini et al. (2024), which found evidence that both passive and evocative/active rGE mechanisms appear to contribute to the association between non-cognitive genetic factors and academic achievement in a sample of adolescents (Malanchini et al., 2024). Our conflicting results may be due to differences in statistical power, and/or potentially explained by a combination of social, educational, and cultural differences between the participants of the ABCD study (American) and the Malanchini study (England and Wales), which may influence related non-cognitive factors of educational attainment (Breinholt & Jaeger, 2020; Mendez & Zamarro, 2018).
We also note that covarying for two measures of SES (caregiver-reported income and neighborhood deprivation index) caused associations between the PGSs and outcomes to become somewhat attenuated. This suggests that SES-related factors may partly explain some of the observed associations between the cognitive and non-cognitive PGSs and outcomes in this study, a finding consistent with potential rGE and previous demonstrations of SES-related gene-environment correlations affecting genetic associations (Abdellaoui et al., 2019).
Findings in middle/late childhood largely mirror findings in adult samples
The current findings in this middle/late childhood sample largely align with findings in Demange et al. (2021) and other majority adult samples (Mitchell et al., 2022). Our neuroimaging results largely mirror those of the Demange et al., as well as prior research on academic achievement that have found that larger global brain volumes were positively associated with PGS for educational attainment in a sample of young adults (Mitchell et al., 2020). Given the congruent findings between previous studies performed in samples of adults (de Zeeuw et al., 2014; Demange et al., 2021), it may be that neural mechanisms associated with academic achievement are developed as early as middle/late childhood and may be temporally stable until later in life (Lövdén et al., 2020).
However, there were several domains where findings between adolescent and adult samples partly diverged. Demange et al. (2021) found that their cognitive factor was genetically correlated with lower risk tolerance, whereas we observed a positive association between the cognitive PGS and risk tolerance in this sample. These diverging findings may be explained by age differences across the samples. For example, in adolescents, cognitive abilities have previously been linked to greater risk tolerance (Andreoni, Di Girolamo, List, Mackevicius, & Samek, 2020). Another reason for this divergence could be typical adolescent neurodevelopment characterized by delayed cognitive control development (Shulman et al., 2016), leading to links between genetic propensity for higher cognitive skills and greater adaptative risk taking during middle/late childhood that may not persist to adulthood.
Additionally, Demange et al. (2021) found negative genetic correlations between the non-cognitive factor and health-risk behaviors (e.g. substance use), whereas the current study did not. Our null findings may be due to a combination of low endorsement of risky behaviors in our younger adolescent sample, as well as limitations of these measures in ABCD (i.e. the ABCD study has fewer assessments of health-risk behaviors, and we only analyzed alcohol initiation due to low endorsement of other substance use). In the domain of psychopathology, we found that both the PGSs were negatively associated with PLE severity scores, diverging from Demange et al.'s observed positive relationship between the non-cognitive factor and schizophrenia risk, as well as other studies that have linked greater creativity (Rajagopal et al., 2023) and other aspects of academic success to greater risk for schizophrenia (Karlsson, 2004). However, in post-hoc within-sibling analyses, the non-cognitive PGS score was positively associated with PLE severity, while the cognitive PGS remained negatively associated with PLE severity. These findings suggest that genetic propensity to cognitive skills may capture age-specific aspects of psychosocial functioning that are protective against development of prodromal psychosis in this adolescent sample (Park et al., 2023). Conversely, the positive association between within-sibling differences in the non-cognitive PGS and PLE severity suggests that this relationship may be consistent across child and adult samples when confounding factors (e.g. passive rGE, assortative mating) are accounted for.
Differentiation of cognitive and non-cognitive factors
Demange et al. (2021) saw differences in the cognitive and non-cognitive factors in terms of their genetic correlations with other phenotypes (e.g. schizophrenia and personality), suggesting some potential insights into the components of the ‘non-cognitive’ factor. Conversely, our study only found substantial differences between the cognitive and non-cognitive PGS associations with cognition-related outcomes and brain imaging metrics. One potential explanation is that there may not be enough variance in the psychopathology and health risk-related behavior outcomes in adolescence for these differences to manifest in the current sample. As such, the makeup of the ‘non-cognitive’ factor remains unclear, complicating the ability to interpret the observed associations. Future studies should focus on explicating the nature of traits involved in the non-cognitive factor, including assessing more in-depth ‘non-cognitive’ phenotypes and other relevant characteristics that contribute to academic achievement. Studies that employ multiple PGS including hypothesized components of non-cognitive skills (e.g. risk tolerance, personality, & decision-making) may be better able to distinguish the effects of genetic propensities for cognitive and non-cognitive factors relating to educational attainment. Further exploration across diverse age groups and subpopulations, including adolescents and adults at varying stages of educational attainment, may further elucidate how cognitive v. non-cognitive components of educational attainment differ in their relationship with complex phenotypes across the lifespan.
Strengths and limitations
Strengths of this study include a relatively large sample size, assessments taken during a developmentally important time period (middle/late childhood), deep phenotyping across multiple psychosocial and neuroimaging measures, and the incorporation of within-family analyses to control for potential confounding factors. However, this study is not without limitations. First, this study was performed only in a European ancestry subsample of individuals (due to the absence of available large-scale GWAS of educational attainment or cognitive performance in non-European ancestry samples, although small (N < 10 000) GWAS of these phenotypes in other ancestries do exist) whose caregivers volunteered to participate in the research study. This limits study generalizability (Schoeler et al., 2023), which may be especially important for better understanding potential relationships between facets of academic achievement and factors related to health and well-being. Furthermore, continuing to limit genetic analyses to individuals of European ancestry may exacerbate disparities in health-related research. However, given the small size of relevant GWAS in non-European ancestry samples, the resulting PGS would have low statistical power, and this could potentially make the interpretation of results difficult. We felt that the inclusion of what would have been a very under-powered PGS to extend our analyses to other ancestries was more likely to be harmful to those groups, especially given the potentially sensitive nature of phenotypes like cognition and educational attainment. However, new approaches are being developed to extend PGS to all populations (Lennon et al., 2024), even when the discovery GWAS are primarily of European ancestry, and this is an important topic worth further discussion in the behavior genetics field. Second, while an additional post hoc analysis was included to see whether non-cognitive and cognitive PGS associations were changed when covarying for SES, we suggest cautious interpretation of these results, as previous studies (Akimova, Breen, Brazel, & Mills, 2021) have highlighted that heritable covariates can potentially lead to epidemiological biases (e.g. collider bias). Third, the outcome measures studied were constrained by what was available in the ABCD Study; we were therefore unable to include potential measures of interest such as all of the big five personality traits, which are considered to be important to non-cognitive aspects of educational attainment and success, but were not assessed by the ABCD Study; Humphries & Kosse, 2017; Malanchini et al., 2024; Tucker-Drob et al., 2016; von Stumm, Hell, & Chamorro-Premuzic, 2011). However, we assessed aspects of the Big 5 by including the BIS sum scale to capture components of Neuroticism. Relatedly, we were unable to assess all the same imaging modalities that prior papers have reported on, including mode of anisotropy. Fourth, it is still unclear what exactly is represented by the ‘non-cognitive’ factor GWAS used to create PGS for the current study, and whether this non-cognitive factor would differ if the input GWASs were derived from samples of different ages (e.g. childhood, adolescence, older age). The authors of the original study (Demange et al., 2021) posit that preferences of risk tolerance, personality traits, decision-making, and socially desirable behaviors contribute to the non-cognitive factor of educational attainment captured by their study, as evidenced by patterns of genetic correlations with relevant phenotypes. However, as the authors note, the original GWAS of cognitive performance (Lee et al., 2018) may not have captured all relevant aspects of cognitive ability across the lifespan, and thus the separation of ‘cognitive’ v. ‘non-cognitive’ may be incomplete. Fifth, while the post hoc within-sibling analyses were designed to be unconfounded by environmental factors that vary between families, it is important to note that the original educational attainment GWAS (Okbay et al., 2022) is complex as it reflects direct and indirect genetic effects, such as genetic nurture, along with other gene-environment correlations and assortative mating. These factors may potentially inflate the strength of the associations observed with the resulting PGS. Sixth, these analyses do not control for all potential heritable components of environmental covariates that could influence these results (Allegrini et al., 2020). As such, existence of third variable confounding should not be ruled out from our work.
Conclusion
Overall, the results of this study provide evidence that as early as middle/late childhood, PGS for cognitive and non-cognitive facets of educational attainment share both overlapping and unique associations with psychosocial outcomes and neuroimaging measures. We speculate that the majority of these PGS associations are stable across adolescence and adulthood; however, further studies are needed before such a conclusion can be made. As the participants of the ABCD Study continue to age, this will be an invaluable sample in which to characterize the degree to which genetic and environmental effects on academic achievement, psychopathology, health behaviors, and neural phenotypes change across development into adulthood.
Supporting information
Gorelik et al. supplementary material
Gorelik et al. supplementary material
Gorelik et al. supplementary material
Acknowledgements
We are thankful to families who have participated in the ABCD Study as well as study staff and investigators.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0033291724002174
Authors’ contributions
ECJ, AJG, NRK, WZ, JB, APM, SEP, and SL conceptualized the study and planned analyses. AJG, SEP, APM, and NRK curated the phenotypic data. SL and ECJ created the PGS, and AJG, SL, ASH, and ECJ performed statistical analyses. PA, DAAB, AJG, and ECJ contributed to data visualizations. AJG, ECJ, RB, SL, JB, NM, and HM drafted the initial manuscript, and all coauthors edited the manuscript and provided important intellectual feedback.
Funding statement
This study was funded by R01DA054750 (RB, AA). ECJ was supported by K01DA051759. AJG was supported by NSF DGE-213989. NRK was supported by K23MH12179201. ASH was supported by K01AA030083. DAAB was supported by K99AA030808. APM was supported by T32DA015035. SEP was supported by F31AA029934. DAB was supported by R21AA027827. Data for this study were provided by the Adolescent Brain Cognitive Development (ABCD) study, which was funded by the National Institutes of Health (grants U01DA041022, U01DA041025, U01DA041028, U01DA041048, U01DA041089, U01DA041093, U01DA041106, U01DA041117, U01DA041120, U01DA041134, U01DA041148, U01DA041156, U01DA041174, U24DA041123, and U24DA041147) and additional federal partners (https://abcdstudy.org/federal-partners.html).
Competing interests
None.
Ethical standards
Working with ABCD NDA data was approved by the Washington University in St. Louis Institutional Review Board: IRB ID#201708123.
Data availability
All ABCD data used in this study are available through the National Institute of Mental Health Data Archive (NDA), which may be accessed here (https://nda.nih.gov).
Code availability
References
- 1000 Genomes Project Consortium, Auton, A., Brooks, L. D., Durbin, R. M., Garrison, E. P., Kang, H. M., Korbel, … Abecasis, G. R. (2015). A global reference for human genetic variation. Nature, 526(7571), 68–74. 10.1038/nature15393 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abdellaoui, A., Hugh-Jones, D., Yengo, L., Kemper, K. E., Nivard, M. G., Veul, L., … Visscher, P. M. (2019). Genetic correlates of social stratification in Great Britain. Nature Human Behaviour, 3(12), 1332–1342. 10.1038/s41562-019-0757-5 [DOI] [PubMed] [Google Scholar]
- Akimova, E. T., Breen, R., Brazel, D. M., & Mills, M. C. (2021). Gene-environment dependencies lead to collider bias in models with polygenic scores. Scientific Reports, 11(1), 9457. 10.1038/s41598-021-89020-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allegrini, A. G., Karhunen, V., Coleman, J. R. I., Selzam, S., Rimfeld, K., von Stumm, S., … Plomin, R. (2020). Multivariable G-E interplay in the prediction of educational achievement. PLOS Genetics, 16(11), e1009153. 10.1371/journal.pgen.1009153 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almy, B., Kuskowski, M., Malone, S. M., Myers, E., & Luciana, M. (2018). A longitudinal analysis of adolescent decision-making with the Iowa gambling task. Developmental Psychology, 54(4), 689–702. 10.1037/dev0000460 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andreoni, J., Di Girolamo, A., List, J. A., Mackevicius, C., & Samek, A. (2020). Risk preferences of children and adolescents in relation to gender, cognitive skills, soft skills, and executive functions. Journal of Economic Behavior & Organization, 179, 729–742. 10.1016/j.jebo.2019.05.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Austin, P. C. (2010). Estimating multilevel logistic regression models when the number of clusters is low: A comparison of different statistical software procedures. The International Journal of Biostatistics, 6(1), 16. 10.2202/1557-4679.1195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barch, D. M., Albaugh, M. D., Avenevoli, S., Chang, L., Clark, D. B., Glantz, M. D., … Sher, K. J. (2018). Demographic, physical and mental health assessments in the adolescent brain and cognitive development study: Rationale and description. Developmental Cognitive Neuroscience, 32, 55–66. 10.1016/j.dcn.2017.10.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartóń, K. (2024). MuMIn: Multi-Model Inference (Version 1.48.4) [Computer software]. CRAN. https://CRAN.R-project.org/package=MuMIn
- Basser, P. J., Mattiello, J., & Lebihan, D. (1994). Estimation of the effective self-diffusion tensor from the NMR spin echo. Journal of Magnetic Resonance, Series B, 103(3), 247–254. 10.1006/jmrb.1994.1037 [DOI] [PubMed] [Google Scholar]
- Bates, D., Maechler, M., Bolker, B., & Walker, S. (2015). Using lme4 [Software]. Journal of Statistical Software, 67, 1–48. 10.18637/jss.v067.i01 [DOI] [Google Scholar]
- Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society: Series B (Methodological), 57(1), 289–300. 10.1111/j.2517-6161.1995.tb02031.x [DOI] [Google Scholar]
- Billieux, J., Heeren, A., Rochat, L., Maurage, P., Bayard, S., Bet, R., … Baggio, S. (2021). Positive and negative urgency as a single coherent construct: Evidence from a large-scale network analysis in clinical and non-clinical samples. Journal of Personality, 89(6), 1252–1262. 10.1111/jopy.12655 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breinholt, A., & Jaeger, M. M. (2020). How does cultural capital affect educational performance: Signals or skills? The British Journal of Sociology, 71(1), 28–46. 10.1111/1468-4446.12711 [DOI] [PubMed] [Google Scholar]
- Brumpton, B., Sanderson, E., Heilbron, K., Hartwig, F. P., Harrison, S., Vie, GÅ, … Davies, N. M. (2020). Avoiding dynastic, assortative mating, and population stratification biases in Mendelian randomization through within-family analyses. Nature Communications, 11(1), Article 1. 10.1038/s41467-020-17117-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carver, C. S., & White, T. L. (1994). Behavioral inhibition, behavioral activation, and affective responses to impending reward and punishment: The BIS/BAS scales. Journal of Personality and Social Psychology, 67(2), 319–333. 10.1037/0022-3514.67.2.319 [DOI] [Google Scholar]
- Casey, B. J., Cannonier, T., Conley, M. I., Cohen, A. O., Barch, D. M., Heitzeg, M. M., … ABCD Imaging Acquisition Workgroup. (2018). The Adolescent Brain Cognitive Development (ABCD) study: Imaging acquisition across 21 sites. Developmental Cognitive Neuroscience, 32, 43–54. 10.1016/j.dcn.2018.03.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chamorro-Premuzic, T., & Furnham, A. (2003). Personality predicts academic performance: Evidence from two longitudinal university samples. Journal of Research in Personality, 37(4), 319–338. 10.1016/S0092-6566(02)00578-0 [DOI] [Google Scholar]
- Chun, C. A., Cooper, S., & Ellman, L. M. (2020). Associations of psychotic-like experiences, related symptoms, and working memory with functioning. European Psychiatry: The Journal of the Association of European Psychiatrists, 63(1), e20. 10.1192/j.eurpsy.2020.21 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cyders, M. A., Littlefield, A. K., Coffey, S., & Karyadi, K. A. (2014). Examination of a short version of the UPPS-P impulsive behavior scale. Addictive Behaviors, 39(9), 1372–1376. 10.1016/j.addbeh.2014.02.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demange, P. A., Malanchini, M., Mallard, T. T., Biroli, P., Cox, S. R., Grotzinger, A. D., … Nivard, M. G. (2021). Investigating the genetic architecture of noncognitive skills using GWAS-by-subtraction. Nature Genetics, 53(1), Article 1. 10.1038/s41588-020-00754-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson, B. C., Blacker, D., … Killiany, R. J. (2006). An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage, 31(3), 968–980. 10.1016/j.neuroimage.2006.01.021 [DOI] [PubMed] [Google Scholar]
- de Zeeuw, E. L., van Beijsterveldt, C. E. M., Glasner, T. J., Bartels, M., Ehli, E. A., Davies, G. E., … Boomsma, D. I. (2014). Polygenic scores associated with educational attainment in adults predict educational achievement and ADHD symptoms in children. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 165(6), 510–520. 10.1002/ajmg.b.32254 [DOI] [PubMed] [Google Scholar]
- Donnellan, M. B., Martin, M. J., & Senia, J. M. (2021). Genetic influences on the interactionist model of socioeconomic development: Incorporating polygenic scores for educational attainment into developmental research using the Family Transitions Project (FTP). Developmental Psychology, 57(2), 180–190. 10.1037/dev0000901 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duckworth, A. L., Peterson, C., Matthews, M. D., & Kelly, D. R. (2007). Grit: Perseverance and passion for long-term goals. Journal of Personality and Social Psychology, 92(6), 1087–1101. 10.1037/0022-3514.92.6.1087 [DOI] [PubMed] [Google Scholar]
- Duckworth, A. L., Quirk, A., Gallop, R., Hoyle, R. H., Kelly, D. R., & Matthews, M. D. (2019). Cognitive and noncognitive predictors of success. Proceedings of the National Academy of Sciences, 116(47), 23499–23504. 10.1073/pnas.1910510116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duell, N., & Steinberg, L. (2019). Positive risk taking in adolescence. Child Development Perspectives, 13(1), 48–52. 10.1111/cdep.12310 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duell, N., & Steinberg, L. (2020). Differential correlates of positive and negative risk taking in adolescence. Journal of Youth and Adolescence, 49(6), 1162–1178. 10.1007/s10964-020-01237-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duncan, L. E., Shen, H., Ballon, J. S., Hardy, K. V., Noordsy, D. L., & Levinson, D. F. (2018). Genetic correlation profile of schizophrenia mirrors epidemiological results and suggests link between polygenic and rare variant (22q11.2) cases of schizophrenia. Schizophrenia Bulletin, 44(6), 1350–1361. 10.1093/schbul/sbx174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elliott, M. L., Knodt, A. R., Ireland, D., Morris, M. L., Poulton, R., Ramrakha, S., … Hariri, A. R. (2020). What is the test-retest reliability of common task-functional MRI measures? New empirical evidence and a meta-analysis. Psychological Science, 31(7), 792–806. 10.1177/0956797620916786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elliott, M. V., Johnson, S. L., Pearlstein, J. G., Muñoz Lopez, D. E., & Keren, H. (2023). Emotion-related impulsivity and risky decision-making: A systematic review and meta-regression. Clinical Psychology Review, 100, 102232. 10.1016/j.cpr.2022.102232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan, C. C., Marshall, A., Smolker, H., Gonzalez, M. R., Tapert, S. F., Barch, D. M., … Herting, M. M. (2021). Adolescent Brain Cognitive Development (ABCD) study Linked External Data (LED): Protocol and practices for geocoding and assignment of environmental data. Developmental Cognitive Neuroscience, 52, 101030. 10.1016/j.dcn.2021.101030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ge, T., Chen, C.-Y., Ni, Y., Feng, Y.-C. A., & Smoller, J. W. (2019). Polygenic prediction via Bayesian regression and continuous shrinkage priors. Nature Communications, 10(1), Article 1. 10.1038/s41467-019-09718-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grotzinger, A. D., Rhemtulla, M., de Vlaming, R., Ritchie, S. J., Mallard, T. T., Hill, W. D., Ip, H. F., … Tucker-Drob, E. M. (2019). Genomic structural equation modelling provides insights into the multivariate genetic architecture of complex traits. Nature Human Behaviour, 3(5), Article 5. 10.1038/s41562-019-0566-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutacker, N., Kinge, J. M., & Olsen, J. A. (2023). Inequality in quality-adjusted life expectancy by educational attainment in Norway: An observational study. BMC Public Health, 23(1), 805. 10.1186/s12889-023-15663-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagler, D. J., Hatton, S., Cornejo, M. D., Makowski, C., Fair, D. A., Dick, A. S., … Dale, A. M. (2019). Image processing and analysis methods for the adolescent brain cognitive development study. NeuroImage, 202, 116091. 10.1016/j.neuroimage.2019.116091 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heath, A. C., Berg, K., Eaves, L. J., Solaas, M. H., Corey, L. A., Sundet, J., … Nance, W. E. (1985). Education policy and the heritability of educational attainment. Nature, 314(6013), Article 6013. 10.1038/314734a0 [DOI] [PubMed] [Google Scholar]
- Hicks, B. M., Clark, D. A., Deak, J. D., Schaefer, J. D., Liu, M., Jang, S., … Vrieze, S. I. (2021). Polygenic scores for smoking and educational attainment have independent influences on academic success and adjustment in adolescence and educational attainment in adulthood. PLoS ONE, 16(8), e0255348. 10.1371/journal.pone.0255348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horwitz, T. B., Balbona, J. V., Paulich, K. N., & Keller, M. C. (2023). Evidence of correlations between human partners based on systematic reviews and meta-analyses of 22 traits and UK Biobank analysis of 133 traits. Nature Human Behaviour, 7(9), 1234–1245. 10.1038/s41562-023-01672-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howe, L. J., Nivard, M. G., Morris, T. T., Hansen, A. F., Rasheed, H., Cho, Y., … Davies, N. M. (2022). Within-sibship genome-wide association analyses decrease bias in estimates of direct genetic effects. Nature Genetics, 54(5), 682–690. 10.1038/s41588-022-01062-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphries, J. E., & Kosse, F. (2017). On the interpretation of non-cognitive skills – what is being measured and why it matters. Journal of Economic Behavior & Organization, 136, 174–185. 10.1016/j.jebo.2017.02.001 [DOI] [Google Scholar]
- Karcher, N. R., & Barch, D. M. (2021). The ABCD study: Understanding the development of risk for mental and physical health outcomes. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology, 46(1), 131–142. 10.1038/s41386-020-0736-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlsson, J. L. (2004). Psychosis and academic performance. The British Journal of Psychiatry: The Journal of Mental Science, 184, 327–329. 10.1192/bjp.184.4.327 [DOI] [PubMed] [Google Scholar]
- Kaufman, J., Birmaher, B., Brent, D., Rao, U., Flynn, C., Moreci, P., … Ryan, N. (1997). Schedule for affective disorders and schizophrenia for school-age children-present and lifetime version (K-SADS-PL): Initial reliability and validity data. Journal of the American Academy of Child and Adolescent Psychiatry, 36(7), 980–988. 10.1097/00004583-199707000-00021 [DOI] [PubMed] [Google Scholar]
- Kohler, R. J., Lichenstein, S. D., & Yip, S. W. (2022). Hyperbolic discounting rates and risk for problematic alcohol use in youth enrolled in the adolescent brain and cognitive development study. Addiction Biology, 27(2), e13160. 10.1111/adb.13160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kovas, Y., Garon-Carrier, G., Boivin, M., Petrill, S. A., Plomin, R., Malykh, S. B., … Vitaro, F. (2015). Why children differ in motivation to learn: Insights from over 13000 twins from 6 countries. Personality and Individual Differences, 80, 51–63. 10.1016/j.paid.2015.02.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lam, M., Awasthi, S., Watson, H. J., Goldstein, J., Panagiotaropoulou, G., Trubetskoy, V., … Ripke, S. (2020). RICOPILI: Rapid imputation for consortias pipeline. Bioinformatics (Oxford, England), 36(3), 930–933. 10.1093/bioinformatics/btz633 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, N. C., Krabbendam, L., Dekker, S., Boschloo, A., de Groot, R. H. M., & Jolles, J. (2012). Academic motivation mediates the influence of temporal discounting on academic achievement during adolescence. Trends in Neuroscience and Education, 1(1), 43–48. 10.1016/j.tine.2012.07.001 [DOI] [Google Scholar]
- Lee, J. J., Wedow, R., Okbay, A., Kong, E., Maghzian, O., Zacher, M., Nguyen-Viet, T. A., … Cesarini, D. (2018). Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nature Genetics, 50(8), 1112–1121. 10.1038/s41588-018-0147-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lennon, N. J., Kottyan, L. C., Kachulis, C., Abul-Husn, N. S., Arias, J., Belbin, G., … Kenny, E. E. (2024). Selection, optimization and validation of ten chronic disease polygenic risk scores for clinical implementation in diverse US populations. Nature Medicine, 30(2), 480–487. 10.1038/s41591-024-02796-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loe, I. M., Adams, J. N., & Feldman, H. M. (2019). Executive function in relation to white matter in preterm and full term children. Frontiers in Pediatrics, 6, 418. 10.3389/fped.2018.00418 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loughnan, R. J., Palmer, C. E., Thompson, W. K., Dale, A. M., Jernigan, T. L., & Fan, C. (2023). Intelligence polygenic score is more predictive of crystallized measures: Evidence from the adolescent brain cognitive development (ABCD) study. Psychological Science, 34(6), 714–725. 10.1177/09567976231160702 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lövdén, M., Fratiglioni, L., Glymour, M. M., Lindenberger, U., & Tucker-Drob, E. M. (2020). Education and cognitive functioning across the life span. Psychological Science in the Public Interest, 21(1), 6–41. 10.1177/1529100620920576 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luciana, M., Bjork, J. M., Nagel, B. J., Barch, D. M., Gonzalez, R., Nixon, S. J., & Banich, M. T. (2018). Adolescent neurocognitive development and impacts of substance use: Overview of the adolescent brain cognitive development (ABCD) baseline neurocognition battery. Developmental Cognitive Neuroscience, 32, 67–79. 10.1016/j.dcn.2018.02.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackey, A. P., Finn, A. S., Leonard, J. A., Jacoby Senghor, D. S., West, M. R., Gabrieli, C. F. O., & Gabrieli, J. D. E. (2015). Neuroanatomical correlates of the income achievement gap. Psychological Science, 26(6), 925–933. 10.1177/0956797615572233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malanchini, M., Engelhardt, L. E., Grotzinger, A. D., Harden, K. P., & Tucker-Drob, E. M. (2019). “Same but different”: Associations between multiple aspects of self-regulation, cognition, and academic abilities. Journal of Personality and Social Psychology, 117(6), 1164–1188. 10.1037/pspp0000224 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malanchini, M., Allegrini, A. G., Nivard, M. G., Biroli, P., Rimfeld, K., Cheesman, R., … Plomin, R. (2024). Genetic associations between non-cognitive skills and academic achievement over development. Nature Human Behaviour. 10.1038/s41562-024-01967-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin, A. R., Gignoux, C. R., Walters, R. K., Wojcik, G. L., Neale, B. M., Gravel, S., … Kenny, E. E. (2017). Human demographic history impacts genetic risk prediction across diverse populations. American Journal of Human Genetics, 100(4), 635–649. 10.1016/j.ajhg.2017.03.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin, A. R., Kanai, M., Kamatani, Y., Okada, Y., Neale, B. M., & Daly, M. J. (2019). Clinical use of current polygenic risk scores may exacerbate health disparities. Nature Genetics, 51(4), Article 4. 10.1038/s41588-019-0379-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClelland, M. M., & Cameron, C. E. (2011). Self-regulation and academic achievement in elementary school children. New Directions for Child and Adolescent Development, 2011(133), 29–44. 10.1002/cd.302 [DOI] [PubMed] [Google Scholar]
- Mendez, I., & Zamarro, G. (2018). The intergenerational transmission of noncognitive skills and their effect on education and employment outcomes. Journal of Population Economics, 31(2), 521–560. 10.1007/s00148-017-0661-0 [DOI] [Google Scholar]
- Miller, A. P., Baranger, D. A. A., Paul, S. E., Garavan, H., Mackey, S., Tapert, S. F., … Bogdan, R. (2024). Neuroanatomical variability associated with early substance use initiation: Results from the ABCD study [Preprint]. medRxiv. 10.1101/2024.03.06.24303876 [DOI] [Google Scholar]
- Mischel, W., Shoda, Y., & Rodriguez, M. I. (1989). Delay of gratification in children. Science (New York, N.Y.), 244(4907), 933–938. 10.1126/science.2658056 [DOI] [PubMed] [Google Scholar]
- Mitchell, B. L., Cuéllar-Partida, G., Grasby, K. L., Campos, A. I., Strike, L. T., Hwang, L.-D., … Rentería, M. E. (2020). Educational attainment polygenic scores are associated with cortical total surface area and regions important for language and memory. NeuroImage, 212, 116691. 10.1016/j.neuroimage.2020.116691 [DOI] [PubMed] [Google Scholar]
- Mitchell, B. L., Hansell, N. K., McAloney, K., Martin, N. G., Wright, M. J., Renteria, M. E., & Grasby, K. L. (2022). Polygenic influences associated with adolescent cognitive skills. Intelligence, 94, 101680. 10.1016/j.intell.2022.101680 [DOI] [Google Scholar]
- Moffitt, T. E., Arseneault, L., Belsky, D., Dickson, N., Hancox, R. J., Harrington, H., … Caspi, A. (2011). A gradient of childhood self-control predicts health, wealth, and public safety. Proceedings of the National Academy of Sciences, 108(7), 2693–2698. 10.1073/pnas.1010076108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagelkerke, N. J. D. (1991). A note on a general definition of the coefficient of determination. Biometrika, 78(3), 691–692. 10.1093/biomet/78.3.691 [DOI] [Google Scholar]
- Noftle, E. E., & Robins, R. W. (2007). Personality predictors of academic outcomes: Big five correlates of GPA and SAT scores. Journal of Personality and Social Psychology, 93(1), 116–130. 10.1037/0022-3514.93.1.116 [DOI] [PubMed] [Google Scholar]
- Okbay, A., Wu, Y., Wang, N., Jayashankar, H., Bennett, M., Nehzati, S. M., Sidorenko, J., … Young, A. I. (2022). Polygenic prediction of educational attainment within and between families from genome-wide association analyses in 3 million individuals. Nature Genetics, 54(4), Article 4. 10.1038/s41588-022-01016-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pagliaccio, D., Luking, K. R., Anokhin, A. P., Gotlib, I. H., Hayden, E. P., Olino, T. M., … Barch, D. M. (2016). Revising the BIS/BAS scale to study development: Measurement invariance and normative effects of age and sex from childhood through adulthood. Psychological Assessment, 28(4), 429–442. 10.1037/pas0000186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park, J., Lee, E., Cho, G., Hwang, H., Kim, B., Kim, G., … Cha, J. (2023). Gene-environment pathways to cognitive development and psychotic-like experiences in children. eLife, 12, Article e88117. 10.7554/eLife.88117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raghupathi, V., & Raghupathi, W. (2020). The influence of education on health: An empirical assessment of OECD countries for the period 1995–2015. Archives of Public Health, 78(1), 20. 10.1186/s13690-020-00402-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajagopal, V. M., Ganna, A., Coleman, J. R. I., Allegrini, A., Voloudakis, G., Grove, J., … Demontis, D. (2023). Genome-wide association study of school grades identifies genetic overlap between language ability, psychopathology and creativity. Scientific Reports, 13(1), Article 1. 10.1038/s41598-022-26845-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schoeler, T., Speed, D., Porcu, E., Pirastu, N., Pingault, J.-B., & Kutalik, Z. (2023). Participation bias in the UK Biobank distorts genetic associations and downstream analyses. Nature Human Behaviour, 7(7), Article 7. 10.1038/s41562-023-01579-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selzam, S., Ritchie, S. J., Pingault, J.-B., Reynolds, C. A., O'Reilly, P. F., & Plomin, R. (2019). Comparing within- and between-family polygenic score prediction. The American Journal of Human Genetics, 105(2), 351–363. 10.1016/j.ajhg.2019.06.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shipp, S. (2017). The functional logic of corticostriatal connections. Brain Structure and Function, 222(2), 669–706. 10.1007/s00429-016-1250-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shulman, E. P., Smith, A. R., Silva, K., Icenogle, G., Duell, N., Chein, J., & Steinberg, L. (2016). The dual systems model: Review, reappraisal, and reaffirmation. Developmental Cognitive Neuroscience, 17, 103–117. 10.1016/j.dcn.2015.12.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silventoinen, K., Jelenkovic, A., Sund, R., Latvala, A., Honda, C., Inui, F., Tomizawa, R., … Kaprio, J. (2020). Genetic and environmental variation in educational attainment: An individual-based analysis of 28 twin cohorts. Scientific Reports, 10(1), 1. 10.1038/s41598-020-69526-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snyder, W., Uddin, L. Q., & Nomi, J. S. (2021). Dynamic functional connectivity profile of the salience network across the life span. Human Brain Mapping, 42(14), 4740–4749. 10.1002/hbm.25581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taliun, D., Harris, D. N., Kessler, M. D., Carlson, J., Szpiech, Z. A., Torres, R., … Abecasis, G. R. (2021). Sequencing of 53831 diverse genomes from the NHLBI TOPMed program. Nature, 590(7845), 7845. 10.1038/s41586-021-03205-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tucker-Drob, E. M., & Harden, K. P. (2012). Learning motivation mediates gene-by-socioeconomic status interaction on mathematics achievement in early childhood. Learning and Individual Differences, 22(1), 37–45. 10.1016/j.lindif.2011.11.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tucker-Drob, E. M., Briley, D. A., Engelhardt, L. E., Mann, F. D., & Harden, K. P. (2016). Genetically-mediated associations between measures of childhood character and academic achievement. Journal of Personality and Social Psychology, 111(5), 790–815. 10.1037/pspp0000098 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vervoort, L., Vandeweghe, L., Vandewalle, J., Van Durme, K., Vandevivere, E., Wante, L., … Braet, C. (2015). Measuring punishment and reward sensitivity in children and adolescents with a parent-report version of the BIS/BAS scales. Personality and Individual Differences, 87, 272–277. 10.1016/j.paid.2015.08.024 [DOI] [Google Scholar]
- Volkow, N. D., Koob, G. F., Croyle, R. T., Bianchi, D. W., Gordon, J. A., Koroshetz, W. J., Pérez-Stable, E. J., … Weiss, S. R. B. (2018). The conception of the ABCD study: From substance use to a broad NIH collaboration. Developmental Cognitive Neuroscience, 32, 4–7. 10.1016/j.dcn.2017.10.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Stumm, S., Hell, B., & Chamorro-Premuzic, T. (2011). The hungry mind: Intellectual curiosity is the third pillar of academic performance. Perspectives on Psychological Science: A Journal of the Association for Psychological Science, 6(6), 574–588. 10.1177/1745691611421204 [DOI] [PubMed] [Google Scholar]
- Watts, A. L., Smith, G. T., Barch, D. M., & Sher, K. J. (2020). Factor structure, measurement and structural invariance, and external validity of an abbreviated youth version of the UPPS-P Impulsive Behavior Scale. Psychological Assessment, 32(4), 336–347. 10.1037/pas0000791 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wulfert, E., Block, J. A., Santa Ana, E., Rodriguez, M. L., & Colsman, M. (2002). Delay of gratification: Impulsive choices and problem behaviors in early and late adolescence. Journal of Personality, 70(4), 533–552. 10.1111/1467-6494.05013 [DOI] [PubMed] [Google Scholar]
- Yengo, L., Robinson, M. R., Keller, M. C., Kemper, K. E., Yang, Y., Trzaskowski, M., … Visscher, P. M. (2018). Imprint of assortative mating on the human genome. Nature Human Behaviour, 2(12), 948–954. 10.1038/s41562-018-0476-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young, A. I., Frigge, M. L., Gudbjartsson, D. F., Thorleifsson, G., Bjornsdottir, G., Sulem, P., … Kong, A. (2018). Relatedness disequilibrium regression estimates heritability without environmental bias. Nature Genetics, 50(9), Article 9. 10.1038/s41588-018-0178-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zajacova, A., & Lawrence, E. M. (2018). The relationship between education and health: Reducing disparities through a contextual approach. Annual Review of Public Health, 39, 273–289. 10.1146/annurev-publhealth-031816-044628 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Gorelik et al. supplementary material
Gorelik et al. supplementary material
Gorelik et al. supplementary material
Data Availability Statement
All ABCD data used in this study are available through the National Institute of Mental Health Data Archive (NDA), which may be accessed here (https://nda.nih.gov).