

Abstract
Changes in climate and atmospheric nitrogen (N) deposition caused pronounced changes in soil conditions and habitat suitability for many plant species over the latter half of the previous century. Such changes are expected to continue in the future with anticipated further changing air temperature and precipitation that will likely influence the effects of N deposition. To investigate the potential long-term impacts of atmospheric N deposition on hardwood forest ecosystems in the eastern United States in the context of climate change, application of the coupled biogeochemical and vegetation community model VSD+PROPS was explored at three sites in New Hampshire, Virginia, and Tennessee. This represents the first application of VSD+PROPS to forest ecosystems in the United States. Climate change and elevated (above mid-19th century) N deposition were simulated to be important factors for determining habitat suitability. Although simulation results suggested that the suitability of these forests to support the continued presence of their characteristic understory plant species might decline by the year 2100, low data availability for building vegetation response models with PROPS resulted in uncertain results at the extremes of simulated N deposition. Future PROPS model development in the United States should focus on inclusion of additional foundational data or alternate candidate predictor variables to reduce these uncertainties.
Keywords: Forest understory, biodiversity, nitrogen, climate change, acidification
Capsule:
Climate change and elevated N deposition were simulated to be important factors for determining habitat suitability for plants, and are expected to interact with changes in soil chemistry
Graphical Abstract
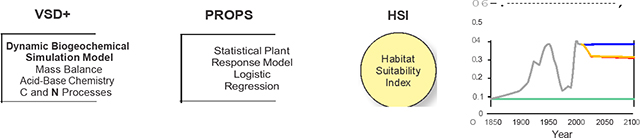
INTRODUCTION
The cycling of nitrogen (N) and carbon (C) has been significantly altered since preindustrial times, primarily through activities associated with the growth of the human population, production and application of N-based fertilizers, the prevalence of concentrated livestock operations with associated emissions of ammonia, and fossil fuel combustion (Schlesinger 1997, Vitousek et al. 1997, Galloway et al. 2008). Emissions of N-containing compounds such as NOx, NH3, and N2O and atmospheric deposition of N to terrestrial ecosystems have altered competitive relationships among plant species and decreased the cover of some N-efficient species by creating an environment that favors nitrophilous (prefer high N) species (Bobbink et al. 2010). Emissions of C and N, as components of greenhouse gases, have also contributed to changes in temperature and precipitation, which will likely continue in the future (Intergovernmental Panel on Climate Change [IPCC] 2013).
The bioavailability of N and patterns in air temperature and precipitation affect plant understory communities in multiple ways (Parmesan 2006, Bobbink et al. 2010, Porter et al. 2013). Increased N supply and climate change are recognized as two of the most important stressors to terrestrial plant biodiversity in the United States (Sala et al. 2000, Millennium Ecosystem Assessment [MEA] 2005). Changes in plant community structure caused by N and/or climate change contribute to declining species richness and biodiversity in many regions (Bobbink et al. 2010). Increased temperature and changes in precipitation affect habitat suitability and can cause additional loss of species (Parmesan 2006, Lenoir et al. 2008). Warmer climate may also increase soil mineral weathering and litter decomposition and thus enhance recovery of soils and vegetation negatively affected by past soil acidification caused by sulfur (S) and N deposition (Reinds et al. 2009, Aherne et al. 2012, Gaudio et al. 2015).
Effects of changes in N supply and climate on sensitive plant species have become important management issues for national parks and wilderness areas (Porter et al. 2005, 2012, 2013). Adverse effects of atmospheric N deposition on plant communities can be compounded by climate change (Davidson et al. 2011, Porter et al. 2013), which will affect virtually all aspects of N cycling (Suddick et al. 2013), and will have impacts on both terrestrial and aquatic ecosystems. Spatial and temporal patterns in temperature and precipitation are expected to change substantially over the next century (IPCC 2013, Melillo et al. 2014). Even as N deposition levels continue to decrease in the eastern US, the potential for forest ecosystem recovery from high cumulative historic N deposition is uncertain, especially in the context of a changing climate (Phelan et al. 2016).
Despite reductions in N emissions throughout much of the eastern US during recent decades, air concentrations and deposition of reactive N are several-fold higher than under preindustrial conditions in this region (Volpe Horii et al. 2005, Galloway et al. 2008, Sullivan 2017). Mitigation strategies for N emissions have not been as successful as those for S emissions (Aguillaume et al. 2016, Ochoa-Hueso et al. 2017, Sullivan 2017). Excess N supply increases growth of some plant species at the expense of others (Suding et al. 2005). Addition of N can affect terrestrial plant communities that are commonly limited or co-limited by N availability, including temperate hardwood forests (Elser et al. 2007).
Dynamic biogeochemical-ecological coupled models have been developed to make long- term projections of ecosystem response to multiple stressors at the species level (deVries et al. 2010). Two prominent examples of coupled models to examine the impacts from climate change and N deposition are ForSAFE-Veg (Belyazid et al. 2011a, Belyazid et al. 2011b) and the more recently developed VSD+PROPS (Reinds et al. 2014, Bonten et al. 2016). ForSAFE-Veg predates VSD+PROPS, and the latter offers potential for addressing some challenges associated with ForSAFE-Veg in two key areas. First, the biogeochemical components differ in that ForSAFE is more process oriented, and as such has many more data requirements that currently restrict its applicability across large areas compared with the simpler VSD+. Second, the vegetation components differ in that species in the Veg component of ForSAFE-Veg are defined by mathematical equations based on expert opinion to represent unobservable fundamental niches, whereas species in the PROPS component of VSD+PROPS are defined using statistical relationships based on empirical data and thus more closely approximate realized niches influenced by competition among species. ForSAFE-Veg has been more heavily used than VSD+PROPS in Europe (Belyazid et al. 2011a, Belyazid et al. 2011b, Rizzetto et al. 2016) and exclusively used in the United States (McDonnell et al. 2014, Phelan et al. 2016) to model the interactive effects from climate change and N deposition on plant communities. The extent to which modeling with VSD+PROPS would contribute to an improved understanding of vegetation community responses in the United States, where only ForSAFE-Veg has been applied thus far, is unknown.
The goal of this research was to assess the feasibility of VSD+PROPS for evaluating the effects of simultaneous changes in N deposition and climate on the habitat suitability of characteristic plant species in the eastern United States. Specific objectives included: (1) parametrization and calibration of the VSD+ model at target sites distributed across the eastern United States, (2) development of statistical models to estimate the probability of occurrence for plant species (PROPS) that can be found in the United States, (3) identification of data gaps that limit model application and development, and (4) linkage of VSD+ with PROPS to examine the interactive influences of climate and N deposition on plant communities through time. The N deposition loads at which plant species losses occur have not previously been determined except for a few correlational and site specific studies (Pardo et al. 2011, Simkin et al. 2016). However, such knowledge is critical to maintenance of biodiversity and biodiversity-related ecosystem services that affect human well-being (U.S. EPA, 2009).
METHODS
We developed PROPS models for 327 species and applied the VSD+PROPS model chain to three hardwood forest ecosystems located in different portions of this large region (in New Hampshire, Virginia, and Tennessee). The PROPS models were developed to represent statistical relations between species occurrence and five abiotic drivers of plant occurrence (Figure 1). Three of these drivers (air temperature, precipitation, and N deposition) are provided as direct input to PROPS. The other two drivers (soil solution pH and C:N) require output from a dynamic biogeochemical model to simulate change in species occurrence through time. The VSD+ model was used to generate time-series of soil solution pH and soil C:N to use as input for the PROPS model applications.
Figure 1.

General depiction of the VSD+ model framework including input requirements and outputs generated. Acronym definitions shown in the figure include: N (nitrogen), S (sulfur), BC (base cation), Cl (Chloride), CEC (cation exchange capacity), C (carbon), VSD+ (very simple dynamic model with carbon and nitrogen cycling), PROPS (probability of occurrence for plant species model)
Site Descriptions
Three sites were selected for VSD+PROPS modeling across a broad geographical region on different forest types, based partly on availability of input data for the models derived from field measurements, laboratory analyses, and other model results (e.g., MAGIC, PROFILE). Landscape protection status was used to evaluate candidates to include sites managed to maintain a primarily natural state. The selected sites are a northern hardwood site (Hubbard Brook) located in the White Mountains National Forest (NF) at the HBEF Long Term Ecological Research Station in New Hampshire; a mixed oak site (Piney River) in Shenandoah National Park (NP), Virginia; and a sugar maple-mixed oak site (Cosby Creek) in Great Smoky Mountains NP, Tennessee. Location and general site attributes are summarized in the Supplementary Material, SM1.
US PROPS Model Development
The vegetation model (PROPS) of the VSD+PROPS model chain consists of species- specific statistical response functions that define niche requirements related to aspects of climate, N deposition, and soil chemistry. A database of PROPS functions has previously been developed for a set of plant species in Europe (Wamelink et al. 2011, Reinds et al. 2014). We developed an analogous set of functions for plant species found in the United States to facilitate PROPS model application at our sites in the eastern United States using the following steps:
Compile a database of species occurrences and available associated abiotic predictor data from published literature and other databases (see Model Input below).
For plant species having sufficient occurrence data, use logistic regression to derive combined one-dimensional response functions based on a set of abiotic predictor variables, including aspects of N deposition, climate, and soil chemistry (see Statistical Modeling below).
Compile all response functions into a US PROPS database.
Model Input
Species occurrence data were primarily derived from a database developed for evaluating relations between herbaceous plant diversity and N deposition by Simkin et al. (2016). They compiled understory plant species data from 12 existing datasets, including the national Forest Inventory and Analysis (FIA) database for herbaceous species and other regional and state-based datasets. We augmented these data with tree seedling data from 2,016 plots included in the FIA database and additional understory plant occurrence data from 30 sites sampled along the Appalachian Trail corridor by Lawrence et al. (2015), which included two of the three model sites in this study (Piney River and Cosby Creek). Each vegetation survey was attributed with a set of variables for use in developing predictive models of plant occurrence The list of explanatory variables included N deposition, temperature, precipitation, soil pH, and soil C:N (see Supplementary Material, SM2 for additional detail).
Raw data of plant occurrences were assembled for more than 5,000 species, aggregated from more than 20,000 plots. Not all plots had all data necessary for modeling. Fewer than 10% of the ~20,000 plots were characterized with field measured soil pH and C:N. The final input database included 1,214 plots having presence/absence response data for 1,875 plant understory species and abiotic predictor data for all five drivers. A PROPS model was fit for each species that had 25 or more occurrences in the plant occurrence database. This resulted in the inclusion of 327 plant species in the initial version of the US PROPS database (275 herbaceous plants and the seedlings of 52 tree species). Of these 327 species, 181 had at least 50 occurrences and 76 had at least 100 occurrences.
Prior to fitting the PROPS models (see Statistical Modeling below for more details on PROPS model development), the abiotic factors were checked for normality and were considered skewed if the difference between mean and minimum was more than three times, or less than a third, of the difference between maximum and mean (Reinds et al. 2014). The distributions of precipitation and C:N were both determined to be skewed and these factors were log-transformed to obtain normal distributions. All transformed and untransformed abiotic conditions were then further normalized according to:
| (1) |
,where is the standard deviation of the variable . Among the five predictors, there was a moderate correlation between N deposition and temperature (r = 0.53) across the entire dataset; all other inter-predictor correlations were < 0.2. Relationships among N deposition, temperature, and biotic response will require further investigation.
Statistical Modeling
To populate the US PROPS database, logistic regression was used to fit one-dimensional occurrence probability curves individually by species for each of the five explanatory variables (given in the Supplementary Material, SM2). The individual one-dimensional curves were fitted with quadratic response functions constrained by a negative (or zero value) coefficient for the quadratic parameter to avoid a U-shaped curve. For each explanatory variable, a response function was derived, according to a logit function:
| (2) |
where is given by a quadratic equation:
| (3) |
where is the probability that a species will occur, and are the abiotic conditions. The parameters , and were determined by the model fit. All models were retained regardless of the extent to which plant occurrence was related to a given explanatory variable. The individual one-dimensional models were used to determine an overall occurrence probability based on the combined probability among all five abiotic factors as described in the Supplementary Material, SM3.
Measures of goodness of fit are not reported for the PROPS models. Such measures are difficult to estimate for logistic regression because the observed values are binominal (0 or 1) and the predictions are continuous values (probabilities between 0 and 1). Pseudo R-squared values (such as McFadden’s pseudo R-squared) are not reliable because they do not give a correlation between observed and predicted values and cannot be interpreted independently. Using an ROC (receiver operating characteristic) curve is not an option because in order to compare conditions (i.e. true positive, true negative, false positive, and false negative) a threshold value above which species occur is needed in order to decide whether or not a species occurs. Such a threshold value is difficult to select.
Habitat Suitability Index
The Habitat Suitability Index (HSI; Rowe et al. 2016) was used as an expression for plant diversity from PROPS modeling results:
| (4) |
where is the total number of indicator species, is the occurrence probability of species , and is the maximum occurrence probability of species . The PROPS model computes the occurrence probability, which is not a direct surrogate for abundance. Thus, the HSI is conceptually appropriate here. As local habitat conditions are simulated to change (i.e., variables in Table SM2–1), the probability of occurrence for individual species changes, as does the suitability of a site or habitat for that species. Sites that are favorable for many species are predicted to have a higher diversity and a higher HSI. Other indices like Simpson or Bray-Curtis can also be used to estimate diversity (Simpson 1949, Bray and Curtis 1957), but these are mostly based on species abundances (e.g., percent abundance ranging from 0–100%).
An increase in the value of HSI means that abiotic conditions have become more favorable for the selected set of indicator species. The HSI assumes a value between 0 and 1, where HSI = 1 indicates environmental conditions that are optimal for all indicator species included. In reality, HSI will be less than 1 because different species typically have different optimal abiotic conditions. Varying the membership of the n species allows the user to focus on different groups of species (e.g. N-fixers, disturbance-colonizers, etc.). However, even if the abiotic conditions are near optimal for all species (e.g., HSI approaches 1.0), various species may not be present if a factor not included in the model (e.g., light availability, herbivory) is limiting.
For the three model sites, vegetation surveys conducted in 2013 at Hubbard Brook (Phelan et al. 2016) and 2010 at Piney River and Cosby Creek (Lawrence et al. 2015) were used to identify candidate indicator species to use as the basis for calculating HSI. The list of candidate indicator species was compared with species included in the US PROPS database to select a set of indicator species for the HSI metric (Table 1). In general, selected indicator species were those considered to be characteristic of the vegetation association in which the site was contained. Details of methods used to select indicator species are detailed in the Supplementary Material, SM4.
Table 1.
List of indicator species used for VSD+PROPS modeling at Hubbard Brook, Piney River, and Cosby Creek.
| Indicator Species |
||||||
|---|---|---|---|---|---|---|
| Scientific Name | Common Name | Life Form | # of Occurrences for PROPS | Hubbard Brook | Piney River | Cosby Creek |
|
| ||||||
| Acer pensylvanicum | striped maple | tree | 103 | x | x | x |
| Acer saccharum | sugar maple | tree | 266 | x | x | |
| Actaea racemosa | black baneberry | herb | 34 | x | ||
| Ageratina altissima | white snakerooot | herb | 207 | x | ||
| Carya ovata | shagbark hickory | tree | 63 | x | ||
|
| ||||||
| Dennstaedtia punctilobula | eastern hayscented fern | fern | 117 | x | ||
| Dryopteris intermedia | intermediate woodfern | fern | 168 | x | ||
| Fagus grandifolia | American beech | tree | 196 | x | ||
| Fraxinus americana | white ash | tree | 284 | x | x | |
| Hydrophyllum virginianum | eastern waterleaf | herb | 36 | x | ||
|
| ||||||
| Laportea canadensis | wood nettle | herb | 72 | x | ||
| Maianthemum racemosum | feathery false lily of the valley | herb | 317 | x | x | |
| Medeola virginiana | Indian cucumber | herb | 128 | x | ||
| Oxalis montana | mountain woodsorrel | herb | 89 | x | ||
| Picea rubens | red spruce | tree | 68 | x | ||
|
| ||||||
| Prunus virginiana | choke cherry | tree | 133 | x | ||
| Quercus alba | white oak | tree | 151 | x | ||
| Quercus rubra | northern red oak | tree | 185 | x | ||
| Trientalis borealis | starflower | herb | 427 | x | ||
| Trillium undulatum | painted trillium | herb | 78 | x | ||
VSD+ Modeling
The Very Simple Dynamic (VSD) biogeochemical soil acidification model (Posch and Reinds 2009) is a simple dynamic extension of the steady-state Simple Mass Balance model (SMB model). Because of its simplicity, lower data requirements compared with other dynamic models (e.g. ForSAFE), and reasonable performance (Tominaga et al. 2010), the VSD model has been used for regional-scale assessments of air pollution impacts in Europe (e.g., Hettelingh et al. 2007, Achermann et al. 2008, Reinds et al. 2009). Such regional assessments are also desirable for US systems. The model VSD+ is an extension of the VSD model with added C and N dynamics (Bonten et al. 2016). This extension enables examination of acidification and eutrophication effects in a single modeling platform, including the effects of climate change on the sequestration of C in soils and the impacts of N supply on the diversity of plant species. VSD+ is used here to develop input data (soil solution pH and soil C:N) for PROPS application. Additional model description and calibration details are included in Supplementary Material, SM5).
Much of the VSD+ input data for the Hubbard Brook site was developed for a related modeling project previously conducted at the same site using the ForSAFE-Veg model (Phelan et al. 2016). Data from Piney River and Cosby Creek were acquired by Lawrence et al. (2015) for MAGIC (Cosby et al. 1985) modeling of stream chemistry at these two national park sites. Remaining input data were derived from nationally available datasets or from the pre-processor models MetHyd and GrowUp (Bonten et al. 2016), as described below. Data sources used to derive VSD+ inputs related to atmospheric deposition, soils, forest nutrient uptake, and N cycling are listed in Supplementary Material, SM6.
Soil Physiochemical Parameters
The upper 0.5 m of soil was assumed to represent the primary plant rooting depth. Soil physiochemical characteristics (bulk density, texture, cation exchange capacity, cation exchange constants) and soil mineral base cation (BC) weathering (BCw) for the Hubbard Brook site were obtained from Phelan et al. (2016; Supplementary Material Figure SM7–1). The Hubbard Brook soils data were derived from laboratory analyses of horizon-specific soil samples taken from a soil pit located on the vegetation plot (available on request from Scott Bailey swbailey@fs.fed.us). Because there were sufficient data for running PROFILE to estimate BCw at Hubbard Brook, BCw was simulated dynamically at that site under four future scenarios (Phelan et al. 2016), described below. For Piney River and Cosby Creek, soil physiochemical characteristics and soil BCw estimates were obtained from Lawrence et al. (2015). The Lawrence et al. (2015) study derived soil characteristics from individual soil horizons to a depth of 1.0 m, or until bedrock was encountered, from three soil pits located within the associated vegetation plot. An average of the data from the three pits was used for the modeling reported here. Estimates of BCw were determined with the MAGIC model at Piney River and Cosby Creek. Adequate data for BCw estimation with PROFILE were not available for these two sites. For all three sites, soil texture was obtained from SSURGO (http://www.nrcs.usda.gov/wps/portal/nrcs/main/soils/survey/. U.S. Geological Survey (USGS) geology maps (http://ngmdb.usgs.gov/ngmdb/ngmdb_home.html) were used to determine the absence of calcareous bedrock.
Reduction Factors
Maximum rates for mineralization, nitrification, and denitrification are established in VSD+. Reduction factors were used to decrease these maximum rates according to simulated soil temperature, moisture, and pH. The MetHyd pre-processor model was used to develop these reduction factors (Bonten et al. 2016). MetHyd is a simple soil water model that computes soil water content and leaching fluxes, both required by VSD+, using a bucket approach. It incorporates a snow-melt module for cold climates. The model is driven by input data regarding soil texture, organic matter, temperature, precipitation, and solar radiation. The effects of pH on mineralization, nitrification, and denitrification were incorporated during VSD+ simulation, using the simulated pH from the previous time-step.
Forest Nutrient Cycling
Tree N uptake and the content of C and N in litterfall contribute to the rate of N cycling and the fate of N in the soil environment. The GrowUp tool (v1.3.2; Bonten et al. 2016) and Forest Vegetation Simulator (FVS; Crookston and Dixon 2005, Keyster 2015) were used in conjunction with known disturbance events (e.g. tree harvests, windthrow) to derive these VSD+ inputs at all three sites, as described in Supplementary Material, SM8.
Atmospheric Deposition
Deposition inputs for Hubbard Brook were taken directly from the previous modeling at this site by Phelan et al. (2016) to facilitate future comparison between the ForSAFE-Veg and VSD+PROPS models (not explored here). The deposition (and climate; see below) scenario inputs at Hubbard Brook included both interannual variability and long-term trends. Modeling at Piney River and Cosby Creek was based solely on linear trends between specified points in time for all scenarios of deposition and climate (see below). The hindcast deposition estimates used for Piney River and Cosby Creek were only available at 10 year intervals and linear ramps between years were used to fill gaps. All atmospheric deposition inputs were determined as annual total (wet + dry) deposition. Historical reconstructions of total N, S, and BC deposition at Hubbard Brook were used as described by Phelan et al. (2016). Historical reconstructions of total N, S, and BC deposition at Piney River and Cosby Creek were developed with data from the Total Deposition Program of the NADP (TDEP; Schwede and Lear 2014), the Advanced Statistical Trajectory Regional Air Pollution (ASTRAP; Shannon 1998) model, Baker (1991), and interpolated NADP wet deposition (http://nadp.sws.uiuc.edu/ntn/maps.aspx). Additional details regarding deposition input data can be found in Supplementary Material, SM9.
In addition to historical reconstructions (HR) of atmospheric deposition, estimates of future deposition were based on anticipated emissions reductions associated with current emissions control policy (Tier III) in the eastern United States (U.S. EPA 2014) implemented by 2030 and held constant to the year 2100. Future deposition scenarios (Figure 2a) were specified as:
Figure 2.

Historical reconstructions (HR) and alternate scenarios of a) atmospheric deposition and b) climate used for VSD+PROPS simulations at Hubbard Brook (HB), Piney River (PR), and Cosby Creek (CC). Scenarios are labelled as: pre-industrial deposition (PID), modern deposition (MD), anticipated deposition (AD), modern climate (MC), and RCP6 climate (RCP6).
Pre-Industrial Deposition (PID): 1.0 kg N ha−1 yr−1 and 3.0 kg S ha−1 yr−1 held constant between 1850 and 2100 (Phelan et al. 2016)
Current Deposition (MD) – Historical reconstructions from 1850 to 2011, then TDEP 5-year (2009 – 2013) average held constant to 2100
Anticipated Deposition (AD) – Historical reconstructions from 1850 to 2011, linear ramp between 2011 and 2025, then Tier III from 2025 to 2100.
Climate
The PRISM database (800 m resolution; http://www.prism.oregonstate.edu/) and data from the NASA Earth Exchange Downscaled Climate Projections (NEX-DCP30 ensemble median; https://cds.nccs.nasa.gov/nex/) were used to develop time-series inputs of historical and future monthly air temperature and precipitation between 1850 and 2100. Two future climate change scenarios were developed and appended to estimates of historical climate: continuation of a modern (contemporary) climate and an anticipated future climate as estimated by IPCC (2013; RCP6.0) as described below. Alternate climate inputs are specified in Figure 2b, as:
Modern Climate (MC): PRISM data were aggregated to represent year 2010 as a 9- year average (2006 – 2014; PRISM data for 2015 were not available at the time of this study) and held constant to the year 2100.
RCP 6.0 (RCP6): Differences between averages of the last 10 years (2090–2099) and first 10 years (2006 – 2015) of the downscaled RCP6.0 output (multi-model median) were added to MC to represent year 2100 climate.
Air temperature estimates showed some historical increase, mainly at Hubbard Brook, and more marked future increases at all three sites under scenario RCP6, while precipitation amounts are projected to generally increase (Figure 2b).
Modeled Scenarios of Deposition and Climate
Four scenarios were modeled with VSD+PROPS by combining the deposition and climate inputs as: MC-PID, MC-MD, MC-AD, and RCP6-AD (Table 2). These scenarios capture the range of expected changes in key response drivers over time.
Table 2.
Combined deposition and climate scenarios used for VSD+PROPS modeling at Hubbard Brook (HB), Piney River (PR), and Cosby Creek (CC).
| Combined Scenario ID | Climate Scenario ID | Deposition Scenario ID | Climate Scenario Name | Deposition Scenario Name |
|---|---|---|---|---|
|
| ||||
| MC-PID | MC | PID | Modern Climate | Pre-Industrial Deposition |
| MC-MD | MC | MD | Modern Climate | Modern Deposition |
| MC-AD | MC | AD | Modern Climate | Anticipated Deposition |
| RCP6-AD | RCP6 | AD | RCP 6 Climate | Anticipated Deposition |
RESULTS
PROPS Model Development
Models were fit for 327 plant species that were represented in the database by at least 25 occurrences. The individual 1-dimensional models (e.g. Brachyelytrum erectum, Athyrium filix- femina, Agrostis perennans, and Poa pratensis) and the associated analysis indicated that the overall (five-dimensional) probabilities corresponded well with the one-dimensional curves for each predictor variable (Supplementary Material, SM10).
Total annual contemporary N deposition (2000 – 2013 average) used as the basis for the PROPS models ranged between 1.0 and 19.7 kg/ha/yr at plant survey locations (Figure 3a, Figure 3b). The majority of sites (83%) were located in the northern and eastern United States (red outline shown in Figure 3a), which ranged in N deposition from 3.8 to 19.7 (four sites with less than 4) kg N/ha/yr; Figure 3c). This has implications for PROPS model development, as will be discussed below.
Figure 3.

a) Total N deposition (2000 – 2013 average) for plots with co-located vegetation surveys and soil chemistry data (n = 1,214) used to develop the U.S. PROPS database. The red outline encompasses the plots located in the northern and eastern U.S. (n = 1,057). Survey data from plots located in the west are not relevant for the indicator species analyzed in this study. Among the modelled indicator species, only Maianthemum canadense is distributed throughout the geographic extent represented by all 1,214 plots. b) Distribution of total N deposition among all sites (n = 1,214) and c) distribution of total N deposition among sites located in the northern and eastern U.S. (n = 1,057) used for US PROPS model development.
Ecosystem Effects of Model Scenarios
Changes in Soil Chemistry
Changes in N deposition and climatic conditions across scenarios affected simulated values of soil solution pH and ANC, soil base saturation, soil C:N, and soil solution NO - concentration (Figure 4). Although soil solution ANC, soil base saturation, and soil solution NO3- concentration are not directly used to estimate HSI with PROPS, results for these variables are shown to provide additional context related to simulated soil acid-base chemistry and N availability. For soil solution pH and ANC, each modeled site showed a decrease for the historical reconstruction (HR) simulation period due to past increases in S and N deposition. Under the MC-MD scenario, soil solution pH and ANC either remained at approximately current levels (Hubbard Brook and Cosby Creek), or increased slightly (Piney River). Both variables moderately recovered under the MC-AD scenario across the three sites. Future climate change (RCP6-AD) resulted in soil solution recovery close to pre-industrial levels at Piney River but had little effect at Hubbard Brook and Cosby Creek.
Figure 4.

VSD+ modeled biogeochemical scenario responses at Hubbard Brook (HB), Piney River (PR), and Cosby Creek (CC). Scenarios are labeled as: historical reconstructions (HR), pre-industrial deposition (PID), modern deposition (MD), anticipated deposition (AD), modern climate (MC), and RCP6 climate (RCP6).
Soil base saturation remained above about 20% at all three sites under assumed long-term deposition at preindustrial levels (MC-PID). Base saturation was projected to have decreased during the 20th century in response to increasing acidic deposition at all three sites, with decreasing effects from north to south in the order of Hubbard Brook>Piney River>Cosby Creek. Base saturation at Hubbard Brook reached especially low values (<15%), well below the approximate 20% threshold for Al mobilization (Reuss and Johnson 1986), by the year 2000 (Figure 4). Base saturation at Piney River and Cosby Creek declined to near the 20% threshold over the historical record. Some base saturation recovery in the future was projected under the MC-AD scenario at all three sites. Further recovery under climate change (RCP6-AD) was only projected for Piney River.
Simulated soil C:N diverged after the early harvest events between MC-PID, which had a relatively stable soil C:N, and the other scenarios, where soil C:N declined through time. The lowest future value for soil C:N was simulated for the RCP6-AD scenario, indicating future climate change will likely increase soil nutrient availability. Future simulated N deposition reduced soil C:N appreciably, as compared with assumed preindustrial deposition at all three sites, due to N inputs that remain higher than N removal.
Soil solution NO3- concentrations, reflecting nutrient availability, were low across the time period simulated for the MC-PID scenario, and showed increases under historical reconstructions of N deposition during the latter part of the 20th century at all three sites. There was relatively little future change in soil solution NO3- concentration in response to any of the scenarios at all three sites, other than an increase under climate change (RCP6-AD) at Piney River. Short-term effects of clearcut logging are evident mainly at Piney River in the early 1900s for soil solution NO3- concentration, ANC, soil C:N, and base saturation. Simulations suggested a drop in nutrient BC and N uptake by the forest after clearcutting.
Changes in Habitat Suitability for Indicator Species
The HSI for the historical period (1850 – 2010) showed similar patterns among the model sites (Figure 5). At all sites, HSI increased during the first half of the 20th century and then showed a sharp drop during the period of highest N deposition (~ 1960 – 1990), followed by recovery in HSI with deposition reductions between 1990 and 2010. Between 2011 and 2100, HSI showed variable responses under the alternative future scenarios of deposition and climate.
Figure 5.

VSD+PROPS modeled HSI scenario responses at Hubbard Brook (HB), Piney River (PR), and Cosby Creek (CC). Scenarios are labeled as: historical reconstructions (HR), pre-industrial deposition (PID), modern deposition (MD), anticipated deposition (AD), modern climate (MC), and RCP6 climate (RCP6).
At Hubbard Brook, future HSI remained relatively constant under the ambient deposition scenario (MC-MD). Future decreases in N deposition (MC-AD and RCP6-AD) caused a decrease in HSI, which was amplified with future climate change (RCP6-AD). Nevertheless, HSI remained above the low values attained during the period that had the highest N inputs (1970– 1990) across all scenarios.
At Piney River, future HSI increased slightly under the ambient deposition scenario (MC-MD). Decreases in future N deposition (MC-AD and RCP6-AD) decreased HSI approximately to the year when deposition stopped declining (2025), and thereafter showed little change under constant N deposition, and a slight increase with future climate change (RCP6-AD).
At Cosby Creek, future HSI remained relatively constant under the ambient deposition and climate scenario (MC-MD). Future decreases in N deposition (MC-AD and RCP6-AD) again decreased HSI out to the year when deposition stopped declining (2025), with little effect afterward for any scenario.
DISCUSSION
In this modeling study, we set out to explore the feasibility of using VSD+PROPS to examine the potential effects of climate change and air pollution on temperate forest vegetation in the eastern United States. Input data were sufficiently available to calibrate and apply of VSD+ and estimate probability of species occurrence and HSI with PROPS at the three model sites. Simulated patterns of soil and soil solution acid-base chemistry (Figure 4) were in general agreement with available observations (Supplementary Material, SM11) and other dynamic modeling studies conducted at Hubbard Brook (Phelan et al. 2016; ForSAFE), Piney River, and Cosby Creek (Lawrence et al. 2015; MAGIC).
The VSD+ simulation results for soil solution pH and soil C:N were used as input to the PROPS models for simulating changes in HSI for the full simulation period (1850 – 2100). At all three sites, simulated HSI increased from relatively low levels during the pre-industrial period to peak values in the mid-20th century and decreased markedly during the period of highest N deposition (~1970–1990). Controlled experiments have found that N additions approximating N deposition within the range simulated by VSD+PROPS in our study can lead to reductions in vegetation diversity in just over one decade and that higher rates of N deposition can cause more rapid reductions in diversity (Clark and Tilman 2008). A 25-year N addition (35 kg N/ha/yr) experiment at the Fernow Experimental Forest of West Virginia suggested N-mediated changes in the herbaceous layer were primarily driven by increases in nitrophilous Rubus spp. (Gilliam et al. 2016). However, increases in Rubus spp. was dependent on light availability (Walter et al. 2016), which highlights the need for examining interactive effects between N deposition and light availability in future iterations of PROPS model development. Unfortunately, there were no Rubus species with sufficient occurrences with which to develop PROPS model functions for comparison with these empirical results. The observed patterns in HSI response in our study were affected, at least in part and perhaps in large part, by the range of available N deposition data for building PROPS models and associated low data availability at the tails of this distribution. The effect of this data limitation is discussed in more detail in the following section.
Overall, the modeling results for the historical period of deposition at each of the model sites suggested that N deposition, relative to the other four PROPS drivers, has had the largest impact on species occurrence probability and HSI (cf. correspondence between HSI and N deposition; Figure 6). The deposition of N was the primary driver of HSI response mainly because simulated changes in N deposition were largest relative to the other drivers. Annual average air temperature, for example, exhibited only a weak temporal trend during the historical period, in contrast with N deposition which increased from background levels (1 kg N ha−1 yr−1) to nearly 20 kg N ha−1 yr−1 during the same time period. A similar discrepancy in the importance of changes in temperature and atmospheric deposition regimes was found in other studies attributing vegetation changes to dramatically changing historical deposition levels more so than to the recent climate warming trends (Wason et al. 2017). The correspondence between changes in N deposition and HSI can be explained by the availability of input data for PROPS model development. The N deposition niches were necessarily narrow due to the available distribution of N deposition input used for building the PROPS models (see below). The US PROPS models were developed primarily from vegetation survey data centered roughly on the year 2000 (Simkin et al. 2016) and N deposition at this time was declining from historical peaks, but remained moderately high (> 8 kg/ha/yr) across much of the region except for the far Northeast (Figures 3b and 3c). Therefore, there are few species observations at low N deposition levels and consequently PROPS predicts low occurrence probabilities under low N deposition conditions. Thus, predicted occurrence probabilities under low future N deposition conditions may not be reliable. The uncertainty introduced by low data availability at either end of the input range of N deposition for building the PROPS models is further discussed in the example case study below.
Figure 6.

Simulated time-series of habitat suitability index (HSI; primary y-axes) and each of the five presumed main drivers of biological response (secondary y-axes) as represented in the MC-MD scenario at Cosby Creek.
Exploring Model Uncertainty: Cosby Creek Case Study
The Cosby Creek site is used here as an illustrative example of how HSI was related to each of the five PROPS drivers. The Cosby Creek site under the modern climate/deposition (MC-MD) scenario is used as an example because no major historical disturbance (e.g. tree harvesting, extensive blowdown) has occurred at this site, thus avoiding the confounding impacts on HSI due to the relatively large simulated changes in soil pH and C:N caused by the tree harvesting that occurred at Hubbard Brook and Piney River in the early 1900s.
The pattern of historical (1850 – 2011) N deposition throughout much of the eastern United States can be generally described as increasing from 1850 until 1970, peaking between 1970 and 1990, and then decreasing from 1990 to a level in 2011 that approximated the mid-20th century. This pattern is shown in the historical reconstructions of N deposition used for VSD+PROPS modeling at Hubbard Brook, Piney River, and Cosby Creek (Figure 2a) and it is generally widely documented in the literature in the eastern United States (e.g., Driscoll et al. 2001).
Among the five drivers that influence HSI at Cosby Creek during the HR period (Figure 6), estimated N deposition corresponds most closely with HSI and does so in two ways: 1) direct correspondence with HSI at deposition rates below 11.2 kg N/ha/yr and 2) inverse correspondence with HSI at deposition rates above 11.2 kg N/ha/yr. This might suggest increased habitat suitability as N deposition increases, until it reaches a tipping point. Further increases in N deposition decrease the suitability of the habitat. However, input data availability for building the PROPS models was limited at this high (> 14 kg N/ha/yr) level of N deposition, which may have contributed to the observed decline under high levels of simulated N deposition and obfuscated identification of a threshold of response. Furthermore, the HSI even at the peak deposition levels of the 1970s-1990s did not decline to levels below the HSI estimated for the 1850s.
The simulated probability of occurrence for individual species at Cosby Creek showed that each of the indicator plants responded similarly to peak deposition (Figure 7; analogous plots for Hubbard Brook and Piney River are shown in Supplementary Material, SM12). Deposition during this time period was in the range of 15 – 20 kg N/ha/yr. Very few data points from either this range or the lower end of the distribution (< 4 kg N/ha/yr) were used to build the PROPS functions (Figure 4). The PROPS model predicts that, for species distributed within the eastern United States (Figure 3; red box) few would thrive at low (< 4 kg N/ha/yr) deposition, partly because plant occurrences were not available within this range of deposition. Because the PROPS functions were derived from a logistic model, no/low input data availability in a given range for an abiotic driver effectively means zero/little chance for any species to occur when simulating within that range. Simulations of habitat suitability with VSD+PROPS based on soil solution pH, soil C:N, deposition, or climate inputs near to or beyond the extreme values used to build the PROPS models should be considered highly uncertain.
Figure 7.

Simulated probability of occurrence for the individual indicator species at Cosby Creek under scenario MC-MD (Modern Climate – Modern Deposition).
Future work on PROPS model development might focus on addressing the reduction in HSI at the extremes of the available deposition data, which may be partly a result of data availability as well as mechanistic effects. This suggests that PROPS, because of its reliance on empirical data, may only be suitable in its current form for assessing effects of future decreases in N deposition in the eastern United States if these loading values occur within the range of observed N deposition. PROPS may not be suitable for assessing time to recovery, historical patterns of vegetation, or other questions that consider deposition levels beyond the range experienced by the available regional vegetation data. VSD+ does not have these limitations because the biogeochemistry is not dependent on the availability of understory vegetation data. Uncertainty in pH and C/N ratio simulated by VSD+ has been shown to be limited (Bonten et al. 2016).
There may be several opportunities for improving PROPS for application to the eastern United States, particularly, inclusion of additional sites at the upper and especially at the lower end of the N deposition distribution. Alternate candidate predictor variables might also be considered, such as light availability, cumulative N deposition, site factors influencing N availability, seasonal climate metrics, and soil moisture indices. Extending the tails of the observational record on the high and low end of the N deposition gradient to match the deposition range of the simulations should remove the biases towards lower HSI that inflate the number of predicted zero or near-zero probabilities of occurrence. Evaluating future iterations of PROPS models against independent test data will be required to further improve confidence in the model results. Data that reflect changes in plant communities over time will be particularly important. Testing the feasibility of a regional VSD+ model application is also an appropriate next step for evaluating biological response with VSD+PROPS at a scale more relevant to environmental policy and management. Given the current state of model development and availability of input data, results of this study should not be used as the basis for resource management or policy decisions. However, observed patterns are appropriate for guiding further studies of N deposition and climate change effects on vegetation.
CONCLUSIONS
The VSD+ model was able to reproduce values consistent with observations of soil chemistry at the study sites. Simulations using VSD+PROPS at these sites suggested that historical changes in N deposition had pronounced impacts on HSI. Following the strong recovery of HSI between 1990 and 2010 caused by decreases in N deposition, future changes in climate and further decreases in N deposition were projected to slightly decrease habitat suitability for the selected indicator plants. The simulated HSI response was driven primarily by N deposition, in part because this driver was the most variable through time. Lack of data at the low and high ends of the distribution of N deposition data used for building the PROPS functions contributed to uncertainty in HSI response for these simulated ranges of N deposition.
Supplementary Material
Highlights.
Changes in climate and N deposition are important factors affecting biodiversity We assess past and future impacts on plant diversity with a soil-plant model chain Species occurrence probability was mostly driven by changes in N deposition Further development of statistical plant response functions is needed
ACKNOWLEDGEMENTS
We thank R. Waring, J. Lynch, and J. Compton for reviewing this manuscript and providing insight. Model input and validation data were provided by S. Bailey, S. Belyazid, B. Cosby, C. Driscoll, G. Lawrence, J. Phelan, J. Quant, S. Simkin, and the Hubbard Brook Ecosystem Study of the USDA Forest Service. T. Evans, A. Hyduke, and W. Cass assisted with indicator species selection. This research was funded by a contract (EP-15-C-000044) between the U.S. Environmental Protection Agency and E&S Environmental Chemistry, Inc. The views expressed in this article are those of the authors and do not necessarily represent the views or policies of the U.S. Environmental Protection Agency.
REFERENCES
- Achermann B, Rihm B, and Kurz D. 2008. Switzerland. In Hettelingh J-P, Posch M. and Slootweg J. (Eds.). Critical Load, Dynamic Modelling and Impact Assessment in Europe. CCE Status Report 2008. Bilthoven, Netherlands. pp. 205–210. [Google Scholar]
- Aguillaume L, Rodrigo A, and Avila A. 2016. Long-term effects of changing atmospheric pollution on throughfall, bulk deposition and streamwaters in a Mediterranean forest. Sci. Total Environ. 544(Supplement C):919–928 10.1016/j.scitotenv.2015.12.017. [DOI] [PubMed] [Google Scholar]
- Aherne J, Posch M, Forsius M, Lehtonen A, and Härkönen K. 2012. Impacts of forest biomass removal on soil nutrient status under climate change: a catchment-based modelling study for Finland. Biogeochemistry 107(1):471–488 10.1007/s10533-010-9569-4. [DOI] [Google Scholar]
- Baker LA 1991. Regional estimates of dry deposition. Appendix B. In Charles DF (Ed.) Acidic Deposition and Aquatic Ecosystems: Regional Case Studies. Springer-Verlag, New York. pp. 645–652. [Google Scholar]
- Belyazid S, Kurz D, Braun S, Sverdrup H, Rihm B, and Hettelingh J-P. 2011a. A dynamic modelling approach for estimating critical loads of nitrogen based on plant community changes under a changing climate. Environ. Pollut 159:789–801 [DOI] [PubMed] [Google Scholar]
- Belyazid S, Sverdrup H, Kurz D, and Braun S. 2011b. Exploring ground vegetation change for different deposition scenarios and methods for estimating critical loads for biodiversity using the ForSAFE-Veg model in Switzerland and Sweden. Water Air & Soil Pollution 216:289–317 [Google Scholar]
- Bobbink R, Hicks K, Galloway J, Spranger T, Alkemade R, Ashmore M, Bustamante M, Cinderby S, Davidson E, Dentener F, Emmett B, Erisman J-W, Fenn M, Gilliam FS, Nordin A, Pardo L, and DeVries W. 2010. Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol. Appl 20(1):30–59 [DOI] [PubMed] [Google Scholar]
- Bonten LTC, Reinds GJ, and Posch M. 2016. A model to calculate effects of atmospheric deposition on soil acidification, eutrophication and carbon sequestration. Environ. Model. Software 79:75–84 10.1016/j.envsoft.2016.01.009. [DOI] [Google Scholar]
- Bray JR and Curtis JT. 1957. An ordination of upland forest communities of southern Wisconsin. Ecol. Monogr 27:325–349 [Google Scholar]
- Clark CM and Tilman D. 2008. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 451:712–715 [DOI] [PubMed] [Google Scholar]
- Cosby BJ, Hornberger GM, Galloway JN, and Wright RF. 1985. Modelling the effects of acid deposition: assessment of a lumped parameter model of soil water and streamwater chemistry. Water Resour. Res 21(1):51–63 [Google Scholar]
- Crookston NL and Dixon GE. 2005. The forest vegetation simulator: A review of its structure, content, and applications. Computers and Electronics in Agriculture 49(1):60–80 10.1016/j.compag.2005.02.003. [DOI] [Google Scholar]
- Davidson EA, David MB, Galloway JN, Goodale CL, Haeuber R, Harrison JA, Howarth RW, Jaynes DB, Lowrance RR, Thomas NB, Peel JL, Pinder RW, Porter E, Snyder CS, Townsend AR, and Ward MH. 2011. Excess nitrogen in the U.S. environment: Trends, risks, and solutions. Issues in Ecology(15) [Google Scholar]
- deVries W, Wamelink GWW, van Dobben H, Kros J, Reinds GJ, Mol-Dukstra JP, Smart SM, Evans CD, Rowe EC, Belyazid S, Sverdrup HU, van Hinsberg A, Posch M, Hettelingh J-P, Spranger T, and Bobbink R. 2010. Use of dynamic soil-vegetation models to assess impacts of nitrogen deposition on plant species composition: an overview. Ecol. Appl 20(1):60–79 [DOI] [PubMed] [Google Scholar]
- Driscoll CT, Lawrence GB, Bulger AJ, Butler TJ, Cronan CS, Eagar C, Lambert KF, Likens GE, Stoddard JL, and Weathers KC. 2001. Acidic deposition in the northeastern United States: sources and inputs, ecosystem effects, and management strategies. BioScience 51(3):180–198 [Google Scholar]
- Elser JJ, Bracken MES, Cleland EE, Gruner DS, Harpole WS, Hillebrand II H, Ngai JT, Seabloom EW, Shurin JB, and Smith JE. 2007. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine, and terrestrial ecosystems. Ecol. Lett 10(12):1135–1142 [DOI] [PubMed] [Google Scholar]
- Galloway JN, Townsend AR, Erisman JW, Bekunda M, Cai Z, Freney JR, Martinelli LA, Seitzinger SP, and Sutton MA. 2008. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 320(5878):889–892 10.1126/science.1136674. [DOI] [PubMed] [Google Scholar]
- Gaudio N, Belyazid S, Gendre X, Mansat A, Nicolas M, Rizzetto S, Sverdrup H, and Probst A. 2015. Combined effect of atmospheric nitrogen deposition and climate change on temperate forest soil biogeochemistry: A modeling approach. Ecol. Model 306:24–34 10.1016/j.ecolmodel.2014.10.002. [DOI] [Google Scholar]
- Gilliam FS, Welch NT, Phillips AH, Billmyer JH, Peterjohn WT, Fowler ZK, Walter CA, Burnham MB, May JD, and Adams MB. 2016. Twenty-five-year response of the herbaceous layer of a temperate hardwood forest to elevated nitrogen deposition. Ecosphere 7(4):Article e01250 10.1002/ecs2.1250. [DOI] [Google Scholar]
- Hettelingh J-P, Posch M, Slootweg J, Reinds GJ, Spranger T, and Tarrason L. 2007. Critical loads and dynamic modelling to assess European areas at risk of acidification and eutrophication. Water Air Soil Pollut: Focus 7:379–384 [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). 2013. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. (Stocker TF, Qin D, Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V. and Midgley PM [Eds.]). Cambridge University Press, Cambridge, United Kingdom & New York, NY, USA. [Google Scholar]
- Keyster CE 2015. Variant Overview. Forest Vegetation Simulator Internal Report. U. S. Department of Agriculture, Forest Service, Forest Management Service Center, Fort Collins, CO. [Google Scholar]
- Lawrence GB, Sullivan TJ, Burns DA, Bailey SA, Cosby BJ, Dovciak M, Ewing HA, McDonnell TC, Minocha R, Quant J, Rice KC, Siemion J, and Weathers K. 2015. Acidic Deposition along the Appalachian Trail Corridor and its Effects on Acid-Sensitive Terrestrial and Aquatic Resources. Results of the Appalachian Trail MEGA-Transect Atmospheric Deposition Effects Study. Natural Resource Report NPS/NRSS/ARD/NRR—2015/996. National Park Service, Fort Collins, CO. [Google Scholar]
- Lenoir J, Gegout JC, Marquet PA, de Ruffray P, and Brisse H. 2008. A significant shift in plant species optimum elevations during the 20th century. Science 320:1768–1771 [DOI] [PubMed] [Google Scholar]
- McDonnell TC, Belyazid S, Sullivan TJ, Sverdrup H, Bowman WD, and Porter EM. 2014. Modeled subalpine plant community response to climate change and atmospheric nitrogen deposition in Rocky Mountain National Park, USA. Environ. Pollut. 187:55–64 [DOI] [PubMed] [Google Scholar]
- Melillo JM, Richmond TC, and Yohe GW (Eds.). 2014. Climate Change Impacts in the United States: The Third National Climate Assessment. U.S. Global Change Research Program 841 pp. doi: 10.7930/J0Z31WJ2. [DOI] [Google Scholar]
- Millennium Ecosystem Assessment (MEA). 2005. Ecosystems and Human Well-Being: The Assessment Series (Four Volumes and Summary). Island Press, Washington, DC. [Google Scholar]
- Ochoa-Hueso R, Munzi S, Alonso R, Arróniz-Crespo M, Avila A, Bermejo V, Bobbink R, Branquinho C, Concostrina-Zubiri L, Cruz C, Cruz de Carvalho R, De Marco A, Dias T, Elustondo D, Elvira S, Estébanez B, Fusaro L, Gerosa G, Izquieta-Rojano S, Lo Cascio M, Marzuoli R, Matos P, Mereu S, Merino J, Morillas L, Nunes A, Paoletti E, Paoli L, Pinho P, Rogers IB, Santos A, Sicard P, Stevens CJ, and Theobald MR. 2017. Ecological impacts of atmospheric pollution and interactions with climate change in terrestrial ecosystems of the Mediterranean Basin: Current research and future directions. Environ. Pollut. 227(Supplement C):194–206 10.1016/j.envpol.2017.04.062. [DOI] [PubMed] [Google Scholar]
- Pardo LH, Fenn ME, Goodale CL, Geiser LH, Driscoll CT, Allen EB, Baron JS, Bobbink R, Bowman WD, Clark CM, Emmett B, Gilliam FS, Greaver TL, Hall SJ, Lilleskov EA, Liu L, Lynch JA, Nadelhoffer KJ, Perakis SS, Robin-Abbott MJ, Stoddard JL, Weathers KC, and Dennis RL. 2011. Effects of nitrogen deposition and empirical nitrogen critical loads for ecoregions of the United States. Ecol. Appl 21(8):3049–3082 [Google Scholar]
- Parmesan C. 2006. Ecological and evolutionary responses to recent climate change. Annual Review of Ecology, Evolution, and Systematics 37:637–669 [Google Scholar]
- Phelan J, Belyazid S, Jones P, Cajka J, Buckley J, and Clark C. 2016. Assessing the effects of climate change and air pollution on soil properties and plant diversity in sugar maple- beech-yellow birch hardwood forests in the northeastern United States: Model simulations from 1900–2100. Water Air & Soil Pollution 227:84 [Google Scholar]
- Porter E, Blett T, Potter DU, and Huber C. 2005. Protecting resources on federal lands: implications of critical loads for atmospheric deposition on nitrogen and sulfur. BioScience 55(7):603–612 [Google Scholar]
- Porter E, Sverdrup H, and Sullivan TJ. 2012. Estimating and mitigating the impacts of climate change and air pollution on alpine plant communities in national parks. Park Science 28(2):58–64. Available online at http://www.nature.nps.gov/ParkScience/index.cfm?ArticleID=513 [Google Scholar]
- Porter EM, Bowman WD, Clark CM, Compton JE, Pardo LH, and Soong JL. 2013. Interactive effects of anthropogenic nitrogen enrichment and climate change on terrestrial and aquatic biodiversity. Biogeochemistry 114:93–120 [Google Scholar]
- Posch M. and Reinds GJ. 2009. A very simple dynamic soil acidification model for scenario analyses and target load calculations. Environ. Model. Software 24(3):329–340 10.1016/j.envsoft.2008.09.007. [DOI] [Google Scholar]
- Reinds GJ, Posch M, and Leemans R. 2009. Modelling recovery from soil acidification in European forests under climate change. Sci. Total Environ 407(21):5663–5673 10.1016/j.scitotenv.2009.07.013. [DOI] [PubMed] [Google Scholar]
- Reinds GJ, Mol-Dijkstra J, Bonten L, Wamelink W, deVries W, and Posch M. 2014. VSD+PROPS: Recent developments. Chapter 4. In Slootweg J, Posch M, Hettelingh J-P and Mathijssen L. (Eds.). Modelling and Mapping the Impacts of Atmospheric Deposition on Plant Species Diversity in Europe. CCE Status Report 2014. Coordination Centre for Effects pp. 47–53. [Google Scholar]
- Reuss JO and Johnson DW. 1986. Acid Deposition and the Acidification of Soils and Waters. Ecol. Series, Vol. 59, Springer-Verlag, New York. [Google Scholar]
- Rizzetto S, Belyazid S, Gégout J-C, Nicolas M, Alard D, Corcket E, Gaudio N, Sverdrup H, and Probst A. 2016. Modelling the impact of climate change and atmospheric N deposition on French forests biodiversity. Environ. Pollut 213:1016–1027 10.1016/j.envpol.2015.12.048. [DOI] [PubMed] [Google Scholar]
- Rowe EC, Ford AES, Smart SM, Henrys PA, and Ashmore MR. 2016. Using Qualitative and Quantitative Methods to Choose a Habitat Quality Metric for Air Pollution Policy Evaluation. PLOS ONE 11(8):e0161085 10.1371/journal.pone.0161085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sala OE, Stuart Chapin III F, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber- Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M.n., Poff NL, Sykes MT, Walker BH, Walker M, and Wall DH 2000. Global biodiversity scenarios for the year 2100. Science 287(5459):1770–1774 10.1126/science.287.5459.1770. [DOI] [PubMed] [Google Scholar]
- Schlesinger WH 1997. Biogeochemistry : An Analysis of Global Change Academic Press, San Diego. [Google Scholar]
- Schwede DB and Lear GG. 2014. A novel hybrid approach for estimating total deposition in the United States. Atmos. Environ 92:207–220 10.1016/j.atmosenv.2014.04.008. [DOI] [Google Scholar]
- Shannon JD 1998. Calculations of trends from 1900 through 1990 for sulfur and NOx-N deposition concentrations of sulfate and nitrate in precipitation, and atmospheric concentrations of SOx and NOx species over the southern Appalachians. Report to SAMI. [Google Scholar]
- Simkin SM, Allen EB, Bowman WD, Clark CM, Belnap J, Brooks ML, Cade BS, Collins SL, Geiser LH, Gilliam FS, Jovan SE, Pardo LH, Schulz BK, Stevens CJ, Suding KN, Throop HL, and Waller DM. 2016. Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States. Proceedings of the National Academy of Science 113(15):4086–4091 10.1073/pnas.1515241113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson EH 1949. Measurement of diversity. Nature 163:688 10.1038/163688a0. [DOI] [Google Scholar]
- Suddick EC, Whitney P, Townsend AR, and Davidson EA. 2013. The role of nitrogen in climate change and the impacts of nitrogen–climate interactions in the United States: foreword to thematic issue. Biogeochemistry 114(1):1–10 10.1007/s10533-012-9795-z. [DOI] [Google Scholar]
- Suding KN, Collins SL, Gough L, Clark C, Cleland EE, Gross KL, Milchunas DG, and Pennings S. 2005. Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proc. Nat. Acad. Sci 102(12):4387–4392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sullivan TJ 2017. Air Pollution and Its Impacts on U.S. National Parks CRC Press, Boca Raton, FL. 638 pp. [Google Scholar]
- Tominaga K, Aherne J, Watmough SA, Alveteg M, Cosby BJ, Driscoll CT, Posch M, and Pourmokhtarian A. 2010. Predicting acidification recovery at the Hubbard Brook Experimental Forest, New Hampshire: evaluation of four models. Environ. Sci. Technol 44(23):9003–9009 [DOI] [PubMed] [Google Scholar]
- U.S. Environmental Protection Agency. 2009. Risk and Exposure Assessment for Review of the Secondary National Ambient Air Quality Standards for Oxides of Nitrogen and Oxides of Sulfur: Final. EPA-452/R-09–008a. Office of Air Quality Planning and Standards, Health and Environmental Impacts Division, Research Triangle Park, NC. [Google Scholar]
- U.S. Environmental Protection Agency. 2014. Technical Support Document (TSD) Preparation of Emissions Inventories for the Version 6.1, 2011 Emissions Modeling Platform. U.S. EPA, Office of Air and Radiation Office of Air Quality Planning and Standards Air Quality Assessment Division Available online at: https://www.epa.gov/air-emissions-modeling/2011-version-61-technical-support-document, Research Triangle Park, NC. [Google Scholar]
- Vitousek PM, Aber JD, Howarth RW, Likens GE, Matson PA, Schindler DW, Schlesinger WH, and Tilman DG. 1997. Human alteration of the global nitrogen cycle: sources and consequences. Ecol. Appl 7(3):737–750 [Google Scholar]
- Volpe Horii C, William Munger J, Wofsy SC, Zahniser M, Nelson D, and Barry McManus J. 2005. Atmospheric reactive nitrogen concentration and flux budgets at a Northeastern U.S. forest site. Agri. For. Meteorol 133(1–4):210–225 10.1016/j.agrformet.2004.08.009. [DOI] [Google Scholar]
- Walter CA, Raiff DT, Burnham MB, Gilliam FS, Adams MB, and Peterjohn WT. 2016. Nitrogen fertilization interacts with light to increase Rubus spp. cover in a temperate forest. Plant Ecol. 217(4):421–430 10.1007/s11258-016-0583-z. [DOI] [Google Scholar]
- Wamelink GWW, Goedhart PW, Malinowska AH, Frissel JY, Wegman RJM, Slim PA, and van Dobben HF. 2011. Ecological ranges for the pH and NO3 of syntaxa: a new basis for the estimation of critical loads for acid and nitrogen deposition. J. Veg. Sci 22(4):741–749 10.1111/j.1654-1103.2011.01286.x. [DOI] [Google Scholar]
- Wason JW, Dovciak M, Beier CM, and Battles JJ. 2017. Tree growth is more sensitive than species distributions to recent changes in climate and acidic deposition in the northeastern United States. J. Appl. Ecol.:n/a-n/a 10.1111/1365-2664.12899. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.