

Abstract
Behavioral studies suggest that immersion in nature improves affect and executive attention. However, the neural mechanisms underlying these benefits remain unclear. This randomized controlled trial (N = 92) explored differences in self-reported affect and in frontal midline theta (FMθ), a neural oscillation linked to executive attention, between a 40-min, low-intensity nature walk and an urban walk of comparable time and distance—controlling for ambient temperature, humidity, elevation change, walking pace, heart rate, calories burned, and moving time between the two groups. While affect improved for both groups, the nature walkers showed a significantly greater boost in positive affect than the urban walkers. Electroencephalography (EEG) data revealed significantly greater FMθ activity following the urban walk compared to the nature walk, suggesting that the urban walk placed higher demands on executive attention. In contrast, the nature walk allowed executive attention to rest, as indicated by the lower FMθ activity observed after the walk. This study suggests that changes in FMθ may be a potential neural mechanism underlying the attentional strain of urban environments in contrast to the attentional rest in nature.
Keywords: Environmental neuroscience, Nature, Attention restoration theory, Executive control, Frontal midline theta
Subject terms: Attention, Psychology and behaviour, Human behaviour
Introduction
Our brains are products of the environments in which they develop and exist. Many of our sensory, motor, and cognitive processes develop over time through interaction with our physical environment. In fact, some argue that our surroundings have such a profound effect on our brains and behavior that we cannot study cognition without considering our physical environment1. This idea forms the basis of the emerging field of environmental neuroscience, which bridges the gap between environmental psychology and cognitive neuroscience to explore the interaction between physical environments, the brain, and behavior—with the goal of understanding how our day-to-day environments can be enriched to improve both psychological and physiological well-being.
More than 55% of people reside in urban areas worldwide, with this number growing steadily and expected to reach 70% by 20502. In the United States, 80% of the population currently resides in urban areas3. While urban living often offers greater access to education, healthcare, and other cultures, many qualities of these environments negatively impact health and cognition. For example, population-based cohort studies have linked qualities of urban environments (such as low greenspace and high air pollution) to increased rates of depression4, anxiety5, and attention disorders6–8. Furthermore, chronic health conditions like cardiovascular disease, high blood pressure, and obesity are disproportionately common in cities9. Thus, researchers from multiple disciplines have allocated considerable effort to exploring the potential benefits of exposure to nature to combat some of the detriments of our day-to-day urban environments and improve physical and mental health.
Behavioral studies have revealed that exposure to nature confers cognitive benefits, including but not limited to enhancing executive attention10,11, mood12, and creativity13. Furthermore, immersion in nature has been linked to physical health benefits by encouraging exercise14, improving immune function15, aiding in recovery from stress16,17, and lowering blood pressure18. Beyond these documented cognitive and physical health benefits, exposure to nature serves as a communal area where people gather, fostering social interactions and a sense of community coupled with a reduced sense of loneliness19. These findings hold promise for a low-cost lifestyle intervention to improve human health and well-being; however, as this health intervention gains popular interest and traction, there is an ever-growing need to understand the biological mechanisms underlying these benefits20.
The current study leverages methods from cognitive neuroscience to understand the influence of immersion in nature on the brain, particularly as it relates to the documented attentional benefits. This work is grounded in Attention Restoration Theory (ART)21, which posits that our modern, urban environments strain our attention by bombarding us with many competing stimuli at once, forcing us to select task-relevant information and ignore task-irrelevant information. This type of flexible, effortful control is highly demanding and, over time, can lead to mental fatigue and impairments in control processes such as self-regulation and executive functioning22. Natural environments, ART proposes, engage our attention in a less demanding way. Rather than the constant task-switching and information filtering that is required in urban environments, natural environments are engaging to look at, but in a way that does not place much demand on executive attention resources. This provides relief for our executive attention, allowing it to rest and be replenished, leading to a subsequent improvement in executive attention capacity.
These theoretical claims have been validated by behavioral evidence indicating that performance on executive attention tasks improves after exposure to nature10,11. The reasoning is that executive attention networks in the brain, having had the opportunity to rest during nature exposure, become more effective when re-engaged for subsequent cognitive tasks, resulting in enhanced task performance compared to before the nature exposure or compared to exposure to a control, urban environment. Therefore, in most prior work, participants complete computer-administered cognitive tasks such as the Flanker Task23 or the Attention Network Task24 before and after exposure to nature to quantify changes in executive attention performance in a standardized and highly controlled manner. While this approach has the strength of linking behavioral performance to underlying neural metrics, it also introduces a theoretical conundrum such that completing an artificial cognitive task may disrupt the process of cognitive restoration that we seek to understand in the first place. In other words, there is a level of artificiality to this approach that confounds the brain’s spontaneous response to immersion in nature. The current study overcomes this limitation by measuring a neural signature of executive attention at rest to better understand the biological mechanisms that underlie restoration without the confound of task-related interference. In doing so, we aim to minimize external influences and improve the ecological validity of the findings, making results more applicable to real-world experiences and practical applications for cognitive health.
In a single-blind, randomized controlled trial, we record resting brain activity using electroencephalography (EEG) before and after a 40-min nature walk and a control, urban walk of comparable time, distance, and elevation change. The EEG signal is comprised of oscillations that reflect the synchronized activity of large groups of neurons in the brain. Using various signal-processing techniques, these oscillations can be separated into different frequency bands (e.g., delta [2–4 Hz], theta [4–8 Hz], alpha [8–12 Hz], beta [15–30 Hz], gamma [30–150 Hz]) that are each involved in different cognitive processes25. We specifically focus on power in the frontal midline theta (FMθ) frequency band, which refers to theta oscillations (4–8 Hz) recorded from scalp electrodes placed over the frontal cortex. Importantly, FMθ oscillations are thought to reflect neural activity generated from the dorsal anterior cingulate cortex (dACC)26—a region of the brain highly implicated in executive attention. The dACC interacts with the prefrontal cortex to regulate attention and prioritize goal-relevant information. It is involved in allocating attentional resources based on task demands and goals, especially in situations where attention needs to be focused amidst competing stimuli or distractions. FMθ has been extensively linked to attention and effort such that FMθ increases with increases in task difficulty and working memory load27. Overall, FMθ serves as a neural signature of the dynamic interplay of the dACC and associated brain networks in attention allocation procedures, providing insight into the underlying neural mechanisms of executive attention.
In line with Attention Restoration Theory and the mounting behavioral evidence in support of it, we hypothesized that immersion in nature would place less demand on executive attention, resulting in lower FMθ associated with the 40-min nature walk compared to the urban walk. We expected this decrease in FMθ to be complemented by positive changes in affect associated with the nature walk—one of the most reliable findings in the nature and cognition literature12.
Methods and materials
This research complied with the APA Code of Ethics and was approved by the University of Utah Institutional Review Board. Informed consent was obtained from each participant and the reported methods were performed in accordance with relevant guidelines and regulations of this institution.
Participants
Participants (N = 92; 71 female, 20 male, 1 gender non-binary; 80% White, 10% Asian, 7% Latinx/Hispanic, 3% Black) between the ages of 18 and 57 (M = 29.43, SD = 10.52) were recruited from the University of Utah Department of Psychology research participant pool (23% of sample) as well as from the greater Salt Lake City community (77% of sample). While university-affiliated participants were considered a sample of convenience, we recruited from the broader community to increase the generalizability of the results. Participants were informed that they would have their brain activity recorded before and after a 40-min, low-intensity walk, but they were not informed that there were two walking conditions at any point during recruitment or participation. University participants received course credits for their participation and community participants received $50 USD. All participants reported normal or corrected-to-normal vision, getting at least 6 h of sleep the night before, having no known neurological conditions, and being physically capable of walking for 40-min at a comfortable pace (~ 20 min/mile).
The sample size for this study was determined by an a priori power analysis conducted in PANGEA v0.228. The analysis indicated that having 43 participants in each condition (for a total of 86 participants) would provide sufficient statistical power at the recommended level of 0.80 to detect within-subject by between-group interactions with a medium effect size (Cohen’s d = 0.50). This effect size was chosen based on prior literature that has identified medium-sized effects of nature exposure on neural activity23,24. To account for potential data loss, we collected data from an overestimate of 92 participants.
One participant’s nature walk was disrupted by rain, ending their testing session early and rendering their post-walk affect score and EEG recording unavailable for analysis. Another participant’s EEG data exhibited poor data quality suggesting that either the reference or ground electrode was not properly secured on the scalp, rendering their EEG data unusable. Therefore, the final affect analyses were run on 92 baseline measurements (46 nature, 46 urban), 92 pre-walk measurements (46 nature, 46 urban), and 91 post-walk measurements (45 nature, 46 urban). The final EEG analyses were run on 91 baseline measurements (45 nature, 46 urban), 91 pre-walk measurements (45 nature, 46 urban), and 90 post-walk measurements (44 nature, 46 urban).
Positive and negative affect schedule
The 20-item Positive and Negative Affect Schedule (PANAS)29 assessed momentary (state) affect along positive and negative dimensions and was administered via Qualtrics on an iPad. Participants were presented with 20 words (e.g., “excited”, “distressed”, “scared”, “strong”) and instructed to rate how much they felt that word in the present moment on a scale of 1–5 (1 being “very slightly or not at all” and 5 being “extremely”). A total score for self-reported negative affect was calculated by adding the scores for the ten negatively valanced items, and a total score for positive affect was calculated by adding the scores for the ten positively valanced items.
Depletion task
A counting backwards task used to induce high cognitive load30–33 was administered as a cognitive depletion manipulation prior to participants’ walk. This was intended to experimentally recreate the state of mental fatigue from which individuals may seek restoration in nature21. This task has been utilized as a depletion task in the nature and cognition literature before32,34. For this task, participants were instructed to count backward from 1000 to 0 by 7’s for 10 min. Participants counted out loud (as opposed to in their head) so that the researcher could ensure they were doing the task.
Manipulation check questions related to fatigue, effort, pleasantness, and frustration were administered via Qualtrics on an iPad immediately following the depletion task. These manipulation check questions were adopted from Bray and colleagues35 and consisted of four, 7-point Likert-scale questions: “how tired do you feel after completing this task?”, “how much effort did you exert while performing this task?”, “how pleasant did you find performing this task?”, and “how frustrated did you feel while performing this task?”. These questions ensured that participants were, in fact, depleted by the task and that reported levels of depletion were comparable across the nature and urban conditions. As expected, the two conditions did not significantly differ on any of the four questions (tiredness, t(90) = − 0.39, p = 0.700, 95% CI [− 0.80, 0.54]; effort, t(90) = 0.57, p = 0.571, 95% CI [− 0.38, 0.68]; pleasantness, t(90) = 1.43, p = 0.157, 95% CI [− 0.17, 1.04]; frustration, t(90) = − 0.32, p = 0.750, 95% CI [− 0.94, 0.68]).
Procedure
Procedural steps of the study can be visualized in Fig. 1. Upon arrival to the laboratory, participants completed a demographic questionnaire while the EEG cap and electrodes were set up. Participants then completed the PANAS to capture baseline levels of affect, followed by a 2-min baseline resting EEG recording. Participants then completed the 10-min counting backwards depletion task followed by the four manipulation check questions. After the depletion manipulation, participants completed a second 2-min resting EEG recording and PANAS survey. We then randomly assigned participants to walk for 40-min in either a natural environment (46 participants) or an urban environment (46 participants) of comparable time, distance, and elevation change. After the walk, participants returned to the lab and completed the PANAS for a third time followed by their third and final 2-min resting EEG recording. The nature walk took place in Red Butte Garden and Arboretum at the University of Utah and the urban walk took place on the adjacent medical campus. Both walking routes departed from the EEG recording laboratory (located in the Kay and Zeke Dumke, Jr. Horticultural Operations Center at the University of Utah), but in opposite directions. On their walk, participants wore a Garmin Forerunner 45S watch for GPS, time, pace, elevation, and distance tracking as well as a GoPro HERO 7 video camera to verify that they followed the correct route.
Fig. 1.

Study procedure.
Walk and environmental characteristics
Both groups completed a ~ 40-min, low-intensity walk, covering approximately two miles. Participants maintained an average pace of ~ 20 min per mile (see Table 1), which is consistent with the classification of low-intensity physical activity according to the American College for Sports Medicine’s Guidelines for Exercise Testing and Prescription36 and existing literature37. To further ensure the two walk conditions were matched on exercise-related metrics and various environmental characteristics, the Garmin Forerunner 45S watch tracked exercise metrics and researchers recorded the temperature, wind speed, and humidity while participants were on their walks. Descriptive statistics for each metric can be seen in Table 1.
Table 1.
Descriptive statistics (mean and standard error) of exercise metrics generated from the Garmin GPS watch and environmental characteristics as a function of walk condition.
| Nature | Urban | |||
|---|---|---|---|---|
| Mean | SE | Mean | SE | |
| Distance (miles) | 1.93 | 0.02 | 1.99 | 0.01 |
| Moving time (min:sec) | 42:19 | 00:36 | 39:10 | 00:20 |
| Pace (min:sec/mile) | 23:25 | 00:20 | 20:00 | 00:11 |
| Avg. Heart rate | 108.24 | 2.40 | 110.41 | 2.22 |
| Max. Heart rate | 131.71 | 2.60 | 137.26 | 2.72 |
| Calories burned | 217.27 | 6.75 | 199.85 | 6.01 |
| Total ascent (ft) | 146.00 | 7.44 | 123.37 | 11.31 |
| Total descent (ft) | 138.62 | 7.84 | 132.52 | 8.75 |
| Temperature (°F) | 74.36 | 1.92 | 72.28 | 1.86 |
| Humidity (%) | 29.84 | 1.85 | 30.52 | 1.72 |
| Wind speed (mph) | 8.31 | 0.80 | 8.20 | 0.63 |
EEG recording and processing
EEG data were collected via a 32-channel, active electrode actiCap manufactured by BrainVision (BrainVision Systems, Morrisville, NC, USA). Channel locations were consistent with the International 10–20 system38 and impedances were kept below 25 kOhms. The signal was amplified with the BrainVision actiCHamp Plus amplifier with an online sampling rate of 500 Hz and the signal was acquired by BrainVision Recorder (Version 1.20.0601). The right mastoid electrode (TP10) served as the online reference. Vertical electrooculographic (VEOG) activity was recorded with two auxiliary electrodes manufactured by BrainVision, placed above and below the right eye, in line with the middle of the pupil, to record participant blinks and eye movements for later data processing.
During each resting EEG recording, participants were instructed to simply sit still and keep their eyes open for 2-min. A black fixation cross on a grey background remained in the middle of the screen for the duration of each recording. This fixation cross served to remind the participant to keep their eyes open and facing forward for the duration of the 2-min. EEG was also recorded during the counting backwards depletion task to serve as a manipulation check that FMθ power is positively correlated with increases in cognitive workload. During this recording, participants were instructed to count out loud so that the researcher could ensure they were doing the task correctly, but to do so without excessive face or jaw movement so as to not introduce extreme muscle artifacts into the EEG recording.
EEG data were processed offline in MATLAB using the EEGLab39 and ERPLab40 toolboxes. Data were downsampled to 250 Hz and band pass filtered from 0.1 to 30 Hz with a Butterworth filter and a roll-off of 12 dB/octave. Data were re-referenced offline to the average of the left (TP9) and right (TP10) mastoid electrodes. Artifacts created by eye movements and blinks were corrected using Gratton et al.'s41 eye movement correction procedure (EMCP)42,43. Subsequent artifact rejection was performed using a moving window to reject sections of data containing flatlines or peak-to-peak activity greater than 200 μV40. On average, 4.76% of data was lost after this artifact correction and rejection procedure: 2.08% at baseline rest recording, 12.40% during the depletion task, 2.19% at the pre-walk rest recording, and 2.37% at post-walk rest recording.
Artifact-free, continuous data from each recording were epoched into 1-s intervals using a Hanning window. We used a Fast Fourier Transform25 to convert the artifact-free data from the time domain to the frequency domain, and the average power at each frequency from 1 to 30 Hz was extracted for each EEG recording. The average power from 4 to 8 Hz at the series of frontocentral electrodes (Fp1, F3, Fz, F4, FC1, FC2, C3, Cz, C4, CP1, CP2, Fp2) was used to quantify FMθ power25.
Statistical analyses
Analyses were conducted in R version 4.0.244. Descriptive statistics for all outcomes were extracted using the describeBy( ) function in the psych package. Data were analyzed with linear mixed effects models45 to control for non-independence in the data (i.e., repeated measures within each participant) and any potential missingness in the data (i.e., if a participant was missing data from any time point). For each dependent measure (positive affect, negative affect, and FMθ), we ran three models with Participant ID as a random intercept. These three models tested a fixed effect of condition (nature, urban), a fixed effect of time (baseline, pre-walk, post-walk), and the interaction between the two to test whether any changes over time significantly differed between the two walking conditions. Statistical inference on fixed effects was conducted using separate likelihood ratio tests for each parameter, computed using the mixed( ) function from the afex package46. For follow-up tests, we calculated pairwise contrasts on the estimated marginal means (also known as least-squares means) using the pairs( ) function from the emmeans packages47 with a Tukey adjustment to account for multiple comparisons. Scalp map visualizations were created in Python using the plot_topomap( ) function in the mne software48.
Results
Self-reported affect
Descriptive statistics from the PANAS as a function of time (baseline, pre-walk, and post-walk) and condition (nature and urban) are presented in Table 2 and visualized in Fig. 2. Linear mixed effects models were run to test the main effect of time, main effect of condition, and the interaction between the two. In terms of positive affect, there was a significant main effect of time (χ2(2) = 122.95, p < 0.001) but no main effect of condition (χ2(1) = 2.06, p = 0.152). These effects were qualified by a significant interaction between time and condition (χ2(2) = 9.11, p < 0.05). We explored this interaction by calculating contrasts between the estimated marginal means. The pairwise comparison results can be seen in the top panel of Table 3. These post-hoc contrasts revealed that both the nature and urban condition showed a decrease in positive affect from baseline to pre-walk followed by an increase in positive affect from pre-walk to post-walk. However, the increase in positive affect after the walk was significantly greater for the nature condition compared to the urban condition.
Table 2.
Descriptive statistics. Positive affect, negative affect, and FMθ descriptive statistics (mean and standard error) as a function of time and condition.
| Nature | Urban | |
|---|---|---|
| Positive affect | ||
| Baseline | 31.39 (1.20) | 30.39 (1.11) |
| Pre-walk | 24.26 (1.26) | 23.07 (1.12) |
| Post-walk | 32.98 (1.36) | 28.80 (1.24) |
| Negative affect | ||
| Baseline | 11.83 (0.35) | 12.43 (0.36) |
| Pre-walk | 12.35 (0.49) | 13.26 (0.67) |
| Post-walk | 10.78 (0.21) | 11.30 (0.27) |
| FMθ power (μV2/Hz) | ||
| Baseline | 1.96 (0.06) | 2.07 (0.06) |
| Depletion | 3.41 (0.16) | 3.58 (0.15) |
| Pre-walk | 2.41 (0.12) | 2.34 (0.12) |
| Post-walk | 2.26 (0.09) | 2.99 (0.13) |
Fig. 2.

PANAS results. Average positive (left) and negative (right) affect scores as a function time and condition. Error bars represent one standard error of the mean. Although each subscale of the PANAS has a possible score range from 10 to 50, the figure is only displayed up to a score of 35 to focus on the range where the average scores are observed, making the differences between groups more visible and easier to interpret.
Table 3.
Pairwise post-hoc contrasts for the three univariate models. The top panel reports the positive affect results generated from the PANAS. The middle panel reports the negative affect results generated from the PANAS. The bottom panel reports the FMθ results.
| Contrast | Dif. Est | df | t | p | 95% CI |
|---|---|---|---|---|---|
| Positive affect model post-hoc contrasts | |||||
| Nature: baseline—pre-walk | 7.13 | 179 | 7.86 | < 0.001*** | [4.99, 9.28] |
| Urban: baseline—pre-walk | 7.33 | 179 | 8.07 | < 0.001*** | [5.18, 9.47] |
| Nature: baseline—post-walk | − 1.89 | 179 | − 2.06 | 0.100 | [− 4.05, 0.27] |
| Urban: baseline—post-walk | 1.59 | 179 | 1.75 | 0.190 | [− 0.56, 3.73] |
| Nature: pre-walk—post-walk | − 9.02 | 179 | − 9.86 | < 0.001*** | [− 11.18, − 6.86] |
| Urban: pre-walk—post-walk | − 5.74 | 179 | − 6.32 | < 0.001*** | [− 7.88, − 3.59] |
| Baseline: nature—urban | 1.00 | 132 | 0.58 | 0.563 | [− 2.41, 4.41] |
| Pre-walk: nature—urban | 1.20 | 132 | 0.69 | 0.490 | [− 2.22, 4.61] |
| Post-walk: nature—urban | 4.48 | 133 | 2.59 | < 0.05* | [1.05, 7.90] |
| Negative affect model post-hoc contrasts | |||||
| Baseline—Pre-walk | − 0.67 | 181 | − 1.94 | 0.131 | [− 1.50, 0.15] |
| Baseline—Post-walk | 1.10 | 181 | 3.14 | < 0.01** | [0.27, 1.92] |
| Pre-walk—Post-walk | 1.77 | 181 | 5.07 | < 0.001*** | [0.95, 2.60] |
| Contrast | Dif. Est | df | z | p | 95% CI |
|---|---|---|---|---|---|
| FMθ model post-hoc contrasts | |||||
| Nature: baseline—depletion | − 1.45 | Inf | − 10.30 | < 0.001*** | [− 1.82, − 1.09] |
| Urban: baseline—depletion | − 1.51 | Inf | − 10.78 | < 0.001*** | [− 1.86, − 1.15] |
| Nature: baseline—pre-walk | − 0.45 | Inf | − 3.17 | < 0.01** | [− 0.81, − 0.08] |
| Urban: baseline—pre-walk | − 0.27 | Inf | − 1.94 | 0.212 | [− 0.63, 0.09] |
| Nature: baseline—post-walk | − 0.28 | Inf | − 1.97 | 0.201 | [− 0.65, 0.09] |
| Urban: baseline—post-walk | − 0.92 | Inf | − 6.59 | < 0.001*** | [− 1.28, − 0.56] |
| Nature: depletion—pre-walk | 1.01 | Inf | 7.13 | < 0.001*** | [0.64, 1.37] |
| Urban: depletion—pre-walk | 1.23 | Inf | 8.85 | < 0.001*** | [0.88, 1.59] |
| Nature: depletion—post-walk | 1.17 | Inf | 8.26 | < 0.001*** | [0.81, 1.54] |
| Urban: depletion—post-walk | 0.59 | Inf | 4.19 | < 0.001*** | [0.23, 0.94] |
| Nature: pre-walk—post-walk | 0.17 | Inf | 1.18 | 0.643 | [− 0.20, 0.53] |
| Urban: pre-walk—post-walk | − 0.65 | Inf | − 4.65 | < 0.001*** | [− 1.01, − 0.29] |
| Baseline: nature—urban | − 0.11 | Inf | − 0.36 | 0.717 | [− 0.73, 0.50] |
| Depletion: nature—urban | − 0.17 | Inf | − 0.53 | 0.597 | [− 0.78, 0.45] |
| Pre-walk: nature—urban | 0.06 | Inf | 0.20 | 0.841 | [− 0.55, 0.68] |
| Post-walk: nature—urban | − 0.75 | Inf | − 2.41 | < 0.05* | [− 1.37, − 0.14] |
*p < 0.05, **p < 0.01, ***p < 0.001. Significant values are in bold. The transition from t-values to z-values in the pairwise comparisons for the FMθ model indicates that the sample size is sufficiently large for the Central Limit Theorem to apply (achieved by estimating values across a series of frontocentral electrodes in the FMθ analysis), allowing the use of normal approximations. In this context, the degrees of freedom reported as Inf signifies that the sample size is large enough to produce stable estimates, justifying the asymptotic approach. Thus, the z-values reflect a more precise estimation of the differences between conditions under these circumstances. For more information, see Lenth et al.47.
In terms of negative affect, there was a significant main effect of time (χ2(2) = 24.77, p < 0.001) but no main effect of condition (χ2(1) = 2.51, p = 0.113) nor a significant interaction between the two (χ2(2) = 0.32, p = 0.850). The pairwise comparison results can be seen in the middle panel of Table 3. Post-hoc contrasts revealed that there was no significant change in negative affect from baseline to pre-walk, but there was a significant decrease in negative affect from pre-walk to post-walk, and post-walk negative affect was even significantly lower than baseline. This pattern held for both the nature condition and the urban condition.
Frontal midline theta power
FMθ descriptive statistics as a function of time (baseline, depletion, pre-walk, and post-walk) and condition (nature and urban) are reported in Table 2 and visualized in Fig. 3. EEG scalp maps as a function of time and condition are presented in Fig. 4.
Fig. 3.

FMθ results. FMθ power (μV2/Hz) as a function of time and condition. Error bars represent one standard error of the mean.
Fig. 4.
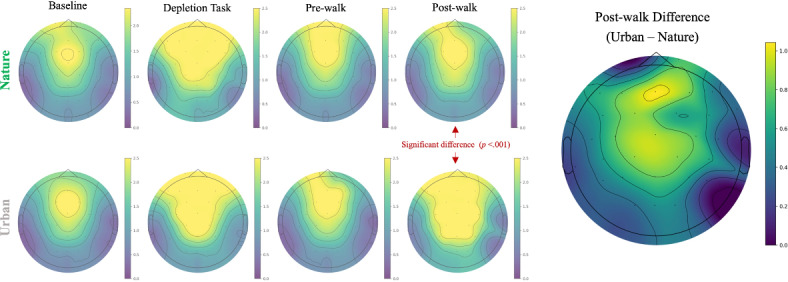
EEG scalp topography maps. Left: Theta power (4–8 Hz; μV) distribution over the scalp as a function of time and condition. Right: Difference scalp topography map (urban–nature; μV) between the two conditions at post-walk, the time point in which the two conditions showed a significant difference in FMθ power.
Linear mixed effects models were run to test the main effect of time, main effect of condition, and the interaction between the two. There was a significant main effect of time on FMθ power (χ2(3) = 235.52, p < 0.001). Follow-up comparisons exploring this main effect of time revealed that FMθ significantly increased from baseline to during the depletion task (Baseline—Depletion: difference estimate = − 1.48, z = − 14.88, p < 0.001), and significantly decreased from the depletion task to pre-walk (Depletion—Pre-walk: difference estimate = 1.12, z = 11.28, p < .001). This validates the point that FMθ increases during engagement of executive attention resources (i.e., during the depletion task). Importantly, FMθ remained significantly elevated above baseline at the pre-walk recording (Baseline—Pre-walk: difference estimate = -0.36, z = -3.60, p < .01), substantiating two key points: 1) that participants entered their walk in a state of mental fatigue (i.e., elevated FMθ power at pre-walk relative to baseline), validating the intended effect of the depletion manipulation. Secondly, the sustained elevation of resting FMθ above baseline after the depletion task underscores the enduring, or residual, impact of attentionally-demanding activities—even after the brain returns to rest. There was no main effect of condition on FMθ power (χ2(1) = 0.72, p = 0.396). These effects were qualified by a significant interaction between time and condition (χ2(3) = 18.94, p < 0.001). We explored these effects by calculating contrasts between the estimated marginal means. The pairwise comparison results can be seen in the bottom panel of Table 3. Post-hoc contrasts revealed that both the nature and urban conditions displayed a significant increase in FMθ power from baseline to during the depletion task, followed by a significant decrease in FMθ power after the depletion task (at the pre-walk rest recording). Importantly, the urban condition displayed a significant increase in FMθ power from pre-walk to post-walk while the nature condition did not.
Exploratory analyses
To better understand the relationship between changes in affect and FMθ in nature, we ran Pearson’s r correlations between the change in positive and negative affect (computed as the difference between pre-walk and post-walk scores) and the change in FMθ power (computed as the difference between pre-walk and post-walk FMθ). There was no significant correlation between change in positive affect and change in FMθ power (r(88) = − 0.05, p = 0.662) nor between change in negative affect and change in FMθ power (r(88) = 0.07, p = 0.535). This suggests that nature’s influence on affect and attention arise from independent mechanisms, replicating the findings of previous work49,50.
We also explored a series of covariates that may have played a role in one’s experience on their walk, including participant age, ambient temperature, and exact distance walked. We did so by adding each of these continuous variables as fixed effects (one at a time) into each univariate model, allowing us to test whether the changes in positive affect, negative affect, and FMθ power persist even after statistically controlling for each of these variables. After controlling for participant age, the significant main effect of time on positive affect (χ2(2) = 123.86, p < 0.001) and the significant interaction between time and condition on positive affect (χ2(2) = 9.44, p < 0.01) persisted. Additionally, the significant main effect of time on negative affect (χ2(2) = 25.03, p < 0.001) persisted and the lack of significant interaction between time and condition on negative affect (χ2(2) = 0.30, p = 0.859) remained. Lastly, the significant main effect of time (χ2(3) = 235.53, p < 0.001) and the significant interaction between time and condition on FMθ (χ2(3) = 19.02, p < 0.001) also persisted. This suggests that nature influences affect and FMθ above and beyond the role that participant age may play.
After controlling for ambient temperature, the significant main effect of time on positive affect (χ2(2) = 123.15, p < 0.001) and the significant interaction between time and condition on positive affect (χ2(2) = 9.39, p < 0.01) persisted. Additionally, the significant main effect of time on negative affect (χ2(2) = 25.02, p < 0.001) persisted and the lack of significant interaction between time and condition on negative affect (χ2(2) = 0.26, p = 0.878) remained. Lastly, the significant main effect of time (χ2(3) = 234.35, p < 0.001) and the significant interaction between time and condition on FMθ (χ2(3) = 18.69, p < 0.001) also persisted. This suggests that nature influences affect and FMθ above and beyond the role that ambient temperature may play.
After controlling for distance walked, the significant main effect of time on positive affect (χ2(2) = 123.15, p < 0.001) and the significant interaction between time and condition on positive affect (χ2(2) = 9.39, p < 0.01) persisted. Additionally, the significant main effect of time on negative affect (χ2(2) = 25.02, p < 0.001) persisted and the lack of significant interaction between time and condition on negative affect (χ2(2) = 0.26, p = 0.878) remained. Lastly, the significant main effect of time (χ2(3) = 234.35, p < 0.001) and the significant interaction between time and condition on FMθ (χ2(3) = 18.69, p < 0.001) also persisted. This suggests that nature influences affect and FMθ above and beyond the role that the distance walked may play.
Discussion
This randomized controlled trial explored the influence of immersion in nature on affect and resting oscillatory activity in the brain. Mounting evidence suggests that exposure to nature enhances executive attention, though the underlying neural mechanisms driving this effect remain unknown. Attention Restoration Theory (ART)21 proposes that qualities of natural environments engage executive attention in a less demanding way than urban environments, allowing our attentional resources to rest and recover, which in turn leads to improved attentional performance when attention is required again. In this study, we measured changes in self-reported affect and in a neural signature of executive attention (FMθ) associated with a 40-min walk in nature versus an urban walk of comparable time and distance. We hypothesized that participants who walked in nature would show greater improvements in affect and lower FMθ than participants who walked in the urban environment, indicative of less attentional demand placed on individuals walking in nature.
Participants self-reported their levels of positive and negative affect at three time points: baseline, before their walk (after the depletion task), and after their walk. Results confirm that while both conditions decreased in negative affect and increased in positive affect after their walk, the boost in positive affect was significantly greater for the nature walkers compared to the urban walkers. Exploratory analyses revealed that these effects persisted even after statistically controlling for participant age, ambient temperature, and distance walked. It is likely that both conditions showed mood improvements after the walk due to relief from mental depletion and to the 40-min of low-intensity exercise (for a review of the mood benefits associated with exercise see Chan et al.51). However, the nature walkers likely showed a greater boost in positive affect due to specific qualities of natural environments above and beyond simply exercise. For example, it is possible that the sights, scents, and sounds unique to nature may be more aesthetically pleasing than urban equivalents, and this appreciation for beauty may have enhanced positive affect to a greater degree for the nature walkers compared to the urban walkers. This would be supported by the environmental aesthetics literature, which proposes that the aesthetic qualities of nature evoke positive emotions such as awe, tranquility, and happiness52,53. It is also possible that the nature walk allowed for a greater sense of “being away” from the daily grind and routine of urban life more so than the urban walk did, thus exhibiting a stronger impact on positive affect. This would be supported by Attention Restoration Theory21, which underscores the necessity of disengaging from the demands of our daily routines—both mentally and physically—to improve mood and promote restoration.
The EEG findings confirm that resting FMθ power serves as a robust neural marker that differentiates between immersion in natural and urban environments. Participants underwent EEG recordings at four timepoints: baseline (resting EEG), during the depletion task (task-related EEG), before their walk (resting EEG), and after their walk (resting EEG). As expected, there was a significant increase in FMθ from baseline to during the depletion task (see Figs. 3 and 4), confirming that FMθ increases with the effortful engagement of executive attention27. Following the depletion task, FMθ levels declined while remaining elevated above baseline (see Figs. 3 and 4). This observation substantiates two key points: First, participants entered their walk in a state of mental fatigue (relative to baseline), validating the intended effect of the depletion manipulation. Second, the sustained elevation of resting FMθ above baseline after the depletion task underscores the enduring, or residual, impact of attentionally-demanding activities—even after the brain returns to rest. This is an important point that we return to in our interpretation of the post-walk FMθ results.
Consistent with our hypotheses, we observed a notable difference in post-walk FMθ power between conditions such that urban walkers exhibited significantly greater FMθ power after their walk than nature walkers did. We interpret these post-walk, resting FMθ recordings as indicative of lingering brain activity following the walk experience. In other words, immersion in the urban environment imposed greater demands on executive attention resources, as participants navigated through construction zones, pedestrian traffic, vehicular hazards, and other typical urban sensory inputs like artificial sights, sounds and scents21. The enduring elevation of resting FMθ recorded back in the laboratory following the urban walk may suggest residual effects of this cognitively demanding experience on this neural signature of executive attention. In contrast, the nature walk was devoid of such demanding, artificial stimuli; rather, participants walked on trails lined with green foliage, a trickling creek and waterfall, and a pond with fish and ducks21. The relative absence of executive attentional demands in nature would have afforded these cognitive resources a respite, reflected as lower FMθ levels recorded immediately following the walk. This conclusion is further supported by recent vision science work suggesting that processing the low-level features of nature scenes consumes fewer cognitive resources than processing urban scenes54. For example, nature scenes typically contain fewer strong edges and have a larger proportion of their spectral energy in high-frequencies compared to urban scenes, both of which demand less cognitive effort for the visual system to process55,56. Therefore, we conclude that the significantly lower FMθ following the nature walk compared to the urban walk reflects the attentional respite afforded in nature. While not tested in this study, we theorize that this attentional rest would subsequently facilitate enhanced executive attention capacity when tasked with coming back online via administration of a task, as shown in prior work24.
Importantly, these FMθ differences between environments cannot be attributed to exercise alone, as both groups walked the same distance, for the same amount of time, on routes with comparable elevation profiles. Furthermore, the groups were matched on exercise-related metrics such as average heart rate, calories burned, and pace as well as nuisance environmental variables such as temperature, humidity, and wind speed (see Table 1). In a series of exploratory analyses, we explored the role that participant age, ambient temperature, and distance walked may have played in driving post-walk differences in FMθ—three variables that may potentially influence a participant’s experience on their walk. The significant interaction between time and condition on FMθ persisted even when statistically controlling for each of these potential nuisance variables, strengthening our conclusion that there is something unique about the characteristics of nature that differentially influence neural oscillations related to executive attention.
These results hold promise for future research to explore spontaneous changes in brain activity in response to immersion in nature. As previously discussed, prior research exploring the cognitive impacts of nature exposure require participants to engage in artificial computer tasks during or after the exposure to be able to index changes in task-related attentional performance23,24,57,58. We believe that requiring participants to engage in cognitively demanding tasks during or immediately after immersion in nature is likely to alter or disrupt the process of restoration and compromises the face validity of the work. The current study advances the literature by identifying an emergent signature of attention in nature by measuring the brain at rest, allowing for the observation of neural activity in a more authentic context, free from the influence of artificial task demands or external stimuli.
Given the reliable link between FMθ and executive attention in the cognitive neuroscience literature at large27, it is very likely that the post-walk condition differences in FMθ are linked to differences in attentional demand between the two environments. This conclusion is supported by both Attention Restoration Theory21 and extensive behavioral evidence. However, it is possible that other unaccounted variables could be contributing to the changes in FMθ observed in this study. For example, condition differences in stress levels might play a role. Stress Recovery Theory59 complements Attention Restoration Theory by suggesting that natural environments promote recovery from stress. Although we did not measure or manipulate stress directly, it is possible that stress recovery occurs alongside attention restoration, a concept supported by a recent unified framework on nature-based restoration presented by Scott and colleagues60. While we carefully controlled for many factors, caution is still warranted when interpreting the findings, as additional influences on FMθ beyond varying attentional demand may exist.
Future directions
We explored theta oscillations in frontal areas of the brain based on both theory and prior evidence surrounding the attention-related benefits of exposure to nature. However, future work should take a similar, randomized controlled trial approach to explore changes in other neural oscillations that may be sensitive to environmental conditions, such as alpha power (~ 8–12 Hz)61 or beta power (~ 12–35 Hz)62. Furthermore, investigating the synchronization between certain oscillations—such as the interaction between theta and alpha oscillations, as demonstrated by Chen et al.63—could reveal changes in functional connectivity associated with immersion in different environments.
Future research should also aim to extend the applicability of these findings to more diverse populations. Although our recruitment efforts included both a university-affiliated sample and members of the broader community to enhance the generalizability of the results, the demographic composition of our sample remained predominantly White, educated, and middle-aged. This limitation highlights the need for studies that include participants from a wider range of racial, socioeconomic, and age groups to better understand how these effects may vary across different backgrounds, lived experiences, cognitive baselines, and relationships to nature. By broadening the demographic scope, we can ensure that the insights gained from this research are more inclusive and relevant to a wider segment of the population.
Future research could also expand on this study by utilizing recent advances in mobile, noninvasive neuroimaging technologies to more precisely capture oscillatory activity during real-time immersion in nature64, rather than relying solely on a pre-post design as we do in this study. This approach could be applied to participants while they sit or walk in natural versus urban environments, offering deeper insights into the neural correlates of environmental exposure. While EEG is particularly sensitive to muscle activity generated from movement, advancements in artifact correction procedures (e.g., independent components analysis65) make it increasingly possible to extract meaningful information in real-world contexts. This approach will increase the ecological validity of the field even further by allowing researchers to capture real-time neural responses in nature, thereby providing more valid and applicable insights into the brain in situ.
Conclusion
This randomized controlled trial (N = 92) explored changes in self-reported affect and in resting frontal midline theta (FMθ), a neural oscillation related to executive attention, associated with a 40-min walk in nature (N = 46) compared to a 40-min walk in an urban environment (N = 46). The two conditions were carefully matched on variables that may influence one’s experience on a walk, including but not limited to participant age, distance walked, walking pace, elevation change, participant heart rate, ambient temperature, and humidity. While both conditions showed improvements in affect, the nature condition showed a greater boost in positive affect than the urban condition. Additionally, the urban condition showed a significant increase in resting FMθ power from before their walk to after their walk, while the nature condition did not. This reflects the attentional strain of urban environments (higher FMθ power) in contrast to the attentional rest in nature (lower FMθ power). We conclude that FMθ serves as a robust neural marker that differentiates between the varying attentional demands present in natural and urban environments.
Acknowledgements
The authors would like to thank Red Butte Garden, located in Salt Lake City, Utah, for their hospitality and generosity in providing laboratory space for this study.
Author contributions
A.S.M. conceptualized the study, programmed the experiment, collected and analyzed the data, and wrote the original draft of the manuscript. D.L.S. aided with conceptualization, provided supervision, and edited the manuscript.
Data availability
The datasets generated during the current study are available from the corresponding author on reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Berman, M. G., Kardan, O., Kotabe, H. P., Nusbaum, H. C. & London, S. E. The promise of environmental neuroscience. Nat. Hum. Behav.3(5), 414–417 (2019). [DOI] [PubMed] [Google Scholar]
- 2.United Nations, Department of Economic and Social Affairs, Population Division. World urbanization prospects: The 2018 revision (Report No. ST/ESA/SER.A/420). https://population.un.org/wup/publications/Files/WUP2018-Report.pdf (2019).
- 3.United States Census Bureau. Nation’s urban and rural populations shift following 2020 census [Press release].https://www.census.gov/newsroom/press-releases/2022/urban-rural-populations.html (2022).
- 4.Rautio, N., Filatova, S., Lehtiniemi, H. & Miettunen, J. Living environment and its relationship to depressive mood: A systematic review. Int. J. Soc. Psychiatry64(1), 92–103 (2018). [DOI] [PubMed] [Google Scholar]
- 5.McKenzie, K., Murray, A. & Booth, T. Do urban environments increase the risk of anxiety, depression and psychosis? An epidemiological study. J. Affect. Disord.150(3), 1019–1024 (2013). [DOI] [PubMed] [Google Scholar]
- 6.Markevych, I. et al. Outdoor air pollution, greenspace, and incidence of ADHD: A semi-individual study. Sci. Total Environ.642, 1362–1368 (2018). [DOI] [PubMed] [Google Scholar]
- 7.Thygesen, M. et al. The association between residential green space in childhood and development of attention deficit hyperactivity disorder: A population-based cohort study. Environ. Health Perspect.128(12), 127011 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yuchi, W. et al. Neighborhood environmental exposures and incidence of attention deficit/hyperactivity disorder: a population-based cohort study. Environ. Int.161, 107120 (2022). [DOI] [PubMed] [Google Scholar]
- 9.Dye, C. Health and urban living. Science319(5864), 766–769 (2008). [DOI] [PubMed] [Google Scholar]
- 10.Ohly, H. et al. Attention restoration theory: A systematic review of the attention restoration potential of exposure to natural environments. J. Toxicol. Environ. Health Part B19(7), 305–343 (2016). [DOI] [PubMed] [Google Scholar]
- 11.Stevenson, M. P., Schilhab, T. & Bentsen, P. Attention restoration theory II: A systematic review to clarify attention processes affected by exposure to natural environments. J. Toxicol. Environ. Health Part B21(4), 227–268 (2018). [DOI] [PubMed] [Google Scholar]
- 12.McMahan, E. A. & Estes, D. The effect of contact with natural environments on positive and negative affect: A meta-analysis. J. Posit. Psychol.10(6), 507–519 (2015). [Google Scholar]
- 13.Atchley, R. A., Strayer, D. L. & Atchley, P. Creativity in the wild: Improving creative reasoning through immersion in natural settings. PloS One7(12), e51474 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Maddock, J. E. & Frumkin, H. Physical activity in natural settings: An opportunity for lifestyle medicine. Am. J. Lifestyle Med. 15598276241253211 (2024).
- 15.Andersen, L., Corazon, S. S. & Stigsdotter, U. K. Nature exposure and its effects on immune system functioning: A systematic review. Int. J. Environ. Res. Public Health18(4), 1416 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Park, B. J., Tsunetsugu, Y., Kasetani, T., Kagawa, T. & Miyazaki, Y. The physiological effects of Shinrin-yoku (taking in the forest atmosphere or forest bathing): Evidence from field experiments in 24 forests across Japan. Environ. Health Prev. Med.15, 18–26 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Yao, W., Zhang, X. & Gong, Q. The effect of exposure to the natural environment on stress reduction: A meta-analysis. Urban For. Urban Green.57, 126932 (2021). [Google Scholar]
- 18.Ideno, Y. et al. Blood pressure-lowering effect of Shinrin-yoku (Forest bathing): A systematic review and meta-analysis. BMC Complement. Altern. Med.17, 1–12 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sachs, A. L. et al. Connecting through nature: A systematic review of the effectiveness of nature-based social prescribing practices to combat loneliness. Landsc. Urban Plan.248, 105071 (2024). [Google Scholar]
- 20.Frumkin, H. et al. Nature contact and human health: A research agenda. Environ. Health Perspect.125(7), 075001 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kaplan, S. The restorative benefits of nature: Toward an integrative framework. J. Environ. Psychol.15(3), 169–182 (1995). [Google Scholar]
- 22.Kaplan, S. & Berman, M. G. Directed attention as a common resource for executive functioning and self-regulation. Perspect. Psychol. Sci.5(1), 43–57 (2010). [DOI] [PubMed] [Google Scholar]
- 23.LoTemplio, S. B. et al. Nature as a potential modulator of the error-related negativity: A registered report. Int. J. Psychophysiol.156, 49–59 (2020). [DOI] [PubMed] [Google Scholar]
- 24.McDonnell, A. S. & Strayer, D. L. Immersion in nature enhances neural indices of executive attention. Sci. Rep.14(1), 1845 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cohen, M. X. Analyzing Neural Time Series Data: Theory and Practice (MIT Press, 2014). [Google Scholar]
- 26.Gevins, A., Smith, M. E., McEvoy, L. & Yu, D. High-resolution EEG mapping of cortical activation related to working memory: Effects of task difficulty, type of processing, and practice. Cereb. Cortex7(4), 374–385 (1997). [DOI] [PubMed] [Google Scholar]
- 27.Cavanagh, J. F. & Frank, M. J. Frontal theta as a mechanism for cognitive control. Trends Cogn. Sci.18(8), 414–421 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Westfall, J. PANGEA: Power analysis for general ANOVA designs. http://jakewestfall.org/publications/pangea.pdf. (2015)
- 29.Watson, D., Clark, L. A. & Tellegen, A. Development and validation of brief measures of positive and negative affect: The PANAS scales. J. Personal. Soc. Psychol.54(6), 1063 (1988). [DOI] [PubMed] [Google Scholar]
- 30.Eysenck, M., Payne, S. & Derakshan, N. Trait anxiety, visuospatial processing, and working memory. Cogn. Emot.19(8), 1214–1228 (2005). [Google Scholar]
- 31.Pellecchia, G. L. Dual-task training reduces impact of cognitive task on postural sway. J. Mot. Behav.37(3), 239–246 (2005). [DOI] [PubMed] [Google Scholar]
- 32.Jiang, B., Schmillen, R. & Sullivan, W. C. How to waste a break: Using portable electronic devices substantially counteracts attention enhancement effects of green spaces. Environ. Behav.51(9–10), 1133–1160 (2018). [Google Scholar]
- 33.Strayer, D. L., Castro, S. C., Turrill, J. & Cooper, J. M. The persistence of distraction: The hidden costs of intermittent multitasking. J. Exp. Psychol. Appl.28(2), 262 (2022). [DOI] [PubMed] [Google Scholar]
- 34.Li, D. & Sullivan, W. C. Impact of views to school landscapes on recovery from stress and mental fatigue. Landsc. Urban Plan.148, 149–158 (2016). [Google Scholar]
- 35.Bray, S. R., Martin Ginis, K. A., Hicks, A. L. & Woodgate, J. Effects of self-regulatory strength depletion on muscular performance and EMG activation. Psychophysiology45(2), 337–343 (2008). [DOI] [PubMed] [Google Scholar]
- 36.American College for Sports Medicine. ACSM’s Guidelines for Exercise Testing and Prescription (Lippincott Williams & Wilkins, 2020). [Google Scholar]
- 37.Varma, V. R. et al. Low-intensity walking activity is associated with better health. J. Appl. Gerontol.33(7), 870–887 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jasper, H. H. The ten-twenty electrode system of the international federation. Electroencephalogr. Clin. Neurophysiol.10, 370–375 (1958). [PubMed] [Google Scholar]
- 39.Delorme, A. & Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods134(1), 9–21. 10.1016/j.jneumeth.2003.10.009 (2004). [DOI] [PubMed] [Google Scholar]
- 40.Lopez-Calderon, J. & Luck, S. J. ERPLAB: An open-source toolbox for the analysis of event-related potentials. Front. Hum. Neurosci.8, 1–14. 10.3389/fnhum.2014.00213 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Gratton, G., Coles, M. G. & Donchin, E. A new method for off-line removal of ocular artifact. Electroencephalogr. Clin. Neurophysiol.55(4), 468–484. 10.1016/0013-4694(83)90135-9 (1983). [DOI] [PubMed] [Google Scholar]
- 42.Clements, G. M. et al. Dynamics of alpha suppression and enhancement may be related to resource competition in cross-modal cortical regions. NeuroImage252, 119048 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gyurkovics, M., Clements, G. M., Low, K. A., Fabiani, M. & Gratton, G. Stimulus-induced changes in 1/f-like background activity in EEG. J. Neurosci.42(37), 7144–7151 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.R Core Team. R: A language and environment for statistical computing. R Foundation for statistical computing. https://www.R-project.org/ (2020)
- 45.Bates, D., Maechler, M. & Bolker, B. lme4: Linear mixed-effects models using S4 classes. R package version 0.999999-0 (2012).
- 46.Singmann, H., Bolker, B., Westfall, J., Aust, F. & Ben-Shachar, M. S. afex: Analysis of factorial experiments. R package version 0.13–145 (2015).
- 47.Lenth, R., Singmann, H., Love, J., Buerkner, P. & Herve, M. emmeans: Estimated marginal means, aka least-squares means. R package version 1.7. 2 (2019).
- 48.Gramfort, A. et al. MNE software for processing MEG and EEG data. NeuroImage86, 446–460 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Berman, M. G. et al. Interacting with nature improves cognition and affect for individuals with depression. J. Affect. Disord.140(3), 300–305 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Berman, M. G., Jonides, J. & Kaplan, S. The cognitive benefits of interacting with nature. Psychol. Sci.19(12), 1207–1212 (2008). [DOI] [PubMed] [Google Scholar]
- 51.Chan, J. S. et al. Special issue–therapeutic benefits of physical activity for mood: A systematic review on the effects of exercise intensity, duration, and modality. J. Psychol.153(1), 102–125 (2019). [DOI] [PubMed] [Google Scholar]
- 52.Schindler, I. et al. Measuring aesthetic emotions: A review of the literature and a new assessment tool. PloS One12(6), e0178899 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Van den Berg, A. E., Koole, S. L. & Van Der Wulp, N. Y. Environmental preference and restoration: (How) are they related?. J. Environ. Psychol.23(2), 135–146 (2003). [Google Scholar]
- 54.Rim, N., Kardan, O., Krishnan, S., Bainbridge, W. A. & Berman, M. G. Perceiving natural images may consume less cognitive resources: Evidence from image memorability, edge magnitudes, and spectral content. J. Vis.24(10), 1369–1369 (2024). [Google Scholar]
- 55.Fintzi, A. R. & Mahon, B. Z. A bimodal tuning curve for spatial frequency across left and right human orbital frontal cortex during object recognition. Cereb. Cortex24(5), 1311–1318 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Valtchanov, D. & Ellard, C. G. Cognitive and affective responses to natural scenes: Effects of low-level visual properties on preference, cognitive load and eye-movements. J. Environ. Psychol.43, 184–195 (2015). [Google Scholar]
- 57.McDonnell, A. S. et al. Immersion in nature decreases neural sensitivity to extrinsic reward (revise and resubmit).
- 58.Sudimac, S., Sale, V. & Kühn, S. How nature nurtures: Amygdala activity decreases as the result of a one-hour walk in nature. Mol. Psychiatry27(11), 4446–4452 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ulrich, R. S. et al. Stress recovery during exposure to natural and urban environments. J. Environ. Psychol.11(3), 201–230 (1991). [Google Scholar]
- 60.Scott, E. E., McDonnell, A. S., LoTemplio, S. B., Uchino, B. N. & Strayer, D. L. Toward a unified model of stress recovery and cognitive restoration in nature. Parks Steward. Forum37, 46–60. 10.5070/P537151710 (2021). [Google Scholar]
- 61.Hopman, R. J., LoTemplio, S. B., Scott, E. E., McKinney, T. L. & Strayer, D. L. Resting-state posterior alpha power changes with prolonged exposure in a natural environment. Cogn. Res. Princ. Implic.5, 1–13 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Neale, C. et al. The impact of walking in different urban environments on brain activity in older people. Cities Health4(1), 94–106 (2019). [Google Scholar]
- 63.Chen, Z., He, Y. & Yu, Y. Attention restoration during environmental exposure via alpha-theta oscillations and synchronization. J. Environ. Psychol.68, 101406 (2020). [Google Scholar]
- 64.Stangl, M., Maoz, S. L. & Suthana, N. Mobile cognition: Imaging the human brain in the ‘real world’. Nat. Rev. Neurosci.24(6), 347–362 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Chaumon, M., Bishop, D. V. & Busch, N. A. A practical guide to the selection of independent components of the electroencephalogram for artifact correction. J. Neurosci. Methods250, 47–63 (2015). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated during the current study are available from the corresponding author on reasonable request.