

Abstract
In the last decade, messenger ribonucleic acid (mRNA)-based drugs have gained great interest in both immunotherapy and non-immunogenic applications. This surge in interest can be largely attributed to the demonstration of distinct advantages offered by various mRNA molecules, alongside the rapid advancements in nucleic acid delivery systems. It is noteworthy that the immunogenicity of mRNA drugs presents a double-edged sword. In the context of immunotherapy, extra supplementation of adjuvant is generally required for induction of robust immune responses. Conversely, in non-immunotherapeutic scenarios, immune activation is unwanted considering the host tolerability and high expression demand for mRNA-encoded functional proteins. Herein, mainly focused on the linear non-replicating mRNA, we overview the preclinical and clinical progress and prospects of mRNA medicines encompassing vaccines and other therapeutics. We also highlight the importance of focusing on the host-specific variations, including age, gender, pathological condition, and concurrent medication of individual patient, for maximized efficacy and safety upon mRNA administration. Furthermore, we deliberate on the potential challenges that mRNA drugs may encounter in the realm of disease treatment, the current endeavors of improvement, as well as the application prospects for future advancements. Overall, this review aims to present a comprehensive understanding of mRNA-based therapies while illuminating the prospective development and clinical application of mRNA drugs.
Subject terms: Nucleic-acid therapeutics, Preclinical research, Translational research, Genetics research
Introduction
Recently, messenger ribonucleic acid (mRNA) therapy represents a novel approach for treating a wide range of diseases, encompassing both immune-related and non-immune conditions. Amidst the COVID-19 pandemic, mRNA vaccines have achieved remarkable advancements, owing to the unwavering dedication of numerous scientists who have been at the forefront of mRNA research for decades.
However, despite a plethora of preclinical studies conducted, successful translation of mRNA medicines into clinical applications remains limited, probably due to the suboptimal design and administration of mRNA drugs to patients with specific physiological and pathological conditions. Distinct characteristics of mRNA drugs are demanded in different therapeutic indications.1 Specifically, in immunogenic applications that include cancer immunotherapies and infectious disease vaccines, appropriate incorporation of adjuvants is required for eliciting augmented host immune responses. In contrast, in non-immunotherapies encompassing protein replacement/supplementation therapy, regenerative medicine therapy, and genetic editing, the immunogenicity of mRNA drugs is unfavorable, which may lead to diminished protein expression and even cause adverse reactions. On the other hand, the host individual variations, such as, age, gender, disease and medical history may influence both the efficacy and safety of mRNA medication. For example, compared to healthy adults, immunocompromised patients exhibit inadequate immune responses following initial mRNA vaccine inoculation, where repeated vaccinations are recommended for the establishment of sufficient immune protection.2 Nevertheless, repeated doses may cause potential risks that exacerbate the pathological burden.3 To date, there is still a lack of systemic understanding over the fate and outcomes of mRNA drugs in personalized recipient.
In this review, we first summarize the preclinical and clinical applications of mRNA drugs pertaining to immunotherapy and non-immunotherapy, then discuss the impact from patient physiological and pathological characteristics. Finally, we provide insights into the future directions and research priorities of mRNA drugs.
Evolution and milestones in mRNA-based drugs
Brief history of mRNA drug development
Chronologically (Fig. 1), mRNA was first discovered by pioneering researches in 1961.4 However, it was not until 1990 when Wolff et al. successfully expressed proteins by injecting mRNA into the body5 that mRNA gradually gained recognition as a therapeutic modality.1 In 1999, mRNA-engineered dendritic cells (DCs) entered clinical trial as antitumor vaccines for the first time (NCT00004211).6–8 In 2004, Weide et al. conducted the initial clinical trial involving the direct injection of protamine-stabilized mRNA into the human body to target metastatic melanoma (NCT00204607),9 wherein the mRNA encoded tumor-associated antigens (TAAs)—Melan-A, Mage-A1, Mage-A3, Tyrosinase, gp100, and Survivin.10 In 2005, Karikó et al. demonstrated that RNA could evade immune detection when it was naturally modified with nucleotides such as 5-methylcytosine (m5C), 5-methyluridine (m5U), N6-methyladenosine (m6A), pseudouridine (Ψ), and 2′-O-Methyluridine.11,12 The identification of this approach to mitigate the immunogenicity of mRNA is crucial for broadening the scope of mRNA therapeutics in non-immunological medical fields. In 2013, CureVac conducted an initial evaluation of the safety and immunogenicity of a prophylactic mRNA vaccine coding the rabies virus glycoprotein (CV7201) in a clinical trial (NCT02241135) involving healthy adults.13 This trial represents the first clinical study utilizing mRNA vaccines to combat infectious diseases.12 Then, in 2016, the therapeutic potential of mRNA was first unleashed in the field of protein supplementation therapy (NCT02935712). Notably, in 2020, two antivirus mRNA vaccines—mRNA-1273 by Moderna (NCT04283461) and BNT162b2 by Pfizer/BioNTech (NCT04380701), were quickly approved by the United States (U.S.) Food and Drug Administration (FDA) and put into use during the COVID-19 outbreak, which achieved great success and marked a significant milestone in emergency response against infectious diseases.14 Since then, mRNA drugs have experienced an explosion of development.
Fig. 1.

Chronological development of mRNA drugs. Yellow box, common events of mRNA drugs; green box, mRNA-based non-immunotherapy; red box, mRNA-based immunotherapy. From 1961 to 1990: mRNA discovery and the maturation of IVT mRNA technology, including the discovery of mRNA,4 purified mRNA could be translated into proteins in the mammalian cell-free system,518 discovery of mRNA cap,519 discovery of single-stranded circular RNA,520 cap analog commercialized,18 synthetic mRNA was first produced in the laboratory by IVT,374 T7 RNA polymerases commercialized.335 From 1990 to 2019: the exploration of mRNA vaccines, particularly for cancer therapy, including IVT mRNA injected into the mouse skeletal muscle achieved protein translation,5 vasopressin mRNA injected into the hypothalamus of Brattleboro rats was found to successfully express vasopressin,357 mRNA vaccine encoding tumor antigen in mice,171 first clinical trial of mRNA-engineered DCs vaccine strategy,6–8 first DC vaccine with autologous tumor mRNA was used to treat clinical Phase I/II of advanced malignant melanoma trial (NCT01278940),332,333 first attempt was made to inject an mRNA vaccine directly into humans to fight tumors (NCT00204607),9,10 nucleotide-modified RNA reduced the potential for immune stimulation,11 first personalized cancer mRNA vaccine in clinical trial (NCT02035956),521,522 first clinical trial of prophylactic mRNA vaccine (CV7201) against rabies (NCT02241135),13 NIH called for gender to be included in biological variables in preclinical and clinical studies,435 first mRNA (AZD8601) therapy encoding VEGF-A to enter the clinic (NCT02935712),523 first clinical trial of mRNA-encoding immunostimulant (mRNA-2416, NCT03323398).524 From 2019 to the present: rapid development of mRNA-based therapeutics, including two mRNA vaccines (mRNA-1273486 and BNT162b2) have been approved for emergency use by the FDA,123,124 first to use unmodified mRNA in regenerative medicine,370 forty-three COVID-19 mRNA vaccines were in clinical trials,125 first combining mRNA therapy with photodynamic therapy to fight tumors,290 Phase III trial of mRNA-4157 plus Pembrolizumab in the treatment of melanoma.501 IVT in vitro transcription, NIH the National Institutes of Health, DCs dendritic cells, FDA the Food and Drug Administration. The graphic is created with Adobe Illustrator
Classification, advantages, and limitations of mRNA drugs
mRNA, a single-stranded ribonucleic acid, acts as a transient carrier for the genetic information transcribed from DNA to guide protein synthesis.15 The execution of mRNA function in eukaryotes involves multiple stages, including transcription, post-transcriptional processing, intracellular transport, and translation.16 Prior to the maturation of mRNA, precursor messenger RNA (pre-mRNA) must undergo three essential processing steps: addition of a 5’ cap structure, cleavage mediated by numerous protein factors, and 3’-end processing which includes adding a polyadenosine [poly(A)] tail. These pre-mRNA processing steps are crucial for the generation of mature mRNA.17 With the commercialization of Cap analogs and T7 RNA polymerase in 1983 and 1985, respectively, the technology for in vitro transcription (IVT) of mRNA has steadily matured.18 The structural characteristics of IVT mRNA closely resemble those of endogenous mRNA found in natural eukaryotic cells.19
The linear non-replicating mRNA, representative of IVT mRNA, is composed of five segments in the 5’ to 3’ direction: the 5’ cap, the 5’ untranslated region (UTR), an open reading frame (ORF) encoding the target protein, the 3’ UTR, and a poly(A) tail.20–22 The ORF functions as the coding sequence for protein translation and is an indispensable element of IVT mRNA. The UTR, although non-coding, plays a pivotal role in ribosome recruitment and successful mRNA translation. Furthermore, the 5’ cap and 3’ Poly(A) tail structures significantly contribute to enhancing mRNA stability and translation efficiency.23,24 One of the key attributes of mRNA is its highly customizable nature. The optimization of mRNA involves nucleotide sequence refinement, chemical modifications,25 and mRNA purification. Various factors must be taken into consideration during mRNA design, including the influence of nucleotide sequence on RNA folding, immunogenicity, enhancement of mRNA stability, and maximization of the expression of the target protein.26,27 The immunogenicity of exogenous mRNA elicits an innate immune response, leading to the suppression of exogenous mRNA translation in the body.28 Substituting uridine with modified nucleotides such as m5U and Ψ, which are not recognized by pattern-recognition receptors, can mitigate the immunogenicity of mRNA.26 In addition, several studies suggest that Ψ modification may enhance the stability of transcripts.29 Nevertheless, there have been investigations suggesting that specific circumstances may disrupt the stability of mRNA.30 It is noteworthy that there are currently more than 170 identified types of RNA modifications.31 Exploring the optimization of mRNA holds substantial research potential.32
Compared to recombinant protein drugs, mRNA drugs have a shorter production cycle33 and are not limited to short peptide sequences.34 mRNA vaccines can easily deliver numerous tumor antigen fragments simultaneously, thereby increasing the diversity of antigenic epitopes.33 In contrast to DNA vaccines, mRNA vaccines circumvent the need to enter the nucleus, thereby obviating the potential for gene insertion and subsequent mutations.33,35 In addition, mRNA exhibits modifiable immunogenicity36 and can be effectively potentiated by adjuvants to efficiently elicit humoral and cellular immunity.35 Moreover, mRNA vaccines have the potential to become personalized therapeutic drugs, in which they have application prospects for targeting specific tumors37 and treating rare diseases. Nevertheless, mRNA is susceptible to degradation by nucleases33 and suffers from thermal instability, necessitating a cold-chain infrastructure for storage and transportation, thereby increasing the overall cost.38 Furthermore, mRNA, being a large polyanionic structure, presents challenges in traversing cell and tissue barriers.33 Overcoming these limitations necessitates the development of appropriate delivery strategies.
Two innovative mRNA structures have been developed for specific purposes, namely self-amplifying mRNA (saRNA)39 and circular mRNA (circRNA).40 saRNA can confer equivalent vaccine protection effects to those of non-replicating mRNA at a reduced dosage.41–43 The primary distinction between saRNA and linear non-replicating mRNA is that saRNA contains additional self-amplifying replicon genes that originate from multiple positive-strand RNA viruses,44–46 resulting in a larger molecular size.47 Owing to the extensive anionic structure of saRNA, its delivery vehicle often comprises polycations. Nevertheless, the use of such delivery vectors may lead to high charge density that induces cytotoxicity. Hence, it is imperative to optimize a suitable delivery method for enhancing both the loading efficiency and the overall safety of saRNA drugs. Dastgerdi et al.48 demonstrated that optimizing the ratio of polyanions, such as γ-polyglutamic acid, to polycations in RNA formulations is an effective approach for improving the delivery efficiency and safety of saRNA. Furthermore, given the continuous generation of new mRNA within the body by saRNA vaccines, it is important to remain vigilant about safety concerns such as the potential for severe systemic or local inflammatory responses.49 CircRNA is a class of covalently closed-loop single-stranded RNA molecule that is not initiated by the classical translation pathway due to the absence of a 5’ cap and a 3’ tail.50 The internal loading of the ribosome may be the sole means for its translation to commence.51 The capability to encode proteins without the classical pathway is granted by circRNA’s ability to translate without a 5’ cap.50 Meanwhile, circRNA exhibits better structural and biochemical stability due to its inherent resistance against exonucleases-mediated degradation.52 Therefore, the average half-life of circRNA in cells significantly exceeds that of linear mRNA.53 Hence, cirRNA has the capacity to extend antigen production and sustain enduring immune responses.54 Nevertheless, despite the more persistent and robust expression of encoded proteins, circRNA seems to perform poorly in targeted delivery.55 At present, non-replicating IVT mRNA, which has been well-established in production and quality control, is primarily used in preclinical and clinical practice.
Delivery system and preparation strategy of mRNA drugs
Due to the susceptibility of IVT mRNA to degradation by nucleases, it is crucial to select an appropriate delivery vehicle to safeguard the integrity of the mRNA prior to its intracellular delivery.56 An innovation initially not intended for mRNA but crucial for mRNA drug development is the advancement of delivery vectors, particularly the development of lipid nanoparticles (LNPs). At present, mRNA delivery systems can be classified into viral and nonviral systems.57
Virus-related delivery systems include both viruses and virus-like particles (VLPs).58 Viral vectors encompass adenoviruses, adeno-associated viruses (AAV), lentiviruses, herpes simplex virus (HSV), and Sendai virus.59,60 The formation of VLPs is facilitated by the structural proteins of a virus, which are viral components devoid of genetic material.61 In comparison to nonviral vectors, viral vectors possess the inherent advantage of efficiently entering cells and delivering nucleic acid drugs.58 This approach holds significant promise for vaccine development.62 Several clinical trials are currently utilizing viral vectors to deliver genetic material for the treatment of diseases. For instance, AAV9-mediated CLN6 gene therapy (AT-GTX-501) has progressed to clinical stages I/II (NCT02725580), and interim findings indicated that AT-GTX-501 demonstrated favorable tolerability and could ameliorate the deterioration in motor and language function among pediatric patients with variant late infantile ceroid lipofuscinosis 6 (vLINCL6).63 Nevertheless, virus-mediated delivery systems pose potential biosecurity concerns (e.g., tumorgenicity and immunogenicity), exhibit low packaging efficiency, and entail high manufacturing costs.59 These limitations have driven researchers to investigate alternative delivery methods for mRNA vaccines, such as nonviral delivery systems.62
Various nonviral delivery vectors, such as LNPs,64–68 polymeric nanoparticles,67,69,70 lipid enveloped hybrid nanoparticles,66 protein/peptide-based nanoparticles,67,68 lyotropic liquid crystalline lipid nanoparticles (LCNPs),71,72 inorganic nanoparticles,65 nanoemulsions,73,74 exosomes,64,75 hydrogels,76,77 polymeric nanoparticle gel,78 and biological membrane-based vesicles,79 have been developed for efficient and versatile mRNA delivery. Optimizing the route of mRNA drug administration is essential for enhancing therapeutic efficacy, as it can significantly impact the biodistribution of the drug within tissues. Generally, intravenously injected mRNA drugs tend to accumulate in the liver, whereas locally administrated ones often prolong the duration of protein expression at the injection site and deliver sustained therapeutic effects. Subcutaneous and intramuscular injections represent the predominant routes for administering mRNA vaccines.80 Alternative mRNA drug delivery methods include aerosol inhalation to directly target the lungs,81–83 eye drops,84 intravitreal administration, subretinal administration,80 oral administration,85,86 transcutaneous route,87 and in utero delivery.88
Despite the development of various types of delivery vectors in preclinical studies, LNPs remain the established preferred delivery system for mRNA due to its clinical validation.89 Therefore, a comprehensive understanding of the design strategies employed in LNPs is essential for the effective implementation of mRNA therapies. The current formulation of LNPs for nucleic acid delivery typically comprises four key components: ionizable lipids, helper lipids, cholesterol, and polyethylene glycol (PEG)-lipids,90,91 with the ratio of ~50:10:38.5:1.5 mol%.92 Ionizable lipids play a crucial role in encapsulating mRNA and facilitating the successful transfection of mRNA into cells.93 The incorporation of PEG-lipids serves primarily to evade macrophage phagocytosis and ensure prolonged systemic circulation of LNPs. The inclusion of helper lipids and cholesterol allows for modulation of LNPs rigidity.94 Kulkarni et al.95 demonstrated that helper lipids and cholesterol play crucial roles in the formulation of LNPs, aiding in the encapsulation of genetic material.96 However, Su et al.97 broke away from the traditional LNPs model and discovered that cholesterol and phospholipids may not be essential for LNPs.
It is essential to elucidate the considerations for designing LNPs delivery systems, encompassing payload efficiency, stability, circulation time, facilitation of endosomal escape, biodegradability, immunogenicity, safety, and targeting capability.98 The precise structure of cationic lipids is pivotal, typically comprising a positively charged head and multiple hydrophobic tails. Even minor alterations, such as the addition or removal of an atom or a functional group, can significantly impact tissue targeting, cellular uptake, and endosomal escape.99 The design of ionizable lipids necessitates careful consideration of the pKa values of the headgroup in order to achieve an optimal pH-dependent electrostatic pattern.98 Ionizable lipids are typically positively charged in acidic pH environments and possess the capability to aggregate negatively charged mRNA into LNPs. At physiological pH, they become neutralized to minimize toxicity and enhance the biocompatibility of mRNA LNPs. Upon uptake by cells into acidic endosomes, they undergo protonation, thereby promoting the translocation of mRNA from endosomes into the cytosol.93 The pKa values of ionizable lipids can be modified by altering the chemical composition of the headgroup, such as imidazole, ester, and piperazine.98 The hydrophobic tails of the ionizable lipids play a crucial role in facilitating the assembly of LNPs and ensuring the stable encapsulation of mRNA. The design of the linker region between the headgroup and the lipid tails has a significant impact on the overall pH sensitivity of the ionizable lipids, consequently influencing both the release kinetics and magnitude of mRNA.61 The shape of ionizable lipids can also impact their functionality. More conical ionizable lipids, with a molecular structure that is not compatible with the lipid bilayer, can facilitate mRNA escape from lysosomes.100
The safety and stability concerns associated with mRNA therapy encompass various facets. With regard to safety, it is imperative to assess the biocompatibility of mRNA and its delivery system, minimize direct cellular damage, and mitigate the elicitation of unnecessary immune responses.101 However, the presence of pH-dependent or permanent cationic properties may result in safety concerns, such as disruption of cellular membranes and organelles, release of degradative enzymes from lysosomes, and damage to DNA. As a result, biocompatible molecular components capable of forming covalent ester or amide bonds have been developed for the production of biodegradable ionizable cationic lipids.102 In terms of stability, the carrier design should ensure protection of the mRNA from enzymatic degradation without impacting its release.101 In addition, enhancing the thermal stability of mRNA LNPs is crucial for reducing storage costs. For example, in order to enhance the thermostability of mRNA LNPs, researchers developed a novel ionizable lipid DOG-IM4 modified with imidazole.103
Modifying the physical and chemical characteristics of LNPs, such as their size, surface charge, and surface hydrophobicity, can impact their biological activity.104 Generally, hydrophilic LNPs with a neutral charge and a size smaller than 100 nm exhibit prolonged circulation in the bloodstream. LNPs with a positive charge are more conducive to cellular uptake.105 The morphology and nanostructure of LNPs are correlated with the transfection efficiency of mRNA. Nevertheless, the morphology of mRNA LNPs remains ambiguous, necessitating further research for elucidation.106 Furthermore, the precise localization of the four components within the LNPs remains unclear, posing certain limitations in molecular design.107 In order to tackle more demanding tasks for LNPs, such as targeted functionality, cellular penetration, and endosomal escape, the surface of LNPs can be subject to modification. Polymers like PEG-modified lipids are the predominant surface modifiers utilized for LNPs.108 Nevertheless, surface-modified LNPs may present potential adverse effects, such as allergic responses triggered by lipids modified with PEG.61 Furthermore, the challenges in scaling up the production of surface-modified LNPs could impede their commercialization.108
Presently, the commonly utilized methodologies for formulating LNPs encompass the thin-film hydration approach, solvent injection technique, reverse evaporation method, and microfluidic technology.109 Depending on the specific application, microfluidic chips can be fabricated from a variety of materials such as silicon, glass, polydimethylsiloxane (PDMS), cyclo-olefin polymers and copolymers (COPs/COCs). In addition, various fabrication methods including photolithography, electron beam lithography, wet and dry etching, and embossing can be employed. Microfluidic technology offers numerous advantages in the production of LNPs, facilitating continuous liquid flow within the microfluidic platform to ensure consistent nanoparticle quality over time and mitigate batch-to-batch variations. Furthermore, the design flexibility of microfluidic channels enables adaptation to different rapid mixing modes.110 Microfluidic technology has not only yielded remarkable results in laboratory research, but also been adopted and integrated into industrial production processes. The microfluidic device utilized in the preparation of BNT162b2 is an impingement jet mixer.111 Furthermore, ongoing advancements in automated high-throughput preparation techniques are continuously enhancing the production of LNPs112 and other nanodelivery vectors.113
Preclinical and clinical applications of mRNA-based drugs
Immunotherapy
mRNA vaccines outplayed their traditional counterparts in rapid development and cost-effectiveness trait,114 rendering them a promising tool for immunization against viruses, tumors, bacteria, and parasite. Despite the fact that mRNA was discovered in 1961, the revolutionary breakthrough of using it as a vaccine has only occurred in recent years, due to its susceptibility to enzymatic degradation, inefficient in vivo delivery, and intrinsic immunogenicity.115
In order to achieve antiviral, antitumor, antibacterial, and antiparasitic effects, mRNA-based immunotherapies must elicit a robust and specific host immune response to eliminate pathogens and confer durable protection.28 mRNA vaccines encoding disease-specific antigens function by transfecting antigen-presenting cells (APCs). The protein antigen is translated within the cell and released into the extracellular space or degraded by the proteasome to expose the antigenic sites. Intrinsic antigens can be presented by the cell surface major histocompatibility complex I (MHC I) to induce maturation of cluster of differentiation (CD) 8+ T cells, while extracellular antigen proteins can be recognized by B cells, internalized, degraded by APC cells, and presented through the MHC II class pathway to activate CD4+ T helper (Th) cells.28,116
In addition to disease-specific antigens, proteins related to immunotherapy encompass antibodies, cytokines, ligands, tumor suppressor proteins, and other functional proteins. It is worth noting that mRNA therapies encoding cytokines could be utilized to either enhance or suppress the immune response. Cytokines are a class of proteins that modulate immune cells by activating downstream cytokine receptors.117 Stimulatory cytokines are suitable for combating infectious diseases and cancer, while inhibitory cytokines are appropriate for treating autoimmune diseases. It is noteworthy that certain cytokines, such as Interleukin (IL)-2, do not have a fixed function in regulating the immune response; rather, their function changes with variations in concentration.118,119 Moreover, mRNA-based adoptive cell therapies may not be exclusively utilized for immune modulation or suppression, contingent upon the protein encoded by the mRNA. For instance, BNT211 is employed in the treatment of CLDN6-positive advanced solid tumors,120 while Descartes-08 is designated for autoimmune disorders.121
In general, mRNA possesses the capacity to encode diverse proteins including antigens, antibodies, ligands, and tumor suppressor proteins. In addition, mRNA can also be harnessed in combination with adoptive cell therapy (Fig. 2), thus showcasing immense potential within the realm of immunotherapy (Table 1).
Fig. 2.

mRNA codes for immunotherapy-associated antigen, antibody, cytokine, ligand, tumor suppressor protein, and adoptive cell therapy. RV rotavirus, VZV varicella-zoster virus, RSV respiratory syncytial virus, CMV cytomegalovirus, HPV human papillomavirus, EBV Epstein–Barr virus, RABV rabies virus, HIV human immunodeficiency virus, MPXV monkeypox virus, anti-VEGF anti-vascular endothelial growth factor. The graphic is created with BioRender.com
Table 1.
Representative completed and ongoing clinical studies (immunotherapy)
| Therapy | Subclass | mRNA drug | Application | mRNA-encoded protein | Delivery system | Administration route | Study started | Phase | Status | Result | NCT number | Sponsor | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Disease-specific antigen therapy | Virus antigen | mRNA-1273 | COVID-19 | Prefusion stabilized full-length spike protein | LNPs (SM-102, PEG2000-DMG, DSPC, cholesterol) | Intramuscular injection | 2020-07-27 | Phase III | Completed | Show 94.1% efficacy in preventing COVID-19 disease, with no safety concerns found except for transient local or systemic reactions | NCT04470427 | ModernaTX, Inc. | 486 |
| BNT162b2 | COVID-19 | SARS-CoV-2 full-length spike protein | LNPs (ALC-0315, ALC-0159, DSPC, cholesterol) | Intramuscular injection | 2020-04-29 | Phase II/III | Completed | Good immune efficacy against COVID-19 with a sustained safety profile and acceptable adverse events, but the immune efficacy declines after 6 months | NCT04368728 | BioNTech SE | 487 | ||
| mRNA-1345 | RSV | RSV prefusion stabilized F (preF) glycoprotein | LNPs (ionizable lipid, phospholipid, PEG lipid, sterol) | Intramuscular injection | 2023-10-06 | Phase III | Recruiting | / | NCT06067230 | ModernaTX, Inc. | 479 | ||
| mRNA-1010 | Seasonal influenza | Membrane-bound hemagglutinin (HA) surface glycoproteins of four influenza strains (A/H1N1, A/H3N2, B/Victoria, and B/Yamagata) | LNPs | Intramuscular injection | 2022-09-14 | Phase III | Completed | Acceptable safety and tolerability support continued study | NCT05566639 | ModernaTX, Inc. | 134,488 | ||
| mRNA-1647 | CMV | Two CMV antigens (glycoprotein B and the pentameric glycoprotein complex) | LNPs | Intramuscular injection | 2021-10-26 | Phase III | Active, not recruiting | / | NCT05085366 | ModernaTX, Inc. | 489 | ||
| mRNA-1325 | Zika virus | Premembrane and envelope E structural proteins (prME) from a Micronesia 2007 Zika virus isolate | LNPs | Intramuscular injection | 2016-12-21 | Phase I | Completed | Well tolerated, but poor Zika virus-specific nAb responses | NCT03014089 | ModernaTX, Inc. | 490 | ||
| mRNA-1893 | Zika virus | prME from the RIO-U1 Zika virus isolate | LNPs | Intramuscular injection | 2019-07-30 | Phase I | Completed | Well tolerated, induce strong Zika virus-specific serum nAb responses after two doses that supported the continued study of mRNA-1893 | NCT04064905 | ModernaTX, Inc. | 490 | ||
| mRNA-1388 | CHIKV | Full CHIKV structural polyprotein (capsid and envelope proteins E3, E2, 6 k/TF, and E1) from CHIKV West African strain 37,997 | LNPs (cholesterol, DPSC, ionizable lipid MC3, PEG2000-DMG) | Intramuscular injection | 2017-08-15 | Phase I | Completed | Good safety and immunogenicity | NCT03325075 | ModernaTX, Inc. | 491 | ||
| H10N8 mRNA vaccine (VAL-506440) | Influenza (H10N8) | Full-length, membrane-bound form of the HA glycoprotein from the H10N8 influenza strain (A/Jiangxi-Donghu/346/2013) | LNPs | Intramuscular injection | 2015-12 | Phase I | Completed | Well tolerated, trigger strong humoral immune responses | NCT03076385 | ModernaTX, Inc. | 492 | ||
| H7N9 mRNA vaccine (VAL-339851) | Influenza (H7N9) | Full-length, membrane-bound form of the HA glycoprotein from the H7N9 influenza strain (A/Anhui/1/2013) | LNPs | Intramuscular injection | 2016-05-11 | Phase I | Completed | Well tolerated, trigger strong humoral immune responses | NCT03345043 | ModernaTX, Inc. | 492 | ||
| CV7201 | Rabies | Rabies virus glycoprotein | Cationic protein protamine as stabilizer and adjuvant | Intradermal, intramuscular injection | 2013-10 | Phase I | Completed | Generally safe and reasonably tolerated | NCT02241135 | CureVac | 13 | ||
| CV7202 | Rabies | Rabies virus glycoprotein | LNPs (cholesterol, DSPC, PEGylated lipid, cationic lipid) | Intramuscular injection | 2018-10-12 | Phase I | Completed | Low doses (1 μg or 2 μg) were well-tolerated, whereas the 5 μg dose exhibited unacceptable reactogenicity | NCT03713086 | CureVac | 493 | ||
| mRNA-1644 | HIV | eOD-GT8 60mer | Self-assembling nanoparticles | Intramuscular injection | 2022-05-25 | Phase I | Active, not recruiting | / | NCT05414786 | International AIDS Vaccine Initiative | 156 | ||
| mRNA-1644v2-Core | HIV | Core-g28v2 60mer | LNPs | Intramuscular injection | 2021-11-12 | Phase I | Active, not recruiting | / | NCT05001373 | International AIDS Vaccine Initiative | 494,495 | ||
| BNT166a | MPXV | MPXV antigens A35, B6, H3, and M1 | LNPs | Intramuscular injection | 2023-09-21 | Phase I | Recruiting | / | NCT05988203 | BioNTech SE | 167 | ||
| EBV mRNA vaccine | EBV-positive advanced malignant tumors | Undisclosed | Undisclosed | Intramuscular injection | 2022-11-18 | Phase I | Recruiting | / | NCT05714748 | West China Hospital | 496 | ||
| BNT113 | HPV16 positive head and neck squamous cell carcinoma (HNSCC) that expresses PD-L1 | HPV16 oncoproteins E6 and E7 | Liposomal | Intravenous injection | 2021-01-07 | Phase II | Recruiting | Acceptable safety profile | NCT04534205 | BioNTech SE | 497 | ||
| HBV mRNA vaccine | HBV-associated refractory hepatocellular carcinoma | HBsAg | LNPs | Intramuscular injection | 2023-02-15 (estimated) | Phase I | Recruiting | / | NCT05738447 | West China Hospital | 498 | ||
| Tumor antigen | BNT111 | Melanoma | Melanoma TAAs: New York esophageal squamous cell carcinoma 1 (NY-ESO-1), tyrosinase, melanoma-associated antigen 3 (MAGE-A3), and transmembrane phosphatase with tensin homology (TPTE) | Lipoplexes | Intravenous administration | 2015-03 | Phase I | Completed | Strong immunogenicity and promising clinical activity | NCT02410733 | BioNTech SE | 174 | |
| BNT112 | Prostate cancer | Prostate cancer TAAs: kallikrein-2, kallikrein-3, acid phosphatase prostate, homeobox B13 (HOXB13), and NK3 homeobox 1 | Lipoplexes | Intravenous bolus injection | 2019-12-19 | Phase I/II | Terminated | Acceptable safety profile, induce robust prostate antigen-specific immune responses in patients with advanced prostate cancer | NCT04382898 | BioNTech SE | 499 | ||
| BNT116 | Non-small cell lung cancer | Six shared antigens frequently expressed in non-small cell lung cancer | Undisclosed | Intravenous injection | 2022-06-17 | Phase I | Recruiting | / | NCT05142189 | BioNTech SE | 500 | ||
| mRNA-4157 | Melanoma | Up to 34 patient-specific tumor neoantigens | LNPs | Intramuscular injection | 2019-07-18 | Phase IIb | Recruiting | Significantly extended distant metastasis-free survival in patients with resected high-risk melanoma as compared with Pembrolizumab monotherapy | NCT03897881 | ModernaTX, Inc. | 481 | ||
| mRNA-4157 | Melanoma | Up to 34 patient-specific tumor neoantigens | LNPs | Intramuscular injection | 2023-07-19 | Phase III | Recruiting | / | NCT05933577 | Merck Sharp & Dohme LLC | 501 | ||
| Autogene cevumeran (RO7198457) | Pancreatic cancer | Neoantigen | Lipoplex nanoparticles | Intravenous delivery | 2019-12-13 | Phase I | Active, not recruiting | Preliminarily shown to be safe in combination with Atezolizumab and mFOLFIRINOX and to delay recurrence in patients with surgically removed pancreatic ductal adenocarcinoma (PDAC) | NCT04161755 | Memorial Sloan Kettering Cancer Center | 502 | ||
| Personalized tumor neoantigen mRNA vaccine | Liver cancer | Personalized tumor neoantigen | LNPs | Subcutaneous injection | 2023-04-20 | Not applicable | Not yet recruiting | / | NCT05761717 | Shanghai Zhongshan Hospital | 503 | ||
| Tumor neoantigen mRNA vaccine | Advanced esophageal cancer and non-small cell lung cancer | Tumor neoantigen | LNPs | Subcutaneous injection | 2019-10-18 | Not applicable | Recruiting | / | NCT03908671 | Stemirna Therapeutics | 504 | ||
| GRT-C901 and GRT-R902 | Solid tumors | Neoantigen (#samRNA) | Chimpanzee adenovirus vector | Intramuscular injection | 2019-02-13 | Phase I/II | Completed | Well tolerated, induce neoantigen-specific CD8+ T-cell response in all patients, subsequent Phase II/III initiated (NCT05141721) | NCT03639714 | Gritstone bio, Inc. | 505 | ||
| Bacterial antigen | mRNA-1975 and mRNA-1982 | Lyme disease | Undisclosed | Undisclosed | Intramuscular injection | 2023-07-26 | Phase I/II | Active, not recruiting | / | NCT05975099 | ModernaTX, Inc | 188 | |
| BNT164a1 and BNT164b1 | Tuberculosis | Undisclosed | Undisclosed | Intramuscular injection | 2023-07-31 | Phase I/II | Recruiting | / | NCT05547464 | BioNTech SE | 453 | ||
| Parasite antigen | BNT165b1 | Malaria | Part of the Plasmodium falciparum circumsporozoite protein (PfCSP) | LNPs | Intramuscular injection | 2022-12-15 | Phase I | Active, not recruiting | / | NCT05581641 | BioNTech SE | 201 | |
| Therapeutic antibody therapy | Antibody | mRNA-1944 | CHIKV | Light and heavy chains of a human monoclonal antibody (CHKV-24 IgG) targeting the CHIKV E2 glycoprotein | LNPs (a proprietary IAL; a proprietary high-purity PEG-2k-stearate monoester; cholesterol, DOPE) | Intravenous infusion | 2019-01-22 | Phase I | Completed | Acceptable safety profile, produce high levels of functionally neutralizing antibodies | NCT03829384 | ModernaTX, Inc. | 506 |
| BNT141 | Unresectable or metastatic CLDN18.2-positive gastric, pancreatic, ovarian and biliary tract tumors | Anti-CLDN18.2 antibodies | LNPs | Intravenous injection | 2022-01-18 | Phase I/II | Terminated | / | NCT04683939 | BioNTech SE | 507 | ||
| BNT142 | CLDN6-positive solid tumors | T-cell-engaging bispecific antibody against CLDN6 and the T-cell receptor–associated molecule CD3 | LNPs | Intravenous bolus/infusion | 2022-03-28 | Phase I/II | Recruiting | / | NCT05262530 | BioNTech SE | 508 | ||
| RNA-transfected mature autologous DC | Melanoma | Anti-CTLA-4 and anti-GITR mAb and melanoma TAAs MART, tyrosinase, and gp100, and MAGE-3 | DCs | Intranodal injection | 2010-01 | Phase I | Terminated | / | NCT01216436 | Duke University | 236 | ||
| Cytokine | Immune-potentiating | MEDI1191 | Solid tumors | IL-12 | LNPs | Intratumoral injection | 2019-05-08 | Phase I | Completed | / | NCT03946800 | MedImmune LLC | 259 |
| mRNA-2752 | Advanced solid tumors and lymphoma | OX40L, IL-23, and IL-36γ | LNPs | Intratumoral injection | 2018-11-27 | Phase I | Active, not recruiting | Tolerated, may be associated with tumor shrinkage | NCT03739931 | ModernaTX, Inc. | 509 | ||
| SAR441000 | Solid tumors | IL-12 single chain, IFN ɑ-2b, GM-CSF, and IL-15 sushi | Saline-formulated mixture | Intratumoral injection | 2019-01-03 | Phase I | Terminated | Well tolerated (clinical trial in 17 patients; July 2020), supporting further clinical studies | NCT03871348 | Sanofi | 263 | ||
| BNT153 and BNT152 | Solid tumors | IL-2, IL-7 | LNPs | Intravenous | 2021-06-08 | Phase I | Recruiting | / | NCT04710043 | BioNTech SE | 510 | ||
| Immune-suppressing | mRNA-6231 | Healthy volunteers | Human serum albumin IL-2 mutein fusion protein (HSA-IL2m) | LNPs | Subcutaneous injection | 2021-07-28 | Phase I | Completed | / | NCT04916431 | ModernaTX, Inc. | 511 | |
| Ligand | Ligand | TriMix | Breast cancer | DC-activating [CD40 ligand (CD40L), CD70, and constitutively active Toll-like receptor 4 (TLR4)] proteins | Undisclosed | Intratumoral administration | 2018-11-12 | Phase I | Recruiting | / | NCT03788083 | Universitair Ziekenhuis Brussel | 512 |
| Adoptive cell therapy | CAR-T | CLDN6 CAR-T cells +/− CARVac | CLDN6-positive advanced solid tumors | CLDN6 | Lipoplexes | Intravenous injection | 2020-09-16 | Phase I/II | Recruiting | Good safety profile and encouraging efficacy | NCT04503278 | BioNTech Cell & Gene Therapies GmbH | 120 |
| CAR-T | Descartes-08 | Myasthenia gravis | Anti-BCMA targeting CAR protein | Autogolous T cells | Infusion | 2019-12-04 | Phase Ib/IIa | Recruiting | Safe and well-tolerated | NCT04146051 | Cartesian Therapeutics | 121 | |
| TCR-T | LioCyx-M | Recurrent hepatocellular carcinoma | HBV-specific TCR | Autologous T cells | Infusion | 2015-07-02 | Phase I | Unknown status | Well tolerated, have no adverse effects on the transplanted liver | NCT02719782 | Lion TCR Pte. Ltd. | 513 | |
| CAR-PBMC | MCY-M11 | Advanced ovarian cancer and peritoneal mesothelioma | Anti-mesothelin CAR (Meso-CAR) | PBMCs | Intraperitoneal infusion | 2018-08-27 | Phase I | Terminated | Mediate effective and long-term antitumor response | NCT03608618 | MaxCyte, Inc. | 329 | |
| DCs | AGS-004 | HIV | CD40L and three or four autologous HIV-1 antigens | DCs | Undisclosed | 2008-02 | Phase II | Completed | / | NCT00672191 | Argos Therapeutics | 514 | |
| WT1 antigen-targeted DC vaccine | Acute myeloid leukemia (AML) | WT1 antigen mRNA loaded autologous DCs (by electroporation) | DCs | Undisclosed | 2012-10 | Phase II | Active, not recruiting | / | NCT01686334 | Zwi Berneman | 515 | ||
| DC vaccine (DCs transfected with Survivin, hTERT and p53 mRNA) | Metastatic breast cancer or malignant melanoma | Survivin, hTERT and p53 | DCs | Intradermal injection | 2009-09 | Phase I | Completed | / | NCT00978913 | Inge Marie Svane | 516 | ||
| Human CMV pp65-LAMP mRNA-pulsed autologous DCs | GBM | CMV protein pp65 | DCs | Intradermal and bilateral administration at the groin site (divided equally to both inguinal regions) | 2015-10-12 | Phase II | Completed | Effective GBM inhibition | NCT02366728 | Mustafa Khasraw, MBChB, MD, FRCP, FRACP | 517 |
LNPs lipid nanoparticles, PEG polyethylene glycol, DSPC distearoylphosphatidylcholine, RSV respiratory syncytial virus, CMV cytomegalovirus, CHIKV chikungunya virus, HIV human immunodeficiency virus, MPXV monkeypox virus, EBV Epstein–Barr virus, HPV human papillomavirus, HBV hepatitis B virus, TAAs tumor-associated antigens, DOPE 1,2-dioleyl-sn-glycero-3-phosphoethanolamine, CLDN6 claudin 6, DCs dendritic cells, IL interleukin, IFN interferon, GM-CSF granulocyte-macrophage colony-stimulating factor, TriMix mRNA-encoding CD40L, CD70, and TLR4, CAR chimeric antigen receptor, BCMA B-cell maturation antigen, TCR T-cell receptor, PBMC peripheral blood mononuclear cells, WTI Wilms’ tumor, GBM glioblastoma
Disease-specific antigen therapy
Virus antigen mRNA vaccine
SARS-CoV-2: In 2019, the novel coronavirus emerged globally, leading to widespread transmission and the tragic loss of numerous lives, thereby posing a huge threat to global health and sanitation. The scarcity of a specific therapeutic agent for post-infection treatment with SARS-CoV-2 caused global panic. In response to this challenge, a variety of SARS-CoV-2 vaccines have been propelled into an intense and accelerated research phase. mRNA vaccines, representing an innovative technology for the prevention of infectious diseases, were initially anticipated to require 5–6 years to reach the market.122 Encouragingly, following the public disclosure of the coronavirus RNA sequence in January 2020,122 it took less than 3 months for the mRNA vaccines, mRNA-1273 (NCT04283461, Phase I initiated in March 2020) and BNT162b2 (NCT04380701, Phase I and Phase II initiated in April 2020), to transition from development to clinical trials.123 Remarkably, within a year, both mRNA vaccines (BNT162b2 and mRNA-1273) rose to prominence, securing emergency approval from the U.S. FDA and were authorized for global mass vaccination against SARS-CoV-2.122,124 According to the consolidated information provided by the World Health Organization (WHO), as of March 30, 2023 (Fig. 3a), a total of 183 vaccines had entered the clinical stage, with RNA-based vaccines accounting for about 24% (43 in total), ranking second only to protein subunit vaccines which account for around 32% (59 in total).125
Fig. 3.
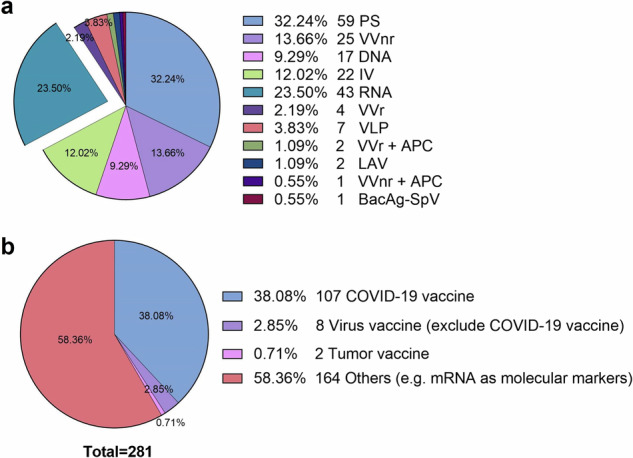
Application of mRNA vaccine in immunotherapy. a Types of COVID-19 vaccines in clinical trials (from WHO; March 30, 2023). b mRNA-based drugs in Phase III/IV trials. On clinicaltrial.gov, search for “mRNA” as the only keyword and the search criteria are limited to “Phase 3” and “Phase 4”. Note that this search method cannot find all mRNA vaccines, and some mRNA vaccines do not contain the word “mRNA”, so the figure should be viewed dialectically. PS protein subunit, VVnr viral vector (non-replicating), IV inactivated virus, VVr viral vector (replicating), VLP virus-like particle, VVr + APC VVr + antigen-presenting cell, LAV live attenuated virus, VVnr + APC VVnr + antigen-presenting cell, BacAg-SpV bacterial antigen-spore expression vector
Rotavirus: The successful application of mRNA vaccines against COVID-19 disease has prompted researchers to swiftly expand them to other antiviral fields. Rotaviruses (RVs) are responsible for causing diarrhea in children worldwide and can result in subsequent gastroenteritis, yet there is no specific treatment for this virus. Lu et al.14 designed an mRNA vaccine encoding the VP7 protein, which is one of the capsid proteins of RVs and plays an important role in the infection of target cells, for preventing RVs infection. The VP7 mRNA vaccine was inoculated to mice via intramuscular or subcutaneous injection (three doses). The findings demonstrated that the VP7 mRNA vaccine effectively stimulated T-cell immune responses and elicited RV-specific antibodies.
Varicella-zoster virus: Varicella-zoster virus (VZV), a member of the alphaherpesvirus subfamily, is a neurotropic human herpesvirus.126 The initial infection manifests as chickenpox symptoms, after which the virus remains latent in the body. Reactivation of the virus occurs when the immune system weakens due to aging or compromised immunity.127 In 2024, Huang et al.128 developed an mRNA-based VZV vaccine called ZOSAL, which employed ionizable LNPs for encapsulating sequence-optimized mRNA encoding the full-length glycoprotein E. In this study, the immunogenicity, safety, and immune mechanisms of ZOSAL were compared with those of the licensed protein-based vaccine Shingrix in mice and rhesus macaques. The findings demonstrated that ZOSAL exhibited superior immunogenicity, safety profile, and capacity to induce virus-specific T-cell immunity when compared to Shingrix.
Respiratory syncytial virus: Respiratory syncytial virus (RSV) is an enveloped, single-stranded RNA virus, which can cause respiratory illnesses.129 RSV can result in fatal outcomes for pediatric, geriatric, and immunocompromised individuals.130 One of the highly progressive vaccines aimed at combating RSV is mRNA-1345, which is presently being developed by ModernaTX, Inc. This vaccine encodes the membrane-anchored RSV prefusion stabilized F (preF) glycoprotein.131 Currently, the mRNA-1345 vaccine for RSV is undergoing Phase III trial (NCT05330975, NCT06067230), and it has already received FDA approval via fast-track designation for administration in individuals aged 60 years and older.132
Influenza virus: Influenza viruses have the ability to infect a wide range of vertebrates and are responsible for causing seasonal influenza as well as influenza pandemics in humans.133 ModernaTX, Inc. developed a quadrivalent mRNA vaccine (mRNA-1010) targeting seasonal influenza, encoding the four hemagglutinin (HA) surface glycoproteins of influenza strains (A/H1N1, A/H3N2, B/Victoria, and B/Yamagata). The vaccine has undergone testing in healthy adults during Phase I/II trial (NCT04956575), with interim results demonstrating favorable safety.134 Further, it has currently enrolled 22,510 adults aged 50 years and older (NCT05566639)135 and 8400 adults aged 18 years and older (NCT05827978)136 to initiate Phase III safety and efficacy studies.
Cytomegalovirus: Cytomegalovirus (CMV) is an incredibly prevalent virus, with estimated incidence rates of 60% in developed countries and 90% in developing countries. Although asymptomatic in healthy individuals, infection with CMV can pose a significant risk to immunocompromised patients (such as those undergoing organ transplantation), potentially leading to graft rejection and even life-threatening complications. In addition, it is imperative to acknowledge the issue of vertical transmission of CMV from pregnant women to fetuses, which can result in premature birth and profound permanent disabilities in neonates, encompassing cognitive impairment and visual deficits. In order to tackle this concern, ModernaTX, Inc. developed an mRNA vaccine (mRNA-1647) that encodes two CMV proteins, namely glycoprotein B (gB) and pentameric gH/gL/UL128/UL130/UL131A glycoprotein complex (pentamer). The vaccine has successfully completed Phase I clinical trial (NCT03382405), demonstrating that administration of three doses of mRNA-1647 (180 µg) could effectively elicit high titers of neutralizing antibodies (nAbs), broad neutralization activity, robust T-cell response, and long-lasting memory B cells in healthy adults,137 while maintaining an acceptable safety profile.138 Hu et al.139 compared mRNA-1647 (NCT03382405) with MF59-adjuvanted gB subunit (gB/MF59) vaccine (NCT00133497), and observed that although the gB-specific IgG responses were lower after inoculation with mRNA-1647 vaccine, it elicited persistent HCMV-specific antibody responses and higher antibody-dependent cellular cytotoxicity responses. In addition, a Phase III trial was conducted in healthy female participants aged 16–40 years with mRNA-1647 (NCT05085366).140
Human papillomavirus: The distinctive properties of certain viruses, such as carcinogenicity, render mRNA vaccines encoding viral proteins a promising strategy for combating tumors. HPV infection is widely recognized as the primary factor contributing to cervical cancer.141,142 The E6 and E7 proteins of human papillomavirus (HPV) have been established to modulate cell cycle and exhibit a high association with cervical cancer. In light of this, Lee et al.143 designed an mRNA vaccine encoding the E6 and E7 proteins, which was subsequently administered to C57BL/6J mice via intramuscular or subcutaneous injection. Their findings revealed that the mRNA vaccine markedly elicited robust T-cell-mediated immune responses in tumor-bearing mice, leading to a significant inhibition of tumor growth. Importantly, the experimental results demonstrated that the vaccine not only exhibited therapeutic potential against existing tumors but also displayed preventive efficacy.
Epstein–Barr virus: Epstein–Barr virus (EBV), an oncogenic virus in humans, is frequently associated with nasopharyngeal carcinoma (NPC). LMP2 is one of the main viral proteins. Xiang et al.144 delivered LMP2-mRNA LNPs to tumor-draining lymph nodes (TDLN) for the purpose of inducing activation and cytotoxicity of CD8+ T cells against tumor cells expressing LMP2. Zhao et al.145 developed three therapeutic EBV mRNA vaccines that encoded truncated latent EBV protein regions containing abundant T-cell epitopes, including truncated forms of latent membrane protein 2A (LMP2A), EBV nuclear antigen 1 (EBNA1), and EBV nuclear antigen 3A (EBNA3A). Their findings demonstrated that these EBV mRNA vaccines could effectively elicit antigen-specific immune responses, thereby suppressing tumor progression and prolonging the survival time of tumor-bearing mice.
Rabies virus: The neurotropic rabies virus (RABV), a member of the Lyssavirus family and a single-stranded RNA virus,146 has the ability to infect all warm-blooded animals,147 including humans, leading to the development of rabies. Rabies is a fatal neurological disease, and once the initial symptoms manifest, there is minimal time to pursue treatment options, as the mortality rate approaches 100%.146 Rabies causes an estimated 59,000 fatalities annually on a global scale.148 The currently most widely used rabies vaccines are all inactivated, requiring individuals to receive three to five doses for optimal immune protection.149,150 In summary, despite advancements in rabies vaccine development, the current availability of vaccines is constrained by factors such as high costs, limited production capacity and storage requirements, as well as the necessity for multiple doses.147 The mRNA vaccines represent a promising approach that has the potential to reduce costs and lower the required dosage for achieving effective immune protection.150 RABV glycoprotein (RABV-G), being the only virion-surface protein of RABV, plays a crucial role as an antigen in vaccine development.151 In 2013, CureVac conducted the initial Phase I clinical trial for an mRNA-based rabies vaccine in healthy adults (NCT02241135). The trial findings indicated that the mRNA vaccine (CV7201) encoding the RABV-G demonstrated a favorable safety profile.13 Subsequently, CureVac also progressed a mRNA vaccine named CV7202, encoding the RABV-G protein, into Phase I clinical trial (NCT03713086). Currently, research on the mRNA vaccine for RABV-G is ongoing. Cao et al.152 developed a novel mRNA LNPs that encodes RABV-G with the H270P mutation, which can stabilize the prefusion conformation of RABV-G. Their findings demonstrated that this vaccine elicited superior humoral and cellular immune responses compared to mRNA LNPs encoding RABV-G, indicating that structured-guided vaccine design may be the future direction for vaccine development.
Human immunodeficiency virus: Acquired immunodeficiency syndrome (AIDS), resulting from infection with the human immunodeficiency virus (HIV), remains a fatal disease for humans.153 Although current antiretroviral therapies154 and therapeutic vaccines can improve HIV-related morbidity and mortality, they are still unable to completely eradicate HIV.155 At present, the HIV mRNA vaccines that have been developed and entered clinical trials include the eOD-GT8 60mer mRNA vaccine (mRNA-1644, NCT05414786)156 and the Core-g28v2 60mer mRNA vaccine (mRNA-1644v2-Core, NCT05001373).157 As of June 2024, these vaccines are still undergoing Phase I clinical trials. Despite the evaluation of various HIV vaccines in preclinical and clinical studies, the overall outcomes have been unsatisfactory.158 Further endeavors are imperative for the development of an effective HIV vaccine. Mandal et al.159 designed LNPs containing mRNA that encoded multiple epitopes of HIV viral protease cleavage sites. The mRNA LNPs exhibited long-term stability at cold-chain temperatures and could elicit potent cellular immunity, making it as a promising candidate for a prophylactic HIV mRNA vaccine. A major obstacle in the eradication of HIV lies in the virus’s ability to conceal itself within host cells, including myeloid cells and CD4+ T cells.160 It appears that vaccines alone may not be sufficient for curing HIV, prompting consideration of combining vaccines with other treatment modalities.155,160 The extensive genetic diversity of HIV and its capacity to integrate into the host cell genome undoubtedly pose significant challenges for the development of HIV vaccines.161,162
Monkeypox virus: Monkeypox is a communicable disease caused by the monkeypox virus (MPXV), which can be transmitted between humans and animals.163,164 MPXV is a type of double-stranded DNA virus163,165 that belongs to the Orthopoxvirus genus.165,166 Currently, a multivalent mRNA vaccine (BNT166) encoding MPXV antigens A35, B6, H3, and M1 has entered Phase I clinical trial (NCT05988203)167. Several preclinical studies are underway to develop novel mRNA vaccines targeting various antigens of the virus. Tian et al.168 designed an mRNA-based vaccine encoding the MPXV A29L antigen. Su et al.169 developed a quadrivalent mRNA vaccine that encoded the antigens A27, L1, A33, and B5 of vaccinia virus. It is noteworthy that the sera from mice immunized with this vaccine demonstrated reactivity with the antigens of various orthopoxviruses in vitro. This suggests that the mRNA vaccine holds promise for safeguarding humans against MPXV and other orthopoxvirus infections.
Tumor antigen mRNA vaccine
Cancer is a major cause of global mortality.170 The inception of antitumor mRNA vaccines can be traced back to 1995, when Conry et al.171 constructed an mRNA encoding the tumor antigen–human carcinogenic antigen (CEA) that could be directly injected into the skin to combat tumors in mouse models. In 2004, a clinical trial was initiated to intradermally administer mRNA-encoding melanoma-associated antigens to stage III/IV melanoma patients (NCT00204607).9 In 2009, Weide et al.10 reported the results of the Phase I/II trial, demonstrating that the feasibility and safety of direct injection of protamine-protected mRNA, thereby encouraging further clinical studies on mRNA vaccines.
A critical step in the development of cancer vaccines involves the identification and selection of appropriate tumor antigens, encompassing both TAAs and tumor-specific antigens (TSAs).172
TAAs are non-mutated proteins that exhibit high expression in tumors while showing no-to-low expression in normal tissues.173 The RNA-LPX vaccine (BNT111), which has successfully completed Phase I clinical trial (NCT02410733) and is now progressing to Phase II (NCT04526899), is a tumor vaccine designed to encode TAAs (New York esophageal squamous cell carcinoma 1, melanoma-associated antigen 3, tyrosinase, and transmembrane phosphatase with tensin homology) for melanoma.174,175 The results of the clinical trial demonstrated that the intravenous administration of BNT111 vaccine in patients can elicit a robust and broad T-cell response against TAAs.117 However, the use of TAAs as tumor antigens is limited by their expression in normal tissues to some extent, which may hinder the development of effective antitumor immune responses owing to the self-tolerance mechanisms and the potential for off-target effects that may lead to autoimmune toxicity.173,176
TSAs, which result from mutations in somatic cells, hold significant potential for inducing specific T-cell responses against tumors.173 For instance, Moderna’s mRNA-4157 is an mRNA vaccine that encodes up to 34 patient-specific tumor neoantigens.177 It became the first mRNA cancer vaccine to enter Phase III clinical trial (NCT05933577). The clinical IIb trial of mRNA-4157 in combination with Pembrolizumab treatment demonstrated a 49% reduction in the risk of mortality among high-risk melanoma patients.176 TSAs are promising targets for cancer vaccines.178 However, only a small fraction of somatic mutations in cancer cells could be recognized by spontaneously occurring T cells, and the efficacy of these neoepitopes in mediating antitumor effects varies, posing challenges for their precise utilization.173
To address the challenge of limited availability of targetable neoantigens and antigen target loss in tumors, it is imperative to investigate and utilize the full spectrum of tumor antigens, to develop personalized cancer vaccines. Trivedi et al.179 established an immunogenomics pipeline called “Open Reading Framework Antigen Network (O.R.A.N.)” to effectively identify immunogenic antigens with high likelihood of becoming therapeutic targets. In addition, they developed a platform called “Tumor Open reading Frames that are Unique (TOFU)” which utilizes IVT mRNA technology for encoding multiple tumor antigens into a single mRNA vaccine, thereby enabling the customization of virtually limitless quantities of antigens specific to each tumor type. Ben-Akiva et al.180 developed a class of bioreducible nanocarriers utilizing lipophilic poly(beta-amino ester) to encapsulate antigen-encoding mRNA and Toll-like receptor (TLR) agonist adjuvants for the treatment of murine melanoma and colon adenocarcinoma. Due to its mRNA sequence-independent encapsulation capacity, this platform can also be extended to address other cancer diseases.
The standalone mRNA cancer vaccine may serve as an effective treatment option for early-stage cancer. However, in the case of advanced cancer patients, the tumor microenvironment often exhibits a high level of immune suppression, thereby diminishing the efficacy of mRNA monotherapy.181 Instead, combining therapeutic cancer mRNA vaccines with other immunotherapies,182 such as the combination of mRNA-4157 and Pembrolizumab to treat high-risk melanoma, holds promise for success.183
Despite the fact that mRNA vaccines encoding tumor antigens entered Phase I trials earlier than viral mRNA vaccines, it is disappointing that few mRNA vaccines against cancer seem to have advanced to Phase III or IV trials (Fig. 3b).184 The reasons are summarized in “Conclusions and prospects” section. We anticipate an influx of approved drugs and methodologies for cancer treatment, instilling renewed optimism in patients.
Bacterial antigen mRNA vaccine
The conventional approach to eradicating bacteria involves the administration of antibiotics. However, due to the overuse of these drugs, we have been alarmed by the emergence of antibiotic-resistant bacteria and even the development of highly resistant strains. Nevertheless, it is important to note that the research and approval process for creating a new antibiotic is exceedingly time-consuming and necessitates substantial financial resources. Regrettably, bacteria can develop resistance at a much faster rate than new drugs can be developed. Consequently, despite our awareness regarding the imperative need for antibiotics control measures, we may find ourselves in a situation where no effective drug exists to combat bacterial infections. Therefore, it is essential to propose an innovative antimicrobial therapy. Given that mammalian organisms are capable of translating mRNA-encoding bacterial proteins, mRNA vaccines offer a promising and efficacious avenue for preventing bacterial diseases.185,186
Lyme disease is a prevalent infectious ailment in the U.S. that currently lacks a specific vaccine. Among various candidates, outer surface protein A (OspA) that expressed on the pathogen Borrelia burgdorferi has emerged as most promising antigen for developing a platform to combat Lyme disease. Therefore, Pine et al.187 developed a mRNA LNPs encoding OspA and observed that its protective efficacy surpassed that of the OspA protein subunit vaccine. The treatment regimen of mRNA-1975 and mRNA-1982 developed by ModernaTX, Inc. for Lyme disease is currently in Phase I/II clinical trial (NCT05975099).188
As a significant infectious disease, Tuberculosis (TB), caused by Mycobacterium Tuberculosis (M. TB),189 continues to present a grave global public health menace. Currently, the only licensed vaccine for TB is Bacillus Calmette–Guerin (BCG), which exhibits limited efficacy in preventing the disease.190 The absence of effective vaccine strategies and the emergence and dissemination of multidrug-resistant TB (MDR-TB) bacteria have further complicated endeavors to prevent and treat TB. Similar to COVID-19, TB also represents a respiratory infection. Can the mRNA vaccine strategy successfully employed for COVID-19 be replicated for TB?191 Fortunately, in 2023, BioNTech SE commenced Phase I/II clinical trial (NCT05547464) for an mRNA approach targeting TB (BNT164a1 and BNT164b1),185,192 and we expect a satisfactory therapeutic efficacy from this strategy.
Currently, the application of antibacterial mRNA vaccines in preclinical and clinical attempts is limited, possibly due to the intricate nature of bacterial infections compared to viral infections. Bacterial infections exhibit distinct molecular expression characteristics at different stages of infection, making it virtually impossible for a single antigen to effectively cover all these stages.193 Moreover, bacterial pathogens display diversity and the variability in antigen expression while also evolving multiple mechanisms to evade or suppress immune responses.185 Therefore, if mRNA vaccines are intended for further application in preventing and treating bacterial infections, careful consideration must be given to rational antigen selection along with the incorporation of adjuvants and other optimization strategies.193 The research conducted by Meulewaeter et al.194 demonstrated that the incorporation of adjuvant αGC into the Listeria monocytogenes mRNA LNPs resulted in a synergistic protective effect against listeriosis, underscoring the therapeutic advantages of co-activating invariant natural killer T (iNKT) cells in antibacterial mRNA vaccines. We are looking forward to the successful development of the first antibacterial mRNA vaccine product.
Parasite antigen mRNA vaccine
In addition to viral infections, parasitic diseases are also infectious illnesses that debilitate individuals. Parasitic diseases can be generally categorized into three groups: ectoparasitic, protozoal, and helminthic diseases. These conditions can be managed through proactive vector control, disease monitoring, the use of effective vaccines/medications, and the implementation of appropriate sanitation infrastructure.195 However, the lack of public awareness and the absence of appropriate treatment options have resulted in widespread parasitic infections.196
Malaria is the most deadly parasitic vector-borne disease197 caused by at least five species of Plasmodium parasites.198–200 Over the years, researchers have endeavored to develop vaccines targeting different stages of the malaria parasite’s life cycle. However, the only approved vaccine to date is Mosquirix, which is based on recombinant proteins. Nevertheless, Mosquirix has encountered several limitations, including suboptimal efficacy in specific age groups and the necessity for multiple booster shots to achieve a satisfactory level of protection. Fortunately, the mRNA vaccine (BNT165b1) developed by BioNTech, which encodes a segment of the Plasmodium falciparum circumsporozoite protein (PfCSP), has progressed to Phase I clinical trial (NCT05581641).201 Kunkeaw et al.202 developed the LNPs platform to deliver nucleotide-modified mRNA encoding the Pvs25 antigen of Plasmodium vivax, a potential vaccine against malaria transmission. Pvs25 is an antigen candidate for blocking malaria transmission. They discovered that the mRNA LNPs platform could elicit stronger and more enduring functional immunity compared to the Pvs25 recombinant protein vaccine. Additional mRNA vaccines against malaria are currently undergoing preclinical research.203–205
The lack of interest in the commercial development of parasite vaccines may be attributed to the fact that parasitic diseases primarily affect economically disadvantaged populations, and most parasites cause chronic illnesses that are non-lethal to the host, with the exception of the deadly malaria. Parasite infections are intricate, and the development of conventional vaccines entails high costs.206 In these regards, mRNA-based vaccines present a promising approach to address these challenges.
Autoimmune disease antigen therapy
The immune system plays a critical role in defending the body against infections caused by viruses, bacteria, fungi, and parasites. Additionally, it surveils and eliminates cancerous cells to maintain the body’s health. Autoimmune disorders arise when the immune system erroneously targets healthy tissues and cells in the body.207,208 An essential characteristic of autoimmune diseases is the presence of autoantibodies that have the ability to specifically bind to the antigens present in the patient’s healthy cells or tissues.207,209 There are more than 80 autoimmune disorders, including multiple sclerosis (MS),210 type 1 diabetes (T1D),211 systemic lupus erythematosus (SLE),212 inflammatory bowel disease (IBD),213 and rheumatoid arthritis (RA).214 The objective of treating autoimmune diseases is to regulate the activity of autoreactive cells in the body without inducing systemic immune suppression.215
Krienke et al.210 utilized a non-inflammatory lipid complex carrier to encapsulate N1-methyl-pseudouridine-modified mRNA (m1Ψ mRNA) encoding disease-related autoantigens. Their findings indicated that the use of this m1Ψ mRNA vaccine could stimulate antigen-specific regulatory T cells (Tregs) and mitigate the severity of multiple sclerosis in a mouse model.216 This strategy seems capable of inducing cross-tolerance in autoimmune T cells while preserving the normal immune response of the immune system.217 This also implied that by reducing unnecessary immunogenicity, mRNA vaccines encoding autoantigens can be a means to induce and sustain natural peripheral immune tolerance.218
Type 1 diabetes is an autoimmune condition in which the immune system mistakenly attacks and destroys the pancreatic islet β-cells, resulting in insufficient insulin production.211,219 Foster et al.220 discovered that the utilization of a novel mRNA vaccine encoding proinsulin II could transiently delay diabetes symptoms in nonobese diabetic mice. This effect may be attributed to the ability of the mRNA vaccine to restore immune tolerance in nonobese diabetic mice.
The mRNA therapies hold significant promise in the realm of autoimmune diseases.221,222 For instance, the mRNA drugs can be tailored to encode proteins or peptides capable of binding and neutralizing autoantibodies. In addition, mRNA can be engineered to specifically target plasma cells and express pro-apoptotic molecules within them, thereby selectively disrupting or eliminating their function and survival. Furthermore, given the pivotal role of Tregs in suppressing immune responses, mRNA can be designed to bolster the functionality of Treg cells.207
Therapeutic antibody therapy
Antibodies, also referred to as immunoglobulins, are effector molecules generated by the humoral immune responses that can neutralize antigens223 and play a crucial role in the prevention, control, and elimination of infections.224 The emergence of the therapeutic antibody can be traced back to 1975, when Köhler and Milstein pioneered the hybridoma technology and laid the foundation for the progress of monoclonal antibodies (mAbs).225 Currently, therapeutic antibodies have emerged as a potent armamentarium against a variety of diseases,223 demonstrating remarkable efficacy in treating viral illnesses, solid tumors, hematological malignancies, autoimmune disorders, and other indications. In the last few decades, antibodies have experienced unprecedented growth as new drugs for these indications.226
However, protein-based drugs such as mAbs encounter challenges in terms of production and cost, including intricate manufacturing process, high purification expenses, and difficulties in post-translational modification of proteins. In addition, the human body harbors a total of 22 distinct amino acids, and due to variations in their types, quantities, and arrangements, proteins can exhibit diverse physicochemical properties. To ensure protein storage stability, specialized buffer solutions optimized for this purpose are indispensable.227 The nucleic acids, in contrast, carry a negative charge and exhibit similar physicochemical characteristics. Therefore, the production and purification process of nucleic acid drugs does not necessitate specific customization, and it is easy to obtain the natural structure of the desired protein owing to the target cell-endogenous expression mechanisms.227
Delivering mRNA into the body for translation into antibodies, rather than directly administering the mAbs, circumvents the hassles associated with protein production and purification. Moreover, this approach enables the encoding of proteins that are challenging to synthesize in vitro. Deal et al.228 engineered an mRNA sequence encoding the pathogen-specific immunoglobulin A (IgA) mAbs, which was encapsulated in LNPs and delivered to mucosal secretions. Intriguingly, the pharmacodynamics of lgAmRNA encoded by this mRNA closely resemble those of endogenous human lgA rather than recombinant IgA (IgAR). These findings suggested that the mRNA antibody technology held promise for intercepting pathogens on mucosal surfaces, offering a novel and effective approach for the prevention and treatment of diseases.
The intravenous administration of anti-vascular endothelial growth factor (VEGF) antibodies typically results in their predominant distribution in the vasculature and interstitial spaces, with limited accumulation in the target organ—the lungs. This limited localization impairs their therapeutic efficacy for non-small cell lung cancers (NSCLCs) and may have adverse effects on normal tissues. A team led by Le et al.229 developed a pulmonary-targeting nanoparticle for delivering mRNA-encoding Bevacizumab, an anti-VEGF antibody widely used in the clinic. These nanoparticles utilized the inherent properties of poly(beta-amino esters) (PBAEs) to achieve their pulmonary-targeting function. This innovative approach effectively suppresses angiogenesis in NSCLCs, offering a promising therapeutic strategy for the treatment of this disease.
The Orthopoxvirus genus, including MPXV and variola virus (VARV), constitutes a significant global health threat that causes severe pox diseases in both humans and animals. Chi et al.230 constructed mRNA combinations encoding four mAbs (mAbs 22, 283, 26, and 301) specifically designed to target and neutralize vaccinia virus (VACV) A33, VACV B5, MPXV M1 and VACV A27 proteins. This was the first application of mRNA antibodies against Orthopoxvirus in vivo. The results indicated that a single injection of LNPs encapsulated with mRNA in mice could rapidly induce the production of corresponding neutralizing antibodies, which demonstrated significant protective effects in the VACV lethal challenge mouse model and reduced mortality rates.
In the last decade, there have been significant advancements in the treatment of advanced cancers with immune checkpoint inhibitors (ICIs). Currently, clinically approved ICIs consist of innovative medications that target programmed cell death protein-1 (PD-1), programmed death ligand-1 (PD-L1), cytotoxic T- lymphocyte-associated protein 4 (CTLA-4), and lymphocyte activation gene-3 (LAG-3).231,232 Nevertheless, ICIs still possess certain limitations, such as the potential for immune-related adverse events.233,234 In order to address these safety concerns, researchers have explored the use of mRNA therapies encoding ICIs.235 Pruitt et al.236 transfected DCs with mRNA encoding the heavy and light chains of anti-CTLA-4 and anti-GITR mAbs, as well as mRNA-encoding tumor antigens. Their findings demonstrated that these mRNA-engineered DC cells significantly augmented antitumor immunity in melanoma-bearing mice without eliciting any signs of autoimmunity. Utilizing this technology, a Phase I clinical trial for patients with metastatic melanoma is presently under assessment (NCT01216436). The IVT mRNA platform also enables the continuous endogenous synthesis of bispecific antibodies that bind to T cells. Zeng et al.237 established and optimized an mRNA sequence called Z15-0-2 that codes for a bispecific nanobody of anti-PD-1 and anti-CTLA-4. Moreover, there are mRNA-based therapeutics encoding anti-PD-L1 mAb for cancer treatment238 and mRNA-6981 encoding PD-L1 for autoimmune diseases.239
Rituximab is a human/murine chimeric glycosylated IgG1-κ mAb with specific affinity for the transmembrane protein CD20 on B lymphocytes.240 Thran et al.241 developed LNPs formulations containing mRNA-encoding Rituximab (anti-CD20 mAb) to combat B-cell lymphoma. They observed that the mRNA LNPs treatment exhibited a pronounced and potent antitumor effect comparable to that of recombinant antibody therapy.79 The mRNA drugs encoding anti-CD20 mAb may also offer promising therapeutic options for autoimmune conditions such as multiple sclerosis and rheumatoid arthritis.207 Antibody-based mRNA therapies hold significant potential for the treatment of tumors, chronic inflammation, and autoimmune disorders.242
Cytokine
Theoretically, mRNA has the potential to encode any protein, including cytokines that can be used as standalone therapies or in combination with other treatment modalities for disease management.243
Cytokines play a pivotal role in regulating various biological processes such as cell survival, proliferation, differentiation and immune cell activity by facilitating crucial intercellular communication within short distances.244 The potential of cytokines in the realm of antitumor research is exemplified by their capacity to impede the proliferation of tumor cells, facilitate the induction of apoptosis, and modulate both innate and adaptive immunity.245 Furthermore, cytokines may exhibit immunosuppressive effects, offering a potential therapeutic approach for conditions derived from hyperactive immune responses such as autoimmune diseases. The cytokine subgroups commonly encompass IL, interferons (IFNs), chemokines, granulocyte-macrophage colony-stimulating factor (GM-CSF) and so on.244
Immune-potentiating cytokine
IL-2, IL-21, and IL-15: The FDA granted approval for the use of recombinant human IL-2 in 1992 for the treatment of metastatic renal cancer, and in 1998 for the management of metastatic melanoma.246 However, intravenous administration of IL-2 exhibits limited tumor accumulation247 and possesses a short half-life (only 5 to 7 min). In order to achieve enhanced antitumor effects, administration of high doses of IL-2 is imperative, leading to elevated concentrations of IL-2 in the bloodstream and subsequent manifestation of adverse reactions in patients, including asthenia, pyrexia, and potentially life-threatening toxicity such as capillary leak syndrome.248 To mitigate the systemic toxicity associated with IL-2 protein delivery, Shin et al.249 proposed a novel approach involving intratumoral administration of IL-2 mRNA NPs. These NPs are composed of polyethyleneimine-modified porous silica and effectively minimize off-target translation of mRNA. Jiang et al.250 developed mRNA sequences encoding fusion proteins consisting of IL-2, CD25 (IL-2Rα), and a cleavable linker, which were encapsulated within ionizable lipid U-101–derived NPs. The designed linker was susceptible to cleavage by tumor-specific matrix metalloproteinase-14 (MMP-14). Furthermore, the researchers observed that U-101–derived NPs exhibited superior transfection efficacy compared to approved ALC-0315–LNPs, thereby contributing to the platform’s ability to achieve enhanced antitumor efficacy with reduced toxicity. Beck et al.251 combined long-lasting mRNA-encoded IL-2 with tumor-targeting mAb therapy, thereby inducing a highly pro-inflammatory TME and overcoming tumor resistance resulting from the absence of MHC I.
In addition to IL-2, other cytokines that belong to the gamma-chain receptor family include IL-4, IL-7, IL-15, IL-21.252 The role of IL-21 in the clearance of hepatitis B virus (HBV) is significant. The mRNA LNPs encoding IL-21, delivered by Shen et al.253, demonstrated remarkable efficacy in inducing effective clearance of HBV and its covalently closed circular DNA (cccDNA).
IL-15 shares a structural resemblance with IL-2254 and plays a crucial role in the regulation of both innate and adaptive immunity.255 IL-15 is widely acknowledged as a T-cell growth factor that governs the activation, proliferation, and cytotoxicity of T cells, B cells, and natural killer (NK) cells, thereby enhancing the production of IFN-γ and TNF-α. Moreover, due to its unlikely induction of Treg cell activity during immune stimulation,254 IL-15 has been regarded as a promising candidate in oncology.255 Nevertheless, IL-15 is associated with significant drawbacks such as severe toxicities and a short half-life. These challenges can be addressed through efficient delivery of mRNA-encoding IL-15 to the tumor. Wang et al.256 designed a nanoplatform to specifically target PD-L1 for the delivery of IL-15 mRNA, and integrated it with an ultrasound-targeted microbubble destruction therapy approach. This therapeutic approach not only effectively facilitates the expression of IL-15, but also significantly enhances intracellular reactive oxygen species (ROS) levels, induces immunogenic cell death (ICD), thereby effectively suppressing tumor growth and recurrence. Furthermore, this integrated therapy offers real-time ultrasound imaging guidance. Gavigan et al.257 developed a combination therapy comprising IFNα, IL-7, IL-15, and a TNF receptor superfamily agonist. Their findings demonstrated that co-delivery of mRNA mixture encoding IFNα, IL-7, a protein fused with IL-15 and its receptor α chain, and OX40L could elicit potent antitumor immune responses in mice.
IL-12 and IL-27: Liu et al.258 endeavored to synergize IL-12 mRNA with Oxaliplatin prodrug for the management of colorectal cancer (CRC). Hewitt et al.259 discovered in preclinical investigations that intratumoral administration of LNPs loaded with mRNA-encoding IL-12 could augment the antitumor effects by inducing the release of IFN-γ and promoting the transformation of Th1 cells within the TME. The Phase I trial of human IL-12 mRNA (MEDI1191) has been successfully completed (NCT03946800). To further enhance tumor infiltration of IL-12 and mitigate systemic toxicity caused by its extratumoral presence, Trani et al.260 engineered mRNAs encoding single-chain IL-12 along with two additional components: a rat anti-mouse AFS98 mAb variable region that targets colony-stimulating factor-1 receptor (CSF1R) and the variable region of Avelumab (anti-PD-L1 antibody), which significantly augment post-transcriptional binding of IL-12 to the tumor.
IL-27, belonging to the IL-12 family, exhibits comparable antitumor effects as IL-12. The activation of T cells and NK cells by IL-12 and IL-27 occurs through distinct mechanisms, whereby IL-12 activates T cells via the signal transducer and activator of transcription (Stat) 4 pathway, while IL-27 engages T cells through both the Stat1 and Stat3 pathways. Liu et al.261 developed LNPs loaded with mRNA and observed that co-delivering IL-12 + IL-27 mRNAs using the same LNPs carrier resulted in superior tumor suppression compared to the single delivery of IL-12 mRNA. Furthermore, the incorporation of IL-27, an anti-inflammatory cytokine, could mitigate potential toxic inflammation associated with mono-administration of IL-12. This combination not only enhanced efficacy but also reduced systemic toxicity.
GM-CSF: GM-CSF is a cytokine that promotes the generation of various myeloid cell subpopulations, such as DCs, monocytes, macrophages, and neutrophils. Excessive or inadequate levels of GM-CSF could enhance the malignancy of cancer.262 The mRNA mixture (SAR441000) encoding IL-12, IFN alpha-2b, GM-CSF, and IL-15 was administered intratumorally as a monotherapy or in combination with Cemiplimab during the Phase I trial (NCT03871348) conducted in 2019 for the treatment of advanced solid tumors. The results demonstrated favorable tolerability and discernible immunomodulatory effects for both SAR441000 monotherapy and its combination with Cemiplimab.263
To extend the serum half-life of GM-CSF, it can be conjugated with biologically safe and structurally stable proteins. Yeapuri et al.264 developed mRNA LNPs encoding the albumin-GM-CSF fusion protein, thereby prolonging the half-life of the target protein and enhancing drug bioavailability to optimize its therapeutic efficacy.
Immune-suppressing cytokine
IL-2: Although mRNA-based cytokine therapies are commonly employed in the field of antitumor research, their potential applications in the treatment of other diseases should not be underestimated. The IL-2 receptor exists in two forms with varying affinities, the form expressed by Treg cells exhibiting a higher affinity for IL-2. Consequently, at low concentrations, IL-2 preferentially activates Tregs.118,119 However, at higher concentrations, it also triggers the activation of pro-inflammatory T cells and NK cells. Treg cells represent a subset of CD4+ T lymphocytes with significant immunosuppressive capabilities.265 Impairments in the function of Treg cells or a deficiency in their numbers can result in the breakdown of immune tolerance and the development of autoimmune diseases within the body.266 Consequently, induced Treg cells hold promise for the treatment of autoimmune diseases and the restoration of immunological self-tolerance.267,268 Picciotto et al.118 engineered mRNA to encode a fusion protein of human serum albumin IL-2 mutein (HSA-IL2m) with an extended half-life. Their findings demonstrated that delivering this lipid-encapsulated mRNA could selectively expand Tregs in vivo without activating NK cells or conventional T cells. This approach offers a novel strategy for suppressing autoreactive T cells and restoring immune balance in autoimmune diseases. The mRNA-6231, encoding HSA-IL2m, has already progressed to Phase I clinical trial (NCT04916431).221
IL-4: IL-4 is involved in the regulation of the activation, proliferation, differentiation, and survival of various T-cell subtypes.269 It supports the differentiation of Th2 cells and antagonizes the function of Th1 cells associated with numerous autoimmune disorders.270 The modulation of Th-cells subgroups, particularly the balance between Th1 and Th2, exerts a significant influence on the pathogenesis of type 1 diabetes.271,272 Creusot et al.270 reported a novel therapy that delivered translationally enhanced IL-4 mRNA into DCs using electroporation. They have shown that modified DCs containing mRNA could modulate autoimmune diabetes in nonobese diabetic mice by boosting the functionality of Treg cells and inducing a shift in the proportion of Th cell populations.
IL-10: IL-10 is a cytokine with anti-inflammatory properties, capable of suppressing the expression of pro-inflammatory cytokines and inhibiting the presentation of alloantigens by APCs.273 Chen et al.274 developed a novel delivery system with self-protection and active targeting capabilities for encapsulating mRNA encoding IL-10. Nucleic acids were combined with polyphenols to mitigate enzymatic degradation. Due to the susceptibility of hyaluronic acid to oxidation in the inflamed colon and subsequent inactivation, it is essential to combine bilirubin, known for its potent antioxidant activity, with hyaluronic acid in order to ensure effective targeting of CD44. They demonstrated that rectal administration of the IL-10 mRNA delivery system could effectively induce therapeutic effects in a murine model of colitis. Atherosclerosis is a chronic inflammatory condition. Bu et al.275 developed an exosome-based delivery platform for loading engineered IL-10 mRNA. They observed that this platform could effectively mitigate atherosclerosis in ApoE−/− (Apolipoprotein E-deficient) mice.
Ligand
The mRNA can also encode immune-stimulating ligands, thereby facilitating an immunogenic response. The TriMix therapy consists of a combination of mRNA encoding three distinct immune-stimulating proteins, namely CD40 ligand (CD40L), CD70, and constitutively active TLR4.276 This innovative mRNA-based vaccine can be administered via intratumoral injection for patients with early-stage, resectable breast cancer, which is currently undergoing Phase I clinical trial (NCT03788083).
The TriMix compound not only serves as a direct mRNA vaccine but also functions as an adjuvant in immunization research due to its capacity to encode DC-activating proteins. The ECI-006 vaccine consists of a combination of mRNA-encoding adjuvant proteins (TriMix) and mRNA-encoding melanoma-specific TAAs (tyrosinase, gp100, MAGE-A3, MAGE-C2, and PRAME). A Phase I clinical trial has been conducted to evaluate this mRNA vaccine in patients with stage IIc/III/IV melanoma who have undergone surgical resection (NCT03394937). The results demonstrated favorable tolerability profiles when administering ECI-006 at doses of 600 or 1800 μg to the patients, and immunogenicity was observed in a subset of patients.277
Tumor suppressor protein
The optimal goal of cancer treatment is to eradicate tumor cells while minimizing the harm to healthy tissues and cells. In pursuit of this objective, suicide gene therapy, a novel approach to cancer treatment, has garnered attention.278 Cancer suicide gene therapies can be categorized into three groups: enzyme/prodrug systems, pro-apoptotic genes, and toxins.279 The enzyme/prodrug system involves the transfer of genetic material encoding enzymes to tumor cells, leading to the conversion of nontoxic prodrugs by these enzymes into cytotoxic metabolites that induce apoptosis in tumor cells.278,280 For instance, the innovative human herpes simplex virus–thymidine kinase (HSV-TK)/ganciclovir suicide gene system shows promise. However, the HSV-TK system is hampered by significant limitations associated with its immunogenicity.281 Furthermore, enzyme/prodrug systems also encounter the challenge of off-target effects on normal tissues, resulting in toxicity, as well as suboptimal efficacy against slow-dividing tumor cells. To address these challenges, the approach of developing genes encoding pro-apoptotic proteins and bacterial toxins as suicide genes was employed.282
Enzyme-encoding mRNA for enzyme/prodrug systems
The combination of cytosine deaminase and uridine phosphoryltransferase (UPRT)/5-fluorocytosine (5-FC) forms an enzyme/prodrug system. The cytosine deaminase is capable of converting the nontoxic 5-FC into the toxic metabolite 5-fluorouracil (5-FU). The gene encoding cytosine deaminase is commonly fused with the UPRT gene to enhance its efficacy.279 Mizrak et al.283 obtained microvesicles (MVs) loaded with cytosine deaminase-UPRT mRNA/protein by transfecting the cytosine deaminase-UPRT-EGFP plasmid into HEK-293T cells. Subsequently, they observed that intratumoral injection of cytosine deaminase-UPRT-mRNA/protein-containing MVs and treatment with 5-FC significantly suppressed tumor growth in tumor-bearing mice.
Inducible caspase-9 (iC9) is an engineered version of caspase-9 that can be dimerized and activated upon exposure to the chemical inducers of dimerization (CID), such as the AP1903 molecule, subsequently triggering downstream apoptotic pathways and leading to cell death.284 Nakashima et al.285 developed LNPs carrying iC9 mRNA. They discovered that the combination of iC9 mRNA and CID exhibited cytotoxic effects on three breast cancer cell lines in vitro. On the other hand, Saito et al.286 engineered liposomes for the delivery of iC9 mRNA and combined them with CID for the treatment of T-cell malignancies. It was found that the treatment regimen exhibited anti-leukemia therapeutic effects both in vitro and in vivo.
Pro-apoptotic protein
p53 and PTEN: It is widely acknowledged that cancer originates from the mutation of somatic cells, and that oncogenic mutations or abnormal expression of cellular proto-oncogenes can trigger carcinogenesis. A distinct category of tumor suppressor gene can counteract these carcinogenic effects and impede tumor development. Notably, p53, retinoblastoma (Rb), and phosphatase and tensin homolog deleted on chromosome ten (PTEN) are three pivotal tumor suppressor proteins with interconnected functions.287 The majority of human cancers exhibit loss-of-function mutations in the p53 protein. However, due to the fact that p53 is a transcription factor, it has long been deemed undruggable.288 Nevertheless, the emergence of mRNA therapies has facilitated the development of numerous p53-based cancer treatments utilizing mRNA.
To augment the therapeutic efficacy of p53 mRNA therapy, it can be synergistically combined with other well-established antitumor modalities. Zhang et al.289 developed paclitaxel amino fat-derived NPs for encapsulating p53 mRNA and chemotherapy drugs, demonstrating remarkable antitumor effects in an orthotopic triple-negative breast cancer (TNBC) mouse model. Zhou et al.290 were the pioneers in reporting the integration of mRNA therapy with photodynamic therapy (PDT) for tumor treatment. They developed a ROS-responsive NPs platform encapsulating p53 mRNA and indocyanine green (ICG). These NPs can be triggered by ROS to release p53 mRNA, thereby augmenting p53 expression and inducing apoptosis in lung cancer cell. Simultaneously, laser irradiation could activate the released ICG to facilitate PDT. To enhance the expression of intracellular target proteins, Cao et al.291 employed a novel strategy by designing the CRISPR/dCasRx-SINEB2 platform to augment the translation of endogenous mRNA-encoding tumor suppressor proteins (including PTEN and p53), thereby inducing suppressed proliferation and increased apoptosis in bladder cancer cells. Liu et al.292 established a nanoparticle platform to encapsulate PTEN mRNA for brain delivery. The NPs exhibited enhanced capability to cross the blood-brain barrier and target tumor cells, demonstrating the potential of using mRNA-based technologies for treating brain diseases.
Bim and Bax: Bim is a significant pro-apoptotic member within the Bcl-2 protein family.293 Gao et al.294 developed a delivery system that combined cell-penetrating peptides (CPPs), cRGD-R9, and cationic nano-sized DOTAP-mPEG-PCL scaffolds to deliver mRNA-encoding Bim. The Bim mRNA delivery system exhibited potent inhibitory effects on tumors in various colon cancer models through the induction of mitochondrial apoptosis.
Bax is a pro-apoptotic molecule that can serve as a tumor suppressor protein. Okumura et al.295 developed liposomes to deliver mRNA-encoding Bax and compared the antitumor effects with those of a Bax plasmid in tumor-bearing mice. Their findings indicated that the use of Bax mRNA therapy via liposomes exhibited stronger antitumor effects than the Bax plasmid.
TRAIL: Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) can trigger apoptotic cell death in tumor cells while sparing normal cells. Gu et al.296 developed mRNA-based LNPs encoding a fusion protein of TRAIL and the receptor-binding domain (RBD) from the SARS-CoV-2 spike protein, which were administered via intratumoral injection in colon cancer mice models. They observed that the RBD-TRAIL mRNA LNPs effectively inhibited tumor growth in mice. Silva et al.297 discovered that the direct intratumoral injection of LNPs loaded with TRAIL mRNA could effectively induce tumor cell death, particularly when combined with Losartan or angiotensin 1–7.
Apoptin: Apoptin is a protein encoded by the gene of the chicken anemia virus.298 It is capable of inducing apoptosis in tumor cells through a p53-independent pathway.299 It is noteworthy that Apoptin can trigger apoptosis in tumor cells while sparing normal diploid cells.298 Therefore, as a tumor suppressor protein, Apoptin holds promise as a safe anticancer medication.300 However, the collection and purification process of this protein is highly intricate and costly.299 Therefore, the gene therapy being developed for antitumor Apoptin primarily relies on DNA301 or viral vectors.298 Tang et al.299 engineered nanospheres with self-assembly capabilities to encapsulate mRNA-encoding Apoptin. The combination of Apoptin mRNA with Doxorubicin exhibited remarkable synergistic antitumor effects.
The mRNA-based therapy that encodes tumor suppressor proteins is currently in its early stages of development. Ongoing explorations are being conducted regarding numerous oncogenes and their corresponding tumor suppressor proteins. Qu et al.302 proposed that Huntingtin-associated protein-1 (HAP1) may function as a potential tumor suppressor in gastric cancer. Zheng and Song303 believed that Synaptopodin 2 (SYNPO2) functions as both a structural protein in muscle tissue and an emerging tumor suppressor protein, supported by its positive correlation with favorable cancer prognosis, thereby indicating its crucial role in cancer prevention and treatment. With the identification of additional tumor suppressor proteins, it is anticipated that mRNA-based therapies encoding these proteins will gradually emerge as a promising avenue in cancer research.
Immunotoxin
Immunotoxins are typically chimeric proteins composed of a protein toxin conjugated to an antibody fragment,304 enabling specific targeting and elimination of antigen-bearing cells.305 The toxic components in immunotoxins typically originate from a variety of sources, including bacteria, plants, and human cells. Bacterial-derived toxins such as diphtheria toxin and pseudomonas exotoxin A (PE), as well as plant-derived toxins like ricin and gelonin, are commonly used. Human-derived toxins include granzymes and RNases. Among these, the application of bacterial and plant toxins is limited due to their potential immunogenicity in humans.306 Various strategies have been developed to mitigate the immunogenicity of immunotoxins. These include employing immunosuppressive therapy, utilizing human-derived toxin proteins, and modifying toxin moiety to evade detection by the human immune system.307
The short half-life of the immunotoxin in the circulatory system may limit its therapeutic activity. To extend the half-life of immunotoxin in circulation, Guo et al.308 fused it with the albumin-binding domain. The production of recombinant immunotoxins is typically conducted in bacteria. However, it is generally challenging to produce soluble proteins with molecular weights exceeding 80 kDa in bacterial systems.309 mRNA-based immunotoxin therapy presents a promising alternative.
The application of immunotoxins for the treatment of cancer is currently under investigation, which is, however, constrained by their large molecular weight, limited tumor penetration, and the presence of cellular resistance. Consequently, immunotoxins have not yet received approval for clinical use against solid tumors. To address these challenges, Granot-Matok et al.310 developed a LNPs system that delivered modified mRNA encoding the PE domain. This innovative approach employed toxin-encoding mRNA to overcome the limitations associated with immunotoxin therapy, offering improved safety and enhanced therapeutic efficacy.
PE38, a derivative of the potent toxin PE produced by Pseudomonas aeruginosa, has been engineered to remove its targeting domain while retaining its toxicity. The toxicity of PE lies in its specific inhibition of elongation factor 2 (EF-2) in the cytosol of eukaryotes. Eggers et al.311 utilized PE38 in the development of immunotoxins by fusing it with human VEGF and an anti-human epidermal growth factor receptor-2 (HER2)/neu single-chain variable fragment (scFv). They transfected human primary T cells with mRNA-encoding immunotoxins and demonstrated that the engineered T cells displayed significantly enhanced cytotoxicity against cancer cells in vitro when combined with bispecific antibodies.
The subtilase cytotoxin, diphtheria toxin, and abrin-a belong to the AB toxin family. Their B-subunit initially binds to the target cell, followed by the entry of the A-subunit into the cell, where it exerts toxicity by inhibiting protein synthesis, ultimately leading to the demise of the target cell. Hirschberger et al.312 developed three chemically modified mRNAs encoding subtilase cytotoxin, diphtheria toxin, and abrin-a. They demonstrated that the mRNA therapy encoding abrin-a exhibited superior efficacy in reducing the viability of tumor cells in vitro compared to the other two mRNA therapies.
Furthermore, immunotoxins offer a promising therapeutic approach for antiviral treatment. Their mechanism involves targeting and eradicating infected cells, rather than the inhibition of viral or cellular pathways essential for virus replication and spread.313 There is great potential for achieving significant advancements in the fields of antitumor and antiviral researches, among others, through the application of immunotoxins.
Adoptive cell therapy
Adoptive cell therapy involves the extraction of the patient’s immune cells, followed by ex vivo editing and activation, and subsequent reintroduction into the patient for disease treatment. This approach aims to enhance the identification and elimination of tumor cells. Additionally, there is ongoing development of in situ adoptive cell therapy.314 It is noteworthy that the subsequent adoptive cell therapies are founded upon RNA methods, as opposed to conventional DNA methodologies. Specifically, CAR-T therapy should be further subdivided into RNA chimeric antigen receptor T cells (rCAR-T).
CAR-T
In the past decade, chimeric antigen receptor (CAR)-expressing T-cell (CAR-T-cell) therapy has made major breakthroughs in the treatment of hematological cancers. Disappointingly, its effectiveness in treating solid tumors is still limited due to a lack of comprehensive understanding of tumor-specific antigens, the suppression of T-cell infiltration and killing ability by the TME, and limited expansion potential of T cells.315 Therefore, Mackensen et al. designed CAR-T cells targeting the tumor-specific antigen Claudin 6 (CLDN6) and demonstrated potential clinical activity. However, due to the limited durability of a single dose, they integrated it with a CAR-T-cell Amplifying RNA Vaccine (CARVac) for administration. This combination approach exhibited good tolerability in subjects and showed promise of enhancing CAR-T-cell expansion and improving tumor treatment efficacy. Currently, the study is in Phase I/II clinical trial (NCT04503278).120,316 Despite promising results observed in clinical trial, the findings fell short of meeting expectations, underscoring the urgent challenges that still need to be addressed in targeting CLDN6 CAR-T therapy combined with CARVac administration.315 The methodology employed in this study for CAR-T-cell production aligns with clinical practice, encompassing ex vivo culturing of CAR-T cells followed by their infusion into patients. This approach entails significant expenditures for manufacturing. Hence, there is an urgent imperative to develop a swift, robust, and economically efficient alternative strategy for manufacturing CAR-T cells. A promising avenue involves the direct delivery of mRNA into the body to induce in situ formation of CAR-T cells.
Despite the challenges associated with transfecting exogenous mRNA into T cells, Tombácz et al.317 discovered that the application of CD4 antibody-conjugated mRNA LNPs encoding Cre recombinase or firefly luciferase significantly augmented the signal of mRNA by ~30-fold in spleen T cells and increased the expression of reporter genes in CD4+ T cells in vivo when compared to non-targeted mRNA LNPs. Zhou et al.318 used CD3 antibody-modified LNPs loaded with IL-6 short hairpin RNA (IL-6 shRNA) and CD19-CAR (based on plasmid DNA) for targeted delivery to CD3+ T cells, which resulted in the downregulation of IL-6 within CAR-T cells and a reduction in cytokine release syndrome (CRS), thereby enhancing the safety profile of leukemia treatment. Rurik et al.319 developed CD5-targeted LNPs encapsulating mRNA encoding a CAR against fibroblast activation protein (FAP). By administering these CD5-targeted LNPs, potent anti-fibrotic CAR-T cells could be generated in vivo, which mitigated cardiac fibrosis and restored cardiac function.320
Given the pivotal role played by autoreactive T and B cells in autoimmune disorders, CAR-T-cell therapy has also become a therapeutic option within the realm of autoimmune diseases.321 The pathogenesis of myasthenia gravis (MG) involves the presence of pathogenic plasma cells and autoantibodies, making it a prototypical autoimmune disorder. The ongoing Phase Ib/IIa trial (NCT04146051) involving Descartes-08 rCAR-T cells for the treatment of generalized MG entailed transfecting mRNA-encoding anti-BCMA (B-cell maturation antigen) into CD8+ T cells. The findings demonstrated that Descartes-08, which expressed anti-BCMA targeting CAR protein on autologous CD8+ T cells, exhibited favorable safety and tolerability profiles in MG patients. Furthermore, sustained reduction in MG severity was observed with continued administration of Descartes-08 infusions over a follow-up period of up to 9 months. These promising outcomes warranted further investigation into the potential therapeutic efficacy of this approach in other autoimmune disorders.121 Excitingly, the Descartes-08 anti-BCMA rCAR-T therapy for systemic lupus erythematosus (SLE-001) has successfully progressed into Phase II as of February 2024 (NCT06038474).322 On the other hand, Thatte et al.323 administered Foxp3 mRNA LNPs to CD4+ T cells in order to generate immunosuppressive Foxp3-T cells. These cells have the capacity to suppress the proliferation of effector T cells, highlighting the potential of mRNA-engineered immunosuppressive T cells in the management of autoimmune disorders.
TCR-T
In addition to CAR-T-cell therapy, there also exist targeted adoptive cell therapies employing T-cell receptor (TCR) engineered T cell (TCR-T cell). To address hepatocellular carcinoma (HCC) associated with HBV following liver transplantation, Yang et al.324 employed electroporation to introduce mRNA-encoding HBV-specific TCR into ex vivo T cells, which were subsequently transferred to the patients. Currently in Phase I clinical trial (NCT02719782), the study demonstrated favorable tolerability of this treatment approach.
Maggadottir et al.325 conducted a study on the treatment of metastatic microsatellite instability-high (MSI-H) CRC, wherein they employed an mRNA TCR-modified T-cell therapy approach. This involved administering autologous T cells electroporated with IVT Radium-1 TCR mRNA, specifically targeting the -1A frameshift mutation in the TGFβRII gene, to patients with advanced MSI-H CRC. The results demonstrated a favorable safety profile for this TCR-T-cell therapy (NCT03431311).
In addition, also for the treatment of viral-associated tumor diseases, such as HPV-associated cervical cancer, Ling et al.326 designed a pH-responsive nonviral nano-carrier to deliver Cas9 mRNA and the guide RNAs (gRNAs) of oncogenes E6 and E7 that target HPV, which can effectively knock out the E6/E7 oncogenes, reverse the tumor’s immunosuppressive environment, and promote CD8+ T-cell survival. Therefore, genome editing therapies co-delivering Cas9 mRNA and gRNAs that target oncogenes can also be combined with adoptive T-cell transfer to improve cancer treatment outcomes.
CAR-M
The therapeutic efficacy of CAR-T therapy in treating solid tumors is limited, which is greatly derived from the limited infiltration capacity of T cells within the TME. Conversely, macrophages possess the ability to infiltrate solid tumor tissue and engage with almost all cells in the TME, including tumor cells and diverse immune cells. Consequently, exploring the potential of CAR-expressing macrophages (CAR-M) represents a novel avenue for immunotherapy.327 The investigational agent CT-0058, a pro-inflammatory macrophage cell product, is currently undergoing Phase I clinical trial for the treatment of solid tumors (NCT04660929). The anti-HER2 CAR expressed in CAR-M cells was obtained via adenoviral vectors rather than mRNA transfection.328 Meanwhile, the MCY-M11, currently undergoing clinical Phase I for advanced ovarian cancer and peritoneal mesothelioma treatment, is derived from the transfection of anti-mesothelin CAR (Meso-CAR) mRNA into peripheral blood mononuclear cells (PBMCs) (NCT03608618).329
CAR-M therapy not only holds immense potential in combating cancer, but also exhibits promising applications in conducting cutting-edge research on bacterial eradication. Tang et al.330 developed LNPs system encapsulating mRNA that encoded a CAR specifically targeting methicillin-resistant Staphylococcus aureus (MRSA). The LNPs surface was modified with the CRV peptide (sequence CRVLRSGSC) to facilitate macrophages targeting. This antibacterial CAR-M therapy demonstrated promising potential in eradicating MRSA.
mRNA-engineered DCs
DCs are a crucial subset of APCs responsible for presenting TAAs to T cells, and their functionality can be compromised by TME. Although engineered DCs therapy has gained attention, its standalone efficacy remains unsatisfactory, necessitating combination with other therapies to counteract the inhibitory effects of the TME.331 The year 2002 witnessed a clinical trial evaluating the efficacy a IVT mRNA DC vaccine against melanoma (NCT01278940).332 In 2006, Kyte et al.333 reported the results of this Phase I/II trial, indicating that the treatment of tumor-mRNA transfected DCs was safe and feasible, and that the DC vaccine was able to induce antigen-specific T-cell responses in ~50% of patients.
To reprogram the TME and initiate tumor-specific T-cell responses, Zhang et al.331 designed two mRNA LNPs. One of these mRNA LNPs encoded CD40 ligand, which elicited potent ICD in tumor tissues, resulting in the expression of TAAs and CD40 ligand on the surface of tumor cells. The other LNPs carried CD40 mRNA with the aim to be internalized by DCs for generating in situ engineered DCs, leading to increased expression of CD40 protein and activation upon interaction with CD40 ligand on tumor cells.
The majority of patients diagnosed with acute myeloid leukemia (AML) exhibit a persistent and recurrent manifestation of the disease attributed to residual leukemic cells, even following standard chemotherapy, resulting in a survival rate of less than 5 years. Consequently, supplementary therapeutic interventions are imperative for eradicating minimal residual disease (MRD). The significant overexpression of Wilms tumor 1 (WT1) in AML and its association with the pathogenesis of the disease render it a promising target for T cells. In light of this, a Phase II clinical trial was conducted in 2010 to evaluate the efficacy of loading autologous DCs with WT1 antigen via mRNA electroporation as an adjunctive treatment strategy for AML patients who had completed chemotherapy (NCT00965224). The findings reported by Anguille et al.334 demonstrated that this vaccination approach effectively elicits robust T-cell immune responses in AML patients at high risk of relapse, thereby establishing both the safety and potential utility of autologous WT1 mRNA-electroporated DC vaccine following chemotherapy.
Non-immunotherapy
The emergence of mRNA therapies has infused considerable hope into the battle against previously incurable diseases. Theoretically, optimally designed IVT mRNA can be introduced into cells either through in vivo or ex vivo transfection and subsequently translated into biologically active peptide/protein.335 The non-immunotherapy described in this article entails directly transcription of mRNA to complement deficient or aberrant proteins within the organism, as well as for gene editing technology, while evading any immune response from the recipient (Fig. 4).
Fig. 4.

Overview diagram of mRNA-based non-immunotherapy. BDNF brain-derived neurotrophic factor, NEP neprilysin, CFTR cystic fibrosis transmembrane conductance regulator, HNF4A human hepatocyte nuclear factor α, COL1A1 extracellular-matrix α1 type-I collagen, VEGF-A vascular endothelial growth factor-A, TE tropoelastin, BMP-2 bone morphogenetic protein-2, RUNX1 runt-related transcription factor 1, OTC ornithine transcarbamylase, G6Pase-α glucose-6-phosphatase-alpha, AGL amylo-α-1,6-glucosidase 4-alpha-glucanotransferase, IGF-1 insulin-like growth factor 1, ZFNs Zinc finger nucleases, CRISPR/Cas9 clustered regularly interspaced short palindromic repeats/associated protein 9. The graphic is created with BioRender.com
The effective treatment of genetic defect diseases with IVT mRNA relies on multiple factors, encompassing the stability and translational potency of the mRNA, the efficacy and biological activity of the encoded protein, as well as its circulation half-life. Importantly, it is worth noting that the therapeutic threshold necessitates an active protein quantity ranging from milligrams to grams, whereas even at the milligram or nanogram level, antigens can provoke an immune response in the body.336 To mitigate the immunogenicity of mRNA therapies, several optimizations can be implemented for IVT mRNA, including refining the nucleotide sequence, employing diverse mRNA nucleoside modifications, optimizing reaction conditions of the IVT conditions to minimize byproducts generation, and purifying the synthesized mRNA.337
The repeated administration remains effective in maintaining high protein expression and ensuring in vivo safety, which is crucial for the transformation of mRNA vaccines into mRNA therapeutics. Long-term or even lifelong repeated administration is necessary for treating chronic diseases.338 However, delivery vectors, such as LNPs, constitute a crucial component in mRNA-based therapeutics, necessitating careful consideration of their immunogenic potential.
The pattern-recognition receptor family on the cell membrane includes TLR4, which is a crucial member known to be activated by damage- and pathogen-associated molecular patterns (DAMPs and PAMPs). PAMPs encompass lipopolysaccharide (LPS)339 and monophosphoryl lipid A (MPLA; a nontoxic TLR4 agonist approved for clinical use).340 Kedmi et al.341 discovered that the systemic administration of positively charged NPs carrying small interfering RNA (siRNA) could trigger TLR4-dependent immune responses, leading to toxicity. Furthermore, they demonstrated that the inflammatory reaction was primarily caused by cationic components rather than the siRNA. The cationic lipid DiC14-amidine, synthesized by Lonez et al.,339 was shown to activate TLR4. Moreover, competition experiments confirmed that the binding site of this cationic lipid to TLR4 differed from that of LPS. Lonez et al.342 synthesized a cationic lipid, RPR206252, which effectively triggered the NF-κB pathway and elicited TNF-α, IL-1β, IL-6, and IFN-γ production in human or mouse macrophages. These cascades were reliant on TLR2/CD14 and NOD-like receptor protein 3 (NLRP3) signaling.
Therefore, it is imperative to develop low-immunogenic LNPs with diverse compositions and enhanced organ-selectivity, or explore alternative delivery vehicles to amplify the therapeutic efficacy of mRNA therapies for non-immune diseases.343 In addition, the combination of mRNA therapy with immunosuppressants such as Dexamethasone (a steroidal anti-inflammatory drug) not only holds the potential to enhance mRNA transfection efficiency344 but also exhibits promise in augmenting the safety and efficacy of non-immunotherapy.
In consideration of the safety of mRNA drugs, it is recommended that preclinical studies encompass the following areas: (1) investigating the correlation between mRNA dosage, carrier properties, and in vivo levels of cytokine secretion as well as complement activation; (2) assessing the potential development of anti-drug antibodies upon repeated administration; (3) evaluating risk of organ damage, particularly focusing on target organs and concerns related to liver toxicity, along with potential accumulation risks associated with carriers.337
Protein replacement therapy
Protein replacement therapy is a broad term encompassing the supplementation of proteins with functional defects or the substitution of deficient proteins to achieve therapeutic efficacy.345 The delivery of therapeutic proteins directly to the target tissue is significantly impeded by factors such as protein size and biochemical properties.346 In contrast, mRNA therapies are more likely to bypass these challenges. At present, the use of mRNA translation in vivo to replace the direct delivery of proteins has become a new pillar of protein replacement therapy, and has been extensively studied in various disease areas, including blood diseases (such as Hemophilia A, HemA), and metabolic diseases (such as ornithine transcarbamylase deficiency, OTCD; Fabry disease, FD; Propionic acidaemia, PA).335
mRNA therapy provides a promising treatment option for refractory genetic diseases. HemA, a form of hemophilia caused by insufficient expression or gene mutation of coagulation factor VIII (FVIII), results in impaired blood clotting ability and can lead to life-threatening bleeding episodes, including intracranial hemorrhage. Current protein replacement therapies are costly and require frequent administration due to the short half-life of FVIII protein.347 In 2023, the FDA approved Roctavian as the first gene therapy drug for severe HemA in adults, utilizing adeno-associated virus vectors; however, concerns arise regarding potential risks such as liver cancer associated with the introduction of DNA sequences from Roctavian products.348 mRNA therapy can express the target protein in the cytoplasm without entering the nucleus or inserting the target cell genome, which is much safer than viral vector and DNA therapy.1 Chen et al.347 designed an LNPs encapsulating mRNA-encoding FVIII protein, which effectively prolonged the duration of FVIII protein expression.
Low levels of key enzyme proteins or lack of enzyme activity resulting from genetic defects can give rise to inherited metabolic disorders (IMDs). Among these, OTCD is caused by the loss of crucial enzymes involved in the urea cycle within the liver, leading to impaired ammonia metabolism. Elevated levels of ammonia in the bloodstream can result in nerve damage and even fatality. Existing approaches, including dietary controls and ammonia scavengers, fail to address the root cause.349 Based on this, Prieve et al.349 designed a Hybrid mRNA Technology delivery system (HMT) that protected OTC-encoding mRNA against degradation by nucleases while exhibiting hepatocellular-specific targeting capabilities. HMT consists of two types of nanoparticles: di-block polymer micelles and inert LNPs. The polymer comprises three functional domains: (1) GalNAc, which serves as a targeting ligand for liver-specific uptake by binding to the asialoglycoprotein receptor abundantly expressed in liver cells; (2) hydrophilic polymer segments that maintain the solubility of the polymer; (3) polymer segments containing butyl methacrylate (hydrophobic monomer), 2-propylacrylic acid, and 2-(dimethylamino)ethyl methacrylate. This polymeric vesicle was designed to achieve liver-specific targeting and further facilitate the pH-dependent release of mRNA from endosomal organelles to the cytosol. The inert LNPs can shield mRNA from nucleases. This platform is not only safe but also extends the therapeutic effects for OTCD treatment. Currently, ARCT-810 (OTC mRNA LNPs) has entered a Phase II clinical trial (NCT05526066) in OCT patients aged 12 years and older.350
FD is an X-linked lysosomal storage disorder (LSD).351 Its root cause lies in the pathogenic mutation of the GLA gene (Xq21.3-q22), which leads to a loss of enzyme activity encoding lysosomal enzyme α-galactosidase A. This results in abnormal metabolism of related glycolipids and the subsequent accumulation of glycolipids throughout the body, causing structural damage and functional loss in various tissues and organs, particularly affecting the heart, kidney, and nervous system. The resulting cardiac dysfunction and cerebrovascular events pose life-threatening risks. Although there are clinically approved enzyme replacement therapies (ERTs), they fail to meet medical needs due to their inability to penetrate the blood-brain barrier.352 To this end, in 2019, DeRosa et al.351 and Zhu et al.353, respectively, reported systemically delivery of mRNA-encoding human alpha-galactosidase A (H-α-gal A) using LNPs, both of which indicated that mRNA LNPs was a potential treatment modality for FD.
PA is a life-threatening IMDs caused by a pathogenic mutation in the propionyl-coenzyme A carboxylase α or β (PCCA or PCCB) subunits gene, and there are currently no approved drugs to address this enzyme deficiency. An innovative approach involving two mRNAs encoding normal human PCCA and PCCB protein subunits has entered Phase I/II clinical trials (NCT04159103). Encapsulated within LNPs, these mRNAs are specifically designed for targeted delivery to the liver to restore PCC enzymes. Promising interim results from this trial have demonstrated a favorable safety profile of mRNA-3927 in PA patients. Notably, dose-optimization studies have revealed a remarkable 70% reduction in relative risk associated with metabolic decompensation events (MDEs), highlighting the therapeutic efficacy.354
Hormone replacement therapy
Hormones produced by various glands in the body serve as crucial signaling molecules that are transported through the bloodstream to target organs, tissues, and cells, playing a pivotal role in maintaining physiological balance. Disruptions in hormonal regulation can give rise to diverse diseases.355 During menopause, hormonal fluctuations lead to the manifestation of menopausal symptoms and an increased risk of cardiovascular disorders. To alleviate these symptoms and slow down the progression of cardiovascular diseases, non-protein hormones like estrogen can be administered orally or topically via gel application. Similarly, for conditions such as androgen deficit in aging men (ADAM) or male hypogonadism, supplementation through similar means may also be considered.356 However, due to their size and spatial structure, protein hormones cannot be effectively administered orally as they would undergo degradation within the digestive system. Surprisingly, in 1992, Jirikowski et al.357 injected mRNA-encoding vasopressin into Brattleboro rats with successful expression of vasopressin in the hypothalamus and showed therapeutic effect (temporary reversal of diabetes insipidus for up to 5 days).
Insulin-like growth factor 1 (IGF-1) is a versatile growth factor primarily synthesized in the liver.358 It can regulate the chondrogenesis of mesenchymal stem cells (MSCs) by stimulating their proliferation and facilitating cartilage differentiation. Wu et al.359 used modified mRNA (modRNA) encoding IGF-1 to engineer adipose-derived stem cells (ADSCs). The therapeutic effect of engineered ADSCs on Osteoarthritis (OA) is superior to that of natural ADSCs.
Although there have been limited preclinical investigations on mRNA-encoded protein-like hormones, it is exciting that that mRNA-0184 (encoding the Relaxin-2-variable Light Chain Kappa) developed by ModernaTX, Inc. for the treatment of chronic heart failure has entered Phase I clinical trials in 2023 (NCT05659264).
Regenerative medicine application
Myocardial infarction
In humans, myocardial infarction results in the occlusion of a portion of the heart’s blood supply and the subsequent death of billions of cardiomyocytes, leading to a high mortality rate due to the challenging regeneration of lost cardiomyocytes. This difficulty hinders the repair of damaged heart function. The vascular endothelial growth factor (VEGF-A) has been identified as having angiogenic properties,360 making it a potential therapeutic agent for patients with ischemic heart disease. However, its systemic administration is associated with long-term side effects and poses challenges in specifically targeting the heart.361 In 2013, Zangi et al.360 designed modRNA encoding VEGF-A and found that a single intracardial infusion of VEGF-A modRNA could increase the density of blood vessels around infarction, reduce infarct size and cell death. In addition, compared to mice treated with VEGF-A DNA, those receiving VEGF-A modRNA showed improved survival rates. This improvement can be attributed to careful consideration regarding timing and dosage since prolonged exposure to VEGF-A in the DNA group resulted in increased vascular permeability and cardiac edema. Furthermore, they discussed how mobilization of epicardial progenitor cells played a crucial role in enhancing cardiac function through expansion and differentiation into cardiovascular lineages.
The VEGF-A165 mRNA (AZD8601), dissolved in biocompatible citrate saline, has commenced clinical trials. Specifically, the Phase IIa clinical trial (NCT03370887) aims to enhance outcomes in coronary artery bypass graft patients through endocardial injection of AZD8601.362 In addition, AZD8601 exhibits promise for treating type 2 diabetes mellitus (T2DM) and is currently undergoing Phase I clinical trials (NCT02935712).363 Nawaz et al.364 compared VEGF-A protein production in LNPs containing the same amount of VEGF-A mRNA or in extracellular vesicles (EVs) secreted by three different types of cells (cardiac progenitor cells, CPCs; human lung epithelial HTB-177 cells; human umbilical vein endothelial cells, HUVECs) treated with VEGF-A mRNA LNPs. Their findings revealed that when delivered to endothelial cells in vitro, the LNPs carrier produced the highest level of VEGF-A while CPC-EVs demonstrated the lowest protein yield. However, CPC-EVs was the most effective in promoting angiogenesis per given amount of VEGF-A produced. Intriguingly, during intramyocardial injection, EVs-based carriers displayed the highest expression of VEDF-A. Considering that CPC-EVs induce minimal inflammatory cytokine expression compared to other vectors, they are better suited for delivering mRNA to the heart.
The T-box transcription factor 18 (TBX18) plays a pivotal role in the formation and differentiation of the sinoatrial node (SAN).365 Wolfson et al.366 discovered that direct injection of naked TBX18 mRNA into the myocardium of rats could effectively transfect cardiomyocytes, leading to transient expression of TBX18 and subsequent reprogramming of the cardiomyocytes into pacemaker cells. Furthermore, they observed that compared to adenoviral vectors, mRNA transfection exhibited a significant reduction in off-target gene expression and lower immunogenicity.
Liver fibrosis and cirrhosis
In the context of liver diseases, human hepatocyte nuclear factor α (HNF4A) has been recognized as a pivotal regulator of liver cell phenotype, exerting its influence on hepatic stellate cells and liver macrophages by targeting paraoxoase-1. This targeted regulation effectively mitigates liver fibrosis. Yang et al.367 designed HNF4A mRNA-loaded LNPs to improve liver fibrosis and cirrhosis in vivo. It was noteworthy that the use of human HNF4A mRNA not only restored the expression of HNF4A in mouse liver but also avoided adverse immune reactions, thereby further enhancing its anti-fibrotic efficacy.
Cystic fibrosis
The cystic fibrosis transmembrane conductance regulator (CFTR) gene encodes epithelial ion channels responsible for the transport of chloride and bicarbonate, playing a crucial role in maintaining normal physiological functions. Cystic fibrosis is a monogenic disease caused by mutations in the CFTR gene that causes the lungs and other organs to produce thick mucus that blocks the airways of the lungs and makes breathing difficult, leading to lung damage, respiratory failure and chronic lung infections. In addition, cystic fibrosis also affects other organs such as the pancreas resulting in digestive problems and malnutrition. Furthermore, cystic fibrosis can give rise to various complications including diabetes, liver disease, and infertility. Unfortunately, there is currently no cure for this lifelong condition.276,368
In 2018, Translate Bio, Inc. conducted a Phase II trial (NCT03375047) involving adults and seniors aged 18 years or older with cystic fibrosis to investigate the safety and therapeutic efficacy of MRT5005—an aerosol containing codon-optimized CFTR mRNA. It was found that the overall safety of patients receiving MRT5005 was good, but fever and allergic reactions occurred. The etiology and mechanism of fever remain unclear, necessitating further research.369 Additionally, two more mRNA therapies, namely ARCT-032 (encoding CFTR protein, NCT05712538) and VX-522 mRNA therapy (NCT05668741), have commenced clinical trials in 2023, with the outcomes yet to be disclosed.
Degenerative disease
The treatment of bone defects resulting from nonunion, trauma, or craniofacial malformations is challenging and affects millions of individuals. Currently, the delivery of bone morphogenetic protein-2 (BMP-2) shows promise as a treatment approach to address these defects. However, it has been observed that excessive administration of BMP-2 can lead to severe side effects such as significant inflammation and swelling. For heterotopic ossification, local delivery and precise translation of BMP-2 mRNA to produce functional proteins can be achieved. Wang et al.370 employed a dual delivery strategy by simultaneously administering BMP-2 mRNA and non-structural protein-1 (NS1) mRNA in a mass ratio of 3:1. Remarkably, their findings demonstrated that this approach induced high expression levels of BMP-2 surpassing previously reported yields. Notably, they are the first to use unmodified mRNA in the field of regenerative medicine since the co-delivered NS1 mRNA encodes an immune evasion protein originally expressed by influenza A viruses (A/Texas/36/1991), which effectively inhibits RNA sensors while reducing the production of immune factors to prevent activation of harmful immune responses.
Wang et al.371 further explored the delivery platform of BMP-2/NS1 mRNA by incorporating a lipopolyplex (Lip100/His-lPEI/RNA ternary complex, LPR) loaded with BMP-2/NS1 mRNA into a collagen-nanohydroxyapatite scaffolds. The resulting ready-to-use mRNA-activated matrices (RAMs), obtained through freeze–drying, exhibit a remarkable enhancement in the release time of mRNA, extending up to 16 days. This approach enables in situ expression and continuous production of BMP-2 protein, thereby promoting bone formation. Geng et al.372 found that compared with the delivery of recombinant protein or single modRNA alone, the co-delivery of human BMP-2 (hBMP-2) and VEGF-A modRNA-modified bone marrow stem cells (BMSCs) increased the expression level of osteogenic-related gene and enhanced the formation of new bone in rats undergoing skull defect surgery. These findings offer a promising therapeutic option for regenerating bone tissue.
Runt-related transcription factor 1 (RUNX1) is a cartilage-anabolic factor, which promotes the proliferation of chondrocytes and increases the expression of cartilage-anabolic markers of chondrocytes. Furthermore, RUNX1 enhances coccygeal disc hydration content while mitigating disc degeneration by minimizing loss of disc cartilage and other constituents. In this regard, Lin et al.373 developed RUNX1 mRNA nanomicelles for the treatment of intervertebral disc (IVD) degeneration. Compared to the control group, administration of RUNX1 mRNA nanomicelles significantly maintained a higher disc height in rats with coccygeal disc degeneration and prevented fibrosis of the disc tissue.
Alzheimer’s disease is a degenerative disorder resulting from an imbalance in the metabolic processes of amyloid-beta (Aβ) synthesis and clearance.374 Neprilysin (NEP) is known to play an important role in the clearance process of Aβ. Therefore, Lin et al.375 designed polyplex nanomicelles based on NEP-encoding mRNA, which significantly reduced the concentration of Aβ in the brain, demonstrating the potential of mRNA-based therapy in brain therapy. In addition, Li et al.376 engineered an mRNA encoding brain-derived neurotrophic factor (BDNF) for AD that was packaged on a poly (β amino esters) (PBAE) nanoplatform and delivered directly to the central nervous system. To prevent excessive neuronal activation caused by high translation of BDNF mRNA, they modified the 3’UTR of this mRNA with neuron-specific miRNA-124 target sequences, ensuring degradation by miRNA upon delivery to neurons. By expressing and releasing BDNF in astrocytes to support neurons, the BDNF mRNA effectively enhanced memory function in AD model mice, thus highlighting the potential of mRNA therapy in the treatment of neurological diseases.
Connective tissue disorder
Elastin, one of the most enduring proteins (with a half-life of approximately 74 years), is predominantly expressed during the neonatal period. However, with age, elastin synthesis gradually diminishes and eventually ceases in adults. Elastin plays a pivotal role in endowing tissues and organs with elasticity, thereby contributing significantly to the normal functioning of elastic connective tissue. Unfortunately, genetic disorders like Williams–Beuren syndrome (WBS) can lead to mutations in elastin-related genes, resulting in a reduction of over 50% in elastin synthesis. Homozygous elastin null mutants (ELN−/−) succumb shortly after birth. In addition, factors such as aging and sunburn can also lead to elastin reduction. Lescan et al.377 synthesized and modified the mRNA-encoding human tropoelastin (TE). Their findings demonstrated that delivery of TE mRNA increased the synthesis of elastin in pig skin by 20%. Furthermore, Golombek et al.378 carried out codon optimization and natural nucleotide modification on TE mRNA to improve the translation efficiency and stability of mRNA, thus improving the expression level of TE protein in vivo and in vitro. This study not only provides valuable insights for other conditions necessitating de novo elastin synthesis but also holds promise for diseases like myocardial infarction.
Another key protein in connective tissue is collagen.379 You et al.380 designed an exosome-based extracellular-matrix α1 type-I collagen (COL1A1) mRNA for anti-aging and treatment of photoaging skin that successfully alleviated UV-induced skin aging through intradermal delivery via microneedle arrays.
Targeted genome engineering
Targeted genome engineering is the use of gene editing technology to knock out, insert and replace the target genome sequence to achieve changes in genetic information,381 which brings a new way for the treatment of diseases (especially genetic diseases caused by gene defects).382 Zinc finger nucleases (ZFNs) and transcriptional activator-like effector nucleases (TALENs) are widely employed in gene editing techniques over a decade ago.383 However, their application is complexed and necessitates the design of proteins that specifically bind to target genes.384 The advent of clustered regularly interspaced short palindromic repeats (CRISPR)/associated protein 9 (CRISPR/Cas9) systems has significantly propelled the advancement of gene editing technology. CRISPR/Cas9 obviates the need for altering the protein sequences and only requires the design of alternative gRNAs for customization purposes.385 There are three typical forms of the CRISPR/Cas9 system: plasmid DNA (pDNA), Cas9 mRNA/sgRNA, and ribonucleoprotein (RNP, Cas9 protein complexed with gRNA). CRISPR/Cas9 pDNA faces the challenge of nuclear localization and has the risks of off-target repeat expression and genome integration; Cas9 RNP delivery has to surmount obstacles related to macromolecular size and instability; while Cas9 mRNA/sgRNA needs to address the instability of single-stranded nucleic acid.386
The CRISPR/Cas9 technology can treat diseases by knocking out abnormal genes. The hallmark of wet age-related macular degeneration (wAMD) is vascular dysplasia. At present, anti-VEGF reagents, the first-line drug for the treatment of wAMD, are expensive and require repeated or even lifelong injection. In light of this, Ling et al.387 designed a platform that used a lentiviral system to package Cas9 mRNA and VEGF-A-targeting gRNA. The area of choroidal neovascularization (CNV) was reduced by 63% with no detectable off-target effect. Legumain, encoded by the LGMN gene located on human chromosome 14, has been identified in various tumors such as breast and gastric cancers where it is associated with tumor aggressiveness, migratory behavior, and poor prognosis. Wang et al.388 used LNPs to co-deliver Cas9 mRNA and LGMN gene-targeting gRNA, and found that this platform held promise as a therapeutic approach to inhibit breast tumor metastasis.
The CRISPR/Cas9 technology can also remove viral DNA from the body. There are 296 million patients with chronic HBV infection in the world, and current clinical treatment can slow the replication of the virus to a certain extent and delay the progression of the disease, but the viral DNA in infected cells cannot be cleared, and HBV DNA integrated into the host body is considered to be a significant source of cancer risk. Moreover, the formation of cccDNA in the liver nucleus and continuous production of progeny virus posed a challenge to the radical treatment of HBV. To address this issue, Yi et al.389 used SM-102-based LNPs to deliver Cas9 mRNA and gRNAs together, and employed this CRISPR/Cas9 gene editing technology to achieve targeted site-specific editing. This approach provided an opportunity for continuous elimination of cccDNA and integrated HBV DNA, thereby facilitating a radical treatment strategy against HBV. In addition, to treat HPV-related cervical cancer, Ling et al.326 designed a pH-responsive nonviral nanonucleus for co-delivery of Cas9 mRNA and gRNAs targeting HPV oncogenes E6 and E7. This platform allowed for combining Cas9 mRNA/gRNA with adoptive cell therapy to effectively treat cancer diseases.
Although CRISPR/Cas9 technology holds immense potential for the treatment of genetically related diseases and virus clearance, its development has been hindered by the challenges associated with delivering mRNA/gRNA. Currently, CRISPR/Cas9 is predominantly used in the form of Cas9 RNP or pDNA and delivered via viral vectors or electroporation in clinical trials.386 However, there are active efforts to develop mRNA/gRNA delivery strategies such as LNPs,388–390 lentivirus, and retroviral vectors.387,391 We look forward to developing various delivery vectors to advance the clinical application of mRNA/gRNA.
Clinical trials based on Cas9 mRNA/gRNA have been conducted. Transthyretin (TTR) amyloidosis (abbreviated ATTR amyloidosis) is a life-threatening disease caused by the accumulation of misfolded TTR proteins in tissues, particularly in the heart and nerves. NTLA-2001 represents a gene editing therapy based on the CRISPR-Cas9 system that includes LNPs encapsulated with mRNA-encoding Cas9 protein and TTR-targeting gRNA. The interim trial results from Phase I study (NCT04601051) demonstrated that NTLA-2001 effectively reduced serum TTR concentration with only mild adverse events.392 Other examples can be found in Tables 2 and 3. Currently, mRNA-based therapies for encoding ZFN, TALEN, and Cas9 are generally in the developmental stage and have the potential to play a crucial role in disease treatment.393
Table 2.
Representative completed and ongoing clinical studies (non-immunotherapy)
| Therapy | Disease | mRNA | mRNA-encoded protein | Delivery system | Administration route | Study started | Phase | Status | Sponsor | NCT number |
|---|---|---|---|---|---|---|---|---|---|---|
| Protein replacement therapy | Ornithine transcarbamylase deficiency (OTCD) | ARCT-810 | Ornithine transcarbamylase (OTC) | LNPs | Intravenous infusion | 2022-07-06 | Phase II | Recruiting | Arcturus Therapeutics, Inc. | NCT05526066 |
| MRT5201 | LNPs | Intravenous administration | 2019-12 | Phase I/II | Withdrawn | Translate Bio, Inc. | NCT03767270 | |||
| Propionic acidaemia (PA) | mRNA-3927 | Propionyl-coenzyme A carboxylase α or β (PCCA or PCCB) subunits | LNPs (SM-86, DSPC, cholesterol, and PEG lipid) | Intravenous infusion | 2021-04-15 | Phase I/II | Recruiting | ModernaTX, Inc. | NCT04159103 | |
| Glycogen storage disease 1a (GSD1a) | mRNA-3745 | Glucose-6-phosphatase-alpha (G6Pase-α) | LNPs | Intravenous infusion | 2022-06-01 | Phase I/II | Recruiting | ModernaTX, Inc. | NCT05095727 | |
| Glycogen storage disease type III | UX053 | Amylo-α-1,6-glucosidase 4-alpha-glucanotransferase (AGL) | LNPs | Intravenous infusion | 2021-10-18 | Phase I/II | Terminated | Ultragenyx Pharmaceutical Inc | NCT04990388 | |
| Methylmalonic acidemia (MMA) | mRNA-3704 | Undisclosed | LNPs | Intravenous infusion | 2019-05-28 | Phase I/II | Withdrawn | ModernaTX, Inc. | NCT03810690 | |
| mRNA-3705 | Methylmalonyl-coenzyme A mutase | LNPs (SM-86, DSPC, cholesterol, and PEG lipid) | Intravenous injection | 2021-08-06 | Phase I/II | Recruiting | ModernaTX, Inc. | NCT04899310 | ||
| Injection | 2022-03-08 | Phase I/II | Recruiting | ModernaTX, Inc. | NCT05295433 | |||||
| Hormone replacement therapy | Chronic heart failure | mRNA-0184 | Relaxin-2-variable light chain kappa | Undisclosed | Intravenous infusion | 2022-12-05 | Phase I | Recruiting | ModernaTX, Inc. | NCT05659264 |
| Healthy participants | Undisclosed | Intravenous infusion | 2024-02-16 | Phase I | Recruiting | ModernaTX, Inc. | NCT06243770 | |||
| Regenerative medicine application | Myocardial infarction | AZD8601 | Vascular endothelial growth factor-A (VEGF-A) | None | Epicardial injection | 2018-02-05 | Phase IIa | Completed | AstraZeneca | NCT03370887 |
| Type 2 diabetes mellitus (T2DM) | None | Intradermal injection | 2016-12-16 | Phase I | Completed | AstraZeneca | NCT02935712 | |||
| Cystic fibrosis | MRT5005 | Cystic fibrosis transmembrane conductance regulator (CFTR) protein | LNPs | Inhalation | 2018-05-10 | Phase I/II | Unknown status | Translate Bio, Inc. | NCT03375047 | |
| ARCT-032 | LNPs | Inhalation | 2023-02-15 | Phase I | Recruiting | Arcturus Therapeutics, Inc. | NCT05712538 | |||
| VX-522 mRNA therapy | Undisclosed | LNPs | Oral inhalation using nebulizer | 2023-02-27 | Phase I/II | Recruiting | Vertex Pharmaceuticals Incorporated | NCT05668741 | ||
| Targeted genome engineering | Transthyretin amyloidosis (ATTR) with cardiomyopathy | NTLA-2001 | Cas9 protein | LNPs (ionizable lipid, DSPC, cholesterol, PEG2000-DMG) | Intravenous infusion | 2023-12-13 | Phase III | Recruiting | Intellia Therapeutics | NCT06128629 |
| Hereditary angioedema (HAE) | NTLA-2002 | LNPs | Intravenous injection | 2021-12-10 | Phase I/II | Active, not recruiting | Intellia Therapeutics | NCT05120830 | ||
| Refractory viral keratitis | BD111 CRISPR/Cas9 mRNA | SpCas9 | Lentiviral particles | Corneal injection | 2020-11-04 | Not applicable | Completed | Shanghai BDgene Co., Ltd. | NCT04560790 | |
| HIV | SB-728mR-HSPC | C-C motif chemokine receptor 5 (CCR5) -specific zinc finger nucleases | Autologous hematopoietic stem and progenitor cells (HSPCs) | Infusion | 2016-03-10 | Phase I | Active, not recruiting | City of Hope Medical Center | NCT02500849 | |
| Transfusion-dependent beta-thalassemia (TDT) | ST-400 investigational product | ZFNs | Autologous hematopoietic stem and progenitor cells (HSPCs) | Infusion | 2018-03-29 | Phase I/II | Completed | Sangamo Therapeutics | NCT03432364 |
LNPs lipid nanoparticles, DSPC distearoylphosphatidylcholine, PEG polyethylene glycol, CRISPR/Cas9 clustered regularly interspaced short palindromic repeats/associated protein 9, HIV human immunodeficiency virus, ZFNs Zinc finger nucleases
Table 3.
Representative preclinical studies (non-immunotherapy)
| Therapy | Target organ/tissue | Disease | mRNA-encoded protein | Delivery system | Administration route | Ref. |
|---|---|---|---|---|---|---|
| Protein replacement therapy | Blood | Hemophilia A | Coagulation factor VIII | LNPs (ionizable lipid: DSPC: cholesterol: PEG lipid, 50:10:38.5:1.5) | Intravenous injection | 347 |
| Liver | Ornithine transcarbamylase deficiency (OTCD) | Ornithine transcarbamylase (OTC) | LNPs (DOTAP, cholesteryl hemisuccinate, cholesterol, PEG lipid); di-block polymer micelle | Intrahepatic delivery | 349 | |
| Whole body (especially heart, kidneys, and brain) | Fabry disease (FD) | α-galactosidase A | LNPs (C12-200, DOPE, cholesterol, dimyristoyl glycerol-polyethylene glycol analog) | Intravenous injection | 351 | |
| Whole body (especially heart, kidneys, and brain) | FD | α-galactosidase A | LNPs (ionizable: helper: structural: PEG, 50:10:38.5:1.5) | Intravenous injection | 353 | |
| Hormone replacement therapy | Brain | Diabetes insipidus | Vasopressin | None | Injection into the hypothalamus | 357 |
| Bone | Osteoarthritis (OA) | Insulin-like growth factor 1 (IGF-1) | ADSCs | Intraarticular injection | 359 | |
| Regenerative medicine application | Heart | Myocardial infarction | Vascular endothelial growth factor-A (VEGF-A) | None | Intramyocardial injection | 360 |
| Liver | Liver fibrosis and cirrhosis | Hepatocyte nuclear factor alpha (HNF4A) | LNPs | Intravenous injection | 367 | |
| Bone | Bone defects | Bone morphogenetic protein-2 (BMP-2) | Lipopolyplex | Subcutaneously implant | 371 | |
| Intervertebral disk (IVD) | IVD degeneration | Runt-related transcription factor 1 (RUNX1) | Polyplex nanomicelles (PEG-polyamino acid block copolymers) | Administered into rat coccygeal disks | 373 | |
| Brain | Alzheimer’s disease | Neprilysin (NEP) | Polyplex nanomicelles (PEG-polyamino acid block catiomer) | Intracerebrovetricular administration | 375 | |
| Nervous system | Alzheimer’s disease | Brain-derived neurotrophic factor (BDNF) | PBAE polymers | Delivery to the central nervous system by brain ventricle pumping | 376 | |
| Skin | Photoaged skin | Extracellular-matrix α1 type-I collagen (COL1A1) | EVs | Intradermal delivery (microneedle array) | 380 | |
| Connective tissue | Williams–Beuren syndrome (WBS) | Tropoelastin (TE) | None | Intradermal injection | 377 | |
| Targeted genome engineering | Eye | Wet age-related macular degeneration | Cas9 | Lentiviral system | Subretinal injection | 387 |
| Tumor | Breast cancer | Cas9 | LNPs | Intravenous injection | 388 | |
| Viral carcinogenesis | HBV | Cas9 | LNPs (SM-102: DSPC: cholesterol: PEG lipid, 50:10:38.5:1.5) | Intravenous injection | 389 |
LNPs lipid nanoparticles, DSPC distearoylphosphatidylcholine, PEG polyethylene glycol, DOTAP (2,3-dioleoyloxy-propyl)-trimethylammonium chloride, DOPE 1,2-dioleyl-sn-glycero-3-phosphoethanolamine, ADSCs adipose-derived stem cells, EVs extracellular vesicles, HBV hepatitis B virus
The impacts from individual physiological and pathological characteristics
Beginning in late 2019, patients infected with SARS-CoV-2 virus will progress to COVID-19 disease, ranging from asymptomatic infection to life-threatening disease, with particularly high mortality among vulnerable populations (such as the elderly with compromised immunity, individuals with chronic diseases, those use immunosuppressants, and cancer patients).394 The U.S. FDA and the European Medicines Agency (EMA) approved mRNA-1273 (Moderna) and BNT162b2 (Pfizer/BioNTech) for eliciting potent B-cell and T-cell immune responses in healthy people to prevent SARS-CoV-2 infection.395,396 However, the protective effect of the vaccine wanes over time, which can be observed in healthy subjects and is more pronounced in immunocompromised subjects. Therefore, it becomes imperative to consider administering a third or even fourth dose of the vaccine.
Specifically, Rizzi et al.394 recruited patients receiving immunosuppressive therapy for a follow-up trial. They found an absence of protective antibody response after two primary vaccinations, and detectable antibody titers were not produced until a third vaccination. Moreover, given that the induced immune response diminishes gradually after the third vaccination, depending on the patient’s condition, even a fourth immunization was necessary. Nugent et al.397 studied the antibody response and pseudovirus neutralization against SARS-CoV-2 wild-type strain, Omicron BA.1, and BA.5 variants in nursing home residents and healthcare workers after primary mRNA vaccination, first and second boosters. They found that antibody titers and neutralizing ability were progressively increasing with each booster, but subsequently diminished over 3–6 months. Considering that individuals with human immunodeficiency virus (HIV) have significantly compromised immunity, many countries have prioritized SARS-CoV-2 mRNA vaccines for this population. It has been observed that after receiving two doses of BNT162b2 vaccine, HIV-infected ones exhibit a more pronounced weakening in the antibody responses than their healthy counterparts, therefore, a third vaccination is recommended. Heftdal et al.398 investigated the persistence of cellular and humoral immunity in HIV patients and controls after the third dose of vaccine and found no significant differences in antibody titers and cellular responses between HIV patients and controls at 4 months after the third dose of BNT162b2 vaccine. In conclusion, repeated vaccination is more imperative in susceptible individuals than in their healthy counterparts for the establishment of enduring immune protection.394
The above results are consistent with the results of epidemiological studies on hospitalized patients with severe and high-risk COVID-19 (including the elderly and patients with other diseases). However, there still exist significant variations in clinical outcomes among epidemiological patients, suggesting that the underlying cause for symptom disparities between severe COVID-19 patients and asymptomatic carriers may be more profound. Through differential gene screening, Zhang et al.399 identified mutations in type-I IFN genes among severe COVID-19 patients, and they found that at least 3.5% of patients with severe COVID-19 had genetic defects in genes associated with TLR3- and regulatory factor 7 (IRF7)-dependent induction and amplification of type-I IFN.
Furthermore, Bastard et al.400 found that some individuals who remained severely affected by COVID-19 even after receiving two doses of SARS-CoV-2 mRNA vaccine exhibited not only neutralizing antibodies to the virus but also autoantibodies targeting type-I IFN. The lack of type-I IFN may be the cause of severe hypoxemic pneumonia of COVID-19. The aforementioned evidence implies that it is imperative to focus more on the distinct genetic immune characteristics of vulnerable populations (Fig. 5).
Fig. 5.

Immunization background map. The variations in individuals’ immunological backgrounds may serve as a potential mechanism for the diversity in vaccine efficacy and adverse reactions following mRNA vaccination. An individual’s immune background is shaped by a multitude of factors, encompassing both physiological conditions and pathological conditions. Moreover, it is crucial to consider the potential adverse effects of repeated or intensive vaccination using the same or multiple vaccines. The graphic is created with BioRender.com
Age
Vaccination, as a powerful weapon in the prevention and treatment of deadly infectious diseases, can save hundreds of millions of lives by harnessing the human immune response.401 However, human immune system is not static, rather, it undergoes profound changes with age.402 Age-dependent immune system alterations in the elderly include immune senescence in innate immune responses (i.e., changes in the number, function, and distribution of neutrophils, NK cells, monocytes, and DCs),403 and adaptive immune responses (e.g., decreased TCR diversity of aging naïve T cells, shortened telomeres of memory T cells, decreased B-cell pool diversity, and decreased response to foreign antigens).404
However, the reasons for decreased vaccine effectiveness in the elderly go far beyond this, that is, it is not only associated with impaired antibody response, decreased T-cell response, and altered antigen presentation of the immune system, but also related to the chronic and systemic aseptic inflammatory state accompanies aging.405 Research conducted by Puzianowska-Kuźnicka et al.406 has shown that two pro-inflammatory cytokines—IL-6 and C-reactive protein (CRP), increase in the elderly in an age-dependent manner, highlighting the connection between chronic inflammation and aging. Nakaya et al.407 conducted studies that revealed an age-related disparity in antibody response following influenza vaccination, irrespective of race or gender, and found that older adults (>65 years) had a relatively reduced B-cell response after vaccination compared to that of younger adults (<65 years). However, these older adults displayed increased monocytes and activated cytotoxic NK cells. Notably, monocytes were also increased in older adults prior to vaccination, suggesting that the baseline state of the immune system in older adults (characterized by age-related low inflammation) is a potential mechanism for the reduced vaccine response.
Age-dependent low-grade chronic inflammation can arise from various potential sources. First, with age, the accumulation of damaged cells and the release of macromolecules such as advanced glycation end-products (AGE) continue to activate the innate immune system and trigger the release of diverse pro-inflammatory factors. Second, immune aging in the elderly may also increase the burden of antigen exposure. For instance, chronic CMV infection not only continuously stimulate the immune system, but also exacerbate both immune aging and inflammation.408 Compared to younger individuals, the elderly and the patients with chronic diseases have a pro-inflammatory status and are more prone to triggering cytokine storms following COVID-19 infection, leading to severe COVID-19 clinical symptoms.408 Immune aging is an inevitable major risk factor in various chronic diseases, particularly chronic inflammatory conditions including neurodegenerative diseases, cardiovascular diseases, and chronic kidney diseases. In addition, chronic inflammation plays a crucial role in the pathogenesis of various cancers that share a common factor—immune aging. The incidence of these pathological conditions significantly increases with age.409 Moreover, immune aging can explain the higher mortality rate observed in older COVID-19 patients due to ineffective T-cell responses and the inability to produce antibodies against SARS-CoV-2. Therefore, identifying markers that reshape immune response in older adults could serve as potential targets for patients with COVID-19 and other age-related ailments.410 Immune senescence leads to the disruption of immune checkpoint molecules, such as PD-1 and CTLA-4, and cytokines such as IL-6 and IL-1β.411,412 Furthermore, immune senescence is also closely associated with many autoimmune diseases. The incidence of autoimmune diseases appears to rise with age. In turn, autoimmune diseases such as rheumatoid arthritis may also accelerate the process of aging. On one hand, aging induces a state of age-dependent low-grade chronic inflammation, which in turn triggers immune suppression. This exacerbates immune senescence and increases the prevalence of various ailments among older individuals, including chronic inflammatory conditions that further hasten human aging.409
Age-related immune senescence and the presence of chronic inflammation in older individuals present significant challenges for disease treatment. Despite the availability of vaccines specifically designed for this population, their overall effectiveness remains inadequate, particularly in infectious diseases such as influenza. Researchers are exploring various therapeutic approaches targeting the pre-vaccination aging characteristics such as inflammation, immune senescence, and mitochondrial dysfunction. Notably, mRNA vaccines have demonstrated promising benefits during the COVID-19 pandemic, including enhanced efficacy among elderly individuals.413
During early childhood, the thymus predominantly harbors a substantial population of naive T cells, which subsequently undergo maturation processes. During this period, when encountering pathogens, memory T cells gradually accumulate and overtake the original T cells.404 Throughout the process of maturation from infancy to adulthood, a gradual decline is observed in the overall percentage of lymphocytes, as well as in the absolute count of T and B cells. In addition, there is a decrease in the number of NK cells. Interestingly, elderly individuals exhibit significantly higher levels of NK cells, pro-inflammatory cytokines (INF-α, IL-6), and monocyte chemoattractant protein-1 (MCP-1), while demonstrating lower levels of epidermal growth factor (EGF).414
The clinical trials of mRNA vaccines are typically conducted across diverse age cohorts, including pediatric, adult, and geriatric populations (Table 4). This may be attributed to variations in their distinct immunological profiles. For instance, mRNA-1273 developed by ModernaTX, Inc. underwent clinical trials in different phases. Initially, it was tested in Phase I (NCT04283461) and Phase III clinical trials (NCT04470427) for individuals aged 18 years and older. Subsequently, it underwent Phase II and III trials for adolescents aged 12 to 18 years (NCT04649151), as well as children aged 6 months to 11 years (NCT04796896). Encouragingly, older adults including participants aged 71 years and older exhibited comparable immune responses to younger adults upon receiving the second dose of the mRNA-1273 vaccine (NCT04283461).415 BNT162b2, developed by BioNTech SE, has encompassed age groups and has completed several clinical trials (NCT04368728, NCT04754594, NCT04816643). The findings revealed a decrease in immunogenicity of BNT162b1 or BNT162b2 with advancing age, resulting in a comparatively attenuated overall humoral response among older adults (aged 65–85 years) compared to their younger counterparts (aged 8 to 55 years). Encouragingly, the administration of the second dose of these vaccines could enhance antibody responses in both young and elderly adults (NCT04368728).416
Table 4.
Clinical trials were conducted according to different age groups
| mRNA | Disease | mRNA encoded protein | Enrollment (actual) | Ages | Sex/gender | Phase | NCT number | Study started (actual) | Status | Sponsor |
|---|---|---|---|---|---|---|---|---|---|---|
| mRNA-1273 | COVID-19 | SARS-CoV-2 spike protein | 30415 | 18 years and older (adult, older adult) | All | Phase III | NCT04470427 | 2020-07-27 | Completed | ModernaTX, Inc. |
| 4331 | 12 years to 18 years (child, adult) | All | Phase II/III | NCT04649151 | 2020-12-09 | Active, not recruiting | ||||
| 11950 | 6 months to 11 years (child) | All | Phase II/III | NCT04796896 | 2021-03-15 | Completed | ||||
| BNT162b1, BNT162b2 | COVID-19 | SARS-CoV-2 receptor-binding domain, SARS-CoV-2 spike protein | 47079 | 12 years and older (child, adult, older adult) | All | Phase II/III | NCT04368728 | 2020-04-29 | Completed | BioNTech SE |
| BNT162b2 | SARS-CoV-2 spike protein | 16385 | 12 years and older (child, adult, older adult) | All | Phase III | NCT04955626 | 2021-07-01 | Completed | ||
| 683 | 18 years and older (adult, older adult, healthy pregnant women) | Female | Phase III | NCT04754594 | 2021-02-16 | Completed | ||||
| 11837 | 6 months to 15 years (child) | All | Phase II/III | NCT04816643 | 2021-03-24 | Completed | ||||
| 0 | 72 days to 102 days (child) | All | Phase I | NCT05630352 | 2025-01-06 (estimated) | Withdrawn | ||||
| mRNA-1345 | RSV | RSV prefusion stabilized F (preF) glycoprotein | 36557 | 60 years and older (adult, older adult) | All | Phase II/III | NCT05127434 | 2021-11-17 | Active, not recruiting | ModernaTX, Inc. |
| 3800 | 50 years and older (adult, older adult) | All | Phase III | NCT05330975 | 2022-04-01 | Active, not recruiting | ||||
| 210 (estimated) | 5 months to 24 months (child) | All | Phase I | NCT05743881 | 2023-02-15 | Active, not recruiting | ||||
| 1150 (estimated) | 18 years and older (adult, older adult) | All | Phase III | NCT06067230 | 2023-10-06 | Recruiting | ||||
| 340 (estimated) | 2 years to 17 years (child) | All | Phase II | NCT06097299 | 2023-10-24 | Active, not recruiting | ||||
| 360 (estimated) | 18 years to 40 years (adult, pregnant women and infants) | All | Phase II | NCT06143046 | 2023-11-15 | Recruiting |
RSV respiratory syncytial virus
Note: Vaccination safety and immunogenicity are closely related to age, and clinical trials at different ages are needed to determine efficacy
Epidemiological data indicates that individuals aged 60 years and above exhibit a disproportionately higher propensity for severe clinical symptoms and increased rates of hospitalization following SARS-CoV-2 infection during the COVID-19 pandemic. While current vaccines have demonstrated favorable efficacy in younger cohorts, they fail to elicit sustained immunity in the elderly population. Despite previous discussions on potential reasons for this phenomenon, further exploration is warranted to uncover deeper underlying factors. The objective of such investigation is to ultimately develop highly effective vaccines with minimal adverse reactions specifically tailored for older adults. To further enhance the efficacy of the vaccine, in addition to exploring novel strategies against immunosenescence and inflammation, it is also imperative to develop and employ more potent adjuvants.405 During the COVID-19 pandemic, mRNA vaccines have accumulated valuable insights into age-related efficacy and adverse reactions of vaccination, thus paving the way for their extensive application in other infectious diseases, tumors, and beyond.
Gender
Gender dimorphisms are pervasive in human development and physiology. The phenotypic discrepancies of gender derive from a complexed mixture of endogenous and exogenous factors, particularly hormones such as estrogen and androgen. In the era of precision medicine, it becomes crucial to consider the relationship between gender and immune responses for disease prevention and treatment.417 The existing literature suggests that male patients demonstrate a higher propensity for developing severe clinical symptoms of SARS-CoV-2 compared to their female counterparts.418 This phenomenon may be attributed to the immunomodulatory effects of estrogen, whereas androgen and progesterone exert immunosuppressive actions.404 However, the relationship between gender and disease severity is intricate, encompassing the influence of sex hormones, expression of X-linked genes, and other immune regulatory factors. Therefore, a comprehensive understanding of the COVID-19 infection and symptom severity can be achieved by investigating genetic, hormonal, and specific behavioral factors associated with gender and age. Moreover, such researches can contribute to the future development of mRNA vaccines.418
Gender plays a pivotal role in determining the risk of HIV infection and its pathogenic progression. Reproductive disparities between genders contribute to an augmented susceptibility of women toward HIV infection. However, during the early stages of HIV infection, women exhibit heightened activation of CD8+ T cells and increased expression of interferon-stimulated genes. Nonetheless, post-HIV infection, women face an elevated risk of non-acquired immunodeficiency syndrome (AIDS) morbidity such as cerebrovascular events.419
Compared to female mice, male mice demonstrate a higher susceptibility to influenza B virus (FLUBV), mirroring the epidemiological features observed in humans. Cardenas-Garcia et al.420 conducted a study encompassing various vaccine designs and found that female mice exhibited more robust lgG and lgA responses and increased CD4+ T cells compared to their male counterparts. Furthermore, they reported that both the quality and quantity of immune response were influenced by factors such as vaccine platform, gender, and inclusion of adjuvants, thereby impacting the vaccine efficacy against FLUBV. The findings suggest that, in general, males demonstrate higher morbidity and mortality rates from viral infectious diseases.421 However, it is noteworthy that in certain infectious diseases such as measles and dengue, females actually exhibit higher morbidity and mortality rates.404
For cardiovascular disease, there are well-established modifiable risk factors such as smoking, obesity, diabetes, hypertension, and hyperlipidemia. In addition to these factors, non-modifiable risk factors such as genetics, age, and gender play a significant role in cardiovascular disease development. The risk of cardiovascular disease significantly increases for males aged 45 years and above, as well as for females who undergo oophorectomy or experience natural menopause.422
It is widely acknowledged that sex hormones play a pivotal role in hormone-dependent organ diseases, such as breast and ovarian cancers in females, as well as prostate and testicular cancers in males. The incidence rates of various non-reproductive organ cancers may also exhibit a significant association with gender.423 Li et al.417 conducted an analysis utilizing data from the UK Biobank and identified 119 diseases exhibiting a gender bias (P adjusted < 0.05). Notably, stomach, kidney and lung malignancies demonstrated higher incidence rates in men, while asthma displayed a higher incidence rate in women.
The immune system is intricately interconnected with cancer treatment, and variations in hormonal and X chromosome gene expression can lead to different immune responses among distinct populations of innate and adaptive immune cells.424 For example, in patients with KRAS mutations in lung adenocarcinoma, females demonstrate a greater abundance of CD4+ T and CD8+ T cells compared to males.425 The study conducted by Thompson et al.426 revealed variations in the functionality of the transcription factor FOXO3, which is associated with tumor-infiltrating DCs tolerance, between genders. These disparities may serve as crucial considerations for augmenting cancer immunotherapy. Observations from clinical trial indicate that immune checkpoint blockade confers greater benefits in the treatment of male cancer patients.427 However, certain studies have reported no statistically significant gender-based differences in the efficacy of ICIs.428 Additionally, clinical investigations have demonstrated that ICIs amplified therapeutic outcomes in female patients when combined with Cisplatin and/or radiotherapy.429 These conflicting experimental findings suggest that the underlying mechanism influencing gender disparities in cancer incidence rates and treatment options are multifaceted and complex.
In summary, women usually exhibit stronger immune responses against infectious diseases and malignant tumors than man. In addition, following vaccination, women demonstrate superior antibody responses, thereby reducing their susceptibility to infections and lowering the mortality rates. However, the better immune response of women also predisposes them to a fourfold increased risk of developing autoimmune diseases.430,431 For example, the incidence ratios of multiple sclerosis, scleroderma, and rheumatoid arthritis in females compared to males range from 2:1 to 3:1. However, the gender distribution of systemic lupus erythematosus is remarkably skewed with a ratio of 9:1, representing the most pronounced gender-based disparity among autoimmune diseases.432 The etiology of the gender disparity in autoimmune disease incidence is multifaceted and intricate. For instance, the potential exacerbating effect of estrogen on systemic lupus erythematosus, does not preclude its potential immune-protective effect on rheumatoid arthritis.433 It is noteworthy that the gender-specific variations in intestinal microbiota may underpin certain gender-related physiological and pathological conditions, thus providing a biological basis for such disparities.434
Despite the higher prevalence of autoimmune diseases in women, there is no significant gender disparity in the incidence of immune-related adverse events (IrAEs) among patients receiving ICIs for skin, gastrointestinal, and lacrimal gland-related conditions. However, notable gender differences exist in specific endocrine-related IrAEs. For example, thyroid dysfunction is more commonly seen in women, whereas hypophysitis is more frequently observed in men.432
Previously, the role of gender in the development and treatment of diseases was overlooked. For instance, in 1977, the U.S. FDA recommended excluding women of reproductive age from Phase I and early Phase II trials under the guise of safeguarding women and children. Nevertheless, it led to a lack of research and insufficient awareness.432 Fortunately, in 2015, the National Institutes of Health (NIH) issued a directive urging researchers to incorporate gender as a biological variable into their research designs, data collection protocols, and outcome analysis.435
Currently, there are relatively few mRNA vaccines designed specifically for gender differences. However, the disparities in immune responses resulting from differences in X-linked genes and sex-based hormonal variations do indeed exist, leading to variations in disease susceptibility and clinical manifestations as well as vaccine efficacy. These differences are increasingly being recognized.436 Research into the interactions of age and gender may aid in the design of personalized mRNA vaccine treatment strategies tailored to individual patient.422
Pathological condition and vulnerable population
In addition to age and gender, there are other intricate factors affect the efficacy of vaccination. Levin et al.437 found that individuals aged over 65 years, males, and groups with immunosuppression-related conditions exhibited significantly lower levels of humoral immunity 6 months after receiving the second dose of BNT162b2. Furer et al.438 revealed that patients with autoimmune inflammatory rheumatic disorders displayed a lower humoral response to the two-dose BNT162b2 mRNA vaccine regimen compared to immunocompetent individuals. Although seropositive levels were restored in immunocompetent individuals after receiving the third vaccine dose, only 80.47% of patients experienced restoration of their humoral response. Notably, all patients who received anti-cytokine biologics, Methotrexate monotherapy, Abatacept and Janus kinase inhibitors had their humoral immunity restored. However, only one-third of patients who received Rituximab were able to regain immune protection. The clinical trials regarding vaccine administration in patients receiving immunosuppressants are listed in Table 5.
Table 5.
Ongoing and completed clinical trials of mRNA vaccines for the treatment of immunosuppressant patients/patients with autoimmune diseases
| Background disease | Trial population | mRNA vaccine | Immunosuppressant | Phase | NCT number | Sponsor | Last update posted |
|---|---|---|---|---|---|---|---|
| Kidney transplant | Recipients who had a kidney transplant at least 6 months ago | Omicron XBB.1.5 vaccine | Everolimus | Phase IV | NCT05924685 | University Medical Center Groningen | 2024-03-04 |
| Relapsing multiple sclerosis (MS) | Ofatumumab treated participants | Pfizer or Moderna mRNA COVID-19 vaccine | Ofatumumab | Phase IV | NCT04878211 | Novartis Pharmaceuticals | 2024-02-08 |
| Secondary progressive MS | Patients with secondary progressive MS | COVID-19 modRNA vaccine | Siponimod | Phase IV | NCT04792567 | Novartis Pharmaceuticals | 2022-12-14 |
| Malignant neoplasms brain | Glioblastoma multiforme patients treated with Temozolomide | Cytomegalovirus (CMV) pp65-LAMP mRNA-loaded dendritic cell (DC) vaccine | Basiliximab | Phase I | NCT00626483 | Gary Archer Ph.D. | 2021-03-09 |
| Inflammatory bowel disease (IBD) | Patients with IBD | COVID-19 vaccine | Non-systemic immunosuppressive therapy (Mesalamine/Vedolizumab/Vedolizumab combination therapy with Methotrexate or Azathioprine); systemic immunosuppression (Azathioprine/Infliximab/Golimumab/Adalimumab/Certolizumab/Ustekinumab/Tofacitinib/Corticosteroid) | Observational | NCT04818892 | University of Wisconsin, Madison | 2022-12-07 |
| Glioblastoma (GBM) | GBM patients who underwent resection, Temozolomide (TMZ) therapy, and radiation therapy were also pre-treated with tetanus and given Basiliximab | CMV-specific DC vaccine | Basiliximab | Phase II | NCT02366728 | Mustafa Khasraw, MBChB, MD, FRCP, FRACP | 2023-06-08 |
| Autoimmune disease | 5 autoimmune diseases in adults [systemic lupus erythematosus (SLE)/rheumatoid arthritis (RA)/MS/systemic sclerosis/pemphigus], 4 autoimmune diseases in pediatric participants [systemic lupus erythematosus (SLE)/juvenile idiopathic arthritis (JIA)/pediatric-onset multiple sclerosis (POMS)/juvenile dermatomyositis (JDM)] | COVID-19 vaccine | / | Phase II | NCT05000216 | National Institute of Allergy and Infectious Diseases (NIAID) | 2024-03-26 |
| Immunosuppress/systematic autoimmunity | Hematological malignancy + immunosuppressant/systematic autoimmunity | COVID-19 vaccine, diphtheria/tetanus toxoids vaccine | e.g., Rituximab, Ocrelizumab, Ofatumumab | Phase III | NCT05415267 | Kirby Institute | 2024-03-15 |
| Solid organ transplant | Kidney transplant recipients, liver transplant recipients | COVID-19 vaccine | Tacrolimus | Phase II | NCT05077254 | NIAID | 2024-03-29 |
| MS | MS patients on immunotherapy | COVID-19 vaccine | Ofatumumab, Ocrelizumab, Fingolimod, Siponimod | Observational | NCT05060354 | Brigham and Women’s Hospital | 2024-01-25 |
| Breast cancer, malignant melanoma | Patients with metastatic breast cancer or malignant melanoma | DC vaccine | Cyclophosphamide (background drugs, as vaccine adjuvants, commonly used as anticancer drugs) | Phase I | NCT00978913 | Inge Marie Svane | 2015-08-19 |
| Immunocompromised, immunosuppressed | Patients with immunocompromising conditions in the United States | COVID-19 vaccine (BNT162b2) | / | Observational | NCT05020145 | Pfizer | 2024-02-12 |
| Brain stem glioma | Brain stem glioma patients treated with radiotherapy and Temozolomide | DC vaccine | / | Phase I | NCT03396575 | University of Florida | 2023-11-18 |
Note: Follow all entries found under “Search for: mRNA vaccine, immunosuppressant |Card Results| ClinicalTrials.gov”
Leston et al.439 classified immunosuppression and correlated it with the mortality rate of COVID-19. Their findings revealed that patients with solid organ transplants and malignant tumors exhibited a significantly higher mortality risk following COVID-19 infection compared to that of immunocompetent individuals. Furthermore, in comparison to immunocompetent individuals, patients with rheumatological conditions and HIV demonstrated a slightly elevated mortality risk. The study established a close association between specific subgroups of immunosuppression and the mortality risk associated with COVID-19 infection. This categorization method holds substantial reference value for targeted treatment strategies and focused vaccination prevention. Meeraus et al.440 discovered that individuals with comorbidities and vulnerable populations exhibited reduced vaccine effectiveness following administration of AZD1222 (ChAdOx1 nCov-19) in comparison to those with normal immune systems, while immunosuppressed individuals demonstrated the lowest vaccine effectiveness after the initial dose of AZD1222. Obeid et al.441 conducted a comparative analysis on the persistence of humoral responses against SARS-CoV-2 and its variants of concern (VOCs) in immunocompromised patients who received two doses of BNT162b2 or mRNA-1273 vaccines, as well as in healthy individuals. The findings revealed that approximately 50% of patients with solid tumors and hematological malignancies, about 70% of autoimmune disease patients and solid organ transplant recipients, and 40% of healthy individuals had lost protective nAbs 6 months after vaccination. Moreover, it was observed that the durability of binding IgG anti-spike antibodies against SARS-CoV-2 was four to nine times greater than that of nAbs. Furthermore, variations were noted in the immune efficacy between different mRNA vaccines, with BNT162b2 inducing lower magnitude and shorter duration of nAbs compared to mRNA-1273. Therefore, it is imperative to select appropriate mRNA vaccines for diverse vulnerable populations in order to ensure effective administration of vaccinations.
In addition to COVID-19 virus infection, similar scenarios seem to arise in other infectious viruses as well. A study conducted by Mbonde et al.442 revealed that immunosuppressed individuals with neuroinvasive West Nile virus (NWNV) exhibited more severe clinical manifestations compared to immunocompetent populations, thereby indicating a worse prognosis and higher risk of adverse reactions. The survival rate of sepsis survivors is significantly diminished following secondary infection, primarily due to the prolonged state of immunosuppression that ensues after acute infection. A comprehensive study conducted by Liao et al.443 revealed a close association between IL-10 production by Siglec-F+ neutrophils and the suppression of T-lymphocyte activity in this phenomenon. Furthermore, their findings indicated that depletion of neutrophils could enhance T-lymphocyte proliferation and improve T-lymphocyte activity, thereby ameliorating the survival rate among immunosuppressed mice subjected to secondary infection. However, further investigation is required to assess the systemic effects associated with neutrophils depletion.
Persistent immune suppression is observed in people with HIV, which is closely associated with low CD4 counts or abnormal CD4/CD8 ratios. The prevention and treatment of cancer in HIV-infected individuals face challenges due to disparities in health status.444 Evidences suggested that individuals infected with HIV might undergo premature aging. The research conducted by Gianesin et al.445 further demonstrated that this phenomenon extended to HIV-infected children, resulting in accelerated biological and immunological senescence, particularly affecting the CD8+ cell subpopulation. Vergori et al.446 discovered that the administration of a third dose of the COVID-19 mRNA vaccine to individuals with HIV could elicit a robust immune response. However, the magnitude of the SARS-CoV-2-specific T-cell response in HIV-positive subjects was comparatively lower than that observed in the HIV-negative control group. In addition, they provided quantitative analysis regarding the potential augmentation of immune levels through extra booster doses for individuals with HIV.
Pregnant women and infants are also considered vulnerable populations. Antibodies derived from COVID-19 vaccines can offer protection against severe virus infections in newborns through the transfer of placental antibodies. There is limited data on the antibody levels of infants, particularly preterm infants, following maternal vaccination. Therefore, Kachikis et al.447 conducted a study which revealed that there was no disparity in maternally derived SARS-CoV-2 anti-spike IgG levels between full-term and preterm infants. However, these levels were closely associated with the concentration of the maternal anti-spike antibodies. In addition, it was observed that receiving two or fewer doses of the COVID-19 vaccine might not provide optimal immune protection for pregnant women, nor for infants via cord blood. Consequently, careful consideration should be given to the timing of vaccination for pregnant women.
Adverse reaction
Johnston et al.448 discovered an adverse reaction manifested as a delayed hypersensitivity at the injection site subsequent to administration of the Moderna COVID-19 vaccine. Cappelletti-Montano et al.449 discovered that the majority of severe adverse reactions reported after COVID-19 vaccination were associated with cardiac complications, particularly among male adolescents. In addition, thrombosis and dyspnea emerged as prevalent serious symptoms following COVID-19 vaccination. In contrast, the adverse reactions after vaccination with HPV and influenza seemed to be more diverse. Dizziness, loss of consciousness, dyspnea, and convulsions occurred after HPV vaccination, and Guillain-Barré syndrome occurred after influenza vaccination. Vaccination with COVID-19 vaccine (BNT162b2 mRNA or mRNA-1273) can also be associated with the development of various autoimmune diseases, such as autoimmune hepatitis or nephritis, rheumatoid arthritis, and new-onset systemic lupus erythematosus. Heil450 endeavored to investigate the underlying factors contributing to a diverse range of life-threatening complications in patients infected with SARS-CoV-2. Although the COVID-19 pandemic seems to be receding, the lessons learned from the development of COVID-19 mRNA vaccines are expected to have implications for non-COVID-19 diseases.
The study conducted by Pettini et al.451 demonstrated that multiple administration of mRNA-1273 vaccine effectively enhanced the specific memory B-cell and antibody responses against SARS-CoV-2 in allogeneic hematopoietic cell transplantation patients. Furthermore, they emphasized the significance of repeated vaccination. The phenomenon of extensive and consecutive immunization has become apparent owing to repetitive vaccinations intended to augment vaccine efficacy, along with advancements in vaccines designed for diverse diseases. Nonetheless, this intricacy presents difficulties in evaluating vaccine safety accurately. Interactions between vaccines have the potential to produce favorable or detrimental outcomes alike. Mawson and Croft452 postulated that multiple vaccinations might induce activation of the retinoid cascade, resulting in a sequence of unfavorable reactions. The uncertainties of the risks associated with vaccination necessitate further research to optimize vaccine safety.
The safety and immunogenicity profiles following vaccination were assessed in clinical trials through the administration of two distinct vaccine formulations in a predetermined sequence. For example, the Phase I trials of the investigational vaccines BNT164a1 and BNT164b1 have recently commenced with volunteers who have previously received the BCG vaccine (NCT05547464).453 Several clinical trials are currently underway to assess the safety and immunogenicity of co-administering two vaccines simultaneously (Table 6). Moreover, mRNA vaccines can be engineered to encode several epitopes from multiple pathogens simultaneously.454 Currently, there is ongoing development of multi-component vaccines aimed at achieving progressively more comprehensive disease prevention (Table 7).
Table 6.
Concurrently administered vaccines
| Vaccine | Disease | Study started (actual) | Phase | Status | Sponsor | NCT number |
|---|---|---|---|---|---|---|
| 9vHPV + mRNA-1273 | Papillomavirus infection and COVID-19 | 2022-03-28 | Phase III | Completed | Merck Sharp & Dohme LLC | NCT05119855 |
| mRNA-1345 + influenza vaccine | RSV and influenza | 2023-09-25 | Phase III | Active, not recruiting | ModernaTX, Inc. | NCT06060457 |
| V110/V114 + mRNA-1273 | Pneumococcal infection and COVID-19 | 2022-01-12 | Phase III | Completed | Merck Sharp & Dohme LLC | NCT05158140 |
| ARCT-2303 + influenza vaccine | COVID-19 and influenza | 2024-03 (estimated) | Phase III | Not yet recruiting | Arcturus Therapeutics, Inc. | NCT06279871 |
| BNT162b2 (Omi XBB.1.5) + RIV | COVID-19 and influenza | 2024-01-31 | Phase II | Active, not recruiting | Pfizer | NCT06237049 |
HPV human papillomavirus, RSV respiratory syncytial virus
Table 7.
Multi-component vaccines
| Vaccine | Disease | Study started (actual) | Phase | Status | Sponsor | NCT number |
|---|---|---|---|---|---|---|
| mRNA-1045 | Influenza and RSV | 2022-10-14 | Phase I | Completed | ModernaTX, Inc. | NCT05585632 |
| mRNA-1230 | Influenza, RSV and COVID-19 | 2022-10-14 | Phase II | Completed | ModernaTX, Inc. | NCT05585632 |
| mRNA-1083 | Influenza and COVID-19 | 2023-04-14 | Phase I/II | Active, not recruiting | ModernaTX, Inc. | NCT05827926 |
| mRNA-1073 | Influenza and COVID-19 | 2022-05-13 | Phase I/II | Completed | ModernaTX, Inc. | NCT05375838 |
RSV respiratory syncytial virus
Conclusions and prospects
Challenges in mRNA drug application
Despite substantial advancements in the field, effectively and safely delivering mRNA therapeutics to the target site remains a significant challenge. The half-life of direct intravenous administration of mRNA is very short due to their susceptibility to degradation by nucleases. The negative charge and high molecular weight of mRNA hinder cellular transcytosis across the biological membrane. Additional barriers include high renal clearance, non-specific tissue distribution, and off-target effects. In addition, mRNA that enter cells face the challenge of escaping from endosomes.455 Furthermore, exogenously introduced mRNA can be recognized by the TLRs and cytosolic nucleic acid receptors, triggering innate immune responses and accelerating the deactivation of IVT mRNA within the body.181
Although the application of mRNA delivery systems, LNPs in particular, mitigates certain of the above obstacles, it is important to remain cautious of the potential hazards associated with delivery vehicles, such as inflammation, immunogenicity, and cytotoxicity.105 In addition to the toxicity caused by the delivery system, it has been discovered that the production methods, routes of administration, and even proteins generated by the complexed mRNA drugs present toxicity concerns.456
Main reasons for failed mRNA application
Following intravenous or intramuscular administration, mRNA LNPs are extensively distributed in the liver and spleen, potentially resulting in pathological alterations within these organs, such as the disruption of hepatic cell regulation of fatty acid metabolism. In addition, mRNA nanomedicines may elicit adverse immune reactions, such as systemic complement activation, hypersensitivity responses, and cytokine-mediated effects post-vaccination.98,456 The adverse immunogenicity caused by LNPs, besides the lipid components, is closely associated with their size, charge, and aggregation characteristics. Smaller LNPs are more advantageous in mitigating the activation of undesired immune responses. On the other hand, it is important to investigate the translation accuracy of nucleotide-modified mRNA therapeutics, as unintended protein synthesis may result in unforeseen outcomes.457 Mulroney et al.458 discovered that the use of m1Ψ-modified IVT mRNA led to a notable increase in the +1 ribosomal frameshift during translation. Understanding the translation process of nucleotide-modified mRNA is essential for future mRNA design and optimization to mitigate potential reductions in efficacy and increases in toxicity.
The success application of mRNA therapies is intricately linked to the complexity and public understanding of targeted disease. Take the extensively studied cancer mRNA vaccine as an example. To date, there is few Phase III mRNA vaccine available for tumors, which may be due to the following reasons: (1) The mRNA encoding a single tumor antigen may inadvertently induce mutations in tumors to evade immune response, necessitating the need for developing mRNA vaccines capable of simultaneously targeting multiple TAAs or TSAs; (2) The surface antigens are distinct across different tumor types, calling for personalized tumor-mRNA vaccines for individual patient. However, the production and implementation of personalized vaccines are often associated with high costs and pose challenges to their widespread clinical application; (3) Tumors exert immunosuppressive effects by impeding the recognition of neoantigens and suppressing the viability of antitumor immune cells, emphasizing the importance of meticulous selection and application of adjuvants for potentiated immune responses; (4) The efficacy of mRNA vaccines is largely limited by its instability, inefficient delivery, and poor transfection efficiency. In these cases, iterative optimization of the mRNA molecule and delivery vehicle is needed; (5) Tumor patients often exhibit compromised immune responses due to a multitude of factors, including advanced age, concurrent medication (e.g., chemotherapy), and comorbidity (e.g., HIV). Hence, in addition to bolstering the immune response with adjuvants, vigilance against potential adverse reactions should be taken into serious consideration; (6) The formidable challenge of eradicating advanced tumors using mRNA vaccines alone necessitates the integration of multiple therapies for synergistic therapeutic effects, such as adoptive cell therapy.459
Efforts and improvements made to address these challenges
The current landscape of mRNA vaccines with promising clinical prospects often comprises three components: the mRNA sequence encoding target protein, delivery vector, and adjuvant.460 In certain cases, the mRNA carrier itself functions as vaccine adjuvant.460 Notably, although the increased immunogenicity of mRNA drugs boosts immune response, it hampers mRNA translation and suppresses target protein production. In this consideration, the technology of immune-silencing IVT mRNA by replacing naturally occurring nucleosides into the mRNA sequence has been developed.461 Employing modified mRNA represents a strategic approach to both enhance protein expression and mitigate aberrant immunogenicity.462 In addition, by optimizing the mRNA sequence, such as introducing multiple adenines at the Poly (A) tail, mRNA stability can be further enhanced with the protein translation efficiency improved. Moreover, purification methods including cellulose and fast protein liquid chromatography can be employed to eliminate the immunogenic double-stranded RNA generated during mRNA manufacturing, thereby reducing the inflammatory responses unfavorable to mRNA stability.463
In the context of immunotherapy, to avoid the insufficient activation of APCs, especially DCs, with nucleoside-modified immune-silenced mRNA following vaccination,115 adjuvants can be incorporated into the delivery system. An ideal adjuvant can lower the dose of antigen-encoding mRNA per vaccine dose, reduce the frequency of vaccination, enhance the robustness and specificity of immune responses, and in certain cases, assume the role of a carrier for the efficient delivery of mRNA and contribute to the stabilization of the vaccine formulation.464,465 Gu et al.466 used TLR2/6 agonist Pam2Cys (a simple synthetic metabolizable lipoamino acid) as an adjuvant to facilitate mRNA encoding CT26 neoantigen peptide or SARS-CoV-2 spike protein antigen for induction of effective immune responses against cancer or infectious diseases. On the other hand, the adjuvant can be incorporated directly into the mRNA sequence. Li et al.395 developed a multiply adjuvanted mRNA vaccine by fusing antigen-encoding mRNA with C3d (the terminal degradation product of mammalian complement component C3)-based mRNA adjuvant, which evoked a protective immune response against SARS-CoV-2 at a dosage ten times lower. Of note, an increasing number of studies have revealed that adjuvant not only enhances the magnitude of immune response to vaccines, but guide the type of adaptive immune response.467 Zhu et al.468 fabricated mRNA LNPs and investigated the differential immune responses elicited by mRNA LNPs supplemented with the natural STING agonist 2'3’-cyclic GMP-AMP (cGAMP) and/or the mucosal adjuvant alpha-galactosylceramide (αGC). The results indicated that mRNA LNPs containing cGAMP alone induced a stronger cellular immune response than mRNA LNPs containing both cGAMP and αGC, which was consistent with higher levels of IgG and IgG2a antibody responses, as well as better protection against homologous viral challenge. The study also demonstrated that mRNA LNPs containing cGAMP could accumulate CD4+ T cells in the lung, while mRNA LNPs containing αGC could enlarge the iNKT cells in the spleen. Activated iNKT cells may exhibit potent cytotoxicity, indicating the paramount importance of the judicious selection and application of adjuvants. At the cellular level, it is well-established that Actinidia eriantha polysaccharide (AEPS) elicits a mixed CD4+ Th1 and CD4+ Th2 immune response, while Alum induces a strict Th2 response. Nevertheless, many intricate molecular mechanisms remain unclear. Du et al.469 conducted a comparative analysis of the innate immune responses elicited by the two adjuvants, and found that AEPS induced higher mRNA expression levels of C-X-C motif chemokine ligand (CCL) 2, CCL3, CCL4, CXCL10, IL-12β, IL-23α in immune effector process, and rapidly recruited neutrophils and monocytes. In comparison, Alum induced IL-7A, IL-17F, and IL-17RA, and recruited neutrophils and eosinophils.
The delivery and storage challenges of mRNA can be resolved through optimization of the delivery vehicle.115 To date, a plethora of delivery systems for mRNA have been developed. Among them, LNPs has progressed in clinical practice most rapidly. LNPs not only possesses the capacity to encapsulate and transport mRNA, but also shields the mRNA from nuclease degradation. However, the use of LNPs still presents certain issues, such as the potential damage to cell membranes and organelles due to the cationic characteristics of the ionizable lipid. The development of biodegradable ionizable lipids appears to offer some solutions to this problem.102,105 It is crucial to recognize that no ionizable lipid is universally optimal for all nucleic acid formats, and variations in nucleic acid modification, size, and structure can impact the effectiveness of LNPs delivery.470 On the other hand, the off-target accumulation of mRNA drugs may lead to suboptimal outcomes. In this case, various stimulus-responsive nanotechnologies have been developed to utilize specific internal stimulus within the cellular microenvironment, including the expression of specific enzymes, increased levels of ROS, hypoxic environments, and changes in the pH or temperature, as well as external stimulus such as light irradiation, magnetic fields, and ultrasound. These stimuli-responsive nanoplatforms have the potential to enhance tissue targeting, promote exosome escape of mRNA, improve the therapeutic effects of mRNA therapy, and reduce toxicity and side effects.471 The active targeting of mRNA drugs to desired biological sites through LNPs surface modification is also widely used. Several preclinical studies have already demonstrated the successful delivery of mRNA LNPs to target tissues and cells.472–474 For instance, Kim et al.475 utilized LNPs modified with tumor-targeting peptides to encapsulate mRNA-encoding PETN. This PETN mRNA LNPs specifically induces ICD in mice 4T1 breast tumor, thereby eliciting effective antitumor immune responses. Tang et al.476 engineered LNPs with sialic acid for DCs targeting and efficient endosomal escape. Furthermore, it is anticipated that the next-generation LNPs with the ability to target drugs specifically to distinct intracellular organelles, such as mitochondria and the nucleus, will serve as an innovative paradigm.472 In addition to the optimization strategies mentioned above, the delivery system for mRNA can be optimized to enhance the precision of mRNA-based therapies for diseases by adjusting factors such as drug dissolution, diffusion rate, pharmacokinetics, toxicity, and half-life.477
Potential of mRNA drugs to treat various diseases
The mRNA vaccine has emerged as a forefront line in disease prevention and treatment.276 The rapid development and regulatory approval of mRNA vaccines against SARS-CoV-2 during the COVID-19 pandemic have demonstrated the advantages of mRNA technology,478 exerting a profound influence on its application in infectious diseases and cancer immunotherapy. The Phase III clinical trial of SARS-CoV-2 mRNA vaccine is currently the most clinically available. Additionally, a series of mRNA vaccines against other infectious diseases are under development. For example, the Phase I study (NCT04528719) of mRNA-1345 encoding the prefusion stabilized F (preF) glycoprotein of RSV demonstrates favorable tolerability profiles in both young and elderly individuals, providing the foundation for further investigations where high-risk adults and solid organ transplant patients are enrolled (Phase III, NCT06067230).479 Meanwhile, compared to mRNA-1345, the development of mRNA-1010 for seasonal influenza is progressing at a faster pace, which has successfully completed a Phase III trial with 22,510 adults aged over 50 years (NCT05566639). mRNA-based immunotherapies have also been extensively researched for antibacterial and anti-parasite applications. Of note, aggressive fungal infections actually result in a higher annual mortality than malaria, yet there are very few mRNA-based treatments available for fungal infections. We aim to increase awareness of the potential of mRNA-based therapies for treating fungal diseases.480
In the context of antitumor immunotherapy, increasing mRNA vaccines has entered the clinic. For example, mRNA-4157 encodes up to 34 patient-specific tumor neoantigens is used in combination with Pembrolizumab for the treatment of melanoma, which has entered Phase IIb clinical trial (NCT03897881).481 Presently, the primary focus of research lies in the identification and design of mRNA sequences that target specific cancer antigens for the purpose of achieving personalized cancer mRNA vaccine therapy. Furthermore, while the majority of existing cancer mRNA vaccines are designed for therapeutic purposes rather than preventive measures, there is also significant potential for prophylactic cancer mRNA vaccines.181 Given the design flexibility and adaptability of mRNA, the development of personalized cancer mRNA vaccines to optimize antitumor effects based on diverse tumor types and individual patient variations represents a crucial avenue for future advancement.482
It should be mentioned that mRNA-based immunotherapy not only has the potential to enhance immune responses against a wide range of infectious diseases and cancers, but also offers the possibility of modulating the immune response for the treatment of autoimmune conditions. mRNA therapeutics designed for autoimmune diseases can be tailored to encode disease-specific antigens or immunosuppressive cytokines. For instance, the mRNA-6231 encoding HSA-IL2m, intended for the treatment of autoimmune disorders, has entered Phase I clinical trial (NCT04916431).221
In contrast to immunotherapy where the incorporation of adjuvants is imperative, it is of vital importance for non-immunotherapy to mitigate the immunogenicity of mRNA drugs and to amplify the expression of encoded target proteins.483 mRNA-based therapeutics function to replenish deficient proteins or substitute abnormal proteins that exhibit diminished functionality/activity. For example, mRNA therapies for OTCD (ARCT-810, Phase II, NCT05526066; MRT5201, Phase I/II, NCT03767270), PA (mRNA-3927, Phase I/II, NCT04159103), MMA (mRNA-3705, Phase I/II, NCT05295433), and myocardial infarction (AZD8601, Phase IIa, NCT03370887) have entered clinical trials (Table 2).
In this review, we summarize the preclinical and clinical progresses of mRNA drugs in both immunotherapy and non-immunotherapy, and highlight the importance of focusing on host-specific variations for maximized efficacy and safety. Although in the early stages of development, the therapeutic use of mRNA has demonstrated great potential in combating multiple diseases.484 The mRNA drugs described in this review primarily encompass traditional IVT mRNA. However, as mRNA technology constantly innovating and advancing, other forms of mRNA drugs, such as saRNA, trans-amplifying mRNA (taRNA), and circRNA, hold immense applicability. It is worth noting that successful clinical implementation of these next-generation mRNA drugs requires the involvement of suitable delivery vehicles and further modifications to the mRNA molecule itself for better tolerability and efficacy.485
Acknowledgements
This work was supported by the National Key R&D Program of China (2023YFC340200), the National Natural Science Foundation of China (No. 82404520, 82273862), the China Postdoctoral Science Foundation (2024M752834), and the China National Postdoctoral Program for Innovative Talents (BX20230321).
Author contributions
Conceptualization: Yingying Shi. Literature collection: Yingying Shi and Meixing Shi. Resources: Yi Wang. Writing—original draft: Yingying Shi and Meixing Shi. Writing—review and editing: Yingying Shi and Meixing Shi. Visualization: Yingying Shi and Meixing Shi. Supervision: Yi Wang and Jian You. Funding acquisition: Jian You and Yingying Shi. All authors have read and approved the article.
Competing interests
The authors declare no competing interests.
Footnotes
These authors contributed equally: Yingying Shi, Meixing Shi
Contributor Information
Yi Wang, Email: zjuwangyi@zju.edu.cn.
Jian You, Email: youjiandoc@zju.edu.cn.
References
- 1.Muslimov, A. et al. The dual role of the innate immune system in the effectiveness of mRNA therapeutics. Int. J. Mol. Sci.24, 14820 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bonelli, M. et al. Additional heterologous versus homologous booster vaccination in immunosuppressed patients without SARS-CoV-2 antibody seroconversion after primary mRNA vaccination: a randomised controlled trial. Ann. Rheum. Dis.81, 687–694 (2022). [DOI] [PubMed] [Google Scholar]
- 3.Mrak, D. et al. Heterologous vector versus homologous mRNA COVID-19 booster vaccination in non-seroconverted immunosuppressed patients: a randomized controlled trial. Nat. Commun.13, 5362 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Brenner, S., Jacob, F. & Meselson, M. An unstable intermediate carrying information from genes to ribosomes for protein synthesis. Nature190, 576–581 (1961). [DOI] [PubMed] [Google Scholar]
- 5.Wolff, J. A. et al. Direct gene transfer into mouse muscle in vivo. Science247, 1465–1468 (1990). [DOI] [PubMed] [Google Scholar]
- 6.Hou, X., Zaks, T., Langer, R. & Dong, Y. Lipid nanoparticles for mRNA delivery. Nat. Rev. Mater.6, 1078–1094 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Duke University. A safety and feasibility study of active immunotherapy in patients with metastatic prostate carcinoma using autologous dendritic cells pulsed with RNA encoding prostate specific antigen, PSA. https://clinicaltrials.gov/study/NCT00004211 (2013).
- 8.Heiser, A. et al. Autologous dendritic cells transfected with prostate-specific antigen RNA stimulate CTL responses against metastatic prostate tumors. J. Clin. Investig.109, 409–417 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.University Hospital Tuebingen. Induction of specific immune responses against melanoma-associated antigens in vivo by intradermal vaccination with stabilized tumor mRNA—a clinical Phase I/II trial. https://clinicaltrials.gov/study/NCT00204607 (2007).
- 10.Weide, B. et al. Direct injection of protamine-protected mRNA: results of a phase 1/2 vaccination trial in metastatic melanoma patients. J. Immunother.32, 498–507 (2009). [DOI] [PubMed] [Google Scholar]
- 11.Karikó, K., Buckstein, M., Ni, H. & Weissman, D. Suppression of RNA recognition by toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity23, 165–175 (2005). [DOI] [PubMed] [Google Scholar]
- 12.Dolgin, E. The tangled history of mRNA vaccines. Nature597, 318–324 (2021). [DOI] [PubMed] [Google Scholar]
- 13.Alberer, M. et al. Safety and immunogenicity of a mRNA rabies vaccine in healthy adults: an open-label, non-randomised, prospective, first-in-human phase 1 clinical trial. Lancet390, 1511–1520 (2017). [DOI] [PubMed] [Google Scholar]
- 14.Lu, C. et al. Safety, Immunogenicity, and mechanism of a rotavirus mRNA-LNP vaccine in mice. Viruses16, 211 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yang, L., Tang, L., Zhang, M. & Liu, C. Recent advances in the molecular design and delivery technology of mRNA for vaccination against infectious diseases. Front. Immunol.13, 896958 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Han, G. et al. Advances in mRNA therapeutics for cancer immunotherapy: from modification to delivery. Adv. Drug Deliv. Rev.199, 114973 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Boreikaitė, V. & Passmore, L. A. 3′-end processing of eukaryotic mRNA: machinery, regulation, and impact on gene expression. Annu. Rev. Biochem.92, 199–225 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shen, G., Liu, J., Yang, H., Xie, N. & Yang, Y. mRNA therapies: pioneering a new era in rare genetic disease treatment. J. Control Release369, 696–721 (2024). [DOI] [PubMed] [Google Scholar]
- 19.Chaudhary, N., Weissman, D. & Whitehead, K. A. mRNA vaccines for infectious diseases: principles, delivery and clinical translation. Nat. Rev. Drug Discov.20, 817–838 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kwon, S., Kwon, M., Im, S., Lee, K. & Lee, H. mRNA vaccines: the most recent clinical applications of synthetic mRNA. Arch. Pharm. Res.45, 245–262 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bourke, A. M., Schwarz, A. & Schuman, E. M. De-centralizing the central dogma: mRNA translation in space and time. Mol. Cell83, 452–468 (2023). [DOI] [PubMed] [Google Scholar]
- 22.Bai, C., Wang, C. & Lu, Y. Novel vectors and administrations for mRNA delivery. Small19, 2303713 (2023). [DOI] [PubMed] [Google Scholar]
- 23.Su, L.-J. et al. Delivery of mRNA for cancer therapy: progress and prospects. Nano Today53, 102013 (2023). [Google Scholar]
- 24.Ye, Z. et al. The mRNA vaccine revolution: COVID-19 has launched the future of vaccinology. ACS Nano17, 15231–15253 (2023). [DOI] [PubMed] [Google Scholar]
- 25.Pan, S., Fan, R., Han, B., Tong, A. & Guo, G. The potential of mRNA vaccines in cancer nanomedicine and immunotherapy. Trends Immunol.45, 20–31 (2024). [DOI] [PubMed] [Google Scholar]
- 26.Metkar, M., Pepin, C. S. & Moore, M. J. Tailor made: the art of therapeutic mRNA design. Nat. Rev. Drug Discov.23, 67–83 (2024). [DOI] [PubMed] [Google Scholar]
- 27.Kawamoto, Y., Wu, Y., Takahashi, Y. & Takakura, Y. Development of nucleic acid medicines based on chemical technology. Adv. Drug Deliv. Rev.199, 114872 (2023). [DOI] [PubMed] [Google Scholar]
- 28.Cao, Q., Fang, H. & Tian, H. mRNA vaccines contribute to innate and adaptive immunity to enhance immune response in vivo. Biomaterials310, 122628 (2024). [DOI] [PubMed] [Google Scholar]
- 29.Sun, H., Li, K., Liu, C. & Yi, C. Regulation and functions of non-m6A mRNA modifications. Nat. Rev. Mol. Cell Biol.24, 714–731 (2023). [DOI] [PubMed] [Google Scholar]
- 30.Rodell, R., Robalin, N. & Martinez, N. M. Why U matters: detection and functions of pseudouridine modifications in mRNAs. Trends Biochem. Sci.49, 12–27 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lin, S. & Kuang, M. RNA modification-mediated mRNA translation regulation in liver cancer: mechanisms and clinical perspectives. Nat. Rev. Gastroenterol. Hepatol.21, 267–281 (2024). [DOI] [PubMed] [Google Scholar]
- 32.Flamand, M. N., Tegowski, M. & Meyer, K. D. The proteins of mRNA modification: writers, readers, and erasers. Annu. Rev. Biochem.92, 145–173 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Qu, Y. et al. Advanced nano-based strategies for mRNA tumor vaccine. Acta Pharm. Sin. B14, 170–189 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fan, Y. et al. Encoding and display technologies for combinatorial libraries in drug discovery: the coming of age from biology to therapy. Acta Pharm. Sin. B14, 3362–3384 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Liu, X., Huang, P., Yang, R. & Deng, H. mRNA cancer vaccines: construction and boosting strategies. ACS Nano17, 19550–19580 (2023). [DOI] [PubMed] [Google Scholar]
- 36.Son, S., Park, M., Kim, J. & Lee, K. ACE mRNA (additional chimeric element incorporated IVT mRNA) for enhancing protein expression by modulating immunogenicity. Adv. Sci.11, 2307541 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kim, Y.-K. RNA therapy: rich history, various applications and unlimited future prospects. Exp. Mol. Med.54, 455–465 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wang, C. & Yuan, F. A comprehensive comparison of DNA and RNA vaccines. Adv. Drug Deliv. Rev.210, 115340 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Minnaert, A.-K. et al. Strategies for controlling the innate immune activity of conventional and self-amplifying mRNA therapeutics: getting the message across. Adv. Drug Deliv. Rev.176, 113900 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Liu, C.-X. & Chen, L.-L. Circular RNAs: characterization, cellular roles, and applications. Cell185, 2016–2034 (2022). [DOI] [PubMed] [Google Scholar]
- 41.Silva-Pilipich, N., Beloki, U., Salaberry, L. & Smerdou, C. Self-amplifying RNA: a second revolution of mRNA vaccines against COVID-19. Vaccines12, 318 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Krähling, V. et al. Self-amplifying RNA vaccine protects mice against lethal Ebola virus infection. Mol. Ther.31, 374–386 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.McGee, J. E. et al. Complete substitution with modified nucleotides in self-amplifying RNA suppresses the interferon response and increases potency. Nat. Biotechnol. 10.1038/s41587-024-02306-z (2024). [DOI] [PMC free article] [PubMed]
- 44.Kim, J., Eygeris, Y., Gupta, M. & Sahay, G. Self-assembled mRNA vaccines. Adv. Drug Deliv. Rev.170, 83–112 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Comes, J. D. G., Pijlman, G. P. & Hick, T. A. H. Rise of the RNA machines—self-amplification in mRNA vaccine design. Trends Biotechnol.41, 1417–1429 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Androsavich, J. R. Frameworks for transformational breakthroughs in RNA-based medicines. Nat. Rev. Drug Discov.23, 421–444 (2024). [DOI] [PubMed] [Google Scholar]
- 47.Blakney, A. K., Ip, S. & Geall, A. J. An update on self-amplifying mRNA vaccine development. Vaccines9, 97 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Dastgerdi, N. K. et al. Charge neutralized poly(β-amino ester) polyplex nanoparticles for delivery of self-amplifying RNA. Nanoscale Adv.6, 1409–1422 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hu, C. et al. Amplifying mRNA vaccines: potential versatile magicians for oncotherapy. Front. Immunol.14, 1261243 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Young, T. L., Wang, K. C., Varley, A. J. & Li, B. Clinical delivery of circular RNA: lessons learned from RNA drug development. Adv. Drug Deliv. Rev.197, 114826 (2023). [DOI] [PubMed] [Google Scholar]
- 51.Hwang, H. J. & Kim, Y. K. Molecular mechanisms of circular RNA translation. Exp. Mol. Med.56, 1272–1280 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Unti, M. J. & Jaffrey, S. R. Highly efficient cellular expression of circular mRNA enables prolonged protein expression. Cell Chem. Biol.31, 163–176 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zhang, F., Li, L. & Fan, Z. circRNAs and their relationship with breast cancer: a review. World J. Surg. Oncol.20, 373 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Niu, D., Wu, Y. & Lian, J. Circular RNA vaccine in disease prevention and treatment. Signal Transduct. Target. Ther.8, 341 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Zhang, J. & Chen, J. Circular mRNAs: More stable RNAs lead to more persistent protein expression. Mol. Ther. Nucleic Acids30, 357–358 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Liu, T. et al. Components, formulations, deliveries, and combinations of tumor vaccines. ACS Nano18, 18801–18833 (2024). [DOI] [PubMed] [Google Scholar]
- 57.Shui, M. et al. Engineering polyphenol-based carriers for nucleic acid delivery. Theranostics13, 3204–3223 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Puri, S. et al. Evolution of nanomedicine formulations for targeted delivery and controlled release. Adv. Drug Deliv. Rev.200, 114962 (2023). [DOI] [PubMed] [Google Scholar]
- 59.Sun, B. et al. Engineering nanoparticle toolkits for mRNA delivery. Adv. Drug Deliv. Rev.200, 115042 (2023). [DOI] [PubMed] [Google Scholar]
- 60.Huayamares, S. G., Loughrey, D., Kim, H., Dahlman, J. E. & Sorscher, E. J. Nucleic acid-based drugs for patients with solid tumours. Nat. Rev. Clin. Oncol.21, 407–427 (2024). [DOI] [PubMed] [Google Scholar]
- 61.Saiding, Q. et al. Nano-bio interactions in mRNA nanomedicine: challenges and opportunities for targeted mRNA delivery. Adv. Drug Deliv. Rev.203, 115116 (2023). [DOI] [PubMed] [Google Scholar]
- 62.Malla, R., Srilatha, M., Farran, B. & Nagaraju, G. P. mRNA vaccines and their delivery strategies: a journey from infectious diseases to cancer. Mol. Ther.32, 13–31 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.de los Reyes, E. et al. Single-dose AAV9-CLN6 gene transfer slows the decline in motor and language function in variant late infantile neuronal ceroid lipofuscinosis 6: interim results from phase 1/2 trial. Mol. Genet. Metab.132, S32–S33 (2021). [Google Scholar]
- 64.Gu, J. et al. Building a better silver bullet: current status and perspectives of non‐viral vectors for mRNA vaccines. Adv. Healthc. Mater.13, 2302409 (2024). [DOI] [PubMed] [Google Scholar]
- 65.Masarwy, R., Stotsky-Oterin, L., Elisha, A., Hazan-Halevy, I. & Peer, D. Delivery of nucleic acid based genome editing platforms via lipid nanoparticles: clinical applications. Adv. Drug Deliv. Rev.211, 115359 (2024). [DOI] [PubMed] [Google Scholar]
- 66.Jiang, Z., Xu, Y., Du, G. & Sun, X. Emerging advances in delivery systems for mRNA cancer vaccines. J. Control Release370, 287–301 (2024). [DOI] [PubMed] [Google Scholar]
- 67.Kong, B., Kim, Y., Kim, E. H., Suk, J. S. & Yang, Y. mRNA: a promising platform for cancer immunotherapy. Adv. Drug Deliv. Rev.199, 114993 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zhong, Y., Du, S. & Dong, Y. mRNA delivery in cancer immunotherapy. Acta Pharm. Sin. B13, 1348–1357 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Yang, W., Mixich, L., Boonstra, E. & Cabral, H. Polymer-based mRNA delivery strategies for advanced therapies. Adv. Healthc. Mater.12, 2202688 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Percec, V. & Sahoo, D. From Frank–Kasper, quasicrystals, and biological membrane mimics to reprogramming in vivo the living factory to target the delivery of mRNA with one-component amphiphilic Janus dendrimers. Biomacromolecules25, 1353–1370 (2024). [DOI] [PubMed] [Google Scholar]
- 71.Zhai, J., Fong, C., Tran, N. & Drummond, C. J. Non-lamellar lyotropic liquid crystalline lipid nanoparticles for the next generation of nanomedicine. ACS Nano13, 6178–6206 (2019). [DOI] [PubMed] [Google Scholar]
- 72.Iscaro, J. et al. Lyotropic liquid crystalline phase nanostructure and cholesterol enhance lipid nanoparticle mediated mRNA transfection in macrophages. Adv. Funct. Mater.10.1002/adfm.202405286 (2024). [Google Scholar]
- 73.Borrajo, M. L. et al. Ionizable nanoemulsions for RNA delivery into the central nervous system – importance of diffusivity. J. Control Release372, 295–303 (2024). [DOI] [PubMed] [Google Scholar]
- 74.Huang, T. et al. Lipid nanoparticle-based mRNA vaccines in cancers: current advances and future prospects. Front. Immunol.13, 922301 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Wang, J., Fang, Y., Luo, Z., Wang, J. & Zhao, Y. Emerging mRNA technology for liver disease therapy. ACS Nano18, 17378–17406 (2024). [DOI] [PubMed] [Google Scholar]
- 76.DeJulius, C. R. et al. Engineering approaches for RNA-based and cell-based osteoarthritis therapies. Nat. Rev. Rheumatol.20, 81–100 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Ji, P. et al. Modular hydrogel vaccine for programmable and coordinate elicitation of cancer immunotherapy. Adv. Sci.10, 2301789 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Neshat, S. Y. et al. Polymeric nanoparticle gel for intracellular mRNA delivery and immunological reprogramming of tumors. Biomaterials300, 122185 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Huang, P., Deng, H., Wang, C., Zhou, Y. & Chen, X. Cellular trafficking of nanotechnology‐mediated mRNA delivery. Adv. Mater.36, 2307822 (2024). [DOI] [PubMed] [Google Scholar]
- 80.Jeong, M., Lee, Y., Park, J., Jung, H. & Lee, H. Lipid nanoparticles (LNPs) for in vivo RNA delivery and their breakthrough technology for future applications. Adv. Drug Deliv. Rev.200, 114990 (2023). [DOI] [PubMed] [Google Scholar]
- 81.Neary, M. T. et al. Nebulised delivery of RNA formulations to the lungs: From aerosol to cytosol. J. Control Release366, 812–833 (2024). [DOI] [PubMed] [Google Scholar]
- 82.Wang, Z. et al. Exosomes decorated with a recombinant SARS-CoV-2 receptor-binding domain as an inhalable COVID-19 vaccine. Nat. Biomed. Eng.6, 791–805 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Liu, M., Hu, S., Yan, N., Popowski, K. D. & Cheng, K. Inhalable extracellular vesicle delivery of IL-12 mRNA to treat lung cancer and promote systemic immunity. Nat. Nanotechnol.19, 565–575 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Cotto, H. A. M. et al. Cationic-motif-modified exosomes for mRNA delivery to retinal photoreceptors. J. Mater. Chem. B12, 7384–7400 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Esih, H. et al. Mucoadhesive film for oral delivery of vaccines for protection of the respiratory tract. J. Control Release371, 179–192 (2024). [DOI] [PubMed] [Google Scholar]
- 86.Luo, P.-K. et al. pH-responsive β-glucans-complexed mRNA in LNPs as an oral vaccine for enhancing cancer immunotherapy. Adv. Mater.36, 2404830 (2024). [DOI] [PubMed] [Google Scholar]
- 87.Wang, K. et al. Delivery of mRNA vaccines and anti-PDL1 siRNA through non-invasive transcutaneous route effectively inhibits tumor growth. Compos. Part B Eng.233, 109648 (2022). [Google Scholar]
- 88.Gao, K. et al. In utero delivery of mRNA to the heart, diaphragm and muscle with lipid nanoparticles. Bioact. Mater.25, 387–398 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Suzuki, Y., Katsurada, Y. & Hyodo, K. Differences and similarities of the intravenously administered lipid nanoparticles in three clinical trials: potential linkage between lipid nanoparticles and extracellular vesicles. Mol. Pharm.20, 4883–4892 (2023). [DOI] [PubMed] [Google Scholar]
- 90.Zimmermann, C. M. et al. Spray drying siRNA-lipid nanoparticles for dry powder pulmonary delivery. J. Control Release351, 137–150 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Zai, W. et al. Optimized RNA interference therapeutics combined with interleukin-2 mRNA for treating hepatitis B virus infection. Signal Transduct. Target. Ther.9, 150 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Albertsen, C. H. et al. The role of lipid components in lipid nanoparticles for vaccines and gene therapy. Adv. Drug Deliv. Rev.188, 114416 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Han, X. et al. An ionizable lipid toolbox for RNA delivery. Nat. Commun.12, 7233 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Han, J. et al. Lipid nanoparticle-based mRNA delivery systems for cancer immunotherapy. Nano Converg.10, 36 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Kulkarni, J. A., Witzigmann, D., Leung, J., Tam, Y. Y. C. & Cullis, P. R. On the role of helper lipids in lipid nanoparticle formulations of siRNA. Nanoscale11, 21733–21739 (2019). [DOI] [PubMed] [Google Scholar]
- 96.Cárdenas, M., Campbell, R. A., Arteta, M. Y., Lawrence, M. J. & Sebastiani, F. Review of structural design guiding the development of lipid nanoparticles for nucleic acid delivery. Curr. Opin. Colloid Interface Sci.66, 101705 (2023). [Google Scholar]
- 97.Su, K. et al. Reformulating lipid nanoparticles for organ-targeted mRNA accumulation and translation. Nat. Commun.15, 5659 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Gyanani, V. & Goswami, R. Key design features of lipid nanoparticles and electrostatic charge-based lipid nanoparticle targeting. Pharmaceutics15, 1184 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Han, X. et al. Fast and facile synthesis of amidine-incorporated degradable lipids for versatile mRNA delivery in vivo. Nat. Chem. 10.1038/s41557-024-01557-2 (2024). [DOI] [PMC free article] [PubMed]
- 100.Carrasco, M. J. et al. Ionization and structural properties of mRNA lipid nanoparticles influence expression in intramuscular and intravascular administration. Commun. Biol.4, 956 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Uchida, S., Lau, C. Y. J., Oba, M. & Miyata, K. Polyplex designs for improving the stability and safety of RNA therapeutics. Adv. Drug Deliv. Rev.199, 114972 (2023). [DOI] [PubMed] [Google Scholar]
- 102.Jörgensen, A. M., Wibel, R. & Bernkop-Schnürch, A. Biodegradable cationic and ionizable cationic lipids: a roadmap for safer pharmaceutical excipients. Small19, 2206968 (2023). [DOI] [PubMed] [Google Scholar]
- 103.Ripoll, M. et al. An imidazole modified lipid confers enhanced mRNA-LNP stability and strong immunization properties in mice and non-human primates. Biomaterials286, 121570 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Tang, Z., Yu, F., Hsu, J. C., Shi, J. & Cai, W. Soybean oil-derived lipids for efficient mRNA delivery. Adv. Mater.36, 2302901 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Yuan, Z., Yan, R., Fu, Z., Wu, T. & Ren, C. Impact of physicochemical properties on biological effects of lipid nanoparticles: are they completely safe. Sci. Total Environ.927, 172240 (2024). [DOI] [PubMed] [Google Scholar]
- 106.Papi, M., Pozzi, D., Palmieri, V. & Caracciolo, G. Principles for optimization and validation of mRNA lipid nanoparticle vaccines against COVID-19 using 3D bioprinting. Nano Today43, 101403 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Lu, J. et al. Screening libraries to discover molecular design principles for the targeted delivery of mRNA with one-component ionizable amphiphilic Janus dendrimers derived from plant phenolic acids. Pharmaceutics15, 1572 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Seo, Y. et al. Recent progress of lipid nanoparticles-based lipophilic drug delivery: focus on surface modifications. Pharmaceutics15, 772 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Li, W., Wang, C., Zhang, Y. & Lu, Y. Lipid nanocarrier‐based mRNA therapy: challenges and promise for clinical transformation. Small20, 2310531 (2024). [DOI] [PubMed] [Google Scholar]
- 110.Lin, Z., Zou, Z., Pu, Z., Wu, M. & Zhang, Y. Application of microfluidic technologies on COVID-19 diagnosis and drug discovery. Acta Pharm. Sin. B13, 2877–2896 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Zhang, H. et al. Microfluidics for nano-drug delivery systems: from fundamentals to industrialization. Acta Pharm. Sin. B13, 3277–3299 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Fan, Y. et al. Automated high-throughput preparation and characterization of oligonucleotide-loaded lipid nanoparticles. Int. J. Pharm.599, 120392 (2021). [DOI] [PubMed] [Google Scholar]
- 113.Wang, H. et al. Combined helical-blade-strengthened co-flow focusing and high-throughput screening for the synthesis of highly homogeneous nanoliposomes. Nano Today56, 102301 (2024). [Google Scholar]
- 114.Chehelgerdi, M. & Chehelgerdi, M. The use of RNA-based treatments in the field of cancer immunotherapy. Mol. Cancer22, 106 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Han, X. et al. Adjuvant lipidoid-substituted lipid nanoparticles augment the immunogenicity of SARS-CoV-2 mRNA vaccines. Nat. Nanotechnol.18, 1105–1114 (2023). [DOI] [PubMed] [Google Scholar]
- 116.Gote, V. et al. A comprehensive review of mRNA vaccines. Int. J. Mol. Sci.24, 2700 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Yang, W. et al. Nanoformulations targeting immune cells for cancer therapy: mRNA therapeutics. Bioact. Mater.23, 438–470 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.de Picciotto, S. et al. Selective activation and expansion of regulatory T cells using lipid encapsulated mRNA encoding a long-acting IL-2 mutein. Nat. Commun.13, 3866 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Xu, L. et al. New therapeutic strategies based on IL-2 to modulate Treg cells for autoimmune diseases. Int. Immunopharmacol.72, 322–329 (2019). [DOI] [PubMed] [Google Scholar]
- 120.Mackensen, A. et al. 958 BNT211: a phase I/II trial to evaluate safety and efficacy of CLDN6 CAR-T cells and vaccine-mediated in vivo expansion in patients with CLDN6-positive advanced solid tumors. J. Immunother. Cancer9, A1008 (2021). [Google Scholar]
- 121.Granit, V. et al. Safety and clinical activity of autologous RNA chimeric antigen receptor T-cell therapy in myasthenia gravis (MG-001): a prospective, multicentre, open-label, non-randomised phase 1b/2a study. Lancet Neurol.22, 578–590 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Hussain, A. et al. mRNA vaccines for COVID-19 and diverse diseases. J. Control Release345, 314–333 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Lamb, Y. N. BNT162b2 mRNA COVID-19 vaccine: first approval. Drugs81, 495–501 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Huang, X. et al. The landscape of mRNA nanomedicine. Nat. Med.28, 2273–2287 (2022). [DOI] [PubMed] [Google Scholar]
- 125.World Health Organization. COVID-19 vaccine tracker and landscape. https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (2023).
- 126.Li, Y. et al. A nanoparticle vaccine displaying varicella-zoster virus gE antigen induces a superior cellular immune response than a licensed vaccine in mice and non-human primates. Front. Immunol.15, 1419634 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Yang, S. et al. Identification and characterization of Varicella Zoster Virus circular RNA in lytic infection. Nat. Commun.15, 4932 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Huang, L. et al. Herpes zoster mRNA vaccine induces superior vaccine immunity over licensed vaccine in mice and rhesus macaques. Emerg. Microbes Infect.13, 2309985 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Meissner, H. C. Disarming the respiratory syncytial virus. New Engl. J. Med.383, 487–488 (2020). [DOI] [PubMed] [Google Scholar]
- 130.Nam, H. H. & Ison, M. G. respiratory syncytial virus. Semin. Respir. Crit. Care Med.42, 788–799 (2021). [DOI] [PubMed] [Google Scholar]
- 131.Shaw, C. A. et al. Safety, tolerability, and immunogenicity of an mRNA-based respiratory syncytial virus vaccine in healthy young adults in a phase 1 clinical trial. J. Infect. Dis. 10.1093/infdis/jiae035 (2024). [DOI] [PMC free article] [PubMed]
- 132.Qiu, X. et al. Development of mRNA vaccines against respiratory syncytial virus (RSV). Cytokine Growth Factor Rev.68, 37–53 (2022). [DOI] [PubMed] [Google Scholar]
- 133.Hutchinson, E. C. Influenza virus. Trends Microbiol.26, 809–810 (2018). [DOI] [PubMed] [Google Scholar]
- 134.Lee, I. T. et al. Safety and immunogenicity of a phase 1/2 randomized clinical trial of a quadrivalent, mRNA-based seasonal influenza vaccine (mRNA-1010) in healthy adults: interim analysis. Nat. Commun.14, 3631 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.ModernaTX, Inc. A phase 3, randomized, observer-blind, active-controlled study to evaluate the safety and efficacy of mRNA-1010 candidate seasonal influenza vaccine in adults 50 years and older. https://clinicaltrials.gov/study/NCT05566639 (2024).
- 136.ModernaTX, Inc. A phase 3, randomized, stratified, observer-blind, active-controlled study to evaluate the immunogenicity, reactogenicity and safety of mRNA-1010 seasonal influenza vaccine in adults 18 years and older. https://clinicaltrials.gov/study/NCT05827978 (2024).
- 137.Wu, K. et al. Characterization of humoral and cellular immunologic responses to an mRNA-based human cytomegalovirus vaccine from a phase 1 trial of healthy adults. J. Virol.98, e01603–e01623 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Fierro, C. et al. Safety and immunogenicity of a messenger RNA-based cytomegalovirus vaccine in healthy adults: results from a phase 1, randomized, clinical trial. J. Infect. Dis. 10.1093/infdis/jiae114 (2024). [DOI] [PMC free article] [PubMed]
- 139.Hu, X. et al. Human cytomegalovirus mRNA-1647 vaccine candidate elicits potent and broad neutralization and higher antibody-dependent cellular cytotoxicity responses than the gb/mf59 vaccine. J. Infect. Dis. 10.1093/infdis/jiad593 (2024). [DOI] [PMC free article] [PubMed]
- 140.ModernaTX, Inc. A phase 3, randomized, observer-blind, placebo-controlled study to evaluate the efficacy, safety, and immunogenicity of mRNA-1647 cytomegalovirus (CMV) vaccine in healthy participants 16 to 40 years of age. https://clinicaltrials.gov/study/NCT05085366 (2023).
- 141.Sivars, L. et al. Cell-free human papillomavirus DNA is a sensitive biomarker for prognosis and for early detection of relapse in locally advanced cervical cancer. Clin. Cancer Res.30, 2764–2771 (2024). [DOI] [PubMed] [Google Scholar]
- 142.Malagón, T., Franco, E. L., Tejada, R. & Vaccarella, S. Epidemiology of HPV-associated cancers past, present and future: towards prevention and elimination. Nat. Rev. Clin. Oncol.21, 522–538 (2024). [DOI] [PubMed] [Google Scholar]
- 143.Lee, S. et al. mRNA-HPV vaccine encoding E6 and E7 improves therapeutic potential for HPV-mediated cancers via subcutaneous immunization. J. Med. Virol.95, e29309 (2023). [DOI] [PubMed] [Google Scholar]
- 144.Xiang, Y. et al. LMP2-mRNA lipid nanoparticle sensitizes EBV-related tumors to anti-PD-1 therapy by reversing T cell exhaustion. J. Nanobiotechnol.21, 324 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Zhao, G.-X. et al. mRNA‐based vaccines targeting the T-cell epitope-rich domain of Epstein Barr virus latent proteins elicit robust anti-tumor immunity in mice. Adv. Sci.10, 2302116 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Armbruster, N., Jasny, E. & Petsch, B. Advances in RNA vaccines for preventive indications: a case study of a vaccine against rabies. Vaccines7, 132 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Aleem, M. T., Munir, F., Shakoor, A. & Gao, F. mRNA vaccines against infectious diseases and future direction. Int. Immunopharmacol.135, 112320 (2024). [DOI] [PubMed] [Google Scholar]
- 148.Li, M. et al. An mRNA vaccine against rabies provides strong and durable protection in mice. Front. Immunol.14, 1288879 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Long, J. et al. A rabies mRNA vaccine provides a rapid and long-term immune response in mice. Nano Today53, 102038 (2023). [Google Scholar]
- 150.Hellgren, F. et al. Unmodified rabies mRNA vaccine elicits high cross-neutralizing antibody titers and diverse B cell memory responses. Nat. Commun.14, 3713 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Wan, J. et al. A single immunization with core–shell structured lipopolyplex mRNA vaccine against rabies induces potent humoral immunity in mice and dogs. Emerg. Microbes Infect.12, 2270081 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Cao, H. et al. A rabies mRNA vaccine with H270P mutation in its glycoprotein induces strong cellular and humoral immunity. Vaccine42, 1116–1121 (2024). [DOI] [PubMed] [Google Scholar]
- 153.Gómez, C. E. et al. Enhancement of the HIV-1-specific immune response induced by an mRNA vaccine through boosting with a poxvirus MVA vector expressing the same antigen. Vaccines9, 959 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Mu, Z., Haynes, B. F. & Cain, D. W. HIV mRNA vaccines—progress and future paths. Vaccines9, 134 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Usero, L. et al. The combination of an mRNA immunogen, a TLR7 agonist and a PD1 blocking agent enhances in-vitro HIV T-cell immune responses. Vaccines11, 286 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Leggat, D. J. et al. Vaccination induces HIV broadly neutralizing antibody precursors in humans. Science378, eadd6502 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Tarrés-Freixas, F., Clotet, B., Carrillo, J. & Blanco, J. Nucleic acid vaccines encoding proteins and virus-like particles for HIV prevention. Vaccines12, 298 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Zhang, P. et al. A multiclade env–gag VLP mRNA vaccine elicits tier-2 HIV-1-neutralizing antibodies and reduces the risk of heterologous SHIV infection in macaques. Nat. Med.27, 2234–2245 (2021). [DOI] [PubMed] [Google Scholar]
- 159.Mandal, S. et al. A long-term stable cold-chain-friendly HIV mRNA vaccine encoding multi-epitope viral protease cleavage site immunogens inducing immunogen-specific protective T cell immunity. Emerg. Microbes Infect.13, 2377606 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Esteban, I. et al. In the Era of mRNA vaccines, is there any hope for HIV functional cure? Viruses13, 501 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Morris, L. mRNA vaccines offer hope for HIV. Nat. Med.27, 2082–2084 (2021). [DOI] [PubMed] [Google Scholar]
- 162.Wu, F., Qin, M., Wang, H. & Sun, X. Nanovaccines to combat virus-related diseases. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol.15, e1857 (2023). [DOI] [PubMed] [Google Scholar]
- 163.Crunkhorn, S. Monkeypox mRNA vaccine protects mice and macaques. Nat. Rev. Drug Discov.23, 251 (2024). [DOI] [PubMed] [Google Scholar]
- 164.Shah, M. et al. Deciphering the immunogenicity of monkeypox proteins for designing the potential mRNA vaccine. ACS Omega8, 43341–43355 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Sang, Y. et al. Monkeypox virus quadrivalent mRNA vaccine induces immune response and protects against vaccinia virus. Signal Transduct. Target. Ther.8, 172 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Hou, F. et al. mRNA vaccines encoding fusion proteins of monkeypox virus antigens protect mice from vaccinia virus challenge. Nat. Commun.14, 5925 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Zuiani, A. et al. A multivalent mRNA monkeypox virus vaccine (BNT166) protects mice and macaques from orthopoxvirus disease. Cell187, 1363–1373 (2024). [DOI] [PubMed] [Google Scholar]
- 168.Tian, Y. et al. An MPXV mRNA-LNP vaccine candidate elicits protective immune responses against monkeypox virus. Chin. Chem. Lett.35, 109270 (2024). [Google Scholar]
- 169.Su, C. et al. A quadrivalent mRNA immunization elicits potent immune responses against multiple orthopoxviral antigens and neutralization of monkeypox virus in rodent models. Vaccines12, 385 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Tadic, S. & Martínez, A. Nucleic acid cancer vaccines targeting tumor related angiogenesis. Could mRNA vaccines constitute a game changer? Front. Immunol.15, 1433185 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Conry, R. M. et al. Characterization of a messenger RNA polynucleotide vaccine vector. Cancer Res55, 1397–1400 (1995). [PubMed] [Google Scholar]
- 172.Sobhani, N. et al. Therapeutic cancer vaccines: from biological mechanisms and engineering to ongoing clinical trials. Cancer Treat. Rev.109, 102429 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Yuan, Y., Gao, F., Chang, Y., Zhao, Q. & He, X. Advances of mRNA vaccine in tumor: a maze of opportunities and challenges. Biomark. Res.11, 6 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Loquai, C. et al. 549 An RNA-lipoplex (RNA-LPX) vaccine demonstrates strong immunogenicity and promising clinical activity in a phase I trial in cutaneous melanoma patients with no evidence of disease at trial inclusion. J. Immunother. Cancer9, A579 (2021). [Google Scholar]
- 175.Sahin, U. et al. An RNA vaccine drives immunity in checkpoint-inhibitor-treated melanoma. Nature585, 107–112 (2020). [DOI] [PubMed] [Google Scholar]
- 176.Yao, R., Xie, C. & Xia, X. Recent progress in mRNA cancer vaccines. Hum. Vaccin Immunother.20, 2307187 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Nierengarten, M. B. Messenger RNA vaccine advances provide treatment possibilities for cancer. Cancer128, 213–214 (2022). [DOI] [PubMed] [Google Scholar]
- 178.Sayour, E. J., Boczkowski, D., Mitchell, D. A. & Nair, S. K. Cancer mRNA vaccines: clinical advances and future opportunities. Nat. Rev. Clin. Oncol.21, 489–500 (2024). [DOI] [PubMed] [Google Scholar]
- 179.Trivedi, V. et al. mRNA-based precision targeting of neoantigens and tumor-associated antigens in malignant brain tumors. Genome Med.16, 17 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Ben-Akiva, E. et al. Biodegradable lipophilic polymeric mRNA nanoparticles for ligand-free targeting of splenic dendritic cells for cancer vaccination. Proc. Natl. Acad. Sci. USA120, e2301606120 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Liu, Y., Yan, Q., Zeng, Z., Fan, C. & Xiong, W. Advances and prospects of mRNA vaccines in cancer immunotherapy. Biochim. Biophys. Acta Rev. Cancer1879, 189068 (2024). [DOI] [PubMed] [Google Scholar]
- 182.Tian, Y., Hu, D., Li, Y. & Yang, L. Development of therapeutic vaccines for the treatment of diseases. Mol. Biomed.3, 40 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Lin, F. et al. Advancing personalized medicine in brain cancer: exploring the role of mRNA vaccines. J. Transl. Med.21, 830 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Search for: mRNA, Phase: 3, 4 | List Results | ClinicalTrials.gov. https://clinicaltrials.gov/search?intr=mRNA&aggFilters=phase:3%204 (2024).
- 185.Bergstrom, C., Fischer, N. O., Kubicek-Sutherland, J. Z. & Stromberg, Z. R. mRNA vaccine platforms to prevent bacterial infections. Trends Mol. Med.30, 524–526 (2024). [DOI] [PubMed] [Google Scholar]
- 186.Maruggi, G. et al. Immunogenicity and protective efficacy induced by self-amplifying mRNA vaccines encoding bacterial antigens. Vaccine35, 361–368 (2017). [DOI] [PubMed] [Google Scholar]
- 187.Pine, M. et al. Development of an mRNA-lipid nanoparticle vaccine against Lyme disease. Mol. Ther.31, 2702–2714 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.ModernaTX, Inc. A phase 1/2, randomized, observer-blind, placebo-controlled, dose-ranging study to evaluate the safety and immunogenicity of heptavalent mRNA-1975 (SR1-7) and Monovalent mRNA-1982 (SR1) in parallel against Lyme disease in healthy participants 18 through 70 years of age. https://clinicaltrials.gov/study/NCT05975099 (2023).
- 189.Matarazzo, L. & Bettencourt, P. J. G. mRNA vaccines: a new opportunity for malaria, tuberculosis and HIV. Front. Immunol.14, 1172691 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Zhou, F. & Zhang, D. Recent advance in the development of tuberculosis vaccines in clinical trials and virus-like particle-based vaccine candidates. Front. Immunol.14, 1238649 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Fan, X.-Y. & Lowrie, D. B. Where are the RNA vaccines for TB? Emerg. Microbes Infect.10, 1217–1218 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Larsen, S. E. et al. An RNA-based vaccine platform for use against mycobacterium tuberculosis. Vaccines11, 130 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Xu, S., Yang, K., Li, R. & Zhang, L. mRNA vaccine era—mechanisms, drug platform and clinical prospection. Int. J. Mol. Sci.21, 6582 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Meulewaeter, S. et al. Alpha-galactosylceramide improves the potency of mRNA LNP vaccines against cancer and intracellular bacteria. J. Control Release370, 379–391 (2024). [DOI] [PubMed] [Google Scholar]
- 195.Singh, B. et al. Host-directed therapies for parasitic diseases. Future Med. Chem.11, 1999–2018 (2019). [DOI] [PubMed] [Google Scholar]
- 196.AlGabbani, Q. Nanotechnology: a promising strategy for the control of parasitic infections. Exp. Parasitol.250, 108548 (2023). [DOI] [PubMed] [Google Scholar]
- 197.You, H. et al. The mRNA vaccine technology era and the future control of parasitic infections. Clin. Microbiol. Rev.36, e00241–21 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Chuang, Y.-M. et al. A mosquito AgTRIO mRNA vaccine contributes to immunity against malaria. NPJ Vaccines8, 88 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Hassert, M. & Harty, J. T. Alt-RNAtive vaccines elicit anti-malarial TRM cells. Nat. Immunol.24, 1397–1398 (2023). [DOI] [PubMed] [Google Scholar]
- 200.Waghela, I. N. et al. Exploring in vitro expression and immune potency in mice using mRNA encoding the Plasmodium falciparum malaria antigen. CelTOS. Front. Immunol.13, 1026052 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Tsoumani, M. E., Voyiatzaki, C. & Efstathiou, A. Malaria vaccines: from the past towards the mRNA vaccine era. Vaccines11, 1452 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Kunkeaw, N. et al. A Pvs25 mRNA vaccine induces complete and durable transmission-blocking immunity to Plasmodium vivax. NPJ Vaccines8, 187 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Makoni, M. mRNA vaccine against malaria effective in preclinical model. Lancet Microbe4, e970 (2023). [DOI] [PubMed] [Google Scholar]
- 204.Ganley, M. et al. mRNA vaccine against malaria tailored for liver-resident memory T cells. Nat. Immunol.24, 1487–1498 (2023). [DOI] [PubMed] [Google Scholar]
- 205.Scaria, P. V. et al. mRNA vaccines expressing malaria transmission-blocking antigens Pfs25 and Pfs230D1 induce a functional immune response. NPJ Vaccines9, 9 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Versteeg, L., Almutairi, M. M., Hotez, P. J. & Pollet, J. Enlisting the mRNA vaccine platform to combat parasitic infections. Vaccines7, 122 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Niazi, S. K. Anti-idiotypic mRNA vaccine to treat autoimmune disorders. Vaccines12, 9 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Cao, W. & Xia, T. mRNA lipid nanoparticles induce immune tolerance to treat human diseases. Med. Rev.3, 180–183 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Pisetsky, D. S. Pathogenesis of autoimmune disease. Nat. Rev. Nephrol.19, 509–524 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Krienke, C. et al. A noninflammatory mRNA vaccine for treatment of experimental autoimmune encephalomyelitis. Science371, 145–153 (2021). [DOI] [PubMed] [Google Scholar]
- 211.Herold, K. C. et al. The immunology of type 1 diabetes. Nat. Rev. Immunol.24, 435–451 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Parayath, N. N. et al. Genetic in situ engineering of myeloid regulatory cells controls inflammation in autoimmunity. J. Control Release339, 553–561 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Kim, A., Xie, F., Abed, O. A. & Moon, J. J. Vaccines for immune tolerance against autoimmune disease. Adv. Drug Deliv. Rev.203, 115140 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Lodde, V., Floris, M., Zoroddu, E., Zarbo, I. R. & Idda, M. L. RNA-binding proteins in autoimmunity: from genetics to molecular biology. Wiley Interdiscip. Rev. RNA14, e1772 (2023). [DOI] [PubMed] [Google Scholar]
- 215.Flemming, A. mRNA vaccine shows promise in autoimmunity. Nat. Rev. Immunol.21, 72 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Wardell, C. M. & Levings, M. K. mRNA vaccines take on immune tolerance. Nat. Biotechnol.39, 419–421 (2021). [DOI] [PubMed] [Google Scholar]
- 217.Furlan, R. A tolerizing mRNA vaccine against autoimmunity? Mol. Ther.29, 896–897 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Villanueva, M. T. Suppressing autoimmunity with mRNA vaccines. Nat. Rev. Drug Discov.20, 99 (2021). [DOI] [PubMed] [Google Scholar]
- 219.Krishnamurthy, B. & Thomas, H. E. METTL3 restrains autoimmunity in β-cells. Nat. Cell Biol.26, 321–322 (2024). [DOI] [PubMed] [Google Scholar]
- 220.Foster, T. P., Haller, M. J., Schatz, D., Wasserfall, C. & Sayour, E. 743-P: delayed diabetes with proinsulin mRNA vaccines in NOD mice. Diabetes72, 743–74 (2023). [Google Scholar]
- 221.Deyhimfar, R. et al. The clinical impact of mRNA therapeutics in the treatment of cancers, infections, genetic disorders, and autoimmune diseases. Heliyon10, e26971 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Shi, J. et al. Delivery of mRNA for regulating functions of immune cells. J. Control Release345, 494–511 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Sharma, P., Joshi, R. V., Pritchard, R., Xu, K. & Eicher, M. A. Therapeutic antibodies in medicine. Molecules28, 6438 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Deal, C. E., Carfi, A. & Plante, O. J. Advancements in mRNA encoded antibodies for passive immunotherapy. Vaccines9, 108 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.An, Z. Magic Bullets” at the center stage of immune therapy: a special issue on therapeutic antibodies. Protein Cell9, 1–2 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Hafeez, U., Gan, H. K. & Scott, A. M. Monoclonal antibodies as immunomodulatory therapy against cancer and autoimmune diseases. Curr. Opin. Pharmacol.41, 114–121 (2018). [DOI] [PubMed] [Google Scholar]
- 227.Van Hoecke, L. & Roose, K. How mRNA therapeutics are entering the monoclonal antibody field. J. Transl. Med.17, 54 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Deal, C. E. et al. An mRNA-based platform for the delivery of pathogen-specific IgA into mucosal secretions. Cell Rep. Med.4, 101253 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Le, N. D. et al. Antiangiogenic therapeutic mRNA delivery using lung-selective polymeric nanomedicine for lung cancer treatment. ACS Nano18, 8392–8410 (2024). [DOI] [PubMed] [Google Scholar]
- 230.Chi, H. et al. Rapid development of double-hit mRNA antibody cocktail against orthopoxviruses. Signal Transduct. Target. Ther.9, 69 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Sun, Q. et al. Immune checkpoint therapy for solid tumours: clinical dilemmas and future trends. Signal Transduct. Target. Ther.8, 320 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Chen, W., Huang, Y., Pan, W., Xu, M. & Chen, L. Strategies for developing PD-1 inhibitors and future directions. Biochem. Pharmacol.202, 115113 (2022). [DOI] [PubMed] [Google Scholar]
- 233.Zhao, Y., Gan, L., Ke, D., Chen, Q. & Fu, Y. Mechanisms and research advances in mRNA antibody drug-mediated passive immunotherapy. J. Transl. Med.21, 693 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Hamilton, A. G. et al. Ionizable lipid nanoparticles with integrated immune checkpoint inhibition for mRNA CAR T cell engineering. Adv. Healthc. Mater.12, 2301515 (2023). [DOI] [PubMed] [Google Scholar]
- 235.Van Hoecke, L. et al. mRNA in cancer immunotherapy: beyond a source of antigen. Mol. Cancer20, 48 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Pruitt, S. K. et al. Enhancement of anti‐tumor immunity through local modulation of CTLA‐4 and GITR by dendritic cells. Eur. J. Immunol.41, 3553–3563 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Zeng, J. et al. Antitumor activity of Z15-0-2, a bispecific nanobody targeting PD-1 and CTLA-4. Oncogene43, 2244–2252 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Chung, E. et al. Abstract 1368: novel mRNA encoding anti-PD-L1 monoclonal antibodies for cancer immunotherapy. Cancer Res.84, 1368 (2024). [Google Scholar]
- 239.ModernaTX, Inc. Moderna builds on clinical validation of systemic delivery with two additional development candidates in new autoimmune therapeutic area. https://investors.modernatx.com/news/news-details/2020/Moderna-Builds-on-Clinical-Validation-of-Systemic-Delivery-with-Two-Additional-Development-Candidates-in-New-Autoimmune-Therapeutic-Area/default.aspx (2020).
- 240.Salles, G. et al. Rituximab in B-cell hematologic malignancies: a review of 20 years of clinical experience. Adv. Ther.34, 2232–2273 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Thran, M. et al. mRNA mediates passive vaccination against infectious agents, toxins, and tumors. EMBO Mol. Med.9, 1434–1447 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Wu, L. et al. Intravenous delivery of RNA encoding anti-PD-1 human monoclonal antibody for treating intestinal cancer. J. Cancer13, 579–588 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Miao, L., Zhang, Y. & Huang, L. mRNA vaccine for cancer immunotherapy. Mol. Cancer20, 41 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Ramesh, P., Shivde, R., Jaishankar, D., Saleiro, D. & Le Poole, I. C. A palette of cytokines to measure anti-tumor efficacy of T cell-based therapeutics. Cancers13, 821 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Bentebibel, S.-E. & Diab, A. Cytokines in the treatment of melanoma. Curr. Oncol. Rep.23, 83 (2021). [DOI] [PubMed] [Google Scholar]
- 246.Cho, J., Ritz, J. & Zylberberg, C. Comparing the functionality of proleukin® and Akron interleukin-2 through an analysis of key T cell subsets. Cytotherapy22, S121–S122 (2020). [Google Scholar]
- 247.Wang, H. et al. Viral vectors expressing interleukin 2 for cancer immunotherapy. Hum. Gene Ther.34, 878–895 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Ko, B., Takebe, N., Andrews, O., Makena, M. R. & Chen, A. P. Rethinking oncologic treatment strategies with interleukin-2. Cells12, 1316 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Shin, H., Kang, S., Won, C. & Min, D.-H. Enhanced local delivery of engineered IL-2 mRNA by porous silica nanoparticles to promote effective antitumor immunity. ACS Nano17, 17554–17567 (2023). [DOI] [PubMed] [Google Scholar]
- 250.Jiang, Y. et al. Tumor-activated IL-2 mRNA delivered by lipid nanoparticles for cancer immunotherapy. J. Control Release368, 663–675 (2024). [DOI] [PubMed] [Google Scholar]
- 251.Beck, J. D. et al. Long-lasting mRNA-encoded interleukin-2 restores CD8+ T cell neoantigen immunity in MHC class I-deficient cancers. Cancer Cell42, 568–582 (2024). [DOI] [PubMed] [Google Scholar]
- 252.Van Acker, H. H. et al. The role of the common gamma-chain family cytokines in γδ T cell-based anti-cancer immunotherapy. Cytokine Growth Factor Rev.41, 54–64 (2018). [DOI] [PubMed] [Google Scholar]
- 253.Shen, Z. et al. Lipid nanoparticle-mediated delivery of IL-21-encoding mRNA induces viral clearance in mouse models of hepatitis B virus persistence. J. Med. Virol.95, e29062 (2023). [DOI] [PubMed] [Google Scholar]
- 254.Lei, S. et al. Efficient colorectal cancer gene therapy with IL-15 mRNA nanoformulation. Mol. Pharm.17, 3378–3391 (2020). [DOI] [PubMed] [Google Scholar]
- 255.Yu, J. et al. Targeted LNPs deliver mRNA encoding IL-15 superagonists to balance efficacy and toxicity in cancer therapy. Preprint at biorxiv10.1101/2024.01.11.575299 (2024).
- 256.Wang, X. et al. A combination of PD-L1-targeted IL-15 mRNA nanotherapy and ultrasound-targeted microbubble destruction for tumor immunotherapy. J. Control Release367, 45–60 (2024). [DOI] [PubMed] [Google Scholar]
- 257.Gavigan, J.-A. et al. Abstract 1711: induction of anti-tumor immunity by intratumoral delivery of mRNA encoding cytokines and TNFRSF agonists. Cancer Res.81, 1711 (2021). [Google Scholar]
- 258.Liu, H. et al. Oxaliplatin lipidated prodrug synergistically enhances the anti-colorectal cancer effect of IL12 mRNA. Drug Deliv. Transl. Res. 10.1007/s13346-024-01540-x (2024). [DOI] [PubMed]
- 259.Hewitt, S. L. et al. Intratumoral IL12 mRNA therapy promotes Th1 transformation of the tumor microenvironment. Clin. Cancer Res.26, 6284–6298 (2020). [DOI] [PubMed] [Google Scholar]
- 260.Di Trani, C. A. et al. Intratumoral injection of IL-12-encoding mRNA targeted to CSF1R and PD-L1 exerts potent anti-tumor effects without substantial systemic exposure. Mol. Ther. Nucleic Acids33, 599–616 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Liu, J.-Q. et al. Intratumoral delivery of IL-12 and IL-27 mRNA using lipid nanoparticles for cancer immunotherapy. J. Control Release345, 306–313 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Kumar, A., Taghi Khani, A., Sanchez Ortiz, A. & Swaminathan, S. GM-CSF: a double-edged sword in cancer immunotherapy. Front. Immunol.13, 901277 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Bechter, O. et al. 391 A first-in-human study of intratumoral SAR441000, an mRNA mixture encoding IL-12sc, interferon alpha2b, GM-CSF and IL-15sushi as monotherapy and in combination with cemiplimab in advanced solid tumors. J. Immunother. Cancer8, A237–A238 (2020). [Google Scholar]
- 264.Yeapuri, P. et al. Development of an extended half-life GM-CSF fusion protein for Parkinson’s disease. J. Control Release348, 951–965 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Eggenhuizen, P. J., Ng, B. H. & Ooi, J. D. Treg enhancing therapies to treat autoimmune diseases. Int. J. Mol. Sci.21, 7015 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266.Beheshti, S. A., Shamsasenjan, K., Ahmadi, M. & Abbasi, B. CAR Treg: a new approach in the treatment of autoimmune diseases. Int. Immunopharmacol.102, 108409 (2022). [DOI] [PubMed] [Google Scholar]
- 267.Mikami, N., Kawakami, R. & Sakaguchi, S. New Treg cell-based therapies of autoimmune diseases: towards antigen-specific immune suppression. Curr. Opin. Immunol.67, 36–41 (2020). [DOI] [PubMed] [Google Scholar]
- 268.Goswami, T. K. et al. Regulatory T cells (Tregs) and their therapeutic potential against autoimmune disorders—advances and challenges. Hum. Vaccin Immunother.18, 2035117 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Dong, C., Fu, T., Ji, J., Li, Z. & Gu, Z. The role of interleukin-4 in rheumatic diseases. Clin. Exp. Pharmacol. Physiol.45, 747–754 (2018). [DOI] [PubMed] [Google Scholar]
- 270.Creusot, R. J. et al. A short pulse of IL-4 delivered by DCs electroporated with modified mRNA can both prevent and treat autoimmune diabetes in NOD mice. Mol. Ther.18, 2112–2120 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Ozgur, B. A. et al. 255-LB: the role of Th1, Th2, Th17, and Treg cells in various clinical phases of type 1 diabetes. Diabetes68, 255–2 (2019). [Google Scholar]
- 272.Vaseghi, H. & Jadali, Z. Th1/Th2 cytokines in type 1 diabetes: relation to duration of disease and gender. Indian J. Endocrinol. Metab.20, 312–316 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 273.Aviña, A. E. et al. IL-10 modified mRNA monotherapy prolongs survival after composite facial allografting through the induction of mixed chimerism. Mol. Ther. Nucleic Acids31, 610–627 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.Chen, Z. et al. A polyphenol-assisted IL-10 mRNA delivery system for ulcerative colitis. Acta Pharm. Sin. B12, 3367–3382 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275.Bu, T. et al. Exosome-mediated delivery of inflammation-responsive IL-10 mRNA for controlled atherosclerosis treatment. Theranostics11, 9988–10000 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276.Zhang, G., Tang, T., Chen, Y., Huang, X. & Liang, T. mRNA vaccines in disease prevention and treatment. Signal Transduct. Target. Ther.8, 365 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 277.Arance Fernandez, A. M. et al. A phase I study (E011-MEL) of a TriMix-based mRNA immunotherapy (ECI-006) in resected melanoma patients: Analysis of safety and immunogenicity. J. Clin. Oncol.37, 2641 (2019). [Google Scholar]
- 278.Wu, T., Huang, C., Yao, Y., Du, Z. & Liu, Z. Suicide gene delivery system mediated by ultrasound-targeted microbubble destruction: a promising strategy for cancer therapy. Hum. Gene Ther.33, 1246–1259 (2022). [DOI] [PubMed] [Google Scholar]
- 279.Nguyen, Q. M., Dupré, P.-F., Haute, T., Montier, T. & d’Arbonneau, F. Suicide gene strategies applied in ovarian cancer studies. Cancer Gene Ther.30, 812–821 (2023). [DOI] [PubMed] [Google Scholar]
- 280.Song, N. et al. Suicide gene delivery by morphology-adaptable enantiomeric peptide assemblies for combined ovarian cancer therapy. Acta Biomater.175, 250–261 (2024). [DOI] [PubMed] [Google Scholar]
- 281.Falcon, C. et al. Combinatorial suicide gene strategies for the safety of cell therapies. Front. Immunol.13, 975233 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Saeb, S. et al. Suicide gene therapy in cancer and HIV-1 infection: an alternative to conventional treatments. Biochem. Pharmacol.197, 114893 (2022). [DOI] [PubMed] [Google Scholar]
- 283.Mizrak, A. et al. Genetically engineered microvesicles carrying suicide mRNA/protein inhibit schwannoma tumor growth. Mol. Ther.21, 101–108 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284.Mohammadi, S., Akbari‐Birgani, S., Nikfarjam, N. & Rasekhian, M. Polyethyleneimine-decorated graphene oxide quantum dot as a carrier for suicide gene delivery to the breast cancer three-dimensional model. J. Drug Deliv. Sci. Technol.88, 104849 (2023). [Google Scholar]
- 285.Nakashima, I. et al. Non-viral inducible caspase 9 mRNA delivery using lipid nanoparticles against breast cancer: an in vitro study. Biochem. Biophys. Res. Commun.635, 144–153 (2022). [DOI] [PubMed] [Google Scholar]
- 286.Saito, S. et al. Tumor-tropic liposome-mediated therapeutic delivery of mRNA for T cell malignancies. Blood136, 21–22 (2020). [Google Scholar]
- 287.Chen, L., Liu, S. & Tao, Y. Regulating tumor suppressor genes: post-translational modifications. Signal Transduct. Target. Ther.5, 90 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 288.Peuget, S., Zhou, X. & Selivanova, G. Translating p53-based therapies for cancer into the clinic. Nat. Rev. Cancer24, 192–215 (2024). [DOI] [PubMed] [Google Scholar]
- 289.Zhang, C. et al. Chemotherapy drugs derived nanoparticles encapsulating mRNA encoding tumor suppressor proteins to treat triple-negative breast cancer. Nano Res.12, 855–861 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 290.Zhou, H. et al. ROS-responsive nanoparticle delivery of mRNA and photosensitizer for combinatorial cancer therapy. Nano Lett.23, 3661–3668 (2023). [DOI] [PubMed] [Google Scholar]
- 291.Cao, C. et al. Enhancement of protein translation by CRISPR/dCasRx coupled with SINEB2 repeat of noncoding RNAs. Nucleic Acids Res.51, e33 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Liu, Y. et al. Non-invasive PTEN mRNA brain delivery effectively mitigates growth of orthotopic glioblastoma. Nano Today49, 101790 (2023). [Google Scholar]
- 293.Ma, P. et al. Local and systemic delivery of the bims gene nano-complex for efficient oral squamous cell carcinoma therapy. Int. J. Nanomed.17, 2925–2941 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 294.Gao, Y. et al. Functionalized DMP-039 hybrid nanoparticle as a novel mRNA vector for efficient cancer suicide gene therapy. Int. J. Nanomed.16, 5211–5232 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Okumura, K. et al. Bax mRNA therapy using cationic liposomes for human malignant melanoma. J. Gene Med.10, 910–917 (2008). [DOI] [PubMed] [Google Scholar]
- 296.Gu, T. et al. Intratumoural delivery of TRAIL mRNA induces colon cancer cell apoptosis. Biomed. Pharmacother.174, 116603 (2024). [DOI] [PubMed] [Google Scholar]
- 297.da Silva, W. N. et al. Ionizable lipid nanoparticle-mediated TRAIL mRNA delivery in the tumor microenvironment to inhibit colon cancer progression. Int. J. Nanomed.19, 2655–2673 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 298.van der Eb, M. M. et al. Gene therapy with Apoptin induces regression of xenografted human hepatomas. Cancer Gene Ther.9, 53–61 (2002). [DOI] [PubMed] [Google Scholar]
- 299.Tang, Y. et al. Self-assembled small messenger RNA nanospheres for efficient therapeutic apoptin expression and synergistic gene-chemotherapy of breast cancer. J. Colloid Interface Sci.603, 191–198 (2021). [DOI] [PubMed] [Google Scholar]
- 300.Castro, J., Ribo, M., Benito, A. & Vilanova, M. Apoptin, A versatile protein with selective antitumor activity. Curr. Med. Chem.25, 3540–3559 (2018). [DOI] [PubMed] [Google Scholar]
- 301.Bae, Y., Lee, J., Kho, C., Choi, J. S. & Han, J. Apoptin gene delivery by a PAMAM dendrimer modified with a nuclear localization signal peptide as a gene carrier for brain cancer therapy. Korean J. Physiol. Pharmacol.25, 467–478 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 302.Qu, Y.-M. et al. Huntingtin-associated protein 1 is a potential tumor suppressor for gastric cancer. Mol. Biol. Rep.50, 1517–1531 (2023). [DOI] [PubMed] [Google Scholar]
- 303.Zheng, Z. & Song, Y. Synaptopodin-2: a potential tumor suppressor. Cancer Cell Int.23, 158 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 304.Polito, L., Djemil, A. & Bortolotti, M. Plant toxin-based immunotoxins for cancer therapy: a short overview. Biomedicines4, 12 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305.Kreitman, R. J. & Pastan, I. Immunotoxins: from design to clinical application. Biomolecules11, 1696 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 306.Akbari, B. et al. Immunotoxins in cancer therapy: review and update. Int. Rev. Immunol.36, 207–219 (2017). [DOI] [PubMed] [Google Scholar]
- 307.Dashtiahangar, M. et al. Anti-cancer immunotoxins, challenges, and approaches. Curr. Pharm. Des.27, 932–941 (2021). [DOI] [PubMed] [Google Scholar]
- 308.Guo, R. et al. Fusion of an albumin-binding domain extends the half-life of immunotoxins. Int. J. Pharm.511, 538–549 (2016). [DOI] [PubMed] [Google Scholar]
- 309.Stöcker, M. et al. Secretion of functional anti-CD30-angiogenin immunotoxins into the supernatant of transfected 293T-cells. Protein Expr. Purif.28, 211–219 (2003). [DOI] [PubMed] [Google Scholar]
- 310.Granot-Matok, Y. et al. Lipid nanoparticles-loaded with toxin mRNA represents a new strategy for the treatment of solid tumors. Theranostics13, 3497–3508 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311.Eggers, R., Philippi, A., Altmeyer, M. O., Breinig, F. & Schmitt, M. J. Primary T cells for mRNA-mediated immunotoxin delivery. Gene Ther.25, 47–53 (2018). [DOI] [PubMed] [Google Scholar]
- 312.Hirschberger, K. et al. Exploring cytotoxic mRNAs as a novel class of anti-cancer biotherapeutics. Mol. Ther. Methods Clin. Dev.8, 141–151 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Spiess, K., Jakobsen, M. H., Kledal, T. N. & Rosenkilde, M. M. The future of antiviral immunotoxins. J. Leukoc. Biol.99, 911–925 (2016). [DOI] [PubMed] [Google Scholar]
- 314.Li, Y. et al. The potential and promise for clinical application of adoptive T cell therapy in cancer. J. Transl. Med.22, 413 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315.Chehrazi-Raffle, A., Budde, L. E. & Pal, S. K. Boosting CAR T cells with anti-tumor mRNA vaccines. Nat. Med.29, 2711–2712 (2023). [DOI] [PubMed] [Google Scholar]
- 316.Mackensen, A. et al. CLDN6-specific CAR-T cells plus amplifying RNA vaccine in relapsed or refractory solid tumors: the phase 1 BNT211-01 trial. Nat. Med.29, 2844–2853 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317.Tombácz, I. et al. Highly efficient CD4+ T cell targeting and genetic recombination using engineered CD4+ cell-homing mRNA-LNPs. Mol. Ther.29, 3293–3304 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 318.Zhou, J. et al. Lipid nanoparticles produce chimeric antigen receptor T cells with interleukin-6 knockdown in vivo. J. Control Release350, 298–307 (2022). [DOI] [PubMed] [Google Scholar]
- 319.Rurik, J. G. et al. CAR T cells produced in vivo to treat cardiac injury. Science375, 91–96 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 320.Liuzzo, G. & Patrono, C. In vivo generated chimeric antigen receptor T cells reduce fibrosis and restore cardiac function in experimental heart failure. Eur. Heart J.43, 1531–1532 (2022). [DOI] [PubMed] [Google Scholar]
- 321.Blache, U., Tretbar, S., Koehl, U., Mougiakakos, D. & Fricke, S. CAR T cells for treating autoimmune diseases. RMD Open9, e002907 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 322.Cartesian Therapeutics. Descartes-08 for patients with systemic lupus erythematosus. https://clinicaltrials.gov/study/NCT06038474 (2024).
- 323.Thatte, A. S. et al. mRNA lipid nanoparticles for ex vivo engineering of immunosuppressive T cells for autoimmunity therapies. Nano Lett.23, 10179–10188 (2023). [DOI] [PubMed] [Google Scholar]
- 324.Yang, F. et al. Messenger RNA electroporated hepatitis B virus (HBV) antigen-specific T cell receptor (TCR) redirected T cell therapy is well-tolerated in patients with recurrent HBV-related hepatocellular carcinoma post-liver transplantation: results from a phase I trial. Hepatol. Int.17, 850–859 (2023). [DOI] [PubMed] [Google Scholar]
- 325.Maggadottir, S. M. et al. Transient TCR-based T cell therapy in a patient with advanced treatment-resistant MSI-high colorectal cancer. Mol. Ther.32, 2021–2029 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 326.Ling, K. et al. Genome editing mRNA nanotherapies inhibit cervical cancer progression and regulate the immunosuppressive microenvironment for adoptive T-cell therapy. J. Control Release360, 496–513 (2023). [DOI] [PubMed] [Google Scholar]
- 327.Chen, Y. et al. CAR-macrophage: a new immunotherapy candidate against solid tumors. Biomed. Pharmacother.139, 111605 (2021). [DOI] [PubMed] [Google Scholar]
- 328.Reiss, K. et al. 951 A phase 1 first in human study of adenovirally transduced anti-HER2 CAR macrophages in subjects with HER2 overexpressing solid tumors: preliminary safety, pharmacokinetics, and TME reprogramming data. J. Immunother. Cancer9, A1000 (2021). [Google Scholar]
- 329.Kasimsetty, S., Gatla, H. & Chinnasamy, D. 108 MCY-M11, a CAR-PBMC cell product transiently expressing a mesothelin targeted mRNA CAR, exhibits desirable functional and immune phenotype attributed to sustained antitumor immunity in vitro. J. Immunother. Cancer8, A67–A68 (2020). [Google Scholar]
- 330.Tang, C. et al. mRNA-laden lipid-nanoparticle-enabled in situ CAR-macrophage engineering for the eradication of multidrug-resistant bacteria in a sepsis mouse model. ACS Nano18, 2261–2278 (2024). [DOI] [PubMed] [Google Scholar]
- 331.Zhang, Y. et al. Close the cancer–immunity cycle by integrating lipid nanoparticle–mRNA formulations and dendritic cell therapy. Nat. Nanotechnol.18, 1364–1374 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 332.Dueland, S. Phase I/II trial of vaccine therapy with mRNA- transfected dendritic cells in patients with advanced malignant melanoma. https://clinicaltrials.gov/study/NCT01278940 (2016).
- 333.Kyte, J. A. et al. Phase I/II trial of melanoma therapy with dendritic cells transfected with autologous tumor-mRNA. Cancer Gene Ther.13, 905–918 (2006). [DOI] [PubMed] [Google Scholar]
- 334.Anguille, S. et al. Dendritic cell vaccination as postremission treatment to prevent or delay relapse in acute myeloid leukemia. Blood130, 1713–1721 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 335.Qin, S. et al. mRNA-based therapeutics: powerful and versatile tools to combat diseases. Signal Transduct. Target. Ther.7, 166 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 336.Sahin, U., Karikó, K. & Türeci, Ö. mRNA-based therapeutics—developing a new class of drugs. Nat. Rev. Drug Discov.13, 759–780 (2014). [DOI] [PubMed] [Google Scholar]
- 337.Vlatkovic, I. Non-immunotherapy application of LNP-mRNA: maximizing efficacy and safety. Biomedicines9, 530 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 338.Rohner, E., Yang, R., Foo, K. S., Goedel, A. & Chien, K. R. Unlocking the promise of mRNA therapeutics. Nat. Biotechnol.40, 1586–1600 (2022). [DOI] [PubMed] [Google Scholar]
- 339.Lonez, C. et al. Critical residues involved in Toll-like receptor 4 activation by cationic lipid nanocarriers are not located at the lipopolysaccharide-binding interface. Cell. Mol. Life Sci.72, 3971–3982 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 340.Romerio, A. & Peri, F. Increasing the chemical variety of small-molecule-based TLR4 modulators: an overview. Front. Immunol.11, 1210 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 341.Kedmi, R., Ben-Arie, N. & Peer, D. The systemic toxicity of positively charged lipid nanoparticles and the role of Toll-like receptor 4 in immune activation. Biomaterials31, 6867–6875 (2010). [DOI] [PubMed] [Google Scholar]
- 342.Lonez, C. et al. Cationic lipid nanocarriers activate Toll-like receptor 2 and NLRP3 inflammasome pathways. Nanomed. Nanotechnol. Biol. Med.10, 775–782 (2014). [DOI] [PubMed] [Google Scholar]
- 343.Qin, Y., Ou, L., Zha, L., Zeng, Y. & Li, L. Delivery of nucleic acids using nanomaterials. Mol. Biomed.4, 48 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 344.Ohto, T. et al. Inhibition of the inflammatory pathway enhances both the in vitro and in vivo transfection activity of exogenous in vitro-transcribed mRNAs delivered by lipid nanoparticles. Biol. Pharm. Bull.42, 299–302 (2019). [DOI] [PubMed] [Google Scholar]
- 345.Vavilis, T. et al. mRNA in the context of protein replacement therapy. Pharmaceutics15, 166 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 346.Schwarze, S. R., Ho, A., Vocero-Akbani, A. & Dowdy, S. F. In vivo protein transduction: delivery of a biologically active protein into the mouse. Science285, 1569–1572 (1999). [DOI] [PubMed] [Google Scholar]
- 347.Chen, C.-Y. et al. Treatment of hemophilia A using factor VIII messenger RNA lipid nanoparticles. Mol. Ther. Nucleic Acids20, 534–544 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 348.Commissioner, O. of the. FDA approves first gene therapy for adults with severe hemophilia A. FDAhttps://www.fda.gov/news-events/press-announcements/fda-approves-first-gene-therapy-adults-severe-hemophilia (2023).
- 349.Prieve, M. G. et al. Targeted mRNA therapy for ornithine transcarbamylase deficiency. Mol. Ther.26, 801–813 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 350.Arcturus Therapeutics, Inc. Phase 2, randomized, double-blind, placebo-controlled, nested single and multiple ascending dose study to evaluate the safety, tolerability and pharmacokinetics of ARCT-810 in adolescent and adult participants with ornithine transcarbamylase deficiency. https://clinicaltrials.gov/study/NCT05526066 (2023).
- 351.DeRosa, F. et al. Improved efficacy in a fabry disease model using a systemic mRNA liver depot system as compared to enzyme replacement therapy. Mol. Ther.27, 878–889 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 352.Rodríguez-Castejón, J., Beraza-Millor, M., Solinís, M. Á., Rodríguez-Gascón, A. & del Pozo-Rodríguez, A. Targeting strategies with lipid vectors for nucleic acid supplementation therapy in Fabry disease: a systematic review. Drug Deliv. Transl. Res.14, 2615–2628 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 353.Zhu, X. et al. Systemic mRNA therapy for the treatment of fabry disease: preclinical studies in wild-type mice, fabry mouse model, and wild-type non-human primates. Am. J. Hum. Genet.104, 625–637 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 354.Koeberl, D. et al. Interim analyses of a first-in-human phase 1/2 mRNA trial for propionic acidaemia. Nature628, 872–877 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 355.Behyar, M. B., Mirzaie, A., Hasanzadeh, M. & Shadjou, N. Advancements in biosensing of hormones: recent progress and future trends. Trends Anal. Chem.173, 117600 (2024). [Google Scholar]
- 356.Kielb, J. et al. Transformation or replacement—effects of hormone therapy on cardiovascular risk. Pharmacol. Ther.254, 108592 (2024). [DOI] [PubMed] [Google Scholar]
- 357.Jirikowski, G. F., Sanna, P. P., Maciejewski-Lenoir, D. & Bloom, F. E. Reversal of diabetes insipidus in Brattleboro rats: intrahypothalamic injection of vasopressin mRNA. Science255, 996–998 (1992). [DOI] [PubMed] [Google Scholar]
- 358.Huang, R., Shi, J., Wei, R. & Li, J. Challenges of insulin-like growth factor-1 testing. Crit. Rev. Clin. Lab. Sci.61, 388–403 (2024). [DOI] [PubMed] [Google Scholar]
- 359.Wu, H. et al. Engineered adipose-derived stem cells with IGF-1-modified mRNA ameliorates osteoarthritis development. Stem Cell Res. Ther.13, 19 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 360.Zangi, L. et al. Modified mRNA directs the fate of heart progenitor cells and induces vascular regeneration after myocardial infarction. Nat. Biotechnol.31, 898–907 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 361.Collén, A. et al. VEGFA mRNA for regenerative treatment of heart failure. Nat. Rev. Drug Discov.21, 79–80 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 362.Anttila, V. et al. Synthetic mRNA encoding VEGF-A in patients undergoing coronary artery bypass grafting: design of a phase 2a clinical trial. Mol. Ther. Methods Clin. Dev.18, 464–472 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 363.Yu, Z. et al. Cell-mediated delivery of VEGF modified mRNA enhances blood vessel regeneration and ameliorates murine critical limb ischemia. J. Control Release310, 103–114 (2019). [DOI] [PubMed] [Google Scholar]
- 364.Nawaz, M. et al. lipid nanoparticles deliver the therapeutic VEGFA mRNA in vitro and in vivo and transform extracellular vesicles for their functional extensions. Adv. Sci.10, 2206187 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 365.Kapoor, N., Galang, G., Marbán, E. & Cho, H. C. Transcriptional suppression of Connexin43 by TBX18 undermines cell-cell electrical coupling in postnatal cardiomyocytes. J. Biol. Chem.286, 14073–14079 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 366.Wolfson, D. W. et al. Transient pacing in pigs with complete heart block via myocardial injection of mRNA coding for the T-box transcription factor 18. Nat. Biomed. Eng. 10.1038/s41551-024-01211-9 (2024). [DOI] [PMC free article] [PubMed]
- 367.Yang, T. et al. Therapeutic HNF4A mRNA attenuates liver fibrosis in a preclinical model. J. Hepatol.75, 1420–1433 (2021). [DOI] [PubMed] [Google Scholar]
- 368.Shteinberg, M., Haq, I. J., Polineni, D. & Davies, J. C. Cystic fibrosis. Lancet397, 2195–2211 (2021). [DOI] [PubMed] [Google Scholar]
- 369.Rowe, S. M. et al. Inhaled mRNA therapy for treatment of cystic fibrosis: interim results of a randomized, double‐blind, placebo‐controlled phase 1/2 clinical study. J. Cyst. Fibros.22, 656–664 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 370.Wang, P. et al. Co-delivery of NS1 and BMP2 mRNAs to murine pluripotent stem cells leads to enhanced BMP-2 expression and osteogenic differentiation. Acta Biomater.108, 337–346 (2020). [DOI] [PubMed] [Google Scholar]
- 371.Wang, P. et al. In Vivo bone tissue induction by freeze-dried collagen-nanohydroxyapatite matrix loaded with BMP2/NS1 mRNAs lipopolyplexes. J. Control Release334, 188–200 (2021). [DOI] [PubMed] [Google Scholar]
- 372.Geng, Y. et al. BMP-2 and VEGF-A modRNAs in collagen scaffold synergistically drive bone repair through osteogenic and angiogenic pathways. Commun. Biol.4, 82 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 373.Lin, C.-Y. et al. Treatment of intervertebral disk disease by the administration of mRNA encoding a cartilage-anabolic transcription factor. Mol. Ther. Nucleic Acids16, 162–171 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 374.Sanger, H. L., Klotz, G., Riesner, D., Gross, H. J. & Kleinschmidt, A. K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA73, 3852–3856 (1976). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 375.Lin, C.-Y. et al. Messenger RNA-based therapeutics for brain diseases: an animal study for augmenting clearance of beta-amyloid by intracerebral administration of neprilysin mRNA loaded in polyplex nanomicelles. J. Control Release235, 268–275 (2016). [DOI] [PubMed] [Google Scholar]
- 376.Li, H. et al. Engineering brain-derived neurotrophic factor mRNA delivery for the treatment of Alzheimer’s disease. Chem. Eng. J.466, 143152 (2023). [Google Scholar]
- 377.Lescan, M. et al. De novo synthesis of elastin by exogenous delivery of synthetic modified mRNA into skin and elastin-deficient cells. Mol. Ther. Nucleic Acids11, 475–484 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 378.Golombek, S. et al. Improved tropoelastin synthesis in the skin by codon optimization and nucleotide modification of tropoelastin-encoding synthetic mRNA. Mol. Ther. Nucleic Acids33, 642–654 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 379.Ghazanfari, S., Khademhosseini, A. & Smit, T. H. Mechanisms of lamellar collagen formation in connective tissues. Biomaterials97, 74–84 (2016). [DOI] [PubMed] [Google Scholar]
- 380.You, Y. et al. Intradermally delivered mRNA-encapsulating extracellular vesicles for collagen-replacement therapy. Nat. Biomed. Eng.7, 887–900 (2023). [DOI] [PubMed] [Google Scholar]
- 381.Sun, N., Abil, Z. & Zhao, H. Recent advances in targeted genome engineering in mammalian systems. Biotechnol. J.7, 1074–1087 (2012). [DOI] [PubMed] [Google Scholar]
- 382.Yang, W. et al. Progress of delivery methods for CRISPR-Cas9. Expert Opin. Drug Deliv.19, 913–926 (2022). [DOI] [PubMed] [Google Scholar]
- 383.Bjurström, C. F. et al. Reactivating fetal hemoglobin expression in human adult erythroblasts through BCL11A knockdown using targeted endonucleases. Mol. Ther. Nucleic Acids5, e351 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 384.Shen, B. et al. Efficient genome modification by CRISPR-Cas9 nickase with minimal off-target effects. Nat. Methods11, 399–402 (2014). [DOI] [PubMed] [Google Scholar]
- 385.Cho, S. W., Kim, S., Kim, J. M. & Kim, J.-S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol.31, 230–232 (2013). [DOI] [PubMed] [Google Scholar]
- 386.Cheng, H., Zhang, F. & Ding, Y. CRISPR/Cas9 delivery system engineering for genome editing in therapeutic applications. Pharmaceutics13, 1649 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 387.Ling, S. et al. Lentiviral delivery of co-packaged Cas9 mRNA and a Vegfa-targeting guide RNA prevents wet age-related macular degeneration in mice. Nat. Biomed. Eng.5, 144–156 (2021). [DOI] [PubMed] [Google Scholar]
- 388.Wang, Y., Peng, Y., Zi, G., Chen, J. & Peng, B. Co-delivery of Cas9 mRNA and guide RNAs for editing of LGMN gene represses breast cancer cell metastasis. Sci. Rep.14, 8095 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 389.Yi, J. et al. Co-delivery of Cas9 mRNA and guide RNAs edits hepatitis B virus episomal and integration DNA in mouse and tree shrew models. Antivir. Res.215, 105618 (2023). [DOI] [PubMed] [Google Scholar]
- 390.Liu, J. et al. Fast and efficient CRISPR/Cas9 genome editing in vivo enabled by bioreducible lipid and messenger RNA nanoparticles. Adv. Mater.31, 1902575 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 391.Yadav, M., Atala, A. & Lu, B. Developing all-in-one virus-like particles for Cas9 mRNA/single guide RNA co-delivery and aptamer-containing lentiviral vectors for improved gene expression. Int. J. Biol. Macromol.209, 1260–1270 (2022). [DOI] [PubMed] [Google Scholar]
- 392.Gillmore, J. D. et al. CRISPR-Cas9 in vivo gene editing for transthyretin amyloidosis. New Engl. J. Med.385, 493–502 (2021). [DOI] [PubMed] [Google Scholar]
- 393.Zhang, H.-X., Zhang, Y. & Yin, H. Genome editing with mRNA encoding ZFN, TALEN, and Cas9. Mol. Ther.27, 735–746 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 394.Rizzi, M. et al. SARS-CoV-2 infection risk is higher in vaccinated patients with inflammatory autoimmune diseases or liver transplantation treated with mycophenolate due to an impaired antiviral immune response: results of the extended follow up of the RIVALSA prospective cohort. Front. Immunol.14, 1185278 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 395.Li, B. et al. Enhancing the immunogenicity of lipid-nanoparticle mRNA vaccines by adjuvanting the ionizable lipid and the mRNA. Nat. Biomed. Eng. 10.1038/s41551-023-01082-6 (2023). [DOI] [PubMed]
- 396.Kon, E., Ad-El, N., Hazan-Halevy, I., Stotsky-Oterin, L. & Peer, D. Targeting cancer with mRNA–lipid nanoparticles: key considerations and future prospects. Nat. Rev. Clin. Oncol.20, 739–754 (2023). [DOI] [PubMed] [Google Scholar]
- 397.Nugent, C. et al. Second monovalent SARS-CoV-2 mRNA booster restores Omicron-specific neutralizing activity in both nursing home residents and health care workers. Vaccine41, 3403–3409 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 398.Heftdal, L. D. et al. Humoral and cellular immune responses eleven months after the third dose of BNT162b2 an mRNA-based COVID-19 vaccine in people with HIV—a prospective observational cohort study. EBioMedicine93, 104661 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 399.Zhang, Q. et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science370, eabd4570 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 400.Bastard, P. et al. Vaccine breakthrough hypoxemic COVID-19 pneumonia in patients with auto-Abs neutralizing type I IFNs. Sci. Immunol.8, eabp8966 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 401.Hou, Y. et al. Insights into vaccines for elderly individuals: from the impacts of immunosenescence to delivery strategies. NPJ Vaccines9, 77 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 402.Goronzy, J. J. & Weyand, C. M. Understanding immunosenescence to improve responses to vaccines. Nat. Immunol.14, 428–436 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 403.Aiello, A. et al. How can we improve vaccination response in old people? Part I: targeting immunosenescence of innate immunity cells. Int. J. Mol. Sci.23, 9880 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 404.Garnica, M. et al. How can we improve the vaccination response in older people? Part II: targeting immunosenescence of adaptive immunity cells. Int. J. Mol. Sci.23, 9797 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 405.Pereira, B., Xu, X.-N. & Akbar, A. N. Targeting inflammation and immunosenescence to improve vaccine responses in the elderly. Front. Immunol.11, 583019 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 406.Puzianowska-Kuźnicka, M. et al. Interleukin-6 and C-reactive protein, successful aging, and mortality: the PolSenior study. Immun. Ageing13, 21 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 407.Nakaya, H. I. et al. Systems analysis of immunity to influenza vaccination across multiple years and in diverse populations reveals shared molecular signatures. Immunity43, 1186–1198 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 408.Tizazu, A. M., Mengist, H. M. & Demeke, G. Aging, inflammaging and immunosenescence as risk factors of severe COVID-19. Immun. Ageing19, 53 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 409.Salminen, A. Clinical perspectives on the age-related increase of immunosuppressive activity. J. Mol. Med.100, 697–712 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 410.Cunha, L. L., Perazzio, S. F., Azzi, J., Cravedi, P. & Riella, L. V. Remodeling of the immune response with aging: immunosenescence and its potential impact on COVID-19 immune response. Front. Immunol.11, 1748 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 411.Müller, L. & Di Benedetto, S. From aging to long COVID: exploring the convergence of immunosenescence, inflammaging, and autoimmunity. Front. Immunol.14, 1298004 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 412.Cisneros, B. et al. Immune system modulation in aging: molecular mechanisms and therapeutic targets. Front. Immunol.13, 1059173 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 413.Cadar, A. N., Martin, D. E. & Bartley, J. M. Targeting the hallmarks of aging to improve influenza vaccine responses in older adults. Immun. Ageing20, 23 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 414.Valiathan, R., Ashman, M. & Asthana, D. Effects of ageing on the immune system: infants to elderly. Scand. J. Immunol.83, 255–266 (2016). [DOI] [PubMed] [Google Scholar]
- 415.Anderson, E. J. et al. Safety and immunogenicity of SARS-CoV-2 mRNA-1273 vaccine in older adults. New Engl. J. Med.383, 2427–2438 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 416.Walsh, E. E. et al. Safety and immunogenicity of two RNA-based Covid-19 vaccine candidates. New Engl. J. Med.383, 2439–2450 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 417.Li, F. et al. Sex differences orchestrated by androgens at single-cell resolution. Nature629, 193–200 (2024). [DOI] [PubMed] [Google Scholar]
- 418.Haitao, T. et al. COVID-19 and sex differences: mechanisms and biomarkers. Mayo Clin. Proc.95, 2189–2203 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 419.Scully, E. P. Sex differences in HIV infection. Curr. HIV/AIDS Rep.15, 136–146 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 420.Cardenas-Garcia, S. et al. Impact of sex on humoral immunity with live influenza B virus vaccines in mice. NPJ Vaccines9, 45 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 421.Forsyth, K. S., Jiwrajka, N., Lovell, C. D., Toothacre, N. E. & Anguera, M. C. The conneXion between sex and immune responses. Nat. Rev. Immunol.24, 487–502 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 422.Kanuri, B., Biswas, P., Dahdah, A., Murphy, A. J. & Nagareddy, P. R. Impact of age and sex on myelopoiesis and inflammation during myocardial infarction. J. Mol. Cell. Cardiol.187, 80–89 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 423.Costa, A. R. et al. The sex bias of cancer. Trends Endocrinol. Metab.31, 785–799 (2020). [DOI] [PubMed] [Google Scholar]
- 424.Bhattacharya, S., Sadhukhan, D. & Saraswathy, R. Role of sex in immune response and epigenetic mechanisms. Epigenet. Chromatin17, 1 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 425.Li, X. et al. Sex-biased molecular differences in lung adenocarcinoma are ethnic and smoking specific. BMC Pulm. Med.23, 99 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 426.Thompson, M. G., Peiffer, D. S., Larson, M., Navarro, F. & Watkins, S. K. FOXO3, estrogen receptor alpha, and androgen receptor impact tumor growth rate and infiltration of dendritic cell subsets differentially between male and female mice. Cancer Immunol. Immunother.66, 615–625 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 427.Xiao, T. et al. Hallmarks of sex bias in immuno-oncology: mechanisms and therapeutic implications. Nat. Rev. Cancer24, 338–355 (2024). [DOI] [PubMed] [Google Scholar]
- 428.Wallis, C. J. D. et al. Association of patient sex with efficacy of immune checkpoint inhibitors and overall survival in advanced cancers: a systematic review and meta-analysis. JAMA Oncol.5, 529–536 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 429.Pinto, J. A., Araujo, J. M. & Gómez, H. L. Sex, immunity, and cancer. Biochim. Biophys. Acta Rev. Cancer1877, 188647 (2022). [DOI] [PubMed] [Google Scholar]
- 430.Dodd, K. C. & Menon, M. Sex bias in lymphocytes: implications for autoimmune diseases. Front. Immunol.13, 945762 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 431.Xing, E., Billi, A. C. & Gudjonsson, J. E. Sex bias and autoimmune diseases. J. Investig. Dermatol.142, 857–866 (2022). [DOI] [PubMed] [Google Scholar]
- 432.Wilkinson, N. M., Chen, H.-C., Lechner, M. G. & Su, M. A. Sex differences in immunity. Annu. Rev. Immunol.40, 75–94 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 433.Moulton, V. R. Sex hormones in acquired immunity and autoimmune disease. Front. Immunol.9, 2279 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 434.Wu, Y. et al. Sex hormones influence the intestinal microbiota composition in mice. Front. Microbiol.13, 964847 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 435.Natl. Inst. Health. NOT-OD-15-102: consideration of sex as a biological variable in NIH-funded research. https://grants.nih.gov/grants/guide/notice-files/not-od-15-102.html (2015).
- 436.Arumugam, G. S., Damodharan, K., Doble, M. & Thennarasu, S. Significant perspectives on various viral infections targeted antiviral drugs and vaccines including COVID-19 pandemicity. Mol. Biomed.3, 21 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 437.Levin, E. G. et al. Waning immune humoral response to BNT162b2 Covid-19 vaccine over 6 months. N. Engl. J. Med.385, e84 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 438.Furer, V. et al. Immunogenicity induced by two and three doses of the BNT162b2 mRNA vaccine in patients with autoimmune inflammatory rheumatic diseases and immunocompetent controls: a longitudinal multicentre study. Ann. Rheum. Dis.81, 1594–1602 (2022). [DOI] [PubMed] [Google Scholar]
- 439.Leston, M. et al. Disparities in COVID-19 mortality amongst the immunosuppressed: a systematic review and meta-analysis for enhanced disease surveillance. J. Infect.88, 106110 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 440.Meeraus, W. et al. AZD1222 effectiveness against severe COVID-19 in individuals with comorbidity or frailty: the RAVEN cohort study. J. Infect.88, 106129 (2024). [DOI] [PubMed] [Google Scholar]
- 441.Obeid, M. et al. Humoral responses against variants of concern by COVID-19 mRNA vaccines in immunocompromised patients. JAMA Oncol.8, e220446 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 442.Mbonde, A. A. et al. Neuroinvasive West Nile virus infection in immunosuppressed and immunocompetent adults. JAMA Netw. Open7, e244294 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 443.Liao, C. et al. Siglec-F+ neutrophils in the spleen induce immunosuppression following acute infection. Theranostics14, 2589–2604 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 444.Suk-Ouichai, C. et al. A clinical overview of people living with HIV and genitourinary cancer care. Nat. Rev. Urol.21, 373–383 (2024). [DOI] [PubMed] [Google Scholar]
- 445.Gianesin, K. et al. Premature aging and immune senescence in HIV-infected children. AIDS30, 1363–1373 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 446.Vergori, A. et al. Immunogenicity to COVID-19 mRNA vaccine third dose in people living with HIV. Nat. Commun.13, 4922 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 447.Kachikis, A. et al. Timing of maternal COVID-19 vaccine and antibody concentrations in infants born preterm. JAMA Netw. Open7, e2352387 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 448.Johnston, M. S., Galan, A., Watsky, K. L. & Little, A. J. Delayed localized hypersensitivity reactions to the Moderna COVID-19 vaccine. JAMA Dermatol157, 716–720 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 449.Cappelletti-Montano, B., Demuru, G., Laconi, E. & Musio, M. A comparative analysis on serious adverse events reported for COVID-19 vaccines in adolescents and young adults. Front. Public Health11, 1145645 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 450.Heil, M. Self-DNA driven inflammation in COVID-19 and after mRNA-based vaccination: lessons for non-COVID-19 pathologies. Front. Immunol.14, 1259879 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 451.Pettini, E. et al. Spike-specific memory B cell response in hematopoietic cell transplantation recipients following multiple mRNA-1273 vaccinations: a longitudinal observational study. Vaccines12, 368 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 452.Mawson, A. R. & Croft, A. M. Multiple vaccinations and the enigma of vaccine injury. Vaccines8, 676 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 453.BioNTech SE. A Phase Ib/IIa two-part, randomized, placebo-controlled, observer-blind, dose-finding evaluation trial to describe the safety, reactogenicity, and immunogenicity of two investigational vaccines against tuberculosis in BCG vaccinated, HIV-negative subjects and people living with HIV. https://clinicaltrials.gov/study/NCT05547464 (2024).
- 454.Park, J. K., Lee, E. B. & Winthrop, K. L. What rheumatologists need to know about mRNA vaccines: current status and future of mRNA vaccines in autoimmune inflammatory rheumatic diseases. Ann. Rheum. Dis.83, 687–695 (2024). [DOI] [PubMed] [Google Scholar]
- 455.Mendonça, M. C. P., Kont, A., Kowalski, P. S. & O’Driscoll, C. M. Design of lipid-based nanoparticles for delivery of therapeutic nucleic acids. Drug Discov. Today28, 103505 (2023). [DOI] [PubMed] [Google Scholar]
- 456.Bitounis, D., Jacquinet, E., Rogers, M. A. & Amiji, M. M. Strategies to reduce the risks of mRNA drug and vaccine toxicity. Nat. Rev. Drug Discov.23, 281–300 (2024). [DOI] [PubMed] [Google Scholar]
- 457.Tursi, N. J. & Weiner, D. B. Modified messenger-RNA components alter the encoded protein. Nature625, 37–38 (2024). [DOI] [PubMed] [Google Scholar]
- 458.Mulroney, T. E. et al. N1-methylpseudouridylation of mRNA causes +1 ribosomal frameshifting. Nature625, 189–194 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 459.Ni, L. Advances in mRNA-based cancer vaccines. Vaccines11, 1599 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 460.Xie, C., Yao, R. & Xia, X. The advances of adjuvants in mRNA vaccines. NPJ Vaccines8, 162 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 461.Buschmann, M. D. et al. Nanomaterial delivery systems for mRNA vaccines. Vaccines9, 65 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 462.Melamed, J. R. et al. Lipid nanoparticle chemistry determines how nucleoside base modifications alter mRNA delivery. J. Control Release341, 206–214 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 463.Swetha, K. et al. Recent advances in the lipid nanoparticle-mediated delivery of mRNA vaccines. Vaccines11, 658 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 464.Zhuang, L., Ye, Z., Li, L., Yang, L. & Gong, W. Next-generation TB vaccines: progress, challenges, and prospects. Vaccines11, 1304 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 465.Pulendran, B., Arunachalam, P. S. & O’Hagan, D. T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov.20, 454–475 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 466.Gu, Y. et al. Incorporation of a Toll-like receptor 2/6 agonist potentiates mRNA vaccines against cancer and infectious diseases. Signal Transduct. Target. Ther.8, 273 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 467.Coffman, R. L., Sher, A. & Seder, R. A. Vaccine adjuvants: putting innate immunity to work. Immunity33, 492–503 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 468.Zhu, W. et al. cGAMP-adjuvanted multivalent influenza mRNA vaccines induce broadly protective immunity through cutaneous vaccination in mice. Mol. Ther. Nucleic Acids30, 421–437 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 469.Du, J., Chen, X., Ye, Y. & Sun, H. A comparative study on the mechanisms of innate immune responses in mice induced by Alum and Actinidia eriantha polysaccharide. Int. J. Biol. Macromol.156, 1202–1216 (2020). [DOI] [PubMed] [Google Scholar]
- 470.Kimura, S. & Harashima, H. Nano–bio interactions: exploring the biological behavior and the fate of lipid-based gene delivery systems. BioDrugs38, 259–273 (2024). [DOI] [PubMed] [Google Scholar]
- 471.Zhou, H. et al. Stimuli-responsive nanotechnology for RNA delivery. Adv. Sci.10, 2303597 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 472.Sato, Y., Nakamura, T., Yamada, Y. & Harashima, H. The impact of, and expectations for, lipid nanoparticle technology: from cellular targeting to organelle targeting. J. Control Release370, 516–527 (2024). [DOI] [PubMed] [Google Scholar]
- 473.Khare, P., Edgecomb, S. X., Hamadani, C. M., Tanner, E. E. L. & Manickam, D. S. Lipid nanoparticle-mediated drug delivery to the brain. Adv. Drug Deliv. Rev.197, 114861 (2023). [DOI] [PubMed] [Google Scholar]
- 474.Lu, J. et al. Targeted and equally distributed delivery of mRNA to organs with pentaerythritol-based one-component ionizable amphiphilic Janus dendrimers. J. Am. Chem. Soc.145, 18760–18766 (2023). [DOI] [PubMed] [Google Scholar]
- 475.Kim, Y. et al. Design of PD-L1-targeted lipid nanoparticles to turn on PTEN for efficient cancer therapy. Adv. Sci.11, 2309917 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 476.Tang, X. et al. Simultaneous dendritic cells targeting and effective endosomal escape enhance sialic acid-modified mRNA vaccine efficacy and reduce side effects. J. Control Release364, 529–545 (2023). [DOI] [PubMed] [Google Scholar]
- 477.Lin, Y., Chen, X., Wang, K., Liang, L. & Zhang, H. An overview of nanoparticle-based delivery platforms for mRNA vaccines for treating cancer. Vaccines12, 727 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 478.Lorentzen, C. L., Haanen, J. B., Met, Ö. & Svane, I. M. Clinical advances and ongoing trials of mRNA vaccines for cancer treatment. Lancet Oncol.23, e450–e458 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 479.Chen, G. L. et al. 234. Safety and immunogenicity of mRNA-1345, an mRNA-based RSV vaccine in younger and older adult cohorts: results from a phase 1, randomized clinical trial. Open Forum Infect. Dis.9, ofac492.312 (2022). [Google Scholar]
- 480.Bruch, A., Kelani, A. A. & Blango, M. G. RNA-based therapeutics to treat human fungal infections. Trends Microbiol30, 411–420 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 481.Khattak, A. et al. Distant metastasis-free survival results from the randomized, phase 2 mRNA-4157-P201/KEYNOTE-942 trial. J. Clin. Oncol.41, LBA9503 (2023). [Google Scholar]
- 482.Wu, L. et al. Lipid nanoparticle (LNP) delivery carrier-assisted targeted controlled release mRNA vaccines in tumor immunity. Vaccines12, 186 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 483.Kowalski, P. S., Rudra, A., Miao, L. & Anderson, D. G. Delivering the messenger: advances in technologies for therapeutic mRNA delivery. Mol. Ther.27, 710–728 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 484.Dong, Y. & Anderson, D. G. Opportunities and challenges in mRNA therapeutics. Acc. Chem. Res.55, 1 (2022). [DOI] [PubMed] [Google Scholar]
- 485.Liu, C. et al. mRNA-based cancer therapeutics. Nat. Rev. Cancer23, 526–543 (2023). [DOI] [PubMed] [Google Scholar]
- 486.Baden, L. R. et al. Efficacy and safety of the mRNA-1273 SARS-CoV-2 vaccine. New Engl. J. Med.384, 403–416 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 487.Thomas, S. J. et al. Safety and efficacy of the BNT162b2 mRNA Covid-19 vaccine through 6 months. New Engl. J. Med.385, 1761–1773 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 488.Soens, M. et al. 1639. A review of clinical safety, reactogenicity, and immunogenicity of an mRNA-based seasonal influenza vaccine (mRNA-1010) in adults. Open Forum Infect. Dis.10, ofad500.1473 (2023). [Google Scholar]
- 489.Panther, L. et al. 2892. Safety and immunogenicity of mRNA-1647, an mRNA-based cytomegalovirus vaccine in healthy adults: results of a phase 2, randomized, observer-blind, placebo-controlled, dose-finding trial. Open Forum Infect. Dis.10, ofad500.2475 (2023). [Google Scholar]
- 490.Essink, B. et al. The safety and immunogenicity of two Zika virus mRNA vaccine candidates in healthy flavivirus baseline seropositive and seronegative adults: the results of two randomised, placebo-controlled, dose-ranging, phase 1 clinical trials. Lancet Infect. Dis.23, 621–633 (2023). [DOI] [PubMed] [Google Scholar]
- 491.Shaw, C. A. et al. A phase 1, randomized, placebo-controlled, dose-ranging study to evaluate the safety and immunogenicity of an mRNA-based chikungunya virus vaccine in healthy adults. Vaccine41, 3898–3906 (2023). [DOI] [PubMed] [Google Scholar]
- 492.Feldman, R. A. et al. mRNA vaccines against H10N8 and H7N9 influenza viruses of pandemic potential are immunogenic and well tolerated in healthy adults in phase 1 randomized clinical trials. Vaccine37, 3326–3334 (2019). [DOI] [PubMed] [Google Scholar]
- 493.Aldrich, C. et al. Proof-of-concept of a low-dose unmodified mRNA-based rabies vaccine formulated with lipid nanoparticles in human volunteers: a phase 1 trial. Vaccine39, 1310–1318 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 494.Parhiz, H., Atochina-Vasserman, E. N. & Weissman, D. mRNA-based therapeutics: looking beyond COVID-19 vaccines. Lancet403, 1192–1204 (2024). [DOI] [PubMed] [Google Scholar]
- 495.International AIDS Vaccine Initiative. A phase 1, randomized, first-in-human, open-label study to evaluate the safety and immunogenicity of eOD-GT8 60mer mRNA Vaccine (mRNA-1644) and Core-g28v2 60mer mRNA vaccine (mRNA-1644v2-Core) in HIV-1 uninfected adults in good general health. https://clinicaltrials.gov/study/NCT05001373 (2024).
- 496.Peng, X. A phase I study of mRNA vaccine for patients with EBV-positive advanced malignant tumors. https://clinicaltrials.gov/study/NCT05714748 (2023).
- 497.Klinghammer, K. et al. 155P BNT113 + pembrolizumab as first-line treatment in patients with unresectable recurrent/metastatic HNSCC: preliminary safety data from AHEAD-MERIT. Immuno-Oncol. Technol.16, 100267 (2022). [Google Scholar]
- 498.Peng, X. A phase I study of mRNA vaccine for patients with HBV-positive advanced hepatocellular carcinoma. https://clinicaltrials.gov/study/NCT05738447 (2023).
- 499.Linch, M. et al. 421 A first-in-human (FIH) phase I/IIa clinical trial assessing a ribonucleic acid lipoplex (RNA-LPX) encoding shared tumor antigens for immunotherapy of prostate cancer; preliminary analysis of PRO-MERIT. J. Immunother. Cancer9, A451 (2021). [Google Scholar]
- 500.BioNTech SE. LuCa-MERIT-1: first-in-human, open label, phase I dose confirmation trial evaluating the safety, tolerability and preliminary efficacy of BNT116 alone and in combinations in patients with advanced non-small cell lung cancer. https://clinicaltrials.gov/study/NCT05142189 (2024).
- 501.Killock, D. Personalized neoantigen mRNA vaccine mitigates melanoma recurrence. Nat. Rev. Clin. Oncol.21, 168 (2024). [DOI] [PubMed] [Google Scholar]
- 502.Rojas, L. A. et al. Personalized RNA neoantigen vaccines stimulate T cells in pancreatic cancer. Nature618, 144–150 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 503.Shanghai Zhongshan Hospital. Clinical study of mRNA personalized tumor vaccine encoding neonatal antigen combined with Sintilimab injection liver cancer. https://clinicaltrials.gov/study/NCT05761717 (2023).
- 504.Stemirna Therapeutics. Clinical trial on the safety and efficacy of neoantigen antigen mRNA tumor vaccine in the treatment of advanced esophageal cancer and non-small cell lung cancer. https://clinicaltrials.gov/study/NCT03908671 (2023).
- 505.Palmer, C. D. et al. Individualized, heterologous chimpanzee adenovirus and self-amplifying mRNA neoantigen vaccine for advanced metastatic solid tumors: phase 1 trial interim results. Nat. Med.28, 1619–1629 (2022). [DOI] [PubMed] [Google Scholar]
- 506.August, A. et al. A phase 1 trial of lipid-encapsulated mRNA encoding a monoclonal antibody with neutralizing activity against Chikungunya virus. Nat. Med.27, 2224–2233 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 507.Papadopoulos, K. P. et al. A phase I/II dose escalation and expansion trial to evaluate safety and preliminary efficacy of BNT141 in patients with claudin-18.2-positive solid tumors. J. Clin. Oncol.41, TPS2670 (2023). [Google Scholar]
- 508.Yap, T. A. et al. A phase I/II dose escalation trial with expansion cohorts to evaluate safety and preliminary efficacy of BNT142 in patients with prospectively confirmed claudin 6-positive solid tumors. J. Clin. Oncol.41, TPS2669 (2023). [Google Scholar]
- 509.Patel, M. R. et al. A phase I study of mRNA-2752, a lipid nanoparticle encapsulating mRNAs encoding human OX40L, IL-23, and IL-36γ, for intratumoral (iTu) injection alone and in combination with durvalumab. J. Clin. Oncol.38, 3092 (2020). [Google Scholar]
- 510.Berraondo, P., Gomis, G. & Melero, I. The liver as a cytokine factory working on mRNA blueprints for cancer immunotherapy. Cancer Cell42, 502–504 (2024). [DOI] [PubMed] [Google Scholar]
- 511.ModernaTX, Inc. A Phase 1, Dose-escalation study to evaluate the safety, tolerability, pharmacokinetics, and pharmacodynamics of mRNA-6231, encoding for a human serum albumin– interleukin-2 mutein fusion protein (HSA-IL2m), in healthy adults. https://clinicaltrials.gov/study/NCT04916431 (2022).
- 512.Universitair Ziekenhuis Brussel. A phase i study on the safety and immune-modulatory effect of intratumoral (i.t.) administration of mRNA (messenger RNA) encoding dendritic cell activating proteins in patients with early, resectable breast cancer. https://clinicaltrials.gov/study/NCT03788083 (2022).
- 513.Chen, W. et al. 273 phase I study of LioCyx-M, autologous hepatitis B virus (HBV)-specific T cell receptor (TCR) T-cells, in recurrent HBV-related hepatocellular carcinoma (HCC) post-liver transplantation. J. Immunother. Cancer8, A167 (2020). [Google Scholar]
- 514.Argos Therapeutics. A phase II study testing the activity and safety of AGS-004 as an immunotherapeutic in successfully ART-treated subjects infected with HIV-1 in combination with ART followed by ART interruption. https://clinicaltrials.gov/study/NCT00672191 (2013).
- 515.Van Acker, H. H. et al. Dendritic cell-based immunotherapy of acute myeloid leukemia. J. Clin. Med.8, 579 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 516.Svane, I. M. Evaluation of dendritic cells transfected with Survivin, hTERT and p53 mRNA as a treatment for patients with metastatic breast cancer or malignant melanoma. https://clinicaltrials.gov/study/NCT00978913 (2015).
- 517.Sampson, J. H. et al. Reproducibility of outcomes in sequential trials using CMV-targeted dendritic cell vaccination for glioblastoma. J. Clin. Oncol.40, 2005 (2022). [Google Scholar]
- 518.Lockard, R. E. & Lingrel, J. B. The synthesis of mouse hemoglobin chains in a rabbit reticulocyte cell-free system programmed with mouse reticulocyte 9S RNA. Biochem. Biophys. Res. Commun.37, 204–212 (1969). [DOI] [PubMed] [Google Scholar]
- 519.Muthukrishnan, S., Both, G. W., Furuichi, Y. & Shatkin, A. J. 5′-Terminal 7-methylguanosine in eukaryotic mRNA is required for translation. Nature255, 33–37 (1975). [DOI] [PubMed] [Google Scholar]
- 520.Steinle, H. et al. Delivery of synthetic mRNAs for tissue regeneration. Adv. Drug Deliv. Rev.179, 114007 (2021). [DOI] [PubMed] [Google Scholar]
- 521.Sahin, U. et al. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature547, 222–226 (2017). [DOI] [PubMed] [Google Scholar]
- 522.BioNTech RNA Pharmaceuticals GmbH. First-in-human study evaluating the safety, tolerability and immunogenicity of i.n. administration of a personalized vaccination with IVAC MUTANOME vaccine w/o initial treatment with RBL001/RBL002 vaccine in patients with advanced melanoma. https://clinicaltrials.gov/study/NCT02035956 (2020).
- 523.Sun, N. et al. Modified VEGF-A mRNA induces sustained multifaceted microvascular response and accelerates diabetic wound healing. Sci. Rep.8, 17509 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 524.Deng, Z., Tian, Y., Song, J., An, G. & Yang, P. mRNA vaccines: the dawn of a new era of cancer immunotherapy. Front. Immunol.13, 887125 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]