

Significance
Globally, ruminant grazing—including cattle for beef production—is the most extensive human land use. Removing cattle from pastures represents a meaningful opportunity to sequester carbon into regrowing vegetation and soils. Yet, carbon sequestration would trade off with beef production. By analyzing these tradeoffs globally in a spatially explicit manner, we identify carbon opportunity areas where removing relatively little pastured beef can result in substantial carbon sequestration, predominantly in high- and upper-middle-income countries. Removing this beef production may be compensated for by improving cattle management in sub-Saharan Africa and Brazil. By providing approaches to identify locations with minimal tradeoffs between food production and ecosystem restoration, this work can aid the design and improvement of policies related to natural climate solutions.
Keywords: ruminant livestock, natural climate solutions, intensification, dietary change, trade-off analysis
Abstract
Pastures, on which ruminant livestock graze, occupy one third of the earth’s surface. Removing livestock from pastures can support climate change mitigation through carbon sequestration in regrowing vegetation and recovering soils, particularly in potentially forested areas. However, this would also decrease food and fiber production, generating a tradeoff with pasture productivity and the ruminant meat production pastures support. We evaluate the magnitude and distribution of this tradeoff globally, called the “carbon opportunity intensity” of pastures, at a 5-arcminute resolution. We find that removing beef–producing cattle from high–carbon intensity pastures could sequester 34 (22 to 43) GtC i.e. 125 (80 to 158) GtCO2 into ecosystems, which is an amount greater than global fossil CO2 emissions from 2021–2023. This would lead to only a minor loss of 13 (9 to 18)% of the global total beef production on pastures, predominantly within high- and upper-middle-income countries. If areas with low–carbon intensity pastures and less efficient beef production simultaneously intensified their beef production to 47% of OECD levels, this could fully counterbalance the global loss of beef production. The carbon opportunity intensity can inform policy approaches to restore ecosystems while minimizing food losses. Future work should aim to provide higher-resolution estimates for use at local and farm scales, and to incorporate a wider set of environmental indicators of outcomes beyond carbon.
Pastures are areas of natural or managed grass, herbs, and other forages grazed by cattle and other ruminant animals to produce a range of goods and services, including meat and dairy. They represent the most extensive human use of land on Earth (1). While many pastures are in longstanding grassland biomes, with few trees, pastures have also been established in areas that would otherwise be forest biomes, which has historically required tree clearing. Maintaining pastures in formerly forested areas incurs a “carbon opportunity cost,” which refers to the quantity of terrestrial carbon sequestration or atmospheric CO2 removal that could occur by forgoing agricultural production and restoring ecosystems on that land (2, 3).
Nature conservation, restoration, and stewardship constitute “natural climate solutions” that can potentially sequester tens of metric gigatons (Gt) CO2, together constituting a substantial fraction of the mitigation necessary to limit anthropogenic global warming to 1.5° or 2 °C (4). A number of these solutions rely on restoring former terrestrial ecosystem carbon stocks that have been cleared or degraded by human land uses. The United Nations has recognized the large potential of ecosystem restoration for climate mitigation and other ecosystem services by launching the UN Decade on Ecosystem Restoration in 2021 (5). The Decade aims to motivate and support commitments made under the Rio Convention and Bonn Challenge by a total of 115 member countries to restore over 1 billion hectares of ecosystems (6). Restoration techniques include reestablishing degraded and eliminated forests and rewilding initiatives (5).
Removing ruminants from pastures and restoring some of these areas to higher-carbon ecosystems can contribute to meeting such restoration goals. Yet, abandoning any parcel of pastureland would entail a tradeoff: forgoing forage availability for ruminant feed and associated food (e.g., beef from cattle) and fiber (e.g., wool from sheep) production. Removing production where forage availability is lowest and potential carbon sequestration is highest would minimize this tradeoff between ruminant production and nature restoration.
Identifying locations with such minimal tradeoffs requires knowledge of both the potential restorable carbon that can be achieved and the quantity of forage forgone by removing ruminants from pastureland. Previous studies have mapped the distribution of carbon in potential vegetation and soils (2, 7, 8) in present-day pasture areas. Advancements in land cover mapping have refined our understanding of pasturelands’ total extent (1, 9). Remotely sensed observations of the productivity or yields (i.e., tons of available dry-equivalent biomass forage per hectare per year) of these pastures have only recently been made available globally on a 5-arcminute basis (10). The tradeoffs between carbon sequestration potential and pasture productivity have not previously been assessed geospatially.
This analysis integrates these data to quantify the tradeoffs between pasture productivity and associated beef production versus the potential carbon sequestered in combined living vegetation and nonliving biomass (soils and debris) by forgoing that productivity to allow ecosystem regrowth in place of pasture. We develop an indicator of this tradeoff—the carbon opportunity intensity (COI)—and we map its distribution globally at a 5-arcminute resolution. The COI is a proportion, wherein the quantity of potential ecosystem carbon sequestration (in kg CO2) is the numerator and quantity of pasture production (of either kg dry biomass pasture or kg protein from beef raised on those pastures) is the denominator. A high COI thus indicates the greatest potential for ecosystem carbon sequestration relative to the least amount of pasture (or beef) productivity forgone. A low COI, conversely, means less potential ecosystem carbon sequestration would follow the abandonment of production on current pastures, in relation to the quantity of current pasture production forgone. Therefore, maximizing carbon sequestration while minimizing lost beef production would entail keeping low-COI pastures in production and restoring high-COI pastures to regrow vegetation to late-successional i.e. climax communities, including forests. We identify high-COI pastures that have potential to be the highest-priority areas for ecosystem restoration and carbon sequestration into vegetation and soils.
In developing regions including much of sub-Saharan Africa, and parts of South America and Asia, beef production on pasture has low efficiency, meaning high forage quantities are required for weight gain, hence beef production. However, across all livestock species and production systems, including exclusively pasture-raised cattle, production is expected to become more efficient over time (11). Further efficiencies could be accomplished with additional investment into managing pastured cattle herds (12). Some improvements are possible without shifting cattle production from pastures onto feedlots—such techniques are referred to as pastoral intensification or semi-intensification (13), which include access to vaccines and veterinary services, more frequent reproduction rates, and younger animal slaughter ages. It may be possible to alleviate production losses from ruminant removal and ecosystem restoration in some areas by increasing production in other low-efficiency areas, e.g., Africa and South America. Increased efficiency could minimize the food-for-carbon tradeoffs (lower COI) and at least partially offset the targeted removal of beef production from high-COI areas.
We quantify and compare the COI of beef production on pastures across three scenarios of pasture forage-to-beef conversion ratios, one of present-day efficiencies and two others of improved efficiencies. This comparison highlights regions where intensification through more efficient herd management could reduce the COI of beef production and replace the productivity lost by restoring the highest-COI pastures. We differentiate native grasslands from areas of potential forest regrowth, where climax vegetation is expected to be dominated by late-successional trees rather than grass-dominated landscapes in current pastures. Existing studies have quantified the carbon opportunity cost of pastures in potential forest areas (2, 3), but have not compared potentially large differences in the productivity of pastures for beef production across these regions. Through this analysis, we aim to advance understanding of how present-day land use for ruminant food production trades off with potential carbon sequestration in restored vegetation and soils of terrestrial ecosystems. Our approach has potential for further use in policy design and scenario analyses that weigh these competing priorities.
Results and Discussion
Carbon Tradeoffs of Global Pastures.
Across global pastures where ruminant animals (i.e., cattle, sheep, and goats) are reared, we find that pastures have on average >7 times more carbon per hectare in restorable ecosystem vegetation and soils when they are established in potential forest areas (170 tons C [tC or MgC] ha−1/624 tCO2 ha−1) than do pastures in native grassland biomes (22.1 tC ha−1/81 tCO2 ha−1) (Table 1). Our analysis of these quantities reflects the carbon stocks in potential restored ecosystems integrated over 75 y, or near the end of the 21st century. Ecosystem carbon pools that have not reached carbon storage saturation throughout that time (namely, tropical forest vegetation and boreal soils) would sequester yet more carbon after this duration.
Table 1.
Carbon sequestration of potential ecosystem restoration, pasture productivity, and area of all pastures in potential vegetation biome types (forest, grassland), expressed in units of carbon
| Removals from Potential Ecosystem restoration | Pasture Productivity |
||||||
|---|---|---|---|---|---|---|---|
| Cumulative to 2100 (MgC ha−1) | Annual Flux (MgC ha−1 y−1) | ||||||
| Potential vegetation type | Soils | Vegetation | Soil + veg combined | Mg DM ha−1y−1 | |||
| Area-weighted | Forest | 64 | 107 | 170 | 3.4 | 3.35 | |
| (50, 68) | (60, 150) | (109, 216) | (2.2, 4.4) | (3.02, 3.68) | |||
| Grassland | 12.7 | 9.5 | 22.1 | 0.62 | 0.99 | ||
| (0.7, 18.7) | (3.4, 17.4) | (4.1, 36.0) | (0.16, 1.22) | (0.89, 1.09) | |||
| Pasture Area | Cumulative to 2100 (GtCO2) | Annual Flux (GtCO2 y−1) | |||||
| Mkm2 | Soils | Vegetation | Soil + veg combined | Gt DM y−1 | |||
| Global sums | Forest | 7.13 | 164 | 281 | 445 | 8.8 | 2.55 |
| (129, 174) | (157, 391) | (286, 565) | (5.7, 11.6) | (2.33, 2.76) | |||
| Grassland | 20.31 | 94 | 71 | 165 | 4.6 | 2.96 | |
| (6, 138) | (25, 129) | (31, 268) | (1.2, 9.1) | (2.68, 3.24) | |||
| Both | 27.44 | 259 | 351 | 610 | 13.4 | 5.51 | |
| (135, 312) | (182, 521) | (316, 833) | (6.8, 20.7) | (5.01, 6.01) | |||
Stocks reflect accumulation rates over 75 y, ending in 2100, with still more sequestration occurring after that point. The first two rows of entries refer to per-hectare metrics, while the lower rows refer to all areas summed globally. Parenthetical ranges reflect 95% confidence intervals across each grid cell, averaged or summed over all grid cells. DM = dry matter.
Removing grazing from all pastures identified as potential forest areas (7.13 Mkm2) could sequester substantial carbon in vegetation and soils through the end of this century—121 (78 to 154) GtC i.e. 445 (286 to 565) GtCO2 (Table 1) (SI Appendix, Figs. S1 and S2). This total quantity is approximately 20% greater than a prior estimate by Hayek et al. (3), which contained a less robust estimate of present and potential soil carbon stocks, but the two estimates have overlapping 95% CI nonetheless. Restoring all potentially forested pastures globally would sequester enough carbon to remove more than the past twelve years of fossil fuel emissions from the atmosphere (14). By comparison, reducing or eliminating grazing in all native grassland areas (20.31 Mkm2) would sequester 45 (8 to 73) GtC i.e. 165 (31 to 268) GtCO2. This quantity is significantly less than the total carbon sequestration of potential forests following livestock removal, and has a larger uncertainty range, by percentage, due largely to more heterogeneous estimated responses of soil carbon (SI Appendix, Fig. S1).
However, pastures in potential forest areas also produce three times more aboveground biomass (AGB) for ruminant consumption than do pastures in native grassland areas (Table 1 and SI Appendix, Fig. S3). Therefore, despite the total global area of pasture in potential forest biomes being approximately one third of the area of pasture in native grasslands, total pasture productivity is comparable between these two domains. Of the annual 5.51 gigatons dry matter (Gt DM y−1) currently produced on pastures globally, 46% is in potential forest areas (2.55 Gt DM y−1) and 54% is in native grassland areas (2.96 Gt DM y−1). Restoring all potential forest areas currently under pasture globally would therefore eliminate almost half of ruminant livestock forage. In summary, the greatest carbon opportunity cost lies in areas that also support the highest pasture productivity and associated ruminant food and fiber production.
Most pastures in native grassland areas have relatively low and positive COIs (Fig. 1A). In some native grasslands (e.g., Northern China, Western Australia), removing livestock may reduce ecosystem stocks of carbon (i.e., lead to net ecosystem carbon emissions rather than sequestration) due to expected soil carbon losses (SI Appendix, Figs. S1 and S2). These areas have a negative COI (Fig. 1B and SI Appendix, Figs. S1 and S2). In our analysis, the COI only reflects differences between present ruminant grazing management versus removing cattle entirely, and cannot account for alternative stocking density or alternative grazing management approaches. However, research suggests that relative to ruminant removal, shifting management to lightened or rotational grazing could lead to positive soil carbon accrual or no net change, and/or confer vegetation cover benefits in certain areas (15, 16). Recent analysis suggests that optimal stocking densities for improving carbon sequestration are well below present densities, resulting in decreased meat and milk production (17), but this analysis did not test alternative (e.g. rotational) grazing techniques due to sparse data on its effects. Additional research is needed to assess whether rotational or other alternative grazing management strategies, if applied across all native grassland regions globally, would result in soil carbon sequestration (i.e. higher pools/stocks), relative to soil carbon under either present management or ruminant removal, across native grasslands globally, and how stocking densities, hence meat production, would change. Nevertheless, the largely positive effects of ruminant exclusion upon grassland soil organic carbon stocks, relative to the current management practices reported here (SI Appendix, Fig. S2), have been confirmed by multiple recent systematic reviews (16, 18, 19).
Fig. 1.

Spatial distribution of carbon opportunity intensity (COI) of pasture production. A large COI corresponds to a high quantity of potential carbon sequestered from regrowing ecosystems after abandoning a given quantity of pasture production in an area. (A) Spatial distribution of COI of pasture production at 5-arcminutes resolution. Pastures with zero or negative COI are visualized here as zero (gray), and can be seen in SI Appendix, Fig. S5. (B) Area-weighted mean carbon opportunity costs of pasture for geographic regions. EAS: East Asia. EUR: Europe (excluding Russia). LAM: Latin America and Caribbean. MNA: Middle East, North Africa, and Central Asia. NAM: North America. OCE: Oceania (excluding Southeast Asia). RF: Russian Federation. SAS: South Asia. SEA: South East Asia. SSA: sub-Saharan Africa. DM = dry matter.
Humid, evergreen tropical forests tend to be more carbon-dense than temperate forests (20, 21). However, we find that pastures in tropical potential forest regions in South America and sub-Saharan Africa have lower COIs than pastures in temperate potential forest regions (Fig. 1B). This is due largely to high, year-round pasture productivity in the tropics (SI Appendix, Fig. S3). At high latitudes, pastures tend to exhibit high COIs (Fig. 1A) due to both lower pasture productivity and moderate potential stores of carbon in vegetation and soils, with especially high potential carbon stores in boreal soils (SI Appendix, Fig. S1). Forests in Maritime Southeast Asia tend to be uniquely inefficient for pastures; they have high COI due to both low pasture productivity and high densities of native ecosystem carbon. Regardless of these general tendencies, there is heterogeneity in both pasture productivity and potential ecosystem carbon in all regions; some pastures in potential forest areas on every continent exhibit high COIs (dark green areas, Fig. 1A).
Pastured Beef Production at Current and Improved Efficiency.
Livestock removal from high-COI pastures potentially offers an opportunity to sequester substantial carbon with little reduction in ruminant production. We explore the consequences specifically of removing beef production from high-COI pastures, as more pasturelands are used to produce cattle for beef than any other ruminant animal-sourced food (17).
The feed conversion ratio (FCR) of cattle raised for beef on pastures is relatively high in many tropical regions (22) (SI Appendix, Fig. S6) signifying a low production efficiency. Some tropical forest regions have both low pasture productivity, high carbon in potential forests (i.e. high pasture COI), and low beef production efficiency (i.e. high FCRs) occurring simultaneously; we thus observe the highest COI values of beef production in these regions (Fig. 2) [e.g., Maritime Southeast Asia (SI Appendix, Fig. S3)]. In other potential tropical forest areas, there is a relatively lower COI of beef production, particularly in sub-Saharan Africa, due to high pasture productivity counterbalancing high FCRs.
Fig. 2.

Area-weighted mean carbon opportunity costs of pasture-raised beef production in areas of potential forest. Light green corresponds to current pasture-raised beef FCRs. Dark green corresponds to improved pasture-raised beef FCRs: assuming 90% yield gap closure between present FCRs and a yield ceiling of area-weighted mean beef FCR from all OECD countries (202 kg DM pasture per kg beef protein). LAM: Latin America and Caribbean. MNA: Middle East, North Africa, and Central Asia. NAM: North America. OCE: Oceania (excluding Southeast Asia). RF: Russian Federation. SAS: South Asia. SEA: South East Asia. SSA: sub-Saharan Africa.
We present a hypothetical scenario of improved FCRs, wherein countries intensify beef production on pastures to a similar level of efficiency to high-income countries. Efficiency improvements may be possible through management changes such as improving stocking densities and rates, reproduction, and animal health. Our comparison suggests that more efficient cattle herd management on pastures could reduce the COIs of each region’s beef production fed on grass in potential forest areas (Fig. 2). The largest relative reductions in these COIs in potential forest areas were by 67% in South Asia, 59% in sub-Saharan Africa, and 46% in South East Asia. Improving herd management on pastures also tends to mitigate emissions of other greenhouse gases such as methane (22).
Opportunities for Potential Carbon Sequestration.
Even if beef FCRs are improved through herd management, COIs of pasture-raised beef would tend to remain comparably large in potential forest areas on all continents relative to native grassland areas (as demonstrated in Table 1 and Fig. 1). Specifically, the area-weighted median COI of beef production, under the scenario of improved FCRs, is 1,893 kg CO2 per kg beef protein across potential forest areas, compared to 641 kg CO2 per kg beef protein across native grassland areas. We map areas of potential forest where the improved-FCR beef production COI is greater than the median value, highlighting high-COI areas where >10% of the grid cell consists of pasture (Fig. 3; orange-red areas). Removing beef-producing cattle from these high-COI pastures and allowing them to revert to forests could lead to sequestration of 34 (22 to 43) GtC i.e. 125 (80 to 158) GtCO2, 37 (24 to 47)% of the total carbon opportunity cost of all beef-producing pastures. This would remove only 184 Mha of pasture, which produces 420 Mt DM of forage per year, corresponding to only 8% of global pasture productivity. The result would be removal of 560 (376 to 773) kilotons [kt] beef protein from the food system, equivalent to 13 (9 to 18)% of the global total beef produced on pastures (SI Appendix, Fig. S8 and Table S1).
Fig. 3.
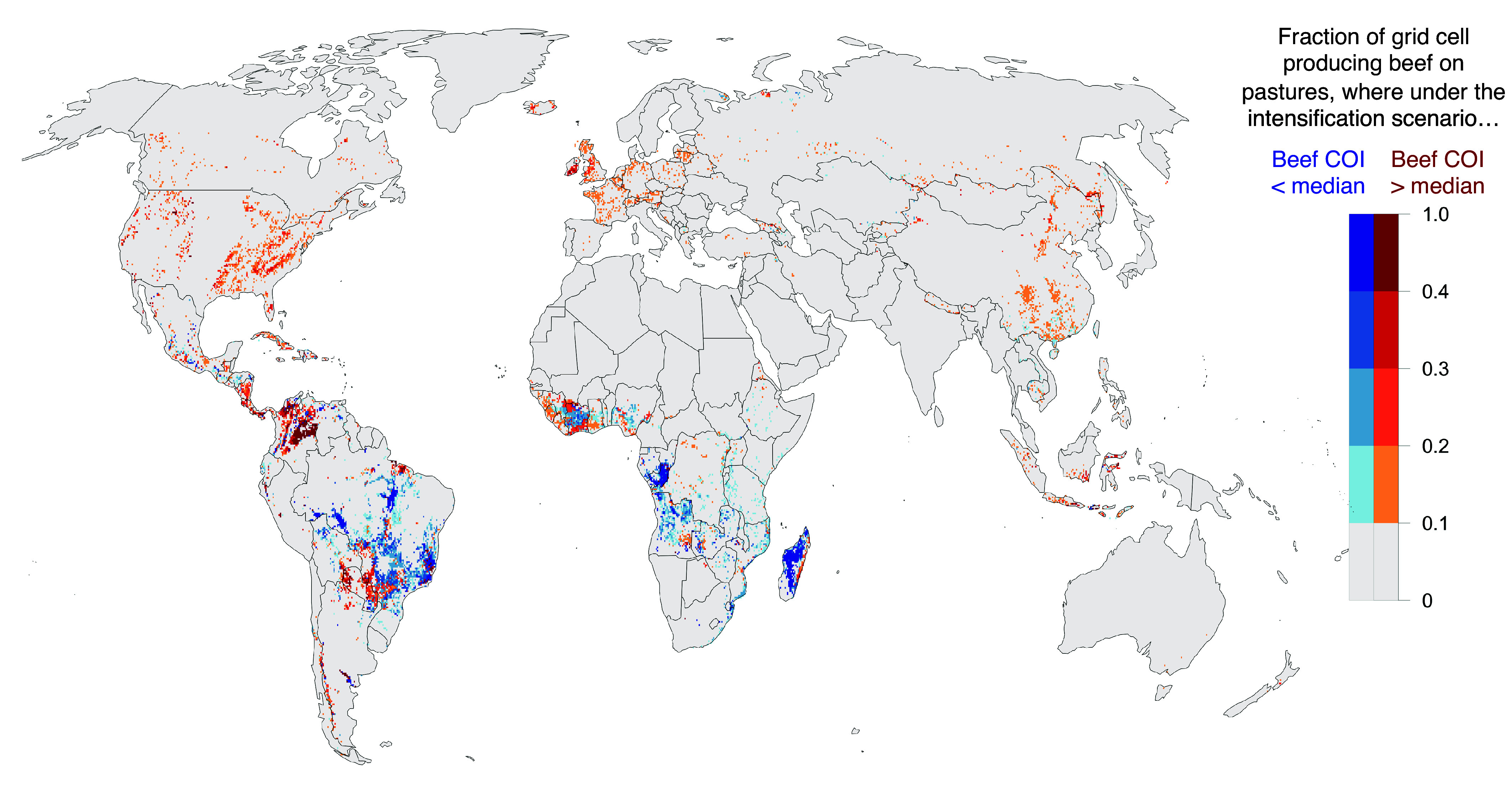
Areas of high carbon opportunity intensity of beef production. High carbon opportunity areas are those greater than the median COI of 1,893 kgCO2 per kg beef protein. Red areas have COIs in the scenario of improved FCRs that still exceed this threshold, and >10% of a gridcell consists of beef-producing pasture area. Blue areas have COIs under current FCRs exceeding this threshold, but COIs are below this threshold in a scenario of improved FCRs, and >10% of the gridcell consists of beef-producing pasture area. In other words, blue areas demonstrate where intensification through improved herd management achieves a more favorable tradeoff of beef production for carbon, whereas red areas demonstrate where these tradeoffs are currently unfavorable, and greater CO2 sequestration could be achieved through removing beef production.
To understand where improved herd management might generate a substantial reduction in the COI of pastured beef, we identify potential forest areas where beef production would be less than the improved-FCR median COI (Fig. 3; blue areas) if herd management and efficiency on pastures were to be greatly improved. This corresponds to an area of 198 Mha of pasture in potential forest areas. These pasture areas are currently responsible for 1,174 kilotons (kt) beef protein production, but could produce 1,920 kt protein if intensified, the difference of which is 745 (515 to 1,005) kt protein, or 18 (12 to 24)% greater beef production on all pastures than present-day (SI Appendix, Fig. S9). This increase is greater than (or approximately equal to, within uncertainty) the amount of beef lost by removing production from high-COI areas (560 kt protein). Many of the areas where intensification has the greatest potential to improve (i.e., lower) COIs are in the tropics, particularly in sub-Saharan Africa (SI Appendix, Table S1). However, pasture beef production could be intensified in all areas including native grasslands, and not merely in areas of potential forest. Intensifying all remaining pastures would produce 1,568 (1,074 to 2,130) kt or 37.3 (25.6 to 50.7)% greater beef protein production on all pastures than present-day (SI Appendix, Fig. S9). This would lead to net carbon sequestration in former pastures without reducing global beef output.
However, this amount of intensification (90% of FCR values observed in OECD countries—Methods) may not be feasible. Economic barriers to intensification include a lack of access to resources such as vaccines, extension services, infrastructure, and supplements, while sociocultural barriers may be present in areas where management or cultural systems are not conducive of minimizing pasture FCRs to such an extent (e.g., due to the role of animals in religious systems, or their value as draft animals). Additionally, our scenario does not account for the presence of different cattle species or breeds, or their potentially varying intensification requirements across regions, which requires further research.
Because such a high degree of intensification in potential forest areas may be infeasible, we also explored the degree of intensification needed for the global addition of pastured beef production on all pastures—in both potential forest and native grassland areas—to equal the quantity of removal from high-COI areas (i.e., 560 kt beef protein). This would require intensifying all pastured beef production, outside of removal areas, to the level of 47% of OECD FCRs (SI Appendix, Table S1). While such a shift would result in the same quantity of beef production globally (intensified beef production minus beef production removals), it would result in a geographic redistribution of production. Decreased beef production would occur predominantly in northern latitudes and in high- and upper-middle-income countries (SI Appendix, Table S1), necessitating reduced beef consumption, reduced beef exports, or increased beef imports to these countries.
As a sensitivity analysis, we analyzed and report the results from a range of other potential COI thresholds for determining high-COI areas besides the median COI value (in 5-percentile intervals, SI Appendix, Fig. S10). A lower threshold would remove less pastured beef and improve the proportion of carbon removal relative to the amount of pastured beef removed, but would sequester less carbon globally. Increasing the COI threshold would reduce the remaining area of pastures available for intensification, hence reducing the quantity of beef that could be increased through intensification on this land (SI Appendix, Fig. S11).
The potential for intensification that we estimate here is an approximation of a technically feasible potential. It does not represent the amount that is economically or otherwise feasible in the face of resource constraints especially by smallholder producers. Nor do we estimate a particular timeframe within which this intensification could occur. The timeframe might be substantially shorter or longer than our chosen timeframe for ecosystem carbon sequestration, which we estimate until approximately the year 2100. Moreover, our estimates for intensification are limited to changes in herd management on the existing pasture resource base (e.g., improving fecundity, or animal health), and do not consider changes to managing the pasture resource itself (e.g., seeding, fertilizing, or irrigating pastures), as we presently lack global, spatially resolved data on pasture management. Nonetheless, our results demonstrate that qualitatively, intensification through cattle herd management and forest restoration can play complementary roles across regions.
Conclusions
Our analysis demonstrates substantial geospatial variation in tradeoffs between the production of forages, ruminants, and associated animal-sourced food, and carbon sequestration in potential ecosystems, globally. We find that simultaneously improving the efficiency of beef production in a number of economically developing regions while restoring potential ecosystem vegetation and soils in high-COI opportunity areas of pastured beef production would have a negligible or positive impact on the global total beef production from pasturelands. Such a scenario would require reducing beef production in some high- and middle-income countries including the United States, China, and European countries (SI Appendix, Table S1). Our results also demonstrate opportunities to restore ecosystems while maintaining or increasing beef production in developing, natively forested tropical regions without further expansion of pastures.
Yet, to date, few policy proposals for reforestation engage seriously with the wide geospatial variation in tradeoffs between agricultural production and carbon sequestration via ecosystem restoration. Policy proposals for natural climate solutions (4, 23), strategies and calls to restore ecosystems (5), and country-level commitments (6) often fail to acknowledge agriculture-restoration tradeoffs. These ecosystem restoration proposals contrast with recent scientific work that has analyzed tradeoffs between carbon sequestration via restoration versus croplands productivity for food and animal feed (24–26).
Here, we have focused on the largest category of land use for beef production—pasturelands—using recent remotely sensed observations of pasture productivity at a relatively high spatial resolution with global coverage. We identify a clear food production/carbon sequestration tradeoff in most pasturelands, and wide spatial variability in this tradeoff. On average, the magnitude of this tradeoff is especially large in areas of potential forest regrowth, but pasture productivity in these potential forest areas varies greatly, across large countries and regions, leading to widely varying tradeoffs.
Our key tradeoff indicator, the COI of pastures, can be used for policy design and analysis to improve the effectiveness of plans to restore nature for carbon sequestration. However, the COI estimates presented here nonetheless have some key limits that should be acknowledged. This analysis only reflects the proportion of beef production that was raised using forages from pastures, while excluding the proportion of beef raised from concentrated crop feeds (e.g. grain, oilseeds), cropland-grown forages (e.g., alfalfa, maize silage), or inedible crop residues (e.g. maize stover, husks) and other occasional feeds. Calculating the full mitigation benefits possible from removing all beef production in a given area additionally requires adding COIs from crop-based feeds to the pasture-based feeds, and also calculating production emissions from the feeds and the ruminant animals themselves. Future work should aim to combine the COI of the total lifecycle of animal feeds with analysis of the fluxes of greenhouse gases from ruminants and their various feed sources. The COI threshold used herein—the median COI of beef-producing pastures in potential forest areas—is only one illuminating threshold for evaluating such tradeoffs. Further consideration should be given to the diverse (e.g., financial; cultural; ethical) values and impacts of agricultural production as compared with multiple social and environmental benefits of ecosystem restoration. Economic and sociocultural research should investigate these potential tradeoffs in further depth.
Additionally, removing and intensifying ruminant production may lead to unintended risks for ecosystems on medium- to longer-term timescales. Two such effects include leakage—wherein ruminant production removed from current pastures in one area leads to deforestation in another—or rebound effects—wherein intensification decreases costs and/or increases profits, thus incentivizing additional land clearing. Regional economic analyses in forested regions such as South America and Central Africa demonstrate that rebound and leakage effects can be partially prevented through public and private forest protection policies (27–31). The COI indicator should be coupled with further integrated economic and ecological research to understand how removal and intensification of agricultural production, along with conservation policies, may dynamically affect ecosystem restoration.
Despite these limitations, we suggest that policymakers who influence management and use of agricultural lands for climate mitigation should align policies to consider COI, among other dimensions, to jointly evaluate the expected ecosystem carbon sequestration from land restoration versus food production under improved management. For countries with ample land in potential forest biomes currently managed as pastures for beef production, and food system stakeholders in these regions, our results can assist in geospatially explicit scenario analyses to visualize and quantify the potential for carbon sequestration from natural climate solutions under different policy instruments and scenarios.
This work extends previous research that suggests multiple policy interventions to reduce land requirements for agriculture are needed to meet climate targets in the agriculture, forestry, and other land use (AFOLU) emissions sector (32). We find that potential ecosystem restoration in many global pasture locations would not produce much carbon removal benefit and/or would substantially reduce ruminant production. We therefore suggest careful targeting of restoration activities across heterogenous pasturelands to optimize the tradeoff between ruminant animal removal and ecosystem carbon restoration.
Future analyses should incorporate a wider range of socioenvironmental tradeoffs and cobenefits associated with pasture intensification or abandonment in areas of potential forest restoration, such as biodiversity, landscape connectivity, hydrological fluxes and water services, and livelihoods. These analyses can benefit the design and implementation of policies that account for the fact that agricultural productivity, and its tradeoffs with ecosystem services such as carbon sequestration, are heterogeneous (24, 33). Because pastures occupy so much land area, and incur the largest opportunity cost in the food system (3), the opportunity-intensity framework can improve conservation outcomes while minimizing food production losses on meaningfully large scales to help meet restoration and climate targets.
Methods
Global Pasture Production Map.
We calculated global pastures productivity, producing a map of ruminant-available aboveground biomass (AGB; units kg available dry weight per ha), in a manner following Piipponen et al. (10). We used publicly available MODIS-derived rasters of annual net primary production (NPP) across a 20-y time series (2001–2020) at 500 m resolution. These were then parameterized to ruminant-available aboveground biomass (AGB) using temperature, precipitation, and terrain slopes using the following equations:
| [1] |
| [2] |
| [3] |
where 0.49 is the conversion of dry matter in vegetation to carbon; slopesmultiplier is a scale factor between zero and one with input from a remotely sensed and modeled 90 m slope estimate (34) and output from Piipponen et al. and references therein (10); fANPP and fBNPP are above- and below-ground fractions of total NPP. MAT and MAP are mean annual temperature and precipitation respectively for 2001–2020 (35). The resulting AGB 500 m grid cells were aggregated to 5-arcminutes by masking nongrassland categories of IGBP (International Geosphere–Biosphere Programme classification system) rasters and averaging to the larger 5-arcminute grids.
Total production of pasture (in weight of dry matter) in each 5-arcminute cell was then parsimoniously calculated as AGB * Pasture Area = weight of dry matter. Pasture AGB methods for this analysis differed from Piipponen et al. (10) in two key ways: [1] a wider range of land classifications were considered as grasslands containing grazable biomass, including open and closed shrublands were considered as grazing lands (further details below) [2] the aboveground fraction of NPP was estimated based on Sun et al. (36), who synthesized extensive field observations and propose that the fraction of aboveground NPP (fANPP) depends on mean annual temperature (MAT) and mean annual precipitation (MAP) as described above in (Eq. 2).
This analysis adopted a different functional classification “pastures” from Piipponen et al. 2022 (10), which includes (a) more International Geosphere–Biosphere Programme (IGBP) classes (37), encompassing Closed Shrublands, Open Shrublands, Woody Savannas, Savannas, Grasslands, and Barren lands (IGBP classes 6 to 10 and 16). This was then constrained by (b) grid cells within these IGBP classifications for all 20 y, and (c) only 5-arcminute cells that are also included in the HYDE v3.3 map, for base year 2020, to reflect estimates of recently cleared pastures in the tropics. We take the sum of combined area of grid cells that are under livestock pastures and rangelands (permanent and cleared) (9). This creates a more accurate representation of exclusively ruminant-grazed areas, and excludes areas that are underutilized or not utilized at all by ruminants, such as highly intensive crop production areas, as well as wildlife preserves where wild ruminants are grazed, and other conservation areas where domesticated ruminants are likely excluded. Nonetheless, we acknowledge, as the HYDE analysis authors do, that there remain limitations to this or any approach for consistently estimating the global distribution of pasturelands, which can create discrepancies between global and local estimates; these limitations include uncertainties in the human population estimates in remote areas with extensive pastureland grazing, and errors in UN FAO census-derived estimates of pastureland area, among others (9). Altogether, the HYDE-derived estimate represents a conservative total land area of pastures (27.4 Mkm2) compared with other previous estimates such as Hayek et al. and references therein (1, 3, 38).
Ecosystem Carbon.
Carbon in present-day and potential vegetation.
Carbon stocks in present-day vegetation were derived from seven different 5-arcminute maps from Erb et al. (7) that utilize ecosystem observations, and additional national and subnational data on tree cover, grazing, forestry, climate, and economic activity, that empirically model and interpolate observations across the grid. These grids were used to model carbon stocks in present-day pastures in native grassland biomes.
For present-day pastures in potential forest areas, we assumed mean vegetation stocks in present-day pastures of 6 ± 3 MgC ha−1, consistent with literature estimates including the Erb et al. analysis (3, 7) because potential biome areas are assumed to contain a heterogenous or patchwork mix of established pasture and forest within 5-arcminute grid cells. This is consistent with our assumptions that treat the tree-covered fraction of present-day 5-arcminute grid cells as nonpasture area. We do not assume any interannual sequestration (or net flux) occurs in present-day ecosystems in vegetation (and soil). Rather, we assume that the (net) flux of carbon into or out of these ecosystems is zero, i.e. a steady-state assumption.
Carbon stocks in potential vegetation were derived from seven maps: six from Erb et al. (7) using a global observations with similar empirical modeling, and reflecting current climatic conditions, and one from Searchinger al. (2) that used a mechanistic global dynamic vegetation model, the Lund–Potsdam–Jena managed land model (LPJmL) run to equilibrium under current climate conditions.
The gridded ecosystem vegetation carbon data described in and derived from Erb et al. (7) come from disparate sources and contained numerous gaps, which were filled by the authors using combinations of remote sensing, census statistics, and environmental correlations. Thus, the data in each map are not strictly comparable, but the mean in each gridcell thus reflects the average across disparate sources of information. The distributions around the means for each grid cell are reflected throughout our results. Uncertainty ranges are provided for all estimates as 95% CI reflecting the joint distribution across all seven maps of actual and potential carbon stocks in vegetation for each grid cell (described in further detail below). In this way, comparisons between potential and actual stocks of vegetation versus the productivity of present-day pastures, performed in our COI analysis, represent robust estimates of the tradeoffs in ecosystem carbon, albeit not strictly harmonized. Future work should aim to compare these estimates to observations derived from in situ measurements and process-based models.
We defined “potential forest” areas in our analysis as any of eight potential forest biome types, as well as savanna areas of sufficiently carbon-dense vegetation, as a proxy for high potential tree cover (corresponding to potential carbon stocks ≥75 MgC ha−1) consistent with Hayek et al. (3). “Potential grasslands” were defined as potential biomes types of grassland, shrubland, or tundra, and savannas with comparatively less tree cover (potential carbon stocks < 75 MgC ha−1) to reflect sparse and open-canopy savannas with sufficient open space and available forages for ruminant grazing. Potential vegetation categories and carbon stocks reflect present climate, and do not reflect a range of future anthropogenic climate scenarios.
Carbon in soils and other nonliving pools.
Stocks of soil carbon in potential ecosystems were also taken from Searchinger et al. (2) from the same model and scenario as their potential vegetation results—the LPJmL model run to equilibrium under current climate conditions. These data provide soil carbon stocks in tons per hectare to a depth of one meter.
Present-day soil carbon in pastures was derived from the United Nations Food and Agriculture Organization (FAO) Intergovernmental Technical Panel on Soils (ITPS) GSOCmap product version 1.5 (39). This 30-arcsecond resolution map reflects soil organic carbon stocks in tons per hectare to a depth of 30 cm, across a variety of land uses. To create a product compatible with the 1-meter depth Searchinger et al. soil organic carbon map of potential ecosystems, we extrapolated the present-day GSOC map to 1 m depth using proportions derived from coarser-resolution datasets of soil carbon at two depths (40) between 0 to 30 cm and 30 to 100 cm. Then, to create a product reflecting only soil organic carbon in present-day pastures, we masked the high-resolution cells for pasture areas (similarly scaled from the aforementioned 500 m resolution IGBP map above), and then averaged the masked pasture-only cells to 5-arcminutes resolution, generating a 5-arcminute resolution product reflecting soil organic carbon on present-day pastures only.
Additionally, we added parametric estimates for carbon in coarse woody debris and leaf and fine woody litter, in potential forest areas, consistent with the “nonliving pools” scenario in supplemental methods in Hayek et al. and references therein (3, 41, 42).
Carbon opportunity costs of pasturelands.
Carbon opportunity costs of pasturelands were calculated on every grid cell on a per-hectare basis of both annual fluxes and stocks integrated over 75 y. The latter is intended to approximate a scenario of hypothetically abandoning pastureland starting around year 2026 and estimate the resulting carbon stocks in potential vegetation around the end of 2100. Our pasture area data reflect a conservative estimate for the 2001–2020 time range, but we lack more recent data on pasture areas. Thus, using this 2026–2100 75-y time horizon in our analysis is intended to reflect a conservative estimate of the ecosystem restoration potentials possible in the long term by implementing land use changes resulting from consequential policies (e.g. production shifts, dietary change, conservation mechanisms, etc.) relatively soon. Therefore, in our estimate, the 75-year total C stocks do not necessarily reflect climax vegetation in all cells, but rather, only reflect climax vegetation in biomes that are estimated to recover nearly all of their carbon stocks within years since abandonment. For all other biomes (e.g. evergreen tropical forests, boreal forests), further carbon accumulation would be expected after 2,100, even though this further regrowth is not reflected in our results.
Net ecosystem productivity C flux estimates were derived from Searchinger et al. (2) and reflect approximate rates of reaccumulating ecosystem carbon following abandonment of pastureland. We calculated uncertainty in this flux estimate by (1) calculating the distribution of net C flux across all grid cells in each potential biome type, and then (2) assuming the distribution in each grid cell of C fluxes were proportional to the distribution of C fluxes in the seven potential grassland types. This created a sample distribution of mean flux estimates from which we derived 95% CI.
Carbon stocks through 2100 were calculated by applying estimates of annual ecosystem fluxes to each grid cell, multiplying by 75, and taking the minimum of that integrated carbon estimate and the climax ecosystem carbon stock in each grid cell. Uncertainties were calculated from (1) the net C flux distribution across all grid cells in each biome type and (2) a compounded error distribution of seven potential C stock values in each grid cell with the distribution of seven actual C stock values in each grid cell. For pasture in potential forest areas, errors in actual vegetation were assumed to be 50%, i.e. 6 ±3 MgC ha−1. We then calculated 95% CI from these compounded carbon stock error distributions.
Carbon Opportunity Intensity of Pastures.
We calculated the carbon opportunity intensity (COI) as a quotient: the carbon flux into potential ecosystems over the productivity of present pastures (potential ecosystem flux in MgCO2 ha−1 y−1 over pasture AGB in Mg DM ha−1 y−1). This is distinct from ongoing production emissions, which tend to represent recent land use changes such as deforestation as a present emission (2, 43). This analysis reflects two COI indicators: the COIs of pasture productivity, and the COIs of the proportion of beef production that was raised using forages from these pastures (following section), while excluding the proportion of beef raised from concentrated crop feeds, cropland-grown forages (e.g., alfalfa, maize silage), or occasional feeds. This analysis is limited to the geographic distribution of the pasture-grown component of ruminant production, as well as the proportion of beef that can be raised using only inputs from that pasture.
Feed Conversion Ratios for Cattle Fed on Pasture.
Feed conversion ratios (FCRs) for beef on pastures were derived from a dataset produced by Herrero et al. (22) at the regional level, which we renormalized to account for country-level compositions of herds of large and small ruminants (44). These data proportion pasture land area in each country by beef herds, dairy herds, and small ruminants (goats and sheep) producing dairy and meat. FCRs were calculated at the country on a commodity basis by including beef produced from both dedicated beef herds and beef from dairy-producing cattle herds.
FCRs are expressed as kg edible feed per kg edible protein produced. These values assume a global conversion ratio of carcass weight to edible protein, derived from the FAOSTAT database. The estimates may therefore underestimate protein concentration in meat in regions where beef systems are low-production and/or exclusively grass-fed, where meat is typically leaner, and overestimate protein concentrations where beef systems are high-production and more grain-finished, where meat is typically fattier. FCRs were taken as the weighted median of all grazing-based systems for each region and country, weighted by the proportions of herds producing beef and type of material grazed. These include arid, humid tropical, and temperate grasses, with different FCRs associated both with production efficiency and the quality of grasses available in these pasture regions (e.g., typically higher FCRs in arid systems, lower FCRs in temperate systems) (22). The resulting FCRs are aggregated at the country level. The FCRs and the proportion of pasture land dedicated to each commodity were then distributed evenly across every grid cell in each country. Therefore, while FCRs reflect present-day combinations of forage quality and cattle herds, our scenarios of beef removal and intensification that change these proportions may result in changes that do not fully reflect the distribution of variation of FCRs within countries that have multiple grass qualities and herd types.
These FCRs reflect “grassland-based” systems of cattle according to the FAO classification system (45) which refers to the method of feeding cattle ≥90% local grass and forages on pastures; it does not refer to the native or endemic vegetation type. These FCRs do not reflect production in mixed or intensive systems from nonpasture feeds (which have lower, i.e. more efficient, FCRs of feed:beef). Because many cattle herds are not exclusively pasture-raised, but fed mixed rations, our use of grassland-only (i.e. pasture-fed) Our use of grassland-based FCRs are intended to approximate the beef productivity in each country that only the pasture-grown forages were responsible for, disaggregated from the additional beef production achieved by feeding cattle cropland-grown forages and grains. This approximation was validated against Hayek and Garrett (46), which used a top–down approach and a demographic model to ascertain that pasture-raised forages could produce 27% of United States’ current beef production. The grassland FCR approach in this analysis produced a similar result: 26% of total beef production resulted from pasture-based feeds (407 kt out of 1538 kt total beef protein production). Therefore, this analysis does not fully account for variations in nutritional quality in feeds present in previous work, because we do not consider cropland-grown forages and grains, which tend to be of higher digestibility.
We assume a mean “sustainable grazing fraction” of 42% of AGB for every 5-arcminute grid cell. This figure is based on a literature review of observed sustainable grazing offtake rates (10). Our estimates of the mean and uncertainty of pasture-based beef production are based on the observed AGB adjusted by this offtake rate with a SD in the literature estimates of ±15% (in absolute percentages: 28 to 57% range). This makes 2.33 (1.59 to 3.22) Gt DM available to ruminants. Our estimate of total annual grazed forages on pastures sits in between two global estimates of global pasture offtake by two other sources: Herrero et al. estimated 2.30 Gt DM (22) and the UN FAO Global Livestock Environmental Assessment Model (GLEAM) used an estimate of 2.76 Gt DM (47). While our sustainable grazing fraction is high by some estimates, this may be compensated for other potentially intractable biases in the analysis, including the estimate of overall pasture NPP or the allocation of belowground NPP fraction based on empirical climatic relationships (Eq. 3). A spatially explicit estimate of the sustainable grazing fraction is desirable, but no such geospatial estimates exist to date.
Our approach of using a global grazing fraction was preferable to geospatial methods for imputing current grazing intensity because of data limitations. Some products contain global, spatially explicit estimates of stocking density—e.g. UN FAO’s Gridded Livestock of the World (GLW) (48). However, majority of cattle herds are managed in mixed livestock systems. These systems are defined by cattle consuming anywhere between 10% to 90% of their feeds as crop products or residues. Therefore, the GLW cannot offer a reliable estimate of grazing intensity because multiplying the stocking density by the livestock feed requirements and dividing by annual forage growth would necessarily result in an overestimate of the grazing intensity. If improved and sufficiently disaggregated geospatial estimates of grazing intensity become available, those estimates may be readily incorporated into this analysis.
For “improved FCRs”, we calculated a practical FCR yield ceiling as the area-weighted mean of grazed beef FCRs from all OECD countries (202 kg pasture per kg beef protein). Our “improved FCR” scenario simplistically assumes that all countries achieve a 90% yield gap closure between present-day FCRs and this FCR ceiling, and again, does not account for additional efficiencies from supplemental feed. We also include a “modestly improved FCR” where we calculate the degree of yield gap closure required to produce the equivalent amount of beef to the quantity removed from high-COI opportunity areas, by intensifying pasture-raised beef FCRs in all areas (potential forests and grasslands). In the future, more detailed economic and technological analyses should be used to project a realistic range of yield-gap closure scenarios. In lieu of these data, our high-yield scenario is intended to reflect an intuitive research question “what if all countries produced beef using their pastures with a similar efficiency?” Additionally, neither scenario can resolve subtle but consequential differences in pasture digestibility among countries, for which fine-scaled data to not currently exist. Despite these shortcomings, the “improved FCR” scenario is useful for gauging differences in pasture-raised beef production, and its carbon opportunity intensities, across regions if the presently large discrepancies in pasture use efficiency across regions were to become smaller.
Supplementary Material
Appendix 01 (PDF)
Acknowledgments
J.P. and M.K. acknowledge the support of the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (SOS.aquaterra; Grant agreement No. 819202). M.K. acknowledges the support of the Research Council of Finland’s Flagship Programme under project Digital Waters (359248). K.R.S. acknowledges the support of the Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning Grant No. 2019-00403.
Author contributions
M.N.H., J.P., and K.C. designed research; M.N.H., J.P., M.K., and K.R.S. performed research; M.N.H., J.P., and K.R.S. contributed analytic tools; M.N.H., J.P., M.K., K.R.S., and S.C.M. analyzed data; and M.N.H., J.P., M.K., K.R.S., S.C.M., and K.C. wrote the paper.
Competing interests
The authors declare no competing interest.
Footnotes
This article is a PNAS Direct Submission.
Although PNAS asks authors to adhere to United Nations naming conventions for maps (https://www.un.org/geospatial/mapsgeo), our policy is to publish maps as provided by the authors.
Data, Materials, and Software Availability
Geotiffs and R code data have been deposited in Zenodo (49).
Supporting Information
References
- 1.Ramankutty N., Evan A. T., Monfreda C., Foley J. A., Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Global Biogeochem. Cycles 22, 1–19 (2008). [Google Scholar]
- 2.Searchinger T. D., Wirsenius S., Beringer T., Dumas P., Assessing the efficiency of changes in land use for mitigating climate change. Nature 564, 249–253 (2018). [DOI] [PubMed] [Google Scholar]
- 3.Hayek M. N., Harwatt H., Ripple W. J., Mueller N. D., The carbon opportunity cost of animal-sourced food production on land. Nat. Sustain. 4, 21–24 (2021). [Google Scholar]
- 4.Griscom B. W., et al. , Natural climate solutions. Proc. Natl. Acad. Sci. U.S.A. 114, 11645–11650 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dickson B., et al. , Becoming #GenerationRestoration: Ecosystem restoration for people, nature and climate (United Nations Environmental Programme, 2021).
- 6.Sewell A., Van Der Esch S., Löwenhardt H., Goals and Commitments for the Restoration Decade: A global overview of countries’ restoration commitments under the Rio Conventions and other pledges (PBL Netherlands Environmental Assessment Agency, 2020), pp. 1–37.
- 7.Erb K. H., et al. , Unexpectedly large impact of forest management and grazing on global vegetation biomass. Nature 553, 73–76 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sun Z., et al. , Dietary change in high-income nations alone can lead to substantial double climate dividend. Nat. Food 3, 29–37 (2022). [DOI] [PubMed] [Google Scholar]
- 9.Goldewijk K. K., Beusen A., Doelman J., Stehfest E., Anthropogenic land use estimates for the Holocene - HYDE 3.2. Earth Syst. Sci. Data 9, 927–953 (2017). [Google Scholar]
- 10.Piipponen J., et al. , Global trends in grassland carrying capacity and relative stocking density of livestock. Glob. Chang. Biol. 28, 3902–3919 (2022). 10.1111/gcb.16174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Food and Agriculture Organization, The future of food and agriculture – Alternative pathways to 2050, (Food and Agriculture Organization of the United Nations, 2018), pp. 224.
- 12.Gerber P. J., et al. , Tackling Climate Change Through Livestock–A Global Assessment of Emissions and Mitigation Opportunities (Food and Agriculture Organization of the United Nations (FAO), 2013). [Google Scholar]
- 13.Hayek M. N., The infectious disease trap of animal agriculture. Sci. Adv. 8, 1–9 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Friedlingstein P., et al. , Global carbon budget 2023. Earth Syst. Sci. Data 15, 5301–5369 (2023). [Google Scholar]
- 15.Jiang Z. Y., et al. , Light grazing facilitates carbon accumulation in subsoil in Chinese grasslands: A meta-analysis. Glob. Chang. Biol. 26, 7186–7197 (2020). [DOI] [PubMed] [Google Scholar]
- 16.Byrnes R. C., Eastburn D. J., Tate K. W., Roche L. M., A global meta-analysis of grazing impacts on soil health indicators. J. Environ. Qual. 47, 758–765 (2018). [DOI] [PubMed] [Google Scholar]
- 17.Ren S., et al. , Historical impacts of grazing on carbon stocks and climate mitigation opportunities. Nat. Clim. Chang. 14, 380–386 (2024). [Google Scholar]
- 18.Beillouin D., et al. , A global meta-analysis of soil organic carbon in the Anthropocene. Nat. Commun. 14, 1–10 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Georgiou K., et al. , Global stocks and capacity of mineral-associated soil organic carbon. Nat. Commun. 13, 3797 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Post W. M., Emanuel W. R., Zinke P. J., Stangenberger A. G., Soil carbon pools and world life zones. Nature 298, 156–159 (1982). [Google Scholar]
- 21.Pan Y., Birdsey R. A., Phillips O. L., Jackson R. B., The structure, distribution, and biomass of the world’s forests. Annu. Rev. Ecol. Evol. Syst. 44, 593–622 (2013). [Google Scholar]
- 22.Herrero M., et al. , Biomass use, production, feed efficiencies, and greenhouse gas emissions from global livestock systems. Proc. Natl. Acad. Sci. U.S.A. 110, 20888–20893 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bastin J.-F., et al. , The global tree restoration potential. Science 365, 76–79 (2019). [DOI] [PubMed] [Google Scholar]
- 24.West P. C., et al. , Trading carbon for food: Global comparison of carbon stocks vs. crop yields on agricultural land. Proc. Natl. Acad. Sci. U.S.A. 107, 19645–19648 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Strassburg B. B. N., et al. , Global priority areas for ecosystem restoration. Nature 586, 724–729 (2020). [DOI] [PubMed] [Google Scholar]
- 26.Castonguay A. C., et al. , Navigating sustainability trade-offs in global beef production. Nat. Sustain. 6, 284–294 (2023). [Google Scholar]
- 27.Ceddia M. G., Bardsley N. O., Gomez-Y-Paloma S., Sedlacek S., Governance, agricultural intensification, and land sparing in tropical South America. Proc. Natl. Acad. Sci. U.S.A. 111, 7242–7247 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Garrett R. D., et al. , Intensification in agriculture-forest frontiers: Land use responses to development and conservation policies in Brazil. Glob. Environ. Chang. 53, 233–243 (2018). [Google Scholar]
- 29.Hertel T. W., Ramankutty N., Baldos U. L. C., Global market integration increases likelihood that a future African Green Revolution could increase crop land use and CO2emissions. Proc. Natl. Acad. Sci. U.S.A. 111, 13799–13804 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kreidenweis U., et al. , Pasture intensification is insufficient to relieve pressure on conservation priority areas in open agricultural markets. Glob. Chang Biol. 24, 3199–3213 (2018). 10.1111/gcb.14272. [DOI] [PubMed] [Google Scholar]
- 31.Villoria N., Garrett R., Gollnow F., Carlson K., Leakage does not fully offset soy supply-chain efforts to reduce deforestation in Brazil. Nat. Commun. 13, 5476 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Intergovernmental Panel on Climate Change, “Special Report on Climate Change and Land” (Technical Summary, Cambridge University Press, 2022).
- 33.Carlson K. M., et al. , Greenhouse gas emissions intensity of global croplands. Nat. Clim. Chang. 7, 63–68 (2017). [Google Scholar]
- 34.Amatulli G., McInerney D., Sethi T., Strobl P., Domisch S., Geomorpho90m, empirical evaluation and accuracy assessment of global high-resolution geomorphometric layers. Sci. Data 7, 1–18 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Abatzoglou J. T., Dobrowski S. Z., Parks S. A., Hegewisch K. C., TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 5, 1–12 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sun Y., et al. , Global patterns and climatic drivers of above- and belowground net primary productivity in grasslands. Sci. China Life Sci. 64, 739–751 (2021). [DOI] [PubMed] [Google Scholar]
- 37.Sulla-Menashe D., Gray J. M., Abercrombie S. P., Friedl M. A., Hierarchical mapping of annual global land cover 2001 to present: The MODIS Collection 6 Land Cover product. Remote Sens. Environ. 222, 183–194 (2019). [Google Scholar]
- 38.Klein Goldewijk K., Beusen A., Van Drecht G., De Vos M., The HYDE 3.1 spatially explicit database of human-induced global land-use change over the past 12,000 years. Global Ecol. Biogeogr. 20, 73–86 (2011). [Google Scholar]
- 39.Food and Agriculture Organization, Global Soil Organic Carbon Sequestration Potential Map – GSOCseq v.1.1 (Food and Agriculture Organization of the United Nations, 2022), pp. 1–150.
- 40.Georgiou K., et al. , Global stocks and capacity of mineral-associated soil organic carbon. Nat. Commun. 13, 3797 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Anderson-Teixeira K. J., et al. , Climate-regulation services of natural and agricultural ecoregions of the Americas. Nat. Clim. Chang. 2, 177–181 (2012). [Google Scholar]
- 42.Bajželj B., et al. , Importance of food-demand management for climate mitigation. Nat. Clim. Chang. 4, 924–929 (2014). [Google Scholar]
- 43.Poore J., Nemecek T., Reducing food’s environmental impacts through producers and consumers. Science 360, 987–992 (2018). [DOI] [PubMed] [Google Scholar]
- 44.Resare Sahlin K., et al. , An exploration of biodiversity limits to grazing ruminant milk and meat production. Nat. Sustain. 7, 1160–1170 (2024). 10.1038/s41893-024-01398-4. [DOI] [Google Scholar]
- 45.Robinson T., et al. , Global livestock production systems (Food and Agriculture Organization of the United Nations, 2011), pp. 152.
- 46.Hayek M. N., Garrett R. D., Nationwide shift to grass-fed beef requires larger cattle population. Environ. Res. Letter 13, 084005 (2018). [Google Scholar]
- 47.Food and Agriculture Organization, “Results” in Global Livestock Environmental Assessment Model (GLEAM) 2.0. (Food and Agriculture Organization of the United Nations, 2011), pp. 10.
- 48.Gilbert M., et al. , Global distribution data for cattle, buffaloes, horses, sheep, goats, pigs, chickens and ducks in 2010. Sci. Data 5, 1–11 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hayek M. N., Piipponen J., Carbon opportunity areas in global beef pastures (1.0). Zenodo. 10.5281/zenodo.12688280. Deposited 1 September 2024. [DOI]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix 01 (PDF)
Data Availability Statement
Geotiffs and R code data have been deposited in Zenodo (49).