

Abstract
Cell elongation is a fundamental component of the bacterial cell cycle and has been studied over many decades, in part owing to its mechanisms being a target of numerous antibiotic classes. While several distinct modes of cell elongation have been described, these studies have largely relied on a handful of model bacterial species. Therefore, we have a limited view of the diversity of cell elongation approaches that are employed by bacteria, and how these vary in response to evolutionary and environmental influences. Here, by employing fluorescent D-amino acids (FDAAs) to track the spatiotemporal dynamics of elongation, we reveal previously unsuspected diversity of elongation modes among closely related species of the Caulobacteraceae, with species-specific combinations of dispersed, midcell and polar elongation that can be either unidirectional or bidirectional. Using genetic, cell biology, and phylogenetic approaches, we demonstrate that evolution of unidirectional-midcell elongation is accompanied by changes in the localization pattern of the peptidoglycan synthase PBP2 and infer that elongation complexes display a high degree of phenotypic plasticity, both among the Caulobacteraceae and more widely among the Alphaproteobacteria. Demonstration that even closely related bacterial species employ highly distinct mechanisms of cell elongation reshapes our understanding of the evolution and regulation of bacterial cell growth, with broad implications for bacterial morphology, adaptation, and antibiotic resistance.
Keywords: Elongation modes, midcell elongation, Peptidoglycan, Caulobacteraceae, Caulobacter
INTRODUCTION
Cell elongation is a fundamental process in bacteria, underlying cell growth, morphogenesis and division. Mechanisms of cell elongation are strictly regulated but respond dynamically to developmental and environmental cues1–3. For instance, spatiotemporal patterns of cell elongation are modulated over the cell cycle in many bacterial species. In others, nutrient starvation can trigger specialized modes of elongation, resulting in shape changes to improve nutrient uptake. Over the course of evolution, mechanisms of cell elongation have also diversified, as evidenced by the variety of cell shapes, life cycles and mechanisms of growth seen in bacteria4,5.
At the structural level, bacterial cell elongation is driven by the synthesis of the peptidoglycan (PG) cell wall, a meshwork of glycan strands crosslinked by peptide chains that confers structural integrity to the cell3,6,7. Synthesis of PG is essential to bacterial growth and division8 and is catalyzed by the penicillin binding proteins (PBPs) acting in concert with regulatory protein complexes called the elongasome (for cell elongation) and the divisome (for cell division). Although the PBPs, the elongasome and the divisome are widely conserved among bacteria, different species have been shown to employ distinct sets of proteins to assemble their elongasomes9–14. Such modularity and flexibility in the regulation of cell elongation at the molecular level may underlie the evolution and diversification of bacterial cell elongation mechanisms. Indeed, recent studies have found that disparate mechanisms of cell elongation can yield similar shapes: a rod-shaped cell can be generated by a dispersed, a lateral elongation mode, or by zonal elongation from one or both cell poles or the midcell (Fig. 1a)9,15–19. Conversely, the same elongation mode can generate different cell shapes: spherical, ovoid, rod, and crescent shapes can be generated by bidirectional elongation from the midcell20,21. However, our understanding of the diversity of elongation modes in bacteria is primarily based on a few distantly related model organisms, leading to the assumption that closely related species share the same elongation strategies. Consequently, the true diversity of bacterial cell elongation mechanisms may be greatly underappreciated, and our insights reveal little about the evolutionary mechanisms driving these differences.
Fig. 1: Diversity of growth modes in rod-shaped bacteria and members of the Caulobacteraceae family.

(a) Schematic illustrating different growth modes observed among rod-shaped bacteria. Green lines represent lateral or dispersed elongation, solid green denotes different modes of localized cell elongation, and solid blue marks cell division.
(b) Dimorphic cell cycles and growth modes in C. crescentus, A. excentricus and A. biprosthecum. At the beginning of their cell cycle, C. crescentus swarmer cells elongate by dispersed insertion of new PG material (green lines) before bidirectional elongation near the midcell (green), followed by cell division (blue). In this study, we show that A. excentricus swarmer cells also undergo dispersed elongation (green lines) during the swarmer stage. However, this is followed by a novel, unidirectional-midcell elongation towards the new pole (green) prior to bidirectional cell division (blue). Finally, we identified that A. biprosthecum cells elongate through yet another novel mode of elongation – a combination of polar and unidirectional-midcell elongation.
(c) Schematic depicting the pulse-chase experiment using FDAA. Whole-cell PG was labeled with 500 µm FDAA (the “pulse,” colored in green) over two generations, followed by washes with PYE to remove free FDAA from the medium. Subsequent growth in the absence of FDAA (the “chase”) was observed using time-lapse microscopy. During the chase period, the loss of FDAA signal corresponds to new PG synthesis/turnover. The red dashed line indicates the position of the ZapA fluorescent protein fusion, as a marker of the future division site.
(d) Pulse-chase experiments using FDAAs in C. crescentus and A. excentricus cells carrying fluorescent fusions of the cell division protein ZapA. Images were taken every 5 minutes during the chase period. Kymographs show the loss of FDAA fluorescence as the cells grow. In C. crescentus (left), the FDAA BADA was used in combination with ZapA-mCherry. In A. excentricus (right), the FDAA TADA was used in combination with ZapA-sfGFP. Kymographs from both these species present the FDAA signal in green and the ZapA fluorescent fusion signal in red. White stars indicate cells starting division. See Extended Data Fig.1 for additional kymographs from each species.
(e) Schematic of FDAA signal loss in C. crescentus and A. excentricus cells. The red dashed line indicates the position of ZapA as a marker of the future division site. Green shading represents the old PG labeled with FDAA.
To address this knowledge gap, our study explores how distinct elongation modes evolved within closely related, morphologically diverse bacterial species from the Caulobacteraceae family4,5,22. Within this family, the well-studied model organism Caulobacter crescentus uses bidirectional elongation from the midcell23 (Fig. 1b). By studying its related species, we discover two novel elongation modes – unidirectional midcell elongation in the species Asticcacaulis excentricus, and polar plus unidirectional midcell elongation in Asticcacaulis biprosthecum. Using a multidisciplinary approach that integrates live-cell imaging with genetic and evolutionary analysis, we explore these elongation mechanisms. Our findings reveal that the evolution of these elongation strategies is associated with shifts in the spatial localization of a core elongasome enzyme. Specifically, we show that the penicillin binding protein PBP2, which localizes diffusely in C. crescentus, concentrates at the midcell in A. excentricus, where it serves as the transpeptidase enzyme driving unidirectional PG synthesis. This highlights how the regulation of conserved elongasome machinery can vary even between closely related species, potentially aligning with broader evolutionary pressures. Extending our analysis beyond the Caulobacteraceae, we found that Rhodobacter capsulatus shares the unidirectional midcell elongation pattern seen in A. excentricus, and we further infer from the phylogeny of the Alphaproteobacteria that bacterial cell elongation mechanisms display far greater phenotypic plasticity than previously anticipated. Ultimately, these results challenge the notion that closely related species share the same elongation mode. Instead, they reveal an unexplored diversity in elongation mechanisms, as cells respond to evolutionary forces to generate diversity in growth modes even at close evolutionary scales.
RESULTS
C. crescentus and A. excentricus have different patterns of PG synthesis
The order Caulobacterales exhibits at least three elongation modes – dispersed, polar and midcell4, suggesting that it could be a good model for studying the evolution of cell elongation. The order comprises three families: the Caulobacteraceae, the Hyphomonadaceae, and the Maricaulaceae. The Hyphomonadaceae elongate polarly through budding24–26, but the elongation modes in the Maricaulaceae and the Caulobacteraceae are unknown, with the exception of the model organism C. crescentus, which predominantly grows through bidirectional midcell elongation, with some dispersed elongation during early stages of the cell cycle (Fig. 1b).
To determine the elongation modes in other Caulobacteraceae, we analyzed the genus Asticcacaulis, comparing C. crescentus and A. excentricus using pulse-chase experiments with fluorescent D-amino acids (FDAAs). FDAAs are fluorescent dyes that are incorporated into the PG by PG transpeptidases, serving as highly effective tools to observe the dynamics of cell growth in various colors (in this study, we use the dyes BADA, HADA and TADA, see Methods)27,28. To track PG synthesis/turnover in real-time relative to the cell division site, we marked the division site in C. crescentus using ZapA-mCherry and in A. excentricus using ZapA-sfGFP29, and performed pulse-chase experiments using FDAAs with complementary fluorophores. Briefly, we labeled whole-cell PG with FDAA (the “pulse”) over two generations, washed the cells to remove free FDAA, and observed the cells during a period of growth in the absence of FDAA (the “chase”) by time-lapse microscopy (Fig. 1c). During the chase period, the loss of FDAA labeling reveals spatial patterns of new PG incorporation30.
In dimorphic bacteria such as Asticcacaulis and Caulobacter species, the new pole generated by division gives rise to a motile swarmer cell, while the old pole forms the larger, non-motile stalked cell, reflecting the species’ characteristic asymmetric division. In C. crescentus, visualizing the retention and loss of the FDAA label throughout the cell cycle, we observed that the loss of FDAA labeling occurred from the division plane defined by ZapA-mCherry, and extended bidirectionally towards both cell poles (Fig. 1d and Extended Data Fig. 1a). Conversely, in A. excentricus, the loss of FDAA labeling originated from the division plane but moved predominantly towards the new pole of the dividing cell (Fig. 1d and Extended Data Fig. 1b). Later in the cell cycle, A. excentricus cells also began losing FDAA signal on the old pole side of the ZapA-sfGFP signal once constriction was initiated (Fig. 1d, white stars). This loss of FDAA signal on the old pole side of the division plane during constriction likely corresponds to PG synthesis during septation and cell division. Together, these data indicate that PG synthesis/turnover for cell elongation are spatially localized close to the future site of cell division in both C. crescentus and A. excentricus. Furthermore, they indicate a variation in PG synthesis/turnover dynamics in these two closely related species, in which C. crescentus elongates bidirectionally and A. excentricus towards the new pole (Fig. 1b and 1e).
A. excentricus elongates unidirectionally from the midcell towards the new pole
To further investigate the position and directionality of PG synthesis during midcell elongation in A. excentricus, we conducted sequential short-pulse FDAA labeling experiments with differently colored FDAAs. This approach allows us to track active sites of PG synthesis in growing cells based on the spatial pattern of incorporation of the sequentially applied dyes over time30. If the signal from the first FDAA (red) pulse appears on both sides of the second FDAA (green), PG incorporation can be inferred to occur bidirectionally, whereas if the first FDAA signal appears on one side of the second signal, it would indicate unidirectional elongation (Fig. 2a).
Fig. 2. Sequential FDAA labeling reveals that C. crescentus grows bidirectionally from the midcell, while A. excentricus grows unidirectionally from the midcell towards the new pole.
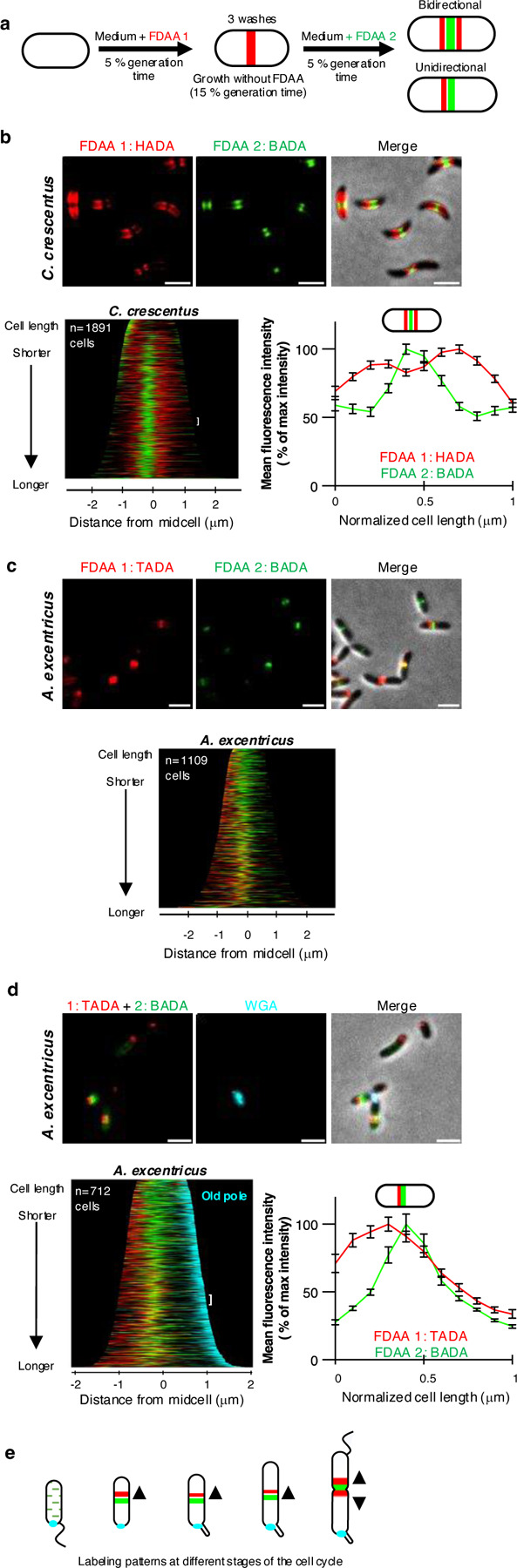
(a) Schematic depicting the dual short-pulse experiment: sequential, dual short FDAA pulses show sites of active PG synthesis. Cells were first labeled with one FDAA (HADA in C. crescentus or TADA in A. excentricus, red) for 5% of their generation time, washed with PYE to remove the free FDAA, allowed to grow for 15 % of their generation time and then labeled with a second FDAA (green) for 5% of their generation time, washed again, and imaged with phase and fluorescence microscopy.
(b-d) Sequential FDAA labeling in C. crescentus (b) and A. excentricus (c-d). Top: Representative images are shown. Left to right: FDAA 1, FDAA 2 and merge images. Scale bars: 2 µm. Middle: population-level demographs showing the localization of the fluorescence intensities of FDAA 1 and FDAA 2. Bottom: fluorescence intensity plots showing the fluorescence signals from both FDAAs. In Panel d, the old pole in A. excentricus cells is additionally labeled with fluorescent WGA (cyan). In all demographs, cells are arranged by length, with 50% of maximum fluorescence intensities shown. Demographs in Panels b-c are oriented with the maximum fluorescence intensity of the second FDAA to the left, and in Panel d with the WGA-labeled old pole (cyan) to the right. In Panels b and d, the white brackets show the 50 cells selected to plot the fluorescence intensities of the two FDAAs along the cell length, shown in the graphs adjacent to the demographs. To generate these graphs, points were selected along the medial axis of each cell, and the normalized signals of FDAA 1 (red) and FDAA 2 (green) were plotted relative to their normalized position along the cell length. The lines represent the mean values, with error bars showing the standard error of the mean (SEM).
See Extended Data Fig. 2 for additional demographs showing each FDAA individually, and at 50% and 100% fluorescence intensities.
(e) Schematic illustrating PG synthesis at different stages of the A. excentricus cell cycle. Smaller swarmer cells undergo dispersed cell elongation (green dots). As they differentiate into stalked cells, they elongate unidirectionally from the midcell towards the new cell pole, with the first FDAA signal (red) located on the new pole side of the second FDAA signal (green). Predivisional cells or cells undergoing septation exhibit bidirectional growth at the midcell, with red signals on both sides of the second FDAA signal (green). The old pole is indicated by the holdfast (cyan).
For these experiments, A. excentricus and C. crescentus cells were first grown in the presence of one FDAA (shown in red) for 5% of their generation time, washed to remove excess dye, and then subjected to a second pulse with a different FDAA (shown in green) (Fig. 2a). In C. crescentus, the first, red FDAA signal appeared on both sides of the second, green signal, which was approximately located at the midcell (Fig. 2b). To analyse these patterns quantitatively, we generated demographs, which are graphical summaries of fluorescence intensities, where cells are sorted by length along the y-axis, and aligned at the center by their midcell as a proxy for cell cycle progression. Demograph analysis of C. crescentus cells confirmed that their first FDAA signal was located on both sides of the second signal in all but the shortest (i.e. most newly divided) cells (Fig. 2b bottom-left and Extended Data Fig. 2b). For further quantification, the fluorescence intensities of the FDAAs were normalized and plotted against their relative positions along the cell length for a subset of cells, from roughly the middle of their cell cycles. As in the demograph, the red signal appeared on both sides of the green signal, confirming bidirectional midcell elongation in C. crescentus (Fig. 2b bottom-right).
Contrastingly, in A. excentricus, the first, red FDAA signal appeared only on one side of the midcell region, whereas the second, green signal appeared at the midcell region, demonstrating unidirectional PG synthesis (Fig. 2c and Extended Data Fig. 2b). To determine whether the unidirectional midcell elongation observed in A. excentricus consistently proceeds towards the new pole, we conducted dual short-pulse FDAA labeling experiments in A. excentricus cells in which the old poles were marked. For this, we utilized the dimorphic cell cycle of this species, which features regulated changes in morphology and surface adhesion within the context of the cell cycle31,32. Specifically, cells produce a holdfast polysaccharide adhesin at the old pole during cell cycle progression, which can be used as an old-pole marker through labeling with fluorescent wheat germ agglutinin (WGA) lectin. Dual short-pulse FDAA labeling in A. excentricus cells with WGA-labeled holdfasts showed that the first FDAA signal was always on the new-pole side of the second FDAA signal (Fig. 2d). These results demonstrate that C. crescentus has a bidirectional midcell elongation mode while A. excentricus has a unidirectional midcell elongation mode in which PG synthesis proceeds towards the new pole (Fig. 2e).
The A. excentricus class B PG synthase PBP2 localizes to sites of new PG synthesis
To understand the evolutionary origins of unidirectional midcell elongation in A. excentricus, we investigated the potential molecular determinants driving this novel elongation mode. Bioinformatic analysis using C. crescentus PBP2 and PBP1a as queries revealed a limited repertoire of PG synthases in A. excentricus: three class A PBPs, including two homologues of PBP1a (Astex_2378 and Astex_2994) and one homologue of PBP1c (Astex_0196); and two monofunctional class B PBPs – PBP2 and PBP3 (FtsI) (Extended Data Fig. 3a). This limited repertoire of PBPs in A. excentricus, and the conserved neighborhood architecture of the cell elongation loci in C. crescentus and A. excentricus (Extended Data Fig. 3b) suggested that the same canonical elongasome proteins may have evolved to generate different modes of elongation in the two species. Therefore, we hypothesized that localization of PBP2, an essential component of the elongasome, may play a role in the evolution of these different elongation modes.
To investigate the role of PBP2 in midcell elongation, we utilized fluorescent protein fusions to track its subcellular localization. We fused PBP2 to mCherry at its native locus in A. excentricus and performed fluorescence microscopy (See Extended Text for validation of the mCherry-PBP2 fusion). Across the population, PBP2 exhibited a patchy localization with enrichment near the midcell (Fig. 3a). Quantifying its localization in a large number of cells in a demograph, or using a population-wide heatmap of subcellular PBP2 localization using the holdfast as a polar marker (Fig. 3a), we found that PBP2 accumulates close to the midcell offset towards the new pole, which is similar to the location of new PG synthesis. To further analyze whether PG synthesis at the midcell coincides with PBP2 localization, we labeled A. excentricus cells expressing mCherry-PBP2 with a short pulse of FDAA. Fluorescence microscopy and quantification using a population-wide heatmap and density map of the maximal fluorescence intensities of the mCherry-PBP2 and FDAA signals showed that the FDAA signal overlapped with the mCherry-PBP2 signal (Fig. 3b and Extended Data Fig. 4a–b). Together, these data indicate that PBP2 localization correlates with sites of PG synthesis during unidirectional midcell elongation in A. excentricus.
Fig 3. Distinct PBP2 localization in A. excentricus and C. crescentus and its correlation with active PG synthesis.
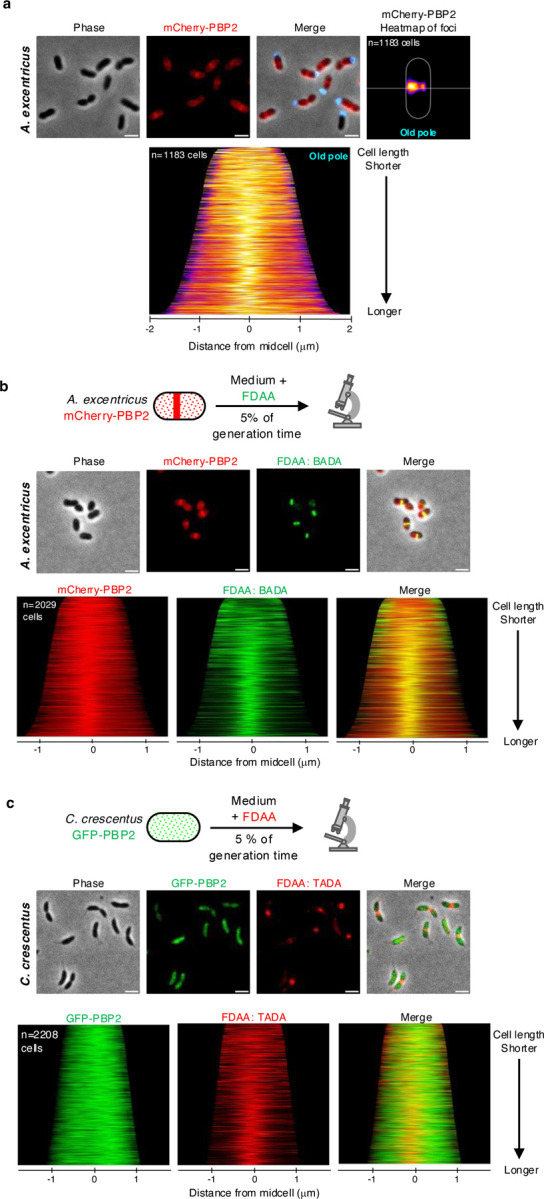
(a) Subcellular localization of mCherry-PBP2 in A. excentricus. Top: Representative images are shown. Left to right: Phase, mCherry-PBP2 fluorescence and merged images. Scale bar: 2 µm. A heatmap of mCherry-PBP2 foci at the population level is displayed. In the heatmap, cells were oriented using the old pole labeled with WGA (cyan), with the white line indicating the midcell. Bottom: A demograph showing the localization of mCherry-PBP2 fluorescence at the population level, with cells arranged by length and oriented with the old pole towards the right.
(b) Short-pulse FDAA (BADA) labeling of the A. excentricus mCherry-PBP2 strain. Top: A schematic depicting the experiment. Cells were labeled with 250 µM BADA for 5% of their generation time, fixed with 70% (v/v) ethanol and imaged. Middle: Representative phase, fluorescence (mCherry-PBP2 and BADA) and merged images are shown. Scale bar: 2 µm. Bottom: Population-level demographs showing the localization of the fluorescence intensities of mCherry-PBP2, BADA and their overlays. See Extended Data Fig. 4a for heatmaps and density maps of mCherry-PBP2 and BADA at the population level.
(c)Short-pulse FDAA (TADA) labeling of the C. crescentus GFP-PBP2 strain. Top: A schematic depicting the experiment. Cells were labeled with 250 µM TADA for 5% of their generation time, fixed with 70% (v/v) ethanol and imaged. Middle: Representative phase, fluorescence (GFP-PBP2 and TADA) and merged images are shown. Scale bar: 2 µm. Bottom: Population-level demographs showing the localization of the fluorescence intensities of GFP-PBP2, TADA and their overlays.
See Extended Data Fig. 4b–c for additional fluorescence images and population-level demographs showing the differences in localization of PBP2 in A. excentricus vs. C. crescentus.
To further probe the link of PBP2’s distinct localization in A. excentricus to its novel mode of cell elongation, we investigated PBP2 localization in C. crescentus, which exhibits bidirectional elongation from the midcell23. Using a GFP fusion to PBP2 at its native locus in C. crescentus, we observed that GFP-PBP2 displayed a dispersed distribution throughout the cell cycle in C. crescentus, consistent with previous studies33, and distinct from PBP2 localization in A. excentricus (Fig. 3c and Extended Data Fig. 4c–d). To determine whether PBP2 colocalizes with sites of PG synthesis in C. crescentus, we performed short-pulse FDAA labeling in cells expressing GFP-PBP2. Fluorescence microscopy and population-level quantification of cells in a demograph revealed that, unlike in A. excentricus, C. crescentus PBP2 did not show significant enrichment at the midcell, and the FDAA labeling did not overlap with the GFP-PBP2 signal (Fig. 3c). This difference in the localization patterns of PBP2 in C. crescentus and A. excentricus suggests that the relocalization of PBP2 may be a critical evolutionary step in the divergence of the elongation modes between these two species.
PBP2 activity is required for unidirectional elongation at the midcell in A. excentricus
To understand the functional role of the essential protein PBP2 in the unidirectional midcell elongation of A. excentricus, we utilized the β-lactam antibiotic mecillinam, a specific inhibitor of PBP2 transpeptidase activity in E. coli34. In C. crescentus, mecillinam causes cell bulging35,36, likely through its inhibition of PBP2 activity (Extended Data Fig. 5a). To determine the target of mecillinam in A. excentricus, we conducted competition assays with the β-lactam Bocillin FL, a fluorescent penicillin that covalently binds all PBPs. Using Bocillin gel analysis, we found that mecillinam predominantly inhibited binding of Bocillin to PBP2, even at high concentrations (100 µg ml−1), while other PBPs in A. excentricus were only partially affected (Extended Data Fig. 5b). These results indicate that mecillinam is a suitable tool to investigate the role of PBP2 in cell elongation in A. excentricus.
To analyze the effect of PBP2 inhibition on cell elongation, we conducted a FDAA pulse-chase experiment in A. excentricus cells expressing ZapA-sfGFP, with or without mecillinam. We labeled whole-cell PG with FDAA over two generations without mecillinam, followed by a wash to remove excess FDAA, and a chase period with or without mecillinam for 120 min. In untreated cells, we found robust FDAA signal loss on the new pole side of the ZapA-sfGFP signal, consistent with unidirectional elongation at the midcell (Fig. 4a, left). In contrast, FDAA signal loss in mecillinam-treated cells was observed in bulges on the new pole side of the cell (Fig. 4a, right). To quantify the FDAA signal loss and cell bulging upon mecillinam treatment, we measured the subcellular localization of these bulges in relation to the fluorescence signals of ZapA and FDAA in a population-wide analysis. We plotted these features relative to the cell center, generating ShapePlots37 for the whole population (Fig. 4b, right panels), as well as for four categories binned by cell length. Bulging was detected consistently on the new pole side of the midcell across all four categories, with its position shifting closer to midcell as cells progressed through the cell cycle and approached division (Fig. 4b, left panels). These analyses confirmed that bulging occurred primarily on the new pole side of the midcell in A. excentricus, i.e. at the site of midcell elongation (Fig. 4b, top panels). Furthermore, loss of FDAA signal peaked at this site, suggesting that the bulging may be a result of abnormal PG synthesis at the midcell when PBP2 is inhibited (Fig. 4b, bottom panels). Together, these results suggest that PBP2 activity is required for the regulation of elongasome activity at the midcell – both to promote unidirectional PG synthesis towards the new pole, and to prevent bulging.
Fig 4. PBP2 is responsible for unidirectional PG synthesis at the midcell in A. excentricus.

(a) Top: Schematic depicting pulse-chase experiments in A. excentricus ZapA-sfGFP cells with (right) or without (left) mecillinam treatment. Whole-cell PG was labeled with TADA (red) over two generations, followed by washing and growth with or without mecillinam (50 µg ml−1) over one generation before imaging. Bottom: Representative phase, fluorescence and merged images from both conditions are shown. The fluorescence images show TADA individually as well as overlaid with ZapA-sfGFP. The merged image shows phase overlaid with TADA, ZapA-sfGFP and WGA fluorescence. Scale bar: 2 µm.
(b) ShapePlots of A. excentricus cells after an FDAA pulse-chase experiment, with mecillinam treatment during the chase period. Top: Shape plots showing bulge localization. Bottom: ShapePlots show ZapA-sfGFP and FDAA signal loss following the chase period. Each ShapePlot is divided longitudinally (black line) to show the loss of FDAA signal exclusively on the left, and overlaid with ZapA-sfGFP on the right.
ShapePlots are presented individually for four categories of cells binned by cell length (left) as well as for the entire population of 160 cells (right). All cells are oriented using WGA-labeling of the holdfast, positioning the old pole at the bottom. Horizontal white lines represent the midcell.
(c) Top: Schematic of a dual short-pulse experiment with or without mecillinam (50 µg ml−1) to show active PG synthesis. A. excentricus cells were first allowed to grow over 120 min with or without mecillinam. Cells were then labeled with TADA (red) for 5% of their generation time, washed to remove excess FDAA, allowed to grow for 18 min, and then labeled with BADA (green) for 5% of their generation time, with or without mecillinam. They were then washed again and imaged with phase and fluorescence microscopy. Middle: Representative phase and merged images from both treatment conditions – with and without mecillinam. Merged images show phase contrast overlaid with fluorescence signals from the two FDAAs as well as WGA-labeled holdfast (cyan). Scale bar: 2 µm. Look up tables (LUTs) were adjusted for each condition to have a visible FDAA signal, and therefore were not identical. Bottom: Population-level demographs showing the fluorescence intensities of both FDAA signals, with and without mecillinam treatment. Cells were arranged by length, with the old pole to the right. 50% of the maximum fluorescence intensities are shown.
(d) Box graphs showing quantification of fluorescence intensities for each FDAA pulse in the presence or absence of mecillinam (for the same cells quantified in the demographs in 4c). **** P<0.0001 (Welch One-Way ANOVA Test). Error bars show the standard error of the mean (SEM).
To further analyze PBP2’s role in the directionality of midcell elongation in A. excentricus, we conducted dual short-pulse experiments using two differently colored FDAAs in the presence or absence of mecillinam (Fig. 4c). In untreated cells, sequential FDAA labeling showed the first, red signal on only one side of the second, green signal, consistent with midcell elongation towards the new pole (Fig. 4c, left panels). However, in mecillinam-treated cells, both FDAA signals were distributed diffusely at the site of bulging, confirming the loss of directionality in PG synthesis upon PBP2 inhibition (Fig. 4c, right panels). Quantification of mean fluorescence intensities showed reduced FDAA incorporation in cells treated with mecillinam (Fig. 4d), consistent with the main transpeptidase of the cell being inhibited. Overall, these observations demonstrate that the inhibition of PBP2 activity severely disrupts the coordination of PG synthesis, leading to bulging at the site of the elongasome, and loss of the novel unidirectional midcell elongation mode of A. excentricus. These findings confirm our hypothesis that PBP2’s localization and activity evolved together with the evolution of unidirectional midcell elongation in A. excentricus, with the enzyme serving as a key organizer of the directionality of elongasome activity in this species.
Diverse modes of midcell elongation within and beyond the Caulobacteraceae family
To determine the elongation modes in other Caulobacteraceae, we analyzed other species in the genera Asticcacaulis, Phenylobacterium, Brevundimonas, and Caulobacter using pulse-chase experiments with FDAA (Fig. 5a). To analyze the patterns of FDAA labeling loss across populations of pulse-chased cells, we generated demographs for each species (Fig. 5b). Smaller cells of all six species showed a loss of signal at one of their poles, which likely corresponds to a recent division (Fig. 5b, white stars). In longer cells, the loss of FDAA labeling during the chase period was located near the midcell, corresponding to midcell elongation at the future site of cell division, or indeed to cell division itself (Fig. 5b). In C. crescentus, C. henricii, and B. diminuta the loss of FDAA labeling was located near the midcell, progressing symmetrically towards both cell poles. This observation is consistent with PG synthesis in C. crescentus occurring bidirectionally from the midcell region in stalked cells23 (Fig. 1b–d, Fig. 2, and Fig. 5c–d). In contrast, the loss of FDAA labeling during the chase period in A. excentricus, A. biprosthecum and P. conjunctum, started in the midcell region but trended towards the left of the demograph (Fig. 5b, red arrows).
Fig. 5. Diversity of cell elongation modes in members of the Caulobacteraceae family.

(a) Schematic depicting the pulse-chase experiment using FDAAs. Whole-cell PG was labeled with TADA for two generations, washed to remove excess FDAA, followed by a chase period of 75 minutes before imaging.
(b) Demograph analysis of pulse-chase experiments in WT cells of C. crescentus, C. henricii, P. conjunctum, B. diminuta, A. excentricus, and A. biprosthecum. Loss of FDAA labeling shows sites of PG synthesis/remodeling. Cells are arranged by length, with each cell oriented so that the pole with the maximum fluorescence intensity is to the right. White stars indicate signal loss at one pole. Red arrows highlight unidirectional midcell elongation in A. excentricus, A. biprosthecum, and P. conjunctum, as observed by greater FDAA signal loss towards the left of the demograph.
(c) Phylogenetic tree of representative species from the Alphaproteobacteria, which includes the family Caulobacteraceae. Taxon label colors correspond to different modes of cell elongation: dispersed cell elongation (green), unidirectional midcell elongation (red), polar elongation (blue), budding (violet), polar and unidirectional midcell elongation (orange and red), bidirectional midcell elongation (magenta), binary fission (black), and unknown (grey). Species for which the cell elongation mode has been studied using FDAAs, TRSE or other methods are highlighted in bold. The node where pbp2 and mreB (and the associated mreCD and rodA genes) are predicted to have been lost within the Rhizobiales is highlighted in blue. The tree, based on a concatenation conserved protein-coding gene sequences, is fully supported, with posterior probabilities of 1 for all clades. See the online Methods section for details on phylogenetic reconstruction and refer to Table S3 for genome IDs and mode of cell elongation.
(d) This schematic, derived from a pruned version of the phylogenetic tree in Fig. 5a to highlight the species of interest, illustrates the identified modes of cell elongation and the possible number of transitions. The transitions are depicted assuming two scenarios: unidirectional cell elongation as the ancestral state (left, red) or bidirectional midcell elongation as the ancestral state (right, magenta). Colored lines and arrows (red or magenta) indicate where the transitions might have occurred.
The loss of FDAA predominantly towards one pole in A. biprosthecum and P. conjunctum suggests a unidirectional elongation mode in these species, similar to A. excentricus. To further investigate this, we performed pulse-chase FDAA labeling and time-lapse microscopy in A. biprosthecum (Fig. 6a). Interestingly, kymograph analysis in this species revealed not only unicellular midcell elongation towards the new pole, as in A. excentricus, but also an additional site of elongation at the new pole itself (Fig. 6b and Extended Data Fig. 6). This observation suggests that A. biprosthecum elongates through yet another novel mode – a combination of polar elongation and unidirectional midcell elongation. To further analyze this mode of cell elongation, we sequentially added differently colored FDAAs during short pulses (5% of a generation time, Fig. 6c). Demograph and fluorescence profile analyses revealed that the second, green FDAA signal was present at both the pole and the midcell, while the first, red signal was located mainly at the pole. At the pole, the second, green signal appeared at the tip, while the red signal was positioned just adjacent away from the tip, indicating apical elongation (Fig. 6d–e and Extended Data Fig. 6). Interestingly, the population-level demographs showed that the FDAA signals at the pole were present throughout the cell cycle, whereas signals at the midcell appeared only in longer cells. This pattern indicates a sequence of polar cell elongation early in the cell cycle followed by unidirectional midcell elongation closer to the time of division. The observation of both polar and unidirectional midcell elongation modes in A. biprosthecum underscores the diverse range of cell elongation mechanisms within the Caulobacteraceae family.
Fig. 6. FDAA labeling experiments demonstrate polar and unidirectional midcell elongation in A. biprosthecum and unidirectional midcell elongation in R. capsulatus.

(a) Schematic depicting the FDAA pulse-chase experiments in A. biprosthecum and R. capsulatus. Whole-cell PG was labeled with 500 µm TADA (green) over two generations, followed by washes with medium to remove free FDAA from the medium. Subsequent growth in the absence of the FDAA (the chase) was followed by time-lapse microscopy. Loss of FDAA signal during the chase period corresponds to new PG synthesis/turnover.
(b,f) Kymographs of the pulse chase experiments showing the loss of FDAA fluorescence during the chase period in (b) A. biprosthecum and (f) R. capsulatus. Images were taken every 5 minutes. See Extended Data Fig. 6 and 7 for additional kymographs.
(c) Schematic depicting the dual short-pulse experiment showing active PG synthesis. Cells were first labeled with TADA for 5% of their generation time, washed with fresh media to remove free FDAA, allowed to grow without FDAA and then labeled with BADA for 5% of their generation time. Cells were then washed again and imaged with phase and fluorescence microscopy.
(d,g) Representative images and demographs showing the fluorescence intensity of both FDAA signals in (d) A. biprosthecum and (g) R. capsulatus. In the demographs, cells were arranged by length, with the old pole (labelled with WGA, not shown) to the right in A. biprosthecum, and with the maximum fluorescence intensity of the first FDAA to the left in R. capsulatus. 50% of the maximum fluorescence intensities are shown. The white brackets indicate the 50 cells selected to show the fluorescence profiles of the two FDAAs in (e) and (f). Scale bars: 2 µm. See Extended Data Fig. 6 and 7 for demographs showing each FDAA individually and with the full range of fluorescence signal.
(e,h) Fluorescence intensity profiles of FDAA 1 (TADA, red line) and FDAA 2 (BADA, green line) in (e) A. biprosthecum cells and (h) R. capsulatus cells, plotted from n = 50 cells for both species. Points were selected along the medial axis of each cell, and the normalized signal was plotted relative to position along the cell length. The lines represent the mean values, with error bars showing the standard error of the mean (SEM).
Given the diversity of elongation modes observed even within closely related species (Fig. 5b–d), we questioned whether unidirectional midcell elongation was a rare occurrence limited to the Caulobacteraceae family or if it could be observed in other species as well. We therefore extended our investigation to the more distantly related R. capsulatus, a member of the alphaproteobacterial order Rhodobacterales. We performed pulse-chase FDAA labeling and time-lapse microscopy as before (Fig. 6a and 6f and Extended Data Fig. 7). Kymograph analysis showed that the loss of FDAA labeling in R. capsulatus originated from the division plane and predominantly moved unidirectionally towards one of the cell poles, suggesting unidirectional midcell elongation. To further investigate the R. capsulatus cell elongation mode, we performed a dual short-pulse experiment and with two differently colored FDAAs (Fig. 6c). Demograph and fluorescence profile analyses revealed that the first FDAA signal in R. capsulatus was only on one side of the second FDAA signal, towards one of the cell poles (Fig. 6g–h). These results show that R. capsulatus also exhibits unidirectional elongation from the midcell, similar to the pattern observed in A. excentricus. This finding indicates that unidirectional midcell elongation is not restricted to the Caulobacteraceae family and suggests that this growth mode may be more widespread within the Alphaproteobacteria.
Overall, the diversity of cell elongation modes identified in this study suggests that there may be more novel modes to discover among bacteria. Our findings also emphasize the need for further exploration of the evolutionary transitions, such as PBP2 relocalization, that may underlie these differences.
DISCUSSION
The existence of different elongation modes in bacteria has been known for decades38, but the mechanisms behind their evolution have remained unknown, largely due to research bias towards a few, distantly related model organisms. In parallel, there has been an assumption that closely related species share similar elongation mechanisms, hampering the study of divergence at shorter evolutionary scales. In this study, we challenge this assumption by revealing significant differences in the elongation strategies of closely related species within the Caulobacteraceae family. Our findings indicate that evolutionary changes in cell elongation can occur more frequently and at shorter evolutionary timescales than previously thought, opening new avenues for research into the evolution of bacterial growth and morphogenesis.
Among the Caulobacteraceae, the diversity and novelty of elongation modes we observe - ranging from bidirectional midcell elongation in C. crescentus, C. henricii and B. diminuta, to the newly described unidirectional midcell elongation in A. excentricus and P. conjunctum as well as the combination of polar and unidirectional midcell elongation in A. biprosthecum (Fig. 1b, Fig. 5 and Fig. 6), indicate remarkable evolutionary flexibility. They suggest a capacity for closely related species to adapt their elongation mechanisms, possibly in response to evolutionary pressures such as nutrient availability, morphological constraints, competition, predation, etc. It is already known that bacteria can regulate their elongation patterns in response to environmental changes. For instance, Streptomyces cells can shift their elongation mechanisms in response to metabolic cues, allowing exploratory growth39,40. Meanwhile, Salmonella typhimurium encodes two differentially regulated elongasomes, one of which is specialized for pathogenesis, functioning under acidic, intracellular conditions, with slower PG synthesis that may help coordinate cell elongation with host metabolism (although the spatial pattern of elongation remains the same)41. Many other bacterial species suppress the highly conserved divisome, using only elongation to produce a filamentous morphology to overcome a range of environmental constraints42–44. Finally, within the Caulobacteraceae, stalk elongation is a well-known response to nutrient limitation, although this specialized mode of PG synthesis at the stalk site is independent of the core elongasome that drives vegetative growth in the cell31,45,46. It would be intriguing to explore whether A. excentricus and related species similarly adjust their core elongation modes in response to environmental conditions. Future studies on this topic may shed light on the selective pressures that have led to the divergent elongation modes among the Caulobacteraceae.
The plasticity of cell elongation strategies is evident not only under different environmental conditions, but even within a single cell under different regulatory states. For instance, among the Caulobacteraceae, the transition from dispersed elongation in the swarmer cell to localized cell elongation at the midcell (Fig. 1) demonstrates how elongasome components may be modulated by the cell cycle. Even in E. coli, which predominantly grows through dispersed elongation along its lateral walls, the elongasome-specific PG synthase PBP2 shows a cell-cycle dependent enrichment at the midcell prior to division, coinciding with a burst of midcell elongation47. Gram-positive coccoid bacteria such as S. aureus offer yet another example of the flexibility of the PG synthesis machinery, which functions both peripherally and for septal sidewall synthesis in these bacteria48. These observations underscore that elongation modes are not static in cells, but rather dynamically adaptable through regulatory networks controlling cell growth. Evolutionary forces may drive changes in bacterial elongation by modulating existing molecular mechanisms rather than by introducing new ones.
In this study, we provide evidence of novel cell elongation modes beyond the Caulobacteraceae (Fig. 5c). Indeed, we discovered that R. capsulatus demonstrates unidirectional midcell elongation similar to A. excentricus (Fig. 6f–h). To understand the evolutionary context of this surprising discovery, we searched the literature for previous reports of elongation modes among the Rhodobacterales and found that Rhodobacter sphaeroides demonstrates bidirectional midcell elongation49. Meanwhile, in another branch of the Alphaproteobacteria, the loss of the key elongation proteins MreB and PBP2 among members of the Rhizobiales has coincided with the evolution of polar elongation (Fig. 5c), further emphasizing the evolutionary plasticity of bacterial elongation systems9. Nonetheless, the conservation of elongasome clusters such as pbp2/rodA across species with different elongation modes such as E. coli, C. crescentus, and A. excentricus (Extended Data Fig. 3), suggests that evolutionary changes in elongation modes stem not only from changes in gene content, but also from differences in the regulation, localization or activity of these conserved proteins.
Here, in characterizing the molecular determinants of unidirectional midcell elongation in A. excentricus, we discovered that the PG synthase PBP2 is central to the regulation of this elongation strategy. In contrast to its dispersed localization in C. crescentus, PBP2 in A. excentricus concentrates at the midcell, and is required for unidirectional PG synthesis. Upon inhibition of PBP2, cells exhibit abnormal PG synthesis leading to bulging at the midcell elongation site. These results suggest that changes in the specific localization of key enzymes such as PBP2 may be associated with the evolution of distinct cell elongation strategies, enabling bacteria to respond to broader pressures acting on morphology and growth. Notably, in C. crescentus, PBP2 shifts to the midcell under osmotic stress, illustrating that its localization is subject to regulation both by environmental cues and evolutionary processes33.
Beyond PBP2, other proteins like FtsZ play versatile roles in regulating localized PG synthesis across bacteria50. In C. crescentus, FtsZ depletion results in dispersed cell elongation23, while in Bacillus subtilis, FtsZ can position itself at the midcell during vegetative growth or closer to the poles for sporulation51, highlighting its flexibility in regulating PG synthesis for different growth modes. Another interesting example is the conserved outer membrane lipoprotein PapS, which in Rhodospirillum rubrum forms molecular cages that confine elongasomes to induce asymmetric cell elongation52. The diversity of mechanisms by which bacteria spatially regulate growth using conserved proteins supports the idea that evolution likely acts by altering the function of existing proteins rather than by introducing new genes. In the future, it will be of interest to determine what scaffolding mechanisms are at play in A. excentricus and A. biprosthecum, driving the unique localization and directionality of PBP2 in their elongasomes.
The observation of diverse elongation mechanisms among bacteria raises the fundamental question of the function of such diversity. Why do some species like A. excentricus evolve unidirectional elongation, while others maintain dispersed, bidirectional or polar growth mechanisms? What evolutionary factors drive these changes? It is clear that midcell and polar elongation are widely distributed in the Alphaproteobacteria (Fig. 5c), suggesting that localized modes of elongation may be ancestral within this class. However, the distribution of elongation modes in our phylogenetic analysis of the Caulobacteraceae (Fig. 5c–d) suggests that their evolution is likely shaped by multiple, independent events, implying a high degree of phenotypic plasticity in the regulation of localized elongation modes. In particular, the midcell, directional elongation identified in the current study is observed in Asticcacaulis and Phenylobacterium, but absent in members of Caulobacter and Brevundimonas, indicating that this growth mode has been gained and/or lost at least twice among the Caulobacteraceae (Fig. 5d). In light of this observed phenotypic diversity, it is intriguing to consider whether localized elongation itself may be a broadly adaptive strategy that bacteria have evolved, allowing greater plasticity under changing evolutionary pressures than dispersed elongation (although these co-exist in many species). Deeper understanding of the diversity of elongation modes among other branches of bacteria should provide further insights into the prevalence and ancestry of localized, dispersed, and septal modes of elongation, helping to identify selective pressures that shape the evolution of growth mode strategies.
Additionally, the evolution of distinct elongation modes has implications for the evolution of bacterial morphology. C. crescentus, A. excentricus, and A. biprosthecum display striking morphological differences, especially in the positioning of their stalks32. This raises the possibility that their distinct elongation modes and stalk positioning mechanisms may have co-evolved. In all three species, the site of stalk synthesis corresponds to the region of the oldest PG in the swarmer cell, as it differentiates into a stalked cell. Could the elongation mechanisms of these species thus be driving the placement of their cellular structures? If future studies find this to be true, it would suggest that elongation modes and cellular morphologies are intricately linked, with changes and selective pressures on one likely to influence the other over evolutionary time.
In summary, the fundamental finding of this study is that bacterial elongation modes are more diverse than previously thought, even at relatively short evolutionary scales. This challenges the assumption that closely related species share similar elongation modes – an assumption that has prevailed largely because we have not had the right tools to explore the diversity of bacterial elongation modes, even at close evolutionary distances from well-studied model organisms. Combining spatiotemporal analyses of PG synthesis using FDAAs alongside genetic and evolutionary approaches now allows a reexamination of previous assumptions about cell elongation modes across bacterial clades. Consequently, we expect these results to be at the forefront of a paradigm shift in our understanding of the diversity of bacterial cell elongation, its regulation and evolution.
Methods
Bacterial strains and growth conditions
All bacterial strains used in this study are listed in Table S1. C. crescentus, B. diminuta, and C. henricii were grown at 30°C in Peptone Yeast Extract (PYE) medium53. A. excentricus, A. biprosthecum and P. conjunctum were grown at 26°C in PYE. R. capsulatus SB1003 cells were grown at 30°C in PYS (3 g l−1 peptone, 3 g l−1yeast extract, 2 mM MgSO4, 2 mM CaCl2) medium54. The culture medium was supplemented with antibiotics as necessary at the following concentrations (μg ml−1; liquid/solid medium): spectinomycin (50/100), kanamycin (5/20), gentamicin (0.5/5), and with sucrose at 3% (w/v) for cloning procedures. For microscopy analysis, cells were grown either from a single colony or from frozen stock. Serial dilutions (1:10, 1:50, 1:100, and 1:1000) were made, and cultures were grown overnight at 26°C or 30°C with shaking at 220 rpm before being imaged in mid-exponential phase. E. coli strains used in this study were grown in liquid lysogeny broth (LB) medium at 37°C supplemented with antibiotics or supplements as necessary (diaminopimelic acid (DAP) 300 μg ml−1, kanamycin 50 μg ml−1, spectinomycin 100 μg ml−1, gentamicin 15 μg ml−1 and streptomycin 30 μg ml−1). Strains were maintained on LB plates at 37°C supplemented with antibiotics as necessary (kanamycin 50 μg ml−1, spectinomycin 100 μg ml−1 and streptomycin 30 μg ml−1).
Plasmid constructions and cloning procedures
All plasmids used in this study were cloned using standard molecular biology techniques and are listed in Table S2. PCRs were performed using A. excentricus CB48 WT or mutant genomic DNA as template. Gibson assemblies were performed using the Gibson Assembly® Master Mix from NEB55. Sequences of the primers used are available upon request.
In-frame deletions and fluorescent fusions in A. excentricus were obtained by double homologous recombination and sucrose counterselection, as previously described56. For deletions, 700-bp fragments from the upstream and downstream regions of the gene to be deleted were amplified by PCR. For the N-terminal mCherry fusion to PBP2, 500-bp of the upstream and N-terminus regions of the pbp2 gene were amplified by PCR, along with the mCherry gene. PCR fragments were gel-purified and cloned using Gibson assembly into the suicide vector pNPTS139 that had been digested by EcoRI and HindIII. The pNPTS139-based constructs were transformed into E. coli DH5α cells, verified by PCR sequencing, and then introduced into A. excentricus via biparental mating using the dap- E. coli strain WM3064 (YB7351)57. The pGFPC-5 plasmid with egfp replaced by sfgfp was used for generating the C-terminal sfGFP fusion to ZapA in A. excentricus. Proper chromosomal integration or gene replacement was verified by colony PCR and Sanger sequencing.
Fluorescent D-amino acids (FDAAs)
In this study, three different FDAAs were used: HADA (7-hydroxycoumarin-3-carboxylic acid-D-alanine; emission peak ~407nm), BADA (BODIPY FL-D-alanine; emission peak ~515nm), and TADA (TAMRA-D-alanine; emission peak ~578nm). FDAA stock solutions were prepared in anhydrous DMSO at a concentration of 100 mM. In fluorescent images and demographs, the FDAAs are false colored in green or red depending on the experiment, as detailed in the figure legends.
FDAA pulse-chase experiments
To label whole cells, 250 μM TADA or BADA was added to early exponential phase cells (OD600 ~0.1). The cells were allowed to grow for two generations, after which they were washed three times with appropriate medium to remove excess FDAA from the medium. Subsequently, growth was monitored following the wash using time-lapse microscopy, or cells were allowed to grow for half their generation times and imaged using phase and fluorescence microscopy to generate demographs (see “Image analysis” below).
For the FDAA pulse-chase experiments with mecillinam treatment, bacterial cells (OD600 ~0.1) were incubated with 250 μM TADA over two generations. Excess FDAA was removed by centrifugation at 6,000g for 3 min, and the cells were washed three times with PYE. Cell pellets were then resuspended in PYE with or without 50 µg ml−1 mecillinam, grown for one additional doubling time, and imaged using phase and fluorescence microscopy.
Dual short-pulse labeling with FDAAs
For dual short-pulse FDAA labeling in C. crescentus, HADA was added to early exponential phase cells (OD600 ~0.25) to a final concentration of 1 mM. The cells were grown in PYE at 30°C for 5% of their doubling time (5 minutes). Then, the excess dye was removed by centrifugation at 6,000g for 3 min, and cells were washed 3 times with PYE. The cell pellets were resuspended in PYE, and the cells were allowed to grow for an additional 15% of their generation time (15 minutes) in fresh PYE. BADA was then added to the culture medium to a final concentration of 1 mM. The cells were grown for an additional 5% of their doubling time for sequential labeling. Excess BADA was then removed by centrifugation at 6,000g for 3 min, and cells were washed 3 times with PYE. The labeled cells were resuspended in PYE and imaged with phase and fluorescence microscopy.
For dual short-pulse FDAA labeling in A. excentricus, TADA was added to early exponential phase cells (OD600 ~0.25) to a final concentration of 500 µM. The cells were grown in PYE at 26°C for 5% of their doubling time (6 minutes). Then, excess TADA was removed by centrifugation at 6,000g for 3 min, and cells were washed 3 times with PYE. The cell pellets were resuspended in PYE, and the cells were allowed to grow for an additional 15% of their generation time (18 minutes) in fresh PYE at 26°C. BADA was then added to the culture medium to a final concentration of 500 µM. The cells were then grown for an additional 5% of their doubling time for sequential labeling. Excess BADA was removed by centrifugation at 6,000g for 3 min, and cells were fixed in ethanol 70% for 1 hour. The fixed cells were washed with PYE twice and imaged with phase and fluorescence microscopy.
For dual short-pulse FDAA labeling in A. biprosthecum, TADA was added to early exponential phase cells (OD600 ~0.25) to a final concentration of 250 µM. The cells were grown in PYE at 26°C for 5% of their doubling time (7 min). Then, excess TADA was removed by centrifugation at 6,000g for 3 min, and cells were washed 3 times with PYE. The cell pellets were resuspended in PYE, and the cells were allowed to grow for an additional 15% of their generation time (21 minutes) in fresh PYE at 26°C. BADA was then added to the culture medium to a final concentration of 250 µM. The cells were then grown for an additional 5% of their doubling time for sequential labeling. Excess BADA was removed by centrifugation at 6,000g for 3 min, and cells were fixed in ethanol 70% for 1 hour. The fixed cells were washed with PYE twice and imaged with phase and fluorescence microscopy.
For dual short-pulse FDAA labeling in R. capsulatus, TADA was added to early exponential phase cells (OD600 ~0.25) to a final concentration of 250 µM. The cells were grown in PYS at 30°C for 5% of their doubling time (6 min). Then, excess TADA was removed by centrifugation at 6,000g for 3 min, and cells were washed 3 times with PYS. The cell pellets were resuspended in PYS, and the cells were allowed to grow for an additional 15% of their generation time (18 minutes) in fresh PYS at 26°C. BADA was then added to the culture medium to a final concentration of 250 µM. The cells were then grown for an additional 5% of their doubling time for dual sequential labeling. Excess BADA was removed by centrifugation at 6,000g for 3 min, and cells were fixed in ethanol 70% for 1 hour. The fixed cells were washed with PYS twice and imaged with phase and fluorescence microscopy
To orientate A. excentricus and A. biprosthecum cells using the old pole, holdfasts were detected with CF®405S conjugated wheat germ agglutinin (CF®405S WGA, 0.5 μg ml−1 final concentration) since WGA binds specifically to the acetylglucosamine residues present in their holdfasts58.
For FDAA dual short-pulse labeling in the presence of mecillinam, A. excentricus cells (OD600 ~0.25) were treated with mecillinam (50 µg ml−1) for 120 min. The treated cells were labeled with FDAAs as described above, but in presence of 50 µg ml−1 mecillinam for two sequential pulses of 5 min.
Single short-pulse labeling with FDAA in A. excentricus and C. crescentus PBP2 fluorescent fusion cells
BADA and TADA were added to early exponential phase (OD600 ~0.25) C. crescentus gfp-pbp2 and A. excentricus mCherry-pbp2 cells, to a final concentration of 250 µM. The C. crescentus and A. excentricus cells were grown in PYE at 30°C and at 26°C, respectively, for 5% of their doubling time. Excess FDAA was removed by centrifugation at 6,000g for 3 min and cells were fixed with 70% ethanol for 1 hour. Cells were washed 2 times with PYE, and labeled cells were then resuspended in PYE and imaged with phase and fluorescence microscopy.
Microscopy
For light microscopy analysis, 24 mm × 50 mm coverslips (#1.5) were used as imaging supports for an inverted microscopy system. An agarose pad was made of 1% SeaKem LE Agarose (Lonza, Cat. No. 50000) in dH2O. Cell samples were loaded onto the coverslips. Then, an 8 mm × 8 mm × 2 mm (length, width, thickness) dH2O-agar pad was laid on top of the cells. The coverslip–pad combination was placed onto a customized slide holder on microscopes with the pad facing upwards.
For time-lapse, 1 μl FDAA-labeled cells were spotted onto pads made of 0.7% Gelrite (Research product international, CAS. No. 71010-52-1) in PYE for C. crescentus, A. excentricus and A. biprosthecum cells and topped with a glass coverslip. 1 μl FDAA-labeled R. capsulatus cells were spotted onto pads made of 0.7% Gelrite in PYS and topped with a glass coverslip. The coverslip was sealed with VALAP (vaseline, lanolin, and paraffin at a 1:1:1 ratio).
Images were recorded with inverted Nikon Ti-E or Ti2 microscopes using a Plan Apo 60X 1.40 NA oil Ph3 DM objective with DAPI/FITC/Cy3/Cy5 or CFP/YFP/mCherry filter cubes and a Photometrics Prime 95B sCMOS camera. Images were processed with the NIS Elements software (Nikon).
Image analysis
Cell dimensions were obtained using FIJI59 and the plugin MicrobeJ37. To quantitatively analyze the pattern of FDAA loss in pulse-chase experiments, or of FDAA incorporation during dual short-pulse experiments, we generated kymographs and/or demographs using the MicrobeJ results interface. In these demographs, each cell is oriented such that the pole with the maximum mean fluorescence is set to the left and cells are aligned at the midcell. Alternatively, where holdfast staining was used, the holdfast signal (old pole) was set to the right. All cells were aligned at the midcell. Using the MicrobeJ demograph tool, 50% of the maximum intensity of each pulse was displayed in the main figures, while the total fluorescence signals are presented in the extended data. Demographs of the pulse-chase experiments of the different Caulobacteraceae species show 80% of the maximum fluorescence intensity.
Subcellular localization heatmaps and density maps for mCherry-PBP2 and FDAA foci were generated using MicrobeJ using the “Maxima” detection option. Density map merges were produced by importing each density map into Adobe Illustrator CC 2023 (Adobe Inc.) and manually merging the plots.
To quantitatively analyze the patterns of FDAA loss and cell bulging during pulse-chase experiments with mecillinam treatment, we quantified the subcellular localization of bulges along with the fluorescence intensities of ZapA and TADA. Localization of the bulge was determined using the “feature” option of MicrobeJ. Instead of looking for constriction, we looked for bulging, using the option “inverted” in the feature parameters interface. Using the subcellular localization charts function of the MicrobeJ results interface, we plotted bulge distribution relative to the cell center and generated a ShapePlot based on cell length to localize ZapA fluorescence intensity and FDAA signal loss as a readout for PG synthesis. GraphPad Prism (v. 10.3.0) was used to generate histograms and fluorescence intensity profiles and to perform statistical analysis.
In vitro mecillinam titration against PBPs
In vitro mecillinam titration against PBPs was performed with modifications to a previous protocol for PBP detection in E. coli60. Specifically, A. excentricus cells in exponential phase (OD600 ~0.5) were harvested by centrifugation at 10,000g for 4 min at room temperature. The cell pellets were washed twice with 1 ml PBS (pH 7.4). Cells were then resuspended in 50 μl PBS containing 1, 10, or 100 μg ml−1 of the antibiotic, while a reference sample was resuspended in 50 μl PBS without antibiotics. After 4 h of incubation at room temperature, cells were pelleted, washed with PBS, and resuspended in 50 μl PBS containing 5 μg ml−1 Boc-FL. Following a 30-min incubation at room temperature, cells were pelleted, washed with 1 ml PBS, and then resuspended in 100 μl PBS. The cells were sonicated on ice using a Branson Sonifier 250 instrument (power setting 60 A, 30 s cycle for three 10 s intervals with 10 s of cooling time between rounds) to isolate the membrane proteome. The membrane pellet was then resuspended in 100 μl PBS and homogenized by sonication (power setting 20 A for 1 s). The protein concentration was measured using a NanoDrop 1000 Spectrophotometer and adjusted to 2.5 mg ml−1 using PBS. Proteome samples (20 μl) were dispensed into clean 1.5 ml microcentrifuge tubes, and 10 μl of 2× SDS-PAGE loading buffer was added to each sample. The samples were heated for 5 min at 90 °C to denature the proteins, cooled to room temperature, and then 25 μl of each sample was loaded onto a 4–15% SDS-PAGE precast gel. The gel was rinsed with distilled water three times and scanned using a Gel Doc XR system (Bio-Rad Laboratories, Inc) with a 526 nm short-pass filter.
Phylogenetic analysis of selected Alphaproteobacteria
Whole-genome data were obtained from the genome database maintained by the National Center for Biotechnology Information61. From each genome, a set of 37 conserved genes was identified and the translated amino acid sequences aligned and concatenated using Phylosift62 Phylogenetic reconstruction with MrBayes63 used a mixed amino acid model including a four-category approximation of gamma-distributed rate variation and an invariant site category. Two simultaneous Markov chain runs were performed for 3,000,000 generations, discarding the initial 25% for burn-in. The tree was visualized and formatted using iTOL64.
Bioinformatic analyses and the phylogeny of PBPs
Identification of putative PBPs in A. excentricus was performed by BLAST analysis using C. crescentus PBP2 and PBP1a as queries. Amino acid sequences of homologues (including from other model organisms) were collected from UniProt (https://www.uniprot.org): PBP5 (BSU00100) in B. subtilis, DacD (JW5329), PBP1a (JW3359), PBP1b (JW0145), PBP1c (JW2503), PBP2 (JW0630), PBP3 (JW0082) and MTG (JW3175) in E. coli, PBP3 (Astex_1844), PBP2 (Astex_1631), PBP1a (Astex_2994), PBP1c (Astex_0196), PBP1b (Astex_2378) and MTG (Astex_0406) in A. excentricus, and PBP3 (CCNA_02643), PBP2 (CCNA_01615), PBPz (CCNA_93685), PBPc (CCNA_03386), PBPx (CCNA_01584), PBP1a (CCNA_01584) and MTG (CCNA_00328) in C. crescentus. Sequences were then aligned using MUSCLE v.3.8.31 (Fig. S5.1), and PhyML 3.0 was used to reconstruct the maximum likelihood phylogenetic tree, with automatic model selection by Smart Model Selection (SMS)65 and Akaike information criterion. Phylogenetic reconstruction was performed by RAxML version 8.2.1066 with 100 rapid bootstraps replicates to assess node support. The tree was visualized and formatted using iTOL64. Taxonomic assignments were based on the taxonomy database maintained by NCBI (https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi).
Genomic organization of the pbp2 and mreB genes
The genomic organization of the pbp2 and mreB operons was analyzed using a combination of annotation, sequence search, and synteny visualization tools. Genome assemblies were first annotated with prokka67. The annotated files were organized into folders according to their formats (FAA, FNA, GFF, and GBK). For each FAA file, a BLAST protein database was created using the makeblastdb function, and the FNA files were indexed using samtools. GBK files were separated by contig using a Python script with BioPython’s SeqIO library68 to save each contig as an individual GBK file.
When protein files contained multiple sequences, they were split into individual files using SeqKit69. Each protein sequence was then identified by performing a BLAST search against the FAA databases. The blast search results were filtered to retain the first hit. Locus tags from the BLAST hits were extracted from the GFF files and converted to BED format. For analyses requiring extended genomic regions, the coordinates were expanded by 5000 bp upstream and downstream using bedtools slop.
To conduct synteny analysis, locus tags from the BED regions were extracted, and the corresponding protein sequences were retrieved from the FAA files using SeqKit69. A BLAST database was generated from these sequences, followed by an all-vs-all BLAST search. The matches were clustered based on sequence identity using the MCL algorithm70, grouping homologous sequences.
Clusters were assigned specific colors, and the GBK files were edited by adding these colors for each corresponding locus tag. Synteny diagrams were created with EasyFig71, with coding regions represented as arrows and other genomic features (e.g., tRNA and rRNA genes) depicted as rectangles. Multiple synteny figures were arranged in the desired order and merged using Adobe Illustrator CC 2023 (Adobe Inc.).
Illumina sequencing methods
Illumina sequencing was performed by SeqCenter in Pittsburgh, PA.
Sample libraries were prepared using the Illumina DNA Prep kit and IDT 10bp UDI indices, and sequenced on an Illumina NextSeq 2000, producing 2×151bp reads. Demultiplexing, quality control and adapter trimming were performed with bcl-convert (v3.9.30).
Variant Calling Methods
Illumina-generated 2×151bp paired-end read data was used as the input for variant calling against the provided GenBank AC48 reference. Variant calling was carried out using Breseq (v0.37.1) under default settings72. Mutations were confirmed by Sanger sequencing.
Supplementary Material
Acknowledgments
The NIH supported this work with grants R35GM122556 to Y.V.B. and R35GM136365 to M.S.V. Y.V.B. is also supported by a Canada 150 Research Chair in Bacterial Cell Biology. M.D. was in part supported by a postdoctoral fellowship from the Swiss National Science Foundation (projet #P2GEP3_191489, and P500PB_206676). F.J.V. received a Junior 1 and Junior 2 research scholar salary award from the Fonds de Recherche du Québec-Santé. Thanks to Yen-pang Hsu for synthesizing the FDAA. We thank Adrien Ducret for help with MicrobeJ. Many thanks to previous and current Brun Lab members for either providing guidance to the project or proofreading the manuscript.
Footnotes
Competing interests
The authors declare no competing interests.
Data Availability
The data that support the findings of this study are available from the corresponding author on request.
Reference
- 1.Curtis P. D. & Brun Y. V. Getting in the loop: regulation of development in Caulobacter crescentus. Microbiol Mol Biol Rev 74, 13–41, doi: 10.1128/MMBR.00040-09 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Woldemeskel S. A. & Goley E. D. Shapeshifting to Survive: Shape Determination and Regulation in Caulobacter crescentus. Trends Microbiol 25, 673–687, doi: 10.1016/j.tim.2017.03.006 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Egan A. J., Errington J. & Vollmer W. Regulation of peptidoglycan synthesis and remodelling. Nature Reviews Microbiology 18, 446–460 (2020). [DOI] [PubMed] [Google Scholar]
- 4.Kysela D. T., Randich A. M., Caccamo P. D. & Brun Y. V. Diversity Takes Shape: Understanding the Mechanistic and Adaptive Basis of Bacterial Morphology. PLoS Biol 14, e1002565, doi: 10.1371/journal.pbio.1002565 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Randich A. M. & Brun Y. V. Molecular mechanisms for the evolution of bacterial morphologies and growth modes. Front Microbiol 6, 580, doi: 10.3389/fmicb.2015.00580 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Garner E. C. Toward a Mechanistic Understanding of Bacterial Rod Shape Formation and Regulation. Annual Review of Cell and Developmental Biology 37, null, doi: 10.1146/annurev-cellbio-010521-010834 (2021). [DOI] [PubMed] [Google Scholar]
- 7.Egan A. J., Biboy J., van’t Veer I., Breukink E. & Vollmer W. Activities and regulation of peptidoglycan synthases. Philosophical Transactions of the Royal Society B: Biological Sciences 370, 20150031 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Egan A. J., Cleverley R. M., Peters K., Lewis R. J. & Vollmer W. Regulation of bacterial cell wall growth. The FEBS journal 284, 851–867 (2017). [DOI] [PubMed] [Google Scholar]
- 9.Brown P. J., de Pedro M. A., Kysela D. T., Van der Henst C., Kim J., De Bolle X., Fuqua C. & Brun Y. V. Polar growth in the Alphaproteobacterial order Rhizobiales. Proc Natl Acad Sci U S A 109, 1697–1701, doi: 10.1073/pnas.1114476109 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cameron T. A., Anderson-Furgeson J., Zupan J. R., Zik J. J. & Zambryski P. C. Peptidoglycan synthesis machinery in Agrobacterium tumefaciens during unipolar growth and cell division. mBio 5, e01219–01214, doi: 10.1128/mBio.01219-14 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cameron T. A., Zupan J. R. & Zambryski P. C. The essential features and modes of bacterial polar growth. Trends in Microbiology 23, 347–353, doi: 10.1016/j.tim.2015.01.003 (2015). [DOI] [PubMed] [Google Scholar]
- 12.Kieser K. J. & Rubin E. J. How sisters grow apart: mycobacterial growth and division. Nature Reviews Microbiology 12, 550–562, doi: 10.1038/nrmicro3299 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sher J. W., Lim H. C. & Bernhardt T. G. Polar Growth in Corynebacterium glutamicum Has a Flexible Cell Wall Synthase Requirement. mBio 12, e0068221, doi: 10.1128/mBio.00682-21 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Williams M. A., Aliashkevich A., Krol E., Kuru E., Bouchier J. M., Rittichier J., Brun Y. V., VanNieuwenhze M. S., Becker A., Cava F. & Brown P. J. B. Unipolar peptidoglycan synthesis in the Rhizobiales requires an essential class A penicillin-binding protein. bioRxiv, 2021.2003.2031.437934, doi: 10.1101/2021.03.31.437934 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hett E. C. & Rubin E. J. Bacterial growth and cell division: a mycobacterial perspective. Microbiol. Mol. Biol. Rev. 72, 126–156 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Baranowski C., Rego E. H. & Rubin E. J. The dream of a Mycobacterium. Gram‐Positive Pathogens, 1096–1106 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Canepari P., Signoretto C., Boaretti M. & Lleò M. D. M. Cell elongation and septation are two mutually exclusive processes in Escherichia coli. Archives of microbiology 168, 152–159 (1997). [DOI] [PubMed] [Google Scholar]
- 18.Kieser K. J. & Rubin E. J. How sisters grow apart: mycobacterial growth and division. Nature Reviews Microbiology 12, 550 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Garner E. C., Bernard R., Wang W., Zhuang X., Rudner D. Z. & Mitchison T. Coupled, circumferential motions of the cell wall synthesis machinery and MreB filaments in B. subtilis. Science 333, 222–225 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pinho M. G., Kjos M. & Veening J. W. How to get (a)round: mechanisms controlling growth and division of coccoid bacteria. Nat Rev Microbiol 11, 601–614, doi: 10.1038/nrmicro3088 (2013). [DOI] [PubMed] [Google Scholar]
- 21.Philippe J., Vernet T. & Zapun A. The elongation of ovococci. Microb Drug Resist 20, 215–221, doi: 10.1089/mdr.2014.0032 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Caccamo P. D. & Brun Y. V. The Molecular Basis of Noncanonical Bacterial Morphology. Trends in Microbiology 26, 191–208, doi: 10.1016/j.tim.2017.09.012 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Aaron M., Charbon G., Lam H., Schwarz H., Vollmer W. & Jacobs-Wagner C. The tubulin homologue FtsZ contributes to cell elongation by guiding cell wall precursor synthesis in Caulobacter crescentus. Molecular microbiology 64, 938 (2007). [DOI] [PubMed] [Google Scholar]
- 24.Leifson E. HYPHOMICROBIUM NEPTUNIUM SP. N. Antonie Van Leeuwenhoek 30, 249–256, doi: 10.1007/bf02046730 (1964). [DOI] [PubMed] [Google Scholar]
- 25.Wali T. M., Hudson G. R., Danald D. A. & Weiner R. M. Timing of swarmer cell cycle morphogenesis and macromolecular synthesis by Hyphomicrobium neptunium in synchronous culture. J Bacteriol 144, 406–412, doi: 10.1128/jb.144.1.406-412.1980 (1980). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Badger J. H., Hoover T. R., Brun Y. V., Weiner R. M., Laub M. T., Alexandre G., Mrázek J., Ren Q., Paulsen I. T., Nelson K. E., Khouri H. M., Radune D., Sosa J., Dodson R. J., Sullivan S. A., Rosovitz M. J., Madupu R., Brinkac L. M., Durkin A. S., Daugherty S. C., Kothari S. P., Giglio M. G., Zhou L., Haft D. H., Selengut J. D., Davidsen T. M., Yang Q., Zafar N. & Ward N. L. Comparative genomic evidence for a close relationship between the dimorphic prosthecate bacteria Hyphomonas neptunium and Caulobacter crescentus. J Bacteriol 188, 6841–6850, doi: 10.1128/jb.00111-06 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kuru E., Hughes H. V., Brown P. J., Hall E., Tekkam S., Cava F., de Pedro M. A., Brun Y. V. & VanNieuwenhze M. S. In Situ probing of newly synthesized peptidoglycan in live bacteria with fluorescent D-amino acids. Angew Chem Int Ed Engl 51, 12519–12523, doi: 10.1002/anie.201206749 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kuru E., Radkov A., Meng X., Egan A., Alvarez L., Dowson A., Booher G., Breukink E., Roper D. I., Cava F., Vollmer W., Brun Y. & VanNieuwenhze M. S. Mechanisms of Incorporation for D-Amino Acid Probes That Target Peptidoglycan Biosynthesis. ACS Chem Biol 14, 2745–2756, doi: 10.1021/acschembio.9b00664 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Woldemeskel S. A., McQuillen R., Hessel A. M., Xiao J. & Goley E. D. A conserved coiled-coil protein pair focuses the cytokinetic Z-ring in Caulobacter crescentus. Mol Microbiol 105, 721–740, doi: 10.1111/mmi.13731 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kuru E., Tekkam S., Hall E., Brun Y. V. & Van Nieuwenhze M. S. Synthesis of fluorescent D-amino acids and their use for probing peptidoglycan synthesis and bacterial growth in situ. Nat Protoc 10, 33–52, doi: 10.1038/nprot.2014.197 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wagner J. K. & Brun Y. V. Out on a limb: how the Caulobacter stalk can boost the study of bacterial cell shape. Molecular microbiology 64, 28–33 (2007). [DOI] [PubMed] [Google Scholar]
- 32.Jiang C., Brown P. J., Ducret A. & Brun Y. V. Sequential evolution of bacterial morphology by co-option of a developmental regulator. Nature 506, 489–493 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hocking J., Priyadarshini R., Takacs C. N., Costa T., Dye N. A., Shapiro L., Vollmer W. & Jacobs-Wagner C. Osmolality-dependent relocation of penicillin-binding protein PBP2 to the division site in Caulobacter crescentus. Journal of bacteriology 194, 3116–3127 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Legaree B. A., Daniels K., Weadge J. T., Cockburn D. & Clarke A. J. Function of penicillin-binding protein 2 in viability and morphology of Pseudomonas aeruginosa. Journal of antimicrobial chemotherapy 59, 411–424 (2007). [DOI] [PubMed] [Google Scholar]
- 35.Koyasu S., Fukuda A., Okada Y. & Poindexter J. S. Penicillin-binding Proteins of the Stalk of Caulobacter crescentus. Microbiology 129, 2789–2799, doi: 10.1099/00221287-129-9-2789 (1983). [DOI] [Google Scholar]
- 36.Seitz L. C. & Brun Y. V. Genetic analysis of mecillinam-resistant mutants of Caulobacter crescentus deficient in stalk biosynthesis. Journal of bacteriology 180, 5235–5239, doi: 10.1128/JB.180.19.5235-5239.1998 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ducret A., Quardokus E. M. & Brun Y. V. MicrobeJ, a tool for high throughput bacterial cell detection and quantitative analysis. Nat Microbiol 1, 16077, doi: 10.1038/nmicrobiol.2016.77 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Brown P. J., Kysela D. T. & Brun Y. V. Polarity and the diversity of growth mechanisms in bacteria. Semin Cell Dev Biol 22, 790–798, doi: 10.1016/j.semcdb.2011.06.006 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zambri M. P., Baglio C. R., Irazoki O., Jones S. E., Garner E. C., Cava F. & Elliot M. A. Bacteria combine polar- and dispersed-growth to power cell elongation and wall width dynamics. bioRxiv, 2024.2007.2030.605496, doi: 10.1101/2024.07.30.605496 (2024). [DOI] [Google Scholar]
- 40.Jones S. E., Ho L., Rees C. A., Hill J. E., Nodwell J. R. & Elliot M. A. Streptomyces exploration is triggered by fungal interactions and volatile signals. Elife 6, doi: 10.7554/eLife.21738 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Castanheira S. & García-del Portillo F. Evidence of two differentially regulated elongasomes in Salmonella. Communications Biology 6, 923, doi: 10.1038/s42003-023-05308-w (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Karasz D. C., Weaver A. I., Buckley D. H. & Wilhelm R. C. Conditional filamentation as an adaptive trait of bacteria and its ecological significance in soils. Environmental Microbiology 24, 1–17, doi: 10.1111/1462-2920.15871 (2022). [DOI] [PubMed] [Google Scholar]
- 43.Justice S. S., Hunstad D. A., Cegelski L. & Hultgren S. J. Morphological plasticity as a bacterial survival strategy. Nat Rev Microbiol 6, 162–168, doi: 10.1038/nrmicro1820 (2008). [DOI] [PubMed] [Google Scholar]
- 44.Justice S. S., Hunstad D. A., Seed P. C. & Hultgren S. J. Filamentation by Escherichia coli subverts innate defenses during urinary tract infection. Proc Natl Acad Sci U S A 103, 19884–19889, doi: 10.1073/pnas.0606329104 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Billini M., Biboy J., Kühn J., Vollmer W. & Thanbichler M. A specialized MreB-dependent cell wall biosynthetic complex mediates the formation of stalk-specific peptidoglycan in Caulobacter crescentus. PLOS Genetics 15, e1007897, doi: 10.1371/journal.pgen.1007897 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wagner J. K., Galvani C. D. & Brun Y. V. Caulobacter crescentus requires RodA and MreB for stalk synthesis and prevention of ectopic pole formation. J Bacteriol 187, 544–553, doi: 10.1128/JB.187.2.544-553.2005 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.van der Ploeg R., Verheul J., Vischer N. O. E., Alexeeva S., Hoogendoorn E., Postma M., Banzhaf M., Vollmer W. & den Blaauwen T. Colocalization and interaction between elongasome and divisome during a preparative cell division phase in scherichia coli. Molecular Microbiology 87, 1074–1087, doi: 10.1111/mmi.12150 (2013). [DOI] [PubMed] [Google Scholar]
- 48.Reichmann N. T., Tavares A. C., Saraiva B. M., Jousselin A., Reed P., Pereira A. R., Monteiro J. M., Sobral R. G., VanNieuwenhze M. S., Fernandes F. & Pinho M. G. SEDS-bPBP pairs direct lateral and septal peptidoglycan synthesis in Staphylococcus aureus. Nat Microbiol 4, 1368–1377, doi: 10.1038/s41564-019-0437-2 (2019). [DOI] [PubMed] [Google Scholar]
- 49.Lakey B. D., Alberge F., Parrell D., Wright E. R., Noguera D. R. & Donohue T. J. The role of CenKR in the coordination of Rhodobacter sphaeroides cell elongation and division. mBio 14, e00631–00623, doi:doi: 10.1128/mbio.00631-23 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Randich A. M. & Brun Y. V. Molecular mechanisms for the evolution of bacterial morphologies and growth modes. Frontiers in Microbiology 6, doi: 10.3389/fmicb.2015.00580 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Khanna K., Lopez-Garrido J., Sugie J., Pogliano K. & Villa E. Asymmetric localization of the cell division machinery during Bacillus subtilis sporulation. eLife 10, e62204, doi: 10.7554/eLife.62204 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Pöhl S., Giacomelli G., Meyer F. M., Kleeberg V., Cohen E. J., Biboy J., Rosum J., Glatter T., Vollmer W., van Teeseling M. C. F., Heider J., Bramkamp M. & Thanbichler M. An outer membrane porin-lipoprotein complex modulates elongasome movement to establish cell curvature in Rhodospirillum rubrum. Nature Communications 15, 7616, doi: 10.1038/s41467-024-51790-z (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Poindexter J. S. Biological properties and classification of the Caulobacter group. Bacteriological reviews 28, 231 (1964). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zappa S. & Bauer C. E. The LysR-type transcription factor HbrL is a global regulator of iron homeostasis and porphyrin synthesis in Rhodobacter capsulatus. Mol Microbiol 90, 1277–1292, doi: 10.1111/mmi.12431 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Gibson D. G., Young L., Chuang R. Y., Venter J. C., Hutchison C. A. 3rd & Smith H. O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat Methods 6, 343–345, doi: 10.1038/nmeth.1318 (2009). [DOI] [PubMed] [Google Scholar]
- 56.Ried J. L. & Collmer A. An nptI-sacB-sacR cartridge for constructing directed, unmarked mutations in gram-negative bacteria by marker exchange-eviction mutagenesis. Gene 57, 239–246, doi: 10.1016/0378-1119(87)90127-2 (1987). [DOI] [PubMed] [Google Scholar]
- 57.Saltikov C. W. & Newman D. K. Genetic identification of a respiratory arsenate reductase. Proc Natl Acad Sci U S A 100, 10983–10988, doi: 10.1073/pnas.1834303100 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Merker R. I. & Smit J. Characterization of the adhesive holdfast of marine and freshwater caulobacters. Applied and environmental microbiology 54, 2078–2085, doi: 10.1128/aem.54.8.2078-2085.1988 (1988). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Schindelin J., Arganda-Carreras I., Frise E., Kaynig V., Longair M., Pietzsch T., Preibisch S., Rueden C., Saalfeld S., Schmid B., Tinevez J.-Y., White D. J., Hartenstein V., Eliceiri K., Tomancak P. & Cardona A. Fiji: an open-source platform for biological-image analysis. Nature Methods 9, 676–682, doi: 10.1038/nmeth.2019 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kocaoglu O. & Carlson E. E. Profiling of β-lactam selectivity for penicillin-binding proteins in Escherichia coli strain DC2. Antimicrobial agents and chemotherapy 59, 2785–2790 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Database resources of the National Center for Biotechnology Information. Nucleic Acids Res 44, D7–19, doi: 10.1093/nar/gkv1290 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Darling A. E., Jospin G., Lowe E., Matsen F. A. t., Bik H. M. & Eisen J. A. PhyloSift: phylogenetic analysis of genomes and metagenomes. PeerJ 2, e243, doi: 10.7717/peerj.243 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ronquist F., Teslenko M., van der Mark P., Ayres D. L., Darling A., Höhna S., Larget B., Liu L., Suchard M. A. & Huelsenbeck J. P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol 61, 539–542, doi: 10.1093/sysbio/sys029 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Letunic I. & Bork P. Interactive tree of life (iTOL) v3: an online tool for the display and annotation of phylogenetic and other trees. Nucleic acids research 44, W242–W245 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lefort V., Longueville J.-E. & Gascuel O. SMS: smart model selection in PhyML. Molecular biology and evolution 6, doi: 10.1093/molbev/msx149 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313, doi: 10.1093/bioinformatics/btu033 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069, doi: 10.1093/bioinformatics/btu153 (2014). [DOI] [PubMed] [Google Scholar]
- 68.Cock P. J. A., Antao T., Chang J. T., Chapman B. A., Cox C. J., Dalke A., Friedberg I., Hamelryck T., Kauff F., Wilczynski B. & de Hoon M. J. L. Biopython: freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 25, 1422–1423, doi: 10.1093/bioinformatics/btp163 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Shen W., Le S., Li Y. & Hu F. SeqKit: A Cross-Platform and Ultrafast Toolkit for FASTA/Q File Manipulation. PLOS ONE 11, e0163962, doi: 10.1371/journal.pone.0163962 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Van Dongen S. Graph Clustering Via a Discrete Uncoupling Process. SIAM Journal on Matrix Analysis and Applications 30, 121–141, doi: 10.1137/040608635 (2008). [DOI] [Google Scholar]
- 71.Sullivan M. J., Petty N. K. & Beatson S. A. Easyfig: a genome comparison visualizer. Bioinformatics 27, 1009–1010, doi: 10.1093/bioinformatics/btr039 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Deatherage D. E. & Barrick J. E. Identification of mutations in laboratory-evolved microbes from next-generation sequencing data using breseq. Methods Mol Biol 1151, 165–188, doi: 10.1007/978-1-4939-0554-6_12 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Letunic I. & Bork P. Interactive tree of life (iTOL) v3: an online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res 44, W242–245, doi: 10.1093/nar/gkw290 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Dion M. F., Kapoor M., Sun Y., Wilson S., Ryan J., Vigouroux A., van Teeffelen S., Oldenbourg R. & Garner E. C. Bacillus subtilis cell diameter is determined by the opposing actions of two distinct cell wall synthetic systems. Nat Microbiol 4, 1294–1305, doi: 10.1038/s41564-019-0439-0 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Evinger M. & Agabian N. Envelope-associated nucleoid from Caulobacter crescentus stalked and swarmer cells. Journal of Bacteriology 132, 294–301 (1977). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Woldemeskel S. A., McQuillen R., Hessel A. M., Xiao J. & Goley E. D. A conserved coiled‐ coil protein pair focuses the cytokinetic Z‐ring in Caulobacter crescentus. Molecular microbiology 105, 721–740 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.West L., Yang D. & Stephens C. Use of the Caulobacter crescentus genome sequence to develop a method for systematic genetic mapping. J Bacteriol 184, 2155–2166, doi: 10.1128/jb.184.8.2155-2166.2002 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Poindexter J. L. S. & Cohen-Bazire G. The fine structure of stalked bacteria belonging to the family Caulobacteraceae. The Journal of cell biology 23, 587–607 (1964). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Pate J. L. & Ordal E. J. The fine structure of two unusual stalked bacteria. J Cell Biol 27, 133–150, doi: 10.1083/jcb.27.1.133 (1965). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Thanbichler M., Iniesta A. A. & Shapiro L. A comprehensive set of plasmids for vanillate-and xylose-inducible gene expression in Caulobacter crescentus. Nucleic acids research 35, e137–e137 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lu H., Chen L., Kong L., Huang L. & Chen G. Asticcacaulis aquaticus sp. nov., Asticcacaulis currens sp. nov. and Asticcacaulis machinosus sp. nov., isolated from streams in PR China. Int J Syst Evol Microbiol 73, doi: 10.1099/ijsem.0.005974 (2023). [DOI] [PubMed] [Google Scholar]
- 82.Vasilyeva L. V., Omelchenko M. V., Berestovskaya Y. Y., Lysenko A. M., Abraham W. R., Dedysh S. N. & Zavarzin G. A. Asticcacaulis benevestitus sp. nov., a psychrotolerant, dimorphic, prosthecate bacterium from tundra wetland soil. Int J Syst Evol Microbiol 56, 2083–2088, doi: 10.1099/ijs.0.64122-0 (2006). [DOI] [PubMed] [Google Scholar]
- 83.Staley J. T., Bont J. A. & Jonge K. Prosthecobacter fusiformis nov. gen. et sp., the fusiform caulobacter. Antonie Van Leeuwenhoek 42, 333–342, doi: 10.1007/bf00394132 (1976). [DOI] [PubMed] [Google Scholar]
- 84.Quan Z. X., Zeng D. N., Xiao Y. P., Roh S. W., Nam Y. D., Chang H. W., Yoon J. H., Oh H. M. & Bae J. W. Henriciella marina gen. nov., sp. nov., a novel member of the family Hyphomonadaceae isolated from the East Sea. J Microbiol 47, 156–161, doi: 10.1007/s12275-008-0290-0 (2009). [DOI] [PubMed] [Google Scholar]
- 85.Schlesner H., Bartels C., Sittig M., Dorsch M. & Stackebrandt E. Taxonomic and phylogenetic studies on a new taxon of budding, hyphal Proteobacteria, Hirschia baltica gen. nov., sp. nov. Int J Syst Bacteriol 40, 443–451, doi: 10.1099/00207713-40-4-443 (1990). [DOI] [PubMed] [Google Scholar]
- 86.URAKAMI T., SASAKI J., SUZUKI K.-I. & KOMAGATA K. Characterization and Description of Hyphomicrobium denitrificans sp. nov. International Journal of Systematic and Evolutionary Microbiology 45, 528–532, doi: 10.1099/00207713-45-3-528 (1995). [DOI] [Google Scholar]
- 87.Moore R. L. The biology of Hyphomicrobium and other prosthecate, budding bacteria. Annu Rev Microbiol 35, 567–594, doi: 10.1146/annurev.mi.35.100181.003031 (1981). [DOI] [PubMed] [Google Scholar]
- 88.Hirsch P. Budding bacteria. Annu Rev Microbiol 28, 391–444, doi: 10.1146/annurev.mi.28.100174.002135 (1974). [DOI] [PubMed] [Google Scholar]
- 89.Cserti E., Rosskopf S., Chang Y. W., Eisheuer S., Selter L., Shi J., Regh C., Koert U., Jensen G. J. & Thanbichler M. Dynamics of the peptidoglycan biosynthetic machinery in the stalked budding bacterium Hyphomonas neptunium. Mol Microbiol 103, 875–895, doi: 10.1111/mmi.13593 (2017). [DOI] [PubMed] [Google Scholar]
- 90.Abraham W. R., Strömpl C., Meyer H., Lindholst S., Moore E. R., Christ R., Vancanneyt M., Tindall B. J., Bennasar A., Smit J. & Tesar M. Phylogeny and polyphasic taxonomy of Caulobacter species. Proposal of Maricaulis gen. nov. with Maricaulis maris (Poindexter) comb. nov. as the type species, and emended description of the genera Brevundimonas and Caulobacter. Int J Syst Bacteriol 49 Pt 3, 1053–1073, doi: 10.1099/00207713-49-3-1053 (1999). [DOI] [PubMed] [Google Scholar]
- 91.Strömpl C., Hold G. L., Lünsdorf H., Graham J., Gallacher S., Abraham W. R., Moore E. R. & Timmis K. N. Oceanicaulis alexandrii gen. nov., sp. nov., a novel stalked bacterium isolated from a culture of the dinoflagellate Alexandrium tamarense (Lebour) Balech. Int J Syst Evol Microbiol 53, 1901–1906, doi: 10.1099/ijs.0.02635-0 (2003). [DOI] [PubMed] [Google Scholar]
- 92.Wu N., Wu Y., Liu L., Zhang Q., Lv Y., Yuan Y., He J. & Shen Q. Peiella sedimenti gen. nov., sp. nov., a novel taxon within the family Caulobacteraceae isolated from sediment of a river. International Journal of Systematic and Evolutionary Microbiology 74, doi: 10.1099/ijsem.0.006344 (2024). [DOI] [PubMed] [Google Scholar]
- 93.Lee K., Lee H. K., Choi T. H. & Cho J. C. Robiginitomaculum antarcticum gen. nov., sp. nov., a member of the family Hyphomonadaceae, from Antarctic seawater. Int J Syst Evol Microbiol 57, 2595–2599, doi: 10.1099/ijs.0.65274-0 (2007). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data that support the findings of this study are available from the corresponding author on request.