

Abstract
Gene set enrichment analysis (GSEA) tools can identify biological insights within gene expression-based studies. Although their statistical performance has been compared, the downstream biological implications that arise when choosing between the range of pairwise or single sample forms of GSEA methods remain understudied. We compare the statistical and biological results obtained from various pre-ranking methods/options for pairwise GSEA, followed by a stand-alone comparison of GSEA, single sample GSEA (ssGSEA) and gene set variation analysis (GSVA). Pairwise GSEA and fGSEA provide similar results when deployed using a range of gene pre-ranking methods. However, pairwise GSEA can overgeneralise biological enrichment, as when the most statistically significant signatures were assessed using single sample approaches, there was a complete absence of biological distinction between these groups. To avoid these issues, we developed a new dualGSEA tool, which provides users with multiple statistics and visuals to aid interpretation of results. This new tool removes the possibility of users inadvertently interpreting statistical findings as equating to biological distinction between samples within groups-of-interest. dualGSEA provides a more robust basis for discovery research, one which allows user to compare both statistical significance alongside biological distinctions in their data.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-024-80534-8.
Keywords: Transcriptional signatures, GSEA, Molecular classification, Computational biology
Subject terms: Cancer genomics, Tumour biomarkers
Introduction
Decreasing costs for sequencing, coupled with an increasing adoption of the FAIR principles1, have provided the cancer research field with a substantial amount of freely available molecular datasets derived from tumour tissue samples. To ensure that these large datasets can reveal important mechanistic insights, increased data availability has been coupled with the development of transcriptional signatures that represent important biological pathways, alongside easy-to-use algorithms that allow users to apply thousands of signatures simultaneously to these data. These are exemplified by the establishment of the Molecular Signatures Database (MSigDB)2 and gene set enrichment analysis (GSEA) tools3, providing the field with a stable set of reference templates and methods to compare across cohorts of interest. The success of these approaches has led to a rapid expansion of established signature collections in both human and mouse, most notably the MSigDB biological “Hallmark” collection4 and development of programming software-based GSEA tools such as the clusterProfiler5 and fast GSEA (fGSEA)6 R packages.
Given that many tumour cohorts have associated metadata linked to important features, such as clinical outcome, the application of these large collections of signatures to cohorts in conjunction with GSEA can serve as the basis for discovery and validation of biomarkers that represent the biological characteristics of the chosen features, such as prognosis. This approach is referred to as a supervised pairwise analysis, as the groups are known prior to application of the GSEA method, and these tools have been tested extensively in terms of the statistical robustness and performance in this setting7,8. Once identified, these biomarkers can be used as the basis for mechanistic investigations, pre-clinical model development, and/or testing of a therapeutic target.
Alongside pairwise GSEA methods, approaches for single sample methods have been developed, which differ from the pairwise approach in that they allow users to apply the same transcriptional signature collections to all samples individually in a cohort, using single sample GSEA (ssGSEA)9 and gene set variation analysis (GSVA)10. While these single sample approaches are based on different statistical models to those in pairwise analyses, the resulting outputs are based on the same gene signatures. Numerous studies have assessed the statistical robustness and performance of this range of pairwise and single sample tools separately7,11. Despite differences being identified between methods when assessed using statistically-driven criteria, few studies have focussed on the consequences in terms of downstream biological approaches. Given that significant pairwise GSEA results can be interpreted as representing the defining biological characteristics of a group of samples, the absence of a comparative study across all approaches means that such an interpretation may be based on incomplete evidence.
In this study, we use a fixed set of transcriptional signatures, in conjunction with a fixed clinical feature (relapse status) within a well-characterised colon cancer (CC) transcriptional cohort12, to perform a series of pairwise and single sample assessments in tandem. Each output is assessed based on the provided statistical values, however the primary focus of this study is to assess how representative and uniform a significant pairwise result is when assessed by single sample methods. Utilising a range of data visualisations and performance measurements, we find that statistical results from a pairwise analysis often do not align with biological distinction when using single sample outputs for the same signature. Moreover, significant signatures identified from pairwise analysis can still be poor predictive biomarkers of the clinical groups they were developed to represent.
To address these issues, we developed the dualGSEA, which provides users with a series of summary statistics and visual outputs enabling direct comparison of both the statistical significance and biological distinction in their data. This new dualGSEA tool has been made freely available via https://github.com/MolecularPathologyLab/Bull-et-al.
Methods
Datasets
The transcriptional dataset used was previously assembled for the development of the FDA-approved stage II ColDx/GeneFx risk-of-recurrence/relapse assay, consisting of n= 215 stage II primary tumours from CC patients profiled on the Almac disease-specific array, and available from ArrayExpress, accession number E-MTAB-86312. This cohort was chosen as it was assembled specifically for the development of a transcriptional classifier to distinguish between relapse and non-relapse cases and has been balanced in terms of the tumours clinical and pathological features, alongside technical factors such as tumour block age, randomisation of reagents. The cohort contained n = 73 tumours from patients that went on to develop distant metastasis within 5-year of surgery to remove the primary tumour (relapse) (R) and n = 142 tumours from patients that did not experience relapse within five years following surgery (non-relapse) (NR). The E-MTAB-863 CEL files were imported into Partek Genomics Suite (PGS; v6.6) and RMA normalised then log2 transformed. The probesets on the array were collapsed by importing the normalised data into R (v3.3.2 or later) and, using the ‘collapseRows’ function from WGCNA (Weighted Gene Co-expression Network Analysis, RRID: SCR_003302) package (v1.61)13, selecting the probeset with the highest mean expression per gene. An independent transcriptional non-cancer cohort (GSE213313), containing 94 samples was accessed and downloaded from GitHub14 to further confirm findings outside the cancer setting.
Differential gene expression analysis
Differential expression analysis (DEA) was performed to measure differentially expressed genes between R and NR CC. DEA was performed using the limma R package (v3.54.2). Following DEA, genes were ranked using three different metrics, (1) the t-statistic (t-stat), (2) the Log Fold Change (LogFC), and (3) the combination of LogFC and p-value (LogFC*-Log10(p-value); hereafter as “combined”). The addition of p-value to LogFC adds statistical significance to the directionality of LogFC. Separately, DEA was also performed for another comparison between tumours classified as PDS1 and PDS3, using the PDSclassifier package15 with resulting groups being assessed using the same metrics and thresholds applied to the R/NR analyses.
Pairwise analysis
To perform pairwise analysis two R packages were used, clusterProfiler5 (v4.6.2) and fgsea6 (v1.24.0) and a random seed of 127 was set. Biological pathways were investigated using the Hallmark gene sets from the MSigDB accessed through the msigdbr package (v7.5.1). Pre-ranked GSEA was first performed using the GSEA function in clusterProfiler with 1000 permutations (nPermSimple = 1000, minGSSize = 1, maxGSSize = Inf). Enrichment plots for GSEA were produced using the gseaplot2 function in the enrichplot R package (v1.18.4). GSEA was next conducted using the fgsea R package with the same parameters as clusterProfiler (nPermSimple = 1000, minSize = 1, maxSize = Inf). Enrichment plots of fGSEA were produced using the plotEnrichment function from the fgsea package. The online tool, GenePattern16, https://cloud.genepattern.org, was also used to perform a pre-ranked pairwise analysis, GSEAPreranked (v7.4.0). The Hallmark gene set collection was selected, ‘h.all.v2023.2.Hs.symbols.gmt’. Default parameters were set except for ‘collapse dataset’ which was set to ‘FALSE’. Normalised enrichment score (NES) and false discovery rate (FDR) values were recorded for each gene set within the two groups (R vs. NR; PDS1 vs. PDS3). A gene set with an FDR q-value below 0.05 was deemed significant.
Single sample analysis
To perform single sample analysis the R/Bioconductor package GSVA (v1.46.0) was used which facilitates ssGSEA9 and GSVA10. ssGSEA was performed with Hallmark4 gene sets from MSigDB2 and method set to “ssgsea”. GSVA was performed with Hallmark gene sets from MSigDB and the default parameters.
Single sample analysis heatmaps
For both ssGSEA and GSVA, matrix was formatted to include only Interferon Alpha Response, Interferon Gamma Response and Epithelial Mesenchymal Transition (EMT), as previously identified to be most significant by GSEA. The single sample scores were converted to Z-scores and were plotted using the ComplexHeatmap (v2.14.0) R package and were grouped using their respective groups (R vs. NR; PDS1 vs. PDS3).
Data visualisation
Additional visualisation R packages used for single sample analysis included: smplot2 (v 0.1.0), ggridges (v 0.5.4), easyGgplot2 (v 1.0.0.9000), pROC (v 1.18.5), randomForest (v 4.7–1.1) and, waterfalls (v 1.0.0).
Statistics
The statistical report was generated on RStudio (4.2.2). A Student’s t-test, from the stats (v 4.2.2) R package, was used to calculate significance of single sample scores between groups (NR compared to R and PDS1 compared to PDS3) and p-values were adjusted with Benjamini-Hochberg correction. The cor.test function from the stats (v 4.2.2) R package, with “pearson” method selected, was used for correlation analysis between single sample enrichment scores for selected significant gene sets. The cutpointr function in the cutpointr (v 1.1.2) R package was used to find the optimal cutpoint for the single sample scores. Once calculated the single sample scores were centred around the cutpoint resulting in a stratification of high and low scores for each of the gene sets being tested.
“dualGSEA” R function
The pairwise method, fGSEA6 and single sample method, ssGSEA9 have been combined to create an open source R-based function named “dualGSEA” available at: https://github.com/MolecularPathologyLab/Bull-et-al. The function enables the user to apply the above statistical analysis and visualisations between two groups-of-interest.
Results
Variations in differential gene expression outputs across a range of methods do not alter overall GSEA results
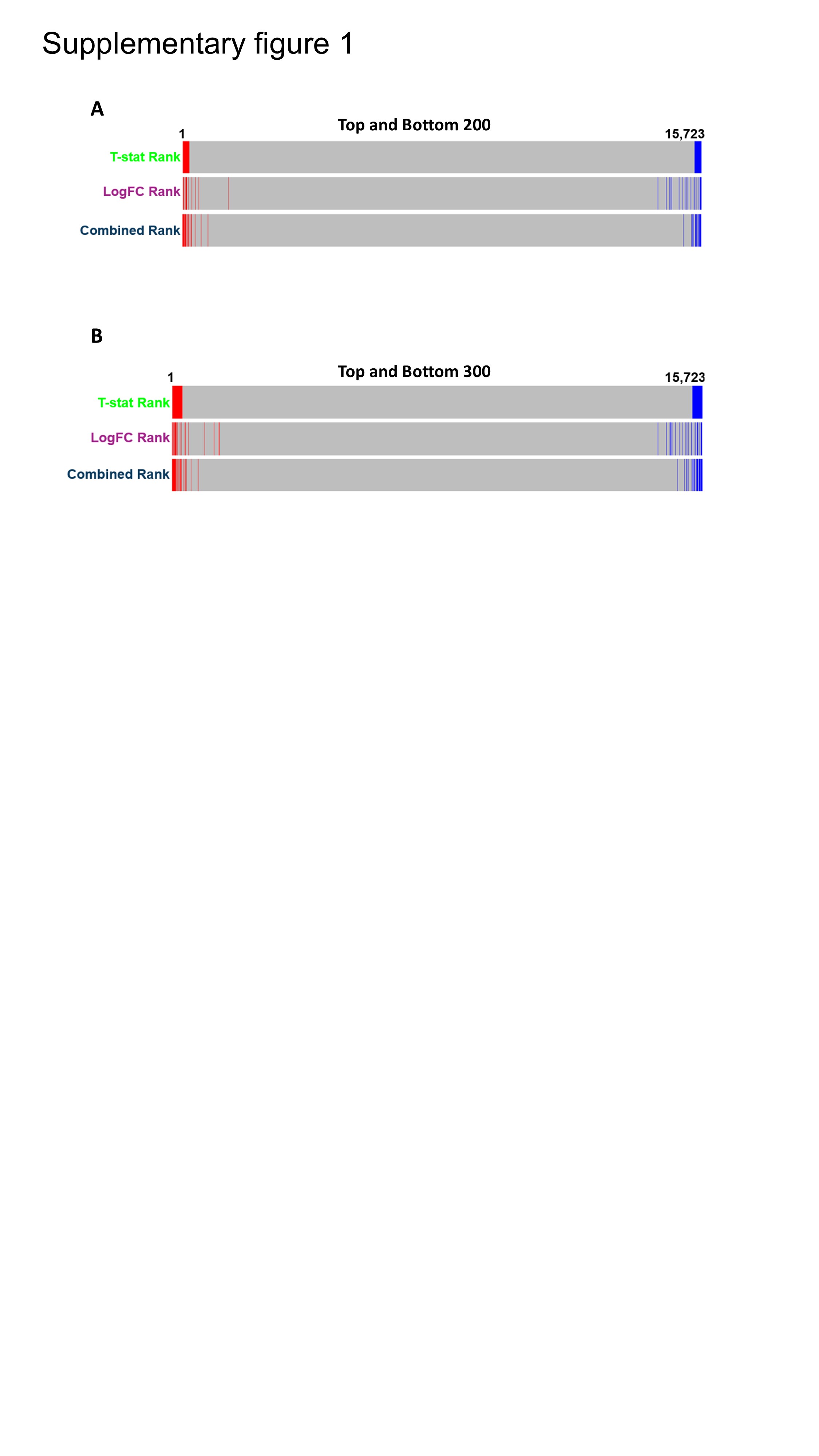
A typical goal when analysing bulk transcriptomic data, is the identification of discriminatory biological signalling cascades that can serve as biomarkers to distinguish between group(s)-of-interest; an output that can rapidly be delivered using transcriptional signatures in conjunction with in silico analytical tools, such as pairwise gene set enrichment analysis (GSEA)3 (Fig. 1A). The initial step in this GSEA process requires all genes in the expression matrix to be ranked based on their differential expression between the groups-of-interest. For example, when using limma17 for microarray or DESeq218 for RNA-seq, a ranked list of genes can be produced based on t-statistics (t-stat) or Log Fold Change (LogFC) values, both of which also provide directionality (up/down) according to the groups used. To assess the outputs from each ranking metric, we compared the ranked order of genes following the application of three approaches based on: (1) t-stat, (2) LogFC, and (3) the combination of LogFC and p-value (LogFC * -Log10(p-value); hereafter stated as combined) on expression profiles from n = 15,723 genes derived from n= 215 FFPE stage II colon cancer samples (E-MTAB-863)12, where patients whose cancer relapsed following surgery (R; n = 73) compared to those who did not (NR; n = 142) was used as an exemplar pairwise GSEA comparison (Fig. 1B). The use of distinct approaches in parallel within the same data/samples enables a direct comparison of both their statistical and biological outputs. Considering a series of the top and bottom 100, 200 or 300 genes ordered based on t-stat (0.6% of genes overall), based on LogFC, or the combined rank, remained remarkably stable. The top/bottom ranked genes identified using each method remain highly enriched at the extremes relative to t-stat ranking (Fig. 1B; Supplementary Fig. 1). When the genes were ranked by LogFC the majority (86%) of the top 100 genes fell within the top 500 genes when ranked by t-stat and the remaining were represented within the top 2,707 genes. With the combined rank, 100% of the top 100 genes were represented within the top 300 genes when ranked by t-stat.
Fig. 1.

Differential gene expression analysis and pairwise analysis of the discovery cohort. (A) Schematic of the differential expression analysis and pairwise analysis. (B) Workflow of differential expression analysis and ranked position of the top 100 differentially expressed genes and bottom 100 genes in NR when ranked by t-stat and the position of these genes when ranked by LogFC and combined. (C) Venn diagram of the significant Hallmark signatures (padj < 0.05) from GSEA when genes were ranked by t-stat, LogFC, and combined. (D) Significant Hallmark signatures (padj < 0.05) identified from clusterProfiler GSEA when genes were ranked by t-stat, LogFC, and LogFC combined with the p-value ordered by NES. (E) clusterProfiler GSEA, fGSEA, and GenePattern pre-ranked GSEA of the significant Hallmark gene sets. Enrichment plots from (F) clusterProfiler GSEA, (G) fGSEA, (H) GenePattern comparing NR CC (n = 142) to R CC (n = 73) for Interferon Alpha Response, Interferon Gamma Response and EMT.
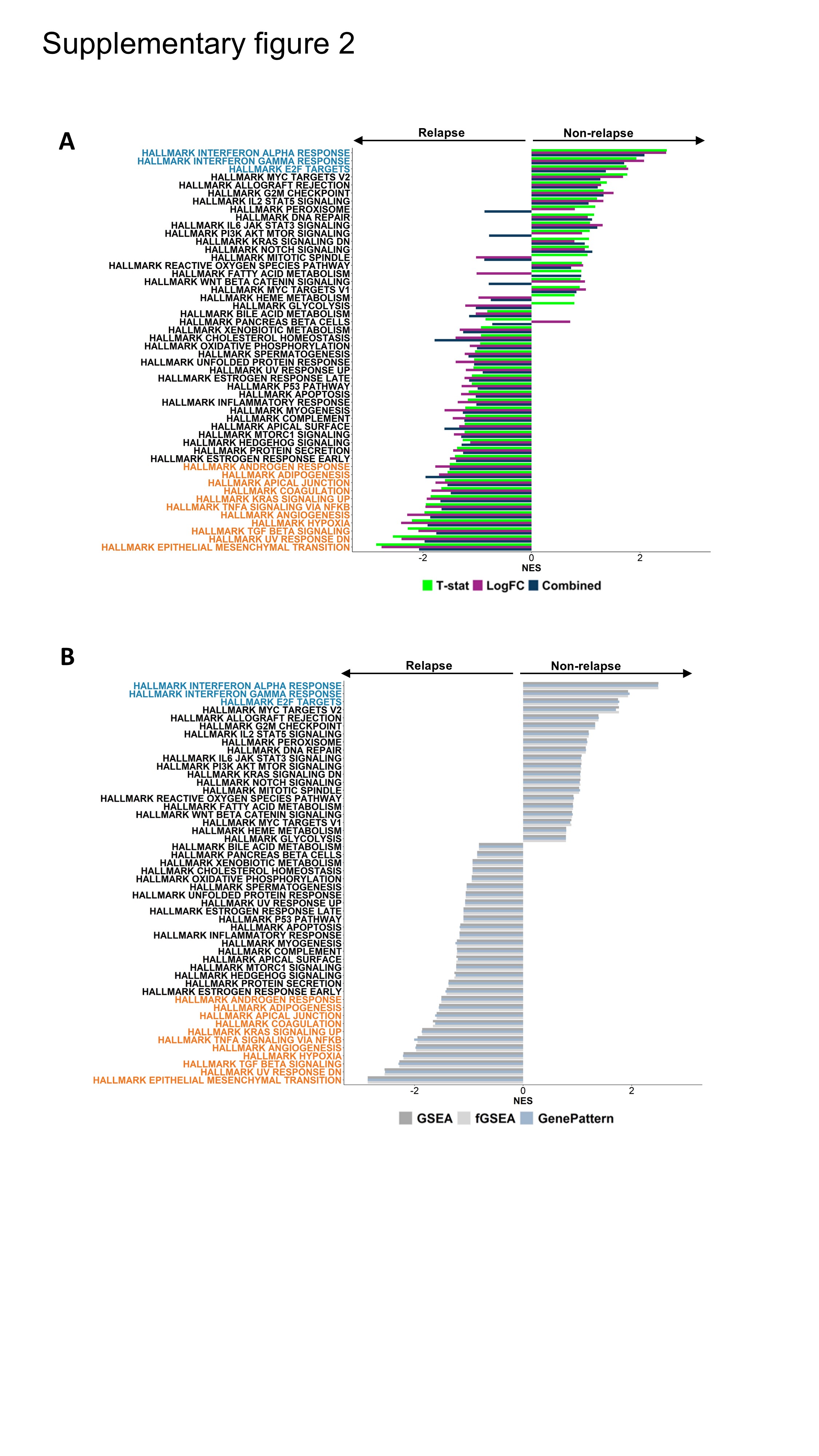
To test if there were more profound downstream consequences of these small pre-ranking gene order fluctuations, GSEA in clusterProfiler was performed5 using each of these ranking metrics on the n = 50 MSigDB ‘Hallmark’ gene sets. These analyses revealed that all three ranking methods resulted in remarkably consistent gene sets being returned as significant (FDR adjusted p-value < 0.05; t-stat = 16/50, LogFC = 21/50, combined = 15/50), n = 14 of the n = 22 total significant gene sets identified as common across from all three ranking methods (Fig. 1C; Supplementary Fig. 2A). When the normalised enrichment score (NES) is assessed to measure directionality, the direction of the n = 14 overlapping significant gene sets identified remained entirely consistent (Fig. 1D), meaning that regardless of the pre-ranking method used for these GSEA analyses, the biological interpretation will remain the same. Furthermore, when gene sets that were identified as significant by one method but not by the others, these were all enriched with the same directionality yet just below the statistical significance threshold: again, confirming the similarities in outputs for GSEA using all three pre-ranking methods (Supplementary Fig. 2A).
Pairwise GSEA methods provide results with consistent downstream interpretation
As there were minimal differences in the GSEA outcome with the three ranking methods, t-stat was used for the remainder of this study. Since the introduction of the original GSEA method, several updated methodologies have been developed and in this study we examined three derivatives of the GSEA method: (1) fast GSEA (fGSEA)6, (2) GSEA via clusterProfiler5 (as used in Fig. 1), which are both R-based tools, and (3) GSEA3 from the Broad Institute GenePattern16 Server. The GSEA tool from GenePattern performs standard GSEA with default signal-to-noise ‘GSEAPreranked’, where users can provide their own pre-ranked gene list prior to analysis. To test outputs from each of these GSEA methods, relapse (R) (n = 73) and non-relapse (NR) (n = 142) groups were compared across the CC cohort previously used (E-MTAB-863), where these methods consistently identify the same common statistically significant gene sets as identified in Fig. 1E, additionally the directionality of the NES for gene sets is consistent (Supplementary Fig. 2B). Between these three methods, we consistently observed that n = 3 gene sets were significantly upregulated in the NR group, including Interferon Alpha Response and Interferon Gamma Response, and n = 11 gene sets were upregulated in the R group, such as EMT (Fig. 1F–H); gene sets that have previously been associated with prognosis in multiple cancer types, including colorectal cancer19,20. Therefore, these three specific pathways were used in subsequent assessments.
Fig. 2.

Comparison of the single sample methods, ssGSEA and GSVA. (A) Schematic of standard single sample analysis workflow. (B) Scatterplot showing the correlation of ssGSEA and GSVA scores for the Hallmark Interferon Alpha Response (Pearson correlation coefficient, R = 0.835), Interferon Gamma Response (R = 0.878) and EMT (R = 0.953). (C) Heatmap of ssGSEA and GSVA scores for Interferon Alpha Response, Interferon Gamma Response and EMT comparing NR and R. Distribution of the ssGSEA and GSVA scores for the Interferon Alpha Response signature in the R (orange) and NR (blue) samples using (D) kernel density and (E) histograms. Distribution of the ssGSEA and GSVA scores for the Interferon Gamma Response signature in the R (orange) and NR (blue) samples using (F) kernel density and (G) histograms. Distribution of the ssGSEA and GSVA scores for the EMT signature in the R (orange) and NR (blue) samples using (H) kernel density and (I) histograms.
Single sample GSEA methods provide biological insights that may be masked when using pairwise GSEA alone
Single sample GSEA (ssGSEA)9 has been proposed as an extension of the GSEA method, one which can provide signature enrichment scores for each individual sample, rather than the summarised “average” scores within groups of samples provided by pairwise GSEA, making it suitable for both biological discovery and post-hoc assessments of individual samples within any established groups-of-interest21,22. Therefore, to compare the results obtained from GSEA (Fig. 1) with those from the single sample approaches, we explored two such methods: (1) ssGSEA9, and (2) gene set variation analysis (GSVA)10 within our discovery cohort (Fig. 2A). These two specific methods were chosen as they represent well described open-source tools for single sample assessments, enabling rapid pathway-level biological exploration within a range of transcriptomics data types. Using the top three significant gene sets identified in Fig. 1E, namely Interferon Alpha Response, Interferon Gamma Response and EMT, these single sample approaches were run using the GSVA R package by selecting either the “ssGSEA” or “gsva” method. A correlative analysis was performed between the resulting ssGSEA and GSVA scores, which revealed that both single sample methods were highly correlated, with a significantly positive correlation across all three gene sets (R > 0.8, p < 0.0001; Fig. 2B). These results suggest that while the algorithms are different, the output of either single sample methods provide consistent results.
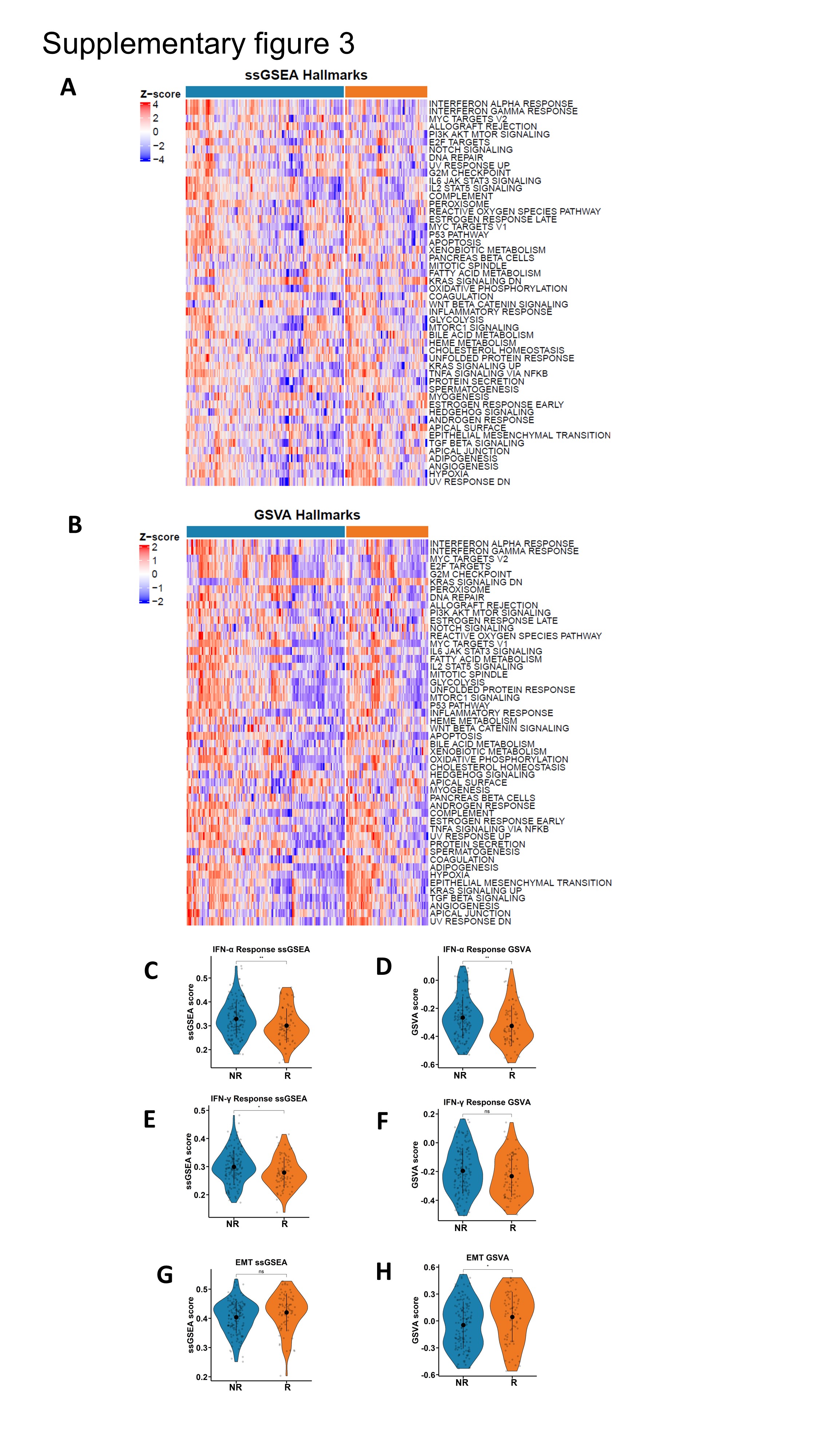
Assessment of the ssGSEA and GSVA scores for the three gene sets that were significantly different between the NR and R groups using GSEA, namely Interferon Alpha Response and Interferon Gamma Response and EMT, revealed that there were comparable quantities of high and low expression samples in each group, as indicated by the blue-to-red colours in the heatmap (Fig. 2C). To test this, a series of quantitative assessments were performed using scores for the significant signatures using GSEA. Although the two clinical groups may appear statistically significant for these single sample scores (Supplementary Fig. 3C-H), both clinical groups fall under the same distribution scale (Fig. 2D–I), thus implying they are not biologically distinct for the signatures, which contradicts with GSEA output. The range of ssGSEA scores showed large overlap between R and NR samples, Interferon Alpha Response had 95.3% overlap between R and NR, Interferon Gamma Response had 97.7% overlap between R and NR and EMT had 99.1% overlap between R and NR. With respect to the GSVA results, Interferon Alpha Response scores had 95.8% overlap between R and NR, Interferon Gamma Response had 98.1% overlap and EMT had 98.6% between R and NR. Overall, these data highlight how even the most statistically significant pairwise GSEA results may not be sufficient to identify transcriptional signalling that is discriminatory between samples across two tumour groups.
Visualisation of ssGSEA score is essential to ensure that statistical significance between sample groups also represents distinct biology
There are a range of biomarker performance metrics that can be used to objectively test and enumerate how well individual signatures represent the signalling within different groups of samples. Therefore, a series of analyses were conducted to test the predictive value of the most significant signatures identified by pairwise GSEA approaches (n = 3) in identifying the specific groups-of-interest that they were enriched in. We performed receiver operating characteristic (ROC) analysis with the ssGSEA/GSVA scores and examined the area under the curve (AUC). NR patients displayed statistically significant enrichment in Interferon Alpha and Interferon Gamma Response, implying that these signatures are contributing factors to favourable outcome in NR patients (Supplementary Fig. 3C–E), albeit GSVA Interferon Gamma Response did not show any statistically significant enrichment in the NR samples (Supplementary Fig. 3F). However, if both interferon response signatures were then to be used to develop a risk stratification tool to predict patient relapse status, the models developed based on these signatures would perform underwhelmingly with the AUC approximately ranging between 0.57 and 0.62 (Fig. 3A and C). Furthermore, although there are more NR (n = 142) than R cases (n = 73), when stratified into high and low groups for the Interferon Alpha and Interferon Gamma Response signature scores using both ssGSEA and GSVA, based on the optimal cut-offs defined by the AUROC analyses, ~ 30–50% of relapse patients have high Interferon Alpha and Interferon Gamma Response scores (Fig. 3B and D). Likewise, regardless of its statistical significance (Supplementary Fig. 3G, H), the EMT ssGSEA and GSVA scores also perform poorly (AUC 0.60), with low sensitivity and specificity as a relapse-specific biological signature for the purpose of risk stratification (Fig. 3E, F).
Taken together, while each of these three signatures have been repeatedly shown to provide statistical significance in terms of association with relapse outcomes, this is primarily due to small (albeit statistically significant) differences in sample distributions, meaning that the biological signalling these signatures are based on cannot be interpreted as reflecting distinct mechanistic phenotypes or biological cascades between the two groups-of-interest.
Fig 3.

Application of single sample analysis as a predictor for relapse. (A) ROC curve using Interferon Alpha Response ssGSEA and GSVA scores to predict NR had an AUC ranging between 0.61 and 0.62. True positive rate is when the sample is classified as high Interferon Alpha Response, and the case was a NR. The true negative rate is the proportion of true negatives, when a sample is a NR cases without high Interferon Alpha Response. (B) Interferon Alpha Response waterfall plots show when stratified into high and low groups for the Interferon Alpha Response with both ssGSEA and GSVA scores, we found a greater number of NR patients classed as high (n = 98 [75.4%], n = 92 [76.0%]) respectively compared to R (n = 32 [24.6%], n = 29 [24.0%]) respectively. (C) Interferon Gamma Response ROC AUC values ranging between 0.57 and 0.61. True positive rate is when the sample is classified as high Interferon Gamma Response, and the case was a NR. The true negative rate is the proportion of true negatives, when a sample is a NR cases with a low Interferon Gamma Response score. (D) Interferon Gamma Response waterfall plots show when stratified into high and low groups for the Interferon Gamma Response with both ssGSEA and GSVA scores, there were greater number of NR patients classed as high (n = 86 [76.1%]; n = 95 [71.4%] respectively compared to R (n = 27 [23.9%]; n = 38 [28.6%] respectively. (E) EMT ROC AUC values of 0.60. True positive rate is when the sample is classified as high EMT, and the case was a relapse. The true negative rate is the proportion of true negatives, when a sample is a NR case with a low EMT score. (F) EMT waterfall plots show when stratified into high and low for EMT for ssGSEA we found that a greater number of R patients classified as high (n = 22 [59.5%], compared to NR (n = 15 [40.5%]). GSVA found a higher number of NR patients with a high EMT score (n = 50 [56.2%]) compared to R (n = 39 [43.8%]).
Pathway-derived subtype serves as an exemplar for performing biological discovery using a single sample approach
As shown above, pairwise methods comparing relapse and non-relapse tumours can provide users with statistically significant results, however these clinically distinct groups do not represent uniform, biologically distinct transcriptional subtypes. Therefore, to test the performance of pairwise and single samples GSEA methodologies in groups of samples that represent biologically distinct entities, we next performed these analyses contrasting tumours based on our recent pathway-derived subtypes (PDS)15 which identified three statistically and biologically distinct subtypes; PDS1-3.
In this current study we now segregate our transcriptional cohort into these three PDS classes (this dataset was not used in the original study) and perform a series of GSEA/ssGSEA assessments on PDS1 (characterised by high MYC signalling) and PDS3 (characterised by low MYC signalling) in conjunction with the performance metrics and visualisations used so far (Fig. 4A). Comparative analysis using the Hallmark gene sets collection and pairwise GSEA, similar to the relapse-based comparisons, highlights a highly significant statistical difference between PDS1 and PDS3 for MYC Targets V1 gene set (hereafter MYC V1; Fig. 4B). Importantly, unlike the assessment on R versus NR in the same cohort (Figs. 1, 2 and 3), comparison of PDS1 to PDS3 clearly shows both statistical significance and biological distinction when using single sample approaches (Fig. 4C). Most importantly, unlike our earlier analyses based on GSEA results comparing R and NR samples, these new assessments across a known biology, reveal a remarkable difference and minimal overlapping distribution for MYC V1 ssGSEA score, with only 6.7% of ssGSEA scores overlapping between PDS1 and PDS3 (Fig. 4D, E), implying that PDS1 and PDS3 can be considered as representing truly distinct biological groups for MYC V1. This is further confirmed using ROC analysis, from both ssGSEA and GSVA MYC V1 scores, which proves a sample will be classified as high MYC V1 when the sample is PDS1 with an AUROC = 0.99 (Fig. 4F, G).
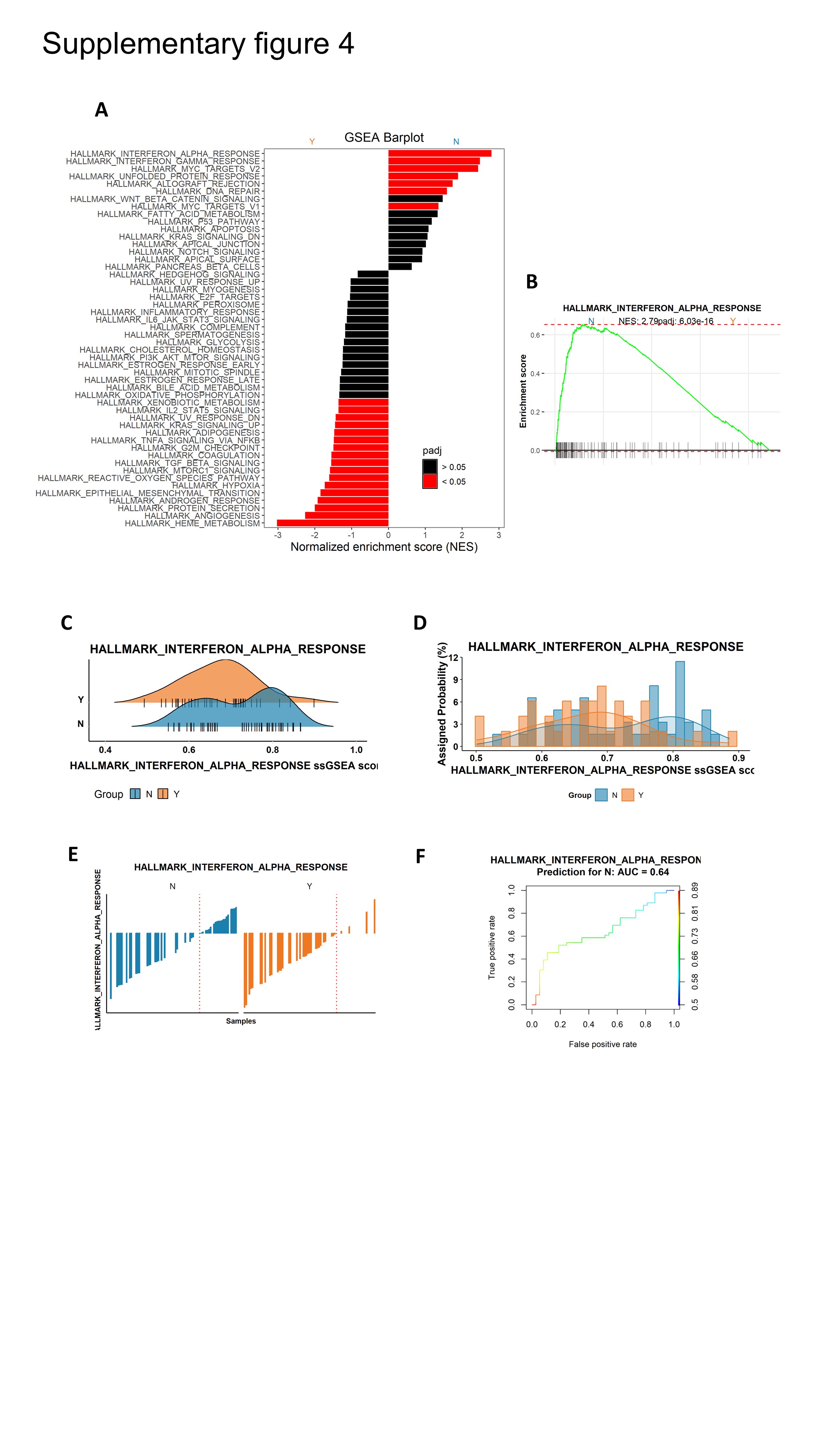
To allow all users to perform these pairwise and single sample approaches in tandem, we have created an open source parallel pairwise/single sample R-based function named “dualGSEA” which is available at: https://github.com/MolecularPathologyLab/Bull-et-al. The function produces multiple visualisations and statistical analysis options that enables users to perform a broad characterisation of their samples and groups-of-interest (Fig. 4H). To provide a wider context for the application of “dualGSEA”, analysis was also performed in an independent non-cancer transcriptional data set (Supplementary Fig. 4), which further confirmed its applicability across different disease settings.
Fig. 4.

The use of single sample analysis provides distinct biology between groups. (A) Schematic of application of pathway analysis methods when applied to PDS classification. (B) GSEA revealed MYC targets V1 is enriched in the PDS1 group compared to the PDS3 group. (C) ssGSEA scores show significant difference of MYC targets V1 between PDS1 and PDS3 groups (****padj < 0.0001). (D & E) ssGSEA scores for PDS1 and PDS3 show little overlap of MYC targets V1 between groups. (F) ROC curve shows that the MYC V1 scores enable discrimination between PDS1 and PDS3. AUC value of 0.99. True positive rate is when the sample is classified as high MYC V1, and the case was PDS1. The true negative rate is the proportion of true negatives, when a sample is a PDS1 without high MYC V1. (G) Stratification of MYC V1 high and MYC V1 low ssGSEA scores showed that PDS1 was classified as high MYC V1 (n = 54 [96.4%] and PDS3 contained only samples with a low MYC V1 score (n = 63 [100%]). (H) Overall schematic for dualGSEA workflow.
Discussion
In this study, we initially set out to provide a comparison of several well-established gene set enrichment analysis (GSEA) methods, with particular emphasis on how choices of standard bioinformatic pipelines can lead to differences in downstream biological interpretation. As an exemplar of this, we assessed how consistent a significant pairwise GSEA result is between pairwise approaches and also when the same signature is assessed using single sample GSEA methods. These analyses highlight concordance within pairwise or single sample approaches, however despite similar statistical performance, data presented here provides a clear indication for how vastly different downstream interpretation of results can be derived when using pairwise or single sample methods for the same transcriptional signatures. Pairwise methods provide the user with strong statistical-based evidence of differences in signature expression between two selected groups of samples, however this can result in confusion when interpreting the biological significance of these differences, as illustrated by enrichment scores across individual samples strongly overlapping between and within groups.
These results strongly support the use of single sample methods for class discovery and mechanistic biomarker development/testing, given their consistency and robustness in identifying distinct biological signalling between defined groups of samples. Many previous studies have focussed on the statistical advantages and limitations of GSEA methods, providing the field with important information on performance metrics for each algorithm7. While these algorithms were developed to identify statistical significance between user-selected groups of samples, they can occasionally be interpreted as representing biologically distinct groups; a point that becomes even more important if the results from GSEA-based methods are used to guide development of new pre-clinical models that are interpreted as faithfully representing the clinical group-of-interest, or used as the basis of developing prognostic/predictive biomarkers to guide clinical decision-making.
Data presented in this paper does not challenge the importance of studies using GSEA methods, as we clearly demonstrate their value in identifying robust statistically distinct groups. Our current study aims to provide an example of the consequence of method selection for biological end-users with a primary interest in using these tools to identify biologically distinct mechanistic signalling between two groups. For such end-users, we propose that emphasis should be placed on more widespread use of visualisation methods at an individual sample resolution, rather than the use of statistical values alone, to ensure there is a clear distinction between the groups being compared23. This point is particularly important for biomarker discovery, where there is a requirement for the most robust and discriminatory features that can be used to predict tumour groups with high sensitivity and specificity. In addition, the identification of representative biological cascades that are both statistically significant and biologically distinct between the two groups across a cohort of tumours is increasingly important in the era of precision medicine, where interrogation of transcriptional data can be used as the basis for development of pre-clinical model systems and testing of subtype-specific therapeutic targets aimed at these patient groups.
An important feature for performing pairwise GSEA is the ranking of differentially expressed genes. Our analyses highlight that the positions of individual differentially expressed genes in an overall list will vary when using different ranking options. These results provide a clear example of how the use of some of the most widely accepted tools for differential gene expression analyses can lead to different users identifying conflicting biomarkers for the same phenotypes in the exact same datasets. However, we find that the effects on using different pre-ranking methods to rank genes for pairwise approaches have minimal effects on biological interpretation when using downstream pathway analyses with any GSEA method. As such, these data again support the use of pathway-level gene signatures as a more representative way of measuring true biological phenotypes in transcriptional data, over the use of individual gene-level biomarkers that can be undermined by technical biases inherent in method choices for gene ranking. This single sample approach was used as basis for class discovery within our recent pathway-derived subtypes (PDS)15 study, which used ssGSEA scores to identify three biologically distinct classes of colorectal cancer that was found to have prognostic value.
The cancer research field is accustomed to the heavy reliance on statistical thresholds as the primary criteria for significance, as they provide users with a quantitative reference in support of their findings. In data presented here we clearly show that additional visualisation of these same data can lead to questions over the true biological significance of such results. In this setting, if GSEA tools were used for discovery, the biological signalling used as the basis for mechanistic studies could be indistinguishable across samples from these different clinical groups, despite such signalling being based on statistically sound evidence. Moving forward, it is essential to find a balance between statistical significance and biological relevance, utilising visualisation techniques and analysis methods, including distribution plots and ROC curves, to validate and contextualise findings.
To ensure users can recapitulate the approaches used here, we have developed an open source parallel pairwise/single sample R-based function, “dualGSEA” https://github.com/MolecularPathologyLab/Bull-et-al, which provides multiple data visualisation outputs and statistical tests, enabling all users to perform a comprehensive assessment of their samples and groups-of-interest as shown in the comparison of PDS1 vs. PDS3 (Fig. 4H).
Overall, our study sheds new light on the nuances between established gene set enrichment methods, highlighting the challenges in interpreting results across different methods, particularly when these data are being used to align with pre-clinical models. The work presented illustrates how a highly significant pairwise result does not always translate to a significant single sample result when the same transcriptional data is analysed using the same gene signatures. By carefully navigating these methods and their implications, researchers can uncover novel meaningful biological insights from transcriptional data.
Electronic supplementary material
Below is the link to the electronic supplementary material.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Acknowledgements
This work was supported by a CRUK early detection grant (A29834), a CRUK International accelerator programme, ACRCelerate, (A26825), a UK Medical Research Council (MRC) National Mouse Genetics Network programme (MC_PC_21042).
Author contributions
Author Contributions: CB: data analysis, data visualisation, writing-original draft, writing-review and editing, RB: writing-review and editing, NCF: writing-review and editing, SMC: writing-review and editing, RA: writing-review and editing, JE: writing-review and editing, LVSH: writing-review and editing, ML: writing-review and editing, AER: writing-review and editing, FL: data analysis, writing-review and editing, PDD: conceptualisation, resources, supervision, writing-original draft, writing-review and editing, SBM: supervision, writing-original draft, writing-review and editing.
Data availability
Data is available in a public, open access repository. The “dualGSEA” scripts used in this current study are publicly available at https://github.com/MolecularPathologyLab/Bull-et-al.
Declarations
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Philip D. Dunne and Sudhir B. Malla.
Contributor Information
Philip D. Dunne, Email: p.dunne@qub.ac.uk
Sudhir B. Malla, Email: s.malla@qub.ac.uk
References
- 1.Wilkinson, M. D. et al. The FAIR guiding principles for scientific data management and stewardship. Sci. Data3, 160018. 10.1038/sdata.2016.18 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Liberzon, A. et al. Molecular signatures database (MSigDB) 3.0. Bioinformatics27, 1739–1740. 10.1093/bioinformatics/btr260 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Subramanian, A. et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci.102, 15545–15550. 10.1073/pnas.0506580102 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Liberzon, A. et al. The molecular signatures database (MSigDB) hallmark gene set collection. Cell. Syst.1, 417–425. 10.1016/j.cels.2015.12.004 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wu, T. et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. The Innovation2, 100141. 10.1016/j.xinn.2021.100141 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Korotkevich, G. et al. Fast gene set enrichment analysis. bioRxiv 060012. 10.1101/060012 (2021).
- 7.Tarca, A. L., Bhatti, G. & Romero, R. A. Comparison of gene set analysis methods in terms of sensitivity, prioritization and specificity. PLOS ONE. 8, e79217. 10.1371/journal.pone.0079217 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Maleki, F., Ovens, K., Hogan, D. J. & Kusalik, A. J. Gene set analysis: Challenges, opportunities, and future research. Front. Genet.11, 654. 10.3389/fgene.2020.00654 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Barbie, D. A. et al. Systematic RNA interference reveals that oncogenic KRAS-driven cancers require TBK1. Nature462, 108–112. 10.1038/nature08460 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hänzelmann, S., Castelo, R. & Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-Seq data. BMC Bioinform.14, 7. 10.1186/1471-2105-14-7 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chang, L. C., Lin, H. M., Sibille, E. & Tseng, G. C. Meta-analysis methods for combining multiple expression profiles: Comparisons, statistical characterization and an application guideline. BMC Bioinform.14, 368. 10.1186/1471-2105-14-368 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kennedy, R. D. et al. Development and independent validation of a prognostic assay for stage II colon cancer using formalin-fixed paraffin-embedded tissue. J. Clin. Oncol.29, 4620–4626. 10.1200/JCO.2011.35.4498 (2011). [DOI] [PubMed] [Google Scholar]
- 13.Langfelder, P. & Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform.9, 559. 10.1186/1471-2105-9-559 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Prebensen, C. et al. Longitudinal whole blood transcriptomic analysis characterizes neutrophil activation and interferon signaling in moderate and severe COVID-19. Sci. Rep.13, 10368. 10.1038/s41598-023-37606-y (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Malla, S. B. et al. Pathway level subtyping identifies a slow-cycling biological phenotype associated with poor clinical outcomes in colorectal cancer. Nat. Genet.10.1038/s41588-024-01654-5 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Reich, M. et al. GenePattern 2.0. Nat. Genet.38, 500–501. 10.1038/ng0506-500 (2006). [DOI] [PubMed] [Google Scholar]
- 17.Ritchie, M. E. et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res.43, e47–e47. 10.1093/nar/gkv007 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Love, M. I., Huber, W. & Anders, S. Moderated estimation of Fold change and dispersion for RNA-seq data with DESeq2. Genome Biol.15, 550. 10.1186/s13059-014-0550-8 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Guinney, J. et al. The consensus molecular subtypes of colorectal cancer. Nat. Med.21, 1350–1356. 10.1038/nm.3967 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Corry, S. M. et al. Activation of innate-adaptive immune machinery by poly(I:C) exposes a therapeutic vulnerability to prevent relapse in stroma-rich colon cancer. Gut71, 2502–2517. 10.1136/gutjnl-2021-326183 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wu, S. et al. Integrated machine learning and single-sample gene set enrichment analysis identifies a TGF-Beta signaling pathway derived score in headneck squamous cell carcinoma. J. Oncol.2022, 3140263. 10.1155/2022/3140263 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yi, M., Nissley, D. V., McCormick, F. & Stephens, R. M. ssGSEA score-based Ras dependency indexes derived from gene expression data reveal potential Ras addiction mechanisms with possible clinical implications. Sci. Rep.10, 10258. 10.1038/s41598-020-66986-8 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yanai, I. & Lercher, M. A hypothesis is a liability. Genome Biol.21, 231. 10.1186/s13059-020-02133-w (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data is available in a public, open access repository. The “dualGSEA” scripts used in this current study are publicly available at https://github.com/MolecularPathologyLab/Bull-et-al.