

ABSTRACT
Migratory species typically undertake demanding long‐distance journeys, across different habitat types during which they are exposed to multiple natural and anthropogenic stressors. Mortality during migration is typically high and may be human induced. Understanding individual responses to these selection pressures is rarely attempted because of the challenges of relating individual phenotypic and genetic data to migration success. Here, we show distinct single nucleotide polymorphism (SNP) sets significantly differentiated between Atlantic salmon smolts making successful migrations to sea and those that failed to migrate, in two different rivers. In contrast, morphological variation was not diagnostic of migration success. Populations from each river were genetically distinct, and while different genes were possibly implicated in migration success in each river, they related to common biological processes (e.g., osmoregulation and immune and stress response). Given that migration failure should quickly purge polymorphism at selected SNPs from a population, the question of how genetic diversity in these populations is maintained is an important one. Standing genetic variation could be maintained by different life history strategies and/or environmentally driven balancing selection. Our work highlights the importance of preserving genetic diversity to ensure evolutionary resilience at the population level and has practical implications for management.
Keywords: animal migration, genome scan, geometric morphometrics, salmon, SNPs, telemetry
Understanding individual responses to selective pressures during animal migration is rarely attempted because of the challenges of relating individual phenotypic and genetic data to migration success. Here, we show distinct single nucleotide polymorphism (SNP) sets significantly differentiated between Atlantic salmon smolts making successful migrations to sea and those that failed to migrate in two different rivers, whereas morphological variation was not diagnostic of migration success. Given that migration failure should quickly purge polymorphism at selected SNPs from a population, the question of how genetic diversity in these populations is maintained is discussed.
1. Introduction
Animal migration has evolved independently many times across the animal kingdom (Dingle and Drake 2007; Bowlin et al. 2010; Shaw 2016). Migration events can involve large numbers of individuals moving between different habitats and regions, and these events play a key ecological and socioeconomic role in natural and human communities (Bauer and Hoye 2014). Migratory species typically rely on multiple habitats to complete their life cycle and often undertake demanding long‐distance journeys exposing themselves to numerous natural and anthropogenic stressors, such as predation, adverse weather conditions, pathogens, pollution, artificial constructions and harvesting (Alerstam, Hedenström, and Åkesson 2003). Mortality during migration is typically high and can be exacerbated by human‐induced pressures, such that it impacts migratory populations and the ecosystems that depend on them (Wilcove and Wikelski 2008; Harris et al. 2009; Middleton et al. 2013; Klaassen et al. 2014; Baker et al. 2020). With an ongoing global decline in migratory species (Wilcove and Wikelski 2008), a better understanding of the factors causing mortality in migration is urgently required to predict responses of migratory populations to future environmental challenges and implement incisive conservation actions.
Recent advances in telemetry technology have made it possible to investigate migratory behaviours of species both temporally and spatially (Doherty et al. 2017; Thorup et al. 2023). This has enabled a better understanding of the exogenous factors directly influencing migration mortality (Thorstad et al. 2013; Hays et al. 2003; Palacín et al. 2017; Weinz et al. 2020). Organisms require a suite of specific morphological, physiological and behavioural adaptive features to successfully complete a migratory cycle (Justen and Delmore 2022). Given the phenotypic and genetic variation found in most populations, it is reasonable to expect that some genetic or phenotypic traits are more likely to increase migration success than others. However, which traits these might be remains poorly understood.
Genomic tools have recently been applied to identify factors regulating migratory behaviour at the population or species level. Several studies have discovered the genetic basis for migratory features such as migration timing and distance, orientation and propensity to migrate, with specific genomic regions linked to these traits (Zhu et al. 2009; Liedvogel, Åkesson, and Bensch 2011; Hecht et al. 2012, 2015; O'Malley et al. 2013; Hess et al. 2014; Pritchard et al. 2018; Waples, Naish, and Primmer 2020; Justen and Delmore 2022). However, understanding the genomic basis on which selection could act at an individual level to dictate migration success has rarely been attempted (but see Bourret, Dionne, and Bernatchez 2014), despite the fundamental insights it could provide into how populations might respond to selection, and the implications for conservation genetics of migratory species.
An important phenotypic trait expected to influence migration success is body morphology (Minias et al. 2013). Morphological variation (i.e., body shape and size) can affect behaviour, resource use, survival and reproductive success of individuals (Wainwright 1994; Skulason and Smith 1995; Fruciano, Tigano, and Ferrito 2011). The effect of morphology on movement is particularly evident in fish because of a direct link to swimming performance (Pakkasmaa and Piironen 2001; Fisher and Hogan 2007; Drinan et al. 2012; Stelkens et al. 2012; Páez and Dodson 2017). Chapman et al. (2015) found a direct correlation between migration propensity and body shape, while other studies have demonstrated an increased ability and ‘motivation’ to pass river barriers in relation to size, fat content and morphology (Newton et al. 2018; Lothian et al. 2020; Goerig et al. 2020). Nevertheless, research on migration survival and mortality as a consequence of body shape variation (as opposed to size; Kennedy, Gale, and Ostrand 2007; Hostetter et al. 2012; Romer et al. 2013; Furey et al. 2016; Lilly et al. 2022) is still lacking.
Atlantic salmon ( Salmo salar Linnaeus) is a migratory species of socioeconomic importance that has suffered substantial declines over the past 40 years (ICES 2024) due to multiple abiotic and biotic factors not yet fully understood (Forseth et al. 2017; Dadswell et al. 2022). The Atlantic salmon has a complex life cycle, which includes two long‐distance migration stages; a feeding migration from freshwater to sea as a juvenile (smolt) and an adult returning spawning migration from sea to freshwater. In addition, it is a philopatric species, accurately homing to its natal spawning grounds (Thorstad et al. 2010). Fidelity to a specific river limits gene flow among populations and has been shown to promote the evolution of local adaption through natural selection, genetic drift and bottlenecks (Garcia De Leaniz et al. 2007; Fraser et al. 2011). The seaward migration of smolts constitutes a key life stage for Atlantic salmon, often characterised by high mortality rates (Thorstad et al. 2012; Lothian et al. 2018) and provides an ideal opportunity to study the genetic and phenotypic components that may differentially affect the ability of individual animals to successfully complete their migration. The identification of genetic and phenotypic traits could play a vital role in local management of Atlantic salmon (Bernos, Jeffries, and Mandrak 2020).
Here, we analysed genomic and morphological data of migrating Atlantic salmon smolts in two rivers. We test to what extent (I) Atlantic salmon populations in the two rivers were genetically distinct, and (II) migration success by seaward migrating smolts could be predicted by specific genomic regions and/or morphological traits.
2. Methods
2.1. Sampling, Tagging and Study Design
The study reported here formed part of a wider acoustic telemetry study to examine migratory behaviours and migration success in juvenile Atlantic salmon (smolts) on their first migration from natal rivers to sea (see Whelan, Roberts, and Gray 2019). Atlantic salmon were captured between 11 April and 3 May 2019 from the rivers Oykel (57°59.640′ N, 4°48.282′ W) and Spey (57°24.960′ N, 3°22.602′ W), Scotland, using 1.5‐m‐diameter rotary screw traps and a box trap (Figure 1). Fish were anaesthetised in MS222 and tagged with Vemco V7‐2L acoustic transmitters (7 mm diameter, 19.5 mm length, 1.5 g in air, 137 dB re 1 μPa @ 1 m, acoustic transmission repeat cycle of 28 ± 10 s, InnovaSea, Bedford, Nova Scotia, Canada). For more details on the tagging and release procedure, see Lilly et al. (2022). Before being tagged, fish were visually inspected to confirm they were in the smolt stage, characterised by silvery colouration and an elongated body shape. Fish were measured (fork length, mm), weighed (g) and photographed. Photographs of the left side of each fish were taken from approximately 30 cm directly above the fish, with a Fujifilm FinePix XP130 Compact Digital Camera on a background reference scale. An adipose fin clip was also taken from every fish and stored in 96% ethanol for later DNA extraction.
FIGURE 1.
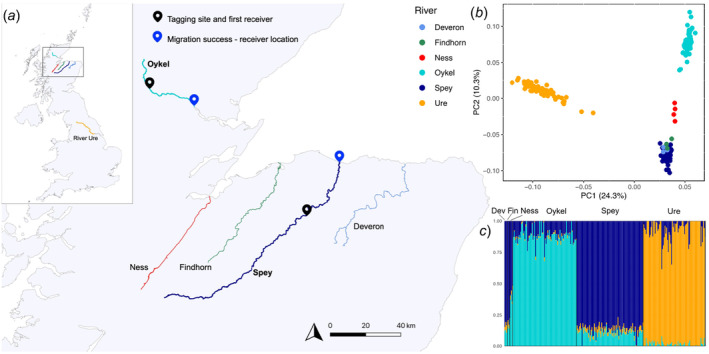
The study area (a) and genetic structuring (b, c) of Atlantic salmon populations from rivers flowing into the Moray Firth (Scotland), with samples from the River Ure (England, (a) top left panel) for comparison. Tagging locations and acoustic receivers are shown in map (a). The principal components analysis (PCA) and the ADMIXTURE analysis plots are based on 44,504 SNPs pruned for linkage disequilibrium. In the PCA scatterplot (b), dots represent individual fish, and variance (%) explained by the first and second axes are shown. Colours correspond to rivers. In the ADMIXTURE plot (K = 3; c), each fish individual is represented by a vertical bar. ‘Dev’ and ‘Fin’ are abbreviations for the Rivers Deveron and Findhorn respectively.
Two acoustic monitoring receivers (InnovaSea VR2Tx) were deployed in each river, one of which was immediately downstream of the tagging site (0.2 and 0.6 km in the rivers Oykel and Spey, respectively; Figure 1). The second receiver was deployed at the river mouth (Figure 1). Of all the salmon tagged and released in the two rivers (Oykel, n = 149, and Spey n = 150), 91.9 and 96.7%, respectively, were detected by the first receiver after release. Of these, 78 (Oykel) and 82 (Spey) smolts were randomly selected for this study and distributed evenly between migratory outcomes ensuring a balanced design. Fish from both rivers were allocated into two groups based on their migratory outcome: (1) fish detected on the second and final river receiver were categorised as ‘successful’ river migrants, and (2) fish only detected on the first receiver were considered as ‘unsuccessful’ river migrants (Table 1, Figure 1). To assess receiver detection efficiency, additional receivers deployed as a part of the broader telemetry study in the marine coastal waters of the Moray Firth were used. Since all smolts detected in marine waters were also detected by the two freshwater receivers, detection efficiency was determined to be 100%, meaning that no fish were wrongly miscategorised as unsuccessful migrants as a result of missed detections at the second river receiver.
TABLE 1.
Classification of smolts (n) in the Rivers Oykel and Spey based on tracking results.
| River | Unsuccessful | Successful | Total river migration distance |
|---|---|---|---|
| Oykel | 36 | 42 | 30.5 km |
| Spey | 35 | 47 | 50.1 km |
To assess whether the study rivers harboured genetically differentiated Atlantic salmon populations, the genetic variation across rivers in the Moray Firth (Figure 1) was investigated. In addition to fish from the study Rivers Oykel and Spey, Atlantic salmon smolts from the Rivers Findhorn (n = 3; 57°25.05′ N, 3°53.35′ W), Deveron (n = 4; 57°30.45′ N, 2°42.35′ W) and Ness (n = 4; 57°27.17′ N, 4°15.35′ W) were included in the analysis. To further contextualise the relative genetic diversity of these rivers, Atlantic salmon samples from the River Ure, England (n = 76; 54°16.19′, N 1°44.57′ W), were also included in the analysis (Figure 1). Fish from the Findhorn, Deveron and Ness were sampled in Spring 2019 using rotary screw traps, while fish from the Ure were captured employing backpack electric fishing equipment (Electracatch 24VDC input, 200–400 V, 100 W, 50 Hz Pulsed DC, variable pulse width output).
2.2. Genomic Analyses
2.2.1. DNA Extraction, Genotyping and Quality Control (QC)
DNA was extracted from adipose fin samples employing a modified Mu‐DNA: Tissue protocol (Sellers et al. 2018) using a solid‐phase reversible immobilisation (SPRI) magnetic bead capture method (adapted from Rohland and Reich 2012) to isolate high‐molecular‐weight DNA. The DNA samples were sent to the Centre for Integrative Genetics (CIGENE, Ås, Norway) for genotyping, including biological and technical replicates to ensure consistency across plates. A custom 220,000 SNP (single nucleotide polymorphism) Affymetrix Axiom array designed for Atlantic salmon (see Barson et al. 2015 for details) was used for data generation. Following the manufacturer's instructions, only SNPs categorised as PolyHighResolution and NoMinorHom were used for analyses, while SNPs with unknown position were excluded from the dataset, leaving 213,945 available loci for genomic investigation. We then performed QC and filtering of SNP data in PLINK versions 1.9 and 2.0 (www.cog‐genomics.org/plink/1.9/ and www.cog‐genomics.org/plink/2.0/; Purcell et al. 2007; Chang et al. 2015). SNPs were filtered for Hardy–Weinberg equilibrium (PLINK 1.9. command: ‐‐hwe 0.001) to remove genotyping errors. Additionally, SNPs were screened for minor allele frequency (‐‐maf 0.05) and genotype missingness (‐‐geno 0.1), and individuals with a high rate of missing SNPs (‐‐mind 0.1) were discarded from analyses. Full siblings were also removed using PLINK 2.0 (‐‐king‐cutoff 0.25). In the migration success analyses, these QC steps were performed separately for the Rivers Oykel and Spey and resulted in the retention of 198,336 SNPs and 82 individual fish from the River Oykel and 201,475 SNPs and 78 individuals from the River Spey. The fish used in this analysis were the same employed for morphometric investigations but included two additional individuals which were not photographed. For the regional population structure analyses (paragraph below), QC was performed on all rivers together, and SNPs in high‐linkage disequilibrium were pruned in PLINK 1.9 (‐‐indep 50 5 1.4) leaving 44,504 unlinked SNPs available for analysis.
2.2.2. Regional Rivers Genetic Structuring
To investigate the genetic variation across rivers in the Moray Firth (Figure 1), a principal component analysis (PCA) was performed in PLINK 1.9 using individuals from the Rivers Oykel, Spey, Findhorn, Deveron, Ness and Ure. Using the same samples, ADMIXTURE v.1.3. (Alexander, Novembre, and Lange 2009) was used to infer the most likely number of genetic clusters (K, testing from K = 1 to K = 6), which was determined based on the lowest cross‐validation error.
2.2.3. Outlier Analysis and Gene Annotation
To detect SNP markers with unusually high levels of allelic differentiation between successful and unsuccessful migrants in each river, two different approaches were computed using the unpruned SNP dataset. In the first approach, the R (R Core Team 2022) package ‘OutFLANK’ v. 0.2 (Whitlock and Lotterhos 2015), which estimates among‐group Fst for each locus, was used. For the second, the allele‐based chi‐squared association test in PLINK 1.9 (command: ‐‐assoc) was implemented. See code (https://tinyurl.com/salmon‐migration‐success) for details about parameters used to run these analyses. Outlier loci in ‘OutFLANK’ were identified by applying a q‐value < 0.05 threshold, while outliers in the association test in PLINK were determined as the top 0.1% SNPs ranked by p‐values. Acknowledging the inherent risk of false positives in genome scan analyses (Luu, Bazin, and Blum 2017), a robust bootstrapping methodology was employed. For each river, 200 bootstrap replicate datasets were generated by randomly removing one fish from each of the migratory groups (successful and unsuccessful). Each of these datasets was examined independently, with 100 analysed using ‘OutFLANK’ and the other 100 using the PLINK association test. Only the outliers that were consistently detected in all 200 bootstrap replicates by both methods were retained for subsequent analysis. Outliers were visualised using the ‘qqman’ v. 0.1.8 R package (Turner 2014). A PCA of the outliers was computed in PLINK on the complete dataset including all fish and the resulting PCA plot was employed to visually test if these outlier SNPs effectively separated successful and unsuccessful migrating salmon smolts in the two rives.
For each river, the 10 genes closest to each outlier SNP were extracted using the closest function in the software bedtools 2.29.1 (Quinlan and Hall 2010) and the genes within 10 kb upstream and downstream of outlier SNPs were filtered in R (Wellband et al. 2019). The potential functions of these genes were assessed by examining the gene ontology (GO) biological process terms associated with each gene, using the R package ‘Ssa.RefSeq.db’ v. 1.2 (Grammes 2016) and literature searches. For these analyses, the NCBI Salmo salar Annotation Release 100 (ICSASG_v2) was used as a reference genome.
2.3. Morphometric Analyses
Fish morphology was analysed using length (mm), weight (g), Fulton's condition factor (K; Nash, Valencia, and Geffen 2006) and geometric morphometrics (GM). The GM analyses were based on photographs of 158 salmon (Spey n = 77 and Oykel n = 81). The images of each fish were imported into tpsUtil v. 1.78 (Rohlf 2019) and randomly shuffled using the Randomly order specimens function so that the operator was blind to the river‐of‐origin of the specimens. Nine fixed and four semi‐landmarks (Figure 2) were digitised on each image by a single operator using tpsDig v. 2.31 (Rohlf 2017) using a subset of landmarks from the scheme proposed by Moccetti et al. (2023). Furthermore, five linear body measurements (Figure 2) used as proxy of body slenderness which has been associated with swimming ability were also included (e.g., Pakkasmaa and Piironen 2001; Drinan et al. 2012). Landmark coordinates were imported into R and analysed using the ‘geomorph’ and ‘RRPP’ v. 4.0.4 packages (Adams et al. 2021; Baken et al. 2021; Collyer and Adams 2021). Preliminary analysis revealed body bending as a major source of shape variation in the dataset. This was corrected by employing landmarks 1, 14, 15 and 11 with the unbend function in tpsUtil. All subsequent analyses were performed on Landmarks 1–13 only. PCA plots were produced with the ‘ggplot2’ package (Wickham 2016).
FIGURE 2.
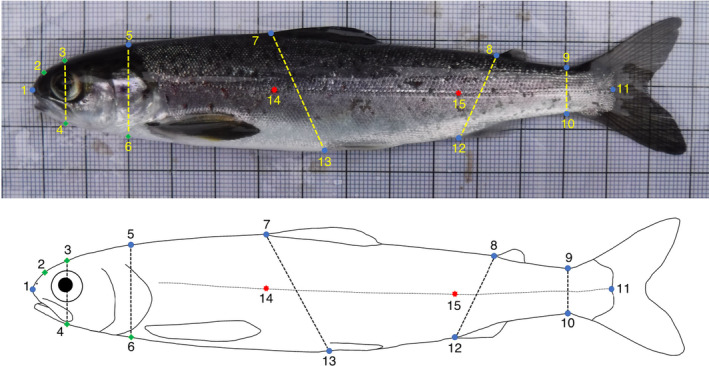
Fixed (blue circles) and semi (green diamonds) landmarks and linear measurements used for the GM analyses of Atlantic salmon smolts (image modified from Moccetti et al. 2023). Landmarks 14 and 15 (red stars) were used to correct for body arching. (1) Tip of snout; (2) midpoint between 1 and 3; (3) directly above middle of eye; (4) perpendicular to lateral line, projected towards 3; (5) dorsal surface posterior of cranium; (6) perpendicular to lateral line, projected towards 5; (7) anterior insertion point of dorsal fin; (8) anterior insertion point of adipose fin; (9) dorsal insertion point of caudal fin; (10) versal insertion point of caudal fin; (11) posterior midpoint of hypural plate; (12) anterior insertion point of anal fin; (13) anterior insertion point of ventral fin; (14) lateral line—perpendicular to 7; and (15) lateral line—perpendicular to 12.
The following analyses were performed separately for each river. First, we tested whether successful and unsuccessful migrating fish in the two rivers were different in length (mm), weight (g) or Fulton's condition factor using t‐ and Mann–Whitney U‐tests depending on the data distribution. Fulton's condition factor (K) was calculated as: K = 100 × W.L −3, where W = weight (g) and L = length (cm). We next tested for a difference in body shape between successful and unsuccessful fish, and whether these differences were consistent across rivers. A generalised Procrustes analysis (GPA) was performed to remove effects not related to body shape through translation, scaling and rotation of the landmark configurations (Rohlf and Slice 1990). The residual effect of fish size on body shape was tested using Procrustes ANOVAs, with Procrustes coordinates used as a response variable and log centroid size used as an independent variable with a randomised residual permutation procedure (10,000 iterations). No significant effect of size on shape was found in either river (p > 0.05). To visualise body shape of successful and unsuccessful fish, a PCA was performed on the Procrustes‐aligned coordinates of fish of each migration category from each river. Procrustes ANOVAs and t‐tests were subsequently used to test for differences in body shape and linear distances between fish with different migratory outcomes. For linear distance and length, weight and condition factor comparisons, significance values were Bonferroni corrected to limit the increased error rate correlated with multiple testing (Rice 1989).
3. Results
3.1. Genomic Analyses
3.1.1. Regional Rivers Genetic Structuring
The study Rivers Spey and Oykel were genetically differentiated from one another (Figure 1). The second principal component (PC2) successfully separated the fish from the River Ness, geographically located between the Oykel and the Spey, from all the other rivers (Figure 1). Fish from the rivers Deveron and Findhorn clustered with the spatially adjacent River Spey (Figure 1). PC1 separated the more geographically distant River Ure, situated in northern England, from all Scottish rivers flowing into the Moray Firth (Figure 1). Similarly, ADMIXTURE analysis identified three different genetic clusters consisting of the Rivers Oykel, Spey and Ure, with the Rivers Deveron and Findhorn clustering with the Spey, while the Ness was admixed with the Rivers Oykel and Spey.
3.1.2. Genomic Regions Linked to Migratory Outcome
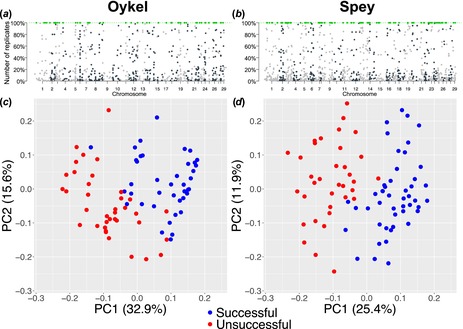
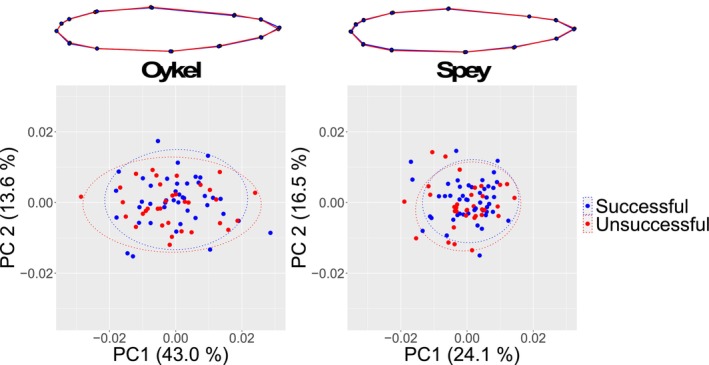
There was a consistently high false discovery rate (FDR) observed across methods and bootstrap replicates in both rivers. On average, only 11.7% of outlier SNPs were detected by ‘OutFLANK’ or PLINK in all 100 replicates. Specifically, the FDR was 89.2% (‘OutFLANK’) and 89.3% (PLINK) in the Oykel dataset and 85.6% (‘OutFLANK’) and 89.3% (PLINK) in the Spey. Seventy outlier SNPs were consistently detected by both methods in all bootstrap replicates for migrating fish in the River Oykel, and 67 outliers for fish from the River Spey (Files S1 and S2). None of the outlier SNPs were found in fish from both rivers. The PCA computed on this subset of outlier loci confirmed their ability to distinguish between successful and unsuccessful fish along PC1 axis for each river (Figure 3).
FIGURE 3.
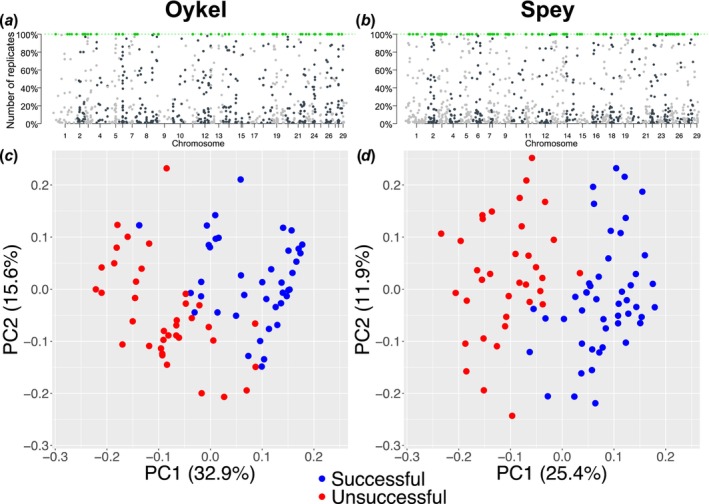
The Manhattan‐style plots (a, b) show all outlier SNPs (dots) identified in bootstrap replicated datasets using ‘OutFLANK’ in each river. The outliers consistently detected in 100% of replicates and used for analysis are highlighted in green. The y‐axis shows the proportion of replicated datasets where each individual outlier SNP was identified. The x‐axis displays the position of the SNPs along the genome with chromosome numbers. (c and d) Principal components analysis scatterplots based on 70 (Oykel) and 67 (Spey) outlier SNPs between successful (blue) and unsuccessful (red) migrant Atlantic salmon smolts. Each dot represents an individual fish. Variance (%) explained by the first and second axes is also shown.
3.1.3. Gene Annotation
There were 50 and 48 putative coding regions (hereafter genes) within 10 kb of the outlier SNPs' locations in the Oykel and Spey fish, respectively (File S3). None of these genes were identified as outliers in fish from both rivers. Eight and 12 genes contained more than one outlier SNP within the 10 kb region in the Oykel and Spey samples, respectively (File S3). The two genes enclosing the highest number of outlier SNPs were the anion exchange protein 2 like (encompassing 18 SNPs, River Oykel) and the collagen alpha‐1(I) chain like (encompassing 10 SNPs, River Spey).
3.2. Morphological Differences Between Successful and Unsuccessful Salmon
There were no differences in length, weight or Fulton's condition factor between successful and unsuccessful migrating smolts (p > 0.07; Figure 4, Tables 2 and 3). Procrustes ANOVAs based on the 13 landmark coordinates did not show significant differences in body shape between successful and unsuccessful smolts in either of the two rivers (p > 0.31; Figure 5, Table 4). Likewise, after Bonferroni correction (new alpha value = 0.005), comparison of body linear measurements did not show any significant difference between migrating groups (p > 0.04; Tables 5 and 6).
FIGURE 4.
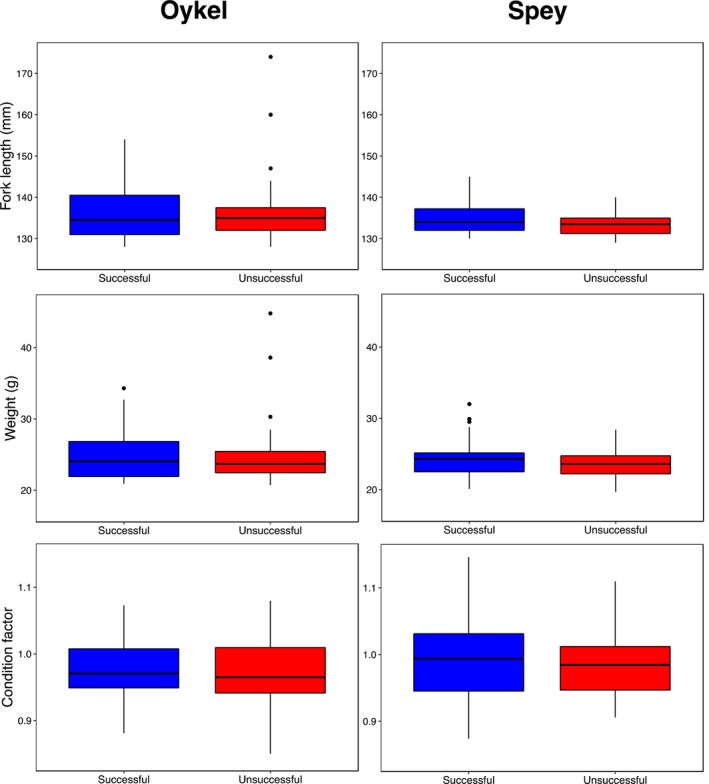
Boxplots of length (mm), weight (g) and Fulton's condition factor to compare successful and unsuccessful Atlantic salmon smolts in the Rivers Oykel and Spey.
TABLE 2.
Effect size and summary statistics of body metrics comparisons between successful and unsuccessful Atlantic salmon smolts in the River Oykel. M–W U = Mann–Whitney U‐test.
| Metric | Mean (±S.D.) successful | Mean (±S.D.) unsuccessful | Test | Df | t | W | p |
|---|---|---|---|---|---|---|---|
| Length (mm) | 135.8 ± 5.6 | 136.7 ± 8.8 | t‐test | 55.2 | −0.513 | — | 0.610 |
| Weight (g) | 24.7 ± 3.1 | 25.0 ± 4.8 | M–W U | — | — | 774 | 0.694 |
| Condition factor | 0.982 ± 0.053 | 0.970 ± 0.048 | t‐test | 74.5 | 1.004 | — | 0.319 |
TABLE 3.
Effect size and summary statistics of body metrics comparisons between successful and unsuccessful Atlantic salmon smolts in the River Spey. M–W U = Mann–Whitney U‐test.
| Metric | Mean (±S.D.) successful | Mean (±S.D.) unsuccessful | Test | Df | t | W | p |
|---|---|---|---|---|---|---|---|
| Length (mm) | 134.9 ± 3.9 | 133.5 ± 2.7 | M–W U | — | — | 951 | 0.145 |
| Weight (g) | 24.5 ± 2.6 | 23.5 ± 2.1 | t‐test | 78.1 | 1.810 | 774 | 0.074 |
| Condition factor | 0.994 ± 0.063 | 0.986 ± 0.052 | M–W U | — | — | 841 | 0.691 |
FIGURE 5.
Mean body shape projections (top) and principal components analysis scatterplots (lower panel) show an absence of shape difference between the migratory groups. Procrustes‐aligned coordinates of successful (blue) and unsuccessful (red) Atlantic salmon, where dots represent individual fish are shown below. Variance (%) explained by the first and second axes and 95% confidence ellipses are displayed. Projections show a complete overlap of the blue (successful) and red (unsuccessful) lines in both rivers despite magnifying morphological differences three times to aid visualisation.
TABLE 4.
Procrustes ANOVA summary statistics of effect of migratory outcome on the body shape of Atlantic salmon smolts in the Rivers Oykel and Spey.
| River | Df | SS | r 2 | F | Z | p |
|---|---|---|---|---|---|---|
| Oykel | 1 | 0.0003318 | 0.0146 | 1.1109 | 0.47321 | 0.313 |
| Spey | 1 | 0.0002305 | 0.01351 | 1.0817 | 0.35458 | 0.366 |
TABLE 5.
Summary statistics of t‐tests comparing body linear measurements between successful and unsuccessful Atlantic salmon smolts in the River Oykel. The alpha value was adjusted to 0.005 following a Bonferroni correction.
| Linear distance (landmark numbers) | Df | t | p |
|---|---|---|---|
| 3–4 | 72.7 | 0.825 | 0.412 |
| 5–6 | 71.7 | 0.819 | 0.416 |
| 7–13 | 74.0 | −1.673 | 0.099 |
| 8–12 | 62.3 | 0.131 | 0.896 |
| 9–10 | 71.7 | 1.985 | 0.051 |
TABLE 6.
Summary statistics of t‐tests comparing body linear measurements between successful and unsuccessful Atlantic salmon smolts in the River Spey. The alpha value was adjusted to 0.005 following a Bonferroni correction.
| Linear distance (landmark numbers) | Df | t | p |
|---|---|---|---|
| 3–4 | 68.9 | −1.517 | 0.134 |
| 5–6 | 73.7 | −2.118 | 0.038 |
| 7–13 | 77.5 | 0.163 | 0.871 |
| 8–12 | 76.6 | −0.938 | 0.351 |
| 9–10 | 73.0 | −0.218 | 0.828 |
4. Discussion
Our work shows that distinct SNP sets were significantly differentiated between Atlantic salmon smolts making successful migrations to sea and those that failed to migrate to sea in two different rivers. In both rivers, the outlier SNPs predicting individual migration success were near several genes that could be relevant for migration, but we found no evidence of phenotypic differences in body shape between successful and unsuccessful Atlantic salmon river migrants.
Categorising genes containing outlier SNPs by biological function highlighted similar processes across the study rivers. In fish from both rivers, genes putatively linked to osmoregulation, immunity, stress and nervous, sensory, muscular, skeletal and cardiovascular system development and activity were detected.
Candidate genes linked to general neuronal, cardiovascular and skeletal functions may play an important role in migration, but a direct link to smolt migration success is hard to determine. Furthermore, given the susceptibility of gene annotation to false positives, it is important to exercise caution when attempting to establish such correlations (Pavlidis et al. 2012). Nevertheless, osmoregulation and immune response are processes shown to play an important part in salmonid migration. In the Oykel and the Spey, outlier SNPs were located within or near (within 10 kb) several candidate osmoregulatory genes. These genes were associated with a range of processes including ion transmembrane and water transport, renal activity, response to salt stress and hyperosmotic response. Noteworthy is the identification of the anion exchange protein 2‐like gene, encompassing 18 of the 70 outlier SNPs detected in the Oykel. This gene is associated with GO terms involved in osmoregulatory processes, such as chloride and bicarbonate transmembrane transport (Wilson, Wilson, and Grosell 2002; Grosell 2006). Osmoregulation and individual ability to undergo physiological changes required for seawater entry have been shown to be important to increase chances of survival and predator avoidance in seaward migrating salmonid smolts (Kennedy, Gale, and Ostrand 2007) and could play a role in migration success of Atlantic salmon smolts in the last tidal kilometres of the Oykel and Spey where transitional zone between freshwater and saltwater occurs.
Immunity‐related and stress‐response genes were also widely detected in association with the outlier SNPs separating successful and unsuccessful river migrants in fish from both the Oykel and Spey. Studies using proteomics in Pacific salmon (Oncorhynchus spp.) have found significant correlation among migratory outcome, expression of specific immune‐related genes and viral‐ and parasite‐induced infection burden (Miller et al. 2011; Jeffries et al. 2014; Furey et al. 2021; Mauduit et al. 2022). The stress hormone cortisol has also been found to be a good predictor of migration success in salmonids (Birnie‐Gauvin et al. 2019; Birnie‐Gauvin, Thorstad, and Aarestrup 2019). Our findings now highlight the potentially important role of pathogens‐driven selection in Atlantic salmon migration success. An additional factor that requires further investigation is the possibility that there are individual differences in response to the tagging process since there are also immune genes annotated with GO terms involved in blood coagulation and response to wounding. To determine migration patterns, all the fish in our study were tagged, so although this is not a confounding factor in our design, this finding warrants further investigation.
While particular SNP sets allow us to predict migratory outcome of Atlantic salmon smolts in the Oykel and Spey, analyses of length, weight, body condition and body shape did not find any significant difference between successful and unsuccessful migrants in either of the rivers. This is somewhat surprising given the importance that morphology plays in swimming performance in fish (Webb 1978, 1984; Fisher and Hogan 2007; Langerhans and Reznick 2010; Assumpção et al. 2012), the specific hydrodynamic characteristics required to effectively migrate in running waters (Langerhans and Reznick 2010; Brodersen et al. 2014) and that size, body shape and condition may also be important in antipredatory behaviour (Domenici et al. 2007). Although a larger sample size might provide greater statistical power and potentially uncover morphological differences if body shape played a role in migration success, it is likely those differences would already be detectable with our current sample size, given the high sensitivity of GM, which can detect even tiny differences in body shape (Webster and Sheets 2010). Morphology, therefore, does not appear to significantly influence migration success of Atlantic salmon smolts in the study rivers. This finding supports the idea that other phenotypic traits, such as physiological performance and immunity described earlier, maybe more critical to migratory success.
Previous work found a difference in smolt morphology between rivers (Moccetti et al. 2023) and there is genetic differentiation of the Oykel and Spey salmon populations indicating reproductive isolation between these geographically close populations, likely facilitated by fine‐scale homing. Different evolutionary histories (evidenced by the geographic structure we find between river populations) combined with different contemporary ecological selection pressures might therefore lead to different traits being linked to migration success. For example, there were no genes that contained outlier SNPs in common in Atlantic salmon from both the Oykel and Spey. It remains possible that increasing the sample size might reveal additional genomic regions associated with migration success that overlap between rivers. However, the bootstrapping approach demonstrated consistent results across migratory groups, suggesting that important genomic regions linked to migration would likely already have been detected with the current sample size, especially given the high‐density 200 K SNP panel used in this study.
Overall, we found that migratory outcomes for individual Atlantic salmon smolts in given rivers, in a given year, could be predicted from a subset of SNPs consistently detected through bootstrapping approach. We next need to understand the ecological and environmental factors which could determine those subsets by adding temporal replication so that we can better understand the limits of our study. From an evolutionary and conservation point of view, the mechanism through which the observed genetic diversity could be maintained needs to be identified, given that migration failure should theoretically quickly purge polymorphism at selected SNPs from a population. We propose that variation in life history could maintain standing genetic variation for environmentally driven balancing selection (Mérot et al. 2020). ‘Partial’ migration (Shaw 2016), where only a portion of the population migrates, is common in several taxa and may be responsible for maintaining high genetic diversity of migratory and resident individuals interbreed (Pulido 2011). Within‐population differences in migratory strategies (e.g., timing, duration and routes) between age classes and sexes are also well‐known phenomena (Cristol, Baker, and Carbone 1999). These subgroups are exposed to different biotic and abiotic conditions potentially selecting different genotypes that may maintain the gene pool diversity within the population (Dingle and Drake 2007; Wittmann et al. 2017; Briedis and Bauer 2018). Alternative migratory and reproductive tactics are well documented in Atlantic salmon (Fleming 1998; Thorstad et al. 2010; Birnie‐Gauvin, Thorstad, and Aarestrup 2019), thus individuals with different life histories experiencing temporally and spatially fluctuating selection can interbreed and induce genetic mixing. Typically, the life history of Atlantic salmon involves seaward migration followed by a return to their natal river to spawn, but a number of males (and occasionally females, Birnie‐Gauvin, Thorstad, and Aarestrup 2019) become sexually reproductive in freshwater as morphological juveniles before migrating to sea (‘precocious male parr’; Lepais et al. 2017). Their contribution to paternity could be substantial (ca. 60% in one study, Saura et al. 2008). The number of years spent in freshwater before smolting and at sea before upstream spawning migration can also vary considerably (Thorstad et al. 2010). Finally, unlike Pacific salmon species, a non‐negligible proportion of Atlantic salmon survive reproduction (especially females), return to the ocean as ‘kelt’ and spawn multiple times (Hedger et al. 2009). Weather and ecological conditions can change dramatically among and within years inducing different selective pressures on migrating smolts and other salmon life stages. For instance, variations in water discharge and temperature may affect ecological factors such as migration timing (Thorstad et al. 2010), predation (Kennedy, Gale, and Ostrand 2007; Hostetter et al. 2012) and pathogen infection (Wagner et al. 2005) as well as passage of artificial migration barriers (Marschall et al. 2011). Clearly, all these variables may differentially alter the allele frequencies under selection and help maintain standing genetic variation. Straying between rivers could also be a source of genetic diversity (Palstra, O'Connell, and Ruzzante 2007; Keefer and Caudill 2014), although we found no evidence of this in our study.
From a conservation point of view, understanding and predicting these selection pressures could be invaluable in managing existing populations, and could inform stock selection where hatchery‐reared individuals are used to augment populations (Jepsen, Nielsen, and Deacon 2003; Koed et al. 2020; Waples, Naish, and Primmer 2020).
Overall, our findings show that migration success could be linked to specific genotypes and highlight the potential importance of preserving genetic diversity for conservation, to allow populations to respond to potential heterogeneity between years, and the increased variability that long‐term climate change may produce. Our next challenge is to understand in detail the selection pressures and associated genetic changes in populations facilitating conservation success and ensuring a future for these iconic species.
Author Contributions
Paolo Moccetti: data curation (equal), formal analysis (lead), investigation (equal), methodology (equal), validation (lead), visualization (lead), writing – original draft (lead), writing – review and editing (lead). Jonathan D. Bolland: conceptualization (equal), funding acquisition (equal), methodology (equal), project administration (equal), supervision (equal), writing – review and editing (equal). Colin E. Adams: conceptualization (equal), funding acquisition (equal), methodology (equal), project administration (equal), resources (equal), supervision (equal), writing – review and editing (equal). Jessica R. Rodger: investigation (equal), methodology (equal), resources (equal), writing – review and editing (equal). Hannele M. Honkanen: investigation (equal), methodology (equal), resources (equal), writing – review and editing (equal). Matthew Newton: investigation (equal), methodology (equal), resources (equal), writing – review and editing (equal). Angus J. Lothian: investigation (equal), methodology (equal), resources (equal), writing – review and editing (equal). Andy D. Nunn: methodology (equal), project administration (equal), supervision (equal), writing – review and editing (equal). Domino A. Joyce: conceptualization (equal), funding acquisition (equal), methodology (equal), project administration (equal), supervision (equal), validation (equal), visualization (supporting), writing – review and editing (equal).
Ethics Statement
For fish samples from the rivers Oykel, Spey, Deveron, Findhorn and Ness, the care and use of experimental animals complied with the UK Home Office animal welfare laws, guidelines and policies (UK Home Office Licence PPL 70/8794) and was approved by the University of Glasgow Animal Welfare and Ethics Review Board (AWERB). Atlantic salmon from the River Ure were treated in compliance with the UK ASPA (1986) Home Office project licence number PD6C17B56.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1.
Acknowledgements
We would like to thank the Spey Fisheries Board and the Kyle of Sutherland Fisheries Trust for their help and support with this project. We would also like to thank Jonathan Archer, Georgios Kyriakou, Fraser Brydon, Jessica Whitney, Mustafa Soganci and Rowan Smith for their help in collecting the data for this study.
Funding: This work was part of the Missing Salmon Project funded by the Atlantic Salmon Trust. Paolo Moccetti was supported by the Leeds‐York‐Hull Natural Environment Research Council (NERC) Doctoral Training Partnership (DTP) Panorama (NE/S007458/1).
Data Availability Statement
Data and code used for analyses are publicly available at: https://tinyurl.com/salmon‐migration‐success.
References
- Adams, D. C. , Collyer M., Kaliontzopoulou A., and Baken E.. 2021. “Geomorph: Software for Geometric Morphometric Analyses. R Package Version 4.0.2.” https://cran.r‐project.org/package=geomorph.
- Alerstam, T. , Hedenström A., and Åkesson S.. 2003. “Long‐Distance Migration: Evolution and Determinants.” Oikos 103: 247–260. 10.1034/j.1600-0706.2003.12559.x. [DOI] [Google Scholar]
- Alexander, D. H. , Novembre J., and Lange K.. 2009. “Fast Model‐Based Estimation of Ancestry in Unrelated Individuals.” Genome Research 19: 1655–1664. 10.1101/gr.094052.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Assumpção, L. D. , Makrakis M. C., Makrakis S., et al. 2012. “The Use of Morphometric Analysis to Predict the Swimming Efficiency of Two Neotropical Long‐Distance Migratory Species in Fish Passage.” Neotropical Ichthyology 10: 797–804. 10.1590/S1679-62252012000400012. [DOI] [Google Scholar]
- Baken, E. K. , Collyer M. L., Kaliontzopoulou A., and Adams D. C.. 2021. “Geomorph v4.0 and gmShiny: Enhanced Analytics and a New Graphical Interface for a Comprehensive Morphometric Experience.” Methods in Ecology and Evolution 12: 2355–2363. 10.1111/2041-210X.13723. [DOI] [Google Scholar]
- Baker, N. J. , Boubée J., Lokman P. M., and Bolland J. D.. 2020. “Evaluating the Impact of Hydropower on Downstream Migrating Anguillid Eels: Catchment‐Wide and Fine‐Scale Approaches to Identify Cost‐Effective Solutions.” Science of the Total Environment 748: 141111. 10.1016/j.scitotenv.2020.141111. [DOI] [PubMed] [Google Scholar]
- Barson, N. J. , Aykanat T., Hindar K., et al. 2015. “Sex‐Dependent Dominance at a Single Locus Maintains Variation in Age at Maturity in Salmon.” Nature 528: 405–408. 10.1038/nature16062. [DOI] [PubMed] [Google Scholar]
- Bauer, S. , and Hoye B. J.. 2014. “Migratory Animals Couple Biodiversity and Ecosystem Functioning Worldwide.” Science 344: 1242552. 10.1126/science.1242552. [DOI] [PubMed] [Google Scholar]
- Bernos, T. A. , Jeffries K. M., and Mandrak N. E.. 2020. “Linking Genomics and Fish Conservation Decision Making: A Review.” Reviews in Fish Biology and Fisheries 30: 587–604. 10.1007/s11160-020-09618-8. [DOI] [Google Scholar]
- Birnie‐Gauvin, K. , Flávio H., Kristensen M. L., et al. 2019. “Cortisol Predicts Migration Timing and Success in Both Atlantic Salmon and Sea Trout Kelts.” Scientific Reports 9: 2422. 10.1038/s41598-019-39153-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birnie‐Gauvin, K. , Thorstad E. B., and Aarestrup K.. 2019. “Overlooked Aspects of the Salmo Salar and Salmo trutta Lifecycles.” Reviews in Fish Biology and Fisheries 29: 749–766. 10.1007/s11160-019-09575-x. [DOI] [Google Scholar]
- Bourret, V. , Dionne M., and Bernatchez L.. 2014. “Detecting Genotypic Changes Associated With Selective Mortality at Sea in Atlantic Salmon: Polygenic Multilocus Analysis Surpasses Genome Scan.” Molecular Ecology 23: 4444–4457. 10.1111/mec.12798. [DOI] [PubMed] [Google Scholar]
- Bowlin, M. S. , Bisson I. A., Shamoun‐Baranes J., et al. 2010. “Grand Challenges in Migration Biology.” Integrative and Comparative Biology 50: 261–279. 10.1093/icb/icq013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Briedis, M. , and Bauer S.. 2018. “Migratory Connectivity in the Context of Differential Migration.” Biology Letters 14: 20180679. 10.1098/rsbl.2018.0679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodersen, J. , Chapman B. B., Nilsson P. A., Skov C., Hansson L. A., and Brönmark C.. 2014. “Fixed and Flexible: Coexistence of Obligate and Facultative Migratory Strategies in a Freshwater Fish.” PLoS One 9: e90294. 10.1371/journal.pone.0090294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang, C. C. , Chow C. C., Tellier L. C. A. M., Vattikuti S., Purcell S. M., and Lee J. J.. 2015. “Second‐Generation PLINK: Rising to the Challenge of Larger and Richer Datasets.” GigaScience 4: 7. 10.1186/s13742-015-0047-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chapman, B. B. , Hulthén K., Brönmark C., et al. 2015. “Shape Up or Ship out: Migratory Behaviour Predicts Morphology Across Spatial Scale in a Freshwater Fish.” Journal of Animal Ecology 84: 1187–1193. 10.1111/1365-2656.12374. [DOI] [PubMed] [Google Scholar]
- Collyer, M. L. , and Adams D. C.. 2021. “RRPP: Linear Model Evaluation With Randomized Residuals in a Permutation Procedure, R Package Version 1.1.2.” https://cran.r‐project.org/package=RRPP.
- Cristol, D. A. , Baker M. B., and Carbone C.. 1999. “Differential Migration Revisited: Latitudinal Segregation by Age and Sex Class.” In Current Ornithology, edited by Nolan V., Ketterson E. D., and Thompson C. F., 33–88. Boston, MA: Springer US. 10.1007/978-1-4757-4901-4_2. [DOI] [Google Scholar]
- Dadswell, M. , Spares A., Reader J., et al. 2022. “The Decline and Impending Collapse of the Atlantic Salmon ( Salmo salar ) Population in the North Atlantic Ocean: A Review of Possible Causes.” Reviews in Fisheries Science & Aquaculture 30: 215–258. 10.1080/23308249.2021.1937044. [DOI] [Google Scholar]
- Dingle, H. , and Drake V. A.. 2007. “What Is Migration?” Bioscience 57: 113–121. 10.1641/b570206. [DOI] [Google Scholar]
- Doherty, P. D. , Baxter J. M., Gell F. R., et al. 2017. “Long‐Term Satellite Tracking Reveals Variable Seasonal Migration Strategies of Basking Sharks in the North‐East Atlantic.” Scientific Reports 7: 42837. 10.1038/srep42837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domenici, P. , Turesson H., Brodersen J., and Brönmark C.. 2007. “Predator‐Induced Morphology Enhances Escape Locomotion in Crucian Carp.” Proceedings of the Royal Society B: Biological Sciences 275: 195–201. 10.1098/rspb.2007.1088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drinan, T. J. , McGinnity P., Coughlan J. P., Cross T. F., and Harrison S. S. C.. 2012. “Morphological Variability of Atlantic Salmon Salmo Salar and Brown Trout Salmo trutta in Different River Environments.” Ecology of Freshwater Fish 21: 420–432. 10.1111/j.1600-0633.2012.00561.x. [DOI] [Google Scholar]
- Fisher, R. , and Hogan J. D.. 2007. “Morphological Predictors of Swimming Speed: A Case Study of Pre‐Settlement Juvenile Coral Reef Fishes.” Journal of Experimental Biology 210: 2436–2443. 10.1242/jeb.004275. [DOI] [PubMed] [Google Scholar]
- Fleming, I. A. 1998. “Pattern and Variability in the Breeding System of Atlantic Salmon ( Salmo salar ), with Comparisons to Other Salmonids.” Canadian Journal of Fisheries and Aquatic Sciences 55: 59–76. 10.1139/d98-009. [DOI] [Google Scholar]
- Forseth, T. , Barlaup B. T., Finstad B., et al. 2017. “The Major Threats to Atlantic Salmon in Norway.” ICES Journal of Marine Science 74: 1496–1513. 10.1093/icesjms/fsx020. [DOI] [Google Scholar]
- Fraser, D. J. , Weir L. K., Bernatchez L., Hansen M. M., and Taylor E. B.. 2011. “Extent and Scale of Local Adaptation in Salmonid Fishes: Review and meta‐Analysis.” Heredity 106: 404–420. 10.1038/hdy.2010.167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fruciano, C. , Tigano C., and Ferrito V.. 2011. “Geographical and Morphological Variation Within and Between Colour Phases in Coris julis (L. 1758), a Protogynous Marine Fish.” Biological Journal of the Linnean Society 104: 148–162. 10.1111/j.1095-8312.2011.01700.x. [DOI] [Google Scholar]
- Furey, N. B. , Bass A. L., Miller K. M., et al. 2021. “Infected Juvenile Salmon Can Experience Increased Predation During Freshwater Migration.” Royal Society Open Science 8: 201522. 10.1098/rsos.201522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furey, N. B. , Hinch S. G., Bass A. L., Middleton C. T., Minke‐Martin V., and Lotto A. G.. 2016. “Predator Swamping Reduces Predation Risk During Nocturnal Migration of Juvenile Salmon in a High‐Mortality Landscape.” Journal of Animal Ecology 85: 948–959. 10.1111/1365-2656.12528. [DOI] [PubMed] [Google Scholar]
- Garcia De Leaniz, C. , Fleming I. A., Einum S., et al. 2007. “A Critical Review of Adaptive Genetic Variation in Atlantic Salmon: Implications for Conservation.” Biological Reviews 82: 173–211. 10.1111/j.1469-185X.2006.00004.x. [DOI] [PubMed] [Google Scholar]
- Goerig, E. , Wasserman B. A., Castro‐Santos T., and Palkovacs E. P.. 2020. “Body Shape Is Related to the Attempt Rate and Passage Success of Brook Trout at In‐Stream Barriers.” Journal of Applied Ecology 57: 91–100. 10.1111/1365-2664.13497. [DOI] [Google Scholar]
- Grammes, F. 2016. “Ssa.RefSeq.db: Annotation Package for the Salmo salar RefSeq (ICSASG_v2).” https://rdrr.io/github/FabianGrammes/Ssa.RefSeq.db/.
- Grosell, M. 2006. “Intestinal Anion Exchange in Marine Fish Osmoregulation.” Journal of Experimental Biology 209: 2813–2827. 10.1242/jeb.02345. [DOI] [PubMed] [Google Scholar]
- Harris, G. , Thirgood S., Hopcraft J. G. C., Cromsigt J. P. G. M., and Berger J.. 2009. “Global Decline in Aggregated Migrations of Large Terrestrial Mammals.” Endangered Species Research 7: 55–76. 10.3354/esr00173. [DOI] [Google Scholar]
- Hays, G. C. , Broderick A. C., Godley B. J., and Luschi P.. 2003. “Satellite Telemetry Suggests High Levels of Fishing‐Induced Mortality in Marine Turtles.” Marine Ecology Progress Series 262: 305–309. 10.3354/meps262305. [DOI] [Google Scholar]
- Hecht, B. C. , Hard J. J., Thrower F. P., and Nichols K. M.. 2015. “Quantitative Genetics of Migration‐Related Traits in Rainbow and Steelhead Trout.” G3: Genes, Genomes, Genetics 5: 873–889. 10.1534/g3.114.016469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hecht, B. C. , Thrower F. P., Hale M. C., Miller M. R., and Nichols K. M.. 2012. “Genetic Architecture of Migration‐Related Traits in Rainbow and Steelhead Trout, Oncorhynchus mykiss .” G3: Genes, Genomes, Genetics 2: 1113–1127. 10.1534/g3.112.003137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedger, R. D. , Hatin D., Dodson J. J., et al. 2009. “Migration and Swimming Depth of Atlantic Salmon Kelts Salmo salar in Coastal Zone and Marine Habitats.” Marine Ecology Progress Series 392: 179–192. [Google Scholar]
- Hess, J. E. , Caudill C. C., Keefer M. L., McIlraith B. J., Moser M. L., and Narum S. R.. 2014. “Genes Predict Long Distance Migration and Large Body Size in a Migratory Fish, Pacific Lamprey.” Evolutionary Applications 7: 1192–1208. 10.1111/eva.12203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hostetter, N. J. , Evans A. F., Roby D. D., and Collis K.. 2012. “Susceptibility of Juvenile Steelhead to Avian Predation: The Influence of Individual Fish Characteristics and River Conditions.” Transactions of the American Fisheries Society 141: 1586–1599. 10.1080/00028487.2012.716011. [DOI] [Google Scholar]
- ICES . 2024. “Working Group on North Atlantic Salmon (WGNAS).” ICES Scientific Reports 6: 36. 10.17895/ices.pub.25730247. [DOI] [Google Scholar]
- Jeffries, K. M. , Hinch S. G., Gale M. K., et al. 2014. “Immune Response Genes and Pathogen Presence Predict Migration Survival in Wild Salmon Smolts.” Molecular Ecology 23: 5803–5815. 10.1111/mec.12980. [DOI] [PubMed] [Google Scholar]
- Jepsen, N. , Nielsen E. E., and Deacon M.. 2003. “Linking Individual Migratory Behaviour of Atlantic Salmon to Their Genetic Origin.” In Aquatic Telemetry: Advances and Applications. Proceedings of the Fifth Conference on Fish Telemetry Held in Europe, Ustica, Italy, 9–13. Rome: FAO/COISPA. [Google Scholar]
- Justen, H. , and Delmore K. E.. 2022. “The Genetics of Bird Migration.” Current Biology 32: R1144–R1149. 10.1016/j.cub.2022.07.008. [DOI] [PubMed] [Google Scholar]
- Keefer, M. L. , and Caudill C. C.. 2014. “Homing and Straying by Anadromous Salmonids: A Review of Mechanisms and Rates.” Reviews in Fish Biology and Fisheries 24: 333–368. 10.1007/s11160-013-9334-6. [DOI] [Google Scholar]
- Kennedy, B. M. , Gale W. L., and Ostrand K. G.. 2007. “Relationship Between Smolt Gill Na+, K+ ATPase Activity and Migration Timing to Avian Predation Risk of Steelhead Trout ( Oncorhynchus mykiss ) in a Large Estuary.” Canadian Journal of Fisheries and Aquatic Sciences 64: 1506–1516. 10.1139/f07-117. [DOI] [Google Scholar]
- Klaassen, R. H. G. , Hake M., Strandberg R., et al. 2014. “When and Where Does Mortality Occur in Migratory Birds? Direct Evidence From Long‐Term Satellite Tracking of Raptors.” Journal of Animal Ecology 83: 176–184. 10.1111/1365-2656.12135. [DOI] [PubMed] [Google Scholar]
- Koed, A. , Birnie‐Gauvin K., Sivebæk F., and Aarestrup K.. 2020. “From Endangered to Sustainable: Multi‐Faceted Management in Rivers and Coasts Improves Atlantic Salmon ( Salmo salar ) Populations in Denmark.” Fisheries Management and Ecology 27: 64–76. 10.1111/fme.12385. [DOI] [Google Scholar]
- Langerhans, R. B. , and Reznick D. N.. 2010. “Ecology and Evolution of Swimming Performance in Fishes: Predicting Evolution With Biomechanics.” In Fish Locomotion: An Eco‐ Ethological Perspective, edited by Domenici O. and Kapoor B. G., 200–248. Enfield, New Hampshire: Science Publisher. [Google Scholar]
- Lepais, O. , Manicki A., Glise S., Buoro M., and Bardonnet A.. 2017. “Genetic Architecture of Threshold Reaction Norms for Male Alternative Reproductive Tactics in Atlantic Salmon ( Salmo salar L.).” Scientific Reports 7: 1–13. 10.1038/srep43552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liedvogel, M. , Åkesson S., and Bensch S.. 2011. “The Genetics of Migration on the Move.” Trends in Ecology & Evolution 26: 561–569. 10.1016/j.tree.2011.07.009. [DOI] [PubMed] [Google Scholar]
- Lilly, J. , Honkanen H. M., McCallum J. M., Newton M., Bailey D. M., and Adams C. E.. 2022. “Combining Acoustic Telemetry With a Mechanistic Model to Investigate Characteristics Unique to Successful Atlantic Salmon Smolt Migrants Through a Standing Body of Water.” Environmental Biology of Fishes 105: 2045–2063. 10.1007/s10641-021-01172-x. [DOI] [Google Scholar]
- Lothian, A. J. , Newton M., Barry J., Walters M., Miller R. C., and Adams C. E.. 2018. “Migration Pathways, Speed and Mortality of Atlantic Salmon ( Salmo salar ) Smolts in a Scottish River and the Near‐Shore Coastal Marine Environment.” Ecology of Freshwater Fish 27: 549–558. 10.1111/eff.12369. [DOI] [Google Scholar]
- Lothian, A. J. , Schwinn M., Anton A. H., et al. 2020. “Are We Designing Fishways for Diversity ? Potential Selection on Alternative Phenotypes Resulting From Differential Passage in Brown Trout.” Journal of Environmental Management 262: 110317. 10.1016/j.jenvman.2020.110317. [DOI] [PubMed] [Google Scholar]
- Luu, K. , Bazin E., and Blum M. G. B.. 2017. “Pcadapt: An R Package to Perform Genome Scans for Selection Based on Principal Component Analysis.” Molecular Ecology Resources 17: 67–77. 10.1111/1755-0998.12592. [DOI] [PubMed] [Google Scholar]
- Marschall, E. A. , Mather M. E., Parrish D. L., Allison G. W., and McMenemy J. R.. 2011. “Migration Delays Caused by Anthropogenic Barriers: Modeling Dams, Temperature, and Success of Migrating Salmon Smolts.” Ecological Applications 21: 3014–3031. 10.1890/10-0593.1. [DOI] [Google Scholar]
- Mauduit, F. , Segarra A., Mandic M., et al. 2022. “Understanding Risks and Consequences of Pathogen Infections on the Physiological Performance of Outmigrating Chinook Salmon.” Conservation Physiology 10: coab102. 10.1093/conphys/coab102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mérot, C. , Llaurens V., Normandeau E., Bernatchez L., and Wellenreuther M.. 2020. “Balancing Selection via Life‐History Trade‐Offs Maintains an Inversion Polymorphism in a Seaweed Fly.” Nature Communications 11: 670. 10.1038/s41467-020-14479-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Middleton, A. D. , Kauffman M. J., McWhirter D. E., et al. 2013. “Animal Migration Amid Shifting Patterns of Phenology and Predation: Lessons From a Yellowstone Elk Herd.” Ecology 94: 1245–1256. 10.1890/11-2298.1. [DOI] [PubMed] [Google Scholar]
- Miller, K. M. , Li S., Kaukinen K. H., et al. 2011. “Genomic Signatures Predict Migration and Spawning Failure in Wild Canadian Salmon.” Science 331: 214–217. 10.1126/science.1196901. [DOI] [PubMed] [Google Scholar]
- Minias, P. , Kaczmarek K., Włodarczyk R., and Janiszewski T.. 2013. “Wing Shape Influences Stopover Strategies in a Migratory Shorebird, the Common Snipe.” Condor 115: 535–542. 10.1525/cond.2013.120137. [DOI] [Google Scholar]
- Moccetti, P. , Rodger J. R., Bolland J. D., et al. 2023. “Is Shape in the Eye of the Beholder? Assessing Landmarking Error in Geometric Morphometric Analyses on Live Fish.” PeerJ 11: e15545. 10.7717/peerj.15545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nash, R. D. M. , Valencia A. H., and Geffen A. J.. 2006. “The Origin of Fulton's Condition Factor ‐ Setting the Record Straight.” Fisheries 31: 236–238. [Google Scholar]
- Newton, M. , Dodd J. A., Barry J., Boylan P., and Adams C. E.. 2018. “The Impact of a Small‐Scale Riverine Obstacle on the Upstream Migration of Atlantic Salmon.” Hydrobiologia 806: 251–264. 10.1007/s10750-017-3364-3. [DOI] [Google Scholar]
- O'Malley, K. G. , Jacobson D. P., Kurth R., Dill A. J., and Banks M. A.. 2013. “Adaptive Genetic Markers Discriminate Migratory Runs of Chinook Salmon ( Oncorhynchus tshawytscha ) Amid Continued Gene Flow.” Evolutionary Applications 6: 1184–1194. 10.1111/eva.12095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Páez, D. J. , and Dodson J. J.. 2017. “Environment‐Specific Heritabilities and Maternal Effects for Body Size, Morphology and Survival in Juvenile Atlantic Salmon ( Salmo salar ): Evidence From a Field Experiment.” Environmental Biology of Fishes 100: 209–221. 10.1007/s10641-016-0572-z. [DOI] [Google Scholar]
- Pakkasmaa, S. , and Piironen J.. 2001. “Water Velocity Shapes Juvenile Salmonids.” Evolutionary Ecology 14: 721–730. 10.1023/A:1011691810801. [DOI] [Google Scholar]
- Palacín, C. , Alonso J. C., Martín C. A., and Alonso J. A.. 2017. “Changes in Bird‐Migration Patterns Associated With Human‐Induced Mortality.” Conservation Biology 31: 106–115. 10.1111/cobi.12758. [DOI] [PubMed] [Google Scholar]
- Palstra, F. P. , O'Connell M. F., and Ruzzante D. E.. 2007. “Population Structure and Gene Flow Reversals in Atlantic Salmon ( Salmo salar ) Over Contemporary and Long‐Term Temporal Scales: Effects of Population Size and Life History.” Molecular Ecology 16: 4504–4522. 10.1111/j.1365-294X.2007.03541.x. [DOI] [PubMed] [Google Scholar]
- Pavlidis, P. , Jensen J. D., Stephan W., and Stamatakis A.. 2012. “A Critical Assessment of Storytelling: Gene Ontology Categories and the Importance of Validating Genomic Scans.” Molecular Biology and Evolution 29: 3237–3248. 10.1093/molbev/mss136. [DOI] [PubMed] [Google Scholar]
- Pritchard, V. L. , Mäkinen H., Vähä J. P., Erkinaro J., Orell P., and Primmer C. R.. 2018. “Genomic Signatures of Fine‐Scale Local Selection in Atlantic Salmon Suggest Involvement of Sexual Maturation, Energy Homeostasis and Immune Defence‐Related Genes.” Molecular Ecology 27: 2560–2575. 10.1111/mec.14705. [DOI] [PubMed] [Google Scholar]
- Pulido, F. 2011. “Evolutionary Genetics of Partial Migration ‐ The Threshold Model of Migration Revis(It)ed.” Oikos 120: 1776–1783. 10.1111/j.1600-0706.2011.19844.x. [DOI] [Google Scholar]
- Purcell, S. , Neale B., Todd‐Brown K., et al. 2007. “PLINK: A Tool Set for Whole‐Genome Association and Population‐Based Linkage Analyses.” American Journal of Human Genetics 81: 559–575. 10.1086/519795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quinlan, A. R. , and Hall I. M.. 2010. “BEDTools: A Flexible Suite of Utilities for Comparing Genomic Features.” Bioinformatics 26: 841–842. 10.1093/bioinformatics/btq033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Core Team . 2022. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. https://www.R‐project.org/. [Google Scholar]
- Rice, W. R. 1989. “Analyzing Tables of Statistical Tests.” Evolution 43: 223–225. 10.2307/2409177. [DOI] [PubMed] [Google Scholar]
- Rohland, N. , and Reich D.. 2012. “Cost‐Effective, High‐Throughput DNA Sequencing Libraries for Multiplexed Target Capture.” Genome Research 22: 939–946. 10.1101/gr.128124.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rohlf, F. 2017. tpsUtil v. 1.78. Stony Brook, NY: Department of Ecology and Evolution, State University of New York. [Google Scholar]
- Rohlf, F. 2019. tpsDig v. 2.31. Stony Brook, NY: Department of Ecology and Evolution, State University of New York. [Google Scholar]
- Rohlf, F. J. , and Slice D.. 1990. “Extensions of the Procrustes Method for the Optimal Superimposition of Landmarks.” Systematic Biology 39: 40–59. 10.2307/2992207. [DOI] [Google Scholar]
- Romer, J. D. , Leblanc C. A., Clements S., et al. 2013. “Survival and Behavior of Juvenile Steelhead Trout ( Oncorhynchus mykiss ) in Two Estuaries in Oregon, USA.” Environmental Biology of Fishes 96: 849–863. 10.1007/s10641-012-0080-8. [DOI] [Google Scholar]
- Saura, M. , Caballero A., Caballero P., and Morán P.. 2008. “Impact of Precocious Male Parr on the Effective Size of a Wild Population of Atlantic Salmon.” Freshwater Biology 53: 2375–2384. 10.1111/j.1365-2427.2008.02062.x. [DOI] [Google Scholar]
- Sellers, G. S. , Di Muri C., Gómez A., and Hänfling B.. 2018. “Mu‐DNA: A Modular Universal DNA Extraction Method Adaptable for a Wide Range of Sample Types.” Metabarcoding and Metagenomics 2: e24556. 10.3897/mbmg.2.24556. [DOI] [Google Scholar]
- Shaw, A. K. 2016. “Drivers of Animal Migration and Implications in Changing Environments.” Evolutionary Ecology 30: 991–1007. 10.1007/s10682-016-9860-5. [DOI] [Google Scholar]
- Skulason, S. , and Smith T. B.. 1995. “Resource Polymorphisms in Vertebrates.” Trends in Ecology & Evolution 10: 366–370. 10.1016/S0169-5347(00)89135-1. [DOI] [PubMed] [Google Scholar]
- Stelkens, R. B. , Jaffuel G., Escher M., and Wedekind C.. 2012. “Genetic and Phenotypic Population Divergence on a Microgeographic Scale in Brown Trout.” Molecular Ecology 21: 2896–2915. 10.1111/j.1365-294X.2012.05581.x. [DOI] [PubMed] [Google Scholar]
- Thorstad, E. B. , Rikardsen A. H., Alp A., and Økland F.. 2013. “The Use of Electronic Tags in Fish Research–an Overview of Fish Telemetry Methods.” Turkish Journal of Fisheries and Aquatic Sciences 13: 881–896. 10.4194/1303-2712-v13_5_13. [DOI] [Google Scholar]
- Thorstad, E. B. , Whoriskey F., Rikardsen A. H., and Aarestrup K.. 2010. “Aquatic Nomads: The Life and Migrations of the Atlantic Salmon.” In Atlantic Salmon Ecology. Aas O, edited by Einum S., Klemetsen A., and Skurdal J., 1–32. Oxford, UK: Wiley‐Blackwell. 10.1002/9781444327755.ch1. [DOI] [Google Scholar]
- Thorstad, E. B. , Whoriskey F., Uglem I., Moore A., Rikardsen A. H., and Finstad B.. 2012. “A Critical Life Stage of the Atlantic Salmon Salmo salar : Behaviour and Survival During the Smolt and Initial Post‐Smolt Migration.” Journal of Fish Biology 81: 500–542. 10.1111/j.1095-8649.2012.03370.x. [DOI] [PubMed] [Google Scholar]
- Thorup, K. , Tøttrup A. P., Willemoes M., et al. 2023. “Resource Tracking Within and Across Continents in Long‐Distance Bird Migrants.” Science Advances 3: e1601360. 10.1126/sciadv.1601360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner, S. D. 2014. “Qqman: An R Package for Visualizing GWAS Results Using Q–Q and Manhattan Plots. bioRxiv:5165.” 10.1101/005165. [DOI]
- Wagner, G. N. , Hinch S. G., Kuchel L. J., et al. 2005. “Metabolic Rates and Swimming Performance of Adult Fraser River Sockeye Salmon ( Oncorhynchus nerka ) After a Controlled Infection With Parvicapsula Minibicornis.” Canadian Journal of Fisheries and Aquatic Sciences 62: 2124–2133. 10.1139/f05-126. [DOI] [Google Scholar]
- Wainwright, P. C. 1994. “Functional Morphology as a Tool in Ecological Research.” In Ecological Morphology: Integrative Organismal Biology, edited by Wainwright P. C. and Reilly S. M., 42–59. Chicago, IL: University of Chicago Press. [Google Scholar]
- Waples, R. S. , Naish K. A., and Primmer C. R.. 2020. “Conservation and Management of Salmon in the Age of Genomics.” Annual Review of Animal Biosciences 8: 117–143. 10.1146/annurev-animal-021419-083617. [DOI] [PubMed] [Google Scholar]
- Webb, P. W. 1978. “Fast‐Start Performance and Body Form in Seven Species of Teleost Fish.” Journal of Experimental Biology 74: 211–226. 10.1242/jeb.74.1.211. [DOI] [Google Scholar]
- Webb, P. W. 1984. “Body Form, Locomotion and Foraging in Aquatic Vertebrates.” American Zoologist 24: 107–120. 10.1093/icb/24.1.107. [DOI] [Google Scholar]
- Webster, M. , and Sheets H. D.. 2010. “A Practical Introduction to Landmark‐Based Geometric Morphometrics.” Quantitative Methods in Paleobiology 16: 168–188. 10.1017/S1089332600001868. [DOI] [Google Scholar]
- Weinz, A. A. , Matley J. K., Klinard N. V., Fisk A. T., and Colborne S. F.. 2020. “Identification of Predation Events in Wild Fish Using Novel Acoustic Transmitters.” Animal Biotelemetry 8: 28. 10.1186/s40317-020-00215-x. [DOI] [Google Scholar]
- Wellband, K. , Mérot C., Linnansaari T., Elliott J. A. K., Curry R. A., and Bernatchez L.. 2019. “Chromosomal Fusion and Life History‐Associated Genomic Variation Contribute to Within‐River Local Adaptation of Atlantic Salmon.” Molecular Ecology 28: 1439–1459. 10.1111/mec.14965. [DOI] [PubMed] [Google Scholar]
- Whelan, K. , Roberts D., and Gray J.. 2019. “The SAMARCH Project International Salmonid Coastal and Marine Telemetry Workshop.” https://www.samarch.org/wp‐content/uploads/2020/05/SAMARCH‐Tracking‐Conference‐Nov‐2019‐final_compressed.pdf.
- Whitlock, M. C. , and Lotterhos K. E.. 2015. “Reliable Detection of Loci Responsible for Local Adaptation: Inference of a Null Model Through Trimming the Distribution of FST.” American Naturalist 186: S24–S36. 10.1086/682949. [DOI] [PubMed] [Google Scholar]
- Wickham, H. 2016. ggplot2: Elegant Graphics for Data Analysis. New York: Springer‐Verlag. [Google Scholar]
- Wilcove, D. S. , and Wikelski M.. 2008. “Going, Going, Gone: Is Animal Migration Disappearing?” PLoS Biology 6: 1361–1364. 10.1371/journal.pbio.0060188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson, R. W. , Wilson J. M., and Grosell M.. 2002. “Intestinal Bicarbonate Secretion by Marine Teleost Fish—Why and How?” Biochimica et Biophysica Acta (BBA) ‐ Biomembranes 1566: 182–193. 10.1016/S0005-2736(02)00600-4. [DOI] [PubMed] [Google Scholar]
- Wittmann, M. J. , Bergland A. O., Feldman M. W., Schmidt P. S., and Petrov D. A.. 2017. “Seasonally Fluctuating Selection Can Maintain Polymorphism at Many Loci via Segregation Lift.” Proceedings of the National Academy of Sciences 114: E9932–E9941. 10.1073/pnas.1702994114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu, H. , Gegear R. J., Casselman A., Kanginakudru S., and Reppert S. M.. 2009. “Defining Behavioral and Molecular Differences Between Summer and Migratory Monarch Butterflies.” BMC Biology 7: 14. 10.1186/1741-7007-7-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data S1.
Data Availability Statement
Data and code used for analyses are publicly available at: https://tinyurl.com/salmon‐migration‐success.