

Abstract
Background
Accumulation of critically short telomeres (CST) is implicated in decreased tissular regenerative capacity and increased susceptibility to degenerative diseases such as Alzheimer’s disease (AD). Telomere shortening has also been associated with age-related brain changes. However, it remains unclear whether CST accumulation is directly associated with AD markers or instead amplifies age-related effects, potentially increasing susceptibility of developing AD in cognitively healthy older adults.
Methods
This cross-sectional study used baseline data of 129 community-dwelling cognitively healthy older adults from the Age-Well trial (NCT02977819), aged 65 years and older enrolled between 2016 and 2018, in France. Using linear regressions, we analyzed the relationship between an innovative marker of telomere shortening, the percentage of CST (%CST), structural, functional and molecular neuroimaging outcomes, and multiple blood-based biomarkers related to AD pathophysiology. The effect of apolipoprotein E ε4 genotype (APOE4) was assessed on these relationships using interaction analysis.
Results
A higher %CST was associated with lower global kurtosis fractional anisotropy (β = -.230; P = .010), particularly in frontal and temporal regions. A higher %CST was also related to higher plasma levels of Neurofilament light chain (β = .195; P = .020) and a lower subiculum volume (β = -.206; P = .020), although these associations did not meet the threshold for multiple comparisons. %CST was not associated with AD-related neuroimaging markers, including the AD-sensitive gray matter pattern (β = -.060; P = .441), glucose metabolism pattern (β = -.099; P = .372), brain perfusion pattern (β = -.106; P = .694) or hippocampus volume (β = -.106; P = .194). In APOE4 carriers, higher %CST was associated with lower subiculum (β = -.423; P = 0.003), DG (β = -.410; P = 0.018) and CA1 volumes (β = -.373; P = 0.024), even though associations with DG and CA1 volumes did not survive multiple comparison.
Conclusions
Although an increase in %CST does not appear to be directly linked to the pathophysiology of AD in cognitively healthy older adults, it could heighten the susceptibility of APOE4 carriers to develop AD plausibly due to greater vulnerability to age-related effects. However, longitudinal studies would be necessary to determine whether %CST influences the development and progression of AD later in life.
Supplementary Information
The online version contains supplementary material available at 10.1186/s13195-024-01635-0.
Keywords: Telomeres, Biomarkers, Neuroimaging, Ageing, Alzheimer’s disease
Introduction
Telomeres are the end of chromosomes and are composed of repetitions of a specific sequence (TTAGGG) [1]. Their main function is to protect the genomic DNA from degradation or chromosomal fusions [2]. However, as cellular divisions occur, telomeres progressively shorten due to the end-replication problem [3] and damaging processes such as oxidative damage [4] and inflammation [5]. In this context, telomere length reflects the mechanism of cellular ageing [3] and might be considered a marker of biological age [6]. In clinical studies, the most commonly used telomere marker is peripheral blood mean leukocyte telomere length, which is a relative parameter that has been proposed to reflect cumulative exposure to stress [7] and inflammation [8]. However, when telomeres reach a critical length, they lose the ability to maintain their structure and protective function, leading to cellular senescence through a persistent DNA damage response [1] and compromised tissue regeneration [9]. The accumulation of critically short telomeres has been associated with increased mortality [10] and increased susceptibility to degenerative diseases in numerous types of tissues (e.g. lung, bone, bone marrow, skin, and immune cells) [11]. While senescence can be triggered even when the mean telomere length is longer than expected, the load of short telomeres has been linked to cellular senescence [12], one of the fundamental mechanisms of ageing. Consequently, this accumulation, measured as the percentage of peripheral blood critically short telomeres in humans (%CST), would be more representative of impaired chromosome stability and cell viability than telomere length [13], although these two parameters remain closely related, exhibiting an inverse association [13, 14].
AD is the first form of dementia in the world, with ageing being the most significant risk factor [15], followed by genetic risk factors, such as the presence of the apolipoprotein E ε4 genotype (APOE4) [15]. The pathological progression of the disease starts decades before clinical diagnosis, underscoring the importance of research on risk factors and subclinical manifestations of AD in cognitively healthy older adults. Although samples (i.e. blood or buccal) from AD patients were found to have shorter telomeres than controls [16–18], the role of telomere attrition in AD pathogenesis has not been determined [1]. Recently, a meta-analysis investigating the relationship between telomere length and brain ageing validated that shorter telomere length was associated with lower total brain and hippocampal volumes [19]. However, the relationship between the %CST and brain integrity, as well as its contribution to AD susceptibility remains unknown. Furthermore, it remains uncertain whether the relationship between %CST and increased AD vulnerability arises from a direct link with AD pathophysiological processes or from a reduction in brain reserve caused by an acceleration of age-related brain alterations, heightening the susceptibility of developing AD.
The aim of this study was to investigate the relationships between the %CST and markers as well as brain regions specific to AD compared to those primarily affected by ageing and less sensitive to the disease. This approach was intended to disentangle the associations linking %CST and the increased vulnerability to develop AD in cognitively healthy older adults. We hypothesize that the %CST will be associated with both AD-specific and non-specific markers. We also assumed that these relationships would be stronger in APOE4 carriers, as the effects of %CST and APOE4 could accumulate and potentiate each other.
Materials and methods
Study population
A total of 129 community-dwelling cognitively healthy older adults were included from the baseline visit of the Age-Well randomized controlled trial of the Medit-Ageing European project (NCT02977819) [20], sponsored by the French National Institute of Health and Medical Research (INSERM). The details of the inclusion and exclusion criteria are described in a previous publication [20] and are listed in supplementary 1. Briefly, participants were aged at least 65 years old, were native French speakers, had retired for 1 year or more, had at least 7 years of education, and had performed within the normal range on standardized cognitive tests. All participants underwent structural MRI, [18F]-Florbetapir (AV45) Positron Emission Tomography (PET), and [18F]-FluoroDeoxyGlucose (FDG)-PET scans, blood sampling and a clinical exam within a 3-month period. Baseline data were collected from November 2016 until April 2018. All participants provided written informed consent for the study, and the Age-Well randomized clinical trial was approved by the ethics committee (CPP Nord-Ouest III, Caen; Clinicaltrials.gov Identifier: NCT02977819; trial registration number: EudraCT: 2016–002441-36; IDRCB: 2016-A01767-44; registration date: 2016–11-25).
Percentage of critically short telomeres
%CST was measured in Peripheral Blood Mononuclear Cells (PBMCs). PBMCs were isolated using the Ficoll method with Histopaque®-1077 Hybri-Max™ from Sigma. The samples were subsequently sent to Life Length (Parque Científico de Madrid Calle Faraday, 7; Campus de Cantoblanco 28,049 Madrid SPAIN), where %CST was determined using the Telomere Analysis Technology (TAT) with the high-throughput quantitative fluorescence in situ hybridization (HT Q-FISH) technique as described previously [21]. This technique allows for the measurement of individual telomere lengths, enabling the determination of %CST, the frequency of telomeres shorter than 3,000 base pairs, previously established as the cutoff for “short telomeres” in humans [10, 22].The entire procedure is detailed in supplementary 2.
APOE4 genotype and blood-based markers related to neurodegeneration and AD physiopathology
The plasma concentrations of β-amyloid 40 and 42 (Aβ40 and 42) were measured using an ultrasensitive electrochemiluminescence measurement technique (Meso Scale Discovery, MSD, Rockville, Marylan, USA). Plasma phosphorylated-Tau 181 (p-Tau181), Glial Fibrillary Acidic Protein (GFAP) and Neurofilament Light chain (NfL) were measured using SIMOA technology with commercial kits from Quanterix (p-Tau181 V2 Advantage #103,714, Neurology 2-plexB #103,520) on an HD-X analyzer (Quanterix, Lexington, MA) (PMID: 20,495,550). Details on plasma marker assessments are available in supplementary 2. APOE genotype was determined using a standardized protocol described in supplementary 2. Participants with at least one ε4 allele were considered APOE4 positive, and the others negative.
Neuroimaging examinations
Participants underwent structural T1, Fluid-Attenuated Inversion Recovery (FLAIR) and Diffusion Kurtosis Imaging (DKI) MRI, as well as FDG and AV45-PET scans (early and late acquisitions), to measure Grey Matter (GM) volume, White Matter (WM) integrity (i.e. Mean Kurtosis [MK] and kurtosis Fractional Anisotropy [kFA]), glucose metabolism, brain perfusion, and amyloid burden, respectively. FDG-PET was available only for 88 participants. Two ultra-high-resolution T2-weighted structural images were also acquired perpendicular to the long axis of the hippocampus. All examinations were performed at the Cyceron center (Caen, France). Averaged global GM volume, glucose metabolism, MK, and kFA values were obtained by applying a binary mask of either global GM or WM depending on the neuroimaging modality, on the corresponding preprocessed images. Individual global cortical amyloid load was extracted from a predetermined neocortical mask (including the entire GM, except the cerebellum, occipital and sensorimotor cortices, hippocampi, amygdala and basal nuclei) [23]. Averaged GM volume was also extracted by applying a binary mask characterizing AD alterations (most representative areas: temporal lobe, notably the parahippocampal gyrus, hippocampus, amygdala, and fusiform gyrus) from a previous study [24]. Brain perfusion and glucose metabolism were also extracted by applying a binary mask characterizing AD signature (most representative areas: posterior cingulate and temporoparietal cortex) from a previous study [25]. Hippocampal subfields (Cornu Ammonis [CA1, CA2, CA3], dentate gyrus [DG], subiculum) volumes were automatically estimated on ultra-high-resolution T2-weighted images using the Automated Segmentation for Hippocampal Subfields (ASHS) software along with a custom atlas [26–28]. The hippocampus volume was extracted using ASHS-T1 [29], on structural T1. Details regarding all neuroimaging procedure are described in supplementary 2.
Statistical analyses
The study design is described in a flow diagram (Fig. 1). Demographic statistics were presented using the mean and Standard Deviation (SD), while qualitative variables were expressed as counts and percentages. Linear regressions were conducted using RStudio software with each neuroimaging or blood-based marker as a dependent variable, %CST as an independent variable and demographics (i. e. age, sex, education, and Body Mass Index (BMI)) as covariates. To further assess the regional specificity of the association between %CST and a specific neuroimaging modality, when a significant association was found with the global value, the corresponding analysis was repeated using a voxelwise approach on SPM12, controlling for the same covariates. Results were evaluated for significance at puncorrected < 0.005 combined with a minimum cluster size determined by Monte‐Carlo simulations using the AFNI’s 3dClustSim program to achieve a corrected statistical significance of p < 0.05. To assess the impact of the APOE4 status, interactions between %CST and APOE4 status were performed for each neuroimaging and blood-based markers with the same covariates as in previous analyses. When the interaction was significant, post-hoc linear regression analyses were conducted separately in APOE4 carriers and non-carriers. For %CST and GFAP values, an ANCOVA was carried out to evaluate the APOE4 effect controlling for age, sex, education and BMI.
Fig. 1.

Flow diagram of the study. Abbreviations: DKI, Diffusion Kurtosis Imaging; GFAP, Glial Fibrillary Acidic Protein; p-Tau181, phosphorylated-Tau181, Aβ; β-Amyloid
The significance level was set at p < 0.05 for all statistical analyses except voxel-wise analyses. Bonferroni correction was then applied to control for multiple comparisons so that results surviving a P-value ≤ (0.05/number of comparisons) were indicated. But we also considered uncorrected results to prevent overlooking biologically relevant associations that may not survive Bonferroni correction. %CST was z-scored to address multicollinearity, particularly with APOE4. However, since this transformation did not affect the distribution or the p-values, the graphics were presented using raw values to provide a biologically interpretable representation. All analyses were replicated, additionally adjusting for the APOE4 status. As plasma NfL and p-Tau181 were previously described as being impacted by kidney function [30, 31], analyses involving these markers were also replicated, adding glomerular filtration rate as a covariate. Telomere length can be influenced by blood cells composition, so all analyses were replicated with lymphocyte concentration, the predominant cell type in PBMC, added as covariate. NfL, GFAP and p-Tau181 values were log transformed. Results remained unchanged with raw values. All analyses were performed without one %CST outlier, identified as more than three SD from the mean. However, as the results remained unchanged, the outlier was not removed.
Results
The baseline demographic characteristics of the population are presented in Table 1. The participants ranged in age from 65 to 83 years, with a mean age of 68.84 years. The proportion of females was higher than the proportion of males. A total of 27.13% of the population were APOE4 carriers.
Table 1.
Demographics of the study participants
| Demographics (n = 129) | Mean (SD) |
| Age, years | 68.84 (3.69) |
| Female Sex, No. (%) | 78 (60.47) |
| Education, years | 13.15 (3.14) |
| Body Mass Index, kg/m2 | 26.20 (4.31) |
| APOE4 carriers, No. (%) | 35 (27.13) |
| Short telomeres (< 3 kb), % | 9.86 (3.07) |
| Global neuroimaging outcomes (n = 127) | |
| Global GM volume, mm3 | 546,614.414 (25,805.89) |
| Global glucose metabolism, SUVr (n = 88) | 1.11 (0.06) |
| Neocortical Amyloid load, SUVr | 1.24 (0.15) |
| WM kurtosis fractional anisotropy | 0.35 (0.02) |
| WM mean kurtosis | 0.96 (0.04) |
| Hippocampal subfields normalized volumes (n = 125) | |
| CA1 | 0.95 (0.11) |
| CA2 | 0.02 (0.003) |
| CA3 | 0.05 (0.01) |
| Dentate gyrus | 0.41 (0.06) |
| Subiculum | 0.32 (0.04) |
| Blood-based markers (n = 122) | |
| NfL (pg/mL) | 20.18 (7.66) |
| GFAP (pg/mL) | 155.54 (60.96) |
| p-Tau181 (pg/mL) | 1.95 (0.79) |
| Ratio Aβ42/40 | 0.07 (0.03) |
| Aβ42 (pg/mL) | 10.36 (4.29) |
| Aβ40 (pg/mL) | 156.40 (39.71) |
Abbreviations: SD Standard deviation, BMI Body mass index, APOE4 Apolipoprotein E ε4, GM Gray Matter, SUVr Standardized uptake value, WM White Matter, CA Cornu Ammonis, NfL Neurofilament light chain, GFAP Glial fibrillary acidic protein, Aβ β-amyloid
Multiple regressions between %CST and neuroimaging values
The %CST was negatively associated with the global WM kFA (β = -0.230; 95%CI[-0.234;-0.226]; P = 0.010), and this result survived Bonferroni correction (Table 2).
Table 2.
Association of %CST with global neuroimaging outcomes
| %CST (z-scored) | ||||
|---|---|---|---|---|
| N | Standardized β coefficient | 95% CI | P-value | |
| Global GM volumea | 127 | -.014 | [-3856.3; 3856.3] | .858 |
| Global Glucose metabolisma | 88 | -.088 | [-.103;-.074] | .427 |
| Neocortical Amyloid loadb | 127 | -.173 | [-.200;-.145] | .060 |
| WM kurtosis fractional anisotropyc | 127 | -.230 | [-.234;-.226] | .010** |
| WM mean kurtosisc | 127 | -.080 | [-.088; -.073] | .371 |
*P < 0.05. **P < 0.01. ***P < 0.001. The results are presented from linear regression after adjusting for age, sex, education and BMI. Bonferroni correction for multiple testing (p = 0.05/5 for the five global neuroimaging outcomes) is indicated in bold. Abbreviations: %CST Percentage of critically short telomeres, GM Gray matter, WM White matter, BMI Body mass index, CI Confidence interval
The %CST was negatively associated with WM kFA in frontal regions, including part of the superior frontal white matter, the cingulum and the corpus callosum, bilaterally. We also found a negative association in the left middle temporal white matter, notably in a part of the uncinate, the inferior longitudinal fasciculus, and the fornix (Fig. 2). The %CST was negatively associated with neocortical amyloid load at a trend-level statistical significance (β = -0.173; 95%CI[-0.200;-0.145]; P = 0.060). The %CST was not significantly associated with other global neuroimaging values (Table 2).
Fig. 2.
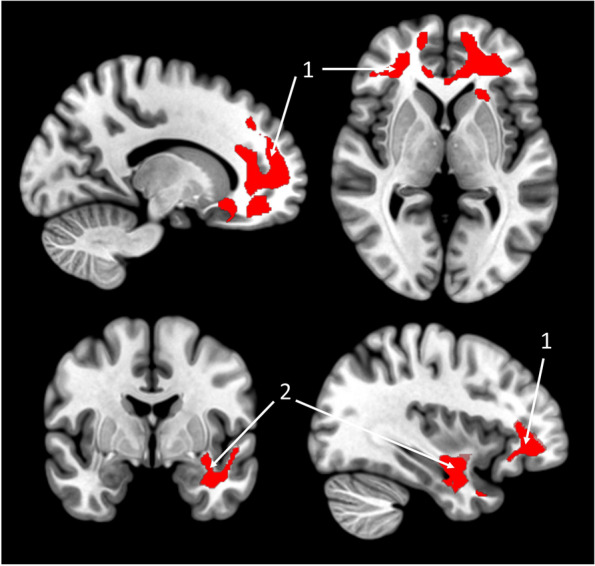
Voxelwise associations between the %CST and kFA. Negative voxel-wise multiple regression between %CST and kFA are presented, controlling for age, sex, education and BMI, in 127 healthy older adults. Results are presented at a puncorrected < 0.005 threshold combined with a cluster-level multiple comparisons correction. (1) Superior frontal gyrus white matter, Cingulum, Corpus Callosum; (2) Left inferior longitudinal fasciculus, uncinate fasciculus, fornix. Abbreviations: %CST, Percentage of Critically Short Telomeres; kFA, kurtosis Fractional Anisotropy; BMI, Body mass index
We did not find any associations between %CST and the AD-sensitive pattern of GM volume, glucose metabolism, brain perfusion, or hippocampal volume (Table 1 in supplementary 3).
The %CST was associated with subiculum volume (β = -0.206; 95%CI [-0.213;-0.200]; P = 0.020), but not with the volume of the other hippocampal subfields (Fig. 3A, Table 2 in supplementary 3). This association did not survive Bonferroni correction. The results remained unchanged when APOE4 status or lymphocyte concentration was added as a covariate (data not shown).
Fig. 3.

Associations of the %CST with hippocampal subfield volumes, in cognitively healthy older adults and according to APOE4 status. A Scatterplots of linear regression between %CST and hippocampal subfield volumes, in 125 healthy older adults. B Scatterplots of linear regression between %CST and hippocampal subfields volumes, according to APOE4 status. Interaction and Post-hoc analyses are indicated. Analysis are corrected by age, sex, education and BMI. Detailed statistics of the analyses are summarized in Table 6 and Table 8 in supplementary 3. 3D representations of hippocampus subfields were obtained using 3D slicer software based on an ASHS segmentation. All hippocampal subfield volumes are TIV normalized. *P < 0.05. **P < 0.01. ***P < 0.001. Abbreviations: %CST, Percentage of critically short telomeres; APOE4, Apolipoprotein E ε4; CA, Cornu Ammonis
Multiple regressions between %CST and blood-based markers related to neurodegeneration and AD physiopathology
A positive relationship was found between the %CST and plasma NfL levels (log transformed) (β = 0.195; 95%CI [0.170; 0.220]; P = 0.020). Plasma Aβ42 levels were positively associated with %CST (β = 0.191; 95%CI [-0.595; 0.978]; P = 0.041). The %CST was positively associated with Aβ42/40 ratio at a trend-level statistical significance (β = 0.183; 95%CI[0.178;0.188]; P = 0.053). %CST was not significantly associated with other blood markers (Fig. 4A, Table 3 in supplementary 3). These relationships did not survive Bonferroni correction. All the results remained consistent after further adjustment for APOE4 status, glomerular filtration rate or lymphocyte concentration (data not shown).
Fig. 4.

Associations of the %CST with neurodegeneration and AD physiopathology-related blood-based markers in cognitively healthy older adults and according to APOE4 status. A Scatterplots of linear regression between %CST and blood-based markers, in 122 healthy older adults. B Scatterplots of linear regression between %CST and blood-based markers, according to APOE4 status. Interaction and post-hoc analyses are indicated. Analysis are corrected by age, sex, education and BMI. Detailed statistics of the analyses are summarized in Table 7 and Table 9 in supplementary 3. *P < 0.05. **P < 0.01. ***P < 0.001. Abbreviations: %CST, Percentage of Critically Short Telomeres; APOE4, Apolipoprotein E ε4; NfL, Neurofilament Light chain, GFAP; Glial Fibrillary Acidic Protein, p-Tau181; phosphorylated-Tau181, Aβ; β-Amyloid
Interaction between %CST and APOE4
There was no group difference in %CST between APOE4-carriers and non-carriers (Fig 1 in supplementary 3). We found significant interactions between %CST and APOE4 status on hippocampal volume (β = -0.202; 95%CI[-158.390; 157.986]; P = 0.019) (Table 5 and Fig. 2 in supplementary 3) and on the volumes of CA1 (β = -0.263; 95%CI[-0.313; -0.213]; P = 0.005), CA2 (β = -0.207; 95%CI[-0.208; -0.205]; P = 0.029), dentate gyrus (β = -0.297; 95%CI[-0.323; -0.272]; P = 0.002), and subiculum (β = -0.238; 95%CI[-0.255; -0.221]; P = 0.010) (Fig. 3B, Table 6 in supplementary 3). All the %CST x APOE4 status interactions survived Bonferroni correction except for hippocampus and CA2 volumes. %CST also interacted with APOE4 status on plasma GFAP levels (β = 0.225; 95%CI[0.149; 0.301]; P = 0.012) (Fig. 4B, Table 7 in supplementary 3). This interaction did not survive Bonferroni correction. Post-hoc analyses revealed that, in APOE4 carriers only, %CST was negatively associated with CA1 (β = -0.373; 95%CI[-0.428; -0.319]; P = 0.024), dentate gyrus (β = -0.410; 95%CI[-0.440;-0.379]; P = 0.018), subiculum volume (β = -0.423; 95%CI[-0.436;-0.409]; P = 0.003) and tented to be associated to plasma GFAP level β = -0.245; 95%CI[0.173;0.316]; P = 0.071). The results of the post-hoc analyses are indicated in the figures and detailed statistics are presented in Table 8 and 9 in supplementary 3. There was no group difference in GFAP levels (log transformed) between APOE4-carriers and non-carriers (Fig. 3 in supplementary 3). No interaction effect was found between the %CST and APOE4 status on other neuroimaging values or blood-based markers (Table 4 and 5 in supplementary 3). Results remained unchanged after controlling for lymphocyte concentration.
Discussion
The objective of the present study was to provide an overview of the association between %CST and age- or AD-related blood and brain imaging markers in cognitively healthy older adults, as well as in individuals at genetic risk for AD (APOE4 carriers) to disentangle the mechanisms linking %CST to an increased vulnerability to develop AD.
Altogether, our results highlight that the %CST is associated with age-related blood and brain imaging markers rather than directly with AD pathological processes. However, the %CST appears to be implicated in a higher vulnerability to AD in APOE4 carriers and is particularly related to the integrity of WM microstructures (kFA).These microstructures are strongly affected by age, particularly in frontal and temporal regions which undergo the most significant changes with age [32, 33]. The regions showing significant associations with the %CST in our study largely coincide with those reported in other studies. Indeed, shorter telomere length was associated with lower FA in the fornix [34], corpus callosum [35], and inferior and superior longitudinal fasciculus [35].This result is further supported by the association found between %CST and blood level of NfL, a well-established marker of neurodegeneration and axonal injury that increases with age [36, 37], although this association did not remain significant after correction for multiple comparisons. It has previously been highlighted that telomere length was negatively associated with cerebrospinal fluid NfL levels [38], and cerebrospinal fluid NfL levels were negatively correlated with cerebral mean FA [39]. Furthermore, shorter telomere length and lower WM integrity has been associated with higher blood level of several inflammatory mediators [8, 40–42]. Additionally, it is already known that one or a few short telomeres can impose senescence, leading to the secretion of proinflammatory cytokines, known as the senescence-associated secretory phenotype [1, 43], suggesting that systemic inflammation could be one of the possible mechanisms triggering the detrimental impact of the %CST on WM integrity. Furthermore, immunosenescence, while being a multifactorial phenomenon, is closely related to telomere attrition, particularly due to the high proliferative potential of immune cells [44, 45]. Peripheral age-related immunosenescence, together with a chronic, low-grade inflammation known as “inflammageing”, has been suggested to alter immune responses within the brain and exacerbate microglial senescence [46–48]. These changes may contribute to the neuroinflammation that enhances brain ageing and neurodegeneration [47, 48]. Similarly, activated glial cells in the brain during neurodegeneration could also influence both brain and peripheral immune cells by secreting pro-inflammatory mediators, promoting immune cells infiltration [49, 50]. Several additional studies examine these mechanisms from a bidirectional perspective, highlighting the complex interplay between the brain and immune system, revealing potential pathways through which systemic and brain ageing processes may be interconnected [49]. Evidence suggests that, although variable, telomere lengths are correlated across tissues, with blood telomere length serving as a proxy for telomere length in various brain regions, such as the hippocampus and cortex [51]. According to post mortem studies, telomere length in brain cells is reportedly associated with age only in WM, and not in GM [52], and telomere length is shorter in WM glial cells than in GM glial cells in adults [53]. Interestingly, unlike GM, the WM is predominantly composed of mitotic cells that undergo telomere attrition [53]. The difference in mitotic activity between GM and WM cells might explain why the %CST is preferentially associated with WM rather than GM integrity and could play a role in the age-related loss of WM integrity. Some studies also revealed a preferential link between telomere length and WM volume and integrity, rather than GM [54, 55]. However, there are also studies demonstrating a consistent association between telomere length and GM volume [56], notably the hippocampal volume [19]. In our study, the %CST did not appear to be related to GM integrity, except for the subiculum, which is the hippocampal subfield most affected by ageing [57]. However, this association did not reach the multiple comparison threshold.
Interestingly, our results suggest that %CST could potentiate the effect of APOE4 on hippocampal volume, especially CA1, DG, and subiculum volumes, potentially increasing the risk of developing AD, as both APOE4 and hippocampus atrophy are risk factors for AD [15]. To date, no study has assessed the relationship between hippocampal subfield volumes and %CST or leukocyte telomere length in APOE4 carriers. Only one study reported that older APOE4 carriers exhibited longer telomere length than noncarriers [58], a finding that we did not replicate with %CST in our study. However, our results appear to support and extend the findings of other studies. Indeed, studies have demonstrated that APOE4 carriers exhibit a lower hippocampal volume compared to non-carriers [59], lower cortical thickness in the subiculum, lower DG volumes [57], and APOE4 interacts with age on CA1, subiculum, and whole hippocampus volumes [28]. APOE4 has been associated with an elevated risk of vascular pathology, a condition known to raise the likelihood of developing AD [60]. It is also linked to higher blood brain barrier leakage, particularly in the medial temporal lobe and hippocampus [61, 62], which could potentially heighten the vulnerability of these regions to the detrimental effects of peripheral %CST.
In APOE4 carriers, a higher %CST was also associated with a higher GFAP blood level, which is a marker of astrocyte activation [63]. This relationship, which did not survive multiple comparison, could be explained by increased levels of GFAP in APOE4 carriers, but there is no consensus on these results. One study with 709 participants showed a difference in GFAP between carriers and noncarriers [64], while another study with 88 participants, similarly to the current study, showed no difference [65]. However, the APOE4 genotype is known to be associated with increased GFAP expression at the blood brain barrier level [66], which could be further amplified by systemic inflammation resulting from telomere shortening [67]. Simultaneously, the increased BBB permeability observed in APOE4 carriers [62] could explain why the relationship between %CST and higher GFAP blood levels is observed primarily in APOE4 carriers.
Contrary to our hypothesis, %CST does not appear to be directly related to AD pathophysiological processes but could potentially lead to an acceleration of brain ageing processes and therefore be implicated in a greater susceptibility to AD, especially individuals at genetic risk for AD, the APOE4 carriers. Thus, in our study, the %CST was not specifically associated with AD markers, i.e. increases in amyloid and tau pathologies, nor with a pattern of neurodegeneration specific to AD. Previous studies reported shorter telomere length in samples from AD patients compared to controls [16]. However, there is no evidence that telomere shortening is specific to AD. Indeed, leukocyte telomere length was not found to be relevant for discriminating between different types of dementia [68]. Additionally, brain regions associated with telomere length in healthy adults are not restricted to regions typically affected by amyloid or tau pathologies in AD, furthermore affecting, for example, the thalamus or the fusiform cortex [19, 56]. Notably, instead of revealing a link between higher %CST and higher amyloid or tau pathologies, we observed trends toward an association between lower %CST and higher blood levels of Aβ42, Aβ42/40 ratio and a lower neocortical amyloid load. Another study in healthy older adults also demonstrated that shorter telomere length was associated with higher cerebrospinal fluid Aβ42/40 ratio [38]. A reduction of amyloid load was also observed in an AD mouse model with short telomeres [69]. The association between telomere shortening and amyloid pathology remains unclear, and further studies are needed to validate and elucidate these unexpected results.
Strengths and limitations
This study is the first, to our knowledge, to investigate the association of %CST, an innovative and absolute marker of telomere shortening, with brain imaging and blood markers. In comparison to the most commonly used telomere measure, which allows for relative quantification of telomere length through a ratio between telomere signal to a reference single copy gene signal (i.e. T/S ratio), telomere length measured by HT Q-FISH provides an absolute quantification of individual telomeres [70]. These markers include age- and AD-related markers, with a multimodal approach including structural, diffusion, functional, and molecular neuroimaging, as well as multiple blood-based markers. Another major strength is the use of a tailored method to estimate hippocampal subfield volumes, consisting of dedicated ultra-high-resolution T2-weighted images along with the ASHS algorithm and a custom atlas based on ex-vivo MRI and histology data. This study has also some limitations, including the cross-sectional nature of the design which prevents us from inferring causal relationships regarding the association of %CST with ageing or AD-related markers. Additionally, we drew a clear distinction between ageing and AD- related markers, though these two phenomena likely exist along a continuum. Moreover, this study is a secondary outcome of a clinical trial [71] originally designed for another objective. In this context, our analyses may have been underpowered to observe weak associations. This study found no significant association between %CST and AD-related markers. However, due to the specific characteristics of our analyses and population size, we may not have been able to fully capture or highlight the real absence of an association. Additionally, participants from the Age-Well trial are in above-average health for their age [71], which may limit the generalizability of our findings and contribute to some of the null results observed. Longitudinal studies are also needed to further disentangle the mechanisms underlying the link between %CST, brain ageing, and greater risk of AD.
Conclusion
Our results support the hypothesis that the %CST is associated with age-related alterations in WM integrity in cognitively healthy older adults. The accumulation of CST does not appear to be directly related to AD pathophysiological processes but may contribute to a higher vulnerability of developing AD, particularly in APOE4 carriers. This increased vulnerability could stem from a heightened sensitivity to age-related effects, resulting in reduced brain reserve and, consequently, an elevated susceptibility to AD.
Supplementary Information
Acknowledgements
The authors thank all the member of the Medit-Ageing Research Group: Claire André, Eider M. Arenaza-Urquijo, Pierre Champetier, Gaël Chételat, Anne Chocat, Fabienne Collette, Robin de Florès, Vincent de La Sayette, Marion Delarue, Francesca Felisatti, Egantine Ferrand Devouge, Antoine Garnier-Crussard, Julie Gonneaud, Marc Heidmann, Frank Jessen, Olga Klimecki, Elizabeth Kuhn, Brigitte Landeau, Gwendoline Le Du, Valérie Lefranc, Antoine Lutz, Natalie L. Marchant, Florence Mézenge, Inès Moulinet, Valentin Ourry, Cassandre Palix, Léo Paly, Géraldine Poisnel, Stefano Poletti, Anne Quillard, Géraldine Rauchs, Stéphane Réhel, Eric Salmon, Siya Sherif, Edelweiss Touron, Matthier Vanhoutte, Denis Vivien, Patrik Vuilleumier, Caitlin Ware and Miranka Wirth. The authors also thank Marine Faure, Jeanne Lepetit, Marie Saville, and the Cyceron MRI-PET staff members for their assistance with recruitment and neuroimaging data acquisition; INSERM administrative, financial, and legal departments; INSERM Transfert (Delphine Smagghe) and Aurélia Cognet for administrative support; the sponsor (Pole de Recherche Clinique at INSERM, Dr. Hélène Espérou); the Euclid Team (Dr Eric Frison) for data management; and the participants of the study and their families.
The Medit-Ageing Research Group: list of Authors and affiliations:
Claire André, PhD1, Eider M. Arenaza-Urquijo, PhD7, Pierre Champetier PhD1, Gaël Chételat, PhD1, Anne Chocat, MD1, Fabienne Collette, PhD8−19, Robin de Florès, PhD1, Vincent de La Sayette MD3, Marion Delarue, MSc1, Francesca Felisatti, PhD1, Egantine Ferrand-Devouge, MD1,9, Antoine Garnier-Crussard, MD-MSc1,10, Julie Gonneaud, PhD1, Marc Heidmann, PhD11, Frank Jessen, MD-PhD12,13, Olga Klimecki, PhD14, Elizabeth Kuhn, PhD12,15, Brigitte Landeau, PhD1, Gwendoline Le Du, MSc1, Valérie Lefranc, BA1, Antoine Lutz, PhD16, Natalie L. Marchant, PhD17, Florence Mezenge, BA1, Inès Moulinet, PhD1, Valentin Ourry, PhD1, Cassandre Palix, PhD1, Léo Paly, MSc1, Géraldine Poisnel, PhD1, Stefano Poletti, MSc16, Anne Quillard, PhD1, Géraldine Rauchs, PhD1, Stéphane Réhel, PhD1, Eric Salmon, MD-PhD8−18, Siya Sherif, PhD8, Edelweiss Touron, PhD1, Matthieu Vanhoutte, PhD1, Denis Vivien, PhD1−4, Patrik Vuilleumier, MD20, Caitlin Ware, MSc1−21, and Miranka Wirth, PhD22.
7Environment and Health Over the Life Course Programme, Climate, Air Pollution, Nature and Urban Health Programme, Barcelona Institute for Global Health (ISGlobal), Barcelona, Spain
8Centre de recherche du Cyclotron, Université de Liège, 4000 Liège, Belgique
9Centre d’Investigation Clinique-Centre de Recherche Biologique 1404, Rouen University Hospital, Rouen, France
10Clinical and Research Memory Center of Lyon, Lyon Institute For Aging, Charpennes Hospital, Hospices Civils de Lyon, Lyon, France
11Institut National de la Santé et de la Recherche Médicale, Lyon, France
12German Center for Neurodegenerative Diseases (DZNE), Bonn, Bonn, Germany
13Department of Psychiatry, University of Cologne, Medical Faculty, Cologne, Germany
14Interim Chair of Biological Psychology, Technische Universität Dresden, Germany
15Department of Old Age Psychiatry and Cognitive Disorders, University of Bonn Medical Center, Bonn, Germany
16EDUWELL team, Lyon Neuroscience Research Centre, INSERM U1028, CNRS UMR 5292, Lyon 1 University, Lyon, France
17Division of Psychiatry, University College London, London, UK
18Centre de la mémoire, université de Liège, CHU, 4000 Liège, Belgique
19Département de psychologie, cognition et comportement, université de Liège, 4000 Liège, Belgique
20Department of Neurosciences and Clinic of Neurology, Laboratory for Neurology and Imaging of Cognition, University of Geneva, 1211 Geneva, Switzerland
21CRPMS, Université de Paris, Paris, France
22German Center for Neurodegenerative Diseases, Dresden, Germany
Abbreviations
- Aβ
β-Amyloid
- AD
Alzheimer’s disease
- AFNI
Analysis of Functional NeuroImages
- APOE4
Apolipoprotein E ε4
- ASHS
Automatical Segmentation for Hippocampal Subfields
- AV45
[18F]-Florbetapir
- BMI
Body Mass Index
- CA
Cornu Ammonis
- CI
Confidence Interval
- CST
Critically Short Telomeres
- DG
Dentate Gyrus
- DKI
Diffusion Kurtosis Imaging
- DNA
DesoxyriboNucleic Acid
- FDG
[18F]-FluoroDeoxyGlucose
- FLAIR
Fluid-Attenuated Inversion Recovery
- GFAP
Glial Fibrillary Acidic Protein
- GM
Grey Matter
- HT Q-FISH
High-Throughput Quantitative Fluorescence In Situ Hybridization
- kFA
kurtosis Fractional Anisotropy
- MK
Mean Kurtosis
- MRI
Magnetic Resonance Imaging
- NfL
Neurofilament Light Chain
- PBMCs
Peripheral Blood Mononuclear Cell
- PET
Positron emission tomography
- pTau181
phosphorylated-Tau181
- SD
Standard Deviation
- SUVr
Standardized Uptake Value ratio
- TAT
Telomere Analysis Technology
- WM
White Matter
Authors’ contributions
L.A. analyzed the data and wrote the manuscript. P.G. conceptualized the study, supervised the analysis, and wrote the manuscript. C.G. conceptualized the study and reviewed the manuscript. K.P. conceptualized the study and reviewed the manuscript. D.F.R. extracted all hippocampus and subfields volumes and reviewed the manuscript. P.C., T.E., T.A.L. and F.S. collected neuroimaging and blood data and reviewed the manuscript. M.F. and L.B. pre-treated all the neuroimaging images and reviewed the manuscript. V.A. and P.C. performed blood-based biomarkers assays and reviewed the manuscript. C.A. conducted inclusion and follow-up visits for the participants and reviewed the manuscript. D.L.S.V. is the principal investigator of the trial and reviewed the manuscript. V.D. reviewed the manuscript. All authors reviewed and approved the final version of the manuscript. The Medit-Ageing Research Group conceptualized the Age-Well clinical Trial and collected data.
Funding
This work was supported by the European Union’s Horizon 2020 Research and Innovation Program [grant 667696], the Institut National de la Santé et de la Recherche Médicale, the Région Normandie (Label d’excellence), the Fondation d’Entreprise MMA des Entrepreneurs du Futur and the ministry of Higher Education and Research. The funders and sponsor had no role in the design and conduct of the study; collection, management, analysis, and interpretation of the data; preparation, review, or approval of the manuscript; and decision to submit the manuscript for publication.
Data availability
The data underlying this report are made available on request following a formal data sharing agreement and approval by the consortium and executive committee https://silversantestudy.eu/2020/09/25/data-sharing). The Material can be mobilized, under the conditions and modalities defined in the Medit-Ageing Charter, by any research team belonging to an academic for carrying out a scientific research project relating to the scientific theme of mental health and well-being in older people. The Material may also be mobilized by non-academic third parties, under conditions, in particular financial, which will be established by separate agreement between Inserm and by the said third party. Data sharing policies described in the Medit-Ageing Charter comply our ethics approval and guidelines by our funding body.
Declarations
Ethics approval and consent to participate
All participants gave their written informed consent to participate in the study. The Age-Well RCT, sponsored by Institut National de la Santé et de la Recherche Médicale (INSERM), was approved by the ethics committee (CPP Nord-Ouest III, Caen).
Competing interests
Dr Chételat reported receiving grants, personal fees, and nonfinancial support from the Institut National de la Santé et de la Recherche Médicale (INSERM), as well as grants from the European Union's Horizon 2020 Research and Innovation Program (grant 667696) during the conduct of the study and grants and personal fees from Fondation Entrepreneurs MMA and Fondation Alzheimer and grants from MMA and Région Normandie outside the submitted work. Dr Poisnel reported grants and nonfinancial support from INSERM grants from European Union's Horizon 2020 Research and Innovation Program [grant 667696] during the conduct of the study and grants from Région Normandie, Ministry of Higher Education and Research and INSERM outside the submitted work; and has participated in the data safety monitoring board of the Age-Well trial and to the executive committee of Medit-Ageing. No other disclosures were reported.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Géraldine Poisnel, Email: poisnel@cyceron.fr.
the Medit-Ageing Research Group:
Cassandre Palix, Edelweiss Touron, Brigitte Landeau, Anne Chocat, Denis Vivien, Vincent de La Sayette, Gaël Chételat, Géraldine Poisnel, Claire André, Eider M. Arenaza-Urquijo, Pierre Champetier, Fabienne Colette, Robin De Flores, Marion Delarue, Francesca Felisatti, Egantine Ferrand Devouge, Antoine Garnier-Crussard, Julie Gonneaud, Marc Heidmann, Frank Jessen, Olga Klimecki, Elizabeth Kuhn, Gwendoline Le Du, Valérie Lefranc, Antoine Lutz, Natalie Marchant, Florence Mezenge, Inès Moulinet, Valentin Ourry, Léo Paly, Stefano Poletti, Anne Quillard, Géraldine Rauchs, Stéphane Rehel, Eric Salmon, Siya Sherif, Matthieu Vanhoutte, Patrik Vuilleumier, Caitliin Ware, and Miranka Wirth
References
- 1.Rossiello F, Jurk D, Passos JF, et al. Telomere dysfunction in ageing and age-related diseases. Nat Cell Biol. 2022;24:135–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.de Lange T. Protection of mammalian telomeres. Oncogene. 2002;21:532–40. [DOI] [PubMed] [Google Scholar]
- 3.Ohki R, Tsurimoto T, Ishikawa F. In Vitro Reconstitution of the End Replication Problem. Mol Cell Biol. 2001;21:5753–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ahmed W, Lingner J. Impact of oxidative stress on telomere biology. Differentiation. 2018;99:21–7. [DOI] [PubMed] [Google Scholar]
- 5.Jurk D, Wilson C, Passos JF, et al. Chronic inflammation induces telomere dysfunction and accelerates ageing in mice. Nat Commun. 2014;5:4172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Maximov VN, Malyutina SK, Orlov PS, et al. Leukocyte telomere length as an aging marker and risk factor for human age-related diseases. Adv Gerontol. 2017;7:101–6. [Google Scholar]
- 7.Mayer SE, Prather AA, Puterman E, et al. Cumulative lifetime stress exposure and leukocyte telomere length attrition: The unique role of stressor duration and exposure timing. Psychoneuroendocrinology. 2019;104:210–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.O’Donovan A, Pantell MS, Puterman E, et al. Cumulative inflammatory load is associated with short leukocyte telomere length in the health, aging and body composition study. PLoS ONE. 2011;6:e19687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Blasco MA. Telomere length, stem cells and aging. Nat Chem Biol. 2007;3:640–9. [DOI] [PubMed] [Google Scholar]
- 10.Vera E, Bernardes de Jesus B, Foronda M, et al. The Rate of Increase of Short Telomeres Predicts Longevity in Mammals. Cell Rep. 2012;2:732–7. [DOI] [PubMed] [Google Scholar]
- 11.Armanios M, Blackburn EH. The telomere syndromes. Nat Rev Genet. 2012;13:693–704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bendix L, Horn PB, Jensen UB, et al. The load of short telomeres, estimated by a new method, Universal S℡A, correlates with number of senescent cells. Aging Cell. 2010;9:383–97. [DOI] [PubMed] [Google Scholar]
- 13.Hemann MT, Strong MA, Hao L-Y, et al. The shortest telomere, not average telomere length, is critical for cell viability and chromosome stability. Cell. 2001;107:67–77. [DOI] [PubMed] [Google Scholar]
- 14.Fernández-Alvira JM, Fuster V, Dorado B, et al. Short telomere load, telomere length, and subclinical atherosclerosis. J Am Coll Cardiol. 2016;67:2467–76. [DOI] [PubMed] [Google Scholar]
- 15.Winblad B, Amouyel P, Andrieu S, et al. Defeating Alzheimer’s disease and other dementias: a priority for European science and society. Lancet Neurol. 2016;15:455–532. [DOI] [PubMed] [Google Scholar]
- 16.Forero DA, González-Giraldo Y, López-Quintero C, et al. Meta-analysis of telomere length in Alzheimer’s disease. J Gerontol Ser A. 2016;71:1069–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Crocco P, De Rango F, Dato S, et al. The shortening of leukocyte telomere length contributes to Alzheimer’s disease: further evidence from late-onset familial and sporadic cases. Biology. 2023;12:1286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Scheller Madrid A, Rasmussen KL, Rode L, et al. Observational and genetic studies of short telomeres and Alzheimer’s disease in 67,000 and 152,000 individuals: a Mendelian randomization study. Eur J Epidemiol. 2020;35:147–56. [DOI] [PubMed] [Google Scholar]
- 19.Gampawar P, Schmidt R, Schmidt H. Telomere length and brain aging: A systematic review and meta-analysis. Ageing Res Rev. 2022;80:101679. [DOI] [PubMed] [Google Scholar]
- 20.Poisnel G, Arenaza-Urquijo E, Collette F, et al. The age-well randomized controlled trial of the Medit-Ageing European project: Effect of meditation or foreign language training on brain and mental health in older adults. Alzheimers Dement Transl Res Clin Interv. 2018;4:714–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Canela A, Vera E, Klatt P, et al. High-throughput telomere length quantification by FISH and its application to human population studies. Proc Natl Acad Sci U S A. 2007;104:5300–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Elvsåshagen T, Vera E, Bøen E, et al. The load of short telomeres is increased and associated with lifetime number of depressive episodes in bipolar II disorder. J Affect Disord. 2011;135:43–50. [DOI] [PubMed] [Google Scholar]
- 23.La Joie R, Perrotin A, de La Sayette V, et al. Hippocampal subfield volumetry in mild cognitive impairment, Alzheimer’s disease and semantic dementia. NeuroImage Clin. 2013;3:155–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Palix C, Felisatti F, Gonneaud J, et al. Relationships between diabetes-related vascular risk factors and neurodegeneration biomarkers in healthy aging and Alzheimer’s disease. Neurobiol Aging. 2022;118:25–33. [DOI] [PubMed] [Google Scholar]
- 25.Besson FL, La Joie R, Doeuvre L, et al. Cognitive and brain profiles associated with current neuroimaging biomarkers of preclinical Alzheimer’s disease. J Neurosci. 2015;35:10402–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yushkevich PA, Pluta JB, Wang H, et al. Automated volumetry and regional thickness analysis of hippocampal subfields and medial temporal cortical structures in mild cognitive impairment. Hum Brain Mapp. 2015;36:258–87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.André C, Kuhn E, Rehel S, et al. Association of sleep-disordered breathing and medial temporal lobe atrophy in cognitively unimpaired amyloid-positive older adults. Neurology. 2023;101:e370–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.de Flores R, Demeilliez-Servouin S, Kuhn E, et al. Respective influence of beta-amyloid and APOE ε4 genotype on medial temporal lobe subregions in cognitively unimpaired older adults. Neurobiol Dis. 2023;181:106127. [DOI] [PubMed] [Google Scholar]
- 29.Xie L, Wisse LEM, Pluta J, et al. Automated segmentation of medial temporal lobe subregions on in vivo T1-weighted MRI in early stages of Alzheimer’s disease. Hum Brain Mapp. 2019;40:3431–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hepatic and renal function impact concentrations of plasma biomarkers of neuropathology - Berry - 2022 - Alzheimer’s & Dementia: Diagnosis, Assessment & Disease Monitoring - Wiley Online Library. [DOI] [PMC free article] [PubMed]
- 31.Schöll M, Verberk IMW, Campo M del et al. Challenges in the practical implementation of blood biomarkers for Alzheimer’s disease. Lancet Healthy Longev 2024;5, 10.1016/j.lanhl.2024.07.013. [DOI] [PubMed]
- 32.Gong N-J, Wong C-S, Chan C-C, et al. Aging in deep gray matter and white matter revealed by diffusional kurtosis imaging. Neurobiol Aging. 2014;35:2203–16. [DOI] [PubMed] [Google Scholar]
- 33.Salat DH, Buckner RL, Snyder AZ, et al. Thinning of the cerebral cortex in aging. Cereb Cortex N Y N. 1991;2004(14):721–30. [DOI] [PubMed] [Google Scholar]
- 34.Staffaroni AM, Tosun D, Lin J, et al. Telomere attrition is associated with declines in medial temporal lobe volume and white matter microstructure in functionally independent older adults. Neurobiol Aging. 2018;69:68–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Topiwala A, Nichols TE, Williams LZJ, et al. Telomere length and brain imaging phenotypes in UK Biobank. PLoS ONE. 2023;18:e0282363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Khalil M, Pirpamer L, Hofer E, et al. Serum neurofilament light levels in normal aging and their association with morphologic brain changes. Nat Commun. 2020;11:812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hampel H, Hu Y, Cummings J, et al. Blood-based biomarkers for Alzheimer’s disease: Current state and future use in a transformed global healthcare landscape. Neuron. 2023;111:2781–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Rodríguez-Fernández B, Vilor-Tejedor N, Arenaza-Urquijo EM, et al. Genetically predicted telomere length and Alzheimer’s disease endophenotypes: a Mendelian randomization study. Alzheimers Res Ther. 2022;14:167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Moore EE, Hohman TJ, Badami FS, et al. Neurofilament relates to white matter microstructure in older adults. Neurobiol Aging. 2018;70:233–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Demissie S, Levy D, Benjamin EJ, et al. Insulin resistance, oxidative stress, hypertension, and leukocyte telomere length in men from the Framingham Heart Study. Aging Cell. 2006;5:325–30. [DOI] [PubMed] [Google Scholar]
- 41.Bettcher BM, Yaffe K, Boudreau RM, et al. Declines in inflammation predict greater white matter microstructure in older adults. Neurobiol Aging. 2015;36:948–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bettcher BM, Watson CL, Walsh CM, et al. Interleukin-6, Age, and Corpus Callosum Integrity. PLoS ONE. 2014;9:e106521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Malaquin N, Martinez A, Rodier F. Keeping the senescence secretome under control: Molecular reins on the senescence-associated secretory phenotype. Exp Gerontol. 2016;82:39–49. [DOI] [PubMed] [Google Scholar]
- 44.Huang L, Zhang M, Bai D, et al. Deciphering the impact of TERT/telomerase on immunosenescence and T cell revitalization. Front Immunol. 2024;15:1465006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Rodriguez IJ, Lalinde Ruiz N, Llano León M et al. Immunosenescence study of T cells: a systematic review. Front Immunol 2021;11. 10.3389/fimmu.2020.604591. [DOI] [PMC free article] [PubMed]
- 46.Deleidi M, Jäggle M, Rubino G. Immune aging, dysmetabolism, and inflammation in neurological diseases. Front Neurosci 2015;9. 10.3389/fnins.2015.00172. [DOI] [PMC free article] [PubMed]
- 47.Di Benedetto S, Müller L, Wenger E, et al. Contribution of neuroinflammation and immunity to brain aging and the mitigating effects of physical and cognitive interventions. Neurosci Biobehav Rev. 2017;75:114–28. [DOI] [PubMed] [Google Scholar]
- 48.von Bernhardi R, Tichauer JE, Eugenín J. Aging-dependent changes of microglial cells and their relevance for neurodegenerative disorders. J Neurochem. 2010;112:1099–114. [DOI] [PubMed] [Google Scholar]
- 49.Bettcher BM, Tansey MG, Dorothée G, et al. Peripheral and central immune system crosstalk in Alzheimer disease — a research prospectus. Nat Rev Neurol. 2021;17:689–701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zang X, Chen S, Zhu J, et al. The emerging role of central and peripheral immune systems in neurodegenerative diseases. Front Aging Neurosci. 2022;14:872134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Demanelis K, Jasmine F, Chen LS, et al. Determinants of telomere length across human tissues. Science. 2020;369:eaaz6876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Nakamura K-I, Takubo K, Izumiyama-Shimomura N, et al. Telomeric DNA length in cerebral gray and white matter is associated with longevity in individuals aged 70 years or older. Exp Gerontol. 2007;42:944–50. [DOI] [PubMed] [Google Scholar]
- 53.Tomita K, Aida J, Izumiyama-Shimomura N, et al. Changes in telomere length with aging in human neurons and glial cells revealed by quantitative fluorescence in situ hybridization analysis. Geriatr Gerontol Int. 2018;18:1507–12. [DOI] [PubMed] [Google Scholar]
- 54.Cao Z, Hou Y, Xu C. Leucocyte telomere length, brain volume and risk of dementia: a prospective cohort study. Gen Psychiatry. 2023;36:e101120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Salih A, Galazzo IB, Petersen SE, et al. Telomere length is causally connected to brain MRI image derived phenotypes: a mendelian randomization study. PLoS ONE. 2022;17:e0277344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.King KS, Kozlitina J, Rosenberg RN, et al. Effect of leukocyte telomere length on total and regional brain volumes in a large population-based cohort. JAMA Neurol. 2014;71:1247–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.de Flores R, La Joie R, Chételat G. Structural imaging of hippocampal subfields in healthy aging and Alzheimer’s disease. Neuroscience. 2015;309:29–50. [DOI] [PubMed] [Google Scholar]
- 58.Wikgren M, Karlsson T, Lind J, et al. Longer leukocyte telomere length is associated with smaller hippocampal volume among non-demented APOE ε3/ε3 subjects. PLoS ONE. 2012;7:e34292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Régy M, Dugravot A, Sabia S, et al. Association of APOE ε4 with cerebral gray matter volumes in non-demented older adults: The MEMENTO cohort study. Neuroimage. 2022;250:118966. [DOI] [PubMed] [Google Scholar]
- 60.Liu C-C, Zhao J, Fu Y, et al. Peripheral apoE4 enhances Alzheimer’s pathology and impairs cognition by compromising cerebrovascular function. Nat Neurosci. 2022;25:1020–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Moon W-J, Lim C, Ha IH, et al. Hippocampal blood–brain barrier permeability is related to the APOE4 mutation status of elderly individuals without dementia. J Cereb Blood Flow Metab. 2021;41:1351–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Montagne A, Nation DA, Sagare AP, et al. APOE4 leads to blood–brain barrier dysfunction predicting cognitive decline. Nature. 2020;581:71–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Benedet AL, Milà-Alomà M, Vrillon A, et al. Differences between plasma and cerebrospinal fluid glial fibrillary acidic protein levels across the Alzheimer disease continuum. JAMA Neurol. 2021;78:1471–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Stocker H, Trares K, Beyer L, et al. Alzheimer’s polygenic risk scores, APOE, Alzheimer’s disease risk, and dementia-related blood biomarker levels in a population-based cohort study followed over 17 years. Alzheimers Res Ther. 2023;15:129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Snellman A, Ekblad LL, Ashton NJ, et al. Head-to-head comparison of plasma p-tau181, p-tau231 and glial fibrillary acidic protein in clinically unimpaired elderly with three levels of APOE4-related risk for Alzheimer’s disease. Neurobiol Dis. 2023;183:106175. [DOI] [PubMed] [Google Scholar]
- 66.Erdő F, Denes L, de Lange E. Age-associated physiological and pathological changes at the blood-brain barrier: A review. J Cereb Blood Flow Metab Off J Int Soc Cereb Blood Flow Metab 2017;37. 10.1177/0271678X16679420. [DOI] [PMC free article] [PubMed]
- 67.Sun Y, Koyama Y, Shimada S. Inflammation From Peripheral Organs to the Brain: How Does Systemic Inflammation Cause Neuroinflammation? Front Aging Neurosci 2022;14. 10.3389/fnagi.2022.903455. [DOI] [PMC free article] [PubMed]
- 68.Boccardi V, Pelini L, Ercolani S, et al. From cellular senescence to Alzheimer’s disease: The role of telomere shortening. Ageing Res Rev. 2015;22:1–8. [DOI] [PubMed] [Google Scholar]
- 69.Rolyan H, Scheffold A, Heinrich A, et al. Telomere shortening reduces Alzheimer’s disease amyloid pathology in mice. Brain J Neurol. 2011;134:2044–56. [DOI] [PubMed] [Google Scholar]
- 70.Lai T-P, Wright WE, Shay JW. Comparison of telomere length measurement methods. Philos Trans R Soc B Biol Sci. 2018;373:20160451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Chételat G, Lutz A, Klimecki O, et al. Effect of an 18-month meditation training on regional brain volume and perfusion in older adults: the age-well randomized clinical trial. JAMA Neurol. 2022;79:1165–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data underlying this report are made available on request following a formal data sharing agreement and approval by the consortium and executive committee https://silversantestudy.eu/2020/09/25/data-sharing). The Material can be mobilized, under the conditions and modalities defined in the Medit-Ageing Charter, by any research team belonging to an academic for carrying out a scientific research project relating to the scientific theme of mental health and well-being in older people. The Material may also be mobilized by non-academic third parties, under conditions, in particular financial, which will be established by separate agreement between Inserm and by the said third party. Data sharing policies described in the Medit-Ageing Charter comply our ethics approval and guidelines by our funding body.