

ABSTRACT
Theory suggests that animals make hierarchical, multiscale resource selection decisions to address the hierarchy of factors limiting their fitness. Ecologists have developed tools to link population‐level resource selection across scales; yet, theoretical expectations about the relationship between coarse‐ and fine‐scale selection decisions at the individual level remain elusive despite their importance to fitness. With GPS‐telemetry data collected across California, USA, we evaluated resource selection of mountain lions ( Puma concolor ; n = 244) relative to spatial variation in human‐caused mortality risk. With hierarchical resource selection, coarse‐scale selection determines availability at finer scales. This simple relationship allowed us to demonstrate that functional responses in resource selection explicitly link individual‐level resource selection decisions across scales. We show that individuals proactively avoiding risk when selecting home ranges are freed to relax this avoidance when making decisions within home ranges. However, individuals also exhibit reactive avoidance of risk at the finest scales along movement paths.
Keywords: functional response, hierarchical resource selection, livestock depredation, Puma concolor
A quantitative framework that links individual‐level responses to factors limiting fitness across scales has been elusive for animal ecology. With hierarchical resource selection, coarse‐scale selection determines availability at finer scales. This simple relationship allowed us to demonstrate that functional responses in resource selection explicitly link individual‐level resource selection decisions across scales, using a large telemetry dataset of mountain lions (n = 244) tracked across the state of California, USA.
1. Introduction
Resource selection is a hierarchical process where the decisions an individual makes at coarse scales constrain what is available when making finer scale decisions (Johnson 1980). Importantly, theory predicts that these decisions correspond to the hierarchy of factors limiting an individual's fitness, such that factors with the greatest potential to reduce fitness should be addressed at the coarsest scale at which they are encountered (limiting factors hypothesis; Rettie and Messier 2000). Pressures that limit fitness should dominate selection decisions at successively finer scales until their effects are overcome (Rettie and Messier 2000). Most often, ecologists evaluate selection of home ranges from the larger landscape (second‐order; Johnson 1980) or selection of locations within home ranges (third‐order; Johnson 1980; Manly et al. 2002). While studies evaluating multiscale resource selection are common (e.g., Boyce 2006; Mayor et al. 2009), explicitly linking and evaluating selection decisions by individuals across scales remain a challenge that has limited understanding of the strategies that animals use to address the hierarchy of factors influencing their fitness.
Traditionally, multiscale resource selection studies have evaluated relative probability of use separately at each order of selection (e.g., Dussault et al. 2005; Johnson, Parker, and Heard 2001; McGarigal et al. 2016; Van Beest et al. 2010). This approach allows ecologists to identify differences in resource selection across scales (e.g., Anderson et al. 2005; Boyce et al. 2003; Ciarniello et al. 2007), but such comparisons are largely descriptive and often fail to explicitly consider that coarse‐scale selection constrains fine‐scale selection (Mayor et al. 2009). More recently, DeCesare et al. (2012) developed the scale‐integrated resource selection function (SRSF), which incorporates a hierarchically nested design wherein locations used at coarse scales serve as available locations at fine scales. SRSFs are an effective tool for modelling habitat suitability that links resource selection across scales at the population level (DeCesare et al. 2012, 2014; McLoughlin, Dunford, and Boutin 2005). However, incorporating a hierarchically nested design that explicitly links responses of animals at the individual level to factors limiting their fitness across multiple spatial scales is an important next‐step in the continued development of methods for evaluating multiscale resource selection.
For prey species, predation and forage are among the most important factors limiting fitness (McNamara 1987). When food availability and predation risk are positively correlated in space and time, individuals must balance the benefit of increased food availability with the cost of increased risk of mortality (Lima and Dill 1990; Sih 1980). Demonstrating that individuals can navigate forage‐predation risk trade‐offs across scales, Hebblewhite and Merrill (2009) found that elk ( Cervus canadensis ) that migrated to avoid predation risk were able to forage more freely at the cost of reduced access to high quality forage. Animals navigating forage‐predation risk trade‐offs also frequently exhibit functional responses in resource selection, where selection of a resource varies as a function of its availability (Godvik et al. 2009; Mysterud and Ims 1998). Despite the frequency with which functional responses are reported as strategies for navigating forage‐predation risk trade‐offs and the strong theoretical basis for the hierarchical nature of resource selection, functional responses in resource selection at a given scale have yet to be explicitly linked to coarser resource selection decisions that determine fine‐scale availability.
Foraging efficiency of predators is influenced not only by prey availability, but also susceptibility of their prey to predation (Sih 2005; Smith et al. 2019). For instance, ambush predators (e.g., large felids) often prioritise selection of stalking cover required to kill prey, rather than areas with the greatest prey availability (Coon et al. 2020; Hopcraft, Sinclair, and Packer 2005). Theoretical and empirical understanding of forage‐predation risk trade‐offs was developed primarily for prey species (Lima and Dill 1990); however, the risk‐disturbance hypothesis suggests human disturbance acts similarly to predation risk and stimulates anti‐predator behaviours that elicit indirect fitness costs in many species (Frid and Dill 2002; Ordiz et al. 2021; Suraci et al. 2019). Indeed, there is increasing empirical evidence that large carnivores face similar trade‐offs in the presence of substantial anthropogenic mortality risk (Basille et al. 2013; Ripari et al. 2022). Human‐caused mortality negatively impacts populations of large carnivores worldwide and often appears to be a greater limiting factor than prey availability (Bleyhl et al. 2021; Chapron et al. 2008; Darimont et al. 2015; Lamb et al. 2020). As individual‐level behavioural responses to forage‐risk trade‐offs may scale up to population‐level effects (Gaynor et al. 2021; Lima 1998; Preisser and Bolnick 2008), understanding how large carnivores balance foraging efficiency with anthropogenic risk as they make resource selection decisions across scales is of both theoretical and practical importance.
Mountain lions ( Puma concolor ) are fully protected from hunting in California, but human‐caused mortality exceeds natural mortality and contributes to lower survival rates for populations with higher rates of human‐caused mortality (Benson et al. 2023). Intentional mortality associated with livestock depredation management is the leading known cause of death for mountain lions in California, although vehicle collisions and other forms of unintentional mortality also contribute to reduced survival (Benson et al. 2023). Mountain lions can only be legally killed while actively attacking livestock or if the California Department of Fish and Wildlife (CDFW) issues a permit following livestock depredation. When a permit is issued, mountain lions are generally killed via gunshot if they return to the site where the depredation incident occurred (see Appendix S1 for additional details regarding mortality rates, depredation management policies and methods of lethal removal). Importantly, cues associated with this method of take may be difficult for mountain lions to reliably perceive, and their behavioural responses to depredation mortality risk are not well understood (Montgomery et al. 2022; Nisi, Benson, and Wilmers 2022). Mountain lions generally avoid humans, but exhibit behaviours to balance mortality risk with the benefits of increased availability of their primary prey, mule deer ( Odocoileus hemionus ), in urban and peri‐urban environments (Furnas et al. 2020; Knopff et al. 2014). In greater Los Angeles, mountain lions appeared to trade off increased risk of human‐caused mortality and prey availability by killing prey closer to development, although they rarely actually entered development (Benson, Sikich, and Riley 2016). However, similar trade‐offs may be more difficult to navigate in sparsely developed areas (Lowry, Lill, and Wong 2013; Nisi, Benson, and Wilmers 2022). Indeed, survival rates were lowest and human‐caused mortality rates were highest in regions of California characterised by intermediate levels of human presence (Benson et al. 2023). Evaluating responses of mountain lions to landscape characteristics influencing both mortality risk and foraging efficiency, and linking these responses across scales at the individual level, provides a valuable opportunity to advance our understanding of how animals respond to multiple fitness‐limiting factors when making hierarchical resource selection decisions.
We investigated mountain lion movement and resource selection relative to spatially varying risk of intentional human‐caused mortality associated with livestock depredation (hereafter depredation mortality risk) and landscape features associated with foraging success across their distribution in California. We hypothesised that stronger avoidance of depredation mortality risk at coarse scales frees large carnivores to relax their avoidance of risk and direct behaviour towards accessing limiting resources when making fine‐scale selection decisions (consistent with the limiting factors hypothesis; Rettie and Messier 2000; Figure 1). If supported, we predicted that mountain lions would exhibit a functional response to depredation mortality risk such that there would be a negative relationship between fine‐scale responses to depredation mortality risk by individuals and the mean level of depredation mortality risk across their home ranges (P1; Figure 1). This prediction highlights that population‐level functional responses in resource selection should provide an explicit link between resource selection decisions across spatial scales for individual animals because decisions at coarse scales (e.g., landscape level) determine availability at finer scales (e.g., within home range) within the framework of hierarchical resource selection (Johnson 1980). We also predicted that strong avoidance of depredation mortality risk at coarse scales should free individual mountain lions to focus fine‐scale selection on features associated with foraging success (i.e., selection of stalking cover used to kill prey, P2; Murphy and Ruth 2009; Smith et al. 2019). Because it is unlikely that they can escape depredation mortality risk entirely, we also hypothesised that large carnivores adopt context‐dependent selection behaviour to mitigate risk they encounter at the finest (local) scales along movement paths. Here, we predicted mountain lions would modify avoidance of depredation mortality risk and selection of cover as a response to variation in risk along movement paths (P3). Our work enhances understanding of how individual animals make hierarchical resource selection decisions to balance the demands of multiple factors assumed to limit fitness by providing a novel framework for linking coarse‐ and fine‐scale resource selection.
FIGURE 1.

A conceptual figure of the hypothesised relationship between coarse‐ and fine‐scale response to depredation mortality risk where the strength of selection or avoidance of risk at each scale influences the level of risk that an individual is exposed to (purple = low risk, yellow = high risk). Our first prediction is best reflected by the top left panel, where strong avoidance of depredation mortality risk at a coarse scale frees an individual to relax avoidance of risk when making fine‐scale selection decisions.
2. Materials and Methods
2.1. Study System and Field Methods
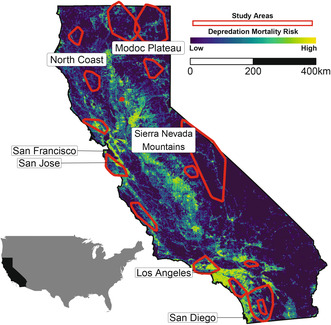
Between 2001 and 2020, we captured mountain lions and deployed GPS collars that obtained locations at fix rates ranging from 1 to 4 h in 16 study areas throughout California (Figure 2; Field Methods described in Appendix S2). The Sierra Nevada Mountains, Modoc plateau and north coast retain large expanses of habitat that support large, genetically diverse populations (Figure 2; Dellinger, Cristescu, et al. 2020). Southern California and the Central Coast contain several major metropolitan areas (e.g., greater Los Angeles, San Francisco and San Diego; Figure 2) and an expansive network of freeways that have contributed to substantial habitat loss and fragmentation. As a result, mountain lion populations in these regions persist within relatively small habitat patches that are below thresholds required to maintain suitable effective population sizes (Dellinger, Cristescu, et al. 2020), which increases risk of extinction from genetic and demographic processes (Benson et al. 2016, 2019).
FIGURE 2.
Relative depredation mortality risk for mountain lions across California. Derived from predictions of relative probability of use from conditional logistic regression models comparing landscape features at locations where mountain lions were killed following livestock depredation (n = 466) with those available across the home ranges of the mountain lions that were killed in California, USA, 2016–2020.
2.2. Overview of Analytical Framework
We used locations where mountain lions were legally killed on depredation permits to predict spatially varying depredation mortality risk throughout California (Figure 2). We then used location data from GPS collared mountain lions to evaluate multiscale resource selection relative to predicted depredation mortality risk. Specifically, we used second‐order resource selection functions (RSFs) to evaluate selection of home ranges from the larger landscape (hereafter coarse‐scale resource selection) and step‐selection functions (SSFs) to evaluate movement‐based resource selection within home ranges (hereafter fine‐scale resource selection). Coarse‐scale resource selection decisions should determine fine‐scale resource availability. Thus, we evaluated whether individuals exhibited functional responses in resource selection to demonstrate the relationship between coarse‐ and fine‐scale resource selection. Where relevant, we considered support for differences in resource selection by sex, time of day and movement state using interactions (details in Appendix S2). We also conducted an exploratory analysis to evaluate whether multiscale resource selection decisions of individuals that were killed on depredation permits differed from those of individuals that were not killed on depredation permits. Our intention was to understand whether behaviour of individual mountain lions influenced this important component of fitness. However, the results of this analysis were inconclusive and uncertain given that the relatively low sample size of GPS collared animals killed on depredation permits may not have been representative (see Appendix S3 for details on exploratory analyses).
2.3. Depredation Mortality Risk
We investigated landscape features influencing spatial variation in risk of mountain lions being killed on depredation permits using a use‐availability resource selection framework similar to Johnson's (1980) third‐order (within home range) selection. Specifically, we used conditional logistic regression to compare landscape attributes at used locations where 466 mountain lions were killed on depredation permits to those at available locations distributed across estimated or approximated home ranges. We estimated home ranges using 100% adaptive local convex hulls (a‐LoCoH; Getz et al. 2007) for individuals killed that were tracked with GPS telemetry (n = 23). For individuals not tracked with GPS telemetry (n = 443), we approximated home ranges based on age‐ and sex‐specific distributions of empirical home range size from our larger mountain lion telemetry dataset collected across the state (n = 244, details in Appendix S2). We used results from the most strongly supported models to generate a probability surface representing spatially varying depredation mortality risk throughout California and included predictions from this surface in subsequent second‐order RSFs and SSFs (see below; details of depredation mortality risk estimation and validation described in Appendix S2).
2.4. Coarse‐Scale Resource Selection
We evaluated coarse‐scale resource selection for 244 mountain lions across California as they selected home ranges from the larger landscape (second‐order selection; Johnson 1980). We estimated 100% a‐LoCoH home ranges and defined the larger landscape as the 100% minimum convex polygon derived from telemetry data of all study animals in each study area (Figure 2; Appendix S2, Table S1). We estimated resource conditions at the centre of every fifth 30‐m2 pixel within home ranges (used) and the larger landscape (availability; Benson 2013; DeCesare et al. 2012; Spitz, Hebblewhite, and Stephenson 2020). At each used and available location, we extracted values for predicted depredation mortality risk, elevation and slope (Farr et al. 2007), and distance to forest, shrub and herbaceous cover (Dewitz and U.S. Geological Survey 2021).
After rescaling all continuous variables by subtracting their mean and dividing by two standard deviations (Gelman 2008), we fit generalised linear mixed models using the ‘glmmTMB’ R package (Brooks et al. 2017) with a binary response variable (1 = used, 0 = available) to estimate the probability of each resource being used relative to its availability. We included a random intercept for each individual nested within study area, which accounted for unbalanced sample sizes of locations among individuals and potential lack of independence of data from the same individuals and areas (Gillies et al. 2006). Lastly, we fit a model including a random slope that allowed the response to depredation mortality risk to vary among individuals to derive individual‐level coefficients for their response to risk (Gillies et al. 2006).
2.5. Fine‐Scale Resource Selection
We used SSFs to evaluate movement decisions within the home ranges of the same 244 mountain lions with respect to anthropogenic features that increased their depredation mortality risk (Fortin et al. 2005). Selection and avoidance inferred from SSFs are strongly influenced by GPS collar fix‐rates (Nisi et al. 2021; Thurfjell, Ciuti, and Boyce 2014). Thus, we based our main inference on models with 2‐h fix‐rates to maximise the number of individuals we could include in the model (n = 144) while maintaining relatively short temporal intervals between locations. However, we present results from models with other fix rates for comparison.
We used hidden Markov models (HMMs) to identify stationary, meandering and directed movement states (HMM methods described in Appendix S2). To address variation in movement characteristics between different states, we generated available steps from individual‐specific distributions of empirically observed step lengths and turning angles that were specific to the movement state of the individual at a given step. We generated five available steps for each used step, which was sufficient to achieve stabilisation of coefficients in our SSFs (Appendix S2, Figure S1). At the endpoint of each used and available step (Thurfjell, Ciuti, and Boyce 2014), we extracted values for predicted depredation mortality risk, elevation and slope (Farr et al. 2007) and distance to forest, shrub and herbaceous cover (Dewitz and U.S. Geological Survey 2021). After rescaling continuous covariates, we fit conditional logistic regression models with a binary response variable (1 = used, 0 = available) using the ‘survival’ package (Therneau and Lumley 2015) in R version 4.3.2 (R Core Team 2023). We used a stratum that linked time‐dependent used and available locations for each individual (Fortin et al. 2005), and also included a cluster term to address possible biases in variance associated with temporally autocorrelated telemetry data and unbalanced sample sizes among individuals (Prima, Duchesne, and Fortin 2017).
2.6. Functional Responses in Resource Selection
We explored whether mountain lions exhibited functional responses in resource selection such that their response to depredation mortality risk or other resources varied as a function of depredation mortality risk within its home range (Mysterud and Ims 1998). First, we fit separate SSFs for each individual to obtain coefficients representing individual responses to depredation mortality risk, forest, shrub and herbaceous cover. We then fit linear models with coefficients for each resource from the individual SSFs as the response variable and mean depredation mortality risk within the individual's home range as a predictor variable. We also fit linear models with individual‐level coefficients derived from a Poisson generalised linear mixed model (GLMM) with stratum‐specific fixed intercepts and random slopes for individual (Muff, Signer, and Fieberg 2020) to verify that our results were robust to our choice of method for deriving individual‐level responses (see Appendix S3 for details).
Including individual‐level coefficients from the SSFs as predictor variables in functional response models without accounting for uncertainty surrounding coefficient estimates could introduce bias (Hadfield et al. 2010). Thus, we evaluated whether observed functional responses were robust to this uncertainty (details in Appendix S2). We used functional response models to test our prediction that differences in the level of depredation mortality risk within an individual's home range influenced variation in their fine‐scale responses to risk and stalking cover. We assumed that selection of home ranges defined risk within home ranges. If true, the functional response provides an explicit link between coarse‐scale (second‐order) and fine‐scale (third‐order) resource selection of individuals. To verify, we estimated correlation coefficients between mean values of depredation mortality risk within home ranges and individual‐level coefficients derived from second‐order RSFs.
The functional response described above is a change in selection across individuals within a population as a function of variation in their home range‐level availability (i.e., a population‐level functional response). Next, we evaluated whether individual mountain lions altered selection as a function of spatial variation in risk along movement paths (Mason and Fortin 2017). Specifically, we fit SSFs with interactions between mean depredation mortality risk at available steps and distance to forest cover, shrub cover, herbaceous cover and depredation mortality risk. Here, we concluded mountain lions exhibited a functional response if interactions were statistically supported and coefficients indicated mountain lions selected resources as a function of local availability of risk. This would represent a functional response exhibited by individual animals that change their selection as a function of variation in local availability (i.e., an individual‐level functional response). We refer to the two types of functional responses throughout as population‐level (change in selection across individuals within a population) and individual‐level (change in selection within individual animals) functional responses in resource selection to avoid confusion.
3. Results
3.1. Depredation Mortality Risk
Risk of mountain lions being killed following livestock depredation increased on gentler slopes (mean β = −0.58, SD = 0.04) and closer to low‐intensity development (mean β = −1.87, SD = 0.21), local roads (mean β = −4.39, SD = 0.43), buildings (mean β = −7.11, SD = 0.42) and vegetative cover (mean β = −0.39, SD = 0.13; n = 466, Figure 2; Figure S2). Our models had good predictive ability (mean ρ = 0.87) and results were consistent when we restricted the model to individuals with empirically derived home ranges (n = 23, Table S3).
3.2. Coarse‐Scale Resource Selection
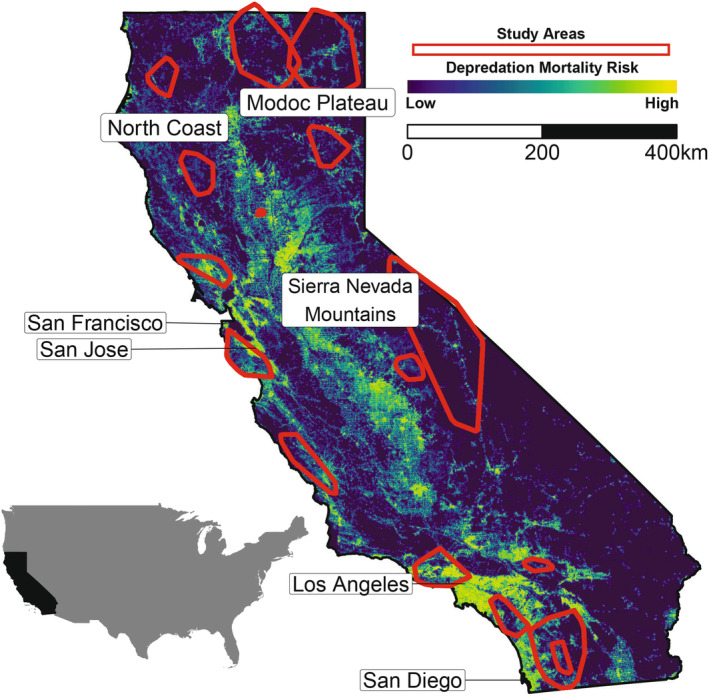
When selecting home ranges from the larger landscape, mountain lions selected forest cover (β = −0.459, 95% CI: −0.464, −0.455), shrub cover (β = −0.315, 95% CI: −0.319, −0.311) and herbaceous areas (β = −0.106, 95% CI: −0.109, −0.103), whereas they avoided higher elevation (β = −0.300, 95% CI: −0.306, −0.294), steeper slopes (β = −0.089, 95% CI: −0.091, −0.086) and depredation mortality risk (β = −0.542, 95% CI: −0.546, −0.539, n = 244, Figure 3A). Interactions between sex and resource variables were statistically supported but did not change inference on resource selection (Tables S4 and S5).
FIGURE 3.
(A) Coefficient plot from the second‐order resource selection function investigating California mountain lions' (n = 244) population‐level responses to landscape features when selecting home ranges from the larger landscape. (B) Coefficient plot from the 2‐h step‐selection function investigating California mountain lions' (n = 144) population‐level responses to landscape features when making selection decisions along movement paths. For classification‐based variables (depredation mortality risk, elevation, slope), values less than 0 indicate avoidance, whereas values greater than 0 indicate selection. For distance‐based variables, values less than 0 indicate selection, whereas values greater than 0 indicate avoidance.
3.3. Fine‐Scale Resource Selection
When making fine‐scale resource selection decisions, mountain lions selected forest cover (β = −0.55, 95% CI: −0.69, −0.40) and shrub cover (β = −0.15, 95% CI: −0.22, −0.07), while they avoided higher elevations (β = −0.91, 95% CI: −1.34, −0.49), steeper slopes (β = −0.47, 95% CI: −0.61, −0.34), herbaceous areas (β = 0.08, 95% CI: 0.01, 0.15) and depredation mortality risk (β = −0.30, 95% CI:‐0.45, −0.15, n = 144, Figure 3B). Despite statistical support, models containing interactions between resource variables and time of day, movement state, or sex did not change inference on fine‐scale resource selection (Tables S6–S9). Thus, we made inference on the simpler model without interactions. Results from SSFs with 1‐, 3‐ and 4‐h intervals were generally consistent with results from models with 2‐h intervals reported here (Figure S3).
3.4. Functional Response Models
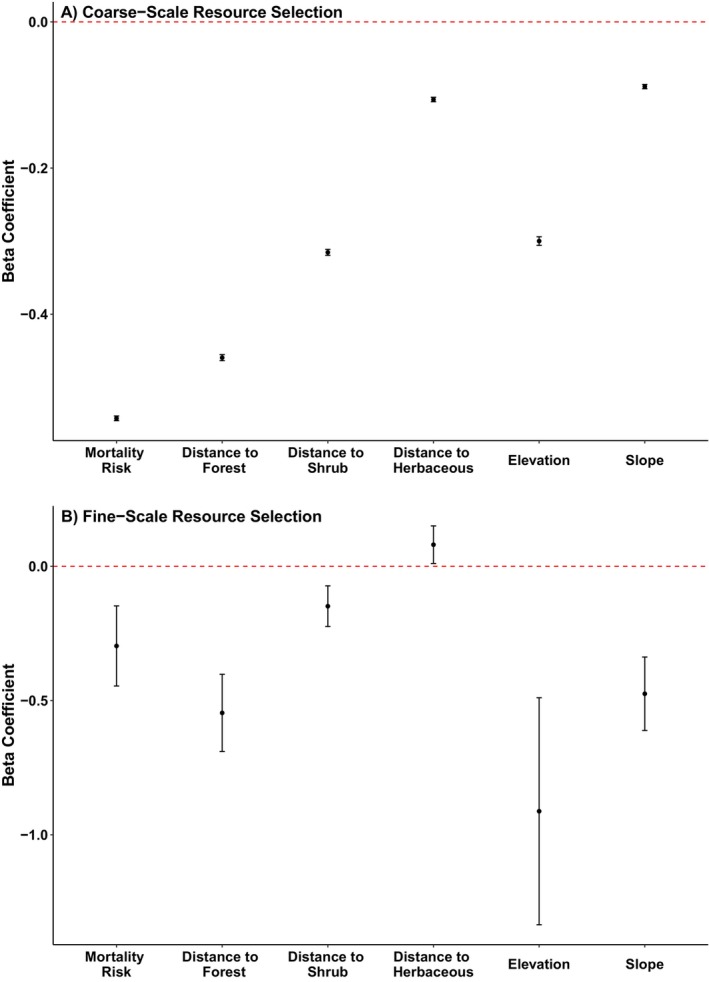
Mountain lions exhibited a population‐level functional response in resource selection where individuals with higher levels of mean depredation mortality risk within their home ranges avoided depredation mortality risk more strongly when making fine‐scale selection decisions (β = −1.50, 95% CI: −2.02, −0.97). The observed functional response was robust to uncertainty around individual‐level coefficients from the SSFs (mean β = −1.50, 95% Highest Posterior Density Interval (HPD): −1.74–1.24, n = 144, Figure 4). We did not find support for sex‐specific functional responses to depredation mortality risk, as model fit was best for the model without interactions (ΔAICc = 0.26). In each study area, mean depredation mortality risk within home ranges of individual mountain lions was highly correlated with individual‐level coefficients from the second‐order RSF, which verified the assumption that coarse‐scale resource selection decisions determined availability at finer scales (mean correlation = 0.88, SD = 0.14, all p < 0.05, n = 14 study areas; Table S10). Mountain lions did not exhibit population‐level functional responses to forest cover (β = 0.16, 95% CI: −0.46, 0.78), shrub cover (β = −0.27, 95% CI: −0.65, 0.11) or herbaceous areas (β = 0.30, 95% CI: −0.16, 0.75) as there were no significant relationships between response to these resources and mean depredation mortality risk within home ranges. However, mountain lions also exhibited individual‐level functional responses where they avoided depredation mortality risk (β = −1.84, 95% CI: −2.07, −1.60) and herbaceous areas (β = 0.17, 95% CI: 0.01, 0.32) more strongly and selected shrub (β = −0.16, 95% CI: −0.29, −0.03) more strongly along their movement paths in areas of locally higher levels of risk (∆QIC = 6934.61, n = 144, Figure 5; Table S11).
FIGURE 4.
Radio collared mountain lions tracked across their distribution in California, USA from 2004 to 2020 (n = 144) exhibited a population‐level functional response to depredation mortality risk as there was a linear relationship between coefficients representing individual mountain lions' fine‐scale responses to depredation mortality risk and mean values of depredation mortality risk within their home ranges. Coefficient values less than 0 indicate avoidance, whereas values greater than 0 indicate selection.
FIGURE 5.
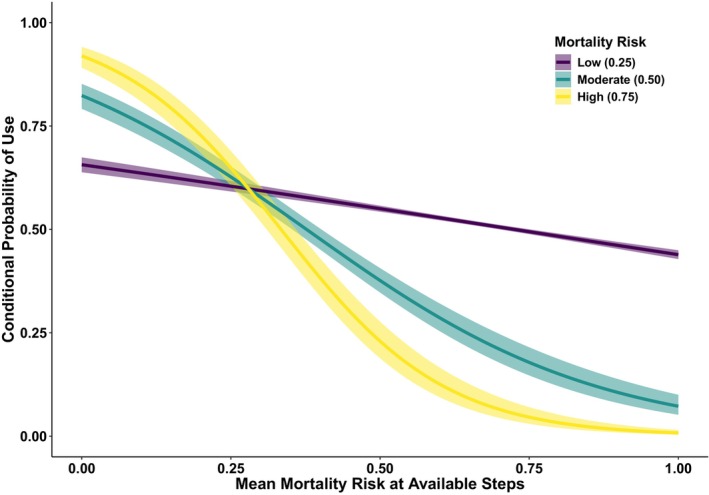
Radio collared mountain lions tracked across their distribution in California, USA from 2004 to 2020 (n = 144) exhibited individual‐level functional responses to depredation mortality risk wherein they avoided risk more strongly when there was higher risk at locations available to them along movement paths. Shown is conditional probability of use for low‐ (0.25), moderate‐ (0.50) and high‐risk (0.75) locations at varying levels of local depredation mortality risk along the movement paths of mountain lions.
4. Discussion
According to the limiting factors avoidance hypothesis (Rettie and Messier 2000), animals making hierarchical resource selection decisions are expected to exhibit patterns of selection and avoidance that reflect the hierarchy of factors limiting their fitness. Unfortunately, elucidating these behavioural responses across multiple scales has been limited by the absence of a quantitative framework that explicitly links selection decisions by individual animals at both coarse‐ and fine scales. We quantified the response of mountain lions to depredation mortality risk and stalking cover that influences their foraging efficiency to test theoretical predictions about resource selection relative to fitness limiting factors associated with risk‐forage trade‐offs across multiple scales (Hebblewhite and Merrill 2009; Rettie and Messier 2000). Mountain lions across the state of California avoided depredation mortality risk more strongly than they selected resources that influence how effectively they can stalk and kill their primary prey (forest and shrub cover; Elbroch and Wittmer 2012; Murphy and Ruth 2009) when selecting home ranges (Figure 3A). Within home ranges, mountain lions avoided depredation mortality risk and selected vegetation that provided stalking cover with similar intensity (Figure 3B). Previous work has suggested that human‐caused mortality is the most important factor limiting fitness of large carnivores (Bleyhl et al. 2021; Chapron et al. 2008). Strong population‐level avoidance of depredation mortality risk when selecting home ranges is consistent with theoretical and empirical work suggesting that animals focus coarse‐scale resource selection decisions on the most important factors limiting their fitness (Gaillard et al. 2010; Owen‐Smith, Fryxell, and Merrill 2010; Rettie and Messier 2000). Still, theoretical expectations about hierarchical resource selection relative to fitness‐limiting factors cannot be explicitly tested without first establishing a link between coarse‐ and fine‐scale selection at the individual level.
Our hypothesis that stronger avoidance of depredation mortality risk at coarse scales would free individuals to relax their avoidance of risk when making fine‐scale selection decisions was supported, as mountain lions across California exhibited a functional response in resource selection (P1). This functional response occurred because individuals that selected home ranges with lower depredation mortality risk had effectively reduced the availability of risk relevant to their finer scale resource selection decisions. Functional responses in resource selection have mainly been used to understand how resource selection decisions at a single scale (usually within home ranges) vary among individuals within populations relative to different environmental context (Mysterud and Ims 1998; Northrup et al. 2022). Classic functional responses in predation describe the relationship between consumption rate of prey and prey density, whereas functional responses in resource selection describe the general relationship between selection of any resource and its availability (Dupke et al. 2021; Mysterud and Ims 1998). As noted by Mysterud and Ims (1998), functional responses in resource selection bear some resemblance to functional responses in predation, although the two concepts were only formally linked more recently (Dupke et al. 2021). Measures of resource availability used to evaluate functional responses in resource selection have been linked to home range size and composition (Herfindal et al. 2009; Paolini et al. 2019). However, functional responses have not been used to quantify the relationship between selection decisions across scales by individual animals, despite the fact that selection at coarse scales constrains or enhances resource availability at finer scales. Indeed, our results reflect the fact that resource attributes of an individual's home range represent both resource use for second‐order selection and resource availability for third‐order selection (DeCesare et al. 2012; Johnson 1980). In California, the mean level of depredation mortality risk for a mountain lion within its home range was conditional on its response to risk when selecting a home range from the larger landscape (Table S10). In demonstrating this relationship, we show that a population‐level functional response in resource selection provides an empirical link between coarse‐ and fine‐scale resource selection decisions by individual animals. With this link established, future research may be able to draw robust conclusions about the relationship between hierarchical resource selection and key components of fitness (see Appendix S3 for our exploratory analysis).
In addition to human‐caused mortality, prey availability and vulnerability are important factors influencing mountain lion fitness. Herbaceous areas, such as grasslands, are likely associated with increased prey availability, as they provide critical forage for mule deer and other ungulates (Bose et al. 2018; Forrester and Wittmer 2019). However, they may not provide suitable cover to successfully stalk and kill prey (Elbroch and Wittmer 2012; Murphy and Ruth 2009). Forest and shrub cover should increase hunting success, especially if foraging prey concentrate in highly productive areas where they are more susceptible to predation (Sih 2005; Smith et al. 2019). Mountain lions consistently selected forest and shrub cover across both scales, but only selected herbaceous cover when selecting home ranges from the larger landscape (Figure 3). Divergent responses to a limiting factor across scales suggest that its influence was overcome at a coarse scale (Rettie and Messier 2000). Thus, our results suggest that mountain lions effectively address prey availability by selecting home ranges that provide access to herbaceous areas, whereas the importance of prey vulnerability persists across scales as dense stalking cover must be present within home ranges and at specific sites where deer are killed. When the influence of limiting factors persists across scales, animals frequently trade‐off between forage quality and risk (Dussault et al. 2005; Hebblewhite and Merrill 2009; Lima and Dill 1990). Across California, mountain lions selected vegetation classes influencing their foraging efficiency independently of the level of depredation mortality risk within their home ranges (Figure 3). Thus, while strong coarse‐scale avoidance of depredation mortality risk allowed individuals to relax their avoidance of risk at finer scales, we did not find evidence that it freed them to select features associated with foraging efficiency more strongly (P2). Trade‐offs are expected when forage and predation risk are correlated in space and time (Lima and Dill 1990). While depredation mortality risk increased slightly with proximity to stalking cover, spatial variation in risk was influenced more strongly by proximity to anthropogenic features. Thus, mountain lions were often able to select locations where their prey were most vulnerable with little consideration for depredation mortality risk.
Coarse‐scale resource selection decisions are believed to have the largest impact on fitness (Gaillard et al. 2010; Owen‐Smith, Fryxell, and Merrill 2010; Rettie and Messier 2000). Thus, mountain lions should select home ranges that provide access to prey, contain suitable habitat for successfully killing prey and minimise risk of human‐caused mortality. While mountain lion populations are unlikely to persist where individuals are unable to efficiently meet their energetic demands (Dellinger, Cristescu, et al. 2020; Dellinger, Gustafson, et al. 2020), individuals frequently select home ranges that expose them to risk of being killed by humans following livestock depredation. Our second hypothesis, that mountain lions would exhibit context‐dependent selection to mitigate depredation mortality risk at fine scales was supported, as they avoided risk more strongly and directed movements towards vegetation classes providing increased cover as they encountered greater risk along their movement paths (P3, Figure 5). Mountain lions therefore exhibited a hierarchically nested functional response to depredation mortality risk, a primary factor limiting their fitness (Benson et al. 2023). The population‐level functional response demonstrates that the level of depredation mortality risk across an individual's home range (availability of risk), which is determined by second‐order selection decisions, exerts a strong influence on their response to risk within the home range. Prey species often exhibit proactive strategies to control exposure to predation risk, but reactive antipredator responses also play an important role in mitigating elements of unpredictable risk (Creel 2018). For mountain lions, human‐caused mortality risk may be less predictable in areas with intermediate levels of development where behavioural cues used to assess risk are unreliable (Nisi, Benson, and Wilmers 2022). Although population‐level responses were consistent with strong proactive avoidance of depredation mortality risk, all mountain lions encountered some level of risk regardless of the strength of their avoidance at the landscape scale and within their home ranges. Thus, responding to risk encountered in ‘real‐time’ at the finest scales along movement paths with individual‐level functional responses is likely an important reactive component of risk‐avoidance strategies of animals.
Results from our analysis of spatially varying depredation mortality risk are consistent with previous research suggesting that mountain lions are at greatest risk of being killed following livestock depredation in areas characterised by intermediate levels of human presence (Benson et al. 2023; Nisi, Benson, and Wilmers 2022). Indeed, local roads, buildings and low‐intensity development are landscape features that are likely associated with the small‐scale hoof stock operations where the majority of depredation incidents that result in retaliatory killings occur (Dellinger et al. 2021). A limitation in our analysis was the lack of direct information about prey availability. However, prey vulnerability influences foraging success of ambush predators like mountain lions and other felids more strongly than availability (Hopcraft, Sinclair, and Packer 2005; Coon et al. 2020). Therefore, as stalking cover facilitates successful killing of prey, it effectively represented spatial variation in foraging efficiency of mountain lions (Elbroch and Wittmer 2012; Murphy and Ruth 2009) in our analysis of forage‐risk trade‐offs.
Mountain lions exhibited divergent responses to herbaceous areas across scales, suggesting open‐structure habitat may be undervalued in assessments of habitat suitability that are derived from analyses of within home range resource selection that do not explicitly consider landscape‐level selection. Maintaining landscapes with productive herbaceous areas interspersed between patches of forest and shrub cover should enable mountain lions to select home ranges providing adequate access to their primary prey, and to effectively stalk and kill prey within their home ranges. For populations that persist within fragmented landscapes that do not provide sufficient habitat to maintain suitable effective population sizes (Dellinger, Cristescu, et al. 2020), human‐caused mortality can lower population‐level survival rates and increase the risk of local extinction (Benson et al. 2016, 2019, 2023). Our results suggest coarse‐scale resource selection decisions are important for mountain lions to mitigate depredation mortality risk and that familiarity of surroundings within home ranges enables more efficient avoidance of risk as it is encountered along movement paths. Recent policies in portions of California mandate that nonlethal methods of livestock depredation management must be attempted before a permit for lethal removal of a mountain lion is issued (see Appendix S1 for details). These policies, along with investment in infrastructure (e.g., road crossing structures) to reduce rates of vehicle mortality, are likely to play an important role in reducing mortality of this species of conservation concern.
Johnson's (1980) hierarchy of resource selection is a foundational framework shaping our understanding of the spatial ecology of animals. Viewed through Johnson's (1980) orders of selection, scale has been widely recognised as a fundamental consideration in animal resource selection that affects inference on the relationship between selection and fitness (Boyce 2006; DeCesare et al. 2014; Mayor et al. 2009). Rettie and Messier (2000) hypothesised that an individual's resource selection decisions across scales should reflect the hierarchy of factors limiting their fitness. However, difficulty connecting individual‐level resource selection decisions across scales has limited understanding of whether coarse‐scale selection decisions can address limiting factors such that their influence does not persist at finer scales. Our work establishes that a functional response provides an intuitive, empirical link between individuals' coarse‐ and fine‐scale resource selection decisions. In support of Rettie and Messier's (2000) hypothesis, we showed that individuals that proactively avoided depredation mortality risk strongly when making coarse‐scale resource selection decisions were able to relax their avoidance of risk at finer scales. However, individuals also made context‐dependent decisions along movement paths within their home ranges, providing a reactive mechanism by which they responded to risk not overcome through coarse‐scale selection decisions.
Author Contributions
K.D.D., J.F.B. and J.A.D. designed the research. J.A.D. initiated the collaboration and compiled the data. K.D.D. and J.F.B. organized the data for analysis and analyzed the data. K.D.D. wrote the paper. All authors performed the research and contributed to revising the paper.
Conflicts of Interest
The authors declare no conflicts of interest.
Peer Review
The peer review history for this article is available at https://www.webofscience.com/api/gateway/wos/peer‐review/10.1111/ele.70035.
Supporting information
Data S1.
Acknowledgements
We thank J. V. Lombardi with the Mountain Lion Conservation Program, Wildlife Health Laboratory, California Department of Fish and Wildlife for providing some of the data used in our analyses. This work was funded and supported by the School of Natural Resources, University of Nebraska‐Lincoln (J.F.B. and K.D.D.); California Department of Fish and Wildlife (B.C., D.J.G., T.R.S., H.U.W. and J.A.D.); California Department of Fish and Wildlife (Federal Wildlife Restoration and Aid Grants #G1698020, G1798020, G1898020, G1998020, G2098020, G2198020, G2298020); US Forest Service; US Department of Agriculture—Wildlife Services; California Department of Parks and Recreation; Hoopa Tribe; and US Department of Defense (J.A.D.); The California State Department of Parks and Recreation, the California Department of Fish and Wildlife, The Nature Conservancy, the McBeth Foundation, the Anza‐Borrego Foundation, The Nature Reserve of Orange County, the NSF, the Foothill/Eastern Transportation Corridor Agency, San Diego County Association of Governments Environmental Mitigation Program, the San Diego Foundation, Felidae Conservation Fund, the Mountain Lion Foundation, the Santa Rosa Plateau Foundation, the Institute for Wildlife Studies and private donors (T.W.V.); the National Park Service, California State Parks, Santa Monica Mountains Fund, Santa Monica Mountains Conservancy, Mountains Recreation and Conservation Authority, and Calabasas Landfill (S.P.D.R. and J.A.S.); Audubon Canyon Ranch and its donors (QEM); NSF grants 0963022 and 1255913 and by the support of the Gordon and Betty Moore Foundation and the Peninsula Open Space Trust (C.C.W.). We thank Peter Mahoney for helpful discussions during data analysis. We also thank the many field technicians, graduate students, volunteers and other organisations that bolstered data collection efforts.
Editor: Andrew Sih
Funding: This study was supported by the School of Natural Resources, University of Nebraska‐Lincoln (J.F.B. and K.D.D.); California Department of Fish and Wildlife (B.C., D.J.G., T.R.S., H.U.W. and J.A.D.); California Department of Fish and Wildlife (Federal Wildlife Restoration and Aid Grants #G1698020, G1798020, G1898020, G1998020, G2098020, G2198020, G2298020); US Forest Service; US Department of Agriculture—Wildlife Services; California Department of Parks and Recreation; Hoopa Tribe; and US Department of Defense (J.A.D.); The California State Department of Parks and Recreation, the California Department of Fish and Wildlife, The Nature Conservancy, the McBeth Foundation, the Anza‐Borrego Foundation, The Nature Reserve of Orange County, the NSF, the Foothill/Eastern Transportation Corridor Agency, San Diego County Association of Governments Environmental Mitigation Program, the San Diego Foundation, Felidae Conservation Fund, the Mountain Lion Foundation, the Santa Rosa Plateau Foundation, the Institute for Wildlife Studies and private donors (T.W.V.); the National Park Service, California State Parks, Santa Monica Mountains Fund, Santa Monica Mountains Conservancy, Mountains Recreation and Conservation Authority, and Calabasas Landfill (S.P.D.R. and J.A.S.); Audubon Canyon Ranch and its donors (QEM); NSF grants 0963022 and 1255913 and by the support of the Gordon and Betty Moore Foundation and the Peninsula Open Space Trust (C.C.W.).
Data Availability Statement
Data and code that support the findings of this study are available on Zenodo (https://doi.org/10.5281/zenodo.14199407).
References
- Anderson, D. P. , Turner M. G., Forester J. D., et al. 2005. “Scale‐Dependent Summer Resource Selection by Reintroduced Elk in Wisconsin, USA.” Journal of Wildlife Management 69: 298–310. [Google Scholar]
- Basille, M. , Moorter B. V., Herfindal I., et al. 2013. “Selecting Habitat to Survive: The Impact of Road Density on Survival in a Large Carnivore.” PLoS One 8: e65493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benson, J. F. 2013. “Improving Rigour and Efficiency of Use‐Availability Habitat Selection Analyses With Systematic Estimation of Availability.” Methods in Ecology and Evolution 4: 244–251. [Google Scholar]
- Benson, J. F. , Dougherty K. D., Beier P., et al. 2023. “The Ecology of Human‐Caused Mortality for a Protected Large Carnivore.” Proceedings of the National Academy of Sciences of the United States of America 120: e2220030120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benson, J. F. , Mahoney P. J., Sikich J. A., et al. 2016. “Interactions Between Demography, Genetics, and Landscape Connectivity Increase Extinction Probability for a Small Population of Large Carnivores in a Major Metropolitan Area.” Proceedings of the Royal Society B: Biological Sciences 283: 20160957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benson, J. F. , Mahoney P. J., Vickers T. W., et al. 2019. “Extinction Vortex Dynamics of Top Predators Isolated by Urbanization.” Ecological Applications 29: e01868. [DOI] [PubMed] [Google Scholar]
- Benson, J. F. , Sikich J. A., and Riley S. P. D.. 2016. “Individual and Population Level Resource Selection Patterns of Mountain Lions Preying on Mule Deer Along an Urban‐Wildland Gradient.” PLoS One 11: e0158006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bleyhl, B. , Ghoddousi A., Askerov E., et al. 2021. “Reducing Persecution Is More Effective for Restoring Large Carnivores Than Restoring Their Prey.” Ecological Applications 31: e02338. [DOI] [PubMed] [Google Scholar]
- Bose, S. , Forrester T. D., Casady D. S., and Wittmer H. U.. 2018. “Effect of Activity States on Habitat Selection by Black‐Tailed Deer.” Journal of Wildlife Management 82: 1711–1724. [Google Scholar]
- Boyce, M. S. 2006. “Scale for Resource Selection Functions.” Diversity and Distributions 12: 269–276. [Google Scholar]
- Boyce, M. S. , Mao J. S., Merrill E. H., et al. 2003. “Scale and Heterogeneity in Habitat Selection by Elk in Yellowstone National Park.” Écoscience 10: 421–431. [Google Scholar]
- Brooks, M. E. , Kristensen K., Van Benthem K. J., et al. 2017. “glmmTMB Balances Speed and Flexibility Among Packages for Zero‐Inflated Generalized Linear Mixed Modeling.” R Journal 9: 378–400. [Google Scholar]
- Chapron, G. , Miquelle D. G., Lambert A., Goodrich J. M., Legendre S., and Clobert J.. 2008. “The Impact on Tigers of Poaching Versus Prey Depletion.” Journal of Applied Ecology 45: 1667–1674. [Google Scholar]
- Ciarniello, L. M. , Boyce M. S., Seip D. R., and Heard D. C.. 2007. “Grizzly Bear Habitat Selection is Scale Dependent.” Ecological Applications 17: 1424–1440. [DOI] [PubMed] [Google Scholar]
- Coon, C. A. C. , Mahoney P. J., Edelblutte E., McDonald Z., and Stoner D. C.. 2020. “Predictors of Puma Occupancy Indicate Prey Vulnerability is More Important Than Prey Availability in a Highly Fragmented Landscape.” Wildlife Biology 2020: 1–12. [Google Scholar]
- Creel, S. 2018. “The Control of Risk Hypothesis: Reactive vs. Proactive Antipredator Responses and Stress‐Mediated vs. Food‐Mediated Costs of Response.” Ecology Letters 21: 947–956. [DOI] [PubMed] [Google Scholar]
- Darimont, C. T. , Fox C. H., Bryan H. M., and Reimchen T. E.. 2015. “The Unique Ecology of Human Predators.” Science 349: 858–860. [DOI] [PubMed] [Google Scholar]
- DeCesare, N. J. , Hebblewhite M., Bradley M., Hervieux D., Neufeld L., and Musiani M.. 2014. “Linking Habitat Selection and Predation Risk to Spatial Variation in Survival.” Journal of Animal Ecology 83: 343–352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeCesare, N. J. , Hebblewhite M., Schmiegelow F., et al. 2012. “Transcending Scale Dependence in Identifying Habitat With Resource Selection Functions.” Ecological Applications 22: 1068–1083. [DOI] [PubMed] [Google Scholar]
- Dellinger, J. , Cristescu B., Ewanyk J., et al. 2020. “Using Mountain Lion Habitat Selection in Management.” Journal of Wildlife Management 84: 359–371. [Google Scholar]
- Dellinger, J. , Gustafson K. D., Gammons D. J., Ernest H. B., and Torres S. G.. 2020. “Minimum Habitat Thresholds Required for Conserving Mountain Lion Genetic Diversity.” Ecology and Evolution 10: 10687–10696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dellinger, J. , Macon D., Rudd J., Clifford D., and Torres S.. 2021. “Temporal Trends and Drivers of Mountain Lion Depredation in California, USA.” Human–Wildlife Interactions 15: 162–177. [Google Scholar]
- Dewitz, J. , and U.S. Geological Survey . 2021. “National Land Cover Database (NLCD) 2019 Products.”
- Dupke, C. , Peters A., Morellet N., and Heurich M.. 2021. “Holling Meets Habitat Selection: Functional Response of Large Herbivores Revisited.” Movement Ecology 9: 45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dussault, C. , Ouellet J.‐P., Courtois R., Huot J., Breton L., and Jolicoeur H.. 2005. “Linking Moose Habitat Selection to Limiting Factors.” Ecography 28: 619–628. [Google Scholar]
- Elbroch, L. M. , and Wittmer H. U.. 2012. “Puma Spatial Ecology in Open Habitats With Aggregate Prey.” Mammalian Biology 77: 377–384. [Google Scholar]
- Farr, T. G. , Rosen P. A., Caro E., et al. 2007. “The Shuttle Radar Topography Mission.” Reviews of Geophysics 45. [Google Scholar]
- Forrester, T. D. , and Wittmer H. U.. 2019. “Predator Identity and Forage Availability Affect Predation Risk of Juvenile Black‐Tailed Deer.” Wildlife Biology 2019: 1–12. [Google Scholar]
- Fortin, D. , Beyer H. L., Boyce M. S., Smith D. W., Duchesne T., and Mao J. S.. 2005. “Wolves Influence Elk Movements: Behavior Shapes a Trophic Cascade in Yellowstone National Park.” Ecology 86: 1320–1330. [Google Scholar]
- Frid, A. , and Dill L. M.. 2002. “Human‐Caused Disturbance Stimuli as a Form of Predation Risk.” Conservation Ecology 6: 11. [Google Scholar]
- Furnas, B. J. , Landers R. H., Paiste R. G., and Sacks B. N.. 2020. “Overabundance of Black‐Tailed Deer in Urbanized Coastal California.” Journal of Wildlife Management 84: 979–988. [Google Scholar]
- Gaillard, J.‐M. , Hebblewhite M., Loison A., et al. 2010. “Habitat–Performance Relationships: Finding the Right Metric at a Given Spatial Scale.” Philosophical Transactions of the Royal Society, B: Biological Sciences 365: 2255–2265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaynor, K. M. , Cherry M. J., Gilbert S. L., et al. 2021. “An Applied Ecology of Fear Framework: Linking Theory to Conservation Practice.” Animal Conservation 24: 308–321. [Google Scholar]
- Gelman, A. 2008. “Scaling Regression Inputs by Dividing by Two Standard Deviations.” Statistics in Medicine 27: 2865–2873. [DOI] [PubMed] [Google Scholar]
- Getz, W. M. , Fortmann‐Roe S., Cross P. C., Lyons A. J., Ryan S. J., and Wilmers C. C.. 2007. “LoCoH: Nonparameteric Kernel Methods for Constructing Home Ranges and Utilization Distributions.” PLoS One 2: e207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillies, C. S. , Hebblewhite M., Nielsen S. E., et al. 2006. “Application of Random Effects to the Study of Resource Selection by Animals.” Journal of Animal Ecology 75: 887–898. [DOI] [PubMed] [Google Scholar]
- Godvik, I. M. R. , Loe L. E., Vik J. O., Veiberg V., Langvatn R., and Mysterud A.. 2009. “Temporal Scales, Trade‐Offs, and Functional Responses in Red Deer Habitat Selection.” Ecology 90: 699–710. [DOI] [PubMed] [Google Scholar]
- Hadfield, J. D. , Wilson A. J., Garant D., Sheldon B. C., and Kruuk L. E. B.. 2010. “The Misuse of BLUP in Ecology and Evolution.” American Naturalist 175: 116–125. [DOI] [PubMed] [Google Scholar]
- Hebblewhite, M. , and Merrill E. H.. 2009. “Trade‐Offs Between Predation Risk and Forage Differ Between Migrant Strategies in a Migratory Ungulate.” Ecology 90: 3445–3454. [DOI] [PubMed] [Google Scholar]
- Herfindal, I. , Tremblay J.‐P., Hansen B. B., Solberg E. J., Heim M., and Sæther B.‐E.. 2009. “Scale Dependency and Functional Response in Moose Habitat Selection.” Ecography 32: 849–859. [Google Scholar]
- Hopcraft, J. G. C. , Sinclair A. R. E., and Packer C.. 2005. “Planning for Success: Serengeti Lions Seek Prey Accessibility Rather Than Abundance.” Journal of Animal Ecology 74: 559–566. [Google Scholar]
- Johnson, C. J. , Parker K. L., and Heard D. C.. 2001. “Foraging Across a Variable Landscape: Behavioral Decisions Made by Woodland Caribou at Multiple Spatial Scales.” Oecologia 127: 590–602. [DOI] [PubMed] [Google Scholar]
- Johnson, D. H. 1980. “The Comparison of Usage and Availability Measurements for Evaluating Resource Preference.” Ecology 61: 65–71. [Google Scholar]
- Knopff, A. A. , Knopff K. H., Boyce M. S., and Clair C. C. S.. 2014. “Flexible Habitat Selection by Cougars in Response to Anthropogenic Development.” Biological Conservation 178: 136–145. [Google Scholar]
- Lamb, C. T. , Ford A. T., McLellan B. N., et al. 2020. “The Ecology of Human–Carnivore Coexistence.” Proceedings of the National Academy of Sciences of the United States of America 117: 17876–17883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lima, S. L. 1998. “Nonlethal Effects in the Ecology of Predator‐Prey Interactions.” Bioscience 48: 25–34. [Google Scholar]
- Lima, S. L. , and Dill L. M.. 1990. “Behavioral Decisions Made Under the Risk of Predation: A Review and Prospectus.” Canadian Journal of Zoology 68: 619–640. [Google Scholar]
- Lowry, H. , Lill A., and Wong B. B. M.. 2013. “Behavioural Responses of Wildlife to Urban Environments.” Biological Reviews 88: 537–549. [DOI] [PubMed] [Google Scholar]
- Manly, B. F. L. , McDonald L., Thomas D. L., McDonald T. L., and Erickson W. P.. 2002. Resource Selection by Animals: Statistical Design and Analysis for Field Studies. Second ed. New York, New York: Kluwer Academic Publishers. [Google Scholar]
- Mason, T. H. E. , and Fortin D.. 2017. “Functional Responses in Animal Movement Explain Spatial Heterogeneity in Animal‐Habitat Relationships.” Journal of Animal Ecology 86: 960–971. [DOI] [PubMed] [Google Scholar]
- Mayor, S. J. , Schneider D. C., Schaefer J. A., and Mahoney S. P.. 2009. “Habitat selection at multiple scales.” Écoscience 16: 238–247. [Google Scholar]
- McGarigal, K. , Wan H. Y., Zeller K. A., Timm B. C., and Cushman S. A.. 2016. “Multi‐Scale Habitat Selection Modeling: A Review and Outlook.” Landscape Ecology 31: 1161–1175. [Google Scholar]
- McLoughlin, P. D. , Dunford J. S., and Boutin S.. 2005. “Relating Predation Mortality to Broad‐Scale Habitat Selection.” Journal of Animal Ecology 74: 701–707. [Google Scholar]
- McNamara, J. M. 1987. “Starvation and Predation as Factors Limiting Population Size.” Ecology 68: 1515–1519. [Google Scholar]
- Montgomery, R. A. , Raupp J., Miller S. A., et al. 2022. “The Hunting Modes of Human Predation and Potential Nonconsumptive Effects on Animal Populations.” Biological Conservation 265: 109398. [Google Scholar]
- Muff, S. , Signer J., and Fieberg J.. 2020. “Accounting for Individual‐Specific Variation in Habitat‐Selection Studies: Efficient Estimation of Mixed‐Effects Models Using Bayesian or Frequentist Computation.” Journal of Animal Ecology 89: 80–92. [DOI] [PubMed] [Google Scholar]
- Murphy, K. , and Ruth T. K.. 2009. “Diet and Prey Selection of a Perfect Predator.” In Cougar: Ecology and Conservation, 118–137. Chicago, Illinois, USA: University of Chicago Press. [Google Scholar]
- Mysterud, A. , and Ims R. A.. 1998. “Functional Responses in Habitat Use: Availability Influences Relative Use in Trade‐Off Situations.” Ecology 79: 1435–1441. [Google Scholar]
- Nisi, A. C. , Benson J. F., and Wilmers C. C.. 2022. “Puma Responses to Unreliable Human Cues Suggest an Ecological Trap in a Fragmented Landscape.” Oikos 2022: e09051. [Google Scholar]
- Nisi, A. C. , Suraci J. P., Ranc N., et al. 2021. “Temporal Scale of Habitat Selection for Large Carnivores: Balancing Energetics, Risk and Finding Prey.” Journal of Animal Ecology 91: 182–195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Northrup, J. M. , Vander Wal E., Bonar M., et al. 2022. “Conceptual and Methodological Advances in Habitat‐Selection Modeling: Guidelines for Ecology and Evolution.” Ecological Applications 32: e02470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ordiz, A. , Aronsson M., Persson J., Støen O.‐G., Swenson J. E., and Kindberg J.. 2021. “Effects of Human Disturbance on Terrestrial Apex Predators.” Diversity 13: 68. [Google Scholar]
- Owen‐Smith, N. , Fryxell J. M., and Merrill E. H.. 2010. “Foraging Theory Upscaled: The Behavioural Ecology of Herbivore Movement.” Philosophical Transactions of the Royal Society, B: Biological Sciences 365: 2267–2278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paolini, K. E. , Strickland B. K., Tegt J. L., VerCauteren K. C., and Street G. M.. 2019. “The Habitat Functional Response Links Seasonal Third‐Order Selection to Second‐Order Landscape Characteristics.” Ecology and Evolution 9: 4683–4691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preisser, E. L. , and Bolnick D. I.. 2008. “The Many Faces of Fear: Comparing the Pathways and Impacts of Nonconsumptive Predator Effects on Prey Populations.” PLoS One 3: e2465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prima, M.‐C. , Duchesne T., and Fortin D.. 2017. “Robust Inference From Conditional Logistic Regression Applied to Movement and Habitat Selection Analysis.” PLoS One 12: e0169779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Core Team . 2023. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. [Google Scholar]
- Rettie, W. J. , and Messier F.. 2000. “Hierarchical Habitat Selection by Woodland Caribou: Its Relationship to Limiting Factors.” Ecography 23: 466–478. [Google Scholar]
- Ripari, L. , Premier J., Belotti E., et al. 2022. “Human Disturbance Is the Most Limiting Factor Driving Habitat Selection of a Large Carnivore Throughout Continental Europe.” Biological Conservation 266: 109446. [Google Scholar]
- Sih, A. 1980. “Optimal Behavior: Can Foragers Balance Two Conflicting Demands?” Science 210: 1041–1043. [DOI] [PubMed] [Google Scholar]
- Sih, A. 2005. “Predator‐Prey Space Use as an Emergent Outcome of a Behavioral Response Race.” In Ecology of Predator‐Prey Interactions, 240–255. Oxford, New York: Oxford University Press. [Google Scholar]
- Smith, J. A. , Donadio E., Pauli J. N., Sheriff M. J., Bidder O. R., and Middleton A. D.. 2019. “Habitat Complexity Mediates the Predator–Prey Space Race.” Ecology 100: e02724. [DOI] [PubMed] [Google Scholar]
- Spitz, D. B. , Hebblewhite M., and Stephenson T. R.. 2020. “Habitat Predicts Local Prevalence of Migratory Behaviour in an Alpine Ungulate.” Journal of Animal Ecology 89: 1032–1044. [DOI] [PubMed] [Google Scholar]
- Suraci, J. P. , Clinchy M., Zanette L. Y., and Wilmers C. C.. 2019. “Fear of Humans as Apex Predators Has Landscape‐Scale Impacts From Mountain Lions to Mice.” Ecology Letters 22: 1578–1586. [DOI] [PubMed] [Google Scholar]
- Therneau, T. M. , and Lumley T.. 2015. “Package ‘survival’.” R Top Doc 128: 28–33. [Google Scholar]
- Thurfjell, H. , Ciuti S., and Boyce M. S.. 2014. “Applications of Step‐Selection Functions in Ecology and Conservation.” Movement Ecology 2: 1–2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Beest, F. M. , Mysterud A., Loe L. E., and Milner J. M.. 2010. “Forage Quantity, Quality and Depletion as Scale‐Dependent Mechanisms Driving Habitat Selection of a Large Browsing Herbivore.” Journal of Animal Ecology 79: 910–922. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data S1.
Data Availability Statement
Data and code that support the findings of this study are available on Zenodo (https://doi.org/10.5281/zenodo.14199407).