

SUMMARY
The fundamental goal of tissue engineering is to functionally restore or improve damaged tissues or organs. Here, in the small bowel, we address this using an in vivo xenograft preclinical acute damage model. We investigated the therapeutic capacity of human intestinal organoids (HIOs), which are generated from human pluripotent stem cells (hPSCs), to repair damaged small bowel. We hypothesized that the HIO’s cellular complexity would allow it to sustain transmural engraftment. To test this, we developed a rodent injury model, where through luminal delivery, we demonstrated that fragmented HIOs engraft, proliferate, and persist throughout the bowel following repair. Not only was restitution of the mucosal layer observed, but significant incorporation was observed in the muscularis and vascular endothelium. Further analysis characterized sustained cell type presence within the regenerated regions, retention of proximal regionalization and the neo-epithelia’s function. These findings demonstrate the therapeutic importance of mesenchyme for intestinal injury repair.
Keywords: human intestinal organoid, enteroid, cell therapy, intestine
eTOC BLURB
Poling et al., have described using human intestinal organoids as a reinforcing material, or cell therapy, for damaged small intestine in a pre-clinical animal model. They observed that the human reinforcing material incorporated into a variety of sustained cell types within the epithelium and mesenchyme of the regenerated small intestine.
Graphical Abstract

INTRODUCTION
Enteroids are in vitro, epithelial-only structures that were first derived from patient intestinal crypts (Enteroids). They can be made from biopsies and full-thickness surgical samples throughout the intestine. Enteroids can also be derived from the epithelium of in vitro HIOs (iv-Enteroids) as well as from the isolated crypts of full-thickness transplanted HIOs (txp-Enteroids)1,2. While they contain the capacity to generate differentiated intestinal epithelial cell subtypes, such as enterocytes, goblet cells, Paneth cells, and enteroendocrine cells, they lack the mesenchymal, neuronal, endothelial, and immune compartments present in the human intestine. Enteroids have been used to replace damaged intestinal epithelium in animal models3–12. Furthermore, an enteroid-based therapeutic is currently being clinically evaluated by Tokyo Medical and Dental University in Japan (Trial UMIN000013524). In the trial, biopsies from healthy gut regions are obtained from patients with inflammatory bowel disease (IBD) with refractory ulcers and used to generate enteroids. Through a second procedure, the patients’ own healthy enteroids, expanded in vitro, are delivered as a reinforcing material/cell therapy product to heal ulcers. Colonic ulcers such as those being treated in the current trial, can also occur throughout the gastrointestinal tract due to many conditions that include peptic ulcers, and ischemic injuries associated with surgical conditions and radiation therapies which result in significant morbidity13. First line treatments of the underlying condition typically restore the damaged epithelium, however, in chronic conditions where the underlying mesenchyme is damaged, transmural bowel injury persists14–16. For this subset of patients, there is a paucity of therapeutic options to regenerate chronically damaged bowel. Even when a treatment successfully controls the underlying damaging condition, it does not always result in bowel regeneration, leaving patients experiencing symptoms impacting quality of life17. A potential cause may be chronic damage within the supporting mesenchymal and vascular compartments which are essential for epithelial regeneration18. Currently, few clinical treatment options exist when this happens. While some patients may benefit from surgical intervention when their diseased region is focal, it is also associated with additional morbidity, and it does not address all patients with complex or multifocal disease. To broaden the treatment modalities available for these patients, it is essential to develop novel therapeutic strategies. We hypothesize that luminal delivery of hPSC organoid products to damaged bowel as a cell therapy source could prove beneficial in sustained transmural restoration beyond the mucosa.
HIOs are generated via stepwise differentiation of hPSCs1,19. HIOs contain both epithelial and mesenchymal cell types in vitro19. Utilizing an immunocompromised host as a bioreactor for engraftment and maturation, transplanted HIOs develop into structures reminiscent of human intestine, including a crypt/villus axis, vasculature, and smooth muscle layers20. In the present study, we tested the HIO as a cell therapy source for damaged small bowel and investigate contributions to repair beyond the mucosa. We speculate that regeneration of the mesenchymal-epithelial niche is required for robust and sustained engraftment of damaged bowel for functional transmural restoration16. These preclinical data are an essential stepping-stone for planning human trials using HIO-based cellular products.
RESULTS
An Acute Injury Model was used to Test the Therapeutic Capacity of HIOs
Our experimentation utilized the H1 embryonic stem cell line, which was modified in house to constitutively express green fluorescent protein (GFP) for ease of downstream xenograft analysis. The modified cell line was characterized and determined to pass quality control metrics (Figure S1A-B). HIOs and txp-Enteroids, both made from the H1-GFP line to reduce genetic variability between groups, were used as cell therapy sources in damaged small bowel limbs of rodents. Immunocompromised Rag1 and Il2rg knockout rats were utilized as host subjects to surgically create an isolated segment of small bowel that underwent chemical and mechanical injury before reseeding with HIOs, txp-Enteroids or media alone as sham (Figure 1A)21,22. The surgical procedure resulted in a blind end-to-side “Y” segment, or loop, which was temporarily tied off with fast absorbing chromic suture such that after a few days the segment could drain distally and be exposed to host enteric content (Figure 1B). A significant advantage of this model is the ability to provide a brief window for HIO/txp-Enteroid retention, epithelial restitution, and engraftment to occur following acute transmural damage in an immunocompromised host. Bowel continuity was retained with this approach, so as not to impair the host’s overall digestive function. Immediately after injury, the isolated segment of bowel was predominantly denuded of epithelium with exposed and damaged mesenchyme, as observed histologically and via topographical micrographs, when compared to healthy bowel (Figure 1 C-D). Mortality and survival rates were similar between the three experimental groups (Figure 1E).
Figure 1. Experimental Design and Mucosectomy Surgical Injury Model.
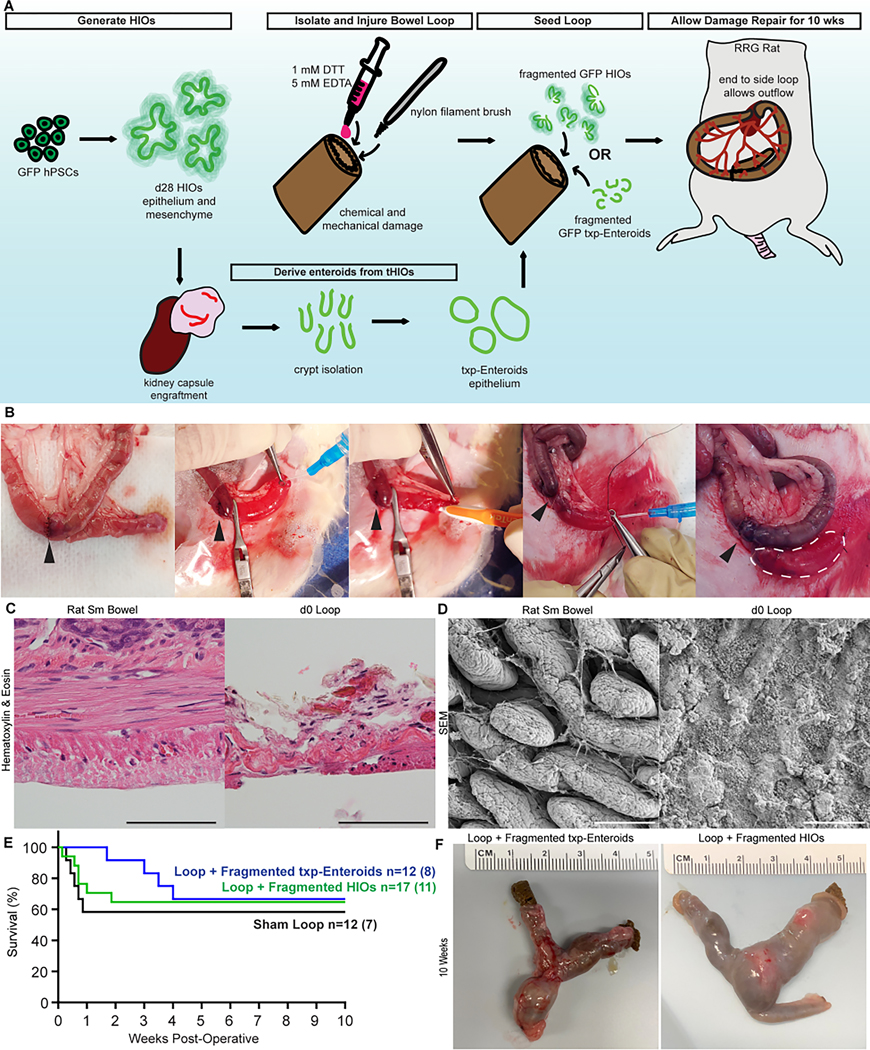
(A) Experimental design schematic. (B) Surgical imagery of loop creation, damage, and application of cell therapy product. Arrowhead indicates anastomosis. White dashed line outlines loop. (C) Histology of healthy and injured rat small bowel. (D) Topography of healthy and injured rat small bowel. (E) Kaplan-Meier curve associated with surgeries. (F) Harvest images of damaged loops reseeded with txp-Enteroids and HIOs after 10 weeks. All scale bars = 100 μm.
Fragmented HIOs and txp-Enteroids Incorporate During Intestinal Repair
At the ten-week post-operative time point, loops were harvested and evaluated (Figure 1F). In loops supplemented with GFP HIOs, we first observed discrete engraftment as organoid units (Figure 2A). Therefore, we devised a fragmentation strategy to promote widespread incorporation of the cellular products as opposed to isolated engraftment (Figure 2B). By doing so, fragments of varying sizes were created, with the highest proportion around 30 μm in size for both fragmented HIOs and fragmented txp-Enteroids (Figure 2C). The average number of cells per structure was also quantified after dissociation, so that consistent amounts of cellular product could be administered (Figure 2D). When administering fragmented GFP HIOs, regions expressing live-GFP were observed and comprised an average of 17% of the loop’s surface area (Figure 2E-F). Immunohistochemistry for a human specific DNA binding protein (KU80), revealed robust human cellular incorporation within the dissected GFP+ regions of HIO fragment reseeded loops (Figure 2G). In addition, KU80+ cells were observed throughout the thickness of the intestine. This was further highlighted by the quantification of KU80+ cells within full-thickness fields of view that contained human cellular incorporation. The average proportion of KU80+ human cells per field was quantified to be 52% (Figure 2H). In loops supplemented with fragmented GFP txp-Enteroids, regions expressing live-GFP were also observed, however they were significantly reduced and comprised 2% of the loop’s surface area on average (Figure 2I-J). KU80+ cell incorporation was observed within the dissected GFP+ regions of txp-Enteroid fragment supplemented loops and presented exclusively within the epithelium (Figure 2K). The proportion of KU80+ cells in full-thickness samples supplemented with fragmented txp-Enteroids was 3% per field of view, a significant reduction compared to loops supplemented with fragmented HIOs (Figure 2L).
Figure 2. Organoid Fragmentation was Essential for Expansion during Regeneration.

(A) Representative histology of a Loop + Whole HIOs and serial section stained for human cells (KU80, brown). (B) Brightfield and live-GFP fluorescence imagery of HIOs and txp-Enteroids before and after fragmentation. (C) Histograms of HIO and txp-Enteroid fragment sizes. (D) Graph of cell number in HIOs and txp-Enteroids represented as mean ± SD. (E) Brightfield and live-GFP fluorescence image of a harvested Loop + Fragmented HIOs after 10 weeks; dashed line indicates tissue edge. (F) Quantification of live-GFP surface area in Loop + Fragmented HIOs samples. (G) Representative section stained for human cells (KU80, brown) in a live-GFP+ region in a Loop + Fragmented HIO. Boxed inset depicts margin/boundary of human (KU80, brown) and rat tissue. (H) Quantification of KU80+ cells as a percentage of cells in each 20x field of view for areas with positive human cellular incorporation in Loop + Fragmented HIOs samples. (I) Brightfield and live-GFP fluorescence image of a harvested Loop + Fragmented txp-Enteroids after 10 weeks; dashed line indicates tissue edge. (J) Quantification of live-GFP surface area in Loop + Fragmented txp-Enteroids samples. (K) Representative section stained for human cells (KU80, brown) in a live-GFP+ region in a Loop + Fragmented txp-Enteroids. (L) Quantification of KU80+ cells as a percentage of cells in each 20x field of view for areas with positive human cellular incorporation in Loop + Fragmented txp-Enteroids samples. All scale bars = 100 μm. For F vs. J, p=0.0023 and for H vs. L, p<0.0001 from Mann-Whitney Tests.
We chose to use txp-Enteroids from the H1-GFP line, for ease of analysis, taking advantage of the live-GFP expression. Because we recognize that enteroids can be made from three sources, baseline analysis of each type was performed in case a phenotypic difference specific to the enteroid source accounted for the low engraftment. Through brightfield and confocal imaging, Enteroids, iv-Enteroids and txp-Enteroids all demonstrated a similar spherical phenotype in WNT-rich media with a high proportion of cells MKI67+ throughout (Figure S2A-B). Quantitative real-time PCR was also performed and there were no differences between groups for the transcription of an intestinal epithelial cell marker, CDX2, a proliferation marker, MKI67, a surrogate marker of stem cell activity and the intestinal crypt, OLFM4, a stem cell marker, LGR5, a Paneth cell marker, LYZ, and a pan-mesenchymal marker, VIM (Figure S2C). Goblet cell marker, MUC2, and enteroendocrine cell marker, CHGA, did not cycle above background in any of the groups. Flow cytometry was performed to quantify the proliferation across groups, which was over 95% for each enteroid type, suggesting that they were all similarly enriched for stemness (Figure S2D-G). After validation of txp-Enteroid engraftment in our damage model, we continued more in-depth analysis using only fragmented HIOs as our cell source, because it allowed us to test our overarching hypothesis.
HIOs Contributed to a Diverse Assortment of Sustained Cell Types after Regeneration
To further characterize the cellular components of the regenerated loops a series of immunostainings was performed. First, to demonstrate the extent of human cellular contributions within loops supplemented with fragmented GFP HIOs, a tile scan was performed on a region of GFP+ loop stained for KU80 (Figure 3A, left). Human cells were observed within the mucosa and the muscularis, which displayed proper alignment (Figure 3A, right). Immunostaining for MKI67 demonstrated that the stereotypical proliferative zonation of the intestine had been reestablished in both sham and HIO supplemented loops (Figure 3B). To further support crypt formation and stem cell activity, an antiapoptotic factor (OLFM4) was stained for and observed to localize correctly in both groups (Figure 3C). Mucin producing goblet cells, antimicrobial Paneth cells, and hormone secreting enteroendocrine cells were also observed in both groups indicating that epithelial cell specification was occurring in both groups (Figure 3D-F). The humanization of the muscularis was further highlighted by co-immunostaining for smooth muscle (ACTA2) and GFP (Figure 3G).
Fig. 3. Fragmented HIOs contribute to tissue regeneration of damaged bowel in vivo after 10 weeks.

(A) Tile scan of a Loop + Fragmented HIOs stained for human cells (KU80, brown); scale bar = 0.5 cm. Higher magnification images of KU80+ cells within the mucosa and muscularis from tile scan; scale bar = 50 μm. (B) Representative images of Sham Loop and Loop + Fragmented HIOs stained for a proliferation marker (MKI67, pink), epithelium (CDH1, white) and human cells (GFP, green). (C) Representative images of Sham Loop and Loop + Fragmented HIOs stained for a surrogate marker of stem cell activity and the intestinal crypt (OLFM4, pink) and epithelium (CDH1, green). (D) Representative images of Sham Loop and Loop + Fragmented HIOs stained for human cells (KU80, brown) and goblet cells (alcian blue for mucin, blue). (E) Representative images of Sham Loop and Loop + Fragmented HIOs stained for human cells (GFP, green) and Paneth cells (DEFA5, pink). (F) Representative images of Sham Loop and Loop + Fragmented HIOs stained for human cells (GFP, green), and enteroendocrine cells (CHGA, white). (G) Representative images of Sham Loop and Loop + Fragmented HIOs stained for human cells (GFP, green), and smooth muscle (ACTA2, pink). (H) Representative images of Sham Loop and Loop + Fragmented HIOs stained for a pan-neuronal marker (TUBB3, brown). Arrow heads indicate neuronal bundles. (I) Representative images of Sham Loop and Loop + Fragmented HIOs stained for human cells (GFP, green), blood vessels (VWF, pink), and human blood vessels (hCD31, white). Single channel insets are within the white dashed box. (B-I) Scale bars = 50 μm.
Next, we explored whether the host innervation, part of the enteric nervous system (ENS), responsible for a variety of functions including motility, maintained its localization within regenerated tissue. Immunostaining for a pan-neuronal marker (TUBB3) confirmed that the host innervation was not displaced after human cellular reconstitution suggesting that its function may also be preserved (Figure 3H). Vascularization was also investigated through co-staining for a cross-species endothelial marker (VWF) and a human specific marker (hCD31) using GFP expression as a secondary confirmation for human cells (Figure 3I). Human vasculature was exclusively present within loops supplemented with HIOs, suggesting HIOs also possess competent endothelial cell precursors that contributed to vascular regeneration.
Clinically relevant regional differences exist throughout the gastrointestinal tract, so we further investigated regionality through protein expression compared to native human bowel. As HIOs are patterned as proximal small bowel and were engrafted into the distal small bowel, we sought to determine if regionalization was maintained 23. Immunostaining for a regional marker expressed in the small intestine but not the colon or rectum (GATA4), revealed positive expression in HIO supplemented loops similar to that in human small bowel samples (Figure S3A)24. Next, brush border enzymes important for digestion and specific to the small bowel were interrogated. Three such enzymes, human specific DPPIV, ALP and SI, were detected in HIO supplemented loops with localization comparable to that of human small bowel (Figure S3B). Finally, distal and colonic markers were also interrogated as a form of further validation of regional specification. A distal transcription factor (SATB2), colonic mucin (MUC5B) and colonocyte marker (MS4A12) were not detected in HIO supplemented loops but present in human colon samples (Figure S3C). Taken together, positive protein expression of proximal intestinal markers along with the absence of distal markers suggest that the regionality of the loop’s HIO derived neo-epithelium was maintained during the engraftment and expansion of the HIO fragments.
Electrophysiology of HIO Neo-epithelia Demonstrates Functionality
To gain functional insight, Ussing chamber assays were performed ten weeks postoperatively on the epithelia of healthy rat small bowel, sham loops, and live-GFP expressing loop regions (Figure 4A). Epithelia were exposed to a series of chemical stimuli while electrophysical properties were continuously recorded. Epithelia from all three groups were responsive to challenges with forskolin, 3-isobutyl-1-methylxanthine (IBMX) and bumetanide as observed in real time recordings (Figure 4B-D). Changes in short circuit current (Isc), a reflection of active ion transport, were consistent with the known method of action of each compound applied for all groups (Figure 4E)25. This demonstrates that appropriate cyclic adenosine monophosphate regulation and function of ion channels in the neo-epithelia was occurring. Baseline readings of transepithelial electrical resistance (TEER) were also measured (Figure 4F). Resistance values without the muscularis and serosal layers have been previously documented between 20 and 45 Ω*cm2 for rat small intestine26. In the adult human small intestine, TEER values have been previously documented between 50 and 100 Ω*cm2 27. While some of the HIO supplemented loop TEER values fall within the established TEER range for adult human small intestine, others were observed to be lower, likely reflective of the immature nature of HIOs and the contribution from regenerated rat bowel28.
Fig. 4. Neo-epithelia of reseeded loops are responsive to chemical stimuli after 10 weeks.

(A) Representative merged brightfield and live-GFP images of healthy rat small bowel (proximal to the loop), Sham Loop, and Loop + Fragmented HIOs mounted in an Ussing assay slider; dashed circle outlines slider opening. (B) Representative time course of healthy rat small bowel Isc measured in Ussing Chamber experiments. (C) Representative time course of Sham Loop Isc measured in Ussing Chamber experiments. (D) Representative time course of Loop + Fragmented HIOs Isc measured in Ussing Chamber experiments. (E) Graphs of calculated changes in Isc in response to 10 μM Forskolin, 100 μM IBMX, and 100 μM Bumetanide across sample groups. (F) Graph of baseline TEER across sample groups. (G) Graph of FITC-dextran permeability across sample groups. (H) Calculated FITC flux from graph in G. Bar graphs are represented as mean ± SD. Kruskal-Wallis test p-values were not significant between groups in E-H.
To further assess the integrity of the loop neo-epithelium, paracellular permeability was evaluated. Fluorescein isothiocyanate (FITC)-dextran, 4kDa in size, was added to the apical chamber of each specimen and samples were collected from the basolateral chamber for evaluation of fluorescence intensity over time (Figure 4G). From this dataset, FITC-dextran flux values were calculated and no difference between groups was observed (Figure 4H), indicating that HIO supplemented loop neo-epithelium established a functional and sufficient barrier during the regeneration and healing process.
Fragmented HIOs were Retained within the Site of Administration
As we demonstrated proliferation of HIO cellular products, we next investigated the biodistribution and potential for undesirable expansion. As a first line assessment of safety, samples were collected from various off target organs from ten-week postoperative rats supplemented with HIOs. Portions of the brain, colon, heart, kidney, liver, lung, and small bowel were collected for histology and PCR. No gross abnormalities or tumor formation was observed, which was consistent with our experience of preforming whole organoid transplants where hosts have been void of tumors20,29–34. When immunostaining for KU80 across serial sections through 2 mm of tissue thickness, KU80+ cells were only observed in the transplanted HIO positive control (Figure S4A). To further investigate the presence of human cells, Alu-based real-time PCR was performed to discern human from rodent cells35,36. To determine the threshold for significance, a titration was performed on a known human gDNA sample and related back to cellular equivalence (Table S1). Then, experimental samples were run, including the H1 GFP cell line and the various off target organs. No Ct data points were found to be below the Ct value threshold for human cell presence (Figure S4B, raw values in Table S2) supporting the lack of human cell migration within the hosts. As an additional measure, tissue samples from slides were rehydrated and their gDNA isolated for Alu-based real-time PCR. Rat small bowel, serial sections from loops with KU80+ cells, and human small bowel samples were all analyzed. Only the samples with known human cellular content were observed below the Ct value threshold for human cell presence (Figure S4C). This served as a third measure to validate human cell incorporation into the regenerated loops receiving HIO fragment therapy.
DISCUSSION
We have established a preclinical model to assess intestinal repair using hPSC generated HIOs. From a basic science standpoint, this model system is advantageous because of the efficient engraftment and related ability to study the regenerated tissue ex vivo. Clinically, these data support the strategy of new treatment paradigms for refractive transmural ulcerative diseases. Here, we report HIO fragment therapy results in an average engraftment of 17% by surface area after ten weeks, compared to 2% when using txp-Enteroids. Likewise, Yui et al. previously reported a limited engraftment/expansion rate of donor cells in colitic mice as only 0.02% of cells or about 100 cells per mouse four weeks post-transplantation when using enteroids3. Several similar reports have also demonstrated the ability of enteroids to contribute to intestinal and colonic epithelial healing, however efficiencies were rarely reported4–9,11,12,37. Consistent with our data on retained regional identity following engraftment, Sugimoto et al. demonstrated sites of ileal enteroid engraftment retained their small bowel identity within the mouse colon37. To our knowledge, this is the first report of using multi-lineage seeding material, epithelial and mesenchymal, to achieve in vivo transmural intestinal repair as a cell therapy without loop/segment removal from the host38,39. This is of high clinical relevance because it allows for an epithelial stem cell niche to be generated exclusively by the cell therapy source in areas where the native mesenchyme has failed to support natural restitution and healing or in which the mesenchyme is itself contributing to the disease etiology. With more recent efforts in generating hPSC derived organoids that represent more diverse and specific regions of the gastrointestinal tract, it is reasonable to speculate that this platform could be extended to additional areas, such as the colon, stomach and esophagus34,40–42.
Limitations of Study
While our preclinical data is promising, the current xenograft model has limited applicability to human gastrointestinal diseases due to inadequate immune representation. Additionally, the HIOs used were not generated in a GMP xeno-free manner, which will be vital for clinical translation. To enhance therapeutic potential, we aim to develop a refined delivery method, potentially using a biological adhesive or hydrogel, to target specific sites of ulcerative damage. Furthermore, next generation organoid models that incorporate neuronal and immune components will be essential for maximal clinical impact.
STAR METHODS
Resource Availability
Lead Contact
Further information and requests for resources should be directed to and will be fulfilled by the lead contact, Michael Helmrath (michael.helmrath@cchmc.org).
Materials Availability
The H1 GFP cell line generated in this study is available, however Material Transfer Agreements between institutions and WiCell would be required.
Data and Code Availability
Data reported in this paper is available from the lead contact upon request.
This paper does not generate original code.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact.
EXPERIMENTAL MODEL AND STUDY PARTICIPANT DETAILS
Human Tissue
Human tissue collection was performed with the prior approval of Cincinnati Children’s Hospital Medical Center’s (CCHMC) Institutional Review Board (Tissue Characterization, Study No. 2014–0427). Surgical samples of pathologically normal adult human small bowel and colon were obtained from patients undergoing bariatric or revision/resection procedures between the ages of 14 and 25 years old. Informed consent or assent was obtained from all patients and/or parent/legal guardians as appropriate. Additional de-identified samples of pathologically normal colon were obtained through CCHMC’s Discover Together Biobank. All human tissue was utilized in accordance with institutional ethics guidelines.
Immunocompromised Rats
All animal procedures and experiments were performed with the prior approval of Cincinnati Children’s Hospital Medical Center’s (CCHMC) Institutional Animal Care and Use Committee (Protocol No. 2021–0060). Both males and females were utilized for experiments. Adult immunodeficient rats with Rag1 and Il2rg gene deletions (RRG) with ages between three and six months of age were used for mucosectomy experiments (founders were purchased from Transgenesis Rat ImmunoPhenomic Platform, Nantes, France, then bred in-house). Rats were housed in CCHMC’s barrier animal vivarium and handled humanely in accordance with the NIH Guide for the Care and Use of Laboratory Animals. RRG rats were primarily fed standard autoclaved chow and provided water bottles supplemented with fluconazole (0.1 mg/ml, NorthStar Rx, LLC.). Both food and water were provided ad libitum before and after surgeries. A single dose of carprofen (5 mg/kg) was administered for pain management at the end of the mucosectomy procedure. Rats were monitored daily for three days postoperatively for signs of pain and distress and additional analgesics administered as needed.
H1-GFP Cell Line Generation
CRISPR-Cas9 was used for introduction of the green fluorescent protein (GFP) sequence to the AAVS1 safe-harbor site in commercially available H1 human embryonic stem cells (hESCs, WiCell Research Institute, Inc.) using modified previously published reagents43. Briefly, single-stranded donor oligonucleotides (ssODNs) encoding the validated guide RNA sequence (5’-GGGGCCACTAGGGACAGGAT-3’) for targeting the AAVS1 locus were annealed and subcloned into PX458M-HF, a modified version of pSpCas9(BB)-2A-GFP (PX458) (Addgene #48138) generated by the CCHMC Transgenic Core, which carries an optimized single guide RNA using TransIT-LT1 according to the manufacturer’s recommendations (Mirus Bio)44,45. Transfection was performed in mTeSR1 media containing 10μM Y27632 on hESC-qualified Matrigel-coated plates (Corning). Four hours post-transfection, media was removed and replaced with mouse embryonic fibroblast (MEF) conditioned hESC media (DMEM/F12, 20% KOSR, 0.1mM nonessential amino acids, 2 mM L-glutamine, 0.1 mM β-Mercaptoethanol and 4 ng/mL bFGF) containing 10μM Y-27632 with daily media changes with MEF-conditioned hESC media. At 2 days post-transfection, a single-cell suspension of cells was generated with Accutase (STEMCELL Technologies) and replated at an approximate density of 10,000 cells/cm2 for Geneticin (G418) selection. Beginning three days post-transfection, G418 selection (100 μg/mL) was performed for eight days, after which daily feeds were performed using mTeSR1. After two weeks, the remaining G418-resistant colonies were harvested using Accutase and plated at cloning density in mTeSR with CloneR supplement (STEMCELL Technologies). Recovered clones were manually excised, expanded in mTeSR1 media, and subjected to genotyping. Correctly targeted insertion of the 2A-NeoR-CAG-GFP cassette in the AAVS1 locus was confirmed by PCR, sanger sequencing, copy-number analysis and GFP expression. This cell line was used in all experiments. Monthly mycoplasma testing was performed using the MycoAlert Plus Detection Kit and Control Set (Lonza #LT07–705 & LT07–518) on all cell cultures and results were consistently negative.
METHOD DETAILS
Generation of Human Intestinal Organoids
HIOs were generated and maintained as previously described 19,20,46. Briefly, H1-GFP cells (CCHMC Pluripotent Stem Cell Core Facility) were grown in feeder-free conditions in Matrigel (BD Biosciences) coated six-well Nunclon surface plates (Nunc) and maintained in mTESR1 media (STEMCELL Technologies). For definitive endoderm (DE) induction, cells were passaged as single cell suspensions generated with Accutase (STEMCELL Technologies) and plated in 24-well Nunc plates at a density of approximately 100,000 cells/well. Cells grew in mTESR1 media for two days before treatment with 100 ng/ml of Activin A for three days. DE was then treated with hindgut induction medium (RPMI 1640, 100x NEAA, 2% dFCS,) for four days with 100 ng/ml FGF4 (R&D) and 3 μM Chiron 99021 (Tocris) to induce mid-hindgut spheroids. Spheroids were then plated in Growth Factor Reduced (GFR) Matrigel and maintained in intestinal growth medium (Advanced DMEM/F-12, N2 supplement, B27 supplement, 15 mM HEPES, 2 mM L-glutamine, penicillin-streptomycin) supplemented with 100 ng/ml EGF (R&D) to generate HIOs. Media was changed twice weekly and HIOs were replated in fresh Matrigel on day 14. HIOs were utilized for surgical transplantation between days 28 and 34.
Generation of Enteroids and txp-Enteroids
Enteroids were generated from human surgical samples and transplanted H1 GFP HIOs for ease of use and downstream analysis. Crypts were isolated from transplanted HIOs as previously described 47. Briefly, segments of HIO tissue were pinned down in a SYLGARD 184 (Dow) coated petri dish, gently scraped to remove villi, washed with 2 mM chelation buffer before a 30 minute incubation in 2 mM chelation buffer. Then, to release the crypts the tissue was again gently scraped. The chelation buffer containing the crypts was removed from the petri dish, filtered through a 150 μm nylon mesh, and spun down at 50 g for 5 min at 4°C to pellet the crypts for use in cell culture. Crypts were plated in Matrigel (Corning) and IntestiCult media (STEMCELL Technologies) was used to generate enteroids. Media was changed twice weekly and passages occurred every 7 to 10 days. txp-Enteroids were used after a minimum of 3 passages to minimize mesenchymal contamination.
Generation of iv-Enteroids
Matrigel bubbles containing singular HIOs were removed from plates and pooled within a 15 mL tube. 500 μL of Cell Recovery Solution (Corning) per HIO was added. Then, incubations took place for 30 min at 4°C with agitation. Tubes were then centrifuged at 300g for 5 min. Supernatant was removed and the HIOs were resuspended in 5 mL of fresh Cell Recovery Solution. Then, using a serological pipette, dissociation was performed by pipetting up and down. Dissociation was achieved when epithelial structures were free from mesenchyme. As a final step, the mixture was filtered through a 40 μm mesh, discarding the flow through, and thoroughly washing with PBS. iv-Enteroids were plated in Matrigel (Corning) and IntestiCult media (STEMCELL Technologies) was used. Media was changed twice weekly and passages occurred every 7 to 10 days. iv-Enteroids were used after a minimum of 3 passages to minimize mesenchymal contamination.
Mucosectomy Surgical Procedure
The mucosectomy procedure was optimized for our purposes from a previously published study 5.
Loop Creation
One day prior to the procedure, chow diet was removed, and rats were placed on GelDiet 76A (ClearH2O), to be continued seven days postoperatively before returning to chow. Rats were anesthetized with 2% inhaled isoflurane (Butler Schein), and their abdomen shaved and prepped in sterile fashion using swabs coated with isopropyl alcohol and povidine-iodine. A midline laparotomy of approximately 3 cm was made. A single dose of piperacillin and tazobactam (100 mg/kg) was administered within the abdominal cavity using an 18G blunt tip fill needle affixed to a 5 mL syringe. Then, the cecum was identified, and the intestines eviscerated, using saline warmed to 37C to maintain tissue moisture throughout the procedure. A suitable stretch of bowel (~2–3 cm) is first identified for blind loop creation. To create the loop, the bowel and mesentery at the proximal end of what will become the loop was transected using scissors and a Bovie pen, as necessary. Then, the distal end of the blind loop was partially transected using scissors. An anastomosis was performed using 7–0 silk suture (PERMA-HAND; Ethicon) between the open end of the fully transected bowel and the partially transected distal end of the blind loop in a simple interrupted fashion. This establishes full continuity for the rat and maintains the blind loops’ distal connection.
Injury Creation
A bulldog clamp is applied to the distal end of the blind loop, blocking flow into the continuous bowel. First, the chemical damage is induced. Using a 20 mL syringe equipped with a cannula, the loop was flushed with saline warmed to 37 C for 2 min. Then, the loop was flushed with 1 mM dithiothreitol (DTT) in saline warmed to 37 C for 2 min using a 20 mL syringe equipped with a cannula. The loop was subsequently flushed with warm saline again before flushing with 5 mM isotonic ethylenediamine tetra-acetic acid (EDTA) buffer warmed to 37 C for 10 min using a 20 mL syringe equipped with a cannula. Approximately 50 mL of EDTA solution was used for flushing over the 10 min period. This series of flushes was repeated and then followed by a final flush with saline warmed to 37 C. To induce the mechanical damage, a dental go-between style brush flosser of appropriate diameter was inserted and removed from the loop, slightly twisting during entry and exit three times. After injury creation, the bulldog clamp was removed, and the distal end of the blind loop was tied off using 4–0 absorbable ChromicGut suture (ETHILON; Ethicon).
Reseeding the Loop
Loops were reseeded with fragmented HIOs or media void of cellular content. To fragment structures, they were collected with their media and pooled. Then, they were drawn into a syringe affixed with an 18G blunt tip fill needle, then the needle was exchanged for a 20G needle and the contents evacuated from the syringe into the well of a 24-well plate. This process was repeated using sequentially smaller needles ending at 25G, doing three syringe evacuations per needle size (Sup. Figure 7a). Then, the fragments were drawn into a 1 mL syringe equipped with a canula and deposited within the loop. Approximately 100,000 cells per 4 mm of bowel length was used for reseeding with fragmented HIO (Sup. Figure 7b-c). After reseeding, the proximal end of the loop was closed using 5–0 silk suture (PERMA-HAND; Ethicon) as the canula was removed. The bowel was then carefully replaced within the abdominal cavity. To close the incision, the muscle was sutured in a running fashion using 4–0 coated absorbable suture (VICRYL RAPIDE; Ethicon). Then, the skin was closed in a buried interrupted fashion again using 4–0 coated absorbable suture (VICRYL RAPIDE; Ethicon). A single dose of carprofen (5 mg/kg) was administered for pain management at the end of the procedure. Tissues were harvested seven days postoperatively as an early time point during the healing process, and ten weeks postoperatively as a long term, recovered time point.
In vitro HIO Dissociation and Cell Quantification
Nuclei were isolated from HIOs and txp-enteroids using the Minute Detergent Free Nuclei Isolation Kit (Invent Biotechnologies, Inc.) following manufacturer guidelines. To have sufficient cell quantities, structures were pooled for isolation. Four d28 HIOs were pooled for single nuclear isolations. Immediately following completion, nuclei were automatically quantified using a TC20 (Bio-Rad Laboratories, Inc.). The total count was then divided by four to determine the number of cells per HIO.
In vitro Enteroid Dissociation and Flow Cytometry
All three enteroid types were dissociated into single-cell suspensions using Accutase (Stemcell Technologies Cat # 07920) according to the manufacturer’s instructions. The single cells were first stained with fixable viability dye eFluor 780 (eBioscience, Cat # 65–0865-14) to assess viability. Following this, cells were fixed and permeabilized using fixation and permeabilization reagents (Invitrogen, Cat # 00-5521-00). Subsequently, the cells were stained with anti-human EpCAM AF647 (Biolegend, Cat # 324212) and anti-MKI67 PeCF594 (BD Biosciences, Cat # 567120) antibodies. After staining, the cells were washed and resuspended in flow buffer. The prepared cell suspensions were analyzed using a BD FACS Canto™ III flow cytometer (BD Biosciences). Data acquisition and analysis were performed using Facs DIVA (BD Biosciences).
Scanning Electron Microscopy
Segments of healthy rat distal small bowel and freshly damaged rat distal small bowel were fixed overnight in 3% glutaraldehyde in 0.175 M sodium cacodylate buffer of pH 7.4. Samples were then buffer rinsed and post fixed in 1% osmium tetroxide in 0.175 M cacodylate buffer for 1 hour at 4°C. After another buffer rinse, samples were put through a graded ethanol series (25%, 50%, 75%, 95%, 3× 100%) for dehydration. Specimens were then critical point dried in an EM CPD300 (Leica), stub mounted and sputter coated 10 nm thick with 60/40 gold palladium using an EM ACE600 (Leica). An SU8010 transmission electron microscope (Hitachi) was used to image samples.
Tissue Processing and Immunostaining
Samples were harvested and fixed overnight in 4% paraformaldehyde (PFA), processed and embedded in paraffin blocks. Sections were deparaffinized and either stained immediately with hematoxylin and eosin or subjected to antigen retrieval, and antibody stained. Antibody incubations took place at 4°C overnight in 1% bovine serum albumin in phosphate buffered saline (PBS). Antibodies used are listed in the Key Resources section. The Vectastain ABC system was used for amplification and the diaminobenzidine substrate kit was used for signal detection (Vector Laboratories). Mucins were demonstrated using 1% Alcian Blue pH 1 (Newcomer Supply, Inc.). Lillie-Mayer’s Hematoxylin (Agilent Technologies) or 1% Nuclear Fast Red (Polysciences, Inc.) was used as a counterstain. For biodistribution, serial sections were made and every tenth slide was stained over 2 mm of tissue thickness.
Key resources table
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| ACTA2; Actin Alpha 2, Smooth Muscle | Abcam | AB_262054 |
| CDH1; Cadherin 1 | BD Biosciences | AB_397581 |
| CDH1; Cadherin 1 | R&D | AB_355504 |
| CDX2; Caudal Type Homeobox 2 | Cell Marque | AB_1516799 |
| CHGA; Chromogranin A | DSHB | AB_1553436 |
| DEFA5; Defensin Alpha 5 | Abcam | AB_2049572 |
| DPPIV, Dipeptidyl peptidase-4 | R&D | AB_354651 |
| F3; Coagulation Factor III, Tissue Factor | Atlas Antibodies | AB_2680701 |
| GATA4; GATA Binding Protein 4 | Santa Cruz | AB_627667 |
| GFP; Green Fluorescent Protein | Abcam | AB_300798 |
| KU80; Ku Autoantigen, 80kDa | Cell Signaling | AB_2218736 |
| MKI67; Marker of Proliferation Ki-67 | Thermo | AB_2341197 |
| MS4A12; Membrane Spanning 4-Domains A12 | Atlas Antibodies | AB_2683491 |
| MUC2; Mucin 2 | Abcam | AB_2889298 |
| MUC5B; Mucin 5B, Oligomeric Mucus/Gel-Forming | Atlas Antibodies | AB_1854203 |
| OLFM4; Olfactomedin 4 | Cell Signaling | AB_2798465 |
| SATB2; SATB Homeobox 2 | Millipore Sigma | Cat# 384R-14 |
| SI; Sucrase-Isomaltase | Atlas Antibodies | AB_1856840 |
| TUBB3; Tubulin Beta 3 Class III | Abcam | AB_727049 |
| VWF; Von Willebrand Factor | Dako | AB_2315602 |
| α-chicken biotin | Vector Labs | AB_2336114 |
| α-goat biotin | Vector Labs | AB_2336123 |
| α-mouse biotin | Vector Labs | AB_2313581 |
| α-rabbit biotin | Vector Labs | AB_2313606 |
| α-chicken Alexa Fluor 647 | Invitrogen | AB_1500594 |
| α-goat Alexa Fluor 647 | Invitrogen | AB_141844 |
| α-mouse Alexa Fluor 488 | Invitrogen | AB_2536161 |
| α-mouse Alexa Fluor 568 | Invitrogen | AB_2534013 |
| α-rabbit Alexa Fluor 568 | Invitrogen | AB_2534017 |
| α-rabbit Alexa Fluor 647 | Invitrogen | AB_2536183 |
| PE-MKI67 | BD Biosciences | AB_2916453 |
| AF647-EPCAM | Biolegend | AB_756086 |
| Bacterial and virus strains | ||
| Biological samples | ||
| Human DNA | SIGMA | 69237-3 |
| Chemicals, peptides, and recombinant proteins | ||
| 1x DMEM | Invitrogen | Cat# 11965-084 |
| 3-isobutyl-1-methylxanthine | Sigma | Cat# I5879 |
| 50x B27 supplement | Thermo Fisher Scientific | Cat# 17504044 |
| 50x B27 supplement w/o Vitamin A | Thermo Fisher Scientific | Cat# 12587-010 |
| A8301 | Torcis | Cat# A83-01 |
| Accutase | Thermo Fisher Scientific | Cat# A11105-01 |
| Activin A | Cell Guidance Systems | Cat# GFH6 |
| Advanced DMEM:F12 | Thermo Fisher Scientfic | Cat# 12634-010 |
| Alcian Blue Stain 1%, pH 1.0 Aqueous | Newcomer Supply, Inc. | Cat# 1005B |
| Atropine sulfate salt monohydrate | Sigma-Aldrich | Cat# A0257 |
| Bethanechol | Sigma-Aldrich | Cat# 1071009 |
| bumetanide | Apexbio | Cat# A1855 |
| C59 | Cellagen Technology | Cat# C7641-2s |
| Cell Recovery Solution | Corning | Cat# 354253 |
| CHIR99021 | ReproCell | Cat# 04-0004-10 |
| Collagen Type IV | Thermo Fisher Scientific | Cat# 17104019 |
| Defined fetal bovine serum (dFBS) | Hyclone | Cat# SH30070.02 |
| Dispase | Thermo Fisher Scientific | Cat# 17105-041 |
| DMEM/F12 | Thermo Fisher Scientific | Cat# 11320033 |
| DMEM/F-12, GlutaMAX | Thermo Fisher Scientific | Cat# 10565018 |
| EGF | Sigma-Aldrich | Cat# 1257-0.1MG |
| F12 Media | Invitrogen | Cat# 11765-054 |
| Fetal Bovine Serum (FBS) | HyClone | Cat# SH30071.03 |
| Fibronectin | Corning | Cat# 354008 |
| Fixable Viability Dye eFluor™ 780 | eBioscience | Cat# 65-0865-14 |
| Forskolin | Cell Signaling | Cat# 3828S |
| Geltrex | Thermo Fisher Scientific | Cat# A1413201 |
| HEPES Buffer | Thermo Fisher Scientific | Cat# 15630080 |
| hESC-qualified Matrigel | BD Biosciences | Cat# 354277 |
| Intesticult Organoid Growth Medium (Human) | Stemcell Technologies | Cat# 06010 |
| Krebs-Henseleit Buffer Modified | Millipore Sigma | Cat# K3753 |
| L-glutamine (100x) | Thermo Fisher Scientific | Cat# 25030-081 |
| Matrigel Matrix Basement Membrane | BD Biosciences | Cat# 354234 |
| mTeSR1 media | Stem Cell Technologies | Cat# 5850 |
| N2 Supplement | Thermo Fisher Scientific | Cat# 17502-048 |
| Neurobasal Medium | Thermo Fisher Scientific | Cat# 21103049 |
| Non-essential Amino Acids (100x) | Thermo Fisher Scientific | Cat# 11140050 |
| Normal donkey serum | Jackson Immunoresearch Labs | Cat# 017-000-121 |
| Normal goat serum | Jackson Immunoresearch Labs | Cat# 005-000-121 |
| Nuclear Fast Red, 1% Aqueous | Polysciences, Inc. | Cat# 24199C-250 |
| PD0325901 | Stem Cell Technologies | Cat# 72184 |
| Pen/Strep (100x) | Thermo Fisher Scientific | Cat# 15140-122 |
| PIK90 | EMD Millipore | Cat# 528117 |
| PMA | Torcis | Cat# 1201 |
| Proteinase K | Bioline | Cat# BIO37084 |
| recombinant human BMP4 | R&D Systems | Cat# 314-BP |
| recombinant human EGF | R&D Systems | Cat# 236-EGF |
| recombinant human FGF10 | R&D Systems | Cat# 345-FG |
| recombinant human FGF2 | R&D Systems | Cat# 233-FB |
| recombinant human FGF4 | R&D Systems | Cat# 235-F4 |
| recombinant human Noggin | R&D Systems | Cat# 6057-NG |
| recombinant human WNT3A | R&D Systems | Cat# 5036-WN |
| Thiazovivin | Tocris | Cat# 3845 |
| Y-27632 dihydrochloride | Tocris | Cat# 1254 |
| Critical commercial assays | ||
| TaqMan™ Universal PCR Master Mix | Applied Biosystem | Cat# 4304437 |
| ISOLATE II Genomic DNA Kit | Bioline | Cat# BIO-52067 |
| Transcription Factor Fixation/Permeabilization Concentrate and Diluent | eBioscience | Cat# 00-5521-00 |
| Experimental models: Cell lines | ||
| H1 GFP | In House | N/A |
| Experimental models: Organisms/strains | ||
| RRG Rats | In House | N/A |
| Oligonucleotides | ||
| hAlu_F: TGGTGGCTCTCTCCTGTAAT | Integrated DNA Technologies | N/A |
| hAlu_R: GATCTCGGCTCACTGCAAC | Integrated DNATechnologies | N/A |
| hAlu_Probe: TGAGGCAGGAGAATCGCTTGAACC | Integrated DNA Technologies | N/A |
| Taqman Probes | ||
| CDX2; Caudal Type Homeobox 2 | Thermo Fisher Scientific | Cat# Hs01078080_m1 |
| CHGA; Chromogranin A | Thermo Fisher Scientific | Cat# Hs00900375_m1 |
| LGR5; Leucine-rich repeat-containing G-protein coupled receptor 5 | Thermo Fisher Scientific | Cat# Hs00173664_m1 |
| LYZ; Lysozyme | Thermo Fisher Scientific | Cat# Hs00426232_m1 |
| MKI67; Marker of Proliferation Ki-67 | Thermo Fisher Scientific | Cat# Hs04260396_g1 |
| MUC2; Mucin 2 | Thermo Fisher Scientific | Cat# Hs03005103_g1 |
| OLFM4; Olfactomedin 4 | Thermo Fisher Scientific | Cat# Hs00197437_m1 |
| VIM; Vimentin | Thermo Fisher Scientific | Cat# Hs00185584_m1 |
| Software and algorithms | ||
| BioRender | BioRender | SCR_018361 |
| ImageJ | NIH | SCR_003070 |
| FACSDiva | BD Biosciences | SCR_001456 |
| NIS-Elements | Nikon | SCR_014329 |
| Prism9 | GraphPad | SCR_002798 |
| Other | ||
| Ussing Chamber System | Physiologic Instruments | EM-8-C |
Image Acquisition
Surgical imagery was acquired using an M80 microscope outfitted with a MC 170HD camera (Leica Microsystems). Gross images of harvested structures were acquired using a V60 ThinQ (LG Electronics, Inc.). Harvests were performed using a M165 FC microscope outfitted with a DCF7000 T camera (Leica Microsystems). Slides were imaged using an Eclipse Ti microscope (Nikon Corporation) an Eclipse Ti2 microscope (Nikon Corporation). Whole mount staining was imaged using an Eclipse Ti1 A1R confocal microscope (Nikon Corporation). Subsequent analysis performed using Nikon Elements Imaging Software, including the Object Count function for cell quantification, (Nikon Corporation) and ImageJ software (NIH) for surface area quantification and measurements.
Ex vivo Epithelial Characteristics and Permeability
The epitheliums of freshly harvested healthy distal small bowel and reseeded loops were carefully dissected through a technique similar to seromuscular stripping as previously reported 25,31,48. Samples were opened and dissection was done in ice cold Kreb’s buffer (NaCl, 117 mM; KCl, 4.7 mM; MgCl2, 1.2mM; NaH2PO4, 1.2 mM; NaHCO3, 25 mM; CaCl2, 2.5 mM and glucose, 11 mM). Full thickness tissue segments were then pinned in a dish containing cured Sylgard (Electron Microscopy Sciences). Reseeded loops were verified to be GFP positive before further dissection and use in the Ussing chamber assay. The seromusculature was then micro-dissected from the epithelium using Dumont #5 and #7 forceps along with Vannas scissors (Fine Science Tools, Inc.). Gross tissue integrity was assessed using the stereoscope’s bottom lighting for uniformity in appearance and any damaged areas were removed. Some remnant subepithelial mucosa remained after dissection, as previously described 31. The central portion of the epithelium, with the least amount of handling, was then positioned for mounting between the hemi-chambers of an Ussing apparatus (Physiologic Instruments). 0.031 cm2 of tissue was exposed to 5 mL of oxygenated Krebs buffer at 37 °C throughout the assay. The transepithelial potential difference was detected with two paired electrodes affixed to a salt bridge containing 3.75% agar in 3 M KCl. The electrodes were connected to a VVC MC8 voltage clamp amplifier (Physiologic Instruments, San Diego). Electrode potential difference and fluid resistance values were offset to zero immediately before sliders were mounted between the chambers. A 30 min period was allowed for the establishment of equilibrium. Then, tissues were voltage-clamped at 0 mV while continuously measuring the short circuit current (Isc) and chemical stimuli applied (10 μM Forskolin, 100 μM IBMX, and 100 μM Bumetanide). For FITC-dextran permeability, 2.2 mg/ml FITC-dextran was added into apical side, and a sample was taken from the basolateral side every 30 minutes for 3 hours, replacing the same amount of fresh modified Kreb’s buffer in the basolateral side to maintain pressure across the sample. Once all aqueous samples were collected, they were quantified with a plate-reader (Synergy 2, BioTek).
Genomic DNA isolation
Samples were harvested 10 weeks post-operatively from reseeded mucosectomy rats (brain, colon, heart, kidney, liver, lung, and small intestine) and flash frozen in liquid nitrogen. As a positive control, H1 GFP cells were also used. Samples were homogenized, filtered, and digested using an ISOLATE II Genomic DNA kit (Meridian Biosciences) according to manufacturer guidelines. After the gDNA was isolated, quality and quantity was analyzed using a NanoDrop Microvolume Spectrophotometer.
Human Specific Alu PCR
Primers and Probes
Detection of human DNA was done using previously described primers and probe 36. Briefly, Alu PCR was performed on gDNA extracted from various organs from reseeded mucosectomy rats and the H1 GFP cell line. The forward primer was designed to anneal upstream of the human specific Alu sequence (5′-TGGTGG CTCTCT CCT GTA AT-3′) and the reverse primer was designed to primarily anneal within the human-specific Alu sequence (5′-GAT CTC GGC TCA CTG CAA C-3′), resulting in a 96 base pair amplicon. The probe was designed to bind between the two primers (5′- TGA GGC AGG AGA ATC GCT TGA ACC-3′) quencher-MGB-6FAM upstream of the hAlu-Fspecific sequence. The primers and probes were custom ordered from Integrated DNA Technologies.
Alu PCR
Quantitative real time PCR was performed using TaqMan Universal PCR Master Mix (Applied Biosystems) on 200 ng of target template gDNA. Each sample was sequenced in triplicate using a OneStep thermocycler (Applied Biosystems). Standard curves were generated by adding 10-fold serial dilutions (200 ng– 0 ng) of hDNA (Millipore Sigma) and H1 GFP cells on each PCR plate. QunatoStudio software (Applied Biosystems) was used to calculate crossing threshold (Ct) values for presence of human cells based on the standard values.
QUANTIFICATION AND STATISTICAL ANALYSIS
For bar graphs, data is represented as a mean ± standard deviation, with all individual data points represented. For violin plots, a dashed line indicates the mean and interquartile ranges, with all individual data points represented. For statistics comparing three groups of data Kruskal-Wallis tests were performed. For statistics comparing unpaired data a student t-test was performed. The statistical significance cutoff was p < 0.05 and confidence 95%.
Supplementary Material
HIGHLIGHTS.
hPSC derived organoids repair damaged bowel in a preclinical animal model
Multi-lineage human organoids engraft more broadly than epithelial-only organoids
Multi-lineage organoids engraft long-term to form mesenchymal and epithelial layers
Functional neo-epithelia is present after intestinal injury repair
ACKNOWLEDGEMENTS
The authors thank Riccardo Barrile and Noah Shroyer for valuable feedback, Tanvi Sumit for assistance with histology, and Leyla Esfandiari for proofreading an early manuscript draft. Research reported in this publication was supported by NIDDK and NIAID of the NIH under grant number U01DK103117 (MAH, JMW). This work was also funded in part by NIH grant number P30DK078292, Cincinnati Digestive Health Center Award (IPRF, PCSF). Additional financial support was provided by an NSF GRFP (HMP) and a generous donation from the Farmer Family Foundation. The graphical abstract was created with BioRender.com.
Footnotes
DECLARATION OF INTERESTS
CCHMC has a patent application in process related to the work in this study.
INCLUSION AND DIVERSITY STATEMENT
We support inclusive, diverse, and equitable conduct of research.
SUPPLEMENTAL INFORMATION
Document S1 including Figures S1-S4 and Tables S1-S2.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
REFERENCES
- 1.Singh A, Poling HM, Spence JR, Wells JM, and Helmrath MA (2020). Gastrointestinal organoids: a next-generation tool for modeling human development. Am J Physiol Gastrointest Liver Physiol 319, G375–G381. 10.1152/ajpgi.00199.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sato T, and Clevers H. (2013). Growing self-organizing mini-guts from a single intestinal stem cell: mechanism and applications. Science 340, 1190–1194. 10.1126/science.1234852. [DOI] [PubMed] [Google Scholar]
- 3.Yui S, Nakamura T, Sato T, Nemoto Y, Mizutani T, Zheng X, Ichinose S, Nagaishi T, Okamoto R, Tsuchiya K, et al. (2012). Functional engraftment of colon epithelium expanded in vitro from a single adult Lgr5(+) stem cell. Nat Med 18, 618–623. 10.1038/nm.2695. [DOI] [PubMed] [Google Scholar]
- 4.Watanabe S, Kobayashi S, Ogasawara N, Okamoto R, Nakamura T, Watanabe M, Jensen KB, and Yui S. (2022). Transplantation of intestinal organoids into a mouse model of colitis. Nat Protoc 17, 649–671. 10.1038/s41596-021-00658-3. [DOI] [PubMed] [Google Scholar]
- 5.Avansino JR, Chen DC, Hoagland VD, Woolman JD, and Stelzner M. (2006). Orthotopic transplantation of intestinal mucosal organoids in rodents. Surgery 140, 423–434. 10.1016/j.surg.2006.03.012. [DOI] [PubMed] [Google Scholar]
- 6.Nakamura T, and Watanabe M. (2017). Intestinal stem cell transplantation. J Gastroenterol 52, 151–157. 10.1007/s00535-016-1288-8. [DOI] [PubMed] [Google Scholar]
- 7.Fukuda M, Mizutani T, Mochizuki W, Matsumoto T, Nozaki K, Sakamaki Y, Ichinose S, Okada Y, Tanaka T, Watanabe M, and Nakamura T. (2014). Small intestinal stem cell identity is maintained with functional Paneth cells in heterotopically grafted epithelium onto the colon. Genes Dev 28, 1752–1757. 10.1101/gad.245233.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fordham RP, Yui S, Hannan NR, Soendergaard C, Madgwick A, Schweiger PJ, Nielsen OH, Vallier L, Pedersen RA, Nakamura T, et al. (2013). Transplantation of expanded fetal intestinal progenitors contributes to colon regeneration after injury. Cell Stem Cell 13, 734–744. 10.1016/j.stem.2013.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sugimoto S, Ohta Y, Fujii M, Matano M, Shimokawa M, Nanki K, Date S, Nishikori S, Nakazato Y, Nakamura T, et al. (2018). Reconstruction of the Human Colon Epithelium In Vivo. Cell Stem Cell 22, 171–176 e175. 10.1016/j.stem.2017.11.012. [DOI] [PubMed] [Google Scholar]
- 10.Sugimoto S, Kobayashi E, Fujii M, Ohta Y, Arai K, Matano M, Ishikawa K, Miyamoto K, Toshimitsu K, Takahashi S, et al. (2021). An organoid-based organ-repurposing approach to treat short bowel syndrome. Nature 592, 99–104. 10.1038/s41586-021-03247-2. [DOI] [PubMed] [Google Scholar]
- 11.Khalil HA, Hong SN, Rouch JD, Scott A, Cho Y, Wang J, Lewis MS, Martin MG, Dunn JCY, and Stelzner MG (2019). Intestinal epithelial replacement by transplantation of cultured murine and human cells into the small intestine. PLoS One 14, e0216326. 10.1371/journal.pone.0216326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Agopian VG, Chen DC, Avansino JR, and Stelzner M. (2009). Intestinal stem cell organoid transplantation generates neomucosa in dogs. J Gastrointest Surg 13, 971–982. 10.1007/s11605-009-0806-x. [DOI] [PubMed] [Google Scholar]
- 13.Lau JY, Sung J, Hill C, Henderson C, Howden CW, and Metz DC (2011). Systematic review of the epidemiology of complicated peptic ulcer disease: incidence, recurrence, risk factors and mortality. Digestion 84, 102–113. 10.1159/000323958. [DOI] [PubMed] [Google Scholar]
- 14.Mammen JM, and Matthews JB (2003). Mucosal repair in the gastrointestinal tract. Crit Care Med 31, S532–537. 10.1097/01.CCM.0000081429.89277.AF. [DOI] [PubMed] [Google Scholar]
- 15.Podolsky DK (1997). Healing the epithelium: solving the problem from two sides. J Gastroenterol 32, 122–126. 10.1007/BF01213309. [DOI] [PubMed] [Google Scholar]
- 16.Powell DW, Mifflin RC, Valentich JD, Crowe SE, Saada JI, and West AB (1999). Myofibroblasts. II. Intestinal subepithelial myofibroblasts. Am J Physiol 277, C183–201. 10.1152/ajpcell.1999.277.2.C183. [DOI] [PubMed] [Google Scholar]
- 17.Kamada T, Satoh K, Itoh T, Ito M, Iwamoto J, Okimoto T, Kanno T, Sugimoto M, Chiba T, Nomura S, et al. (2021). Evidence-based clinical practice guidelines for peptic ulcer disease 2020. J Gastroenterol 56, 303–322. 10.1007/s00535-021-01769-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Elmentaite R, Ross ADB, Roberts K, James KR, Ortmann D, Gomes T, Nayak K, Tuck L, Pritchard S, Bayraktar OA, et al. (2020). Single-Cell Sequencing of Developing Human Gut Reveals Transcriptional Links to Childhood Crohn’s Disease. Dev Cell 55, 771–783 e775. 10.1016/j.devcel.2020.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Spence JR, Mayhew CN, Rankin SA, Kuhar MF, Vallance JE, Tolle K, Hoskins EE, Kalinichenko VV, Wells SI, Zorn AM, et al. (2011). Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature 470, 105–109. 10.1038/nature09691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Watson CL, Mahe MM, Munera J, Howell JC, Sundaram N, Poling HM, Schweitzer JI, Vallance JE, Mayhew CN, Sun Y, et al. (2014). An in vivo model of human small intestine using pluripotent stem cells. Nat Med 20, 13101314. 10.1038/nm.3737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Menoret S, Ouisse LH, Tesson L, Delbos F, Garnier D, Remy S, Usal C, Concordet JP, Giovannangeli C, Chenouard V, et al. (2018). Generation of Immunodeficient Rats With Rag1 and Il2rg Gene Deletions and Human Tissue Grafting Models. Transplantation 102, 1271–1278. 10.1097/TP.0000000000002251. [DOI] [PubMed] [Google Scholar]
- 22.Tait IS, Evans GS, Flint N, and Campbell FC (1994). Colonic mucosal replacement by syngeneic small intestinal stem cell transplantation. American journal of surgery 167, 67–72. 10.1016/0002-9610(94)90055-8. [DOI] [PubMed] [Google Scholar]
- 23.Middendorp S, Schneeberger K, Wiegerinck CL, Mokry M, Akkerman RD, van Wijngaarden S, Clevers H, and Nieuwenhuis EE (2014). Adult stem cells in the small intestine are intrinsically programmed with their location-specific function. Stem Cells 32, 1083–1091. 10.1002/stem.1655. [DOI] [PubMed] [Google Scholar]
- 24.Thompson CA, Wojta K, Pulakanti K, Rao S, Dawson P, and Battle MA (2017). GATA4 Is Sufficient to Establish Jejunal Versus Ileal Identity in the Small Intestine. Cell Mol Gastroenterol Hepatol 3, 422–446. 10.1016/j.jcmgh.2016.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Clarke LL (2009). A guide to Ussing chamber studies of mouse intestine. Am J Physiol Gastrointest Liver Physiol 296, G1151–1166. 10.1152/ajpgi.90649.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ma TY, Hollander D, Bhalla D, Nguyen H, and Krugliak P. (1992). IEC-18, a nontransformed small intestinal cell line for studying epithelial permeability. J Lab Clin Med 120, 329–341. [PubMed] [Google Scholar]
- 27.Srinivasan B, Kolli AR, Esch MB, Abaci HE, Shuler ML, and Hickman JJ (2015). TEER measurement techniques for in vitro barrier model systems. J Lab Autom 20, 107–126. 10.1177/2211068214561025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Finkbeiner SR, Hill DR, Altheim CH, Dedhia PH, Taylor MJ, Tsai YH, Chin AM, Mahe MM, Watson CL, Freeman JJ, et al. (2015). Transcriptome-wide Analysis Reveals Hallmarks of Human Intestine Development and Maturation In Vitro and In Vivo. Stem Cell Reports. 10.1016/j.stemcr.2015.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Workman MJ, Mahe MM, Trisno S, Poling HM, Watson CL, Sundaram N, Chang CF, Schiesser J, Aubert P, Stanley EG, et al. (2017). Engineered human pluripotent-stem-cell-derived intestinal tissues with a functional enteric nervous system. Nat Med 23, 49–59. 10.1038/nm.4233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Singh A, Poling HM, Sundaram N, Brown N, Wells JM, and Helmrath MA (2020). Evaluation of transplantation sites for human intestinal organoids. PLoS One 15, e0237885. 10.1371/journal.pone.0237885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Poling HM, Wu D, Brown N, Baker M, Hausfeld TA, Huynh N, Chaffron S, Dunn JCY, Hogan SP, Wells JM, et al. (2018). Mechanically induced development and maturation of human intestinal organoids in vivo. Nat Biomed Eng 2, 429–442. 10.1038/s41551-018-0243-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pradhan S, Karve SS, Weiss AA, Hawkins J, Poling HM, Helmrath MA, Wells JM, and McCauley HA (2020). Tissue Responses to Shiga Toxin in Human Intestinal Organoids. Cell Mol Gastroenterol Hepatol 10, 171–190. 10.1016/j.jcmgh.2020.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cortez AR, Poling HM, Brown NE, Singh A, Mahe MM, and Helmrath MA (2018). Transplantation of human intestinal organoids into the mouse mesentery: A more physiologic and anatomic engraftment site. Surgery 164, 643–650. 10.1016/j.surg.2018.04.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Munera JO, Sundaram N, Rankin SA, Hill D, Watson C, Mahe M, Vallance JE, Shroyer NF, Sinagoga KL, Zarzoso-Lacoste A, et al. (2017). Differentiation of Human Pluripotent Stem Cells into Colonic Organoids via Transient Activation of BMP Signaling. Cell Stem Cell 21, 51–64 e56. 10.1016/j.stem.2017.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Funakoshi K, Bagheri M, Zhou M, Suzuki R, Abe H, and Akashi H. (2017). Highly sensitive and specific Alu-based quantification of human cells among rodent cells. Sci Rep 7, 13202. 10.1038/s41598-017-13402-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Creane M, Howard L, O’Brien T, and Coleman CM (2017). Biodistribution and retention of locally administered human mesenchymal stromal cells: Quantitative polymerase chain reaction-based detection of human DNA in murine organs. Cytotherapy 19, 384–394. 10.1016/j.jcyt.2016.12.003. [DOI] [PubMed] [Google Scholar]
- 37.Sugimoto S, Kobayashi E, Fujii M, Ohta Y, Arai K, Matano M, Ishikawa K, Miyamoto K, Toshimitsu K, Takahashi S, et al. (2021). An organoid-based organ-repurposing approach to treat short bowel syndrome. Nature. 10.1038/s41586-021-03247-2. [DOI] [PubMed] [Google Scholar]
- 38.Rutherford D, and Ho GT (2023). Therapeutic Potential of Human Intestinal Organoids in Tissue Repair Approaches in Inflammatory Bowel Diseases. Inflamm Bowel Dis. 10.1093/ibd/izad044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kitano K, Schwartz DM, Zhou H, Gilpin SE, Wojtkiewicz GR, Ren X, Sommer CA, Capilla AV, Mathisen DJ, Goldstein AM, et al. (2017). Bioengineering of functional human induced pluripotent stem cell-derived intestinal grafts. Nat Commun 8, 765. 10.1038/s41467-017-00779-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tsai YH, Nattiv R, Dedhia PH, Nagy MS, Chin AM, Thomson M, Klein OD, and Spence JR (2017). In vitro patterning of pluripotent stem cell-derived intestine recapitulates in vivo human development. Development 144, 1045–1055. 10.1242/dev.138453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Eicher AK, Kechele DO, Sundaram N, Berns HM, Poling HM, Haines LE, Sanchez JG, Kishimoto K, Krishnamurthy M, Han L, et al. (2022). Functional human gastrointestinal organoids can be engineered from three primary germ layers derived separately from pluripotent stem cells. Cell Stem Cell 29, 36–51 e36. 10.1016/j.stem.2021.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Shacham-Silverberg V, and Wells JM (2020). Generation of esophageal organoids and organotypic raft cultures from human pluripotent stem cells. Methods Cell Biol 159, 1–22. 10.1016/bs.mcb.2020.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Oceguera-Yanez F, Kim SI, Matsumoto T, Tan GW, Xiang L, Hatani T, Kondo T, Ikeya M, Yoshida Y, Inoue H, and Woltjen K. (2016). Engineering the AAVS1 locus for consistent and scalable transgene expression in human iPSCs and their differentiated derivatives. Methods 101, 43–55. 10.1016/j.ymeth.2015.12.012. [DOI] [PubMed] [Google Scholar]
- 44.Ran FA, Hsu PD, Wright J, Agarwala V, Scott DA, and Zhang F. (2013). Genome engineering using the CRISPR-Cas9 system. Nature protocols 8, 2281–2308. 10.1038/nprot.2013.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Chen B, Gilbert LA, Cimini BA, Schnitzbauer J, Zhang W, Li GW, Park J, Blackburn EH, Weissman JS, Qi LS, and Huang B. (2013). Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas system. Cell 155, 1479–1491. 10.1016/j.cell.2013.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.McCracken KW, Howell JC, Wells JM, and Spence JR (2011). Generating human intestinal tissue from pluripotent stem cells in vitro. Nat Protoc 6, 1920–1928. 10.1038/nprot.2011.410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Mahe MM, Sundaram N, Watson CL, Shroyer NF, and Helmrath MA (2015). Establishment of human epithelial enteroids and colonoids from whole tissue and biopsy. J Vis Exp. 10.3791/52483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Giles DA, Moreno-Fernandez ME, Stankiewicz TE, Graspeuntner S, Cappelletti M, Wu D, Mukherjee R, Chan CC, Lawson MJ, Klarquist J, et al. (2017). Thermoneutral housing exacerbates nonalcoholic fatty liver disease in mice and allows for sex-independent disease modeling. Nat Med 23, 829–838. 10.1038/nm.4346. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data reported in this paper is available from the lead contact upon request.
This paper does not generate original code.
Any additional information required to reanalyze the data reported in this paper is available from the lead contact.