

Abstract
Objective(s):
The aim of this study is to investigate the possible role of the hippocampal BDNF-PI3K-AKT signaling pathway in the antidepressant-like activity of ellagic acid (EA) in mice.
Materials and Methods:
Male BALB/C mice were divided into 5 groups; vehicle (0.1 ml/day), sertraline (5mg/kg), EA (1 mg/kg), EA+BKM120 (PI3K inhibitor), EA+MK2206 (AKT inhibitor). EA, sertraline and vehicle were injected intraperitoneally for 14 days. Locomotor activity was determined by open field test. The tail suspension test was used to detect the antidepressant-like effect. After behavioral tests, hippocampal tissue was obtained and Western blot analyzes were performed for BDNF and pAKT1.
Results:
Sertraline and EA provided a reduction in immobility time in the tail suspension test when compared with the control group. BKM120 and MK2206 administration reversed this effect of EA. No statistical difference was found between groups in terms of locomotor activity. EA treatment caused an increase in hippocampal BDNF and pAKT1 levels in mice. While inhibitory agent administrations did not affect the increase of BDNF induced by EA, MK2206 administration reversed the increase in pAKT1 observed with EA.
Conclusion:
It has shown that EA has an antidepressant-like effect in mice without changing locomotor activity, and this effect may be mediated by the BDNF-PI3K-AKT signaling pathway.
Key Words: AKT, BDNF, Depression, Ellagic acid, PI3K
Introduction
Major depressive disorder (MDD) is a multifactorial psychiatric disorder that can be caused by genetic, social, and biochemical factors. MDD causes a burden. To eliminate this burden, procedures such as pharmacotherapy, psychotherapy, electroconvulsive therapy (ECT), deep brain stimulation, and lifestyle exercises are being used. Antidepressant medications are widely used to treat mood disorders. Antidepressants may cause various side effects (QT prolongation, gastrointestinal toxicity, anticholinergic effects, weight gain, sexual dysfunction, orthostatic hypotension, insomnia/agitation, drowsiness, etc.) (1). In clinic, patient compliance is disrupted due to these side effects and delayed onset and treatment resistance (2, 3). Understanding the pathophysiology of MDD enables us to generate new drugs. In recent years, herbal drugs have been investigated for the treatment of MDD.
Neurotrophins are growth factors that regulate neuronal growth, survival, and differentiation in the nervous system (4). Nerve growth factor and brain-derived neurotrophic factor (BDNF) are the most important members of the neurotrophin family (5). When BDNF binds to its receptor TrkB, three different signaling pathways are activated: Ras-PI3K-Akt, Ras-MAP kinase-Erk, and phospholipase Cγ (6). The PI3K/AKT pathway plays a role in cell survival, proliferation, cytoskeletal restructuring, and secretion events (7).
A meta-analysis conducted in 2017 found that antidepressant treatment for up to 12 weeks increased serum/plasma BDNF levels. (8). A recent meta-analysis showed that ECT application increased blood BDNF levels and significantly reduced depressive symptoms (9). Therefore, antidepressant drugs may provide beneficial effects through their interaction with BDNF.
Ellagic acid (EA) is a symmetric polyphenol. EA is found in strawberry, raspberry, blackberry, blueberry, goji berry, pomegranate, grape, walnut, chestnut, and a type of edible mushroom (Fistulina hepatica) in free form or as a complex derivative of ellagitannins (10-12). Huang et al. showed that EA reversed depression-like phenotypes and hippocampal damage caused by chronic unpredictable mild stress, and EA increased BDNF and serotonin levels (13). Our previous study found that EA has an antidepressant-like effect mediated by hippocampal BDNF increase (14). In this study, we aimed to determine the role of the BDNF-PI3K-AKT pathway among the possible mechanisms underlying the antidepressant-like effect of EA.
Materials and Methods
Male 2-3 month-old Albino mice (15-30 g) were used in the study. During the experiment, the animals were housed in their cages at a room temperature of 22-25 °C and exposed to 12 hours of light and 12 hours of darkness. The animals were kept in cages at a room temperature of 22-25 °C with a 12-hour light and 12-hour darkness cycle during the experiment.Animals were fed standard food and water. This study was carried out according to ethical rules by considering animal welfare. Akdeniz University Animal Experiments Local Ethics Committee approved the study protocol. (Protocol number:927/2019.07.04)
Experimental animals were divided into five groups (n=10 per group). Mice in the first group received 0.1 ml vehicle (10% DMSO); sertraline (5 mg/kg) was administered to the second group, and EA (1 mg/kg) was administered to the third group. All chemicals were given intraperitoneally once a day for 14 days. The fourth group (EA+BKM120) was given 14 days of EA, followed by BKM-120 (PI3K inhibitor) on the last day. In the fifth group (EA+MK2206), after 14 days of EA treatment, MK-2206 (AKT inhibitor) was given on the last day. After the last drug treatment (the 14th day), the animals were brought to the room where behavioral tests would be performed and waited for half an hour to acclimate to the room. Afterward, open field and tail suspension tests (TST) were applied. After the behavioral experiments, the animals were sacrificed, and their hippocampus was removed. Western blot analysis was performed to determine BDNF and AKT1 protein levels in the hippocampus.
Open Field Test: A square area made of black acrylic material 40 cm long * 40 cm high* 40 cm wide was divided into 16 equal parts. The mouse was placed in the middle of the apparatus, and a camera recorded its movements over five minutes as it moved around and explored its environment. After each mouse, the area was cleaned with 70% ethanol. The total number of squares crossed was calculated (15).
Tail Suspension Test: During the test, the mouse was suspended from its tail, and the duration of inactivity was recorded by camera for six minutes. The first two minutes of the experiment were considered the acclimation period, and the time the experimental animal remained immobile in the last four minutes was recorded in seconds (16).
Western blot: Mouse hippocampus tissues were dissected. Proteins were separated using 10% SDS-PAGE and transferred to polyvinylidene fluoride membranes. The membranes were incubated for 2 h with 5% skimmed milk or 5% BSA at room temperature and overnight at 4˚C with primary antibodies against BDNF (Elabscience, E-AB-18244), GAPDH (Elabscience, E-AB-40337), pAKT1 (Thr308) (Elabscience, E-AB-21082) and AKT1 (Elabscience, E-AB-63467) The next morning, the membrane was washed three times with TBST for ten minutes each. A secondary antibody (Anti-rabbit, Elabscience, E-AB-1003) suitable for the primary antibody was prepared at a ratio of 1:5,000 and incubated for one hour in a shaker at room temperature. After washing with washing solution, membranes were incubated with ECL for four minutes at room temperature. The bands were visualized using the bioanalytical imaging system (Azure Biosystems Model: c280), and the necessary calculations were made using the ImageJ software program.
Statistical analysis
The results were statistically analyzed using the Graph-Pad computer program, using one-way analysis of variance (One-way ANOVA) followed by Dunnett’s test. P-values below 0.05 were considered significant. All values are expressed as mean±standard error.
Results
Open field test results
The intervention groups (EA 1 mg/kg, sertraline, EA+BKM120, and EA+MK2206) showed no statistical difference in locomotor activity compared with the control group (Figure 1).
Figure 1.
Effects of EA, EA+BKM120, EA+MK2206, and sertraline treatment in open field test in mice
The mean difference is not statistically significant when compared with the control group, P>0.05; EA: Ellagic acid
Tail suspension test results
It was observed that there was a statistically significant decrease in immobility time with EA 1 mg/kg and sertraline treatment compared to the control group (P<0.01). However, although it reduced the immobility time in the EA+BKM120 and EA+MK2206 groups compared to the control group, no statistically significant difference was detected (Figure 2).
Figure 2.
Effects of EA, EA+BKM120, EA+MK2206, and sertraline treatment on the tail suspension test in mice

**The mean difference is significant when compared with the control group, P<0.01; EA: Ellagic acid
Western blot results for BDNF protein and AKT1 protein activation
BDNF levels in the mouse hippocampus were proportioned to GAPDH, and the ratio of the control group was accepted as one. The other groups were normalized to the control group. There was no statistically significant decrease in BDNF levels in the sertraline group compared to the control group. EA 1 mg/kg, EA+BKM120, and EA+MK2206 groups increased BDNF levels statistically significantly compared to the control group (P<0.05) (Figure 3).
Figure 3.
Effect of EA, EA+BKM120, EA+MK2206, and sertraline treatment on BDNF levels in mouse hippocampus

*The mean difference is significant when compared with the control group, P<0.05; EA: Ellagic acid
pAKT1 levels in the mouse hippocampus were proportioned to total AKT1, the control group’s ratio was accepted as one, and the other groups were normalized to the control group. EA 1 mg/kg and EA+BKM120 groups increased AKT1 phosphorylation statistically significantly compared to the control group (P<0.05). No statistically significant difference was observed in AKT1 phosphorylation in the sertraline and EA+MK2206 group compared to the control group EA dramatically up-regulated levels of p-Akt, and this up-regulation was partially reversed in the presence of the Akt inhibitor MK-2206 (Figure 4).
Figure 4.
Effect of EA, EA+BKM120, EA+MK2206, and sertraline treatment on AKT1 phosphorylation in mouse hippocampus
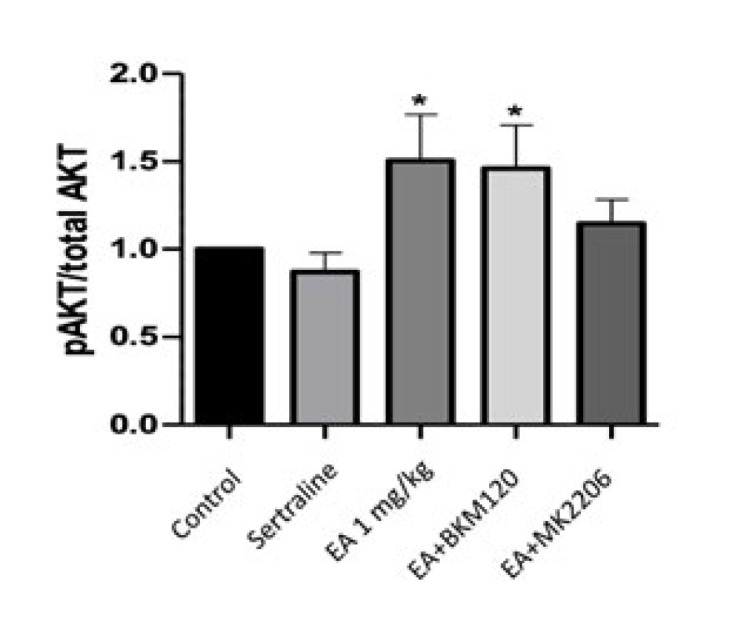
*The mean difference is significant when compared with the control group, P<0.05; EA: Ellagic acid
Discussion
To understand the multifactorial nature of depression, various hypotheses, such as the neuroplasticity hypothesis and the inflammation hypothesis, have been developed. According to the neuroplasticity hypothesis, BDNF levels decrease in depression, and this decrease is reversed with antidepressant treatment. The role of BDNF in the pathophysiology of depression has been intensively investigated for the last 20 years (17). Various studies have shown that bioactive molecules obtained from medicinal plants have antidepressant potential by acting through different pathways (18, 19). EA is a herbal molecule that is generally considered safe. Although there are bioavailability problems, the metabolites produced by the intestinal microbiota have also been shown to affect health positively. (20). EA is a plant secondary metabolite that has attracted attention in recent years with its antioxidant (21), anti-inflammatory (22), and anticarcinogenic (23) properties.
It has been demonstrated that herbal treatments may produce their antidepressant effects with BDNF-mediated responses. For example, liquiritigenin found in licorice root has shown antidepressant-like activity similar to fluoxetine in behavioral tests; this effect has been shown to be associated with BDNF/TrkB-mediated PI3K/AKT/GSK3 signaling in the hippocampus (24). Consumption of carotenoids, lycopene, and zeaxanthin has been associated with antidepressive symptoms with interaction of BDNF in silico (25).
In our study, the effects of EA were investigated by determining mouse hippocampal BDNF protein levels by western blot method, and it was shown that sertraline (5 mg/kg, 14 days) did not change the amount of BDNF in the mouse hippocampus. Consistent with our study, no statistically significant difference was detected in BDNF levels in the hippocampus of rats administered sertraline (5 mg/kg, 14 days) on the same day and dose, according to ELISA test results (26). In another study, fluoxetine, a selective serotonin reuptake inhibitor (SSRI), and sertraline were administered to mice for four weeks. Compared to the control group, BDNF levels in the hippocampus did not create a statistically significant change according to western blot results. However, chronic unpredictable mild stress reduced hippocampal BDNF levels compared to the control group, and this decrease was reversed when fluoxetine was given to the chronic stress group for four weeks (27). As can be understood from these studies, while SSRI group antidepressants do not affect BDNF levels in control group animals, they reverse the BDNF decrease caused by chronic stress.
One of the BDNF intracellular signaling pathways associated with the antidepressant-like effect is the PI3K/AKT pathway. An antidepressant-like effect was observed in the TST and forced swimming test in animals administered total saikosaponin, and it was shown that this effect was mediated by BDNF/pPI3K/pAKT up-regulation (28). In our previous study, we showed that EA has an antidepressant-like effect and increases BDNF levels in the mouse hippocampus. Moreover, in this study, we investigated whether the PI3K/AKT pathway plays a role in the effect of EA. We found that EA treatment reduced the immobility time in TST, and this effect was blocked in the presence of PI3K inhibitor (BKM120) or AKT inhibitor (MK2206). In a similar study, Pochwat et al. found that the immobility time in TST decreased after a single dose of hyperforin administration. The effects seen with hyperforin were blocked in the group given MK2206 (20 mg/kg) at the dose used in our study before hyperforin administration (29). Similarly, the antidepressant-like effects observed with vanillic acid were also eliminated with MK-2206 (60 mg/kg)(30).
In our study, EA increased hippocampal AKT1 phosphorylation. While BKM120 administration to animals receiving EA treatment did not change this increase, MK2206 administration disappeared EA’s increasing effect on AKT1 levels. Consistent with our study, in the study by Roy et al., BKM120 administration did not change AKTT308 and AKTS473 phosphorylation in the hippocampus compared to the control group (31).
It has been shown that Huperzine A (the active ingredient found in the Huperzia serrata plant) corrects the oxidative glutamate toxicity produced in hippocampal HT22 cells, and the modulation of the BDNF/TrkB-dependent PI3K/AKT/mTOR signaling pathway is effective in this neuroprotective effect (32). In an animal study, rats were given oral EA at a dose of 50 mg/kg, and it was determined that hippocampal BDNF, pAKT, and pPI3K levels increased with chronic EA treatment compared to the control group (33). In our study, EA significantly increased BDNF and pAKT1 levels compared to the control group. Therefore, it may be assumed that this pathway may be used in the antidepressant-like effect of EA.
Conclusion
Very few studies exist about the possible antidepressant effectiveness of EA and its underlying mechanisms. Our study showed that EA has an antidepressant-like effect in mice without changing locomotor activity, and the BDNF-PI3K-AKT signaling pathway may mediate this effect. Recently, EA has been subject to clinical trials. Ellagic acid supplementation had a favorable effect on depression in MS patients (34) and also in MDD patients (35). EA seems a promising agent in depression. However, more preclinical and clinical studies are needed to evaluate whether the results obtained from our study can pave the way for clinical application.
Acknowledgment
The results presented in this paper were part of a student thesis. This work was supported by The Scientific Research Projects Coordination Unit of Akdeniz University. Project Number: TDK-2020-5128.
Authors’ Contributions
HA B and C U designed the experiments; HA B performed experiments and collected data; HA B and C U discussed the results and strategy; C U supervised, directed, and managed the study; HA B and C U approved the final version to be published.
Ethics Statement
The current study protocol was approved by the Akdeniz University Animal Experiments Local Ethics Committee (Protocol number: 927/2019.07.04).
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Declaration
We have not used any AI tools or technologies to prepare this manuscript.
References
- 1.Rush AJ. Unipolar major depression in adults: Choosing initial treatment. Retrieved from 2022; https://www.uptodate.com/contents/unipolar-major-depression-in-adults-choosing-initial-treatment.
- 2.Goethe JW, Woolley SB, Cardoni AA, Woznicki BA, Pie DA. Selective serotonin reuptake inhibitor discontinuation: Side effects and other factors that influence medication adherence. J Clin Psychopharmacol. 2007;27:451–458. doi: 10.1097/jcp.0b013e31815152a5. [DOI] [PubMed] [Google Scholar]
- 3.Marasine NR, Sankhi S. Factors associated with antidepressant medication non- adherence. Turk J Pharm Sci. 2021;18:242–249. doi: 10.4274/tjps.galenos.2020.49799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nasrolahi A, Javaherforooshzadeh F, Mahmoudi J, Asl KD, Shabani Z. Therapeutic potential of neurotrophic factors in Alzheimer’s disease. Mol Biol Rep. 2022;49:2345–2357. doi: 10.1007/s11033-021-06968-9. [DOI] [PubMed] [Google Scholar]
- 5.Sathyanesan M, Newton SS. Antidepressant-like effects of trophic factor receptor signaling. Front Mol Neurosci. 2022;15:958797–958811. doi: 10.3389/fnmol.2022.958797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gupta VK, You Y, Gupta VB, Klistorner A, Graham SL. Trkb receptor signalling: Implications in neurodegenerative, psychiatric and proliferative disorders. Int J Mol Sci. 2013;14:10122–10142. doi: 10.3390/ijms140510122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ghigo A, Morello F, Perino A, Hirsch E. Phosphoinositide 3-kinases in health and disease. Subcell Biochem. 2012;58:183–213. doi: 10.1007/978-94-007-3012-0_6. [DOI] [PubMed] [Google Scholar]
- 8.Zhou C, Zhong J, Zou B, Fang L, Chen J, Deng X, et al. Meta-analyses of comparative efficacy of antidepressant medications on peripheral bdnf concentration in patients with depression. PLoS One. 2017;12:e0172270–172287. doi: 10.1371/journal.pone.0172270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Pelosof R, Santos LAD, Farhat LC, Gattaz WF, Talib L, Brunoni AR. BDNF blood levels after electroconvulsive therapy in patients with mood disorders: An updated systematic review and meta-analysis. World J Biol Psychiatry . 2022;24:24–33. doi: 10.1080/15622975.2022.2058083. [DOI] [PubMed] [Google Scholar]
- 10.Do Carmo Barbosa Mendes De Vasconcelos M, Bennett RN, Rosa EA, Ferreira Cardoso JV. Primary and secondary metabolite composition of kernels from three cultivars of Portuguese chestnut (Castanea sativa mill ) at different stages of industrial transformation. J Agric Food Chem. 2007;55:3508–3516. doi: 10.1021/jf0629080. [DOI] [PubMed] [Google Scholar]
- 11.Ribeiro B, Valentão P, Baptista P, Seabra RM, Andrade PB. Phenolic compounds, organic acids profiles and antioxidative properties of beefsteak fungus (Fistulina hepatica) Food Chem Toxicol. 2007;45:1805–1813. doi: 10.1016/j.fct.2007.03.015. [DOI] [PubMed] [Google Scholar]
- 12.Truzzi F, Tibaldi C, Zhang Y, Dinelli G, Eros DA. An overview on dietary polyphenols and their biopharmaceutical classification system (BCS) Int J Mol Sci. 2021;22:5514–5537. doi: 10.3390/ijms22115514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Huang X, Li W, You B, Tang W, Gan T, Feng C, et al. Serum metabonomic study on the antidepressant-like effects of ellagic acid in a chronic unpredictable mild stress-induced mouse model. J Agric Food Chem. 2020;68:9546–9556. doi: 10.1021/acs.jafc.0c02895. [DOI] [PubMed] [Google Scholar]
- 14.Bedel HA, Kencebay Manas C, Özbey G, Usta C. The antidepressant-like activity of ellagic acid and its effect on hippocampal brain-derived neurotrophic factor levels in mouse depression models. Nat Prod Res. 2018;32:2932–2935. doi: 10.1080/14786419.2017.1385021. [DOI] [PubMed] [Google Scholar]
- 15.Yusha’u Y, Muhammad UA, Nze M, Egwuma JM, Igomu OJ, Abdulkadir M. Modulatory role of rutin supplement on open space forced swim test murine model of depression. Niger J Physiol Sci. 2017;32:201–205. [PubMed] [Google Scholar]
- 16.Steru L, Chermat R, Thierry B, Simon P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology (Berl) 1985;85:367–370. doi: 10.1007/BF00428203. [DOI] [PubMed] [Google Scholar]
- 17.Zhao XP, Li H, Dai RP. Neuroimmune crosstalk through brain-derived neurotrophic factor and its precursor pro-BDNF: New insights into mood disorders. World J Psychiatry. 2022;12:379–392. doi: 10.5498/wjp.v12.i3.379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Can ÖD, Turan N, Demir Özkay Ü, Öztürk Y. Antidepressant-like effect of gallic acid in mice: Dual involvement of serotonergic and catecholaminergic systems. Life Sci. 2017;190:110–117. doi: 10.1016/j.lfs.2017.09.023. [DOI] [PubMed] [Google Scholar]
- 19.Diniz LRL, Souza MTS, Barboza JN, Almeida RN, Sousa DP. Antidepressant potential of cinnamic acids: Mechanisms of action and perspectives in drug development. Molecules. 2019;24:4469–4481. doi: 10.3390/molecules24244469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zuccari G, Baldassari S, Ailuno G, Turrini F, Alfei S, Caviglioli G. Formulation strategies to improve oral bioavailability of ellagic acid. Appl Sci. 2020;10:3353–3380. [Google Scholar]
- 21.Kilic I, Yeşiloğlu Y, Bayrak Y. Spectroscopic studies on the antioxidant activity of ellagic acid. Spectrochim Acta A Mol Biomol Spectrosc. 2014;130:447–452. doi: 10.1016/j.saa.2014.04.052. [DOI] [PubMed] [Google Scholar]
- 22.Bhattacharjee A, Kulkarni VH, Chakraborty M, Habbu PV, Ray A. Ellagic acid restored lead-induced nephrotoxicity by anti-inflammatory, anti-apoptotic and free radical scavenging activities. Heliyon. 2021;7:e05921–5928. doi: 10.1016/j.heliyon.2021.e05921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mohammadinejad A, Mohajeri T, Aleyaghoob G, Heidarian F, Kazemi Oskuee R. Ellagic acid as a potent anticancer drug: A comprehensive review on in vitro, in vivo, in silico, and drug delivery studies. Biotechnol Appl Biochem. 2022;69:2323–2356. doi: 10.1002/bab.2288. [DOI] [PubMed] [Google Scholar]
- 24.Tao W, Dong Y, Su Q, Wang H, Chen Y, Xue W, et al. Liquiritigenin reverses depression-like behavior in unpredictable chronic mild stress-induced mice by regulating Pİ3K/AKT/mTOR mediated BDNF/TrKB pathway. Behav Brain Res. 2016;308:177–186. doi: 10.1016/j.bbr.2016.04.039. [DOI] [PubMed] [Google Scholar]
- 25.Park SJ, Jaiswal V, Lee HJ. Dietary intake of flavonoids and carotenoids is associated with antidepressive symptoms: Epidemiological study and in silico-mechanism analysis. Antioxidants (Basel) 2021;11:53–73. doi: 10.3390/antiox11010053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Amidfar M, Réus GZ, Quevedo J, Kim YK, Arbabi M. Effect of co-administration of memantine and sertraline on the antidepressant-like activity and brain-derived neurotrophic factor (bdnf) levels in the rat brain. Brain Res Bull. 2017;128:29–33. doi: 10.1016/j.brainresbull.2016.11.003. [DOI] [PubMed] [Google Scholar]
- 27.Mu RH, Fang XY, Wang SS, Li CF, Chen SM, Chen XM, et al. Antidepressant-like effects of standardized gypenosides: Involvement of brain- derived neurotrophic factor signaling in hippocampus. Psychopharmacology (Berl) 2016;233:3211–3221. doi: 10.1007/s00213-016-4357-z. [DOI] [PubMed] [Google Scholar]
- 28.Zhou Z, Chen H, Tang X, He B, Gu L, Feng H. Total saikosaponins attenuates depression-like behaviors induced by chronic unpredictable mild stress in rats by regulating the Pİ3K/AKT/NF-ΚB signaling axis. Evid Based Complement Alternat Med. 2022;2022:4950414. doi: 10.1155/2022/4950414. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 29.Pochwat B, Szewczyk B, Kotarska K, Rafało-Ulińska A, Siwiec M, Sowa JE, et al. Hyperforin potentiates antidepressant-like activity of lanicemine in mice. Front Mol Neurosci. 2018;11:456–472. doi: 10.3389/fnmol.2018.00456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chuang HW, Wei IH, Lin FY, Li CT, Chen KT, Tsai MH, et al. Roles of akt and erk in mtor-dependent antidepressant effects of vanillic acid. ACS Omega. 2020;5:3709–3716. doi: 10.1021/acsomega.9b04271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Roy A, Skibo J, Kalume F, Ni J, Rankin S, Lu Y, et al. Mouse models of human PİK3CA-related brain overgrowth have acutely treatable epilepsy. ELife. 2015;4:e12703–12725. doi: 10.7554/eLife.12703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mao XY, Zhou HH, Li X, Liu ZQ. Huperzine a alleviates oxidative glutamate toxicity in hippocampal ht22 cells via activating BDNF/TRKB-dependent Pİ3K/AKT/MTOR signaling pathway. Cell Mol Neurobiol. 2016;36:915–925. doi: 10.1007/s10571-015-0276-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Alfaris NA, Alshammari GM, Altamimi JZ, Aljabryn DH, Alagal RI, Aldera H, et al. Ellagic acid prevents streptozotocin-induced hippocampal damage and memory loss in rats by stimulating NRF2 and nuclear factor-κb, and activating insulin receptor substrate/Pİ3K/AKT axis. J Physiol Pharmacol. 2021;72:503–515. doi: 10.26402/jpp.2021.4.02. [DOI] [PubMed] [Google Scholar]
- 34.Hajiluian G, Karegar SJ, Shidfar F, Aryaeian N, Salehi M, Lotfi T, et al. The effects of ellagic acid supplementation on neurotrophic, inflammation, and oxidative stress factors, and indoleamine 2, 3-dioxygenase gene expression in multiple sclerosis patients with mild to moderate depressive symptoms: A randomized, triple-blind, placebo-controlled trial. Phytomedicine. 2023;121:155094. doi: 10.1016/j.phymed.2023.155094. [DOI] [PubMed] [Google Scholar]
- 35.Hosseini SH, Zarghami M, Haghighian HK, Roochi AB, Abediankenari S, Kargar-Soleimanabad S, et al. Effects of ellagic acid supplementation on antioxidant status and symptom ımprovement in patients with major depressive disorder: A double-blind randomized clinical trial. Iran J Psychiatry Behav Sci. 2024;18:e144775. [Google Scholar]