

Abstract
Background
Analyses of Fgf10 and Fgfr2b mutant mice, as well as human studies, suggest that FGF10/FGFR2b signaling may play an essential, nonredundant role during embryonic SMG development. To address this question, we have analyzed the SMG phenotype in Fgf10 and Fgfr2b heterozygous and null mutant mice. In addition, although previous studies suggest that the FGF10/FGFR2b and FGF8/FGFR2c signaling pathways are functionally interrelated, little is known about the functional relationship between these two pathways during SMG development. We have designed in vivo and in vitro experiments to address this question.
Results
We analyzed Fgf10 and Fgfr2b heterozygous mutant and null mice and demonstrate dose-dependent SMG phenotypic differences. Hypoplastic SMGs are seen in Fgf10 and Fgfr2b heterozygotes whereas SMG aplasia is seen in Fgf10 and Fgfr2b null embryos. Complementary in vitro studies further indicate that FGF10/FGFR2b signaling regulates SMG epithelial branching and cell proliferation. To delineate the functional relationship between the FGF10/FGFR2b and FGF8/FGFR2c pathways, we compared the SMG phenotype in Fgfr2c+/Δ/Fgf10+/- double heterozygous mice to that seen in wildtype, Fgf10+/- (Fgfr2c+/+/Fgf10+/-) and Fgfr2c+/Δ (Fgfr2c+/Δ/Fgf10+/+) single heterozygous mutant littermates and demonstrate genotype-specific SMG phenotypes. In addition, exogenous FGF8 was able to rescue the abnormal SMG phenotype associated with abrogated FGFR2b signaling in vitro and restore branching to normal levels.
Conclusion
Our data indicates that FGF10/FGFR2b signaling is essential for the SMG epithelial branching and histodifferentiation, but not earliest initial bud formation. The functional presence of other endogenous signaling pathways could not prevent complete death of embryonic SMG cells in Fgf10 and Fgfr2b null mice. Though we were able to rescue the abnormal phenotype associated with reduced in vitro FGF10/FGFR2b signaling with exogenous FGF8 supplementation, our results indicate that the FGF10/FGFR2b and FGF8/FGFR2c are nonredundant signaling pathways essential for in vivo embryonic SMG development. What remains to be determined is the in vivo functional relationship between the FGF10/FGFR2b signal transduction pathway and other key signaling pathways, and how these pathways are integrated during embryonic SMG development to compose the functional epigenome.
Background
Mouse submandibular gland (SMG) development is initiated with a thickening of the oral epithelium of the mandibular arch around embryonic day 11.5 (E11.5) and is best conceptualized in stages[1,2]. In the PreBud Stage, SMG development begins as thickening of the oral epithelium adjacent to the tongue. During the Initial Bud Stage, this thickening grows down into mandibular arch mesenchyme to form the initial SMG bud. With continued epithelial proliferation and downgrowth, the SMG primordium becomes a solid, elongated epithelial stalk terminating in a bulb. Repeated end-bud branching results in the formation of a network of epithelial branches and terminal buds (the Pseudoglandular Stage). These epithelial branches and terminal buds hollow out by cell apoptosis during the Canalicular and Terminal Bud Stages, respectively, to form the ductal system and presumptive acini, with mucin protein being produced by the presumptive acini.
Morphogenesis of complex organs such as the SMG is regulated by the functional integration of parallel and broadly related signaling pathways which regulate cell proliferation, apoptosis and histodifferentiation [3-6]. To understand the complex interactions within this dynamic signaling network, one must first determine the contribution of individual pathways and identify those which play essential, nonredundant roles during embryonic SMG initial bud formation, branching morphogenesis and histodifferentiation.
The FGF family, with at least 22 members, mediates diverse biological functions such as cell proliferation, branching morphogenesis and histodifferentiation by binding and activating four tyrosine kinase receptors (FGFR 1–4) [see reviews [7-9]]. Tissue-specific alternate splicing of the Fgfr1, Fgfr2 and Fgfr3 genes generates isoforms which differentially bind specific FGF ligands [8,10]. Ligand-receptor binding potentially activates multiple intracellular cascades, including the ERK/RAS/MAPK, P13K, and PLC-γ/PKC pathways [see reviews [8,9,11]].
Functional studies have demonstrated that embryonic SMG epithelial cell proliferation, branching morphogenesis, and histodifferentation are regulated through growth factor, cytokine, and transcription factor-mediated signaling pathways, including EGF, TGF-β, Shh, FGFs, and Eda [2,6,12-21]. Although FGFs have been implicated in embryonic SMG development [13,15,22-24], a more complete understanding of their precise roles will provide insight into the complex network of parallel and broadly-related signaling pathways which regulate SMG organogenesis.
Gene targeting studies have clearly shown the importance of the FGFR2b (FGFR2-IIIb) signaling pathway for embryonic morphogenesis [22,23,25]. Fgfr2b-/- null mice die at birth due to lung insufficiency and exhibit severe dysmorphic organs, including agenesis or dysplasia of the lungs, mammary glands, pancreas, thyroid, teeth, and limbs [22,23,25,26]. Although FGF1, FGF3, FGF7, and FGF10 bind with high affinity to FGFR2b [8,10], the phenotypic similarities between Fgf10 and Fgfr2b null mice [22-25,27] indicate that FGF10 is the major ligand for FGFR2b in vivo. Of particular interest is the absence of SMGs in E14.5 and older Fgfr2b null mice and newborn Fgf10 null mice [23-25]. However, whether this glandular absence is due to the lack of development of a SMG initial bud (i.e. agenesis) or subsequent aplasia of the initial bud was heretofore unknown.
Recently, Entesarian et al. [28] have shown the importance of FGF10 gene dosage for salivary gland development in humans. Individuals with autosomal dominant ALSG (aplasia of lacrimal and salivary gland) exhibit hypoplastic or absent parotid and submandibular glands. ALSG was mapped to 5p13.2-5q13.1 to include the FGF10 gene; heterozygous FGF10 mutations were identified in all family members with ALSG. Complementary study of adult Fgf10+/- mutant mice revealed that Fgf10 heterozygotes have absent parotid glands and smaller SMGs although other organs such as lungs, liver, spleen, pancreas, thyroid, limbs appeared normal.
Taken together, the literature suggests that FGF10/FGFR2b signaling plays an essential, nonredundant, dose-dependent role during embryonic SMG development. To address this postulate, we evaluated SMG development in Fgfr2b and Fgf10 heterozygous mutant and null mice and demonstrate dose-dependent differences in SMG phenotypes. In a complementary set of in vitro experiments, we confirm the importance of FGF10/FGFR2b signaling and demonstrate that enhanced FGF10/FGFR2b signaling significantly induces, and abrogated FGF10/FGFRb signaling significantly diminishes, SMG branching morphogenesis and cell proliferation.
Our previous analyses of mutant mice and functional in vitro studies indicate that the FGF8/FGFR2c signaling pathway is essential for embryonic SMG epithelial branching morphogenesis and histodifferentiation [2,13]. Little is known about the functional relationship between the FGF10/FGFR2b and FGF8/FGFR2c pathways during SMG development. Thus, we evaluated the SMG phenotype in Fgfr2c+/Δ/Fgf10+/- double heterozygous mice, compared them to Fgf10+/- (Fgfr2c+/+/Fgf10+/-) and Fgfr2c+/Δ (Fgfr2c+/Δ/Fgf10+/+) single heterozygous mutant, as well as wildtype (WT), littermates and demonstrate genotype-specific SMG phenotypes. Though we were able to rescue the abnormal phenotype associated with reduced in vitro FGF10/FGFR2b signaling with exogenous FGF8 peptide supplementation, our results indicate that the FGF10/FGFR2b and FGF8/FGFR2c are nonredundant signaling pathways essential for in vivo embryonic SMG development.
Results
To delineate the role of FGF10/FGFR2b signaling during embryonic SMG development, we evaluated the SMG phenotype in Fgf10 and Fgfr2b null and heterozygous mutant mice. The E12.5 normal SMG appears as an elongated solid cord of epithelium terminating in an end-bulb (i.e., Initial Bud Stage) (Fig. 1A). By contrast, the E12.5 Fgfr2b null SMG is severely hypoplastic (compare Fig. 1B to 1A), displaying an extremely small initial bud similar to the earliest Initial Bud Stage [1,2]. A similar phenotype is seen in Fgf10 null mice (compare Fig. 1C to 1A, B). Normally by E13.5, epithelial proliferation results in SMG primordia with end-bulbs characterized by several branches; SMGs from both Fgfr2b and Fgf10 null mice are entirely absent (compare Fig. 2B, C to 2A). On the other hand, Fgfr2b+/- and Fgf10+/- heterozygosity results in SMG branching hypoplasia compared to WT glands, with fewer ducts and terminal buds being seen in heterozygous mutant SMGs than in WT SMG (compare Fig. 3B to 3A, D to 3C). Taken together, our data indicate that FGF10/FGFR2b signaling plays an essential, dose-dependent role during in vivo embryonic SMG branching morphogenesis and histodifferentation, but not earliest initial bud formation. Thus, the pathology in null mice is aplasia, not agenesis.
Figure 1.
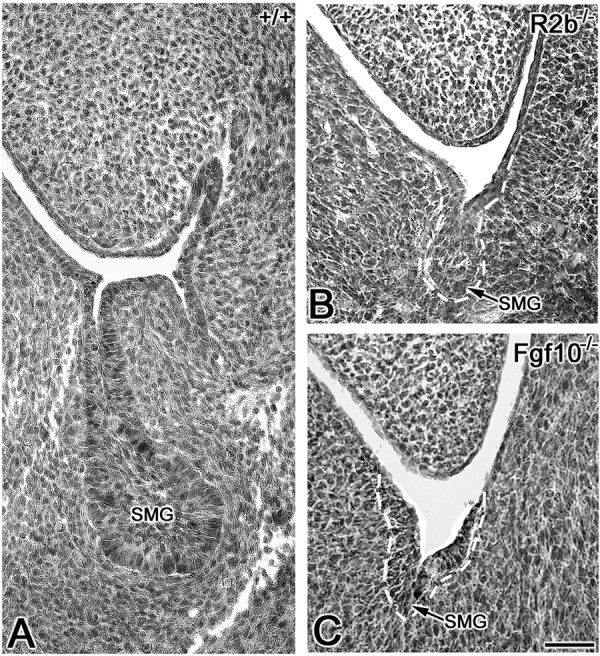
Abnormal SMG phenotypes in E12.5 Fgfr2b and Fgf10 null mutants A. E12.5 WT (+/+) mouse. B. E12.5 Fgfr2b-/- (R2b-/-) null mouse. C. E12.5 Fg10-/- null mouse. In the WT mouse (A), the Initial Bud Stage SMG is seen in the mandible ventrolateral to the tongue. By contrast, the SMG bud (outlined in white) in Fgfr2b-/- (B) and Fg10-/- (C) mice is extremely small, resembling the earliest Initial Bud Stage SMG. Bar, 50 μm.
Figure 2.

SMG aplasia in Fgfr2b and Fgf10 null mice. A. E13.5 WT mouse. B. E13.5 Fgfr2b-/- null mouse. C. E13.5 Fg10-/- null mouse. The E13.5 WT SMG (A) has achieved the Late Initial Bud Stage, with several branches being seen in the end-bulb epithelium; a small sublingual gland (SL) bud is seen lateral to the SMG bud. In Fgfr2b-/- (B) and Fg10-/- (C) embryos, no SMGs are found; undifferentiated mesenchyme is seen in the site normally occupied by SMG epithelia. PS-palatal shelf. Bar, 50 μm.
Figure 3.

Hypoplastic SMGs are seen in Fgfr2b and Fgf10 heterozygous mice. A. Newborn WT Fgfr2b+/+ SMG. B. Newborn Fgfr2b+/- heterozygous SMG. C. Newborn WT Fgf10+/+ SMG. D. Newborn Fg10+/- heterozygous SMG. WT Fgfr2b+/+ (A) and Fgf10+/+ (C) mice exhibit Late Terminal Bud Stage SMGs consisting of ducts and terminal buds displaying distinct lumina. In contrast, fewer ducts and terminal buds are seen in Fgfr2b+/- (B) and Fg10+/- (D) heterozygous SMGs compared to WT littermates. Bar, 50 μm.
Enhanced FGF10/FGFR2b signaling in vitro induces embryonic SMG branching morphogenesis and epithelial cell proliferation
To further delineate the role of FGF10/FGFR2b signaling during embryonic SMG development, we used our well-defined organ culture system to analyze the effect of enhanced FGF10/FGFR2b signaling on embryonic SMG branching morphogenesis. Paired E13 (Initial Bud Stage) or E14 (Pseudoglandular Stage) SMG primordia were cultured for up to 3 days in the presence or absence of FGF10 peptide (200 ng/ml or 500 ng/ml). Since a notable difference in SMG branch number is usually seen among littermates, we compared the number of terminal buds in right and left glands (treated and control) from each embryo. Spooner ratios (end bud number/initial bud number) were determined for each explant, the data were then arcsin transformed, and the mean ratios compared by paired t-test. FGF10 supplementation induced a significant increase in branching morphogenesis (Fig. 4A, B, E): 81% for E13 + 3 [200 ng/ml (P < 0.01); 500 ng/ml (P < 0.001)] and 46% for E14 + 2 [500 ng/ml (P < 0.0001)].
Figure 4.

Enhanced or abrogated FGF10/FGFR2b signaling modulates embryonic SMG branching morphogenesis in vitro. A-B. Enhanced signaling. Paired E13 SMG primordia were cultured for 3 days in the absence (A) or presence (B) of 500 ng/ml FGF10 peptide supplementation. FGF10 induced a significant increase in branching compared to control (CONT). C-D. Abrogated signaling. Paired E13 SMG primordia were cultured for 3 days in 10 ng/ml IgG-Fc (C) or FGFR2b-Fc chimera (D). FGFR2b-Fc chimera treated explant exhibits a significant decrease in branching compared to IgG-Fc control. Bar, 30 μm. E. Comparison of mean Spooner ratios in E13 + 3 and E14 + 2 explants. Exogenous FGF10 peptide supplementation induced a significant 81% increase in E13 +3 explants [200 ng/ml (P < 0.01); 500 ng/ml (P < 0.001)] and a significant 46% increase in E 14 + 2 explants (P < 0.0001) compared to controls. FGFR2b-Fc chimera-mediated interruption of E13 + 3 and E 14 + 2 SMGs significantly reduced branching morphogenesis by 22% [E13 +3: 10 ng/ml (P < 0.01); E 14 + 2: 5 ng/ml (P > 0.0001); 10 ng/ml (P < 0.01)] compared to controls.
Since cell proliferation is not required for early embryonic SMG epithelial branching [29], we determined if this FGF10-induced increase in branching is due to increased epithelial cell proliferation. We cultured E13+3 SMG primordia in the presence or absence of 500 ng/ml FGF10 peptide and calculated the epithelial cell proliferation index (the number PCNA-positive cells/total number of cells). Exogenous FGF10 induced a significant 78% (P < 0.0001) increase in epithelial cell proliferation compared to control (Fig. 5). Our results are similar to enhanced pancreatic epithelial cell proliferation and pancreatic hyperplasia in transgenic mice with persistent Fgf10 expression in developing pancreatic epithelia [30].
Figure 5.
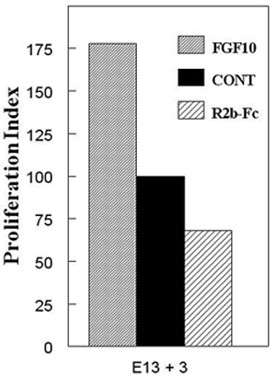
Enhanced or abrogated FGF10/FGFR2b signaling modulates cell proliferation. The cell proliferation index of E13 + 3 explants was calculated as the number of PCNA positive epithelial cells/total epithelial cells. Exogenous FGF10 peptide induced a significant 78% (P < 0.0001) increase, and FGFR2b-Fc chimera resulted in a significant 32% (P < 0.001) decrease, in cell proliferation compared to control.
Abrogated FGF10/FGFR2b signaling in vitro decreases embryonic SMG branching morphogenesis and cell proliferation
We interrupted FGF10/FGFR2b signaling in vitro by adding exogenous soluble FGFR2b-Fc chimera to the culture medium to competitively bind endogenous FGFR2b ligands. This exogenous receptor/ligand binding methodology has been successfully used to interrupt FGFR2b, FGFR2c, FGFR1b and FGFR1c signaling in vitro [13,15,21]. This experiment was designed to provide an in vitro model of the in vivo Fgf10+/- heterozygous mutant SMG phenotype (i.e. SMG hypoplasia). We conducted dose-response studies and determined the concentration of FGFR2b-Fc chimera which induces a clear biologic effect of reduced branching morphogenesis, thereby creating an in vitro model of the in vivo Fgf10 mutant heterozygote. Paired E13+3 or E14+2 SMG primordia were cultured in IgG-Fc (5 ng/ml or 10 ng/ml) or FGFR2b-Fc chimera (5 ng/ml or 10 ng/ml) and Spooner ratios were determined as described above. E13 + 3 FGFR2b-Fc-treated explants exhibit a significant 22% [10 ng/ml (P < 0.01)] decrease in branching morphogenesis compared to controls (Figs. 4C, D, E); a similar 22% reduction [5 ng/ml (P < 0.001); 10 ng/ml (P < 0.01)] was also seen in E14+2-treated explants (Figs. 4E). These results mimic the in vivo heterozygous mutants reported above.
Since exogenous FGF10 supplementation in vitro induced a significant increase in cell proliferation, we then evaluated the epithelial cell proliferation index in E13 +3 SMG primordia with or without abrogated FGF10/FGFR2b signaling. A significant 32% (P < 0.001) decrease in cell proliferation was seen in the presence of 10 ng/ml FGFR2b-Fc chimera compared to control (Fig. 5). Reduction in cell proliferation with FGFR2b-Fc treatment in vitro was also reported by Steinberg et al., [21]. These results indicate that FGF10/FGFR2b signaling modulates embryonic SMG epithelial cell proliferation and branching morphogenesis.
Relationship between FGFR2b and FGFR2c signaling pathways during SMG development
Our previous observation of hypoplastic SMGs in Fgfr2c+/Δ deficient mice suggested that FGFR2c signaling is necessary for embryonic SMG branching morphogenesis and histodifferentation [2]. The functional importance of this signaling pathway was confirmed in vitro; reduction of FGFR2c signaling in vitro resulted in a significant dose-dependent decrease in branching morphogenesis [13]. In addition, FGF8, a major FGFR2c ligand, has been shown to play an essential, nonredundant role during embryonic SMG development [13]. Like the hypoplastic SMG phenotype of Fgfr2c deficient mice, hypoplastic glands are seen in Fgf8 hypomorphic mice [13]. Importantly, SMG aplasia is seen in Fgf8 tissue-specific conditional mutant mice. To investigate the relationship between the FGFR2b and FGFR2c signal transduction pathways during in vivo SMG development, we evaluated the SMG phenotype in mice which are heterozygous for both the Fgf10 and Fgfr2c genes (i.e. Fgfr2c+/Δ/Fgf10+/- mutants). Mutant mice heterozygous for either Fgfr2c (Fgfr2c+/Δ/Fgf10+/+) or Fgf10 (Fgfr2c+/+/Fgf10+/-) exhibit hypoplastic SMGs compared to WT (Fgfr2c+/+/Fgf10+/+) littermates (compare Fig. 6B, C to 6A). In contrast, Fgfr2c+/Δ/Fgf10+/- double heterozygous mutant SMGs are smaller and exhibit fewer ducts and terminal buds than seen in single heterozygous Fgfr2c+/Δ/Fgf10+/+ or Fgfr2c+/+/Fgf10+/- mutant glands (compare Fig. 6D to 4B, C). These results suggest that, during embryonic SMG development, the FGFR2b and FGFR2c signal transduction pathways each induce at least some downstream targets that are idiosyncratic and not coincident.
Figure 6.

Fgfr2c+/Δ/Fgf10+/- double heterozygous mutant SMG is severely hypoplastic. A. Newborn R2c+/+/Fgf10+/+ WT SMG. B. Newborn R2c+/Δ/Fgf10+/+ (Fgfr2c+/Δ/Fgf10+/+) single heterozygous mutant SMG. C. Newborn R2c+/+/Fgf10+/-(Fgfr2c+/+/Fgf10+/-) single heterozygous mutant SMG. D. Newborn R2c+/Δ/Fgf10+/-(Fgfr2c+/Δ/Fgf10+/-) double heterozygous mutant SMG. The Fgfr2c+/Δ/Fgf10+/+ (B) and Fgfr2c+/+/ Fgf10+/- (C) single heterozygous SMGs have fewer ducts and terminal buds than seen in WT littermates (A). A much smaller, severely hypoplastic gland is seen in Fgfr2c+/Δ/Fgf10+/- double heterozygous mice (D) compared to Fgfr2c+/Δ/Fgf10+/+ (B) and Fgfr2c+/+/Fgf10+/- (C) littermates. Bar, 50 μm.
FGF8 rescues SMGs with interrupted FGF10/FGFR2b in vitro
Given that FGF8-mediated signaling plays an essential and unique role during SMG development [13], as well as our observation in Fgfr2c+/Δ/Fgf10+/- double heterozygous mutant mice discussed above, we postulated that exogenous FGF8 peptide would rescue the abnormal SMG phenotype associated with abrogated FGFR2b signaling in vitro. To determine if exogenous FGF8 peptide supplementation could restore branching morphogenesis to normal, we interrupted SMG morphogenesis in vitro with FGFR2b-Fc chimera and attempted to "rescue" these explants and restore branching to the level seen in controls. We cultured paired E13 + 3 SMGs in 10 ng/ml FGFR2b-Fc chimera for an initial period of 3 hours and then in FGFR2b-Fc + 500 ng/ml FGF8 peptide or FGFR2b-Fc alone for a total of 3 days (Fig. 7A, B). A second experiment to confirm that exogenous FGFR2b-Fc chimera decreased SMG branching morphogenesis consisted of paired E13 + 3 cultured in the presence of 10 ng/ml FGFR2b-Fc or IgG-Fc. In this set of experiments, FGFR2b-Fc-treated explants exhibit a significant 20% (P < 0.001) decrease in branching compared to IgG-Fc controls (Fig. 7C). Addition of exogenous FGF8 peptide induced a significant 24% (P < 0.02) increase in branching compared to FGFR2b-Fc treatment alone, thus completely restoring branching morphogenesis to the level seen in controls (Fig. 7C).
Figure 7.

Exogenous FGF8 supplementation in vitro rescues SMG branching morphogenesis. Paired E13 embryonic SMGs were preincubated for 3 hrs in 10 ng/ml FGFR2b-Fc chimera and then cultured for a total of 3 days with/without 500 ng/ml FGF8 peptide. A. E13+3 10 ng/ml FGFR2b-Fc-treated explant. B. E13+3 FGFR2b-Fc chimera + FGF8-treated explant. Bar, 30 μm. C. Comparison of Spooner ratios. A significant 20% (P < 0.001) decrease in branching morphogenesis with FGFR2b-Fc chimera abrogation was seen compared to IgG-Fc control. Exogenous FGF8 supplementation induced a significant 24% (P < 0.02) increase in branching to completely restore branching to the level seen in control.
Discussion
The FGF family of growth factors is critical to normal embryogenesis, regulating cell proliferation, survival and apoptosis [8]. Analyses of Fgfr2b and Fgf10 mutant and null mice clearly demonstrate that the FGF10/FGFR2b signal transduction pathway is essential for the development of branching organs, including the lung, mammary gland, lacrimal gland, pancreas, thyroid gland and salivary gland [23-28,31-33]. Although SMGs were absent from E14.5 or older Fgfr2b null mice and newborn Fgf10 null mice [22-25], the presence of an initial SMG bud in E12.5 Fgf10 and Fgfr2b null embryos (Fig. 1) indicates that this is a true aplasia, and not agenesis. Moreover, the absence of SMGs in E13.5 and older Fgfr2b-/- and Fgf10-/- mutants confirms that FGF10/FGFR2b signaling is essential for earliest initial epithelial branching and subsequent Pseudoglandular Stage and older SMG morphogenesis, but not earliest initial bud formation.
The observed initial SMG bud formation and subsequent aplasia is consistent with the pathogenesis seen in other organs, including the Fgfr2b null mammary gland [26], but differs from that seen in lung and pancreas [24,31,34]. Interestingly, Bellusci and colleagues [26] also detected genotype-specific phenotypic differences in mammary bud formation. A transient single initial mammary gland bud (bud 4) seen in E11.5 Fgfr2b-/- mice is absent in E12.5 mice whereas bud 4 is maintained (but does not branch) in Fgf10-/- mice. This result suggests that FGFR2b signaling is essential to maintain bud 4 and to induce the other mammary placodes whereas another FGFR2b ligand (probably FGF7) acts redundantly with FGF10 to maintain the mammary gland placode. Similarly, differences in pancreatic development were observed between Fgf10 and Fgfr2b null mice [31,35]. Taken together, these results clearly indicate that Fgfr2b and Fgf10 null mice demonstrate tissue-specific differences in affected organs.
Although Fgf10+/- heterozygous mice were described as being normal [24,34], lacrimal, parotid and submandibular gland aplasia or hypoplasia were recently reported in adult Fgf10+/- mice and in ALSG patients with FGF10 heterozygous mutations [28]. To determine if Fgfr2b and Fgf10 gene dosage plays an important role during embryonic SMG development, we evaluated newborn Fgfr2b+/- and Fgf10+/- and found SMG hypoplasia in both (Fig. 3). This is the first report of organ abnormality in Fgfr2b heterozygous mice. Our data indicate that SMG development is Fgf10 and Fgfr2b dose-dependent. Homozygous null mutants exhibit SMG aplasia while heterozygotes exhibit SMG hypoplasia. Moreover, the observations of normal lungs, livers, and limbs [23-25,28,34], but abnormal SMG phenotypes [28], in Fgfr2b+/- and Fgf10+/- mutant mice provides additional evidence of FGF10/FGFR2b tissue-specificity.
Relationship between FGF10/FGFR2b and FGF8/FGFR2c signaling pathways
It is critical to remember that FGF10 binding to FGFR2b is part of a much larger genetic network. Organogenesis is the programmed expression of regulatory genes coupled to downstream structural genes and epigenetic events. Specific signaling pathways are parallel and largely functionally redundant; that is, several pathways differentially and combinatorially compensate for the dysfunction of a given individual pathway. There are some pathways, however, that have unique and nonredundant functions. One has always to ask two key questions: Is our pathway of interest functionally redundant or nonredundant? Will a broadly related, not independent, pathway compensate for the dysfunction of our pathway of interest?
The observation of SMG aplasia in Fgf10 and Fgfr2b null mice indicates that the functional presence of other endogenous FGF/FGFR pathways (e.g., FGF8/FGFR2c, FGF/FGFR1) or other signaling pathways (e.g., TGF-α/EGF/EGFR, Eda/Edar; IGF-II/IGF-IR) could not prevent complete death of embryonic SMG cells in Fgf10-/- and Fgfr2b-/- mice. Interestingly, although SMG aplasia was also seen in Fgf8 (Fgf8C/N;AP2αIRESCre/+) conditional mutant mice in which Fgf8 expression was ablated from first branchial arch epithelium, a small initial SMG bud is still seen in E15.5 embryos [13]. By contrast, the Fgf10and Fgfr2b null SMG bud is much more transient, being seen in E12.5 and absent in E13.5 mice (present study). This suggests that the FGF10/FGFR2b and FGF8/FGFR2c signaling pathways are both essential for branching and that the FGF10/FGFR2b signal is necessary for epithelial bud maintenance at earlier stages than the FGF8/FGFR2c signal.
The absence of SMGs in both Fgf8C/N; AP2αIRESCre/+ conditional mutant [13] and Fgf10-/- null mice (present study) suggests that FGF8 and FGF10 probably induce some of the same downstream targets, although through different receptors (FGF10/FGFR2b and FGF8/FGFR2c). However, the mere fact of ontogenic arrest and SMG aplasia, as well as temporal differences, in Fgf8 conditional mutants and Fgf10 and Fgfr2b null mice indicate that FGF10/FGFR2b and FGF8/FGFR2c signaling pathways induce broadly-related, but unique and nonredundant downstream cascades in SMGs that cannot be compensated by normal function of the other under physiologic conditions. Our observation of smaller and more severely hypoplastic SMGs in Fgfr2c+/Δ/Fgf10+/- double heterozygous mice compared to either the Fgfr2c+/Δ or Fgf10+/- single heterozygous mutants further supports this conclusion.
It is well established that FGF/FGFR signaling can simultaneously activate multiple signaling cascades (e.g., ERK/RAS/MAPK, P13K, and PLC-γ/PKC to mediate epithelial cell proliferation, survival and histodifferentiation [8,9,11]. Inhibition of ERK/RAS/MAPK or P13K signaling significantly reduced embryonic SMG branching morphogenesis in vitro [17-19,21], whereas inhibition of PKC modestly increased morphogenesis [18]. A recent study of FGFR2b signaling in vitro suggests that FGF10/FGFR2b requires ERK activation to mediate cell proliferation and branching whereas FGF7/FGFR2b requires both ERK and P13K activation [21].
Although the importance of ERK/RAS/MAPK, P13K and PKC signaling during embryonic SMG development in vitro has been demonstrated, it is presently unclear which signaling cascades downstream of the FGF10/FGFR2b and FGF8/FGFR2c pathways are essential for epithelial branching morphogenesis, proliferation and survival during in vivo SMG development. This is important because in vitro and in vivo results may not necessarily coincide, suggesting the effect of differing physiologic conditions. Recently, Steinberg et al. [21] demonstrated that, in vitro, once the gland is formed, inhibition of FGF10 does not inhibit branching. This is the precise opposite of that seen in both the Fgf10-/- null mice and the Fgf10+/- heterozygotes in vivo (Fig. 1C, 2C, 3C), as well as FGF10 heterozygous mutant individuals with ALSG syndrome [28].
Fgf10 (Fgf10-/-) or Fgfr2b (Fgfr2b-/-) loss of function is ultimately epistatic to each other and to the epigenome under normal physiologic conditions (i.e. no other gene mutations nor untoward environments), the very reason they are critical to SMG morphogenesis. However, since the epistasis associated with declining Fgf10 or Fgfr2b function is a nonlinear emergent property of the complete functional epigenotype, it can be manipulated in vitro in the manner reported here. Exogenous FGF8 peptide can completely rescue and restore to normal the abnormal phenotype seen with abrogated FGF10/FGFR2 signaling in vitro (Fig. 7). This is not surprising since FGF8 has the ability to simultaneously activate similar, as well as unique and ligand-specific, intracellular cascades which control proliferation, survival, and differentiation. Rescue experiments have always to be only a proof of this principle, not a mimic of the in vivo condition. After all, Fgf10-/- and Fgfr2b-/- mutant SMGs are not rescued in vivo, the very essence of epistatic mutations. What remains to be determined is the in vivo functional relationship between the FGF10/FGFR2b signal transduction pathway and other key downstream signaling pathways, and how these pathways are integrated during embryonic SMG development to compose the functional epigenome.
Conclusion
Our results indicate that FGF10/FGFR2b signaling is essential for the SMG epithelial branching and histodifferentiation, but not earliest initial bud formation. The functional presence of other endogenous FGF pathways or other signaling pathways could not prevent complete death of embryonic SMG cells in Fgf10 and Fgfr2b null mice. Moreover, our analysis of Fgfr2c+/Δ/Fgf10+/- double heterozygous mice indicates that FGF10/FGFR2b and FGF8/FGFR2c signaling pathways induce broadly-related, but unique and nonredundant downstream cascades in SMGs. What remains to be determined is the in vivo functional relationship between the FGF10/FGFR2b signal transduction pathway and other key downstream signaling pathways, and how these pathways are integrated during embryonic SMG development to compose the functional epigenome.
Methods
Fgf10 and Fgfr2b mutant mice
Fgf10 and Fgfr2b mutant mice were generated on a C57Bl/6 background and genotyped by RT-PCR as previously described [23,26,34]. Wildtype mice (WT) (littermates or otherwise) were used as controls. WT (E11.5-E18.5), Fgf10-/- (E11.5-5-E14.5) and Fgfr2b-/- (E11.5-E15.5) embryos were collected. For analysis of heterozygous SMGs, WT and heterozygous mice were mated and newborn Fgf10+/-, Fgfr2b+/- and WT littermates were collected and their genotypes confirmed by RT-PCR. The embryos and newborn heads were fixed in 4% paraformaldehyde in PBS, and stored in 70% ethanol until further processing. E11.5-E14.5 heads and E15 and older SMGs were processed, embedded in paraplast and serial coronal sections were stained with hematoxylin and eosin as previously described [1]. A minimum of 3 SMGs per age per genotype were analyzed.
Generation of Fgfr2c+/Δ/Fgf10 +/- mice
Due to their neonatal lethality, Fgfr2-IIIc+/Δ mice are routinely generated by crossing males in which a copy of FgfR2-exon 9 (IIIc) has been flanked by loxP sites [36], with females carrying a PGK-Cre transgene [37]. However, to obtain the Fgfr2-IIIc +/Δ;Fgf-10 +/- allele, we used PGK-Cre females in to which we had previously introduced a heterozygous Fgf10 null allele (Fgf10 +/-) [27]. For simplicity, the Fgfr2-IIIc +/Δ;Fgf-10 +/- is called Fgfr2c +/Δ/Fgf-10 +/-. This cross resulted in the recovery of the following genotypes in the correct Mendelian ratios: Fgfr2c +/+/ Fgf10 +/+ (WT); Fgfr2c +/+/ Fgf10 +/-, Fgfr2c +/Δ/Fgf-10 +/+ and Fgfr2c +/Δ/Fgf-10 +/- [36]. Lines were maintained and crosses were performed in a C57/black 6 background. E17-newborn WT and mutant mice were generated, SMGs were fixed and processed as described above, and their genotypes confirmed by RT-PCR as previously described [27,36]. All animal studies were conducted with the approval of the appropriate committees regulating animal research.
Culture system
Timed-pregnant females [C57Bl/10 (B10.A)] were sacrificed on day 13 and day 14 of gestation, and embryos were dissected in cold PBS and staged according to Theiler [38]. E13 (Initial Bud Stage) and E14 (Pseudoglandular Stage) SMG primordia were cultured for up to 3 days using a modified Trowell method as previously described [6]. The medium consisted of BGJb (Life Technologies, Rockville, MD) supplemented with 1% BSA, 0.5 mg ascorbic acid/ml and 50 units penicillin/streptomycin (Life Technologies), pH 7.2, and replicate cultures were changed every day. Supplementation studies: paired E13 and E14 SMG primordia were cultured in the absence or presence of exogenous FGF10 peptide (200 or 500 ng/ml, R and D Systems) for 3 days (E13 + 3) or 2 days (E14 + 2), respectively; controls consisted of enriched BGJb alone. Because a notable difference in SMG epithelial branch number is seen between embryos within a given litter and among litters, we calculated the Spooner branch ratios (end bud number/initial bud number) for each explant as previously described [13] and compared the Spooner branch ratios in right and left glands (treated and control) from each embryo. Mean Spooner ratios were determined, the data were arcsin transformed to insure normality and homoscedasticity, and compared by paired t-test for all embryos studied [39]. In this set of experiments, 4–6 explants per treatment were analyzed.
Interruption studies
We interrupted FGF10/FGFR2b signaling using soluble FGFR2b-Fc chimera (R & D Systems, Inc). This method has been successfully used to interrupt FGFR2b, FGFR2c, FGFR1b and FGFR1c signaling during embryonic SMG development [13,15,21]. We conducted dose-response studies in which we cultured paired E13 + 3 or E14 + 2 SMG primordia in the presence of soluble FGFR2b-Fc chimera (5 or10 ng/ml) or control IgG-Fc (5 or 10 ng/ml IgG-Fc; R & D Systems, Inc), analyzed Spooner ratios as described above, and determined the optimal FGFR2b-Fc chimera concentration which results in a hypoplastic gland similar to that seen in the in vivo Fgf10 mutant heterozygotes. In this set of experiments, 4–8 explants per treatment were analyzed. Based on this set of experiments, we used 10 ng/ml FGFR2b-Fc chimera in all subsequent interruption experiments.
Cell proliferation assay
The cell proliferation index was determined as previously described [6]. Paired E13 + 3 were cultured in 10 ng/ml FGFR2b-Fc or 10 ng/ml IgG-Fc, fixed in 10% formalin, embedded in paraffin and serially-sectioned. The sections were stained with anti-PCNA for 1 hr using the Zymed mouse PCNA kit (South San Francisco, CA) and counterstained with hematoxylin as previously described [13]. The development time was adjusted according to experiment: 2–5 min for supplementation studies and 10–15 min for interruption studies. In these experiments, the cytoplasm appears blue and PCNA-positive cells appear brown. Three explants per group were analyzed. Control and treated explants were serially-sectioned and the midpoint of each explant identified. The section showing the explant's mid-point was selected, as well as the fourth section to the right and the fourth section to the left of the midpoint. This design insured that different buds were counted in each of the 3 sections/treatment. In each section, we photographed terminal bud clusters in the upper left and lower right at 400 × and counted a minimum of 3 buds/area.
Cell proliferation was quantitated as the ratio of PCNA-positive epithelial cells/total epithelial cells. The mean cell proliferation index was determined per section and the mean cell proliferation index of the three sections/explant was determined for each treatment. The data was arcsin transformed and the means ratios compared by t-test.
Rescue experiment
Paired E13 SMG primordia were cultured in 10 ng/ml FGFR2b-Fc chimera for an initial period of 3 hrs and then each pair was cultured in FGFR2b-Fc or FGFR2b-Fc + 500 ng/ml FGF8 (R and D Systems, Inc.) for 3 days. This FGF8 peptide concentration was determined to induce a significant increase in branching morphogenesis (data not shown). A concurrent control experiment was conducted as an internal control to verify that FGFR2b-Fc chimera supplementation interrupted branching morphogenesis; these controls consisted of E13 primordia cultured in 10 ng/ml FGFR2b-Fc chimera or 10 ng/ml IgG-Fc for 3 days. The explants were collected and mean Spooner ratios determined and compared as described above. Four explants per treatment were analyzed.
List of abbreviations
ALSG-aplasia of lacrimal and salivary gland
BSA-bovine serum albumen
CONT-control
FGF-Fibroblast growth factor
FGFR2b-FGF receptor 2-IIIb
FGFR2c-FGF receptor 2-IIIc
ERK-p44/p42 mitogen activating kinases (ERK-1/2)
MAPK-mitogen-activating protein kinases
P13K-phosphatidylinositol 3-kinase
PBS-phosphate-buffered saline
PKC-protein kinase C
PLC-γ-phospholipase C γ1
R2b-/--Fgfr2b-/-
R2c-Fgfr2c
RT-PCR-reverse transcriptase-polymerase chain reaction
SMG-submandibular salivary gland
WT-wildtype
Authors' contributions
TJ designed and coordinated the study, was involved in aspects of all experiments, and drafted the manuscript. GA prepared the histological sections for this study and performed some of the morphology and cell proliferation experiments. DW participated in morphological analyses and culture experiments and generated all figures. FGS generated the Fgf10 and Fgfr2b mutant mice and harvested and genotyped the embryos. SB assisted in the analyses of mutant mouse histopathology. MKH generated the Fgfr2c+/Δ/Fgf10 +/- single and double mutant mice, harvested and genotyped these embryos, and assisted in the analysis of their histopatholgy. MM participated in the design and coordination of this study, assisted in the analysis of histopathology, performed the statistical analysis, and helped draft the manuscript. All authors read and approved the final manuscript.
Acknowledgments
Acknowledgements
We gratefully acknowledge Dr. Clive Dickson in whose laboratory the double heterozygous mice were generated and maintained. This research was supported by a grant from American Lung Association (SB), NIH grants RO1 DE014535 (TJ/MM) and RO1 HL074832 (SB).
Contributor Information
Tina Jaskoll, Email: tjaskoll@usc.edu.
George Abichaker, Email: abichake@usc.edu.
Daniel Witcher, Email: witcher@usc.edu.
Frederic G Sala, Email: fsala@chla.usc.edu.
Saverio Bellusci, Email: sbellusci@chla.usc.edu.
Mohammad K Hajihosseini, Email: m.k.h@uea.ac.uk.
Michael Melnick, Email: mmelnick@usc.edu.
References
- Jaskoll T, Melnick M. Submandibular gland morphogenesis: stage-specific expression of TGF-alpha, EGF, IGF, TGF-beta, TNF and IL-6 signal transduction in normal mice and the phenotypic effects of TGF-beta2, TGF-beta3, and EGF-R null mutations. Anat Rec. 1999;256:252–268. doi: 10.1002/(SICI)1097-0185(19991101)256:3<252::AID-AR5>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- Jaskoll T, Zhou YM, Chai Y, Makarenkova HP, Collinson JM, West JD, Hajihosseini MK, Lee J, Melnick M. Embryonic submandibular gland morphogenesis: stage-specific protein localization of FGFs, BMPs, Pax 6 and Pax 9 and abnormal SMG phenotypes in FGF/R2-IIIc+Δ, BMP7-/- and Pax6-/- mice. Cells Tissues Organs. 2002;170:83–98. doi: 10.1159/000046183. [DOI] [PubMed] [Google Scholar]
- Davidson EH, Rast JP, Oliveri P, Ransick A, Calestanim C, Yuh CH, Minokawa T, Armore G, Hinman V, Arenas-Mena C, Otim O, Brown CT, Livi CB, Lee PY, Reville R, Tust AG, Pan ZJ, Schilstra MJ, Clarke PJC, Arnone ML, Rowen L, Cameron RA, McClay DR, Hood L, Bolour H. A genomic regulatory network for development. Science. 2002;295:1669–1678. doi: 10.1126/science.1069883. [DOI] [PubMed] [Google Scholar]
- Davidson EH, McClay DR, Hood L. Regulatory gene networks and the properties of the development process. Proc Natl Acad Sci. 2003;100:1475–1480. doi: 10.1073/pnas.0437746100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner TS, Bernardo DD, Lorenz D, Collins JJ. Inferring genetic networks and identifying compound mode of action via expression profiling. Science. 2003;301:102–105. doi: 10.1126/science.1081900. [DOI] [PubMed] [Google Scholar]
- Melnick M, Chen H, Zhou Y, Jaskoll T. The functional genomic response of developing embryonic submandibular glands to NF-κB inhibition. BMC Dev Biol. 2001;1:15. doi: 10.1186/1471-213X-1-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldfarb M. Signaling by fibroblast growth factors: the inside story. Sci STKE. 2001;2001:PE37. doi: 10.1126/stke.2001.106.pe37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ornitz DM, Itoh N. Fibroblast growth factors. Genome Biol. 2001;2:reviews3005. doi: 10.1186/gb-2001-2-3-reviews3005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsang M, Dawid IB. Promotion and attenuation of FGF signaling through the Ras-MAPK pathway. Sci STKE. 2004. PE17. [DOI] [PubMed]
- Ibrahimi OA, Zang F, Eliseenkova AV, Itoh N, Linhardt RJ, Mohammadi M. Biochemical analysis of pathogenic ligand-dependent FGFR2 mutations suggest distinct pathophysiological mechanisms for craniofacial and limb anomalies. Hum Mol Genet. 2004;13:2313–2324. doi: 10.1093/hmg/ddh235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlessinger J. Common and Distinct Elements in Cellular Signaling via EGF and FGF Receptors. Science. 2004;306:1506–1507. doi: 10.1126/science.1105396. [DOI] [PubMed] [Google Scholar]
- Hoffman MP, Kidder BL, Steinberg ZL, Lakhani S, Ho S, Klienman HK, Larsen M. Gene expression profiles of mouse submandibular gland development: FGFR1 regulates branching morphogenesis in vitro through BMP- and FGF- dependent mechanisms. Development. 2002;129:5767–5778. doi: 10.1242/dev.00172. [DOI] [PubMed] [Google Scholar]
- Jaskoll T, Witcher D, Toreno L, Bringas P, Moon AM, Melnick M. FGF8 dose-dependent regulation of embryonic submandibular salivary gland morphogenesis. Dev Biol. 2004;268:457–469. doi: 10.1016/j.ydbio.2004.01.004. [DOI] [PubMed] [Google Scholar]
- Jaskoll T, Leo T, Witcher D, Ormestad M, Astorga J, Carlsson P, Melnick M. Sonic Hedgehog signaling plays an essential role during embryonic salivary gland Epithelial Branching Morphogenesis. Dev Dyn. 2004;229:722–732. doi: 10.1002/dvdy.10472. [DOI] [PubMed] [Google Scholar]
- Jaskoll T, Melnick M. Embryonic salivary gland branching morphogenesis. In Branching Morphogenesis: Davies J. (Ed.) Georgetown: Landes Biosci. 2004. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=eurekah.chapter.21768
- Kashimata M, Gresik E. Epidermal growth factor system is a physiological regulator of development of the mouse fetal submandibular gland and regulates expression of the α6-integrin subunit. Dev Dyn. 1997;208:149–161. doi: 10.1002/(SICI)1097-0177(199702)208:2<149::AID-AJA2>3.0.CO;2-I. [DOI] [PubMed] [Google Scholar]
- Kashimata M, Sayeed S, Ka A, Onetta-Muda A, Sakagami H, Faraggiana T, Gresik E. The ERK-1/2 signaling pathway is involved in the stimulation of branching morphogenesis of fetal mouse submandibular glands by EGF. Dev Biol. 2000;220:183–196. doi: 10.1006/dbio.2000.9639. [DOI] [PubMed] [Google Scholar]
- Koyama N, Kashimata M, Sakagami H, Gresik EW. EGF-stimulated signaling by means of P13K, PLCγ1, and PKC isozymes regulates branching morphogenesis of the fetal mouse submandibular gland. Dev Dyn. 2003;227:216–226. doi: 10.1002/dvdy.10309. [DOI] [PubMed] [Google Scholar]
- Larsen M, Hoffman MP, Sakai T, Neibaur JC, Mitchell JM, Yamada KM. Role of PI 3-kinase and PIP3 in submandibular gland branching morphogenesis. Dev Biol. 2003;255:178–191. doi: 10.1016/S0012-1606(02)00047-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melnick M, Jaskoll T. Mouse submandibular gland morphogenesis: a paradigm for embryonic signal processing. Crit Rev Oral Biol Med. 2000;11:199–215. doi: 10.1177/10454411000110020401. [DOI] [PubMed] [Google Scholar]
- Steinberg Z, Myers C, Heim V, Lathrop CA, Rebustini IT, Stewart JS, Larsen M, Hoffman MP. FGFR2b signaling regulates ex vivo submandibular gland epithelial cell proliferation and branching morphogenesis. Development. 2005;132:1223–1234. doi: 10.1242/dev.01690. [DOI] [PubMed] [Google Scholar]
- Celli G, LaRochelle WJ, Makem S, Sharp R, Merlino G. Soluble dominant-negative receptor uncovers essential roles for fibroblast growth factors in multi-organ induction and patterning. EMBO J. 1998;17:1642–1655. doi: 10.1093/emboj/17.6.1642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeMoerlooze L, Spencer-Dene B, Revest JM, Hajihosseini MK, Rosewell I, Dickson C. An important role for the III isoform of fibroblast growth factor receptor 2 (FGFR2) in mesenchymal-epithelial signaling during mouse organogenesis. Development. 2000;127:483–492. doi: 10.1242/dev.127.3.483. [DOI] [PubMed] [Google Scholar]
- Ohuchi H, Hori Y, Yamasaki M, Harada H, Sekine K, Kato S, Itoh N. FGF10 acts as a major ligand for FGF Receptor 2 IIIb in mouse multi-organ development. Biochem Biophys Res Commun. 2000;277:643–649. doi: 10.1006/bbrc.2000.3721. [DOI] [PubMed] [Google Scholar]
- Revest JM, Spencer-Dene B, Kerr K, De Moerlooze L, Roswell I, Dickson C. Fibroblast growth factor receptor-2-IIIb acts upstream of Shh and Fgf4 and is required for limb bud maintenance but not for the induction of Fgf8, Msx1, or Bmp4. Dev Biol. 2001;231:47–62. doi: 10.1006/dbio.2000.0144. [DOI] [PubMed] [Google Scholar]
- Mailleux AA, Spencer-Dene B, Dillion C, Ndiaye D, Savona-Baron C, Itoh N, Kato S, Dickson C, Thiery JP, Bellusci S. Role of FGF10/FGFR2b signaling during mammary gland development in the mouse embryo. Development. 2002;129:53–60. doi: 10.1242/dev.129.1.53. [DOI] [PubMed] [Google Scholar]
- Min H, Danilenko DM, Scully SA, Bolon B, Ring BD, Tarpley JE, DeRose M, Simonet WS. Fgf-10 is required for both limb and lung development and exhibits striking functional similarity to Drosophila branchless. Genes Dev. 1998;12:3156–3161. doi: 10.1101/gad.12.20.3156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Entesarian M, Matsson H, Klar J, Bergendal B, Olson L, Arakaki R, Hayashi Y, Ohuchi H, Falahat B, Bolstad AI, Jonsson R, Wahren-Herlenius M, Dahl N. Mutations in the gene encoding fibroblast growth factor 10 are associated with aplasia of lacrimal and salivary glands. Nat Genet. 2005;37:125–127. doi: 10.1038/ng1507. [DOI] [PubMed] [Google Scholar]
- Nakanishi Y, Morita T, Nogawa H. Cell proliferation is not required for the initiation of early cleft formation in mouse embryonic submandibular epithelium in vitro. Development. 1987;99:429–437. doi: 10.1242/dev.99.3.429. [DOI] [PubMed] [Google Scholar]
- Hart A, Papadopoulou S, Edlund H. Fgf10 maintains Notch activation, stimulates proliferation, and blocks differentiation of pancreatic epithelial cells. Dev Dyn. 2003;228:185–193. doi: 10.1002/dvdy.10368. [DOI] [PubMed] [Google Scholar]
- Bhushan A, Itoh N, Kato S, Thiery JP, Czernichow P, Bellusci S, Scharfmann R. Fgf10 is essential for maintaining the proliferative capacity of epithelial progenitor cells during early pancreatic organogenesis. Development. 2001;128:5109–5117. doi: 10.1242/dev.128.24.5109. [DOI] [PubMed] [Google Scholar]
- Govindarajan V, Ito M, Makarenkova H, Lang RA, Overbeek PA. Endogenous and Ectopic Gland induction by FGF-10. Dev Biol. 2000;225:188–200. doi: 10.1006/dbio.2000.9812. [DOI] [PubMed] [Google Scholar]
- Mailleux A, Kelly R, Veltmaat JM, De Langhe SP, Zaffran S, Thiery JP, Bellusci S. Fgf10 expression identifies parabronchical smooth muscle cell progenitors and is required for their entry into the smooth muscle cell lineage. Development. 2005;132:2157–2166. doi: 10.1242/dev.01795. [DOI] [PubMed] [Google Scholar]
- Sekine K, Ohuchi H, Fujiwara M, Yamasaki M, Yoshizawa T, Sato T, Yagishita N, Matsui D, Koga Y, Itoh N, Kato S. Fgf10 is essential for limb and lung formation. Nat Genet. 1999;21:138–141. doi: 10.1038/5096. [DOI] [PubMed] [Google Scholar]
- Pulkkinen MA, Spencer-Dene B, Dickson C, Otonkoski T. The IIIb isoform of fibroblast growth factor receptor 2 is required for proper growth and branching of pancreatic ductal epithelium but not for differentiation of exocrine or endocrine cells. Mech Dev. 2003;120:167–175. doi: 10.1016/S0925-4773(02)00440-9. [DOI] [PubMed] [Google Scholar]
- Hajihosseini MK, Wilson S, De Moerlooze L, Dickson C. A splicing switch and gain-of-function mutation in FgfR2-IIIc hemizygotes causes Apert/Pfeiffer-syndrome-like phenotypes. Proc Natl Acad Sci U S A. 2001;98:3855–3860. doi: 10.1073/pnas.071586898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lallemand Y, Luria V, Haffner-Krausz R, Lonai P. Maternally expressed PGK-Cre transgene as a tool for early and uniform activation of the Cre site-specific recombinase. Transgenic Res. 1998;7:105–112. doi: 10.1023/A:1008868325009. [DOI] [PubMed] [Google Scholar]
- Theiler K. The House Mouse. New York, Springer-Verlag; 1989. [Google Scholar]
- Sokal R, Rohlf FJ. Biometry. New York, Freeman; 1981. [Google Scholar]