

Abstract
Extracellular matrix (ECM) is a vital component of the tumor microenvironment and plays a crucial role in the development and progression of gastric cancer (GC). Co-expression networks were established by means of the “WGCNA” package, the optimal model for extracellular matrix scores (ECMs) was developed and validated, with its accuracy in predicting the prognosis and treatment sensitivity of GC patients assessed. We performed univariate cox regression analysis [HR = 6.8 ( 3.3–14 ), p < 0.001] which demonstrated that ECMs was an independent risk character and perceptibly superior to other factors with further analysis of multivariate Cox regression [HR = 8.68 ( 4.16–18.08 ), p < 0.001]. The nomogram, presenting the clinical prognosis model for GC patients, demonstrated accuracy through KM analysis [HR = 3.97 (2.56–6.16), p < 0.001] and ROC curves with AUC values of 0.70, 0.72, and 0.72 at 1, 3, and 5 years, respectively. Using the ECMs model, we stratified GC patients into high- and low-risk groups, enabling precise predictions of prognosis and drug sensitivity. This stratification provides a new strategic direction for the personalized treatment of GC.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-025-88376-8.
Keywords: Gastric cancer, Extracellular matrix, Weighted gene co-expression network analysis, Prognosis, Immunotherapy
Subject terms: Cancer, Computational biology and bioinformatics, Immunology
Introduction
As the third most common cause of cancer-related death worldwide and the sixth most common disease overall, gastric cancer (GC) continues to pose a serious threat1. With a 5-year survival rate of roughly 25%, patients diagnosed with GC usually have a poor prognosis because of metastases, recurrent recurrences2. Despite the wide variety of medications available, there are still restrictions on available treatment options due to adverse effects, targetability, and drug resistance. For example, savolitinib is a highly selective small molecule inhibitor which can be used to contrapose advanced GC with mesenchymal epithelial transition factor (MET) abnormalities3,4, but it probably leads to acquired resistance in patients with MET-amplified GC for the target mechanisms of resistance MET D1228V/N/H and Y1230C mutations or high copy number MET gene amplifications5,6. Triplet chemotherapy with cisplatin, 5-fluorouracil, and docetaxel can improve survival but usually causing serious neutropenia and febrile neutropenia7. The cooperation between CCDC8 and JMJD2A, along with mechanisms involving ATP7A and histone H2AX, has been implicated in modulating gastric cancer susceptibility and resistance to oxaliplatin-based therapies, despite its efficacy and associated side effects such as polyneuropathy8–10. What’s more, it’s reported that drug resistance is present in 80% of the GC patients, including patients who are administered afatinib or erlotinib, which are important clinical drug11–13. Obviously, it’s difficult to achieve ideal effects merely taking conventional treatments. Therefore, identifying new biomarkers is crucial for advancing precision treatment and improving prognostic assessments for GC patients.
The tumor microenvironment (TME), which has long been linked to tumor growth, immune escape, and the efficacy of immunotherapy14,15, is made up of cancer cells, stromal cells, blood vessels, nerve fibers, extracellular matrix (ECM), and related acellular components16. Numerous research has suggested that expounding the cellular composition stands a chance of providing prognostic information and further improving the applicability of immunotherapy17,18. Thus, it’s necessary to conduct a further analysis to complex environment of TME by bioinformatics, which will be conducive to improve the personalized immunotherapy for GC. Therein, ECM, a three-dimensional, non-cellular and highly dynamic structure, is a complex aggregation which mainly consists of collagen, fibrous proteins, proteoglycans, cytokines, and other biomolecule19,20. ECM is also a momentous factor dictating tumor destiny21. Current evidence has indicated that ECM can lead up various cellular signaling cascades to encourage tumor growth and metastasis by interacting with ambient cells22. According to the clinical observation, changes in the composition of the ECM have much to do with invasiveness of tumors and prognosis23. Research has shown that epithelial-mesenchymal transition (EMT) is a critical process of tumor metastasis while increased ECM stiffness promotes EMT by activating EPHA2/LYN/TWIST1 signaling cascade in breast cancer24,25. In colorectal cancer, it’s confirmed that integrin α2β1 from ECM can boost metastatic capability and stemness of colorectal cancer cells with PI3K/AKT signal activated, besides, as the downstream of PI3K/AKT signaling, snail can further enhance invasion and metastasis of colorectal cancer26,27. However, there is a lack of research on evaluating GC with ECM, so it is crucial to investigate the relationship between ECM and GC in order to investigate ECM-based prognostic and efficacy prediction models for GC. These models would be especially beneficial for patient prognostic assessment and the creation of customized treatment strategies.
Thus, we used WGCNA to determine the key modules that are most pertinent to ECM and stromal abundance in GC in light of this creative thinking in this study. Furthermore, we established an ECM signature prognosis model in GC with its activity to assess the prognoses of GC patients precisely and response to immunotherapy evaluated.
Results
Identification of ECM-nominated genes
All four cohorts (TCGA-STAD, GSE26942, MSigDB, and GSE184198) were processed to meet the requirements of this study. Missing values and outliers were identified and addressed, while duplicate records were checked and removed. The “ESTIMATE” package was used to evaluate the Stromal Score, Immune Score, and ESTIMATE Score for all specimens in the TCGA-STAD cohort, providing metrics for WGCNA module filtering.
Based on the clinical data, the optimal Soft Threshold value was determined to be 7 (Fig. 1A), resulting in the generation of a clustering dendrogram that identified 55 modules (Fig. 1B). Among these, a specific and significant module, designated as cyan, was identified with high correlations to the Stromal Score (0.76), Immune Score (0.31), and ESTIMATE Score (0.58), all with p-values < 0.0001 (Fig. 1C). These findings indicate that this module is highly associated with the extracellular matrix (ECM) and immune infiltration. Further analysis using the MSigDB cohort yielded 201 genes from this module (Fig. 1D), which were subsequently categorized as ECM-related genes.
Fig. 1.

Identification of ECM-nominated Genes. (A) The scale-free topology criterion for TCGA-STAD requires a soft threshold power (β) of option 7. (B) Gene Co-expression Module Analysis recognized by cluster dendrogram. (C) Relevancy analysis of modules with StromalScore, ImmuneScore and ESTIMATEScore. (D) Venn diagram of the 1,026 ECM genes and 937 ECM-associated genes from WGCNA. (E) Forest plots presenting univariate Cox regression analyses of candidate ECM genes as independent prognostic factors. (F, G) Enrichment analysis of the candidate ECM genes by virtue of KEGG Enrichment Analysis (F) and Metascape database (G), respectively.
To refine these candidate genes, univariate Cox regression analysis was conducted to assess both the Hazard Ratio (HR) and p-values, identifying 83 significant genes, with the top 20 listed (Fig. 1E). Further exploration of these genes revealed that ECM-related genes were primarily enriched in the PI3K-Akt signaling pathway, as indicated by KEGG28–30 enrichment analysis (Fig. 1F), and in the NABA CORE MATRISOME category, based on the Metascape database (Fig. 1G).
The establishment of ECMs-score scoring model and prognostic analysis
The candidate genes filtered through univariate analysis underwent further testing using LASSO-Cox regression analysis (Fig. 2A and B), resulting in the identification of six key genes (Table S1) for the construction of the ECMs risk score model, with a λ value of 0.068. The final ECMs risk score was calculated as follows: ECMs risk score = expression of MMP16 * 0.04 + expression of NGF * 0.01 + expression of MMRN1 * 0.012 + expression of ANXA5 * 0.0006 + expression of IGFBP7 * 8.5e-06 + expression of PLOD2 * 0.00013. With that, it was eventually demonstrated that the model made sense in virtue of Kaplan-Meier (KM) curves [HR = 1.90 ( 1.39–2.61 ), p < 0.001 in the training cohort (Fig. 2C), HR = 2.92 ( 1.91–4.46 ), p < 0.001 in the validation (Fig. 2E) ]. Further confirmation was provided by Receiver Operating Characteristic (ROC) curve analysis, which yielded AUC values of 0.62, 0.68, and 0.67 at 1, 3, and 5 years, respectively, for the training cohort (Fig. 2D) and AUC values of 0.63, 0.66, and 0.67 at the same time points for the validation cohort (Fig. 2F), underscoring the model’s accuracy.
Fig. 2.

Establishment of ECMs-Score Scoring Model. (A, B) Coefficient profiles (B) of LASSO-Cox regression analysis, and the adjustment parameter (lambda) was calculated based on the partial likelihood deviance with ten-fold cross validation (A). (C, E) Kaplan-Meier analyses determined GC patients in the high-ECMs-risk group exhibiting worse overall survival both in TCGA-STAD (C) and GSE26942 (E) cohort. (D, F) ROC curves of ECMS in TCGA-STAD (D) and GSE26942 (F) cohort.
Next, we analyzed the clinical characteristics of patients in the low- and high-ECM risk score groups, including 5-year survival rates, age, T stage, N stage, and M stage (Fig. 3A). Significant differences were observed in the distributions of these clinical characteristics between the two groups. Consistent with our previous findings, the high-ECM risk score group exhibited a significantly lower 5-year survival rate (Fig. 3B). Additionally, a higher ECM risk score was strongly associated with a more advanced T stage (Fig. 3C). These findings suggest that the ECM risk score is closely linked to a higher risk of poor clinical outcomes.
Fig. 3.

Correlations between ECMs and clinical characteristics in GC. (A) Heatmap of the clinicopathological characteristics and ECMs. (B, C) The ECMs in different groups classified based on clinical characteristics from TCGA-STAD. (D, E) Violin diagrams of geographically and ethnically diverse cohorts and ECMs from TCGA-STAD. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
To further evaluate the robustness and generalizability of our model, we conducted additional analyses using geographically and ethnically diverse cohorts from TCGA-STAD (Fig. 3D-E). These analyses revealed significant differences in ECM-related characteristics among GC patients from different ethnicities and countries, underscoring the broader applicability of our findings.
We further explored the relationship between age, clinical stage, ECMs, and prognosis through univariate Cox regression analysis, which indicated that ECMs was an independent risk factor for prognosis [HR = 6.8 (3.3–14), p < 0.001; Fig. 4A]. This result demonstrated that ECMs was a stronger predictor of survival compared to other clinical factors. Multivariate Cox regression analysis reinforced these findings, indicating that ECMs maintained its prognostic significance [HR = 8.68 (4.16–18.08), p < 0.001; Fig. 4A].
Fig. 4.

Prognostic Analysis of ECMs. (A) Forest plots of univariate regression analyses and multivariable regression analyses, the ECM, Age, and Stage contained, indicating that ECM is an independent prognostic predictor and superior to other features. (B) Nomogram showing the risk assessment of patients. (C, D) the KM curves (C) and ROC curves (D) of Nomogram calibration predicting the OS at 1, 3, and 5 years.
To improve the clinical application of the ECMs model, we developed a nomogram that combines clinical and ECMs risk score data to predict the survival probability of GC patients (Fig. 4B). KM analysis confirmed the prognostic value of the nomogram, with HR = 3.97 (2.56–6.16), p < 0.001 (Fig. 4C), and ROC curve analysis yielded AUC values of 0.70, 0.72, and 0.72 at 1, 3, and 5 years, respectively (Fig. 4D), further validating the accuracy of the nomogram in predicting patient outcomes.
Functional enrichment analysis
To explore the underlying mechanisms by which ECMs characteristics influence the prognosis of GC, we performed a functional enrichment analysis using ssGSEA to analyze the GO and KEGG gene sets between the high- and low-ECM tisk groups, according to the optimum truncation. Differentially expressed genes (DEGs) were identified using the “limma” package, and the results were submitted to Metascape for further analysis. The results revealed that genes upregulated in the high ECM group were significantly associated with processes such as the mitotic cell cycle, ECM organization, and cell cycle checkpoints (Fig. 5A). In contrast, genes downregulated in the low ECM group were closely related to immunoglobulin production, regulation of membrane potential, and system process regulation (Fig. 5B).
Fig. 5.

Functional enrichment analysis. (A, B) Enrichment analysis of the DEGs in the High (A) and the Low (B) groups through Metascape database; (C, D) GSEA of KEGG gene sets between the High (C) and the Low (D) groups.
Further analysis using Gene Set Enrichment Analysis (GSEA) with gene expression data from the high and low ECM groups confirmed these findings. The DEGs were sorted by fold change to illustrate the trends of gene expression differences between the two groups. Functional enrichment analysis of the high ECM group showed that DEGs were predominantly involved in pathways such as Calcium Signaling, Neuroactive Ligand Receptor Interaction, Vascular Smooth Muscle Contraction, ECM Receptor Interaction, and Focal Adhesion (Fig. 5C). In contrast, the DEGs in the low ECM group were mainly enriched in pathways related to Ribosome, Proteasome, Base Excision Repair, Oxidative Phosphorylation, and DNA Replication (Fig. 5D).
Additional validation of these findings revealed consistent enrichment patterns. In the high ECM group, the DEGs were primarily associated with pathways such as Glycosphingolipid Biosynthesis Ganglio Series, Glycosaminoglycan Biosynthesis Chondroitin Sulfate, ECM Receptor Interaction, Primary Bile Acid Biosynthesis, and Vascular Smooth Muscle Contraction (Figure S1A). For the low ECM group, the dominant pathways included Steroid Biosynthesis, DNA Replication, Homologous Recombination, Proteasome, and Base Excision Repair (Figure S1B).
Immunoinfiltration analysis and prediction of drug susceptibility response
The Estimate score, based on the ssGSEA algorithm, showed significant differences in StromalScore, ImmuneScore, and ESTIMATEScore between the high and low ECM risk groups (p < 0.001). The relative abundance of mesenchymal and immune cells in the high ECM risk group was notably higher than in the low ECM risk group, with significant differences observed in immune cell infiltration and immune response levels (Fig. 6A). This highlights the strong correlation of our ECM model with the relative abundance of immune cells in tumor samples.
Fig. 6.

Immune landscape and Prediction of drug susceptibility response. (A) Box diagram of the “ESTIMATE” arithmetic score. (B, C) TIDE score predicts response to immunotherapy. The ECM signature discrepancies between immunotherapy-responders and non-responders groups. (B); Distributions of responders and non-responders in high- and low- ECMs-risk groups (C). (D)) Box diagram of the proportion of immune cell infiltration. (E) Box diagram comparing IC50 values of several chemotherapy drugs between high- and low- ECMs-risk groups. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001; ns, no significance.
At the same time, we applied the TIDE method to assess whether the ECM risk score could serve as a predictor for immunotherapy response in GC patients. For the TCGA-STAD cohort, the ECM risk score in the non-response subgroup was significantly higher than in the response group (p < 0.001; Fig. 6B). GC patients in the low ECM risk group were more sensitive to immunotherapy (140/251) compared to those in the high ECM risk group (44/134) (p < 0.05; Fig. 6C), suggesting that our ECM model is effective in predicting immunotherapy response.
Furthermore, CIBERSORTx was used to analyze the fraction of 22 immune-infiltrating cell types in patient samples. Significant differences in the distribution of 22 infiltrating immune cells between the high and low ECM risk groups were observed (Fig. 6D), with notable differences in nine subtypes of immune cells (p < 0.05). Four subtypes were downregulated in the high ECM risk group, including plasma cells (p < 0.01), activated memory CD4 + T cells (p < 0.01), follicular helper T cells (p < 0.001), and M0 macrophages (p < 0.05). Conversely, five cell types were upregulated in the high ECM risk group, including monocytes (p < 0.01), M2 macrophages (p < 0.0001), resting dendritic cells (p < 0.05), resting mast cells (p < 0.05), and eosinophils (p < 0.01).
Based on the GDSC database, Wilcoxon analysis revealed significant differences in the IC50 values of commonly used GC drugs between patients with high and low ECM risk. Patients in the low ECM risk group were more sensitive to afatinib, cisplatin, crizotinib, docetaxel, erlotinib, lapatinib, oxaliplatin, savolitinib, sorafenib, telomerase inhibitor IX, uprosertib, and docetaxel (p < 0.001). On the other hand, the high ECM risk group showed greater sensitivity to paclitaxel (p < 0.001). The evaluation of 5-Fluorouracil and Irinotecan showed no significant differences due to outliers (Fig. 6E). These findings underscore the ECM model’s potential in drug susceptibility assessment, offering a new dimension for personalized treatment strategies.
Substantiate the expression of hub genes in GC tissues
Given the findings from CIBERSORT, which indicated a close relationship between ECMs and immune cells, we employed Single-cell Sequencing31 to further explore this connection (Figure S2A-D). Interestingly, the six hub genes all exhibited significant expression in tissue stem cells. Specifically, NGF was predominantly expressed in epithelial cells, ANXA5 in monocytes, MMRN1 and IGFBP7 in endothelial cells, and MMP16 in both epithelial and endothelial cells (Figure S2E). These findings suggest that the hub genes play crucial roles in the TME.
Based on KM analysis derived from optimal truncation of the hub genes, we observed that patients with high expression of NGF and ANXA5 had poorer outcomes compared to those with low expression of these genes (Figure S3A-F). A heatmap of the six hub genes from TCGA-STAD further illustrates their expression levels in both non-cancerous and cancerous tissues at the mRNA level (Fig. 7A). To determine the protein expression levels of these six ECM-related genes in normal versus tumor tissue, we analyzed IHC images from the HPA database. The results revealed that these proteins were upregulated in tumors and exhibited strong staining in the GC matrix (Fig. 7B-F), though NGF was not included in the database. These findings support the notion that the six hub genes may serve as ECM-specific markers, with NGF requiring further validation in future studies.
Fig. 7.

Substantiate the expression of hub genes in GC tissues. (A) The heatmap of the hub genes in Non-cancer tissues and Cancerous tissue, respectively. (B-F) Protein expression levels and expression features of MMP16, MMRN1, ANXA5, IGFBP7 and PLOD2 in normal tissues and gastric cancer specimens from the Human Protein Atlas database.
Discussion
In GC, ECM plays a critical role, with its involvement spanning nearly all stages of the disease, from initiation to progression and metastasis19. For example, increased expression of tenascin has been observed in precancerous cells32,33, while dysregulated collagen expression is evident in more advanced stages of the disease34. Consistently, our findings indicate that patients in the high-ECM subgroup exhibit a poorer prognosis, with significant differences in the protein expression levels of ECM-related genes between normal and cancerous gastric tissues. These results suggest that ECM components have substantial clinical potential as prognostic biomarkers and therapeutic targets in GC. This study is the first to employ weighted gene co-expression network analysis (WGCNA) alongside multiple computational algorithms to investigate the ESTIMATE co-expression network of ECM-related genes. By applying univariate Cox regression and the LASSO-Cox regression algorithm, we identified six hub genes that formed a prognostic ECM model. The accuracy of this model was validated using KM analyses and ROC curves. Furthermore, both univariate and multivariate Cox regression analyses, considering age, stage, and ECM scores, confirmed that ECMs serve as independent prognostic factors, outperforming other clinical characteristics. Additionally, we developed a nomogram to assess patient risk, which demonstrated favorable prognostic performance, validated through KM analyses and ROC curves, further supporting its clinical utility.
The TME of GC contains both the ECM and immune cell components such as macrophages, lymphocytes, and neutrophils. These cellular components coexist and interact with each other, and their secreted metabolites and cytokines are as well as significant components of TME. These components of TME in GC play their role in inducing immune tolerance to promote GC progression14. Consistently, it is demonstrated synchronously that the ECMs model established is significant for the assessment of immune infiltration, immune checkpoints and immune escape. The immune infiltration analysis reveals a strong correlation between ECMs and various immune cells, especially M2 macrophages, suggesting their interaction with ECM components influences GC progression. Cancer-related metabolic reprogramming, including the acidic TME, plays a pivotal role in malignancy, metastasis, and immune evasion35. TME acidosis promotes the polarization of tumor-associated macrophages (TAMs) to the M2 phenotype, enhancing immune suppression36. ECMs also serve as reservoirs for TGF-β, which, through mutations in TGF-β signaling molecules, is involved in tumor formation. Tumor-derived TGF-β induces immune suppression by converting T cells to regulatory T cells (Tregs) and polarizing macrophages to the M2 phenotype37. TAMs, key players in tumor angiogenesis and lymphangiogenesis, secrete factors like VEGF, bFGF, and MMPs, and also contribute to tumor progression by inhibiting dendritic cell maturation38,39. This complex interplay between ECMs, immune cells, and tumor microenvironment significantly influences GC progression. The functional enrichment analysis showed that the candidate ECM genes were mostly enriched in PI3K-Akt signaling pathway which has been implicated in apoptosis, autophagy, and survival in GC40. Investigate thoroughly the differential genes in the High and the Low, it presented that the high group of ECM characteristics is strongly interrelated with the cell cycle, meanwhile, the genes down-regulated in low ECM group suggested a considerable association with immunity. The GSEA manifested that the pathways bound with ECM straightway or indirectly affect the occurrence, growth, invasion, and migration of carcinoma. For example, about the Ca subtypes, TRPV2 and TRPV4 are viewed as remarks of prognosis of patients of GC41. The ECM receptors exert their function by binding to the ECM ligands, mainly by stimulating integrin-dependent signaling, which promotes invasion and proliferation, creating a favorable microenvironment for metastatic cells, and interfering with communication between tumor cells and immune cells and other pathways19. DNA replication is regarded as the important portion of cellular response while uncontrolled DNA replication is one of the typical signs acquired by cancer cells42,43. In proteasome, colorectal neoplasia differentially expressed can promote the degradation of serine and arginine-rich splicing factor 6, which plays a role in regulating the sensitivity of GC to drugs44,45, and the tripartite motif 69 can inhibit the anoikis resistance and metastasis of GC by mediating the degradation of protein kinase C delta through the ubiquitin-proteasome pathway46.
GC treatment typically follows a comprehensive approach, combining radical surgery with adjuvant chemotherapy. The ECM model has been rigorously validated and provides a useful reference for evaluating the drug sensitivity of individual GC patients to commonly used drugs in clinical practice. 5-Fluorouracil (5-FU) is widely used in cancer treatment47. However, gastric carcinoma cells have developed resistance to 5-FU due to increasing methylation of genes and proteins, limiting its clinical application48. This drug resistance issue is also observed with Cisplatin, Lapatinib, and other chemotherapy agents49–51. Patient heterogeneity adds to the complexity of treatment, as individual sensitivities to drugs vary. Our analysis revealed that GC patients with low ECM risk scores exhibited higher sensitivity to treatments with Afatinib, Cisplatin, Crizotinib, Docetaxel, Erlotinib, Lapatinib, Oxaliplatin, Savolitinib, Sorafenib, Telomerase Inhibitor IX, Uprosertib, and Docetaxel. In contrast, subgroups with high ECM scores were more sensitive to Paclitaxel. Evaluation of 5-FU and Irinotecan showed no significant correlation due to outliers, but the reduction of ECM components, such as capillaries and stromal cells, alongside enhanced immune processes, was identified as a key feature of a favorable response to 5-FU, particularly in patients with diffuse GC52. Oxaliplatin exposure induces GC cells to secrete ECM, increasing tissue stiffness and promoting mitochondrial transfer via microvesicles from mesenchymal stem cells. These mitochondria fuse with GC cell mitochondria, reducing autophagy and enhancing cell survival. Single-cell sequencing revealed that both GC cells and mesenchymal stem cells actively remodel TME, driving ECM stiffness and drug resistance53. Therapeutic strategies targeting ECM remodeling, such as integrin inhibitors or ECM-degrading enzymes, show promise in overcoming these barriers19. Furthermore, combining ECM modulation with immune checkpoint inhibitors or anti-angiogenic therapies may enhance treatment efficacy by improving drug penetration and immune cell infiltration in resistant tumors54.
Regarding the six discerned genes in the model, ANXA5 can promote GC by affecting ECM-related processes and immune infiltration55 with studies suggesting that ANXA5 is not only closely associated with tumorigenesis and progression through PI3K/Akt/NF-κB pathway, but also enhances immunogenicity of tumor cells and the efficacy of related chemotherapy drugs56–58. Elevating IGFBP7 level promotes tumor progression by enhancing TAM/M2 macrophage polarization through the FGF2/FGFR1/PI3K/AKT axis in GC59,60. PLOD2 encodes a membrane-bound homo-dimeric enzyme helping to form ECM in tumors61, which can increase invasiveness and resistance to 5-FU of GC cells62. It has been pointed out that NSCLC metastasizes through the EGFR-PI3K/AKT-FOXA1-PLOD2 pathway, providing a basis for PLOD2 as a therapeutic target for NSCLC treatment63. It is reported that NGF recruits sensory nerves to the TME through axonal production64,65, and promotes the proliferation and growth of tumors using PLCγ pathways66, possessing the capability to impact ECM by enhancing the proliferation of basal cells67. In regard to GC, MMRN1 can interact with platelets to promotes the evasion of circulating cancer cells from natural killer cells, recruit ECM and granulocytes, and release angiogenic growth factors, thereby leading to the occurrence and metastasis of GC68. MMP16 is the downstream gene of β-catenin, which is closely associated with Wnt-mediated invasive and metastasis in GC cell69,70. It also can activate pro-Matrix Metalloproteinase 2 inducing the denaturation of type IV collagen and partially degrade type I collagen and other ECM proteins in the basement membrane70. Simultaneously, the expression of IGFBP7, ANXA5, and PLOD2 was significantly up-regulated in GC, while the expression of MMRN1 genes was prominently down-regulated71–73. Both ANXA5 and MMRN1 are considered biomarkers for GC diagnosis and progression68,73, while ANXA5 and MMP16 exhibit potential as predictive markers for lymph node and distant organ metastasis74,75. However, the exact functions of these ECM markers in GC remain unclear, and further studies are needed to elucidate their roles in proliferation, invasion, and metastasis.
Despite the promising findings, there are still several noteworthy limitations to our study. First, as a retrospective bioinformatics analysis based on three publicly available gene expression datasets, the prognostic accuracy and therapeutic value of the ECMs model require further validation with large sample sizes in real clinical settings. Additionally, the specific biological roles of ECM signature biomarkers in GC need to be confirmed through molecular biology and animal experiments. A key challenge in translating bioinformatics findings into clinical practice lies in patient heterogeneity and dataset biases, which must be addressed in future research. Furthermore, there is a lack of robust experimental methods to study ECM, and developing suitable techniques to explore its role in tumorigenesis and progression is crucial. Despite these limitations, our findings provide a foundational framework for future studies on the role of ECM in GC and its potential as a therapeutic target.
Conclusion
In summary, we established an ECMs model consisting of six hub genes and classified GC patients into high- and low-ECMs-risk groups to predict the prognosis and drug sensitivity of GC patients, which might provide new insights and options for individualized treatment of GC patients.
Methods
Data acquisition and processing
RNA-seq data and clinical information for 448 TCGA stomach adenocarcinoma (TCGA-STAD) cases were obtained from the GDC Data Portal of the Cancer Genome Atlas (TCGA) (https://portal.gdc.cancer.gov/). This dataset includes 412 tumor samples and 36 normal samples. The raw data were normalized to transcripts per million (TPM) and log-transformed. GSE2694276, which consists of 202 tumor samples, served as the validation cohort. Additionally, 1026 ECM-related genes were gathered from the Molecular Signatures Database (MSigDB) (https://www.gseamsigdb.org/gsea/msigdb/). These datasets were selected based on the following criteria: (a) primary human gastric cancer samples, (b) intact gene expression matrix, and (c) relevant clinical data including prognosis, stage, gender, and age. Single-cell RNA-seq data containing both GC and normal tissue samples were obtained from GSE18419877.
Weighted gene co-expression network analysis
The “ESTIMATE” package, based on the ssGSEA algorithm, was used to calculate Stromal, Immune, and ESTIMATE Scores for each sample. Co-expression networks were constructed using the “WGCNA” package. We determined the optimal soft threshold for the network by selecting a minimum power that yielded a scale-free topology fit close to 0.9. The co-expression network was then constructed using weighted genes with similar expression profiles, and gene modules were identified. A hierarchical clustering dendrogram was generated after calculating the topological overlap matrix (TOM), with average agglomerative clustering using the median non-weighted pairwise method.
Prognostic model construction and validation
The intersection of ECM-related genes and those identified through WGCNA was used to create a preliminary set of ECM genes. A prognostic model was constructed using univariate Cox regression analysis, followed by refinement using a false discovery rate (FDR) threshold of < 0.05. The model was further optimized by selecting the λ (lambda.min) that minimized the error during 10-fold cross-validation.
Extracellular matrix scores (ECMs)=
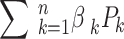 |
where k, βk, Pk refer to the number of dominating kernel genes, the coefficient index, and the gene expression levels, respectively. Patients were then stratified into high- and low-ECM risk groups (henceforth referred to as High and Low, respectively) based on the calculated ECM risk score and the optimal cutoff value. To evaluate the predictive accuracy and robustness of the model, both the training and validation cohorts were analyzed using the “survival” and “survivalROC” R packages. These analyses provided a comprehensive assessment of the model’s prognostic performance and credibility.
Enrichment analysis and immune landscape
Enrichment Analysis respectively coursed and construed via “clusterProfiler” and “org.Hs.eg.db” packages and Metascape database. As the “limma” package made a thorough inquiry about the differential genes, it was adopted that “clusterProfiler”, “enrichplot” and “patchwork” packages were employed to proceed Gene Set Enrichment Analysis (GSEA) to deliberate the discrepancy genes both in the High and Low groups in which pathway of major enrichment. The immune cell abundance analysis was performed using the “CIBERSORT” package, which deconvoluted the immune cell subtype expression matrix. Tumor immune dysfunction and rejection were evaluated using the TIDE algorithm to assess immune escape potential. The “Predict Response” function was used to evaluate responses to immune checkpoint blockade.
Drug susceptibility assessment
Drug sensitivity (IC50) was assessed using the “get_oncePredict_res” function from the “oncoPredict” package77. Expression data and pType data from the GDSC2 database were obtained from OSFHOME (https://osf.io/), which includes both training sets and corresponding drug response data. The “calcPhenoty” function from the “oncoPredict” package was used to predict the drug activity of each sample, enabling the evaluation of ECM classification effects on clinical drug therapy in GC.
Human protein atlas (HPA) databases
For protein level analysis, immunohistochemical (IHC) staining images from GC tissues were retrieved from the HPA online database (https://www.proteinatlas.org/)78, which yielded the localization and degree of expression of the target proteins.
Statistical analysis
All statistical analyses and graphing were performed using R software (version 4.3.1). The “WGCNA” package was used to identify co-expressed gene modules, and the “survival,” “survminer,” and “glmnet” packages were employed to construct the prognostic model, perform univariate and multivariate Cox regression analyses, and assess predictive accuracy using the “survivalROC” package. The “rms” package was used to plot the nomogram. Statistical significance was considered at p < 0.05 unless otherwise specified.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Author contributions
NX and HMZ conceived and designed this study. NX established the methods and collected, processed and analyzed the data. NX, TJZ and WWS drafted and wrote this manuscript. NX, HMZ, TJZ, WWS and CXY made the manuscript revised. All authors read and approved the final manuscript.
Data availability
Publicly available datasets were analyzed in this study. These data can be found here: GDC Data Portal of TCGA (https://portal.gdc.cancer.gov/); GEO datasets (https:// www.ncbi.nlm.nih.gov/geo/): GSE26942 and GSE184198.
Declarations
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Sung, H. et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Cancer J. Clin.71(3), 209–249. 10.3322/caac.21660 (2021). [DOI] [PubMed] [Google Scholar]
- 2.Li, X. et al. Identification of N7-methylguanosine related subtypes and construction of prognostic model in gastric cancer. Front. Immunol.1310.3389/fimmu.2022.984149 (2022). [DOI] [PMC free article] [PubMed]
- 3.Kim, J. S., Kim, M. Y. & Hong, S. Characterization of MET alterations in 37 gastroesophageal Cancer cell lines for MET-Targeted therapy. Int. J. Mol. Sci.25(11), 5975. 10.3390/ijms25115975 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kim, S. T. et al. Combination of Docetaxel Plus Savolitinib in Refractory Cancer patients: a Report on Phase I Trial. Translational Oncol.12(4), 597–601. 10.1016/j.tranon.2018.12.009 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Frigault, M. M. et al. Mechanisms of acquired resistance to savolitinib, a selective MET inhibitor in MET-amplified gastric cancer. JCO Precision Oncol.4(PO.19.00386). 10.1200/PO.19.00386 (2020). [DOI] [PMC free article] [PubMed]
- 6.Zhong, L. et al. Small molecules in targeted cancer therapy: advances, challenges, and future perspectives. Signal. Transduct. Target. Therapy. 6(1), 201. 10.1038/s41392-021-00572-w (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hartgrink, H. H., Jansen, E. P., van Grieken, N. C. & van de Velde, C. J. Gastric cancer. Lancet374(9688), 477–490. 10.1016/S0140-6736(09)60617-6 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nakagawa, T. et al. JMJD2A sensitizes gastric cancer to chemotherapy by cooperating with CCDC8. Gastric Cancer: Official J. Int. Gastric Cancer Association Japanese Gastric Cancer Association. 23(3), 426–436. 10.1007/s10120-019-01024-9 (2020). [DOI] [PubMed] [Google Scholar]
- 9.Sun, Y. et al. In vitro and in vivo evaluation of a novel estrogen-targeted PEGylated oxaliplatin liposome for gastric cancer. Int. J. Nanomed.16, 8279–8303. 10.2147/IJN.S340180 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chen, C. C. et al. Combined modalities of resistance in an oxaliplatin-resistant human gastric cancer cell line with enhanced sensitivity to 5-fluorouracil. Br. J. Cancer. 97(3), 334–344. 10.1038/sj.bjc.6603866 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li, Y. et al. Author correction: proteomic characterization of gastric cancer response to chemotherapy and targeted therapy reveals potential therapeutic strategies. Nat. Commun.13(1), 6749. 10.1038/s41467-022-34238-0 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sun, J. et al. Knockdown of CALM2 increases the sensitivity to afatinib in HER2-amplified gastric cancer cells by regulating the Akt/FoxO3a/Puma axis. Toxicol. Vitro: Int. J. Published Association BIBRA. 87, 105531. 10.1016/j.tiv.2022.105531 (2023). [DOI] [PubMed] [Google Scholar]
- 13.Cha, M. Y. et al. Antitumor activity of HM781-36B, a highly effective pan-HER inhibitor in erlotinib-resistant NSCLC and other EGFR-dependent cancer models. Int. J. Cancer. 130(10), 2445–2454. 10.1002/ijc.26276 (2012). [DOI] [PubMed] [Google Scholar]
- 14.Jin, M. Z. & Jin, W. L. The updated landscape of tumor microenvironment and drug repurposing. Signal. Transduct. Target. Therapy. 5(1), 166. 10.1038/s41392-020-00280-x (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zhang, B. et al. M(6)a regulator-mediated methylation modification patterns and tumor microenvironment infiltration characterization in gastric cancer. Mol. Cancer. 19(1), 53. 10.1186/s12943-020-01170-0 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kang, B. et al. Parallel single-cell and bulk transcriptome analyses reveal key features of the gastric tumor microenvironment. Genome Biol.23(1), 265. 10.1186/s13059-022-02828-2 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cui, K. et al. A novel high-risk subpopulation identified by CTSL and ZBTB7B in gastric cancer. Br. J. Cancer. 127(8), 1450–1460. 10.1038/s41416-022-01936-x (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Liu, J. et al. The combined signatures of the tumour microenvironment and nucleotide metabolism-related genes provide a prognostic and therapeutic biomarker for gastric cancer. Sci. Rep.13(1), 6622. 10.1038/s41598-023-33213-z (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Moreira, A. M. et al. The extracellular matrix: an accomplice in gastric cancer development and progression. Cells9(2), 394. 10.3390/cells9020394 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bonnans, C., Chou, J. & Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol.15(12), 786–801. 10.1038/nrm3904 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Guo, K. S. & Brodsky, A. S. Tumor collagens predict genetic features and patient outcomes. NPJ Genomic Med.8(1), 15. 10.1038/s41525-023-00358-9 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jiang, Y. et al. Targeting extracellular matrix stiffness and mechanotransducers to improve cancer therapy. J. Hematol. Oncol.15(1), 34. 10.1186/s13045-022-01252-0 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yang, Z. et al. Extracellular matrix characterization in gastric cancer helps to predict prognosis and chemotherapy response. Front. Oncol.1110.3389/fonc.2021.753330 (2021). [DOI] [PMC free article] [PubMed]
- 24.Heerboth, S. et al. EMT and tumor metastasis. Clin. Translational Med.4, 6. 10.1186/s40169-015-0048-3 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fattet, L. et al. Matrix rigidity controls epithelial-mesenchymal plasticity and tumor metastasis via a mechanoresponsive EPHA2/LYN complex. Dev. Cell. 54(3), 302–316e7. 10.1016/j.devcel.2020.05.031 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wu, X., Cai, J., Zuo, Z. & Li, J. Collagen facilitates the colorectal cancer stemness and metastasis through an integrin/PI3K/AKT/Snail signaling pathway. Biomed. Pharmacotherapy = Biomedecine Pharmacotherapie. 114, 108708. 10.1016/j.biopha.2019.108708 (2019). [DOI] [PubMed] [Google Scholar]
- 27.Ungewiss, C. et al. The microRNA-200/Zeb1 axis regulates ECM-dependent β1-integrin/FAK signaling, cancer cell invasion and metastasis through CRKL. Sci. Rep.6, 18652. 10.1038/srep18652 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kanehisa, M. et al. KEGG: biological systems database as a model of the real world. Nucleic Acids Res.53(D1), D672–D677. 10.1093/nar/gkae909 (2025). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Science: Publication Protein Soc.28(11), 1947–1951. 10.1002/pro.3715 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kanehisa, M. & Goto, S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res.28(1), 27–30. 10.1093/nar/28.1.27 (2000). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Aran, D. et al. Reference-based analysis of lung single-cell sequencing reveals a transitional profibrotic macrophage. Nat. Immunol.20(2), 163–172. 10.1038/s41590-018-0276-y (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tiitta, O., Sipponen, P., Gould, V. & Virtanen, I. Tenascin expression in inflammatory, dysplastic and neoplastic lesions of the human stomach. Virchows Archiv: Int. J. Pathol.425(4), 369–374. 10.1007/BF00189574 (1994). [DOI] [PubMed] [Google Scholar]
- 33.Jiang, X. et al. COL12A1, a novel potential prognostic factor and therapeutic target in gastric cancer. Mol. Med. Rep.20(4), 3103–3112. 10.3892/mmr.2019.10548 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Webb, B. A. et al. Dysregulated pH: a perfect storm for cancer progression. Nat. Rev. Cancer. 11(9), 671–677. 10.1038/nrc3110 (2011). [DOI] [PubMed] [Google Scholar]
- 35.He, Z. & Zhang, S. Tumor-Associated macrophages and their Functional Transformation in the hypoxic Tumor Microenvironment. Front. Immunol.12, 741305. 10.3389/fimmu.2021.741305 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wakefield, L. M. & Roberts, A. B. TGF-beta signaling: positive and negative effects on tumorigenesis. Curr. Opin. Genet. Dev.12(1), 22–29. 10.1016/s0959-437x(01)00259-3 (2002). [DOI] [PubMed] [Google Scholar]
- 37.Riabov, V. et al. Role of tumor associated macrophages in tumor angiogenesis and lymphangiogenesis. Front. Physiol.5, 75. 10.3389/fphys.2014.00075 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Schlereth, S. L. et al. Impact of the prolymphangiogenic crosstalk in the tumor microenvironment on lymphatic cancer metastasis. Biomed. Res. Int.2014, 639058. 10.1155/2014/639058 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Manning, B. D., Toker, A. A. K. T. P. K. B. & Signaling Navigating the network. Cell169(3), 381–405. 10.1016/j.cell.2017.04.001 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tang, B. et al. VPAC1 couples with TRPV4 channel to promote calcium-dependent gastric cancer progression via a novel autocrine mechanism. Oncogene38(20), 3946–3961. 10.1038/s41388-019-0709-6 (2019). [DOI] [PubMed] [Google Scholar]
- 41.Macheret, M. & Halazonetis, T. D. DNA replication stress as a hallmark of cancer. Annu. Rev. Pathol.10, 425–448. 10.1146/annurev-pathol-012414-040424 (2015). [DOI] [PubMed] [Google Scholar]
- 42.Mughal, M. J., Mahadevappa, R. & Kwok, H. F. DNA replication licensing proteins: saints and sinners in cancer. Sem. Cancer Biol.58, 11–21. 10.1016/j.semcancer.2018.11.009 (2019). [DOI] [PubMed] [Google Scholar]
- 43.An, P. et al. Natural products are an important source for proteasome regulating agents. Phytomedicine: Int. J. Phytotherapy Phytopharmacology. 9310.1016/j.phymed.2021.153799 (2021). [DOI] [PubMed]
- 44.Zhang, F. et al. LncRNA CRNDE attenuates chemoresistance in gastric cancer via SRSF6-regulated alternative splicing of PICALM. Mol. Cancer. 20(1), 6. 10.1186/s12943-020-01299-y (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sun, L. et al. TRIM69 suppressed the anoikis resistance and metastasis of gastric cancer through ubiquitin–proteasome-mediated degradation of PRKCD. Oncogene42(49), 3619–3632. 10.1038/s41388-023-02873-6 (2023). [DOI] [PubMed] [Google Scholar]
- 46.Ge, J. et al. Retrospective Cohort Study of Intraoperative Administration of Sustained-Release 5-Fluorouracil implants in Advanced Gastric Cancer patients. Front. Pharmacol.1210.3389/fphar.2021.659258 (2021). [DOI] [PMC free article] [PubMed]
- 47.Qin, Y. et al. MeCP2 confers 5-fluorouracil resistance in gastric cancer via upregulating the NOX4/PKM2 pathway. Cancer Cell Int.22(1), 86. 10.1186/s12935-022-02489-y (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Trendowski, M. R. et al. Genetic and modifiable risk factors contributing to cisplatin-induced toxicities. Clin. cancer Research: Official J. Am. Association Cancer Res.25(4), 1147–1155. 10.1158/1078-0432.CCR-18-2244 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Choi, K. M. et al. Prolonged MEK inhibition leads to acquired resistance and increased invasiveness in KRAS mutant gastric cancer. Biochem. Biophys. Res. Commun.507(1–4), 311–318. 10.1016/j.bbrc.2018.11.030 (2018). [DOI] [PubMed] [Google Scholar]
- 50.Ning, G. et al. A novel treatment strategy for lapatinib resistance in a subset of HER2-amplified gastric cancer. BMC Cancer. 21(1), 923. 10.1186/s12885-021-08283-9 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dong, S. et al. A combined analysis of bulk and single-cell sequencing data reveals that depleted extracellular matrix and enhanced immune processes co-contribute to fluorouracil beneficial responses in gastric cancer. Front. Immunol.13, 999551. 10.3389/fimmu.2022.999551 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.He, X. et al. Gastric Cancer actively remodels mechanical microenvironment to promote Chemotherapy Resistance via MSCs-Mediated mitochondrial transfer. Advanced science (Weinheim. Baden-Wurttemberg Germany). 11(47), e2404994. 10.1002/advs.202404994 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Jung, B. K. et al. Relaxin-expressing oncolytic adenovirus induces remodeling of physical and immunological aspects of cold tumor to potentiate PD-1 blockade. J. Immunother. Cancer. 8(2), e000763. 10.1136/jitc-2020-000763 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zhang, Z. & Zhu, Y. ANRGs impact on gastric cancer progression and drug efficacy: a comprehensive study. Medicine102(43), e34861. 10.1097/MD.0000000000034861 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ji, C. et al. AnnexinA5 promote glioma cell invasion and migration via the PI3K/Akt/NF-κB signaling pathway. J. Neurooncol.138(3), 469–478. 10.1007/s11060-018-2818-4 (2018). [DOI] [PubMed] [Google Scholar]
- 56.Ejtehadifar, M. et al. Meta-analysis of MS-based proteomics studies indicates interferon regulatory factor 4 and nucleobindin1 as potential prognostic and drug resistance biomarkers in diffuse large B cell lymphoma. Cells12(1), 196. 10.3390/cells12010196 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kang, T. H. et al. Annexin A5 as an immune checkpoint inhibitor and tumor-homing molecule for cancer treatment. Nat. Commun.11(1), 1137. 10.1038/s41467-020-14821-z (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Nowwarote, N. et al. Extracellular matrix derived from dental pulp stem cells promotes mineralization. Front. Bioeng. Biotechnol.910.3389/fbioe.2021.740712 (2021). [DOI] [PMC free article] [PubMed]
- 59.Li, D. et al. Cancer-associated fibroblast-secreted IGFBP7 promotes gastric cancer by enhancing tumor associated macrophage infiltration via FGF2/FGFR1/PI3K/AKT axis. Cell. Death Discovery. 9(1), 17. 10.1038/s41420-023-01336-x (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Li, J., Pu, K., Li, C., Wang, Y. & Zhou, Y. A. Novel six-gene-based Prognostic Model predicts survival and clinical risk score for gastric Cancer. Front. Genet.12, 615834. 10.3389/fgene.2021.615834 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Xu, Q. et al. Pan-cancer analyses reveal oncogenic and immunological role of PLOD2. Front. Genet.1310.3389/fgene.2022.864655 (2022). [DOI] [PMC free article] [PubMed]
- 62.Du, H. et al. PLOD2 regulated by transcription factor FOXA1 promotes metastasis in NSCLC. Cell Death Dis.8(10). 10.1038/cddis.2017.553 (2017). e3143. [DOI] [PMC free article] [PubMed]
- 63.Dou, N. et al. SNRPA enhances tumour cell growth in gastric cancer through modulating NGF expression. Cell Prolif.51(5), e12484. 10.1111/cpr.12484 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Tan, Z., Kang, T., Zhang, X., Tong, Y. & Chen, S. Nerve growth factor prevents arsenic-induced toxicity in PC12 cells through the AKT/GSK-3β/NFAT pathway. J. Cell. Physiol.234(4), 4726–4738. 10.1002/jcp.27255 (2019). [DOI] [PubMed] [Google Scholar]
- 65.Jiang, T. et al. Development of small-molecule tropomyosin receptor kinase (TRK) inhibitors for NTRK fusion cancers. Acta Pharm. Sinica B. 11(2), 355–372. 10.1016/j.apsb.2020.05.004 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Hayakawa, Y. et al. Nerve growth factor promotes gastric tumorigenesis through aberrant Cholinergic Signaling. Cancer Cell.31(1), 21–34. 10.1016/j.ccell.2016.11.005 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Posner, M. G. Multimerin-1 and cancer: a review. Biosci. Rep.42(2), BSR20211248. 10.1042/BSR20211248 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Tatti, O. et al. MMP16 mediates a Proteolytic switch to promote cell-cell adhesion, Collagen Alignment, and Lymphatic Invasion in Melanoma. Cancer Res.75(10), 2083–2094. 10.1158/0008-5472.CAN-14-1923 (2015). [DOI] [PubMed] [Google Scholar]
- 69.Lowy, A. M. et al. beta-Catenin/Wnt signaling regulates expression of the membrane type 3 matrix metalloproteinase in gastric cancer. Cancer Res.66(9), 4734–4741. 10.1158/0008-5472.CAN-05-4268 (2006). [DOI] [PubMed] [Google Scholar]
- 70.Zhao, Q. et al. Increased IGFBP7 expression correlates with poor prognosis and immune infiltration in gastric cancer. J. Cancer. 12(5), 1343–1355. 10.7150/jca.50370 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Su, Z., Shu, K. & Li, G. Increased ANXA5 expression in stomach adenocarcinoma infers a poor prognosis and high level of immune infiltration. Cancer Biomark. A. 35(2), 155–165. 10.3233/CBM-210482 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Li, S. S., Lian, Y. F., Huang, Y. L., Huang, Y. H. & Xiao, J. Overexpressing PLOD family genes predict poor prognosis in gastric cancer. J. Cancer. 11(1), 121–131. 10.7150/jca.35763 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Peng, B., Guo, C., Guan, H., Liu, S. & Sun, M. Z. Annexin A5 as a potential marker in tumors. Clin. Chim. Acta. 427, 42–48. 10.1016/j.cca.2013.09.048 (2014). [DOI] [PubMed] [Google Scholar]
- 74.Xue, Z. et al. Mesenchymal stem cells promote epithelial to mesenchymal transition and metastasis in gastric cancer though paracrine cues and close physical contact. J. Cell. Biochem.116(4), 618–627. 10.1002/jcb.25013 (2015). [DOI] [PubMed] [Google Scholar]
- 75.Oh, S. C. et al. Clinical and genomic landscape of gastric cancer with a mesenchymal phenotype. Nat. Commun.9(1), 1777. 10.1038/s41467-018-04179-8 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Zhao, R. et al. AQP5 complements LGR5 to determine the fates of gastric cancer stem cells through regulating ULK1 ubiquitination. J. Experimental Clin. Cancer Research: CR. 41(1), 322. 10.1186/s13046-022-02532-w (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Maeser, D., Gruener, R. F. & Huang, R. S. OncoPredict: an R package for predicting in vivo or cancer patient drug response and biomarkers from cell line screening data. Brief. Bioinform.22(6), bbab260. 10.1093/bib/bbab260 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Uhlén, M. et al. Proteomics. Tissue-based map of the human proteome. Sci. (New York NY). 347(6220), 1260419. 10.1126/science.1260419 (2015). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Publicly available datasets were analyzed in this study. These data can be found here: GDC Data Portal of TCGA (https://portal.gdc.cancer.gov/); GEO datasets (https:// www.ncbi.nlm.nih.gov/geo/): GSE26942 and GSE184198.