

Abstract
Correct mitotic spindle alignment is essential for tissue architecture and plays an important role in cell fate specification through asymmetric cell division. Spindle tethering factors such as Drosophila Mud (NuMA in mammals) are recruited to the cell cortex and capture astral microtubules, pulling the spindle in the correct orientation. However, how spindle tethering complexes read the cell polarity axis and how spindle attachment is coupled to mitotic progression remains poorly understood. We explore these questions in Drosophila sensory organ precursors (SOPs), which divide asymmetrically to give rise to epidermal mechanosensory bristles. We show that the scaffold protein Meru, which is enriched at the posterior cortex by the Frizzled/Dishevelled planar cell polarity complex, in turn recruits Mud, linking the spindle tethering and polarity machineries. Furthermore, Cyclin A/Cdk1 associates with Meru at the posterior cortex, promoting the formation of the Mud/Meru/Dsh complex via Meru and Dsh phosphorylation. Thus, Meru couples spindle orientation with cell polarity and provides a cell cycle-dependent cue for spindle tethering.
Keywords: Spindle Orientation, Asymmetric Cell Division, Cell Polarity, Drosophila, Development
Subject terms: Cell Adhesion, Polarity & Cytoskeleton; Cell Cycle; Development
Synopsis

How spindle-tethering complexes read the cell polarity axis during asymmetric cell division and couple spindle attachment to mitotic progression remains poorly understood. Here, the scaffold protein Meru is found to provide spatial and temporal cues for mitotic spindle orientation by coupling spindle tethering with cell polarity.
Correct spatial orientation of the mitotic spindle is essential for cell fate determination and tissue integrity.
The scaffold protein Meru is required for spindle orientation of Drosophila sensory organ precursor cells.
Meru receives spatial inputs from the planar cell polarity machinery via Dishevelled and in turn recruits the spindle-tethering factor Mushroom body defect.
Meru senses cell-cycle progression by recruiting Cyclin A to the cell cortex, leading to Meru phosphorylation by Cdk1.
Cdk1 phosphorylation of Meru increases its ability to bind Mud, allowing timely Mud cortical recruitment.
The scaffold protein Meru provides spatial and temporal cues by coupling spindle tethering with cell polarity, and via cell-cycle-dependent input from Cyclin A/Cdk1 during asymmetric cell division.
Introduction
Accurate mitotic spindle orientation is key to the development and adult homoeostasis of multicellular organisms. In symmetrically dividing epithelial cells, the mitotic spindle generally aligns parallel to the tissue plane, allowing the daughter cells to insert themselves seamlessly into the epithelium, thereby maintaining tissue architecture and integrity (Bergstralh et al, 2017; Ragkousi and Gibson, 2014). Correct spindle orientation is also essential for the generation of cell diversity through the process of asymmetric cell division (ACD). During ACD, cell fate determinants (CFDs) are unequally inherited by the daughter cells, promoting progenitor self-renewal or differentiation (Sunchu and Cabernard, 2020). CFD asymmetric segregation depends upon the polarisation of the cell cortex, which directs spindle alignment and ensures unequal CFD inheritance. Thus, cell polarity and spindle orientation must be tightly linked, and aberrant alignment can result in developmental defects and neoplasia (Bergstralh et al, 2017; Ragkousi and Gibson, 2014; Sunchu and Cabernard, 2020).
Mushroom body defective (Mud; Nuclear Mitotic Apparatus Protein/NuMA in mammals; LIN-5 in C. elegans) is a conserved spindle tethering factor that is recruited to the cell cortex by cell polarity proteins and exerts forces on astral microtubules by binding the Dynein/Dynactin motor complex (di Pietro et al, 2016). A core complex comprising the Mud binding partner Pins (Partner of inscrutable)/LGN and Gαi has been implicated in Mud cortical recruitment (Bergstralh et al, 2017; Morin and Bellaïche, 2011; Siller and Doe, 2009). However, how Pins itself is recruited to the correct subcellular location and whether it is even required for Mud localisation is clearly context-dependent. For instance, Drosophila Mud localises to the cortex in a Pins-dependent manner in the ovarian follicular epithelium and via a Pins-independent mechanism in the wing and thoracic epithelia (Bergstralh et al, 2016; Bosveld et al, 2016; Nakajima et al, 2013, 2019; Neville et al, 2023). Thus, how the spatial alignment of the mitotic spindle adapts to the polarity of different symmetrically or asymmetrically dividing cell types remains unclear. Recent work has also proposed that, as well as being cortically recruited, the Mud/NuMA spindle tethering complex must be activated as the cells enter mitosis (Neville et al, 2023). However, our understanding of how the spindle tethering machinery is coupled to cell cycle progression remains limited. Here, we use the asymmetric division of Drosophila sensory organ precursor cells (SOPs) as a model to study these questions.
SOPs (also known as pI), which give rise to the adult sensory bristles of the fly epidermis, are a well-studied example of ACD (Hartenstein and Posakony, 1989; Schweisguth, 2015). The best-studied SOPs are located on the dorsal thorax (notum) and produce the mechanosensory microchaetes that cover this tissue (Hartenstein and Posakony, 1989; Schweisguth, 2015). SOPs undergo consecutive rounds of asymmetric division to produce five distinct cell types: neuron, sheath, shaft, socket and a glial cell that undergoes apoptosis (Hartenstein and Posakony, 1989). The SOP lineage cells then recruit a neighbouring epidermal cell (the F-Cell) and together these assemble into a functional sensory hair (Mangione et al, 2023). In the notum, SOPs are specified at ~12 h APF (hours After Puparium Formation) from the epithelial sheet through the highly conserved Notch pathway (Corson et al, 2017; Gómez-Skarmeta et al, 2003; Simpson, 2007).
Until the SOPs are specified, they share the same polarity as the surrounding epithelial cells, characterised by two polarity axes: (1) planar cell polarity (PCP), whereby the cells align in the tissue plane and (2) apical-basal (A–B) polarity which defines the cellular apical and basal domains separated by the adherens junctions (AJs) (Buckley and St Johnston, 2022; Goodrich and Strutt, 2011). The core PCP pathway is established by three transmembrane proteins that form opposing domains at the AJs through mutual antagonism (Goodrich and Strutt, 2011; Yang and Mlodzik, 2015). In the notum, heterodimers of the seven-pass transmembrane protein Flamingo (Fmi, also known as Starry night/Stan; CELSR1/2/3 in vertebrates) with the four-pass transmembrane protein Van Gogh (Vang, also known as Strabismus; Vang1/2 in vertebrates) are located on the anterior side of each cell, while heterodimers of Fmi with the seven-pass protein Frizzled (Fz; Fz1 in vertebrates) are on the posterior side (Bellaïche et al, 2001; Schweisguth, 2015; Ségalen et al, 2010). Vang then recruits its downstream effector Prickle (Pk) (Bastock et al, 2003; Jenny et al, 2003; Taylor et al, 1998), while Fz recruits Dishevelled (Dsh; DVL1-3 in vertebrates) and Diego (Dgo) (Axelrod et al, 1998; Feiguin et al, 2001; Jenny et al, 2005). In A–B polarity, the apical domain is defined by the Par complex, comprised of Bazooka (Baz; Par3 in vertebrates), atypical Protein Kinase C (aPKC; PKCι/PKCζ in vertebrates) and Par6 which regulates the placement of the AJs (Bilder et al, 2000; Buckley and St Johnston, 2022; Tepass, 2012). The basolateral domain is defined by the septate junction components Scribble (Scrib; SCRIB in vertebrates), Discs large (Dlg; Dlg1-5 in vertebrates) and Lethal giant larvae (Lgl; LLGL1/2 in vertebrates) (Albertson et al, 2004; Bilder et al, 2000; Woods and Bryant, 1991).
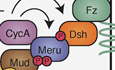
Upon SOP specification, the proneural transcription factors of the Achaete-Scute complex turn on the expression of the N-terminal RASSF (Ras-association domain family) protein Meru (Banerjee et al, 2017; Buffin and Gho, 2010; Reeves and Posakony, 2005). Meru is enriched to the posterior cortex by Dsh, and in turn recruits Baz, leading to its planar asymmetry, specifically in the SOP (Banerjee et al, 2017). Baz then polarises the rest of the Par complex to the posterior cortex, leading to the exclusion of the CFDs Numb and Neuralized (both Notch pathway modulators) from the posterior pole prior to ACD (Bellaïche et al, 2001; Bellaïche et al, 2001; Besson et al, 2015; Le Borgne and Schweisguth, 2003; Roegiers et al, 2001a, 2001b; Smith et al, 2007; Wirtz-Peitz et al, 2008). Thus, Meru provides the initial planar bias that triggers the polarisation of the Par complex and the CFDs along the antero-posterior axis (Banerjee et al, 2017). To ensure accurate segregation of CFDs, the mitotic spindle must also align along the antero-posterior axis. This is achieved by redundant spindle tethering complexes on the anterior and posterior poles (Gomes et al, 2009; Schweisguth, 2015) (Fig. 1A). On the anterior side, Gαi/Dlg/Pins recruit Mud basally, while on the posterior side, Mud is recruited in a Fz/Dsh-dependent manner to the apical cortex (Bellaïche et al, 2001; Bellaïche et al, 2001; David et al, 2005; Johnston et al, 2013; Schaefer et al, 2000; Ségalen et al, 2010). This allows the capture of one centrosome each by the anterior and posterior poles, but also imparts a characteristic A–B tilt to the spindle (David et al, 2005; Schweisguth, 2015; Ségalen et al, 2010) (Fig. 1A). Thus, SOP division is a powerful system to study how spindle orientation adapts to cellular context, since two distinct Mud localisation mechanisms co-exist in this cell type.
Figure 1. Meru localises to the apical–posterior domain of the SOP.

(A) The current model of SOP spindle alignment where Mud is recruited to the anterior cortex by Pins/Gαi and to the posterior cortex through Fz/Dsh. Note that the extent of spindle tilt in the diagram is amplified compared with reality for illustrative purposes. (B, C) Brightfield images of the adult notum (B), wing (C), and anterior wing margin (C’) in animals expressing UAS-mK2-meru under the neurG4 driver. No visible defects (arrows indicating micro/macrochaetes) in the notum and wing sensory organs were observed. (D–F) Maximum intensity projections of pupal nota at 16 h APF of neurG4 > UAS-mK2-meru (magenta, D–F), Ecad-GFP (composite of Fig. EV1A–A’), Pon-GFP and Mud-GFP (green; D, E’ and F’, respectively) at 18 min prior to first indication of cytokinesis. SOP marked by a star. Scale bars = 5 µm. (G) Single brightest slice in (F, F’) plotted as line graph by measuring the grey value across the posterior domain as defined by Meru localisation normalised to the highest value in each channel. High Pearson’s coefficient (r) indicates a positive correlation. P value calculated using a two-tailed test. Source data are available online for this figure.
An important open question remains how Mud is recruited to the posterior cortex as the SOPs enter mitosis. Two potential mechanisms have been proposed: (1) Mud binds directly to Dsh (Ségalen et al, 2010) and (2) the mitotic cyclin, Cyclin A (CycA), is recruited to the posterior cortex in a Dsh-dependent manner and in turn recruits Mud (Darnat et al, 2022). Here, we show that, in parallel to its role in posterior Baz recruitment, Meru acts as a bridging factor between Dsh and Mud. Examination of the Meru sequence revealed a CycA docking site that is required for Meru cortical localisation, Mud localisation and spindle alignment. We also identified multiple Cdk1 (Cyclin-dependent kinase 1) phosphorylation consensus sites that promote the Meru/Mud interaction. Meru therefore links PCP with spindle tethering, and its ability to recruit Mud is coupled to the cell cycle through CycA to ensure timely alignment of the spindle during mitosis.
Results
Meru localises to the apical–posterior pole prior to and during SOP mitosis
To address the potential role of Meru in SOP spindle alignment, we generated a UAS-mKate2(mK2)-meru transgenic line to track its localisation during SOP divisions relative to known polarity and spindle factors. We had previously shown that endogenously tagged GFP-Meru localises to the posterior cortex in late G2, where it remains enriched throughout mitosis and colocalises with Dsh (Banerjee et al, 2017). No bristle defects were observed in animals that had mK2-Meru driven in SOPs by the neurP72-GAL4 driver (neurG4) (Bellaïche et al, 2001), suggesting overexpression does not lead to strong SOP division defects (Fig. 1B–C’). mK2-Meru localised to the posterior–apical cortex at the level of E-cadherin (Ecad) as expected (Banerjee et al, 2017), (Figs. 1D and EV1A,A’). Upon mitotic rounding, Partner of Numb (Pon), the adaptor of the CFD Numb, is rapidly relocalised to the basolateral anterior cortex during prophase, followed by its unequal inheritance into the anterior pIIb daughter cells (Bellaïche et al, 2001; Roegiers et al, 2001b). Co-expression of mK2-Meru and Pon showed that mK2-Meru was present at the posterior domain before the asymmetric localisation of Pon was detectable, indicating SOP polarisation occurs prior to CFD segregation (Fig. 1E,E’). Thus, Meru is localised at the posterior–apical cortex during most of SOP mitosis, consistent with a possible role in spindle positioning.
Figure EV1. Posterior–apical localisation of Meru occurs prior to SOP mitosis and persists throughout division.

(A-B’) Maximum intensity projections of apical-most three sections of pupal nota live-imaged on a confocal microscope at 16 h APF, 0 min marking the first frame indicating cytokinesis, of neurG4 > UAS-mK2-meru (magenta; A, B, see composite in Fig. 1D), co-expressed with Ecad-GFP and Mud-GFP (green; A’, B’, respectively). (C–C”) Single brightest slice in (B, B’) plotted as line graphs by measuring the grey value across the posterior domain as defined by Meru localisation normalised to highest value in each channel. High Pearson’s coefficient (r) indicates positive correlation. P values calculated using a two-tailed test. Scale bars = 5 µm. Source data are available online for this figure.
Meru promotes Mud recruitment to the posterior cortex
Dsh and Mud have been shown to colocalise at the posterior–apical cortex during SOP mitosis (Ségalen et al, 2010). We reported that Meru and Dsh associate in cell culture, colocalise in fixed tissue, and that Dsh is required for Meru posterior localisation (Banerjee et al, 2017). We therefore sought to test whether there was an interaction between Meru and Mud by tracking the localisation of mK2-Meru and a GFP-tagged Mud construct expressed under the mud promoter (GFP-Mud) with live-imaging (Ségalen et al, 2010). Mud and Meru localisations were highly correlated (as indicated by a Pearson’s coefficient r near +1), particularly at the onset of mitosis (Fig. 1F,G). Despite a considerable drop in Mud intensity levels, the two proteins remained positively correlated even after metaphase (Figs. EV1B-C”).
The strong Meru/Mud colocalisation is consistent with a physical interaction between these proteins. To test this possibility, we performed co-immunoprecipitations (co-IPs) from S2 cell lysates expressing full-length Meru together with Mud fragments spanning much of the protein (Ségalen et al, 2010) (Fig. 2A,B). We found that Meru binds to the C-terminal part of Mud (1825–2456), a region which is largely disordered, but contains a short domain that mediates Pins binding, known as Pins-Binding Domain (PBD—aa 1825–1961) and a transmembrane-like (TML) domain. The PBD has been shown to recruit Mud cortically in SOPs and neuroblasts (Ségalen et al, 2010; Siller et al, 2006). As Mud (1452–1961) which contains the PBD binds Meru, but Mud (1452–1824) which lacks the PBD does not, the PBD represents one interaction surface for the Meru/Mud interaction (Fig. 2B). Moreover, two truncations lacking the PBD, Mud (1951–2456) and Mud (2089–2499) can both co-immunoprecipitate Meru, though not as efficiently as Mud (1825–2456), which has both the PBD and C-terminus (Fig. 2B). This suggests that Meru/Mud association is mediated both by the Mud PBD and C-terminus (aa 1951–2456).
Figure 2. Meru is required for Dsh/Mud complex formation.

(A) Schematic of constructs that span Meru, Dsh and Mud used in co-IPs to characterise Meru binding in S2 cells. RA Ras-association domain, DIX DIshevelled and aXin domain, PDZ = PSD96, Dlg, ZO-1 domain, DEP Dishevelled, Egl-10 and Pleckstrin domain, CH Calponin Homology domain, PBD Pins-Binding Domain, TML Trans-Membrane-Like domain. (B–D) Western blots of co-IP experiment using S2 cell lysates from transfected S2 cells, immunoprecipitated using and probed with the indicated antibodies. (B) Full-length Meru immunoprecipitates with Mud fragments containing the PBD and C-terminus. (C) Mud does not co-IP with full-length Dsh. The Meru/Mud interaction is used as a positive control (right-most lane). (D) Meru promotes Dsh/Mud complex formation. Source data are available online for this figure.
A previous report showed an interaction between Mud and Dsh in HEK293T cells (Ségalen et al, 2010). We were unable to detect such an interaction in S2 cells between full-length Dsh and any of the Mud fragments (Fig. 2C). However, Meru is not endogenously expressed in S2 cells, whereas the orthologs of Meru (RASSF9/10) are expressed in HEK293T cells (Hauri et al, 2013). Together with our finding that Meru interacts with both Dsh and Mud in cell culture, this prompted us to test whether Meru could bridge the previously reported Dsh/Mud interaction (Ségalen et al, 2010). Indeed, co-expression of Meru elicited a robust association between Dsh and Mud (1452–1961) (Fig. 2D). Thus, Meru colocalises with Mud and Dsh at the posterior cortex during SOP cell divisions and mediates the assembly of a Mud/Meru/Dsh complex in cell culture.
Meru promotes cortical Mud localisation and proper spindle orientation in vivo
In wild-type SOPs and epithelial cells, Mud localises to three regions: at the cortex (with polarity proteins), at the centrosomes, and at tricellular junctions (Bosveld et al, 2016; Izumi et al, 2006; Ségalen et al, 2010). Specifically, Mud localisation to the posterior cortex in SOPs is dependent on Dsh (Ségalen et al, 2010). As Meru is required for the Dsh/Mud interaction in cell culture, we tested whether posterior cortical Mud recruitment is dependent on Meru in vivo. We quantified Mud-GFP at both the anterior and posterior cortex in meru1 mutants (a 1.6 kb deletion that removes half the coding region, including the RA domain (Banerjee et al, 2017)) compared to wild-type flies (Fig. 3A–C). We noted that, in wild-type SOPs, Mud crescent intensity is consistently higher on the anterior side than the posterior (Fig. 3C). We observed a significant decrease in Mud intensity at the posterior cortex of meru1 mutant SOPs relative to the wild-type (Fig. 3C). However, anterior cortical Mud levels were also decreased, potentially indicating a role for Meru in Mud stability (Fig. 3C). Thus, Meru promotes Mud cortical recruitment, consistent with a role in spindle alignment.
Figure 3. Meru loss leads to Mud mislocalisation and spindle alignment defects.

(A, B) Pupal notal confocal live-imaging of Mud-GFP (green, max projection of 6 slices) and UAS-cd8-mRFP driven by neurG4 (magenta, max projection of 4 slices) in a meru wild-type (A) and meru1 background (B) at 16 h APF at the first indication of cytokinesis. (C) Graph showing the intensity of cortical Mud at the anterior and posterior crescent of each genotype. Measurements were taken at anaphase. Both the anterior and posterior cortical Mud levels were significantly lower in the meru1 flies relative to the wild-type (P = 1.4 × 10−6 and P = 1.4 × 10−6, respectively). ****P < 0.0001 using a Mann–Whitney U test. Large and small dashed line represent the median and Q1/Q3, respectively. (D, E) Confocal live-imaging of neur-H2B-RFP (magenta) and Jupiter-GFP (green) in a meru wild-type (D) and meru1 background at approximate cytokinesis (E). (F) Graphs showing the spindle orientation of each genotype relative to the dorsal midline (F) and epithelial plane (F’). P values indicated using a Kolmogorov–Smirnov test. Scale bars = 10 µm. Number of SOPs imaged indicated on the panels from three nota in (C) or six wild-type or 4 meru1 nota, respectively, in (F, F’). Source data are available online for this figure.
We previously showed that meru1 mutant SOPs display cell division axis defects (Banerjee et al, 2017). However, these measurements were performed in the presence of overexpressed pon, which is known to induce changes in SOP polarity (Perdigoto et al, 2008). We therefore directly measured spindle alignment by labelling the spindle using a GFP-tagged version of the microtubule-binding protein Jupiter (Jup) and quantifying the deviation from the A–P axis in wild-type and meru1 mutant flies (Fig. 3D–F’; Movies EV1 and EV2). As the mitotic spindle remains dynamic until metaphase, when it settles for its final division angle (Bellaïche et al, 2001; Bergstralh et al, 2016; Ségalen et al, 2010), our measurements were carried out post-metaphase. Consistent with previous work (Ségalen et al, 2010), 71% of wild-type cells divided within 30° of the A–P axis (Fig. 3D,F), and the spindle alignment in the A–B axis averaged at 6.8°, displaying the characteristic z-tilt of SOP mitotic spindles (David et al, 2005; Ségalen et al, 2010) (Fig. 3F’). Strikingly, meru1 flies had a nearly random spindle alignment in the A–P axis, with only 45% of cells dividing within 30° of the midline (Fig. 3E,F). The average A–B angle showed a slight increase to 9.0°, with a significantly wider distribution—nearly 20% of cells dividing over 20°, compared to 2% in controls (Fig. 3F’). Ségalen et al observed that the spindle is more planar in fz and dsh mutants (Ségalen et al, 2010) rather than more tilted in the A–B axis as it is in meru mutants (Fig. 3F’) or upon cycA depletion (Darnat et al, 2022). The likely explanation is that the authors analysed tissues that are uniformly mutant for fz and dsh. Because the anterior and posterior PCP complexes stabilise each other across cell junctions as well as antagonise each other intracellularly (Goodrich and Strutt, 2011; Yang and Mlodzik, 2015), global loss of dsh or fz perturbs the organisation of both the anterior and posterior cortices in SOPs which no longer receive PCP input from their neighbours on the anterior side. In these conditions, the cell divisions become planar since spindle tilt is lost (Ségalen et al, 2010). As depletion of meru and cycA do not affect PCP in neighbouring cells, the basal pulling force from Pins/Mud remains and is no longer counterbalanced by the apical–posterior side and spindle tilt increases (Fig. 3F’ and (Darnat et al, 2022)). Consistent with its role in Mud positioning, Meru is therefore required for A–P and planar spindle alignment.
Meru associates with CycA
CycA has recently been reported to be enriched at the apical–posterior cortex at the end of G2 and early prophase in SOPs (Darnat et al, 2022). We therefore wished to investigate whether CycA could provide a link between cell cycle progression and spindle orientation by functioning in concert with Meru. We first tracked Meru/CycA colocalisation in vivo. In late G2 phase, when both proteins have been reported to be present at the posterior cortex during SOP mitosis (Banerjee et al, 2017; Darnat et al, 2022), we observed that the localisations of mK2-Meru and endogenously tagged CycA were highly correlated (Fig. 4A,B—Pearson’s coefficient = 0.71). The colocalisation persisted until metaphase (Fig. 4A), when CycA enters the nucleus, which is followed by its degradation (Lehner and O’Farrell, 1989). We then tested a potential Meru/CycA association by co-IP in S2 cells. Interestingly, CycA robustly immunoprecipitated with Meru, but not Dsh or Mud (Fig. 4C). To determine whether the Meru/CycA interaction is specific, we examined binding to other Cyclins associated with cell cycle progression (A, B, E and D). We observed an interaction with CycB, which is expressed during early M-phase (Lehner and O’Farrell, 1990). However, this CycB association was far weaker than the interaction with CycA (Fig. 4D). Thus, Meru binds to CycA, with which it colocalises at the posterior cortex in SOPs.
Figure 4. Meru and CycA interact at the posterior cortex of SOPs.

(A) Confocal live-imaging of pupal nota (apical-most three sections) expressing neurG4 > UAS-mK2-meru (magenta) and CycA-GFP (green) of an SOP during mitosis at 16 h APF and 0 min indicating the first frame in cytokinesis. (B) Single brightest slice in (A, A’) at −20 min plotted as line graph by measuring the grey value across the posterior domain as defined by Meru localisation. High Pearson’s coefficient (r) indicates a positive correlation. P value calculated using a two-tailed test. (C, D) Western blots of co-IP experiment using cell lysates from transfected S2 cells, immunoprecipitated using α-GFP beads and probed using α-FLAG and α-GFP antibodies. (C) Meru, but not Dsh or C-terminal Mud (1452–1961), immunoprecipitates with CycA. (D) Meru associates with CycA and weakly with CycB, but not CycD or CycE. Scale bar = 10 µm. Source data are available online for this figure.
CycA docking regulates Meru localisation and association with Mud and Dsh
CycA binding is highly regulated through short linear motifs (or SLiMs), the most well-known of which is the RxL motif. This consensus sequence consists of amino acids R/K-x-L-φ or R/K-x-L-x–φ (φ = hydrophobic amino acid) (Tatum and Endicott, 2020) (Fig. EV2A). When we investigated the Meru sequence, we found six possible RxL motifs, two of which (RxL 258 and 259) were very good matches and located immediately adjacent to each other (Figs. 5A and EV2A). We generated mutations at all six RxL motifs, substituting the R/K and L to A. We initially found that Meru/CycA binding was only affected in the RxL 258/9 double mutant (Fig. EV2B). Subsequent mutation of either the 258 or 259 site alone showed that this was sufficient to disrupt the Meru/CycA interaction (Fig. 5B). In all subsequent experiments, we used the double 258/9 mutants (meruRxL).
Figure EV2. The RxL motifs of Meru.

(A) The amino acid (aa) positions of the six RxL motifs identified in the Meru sequence are indicated on the left. On the right is the sequence alignment of all RxL motifs. Orange indicates the R/K and the L is labelled in blue at position 0. The hydrophobic aa is marked in purple and the grey boxes indicate the two aa that were mutated to Alanine in RxL mutants. (B, C) Western blots of co-IP experiment using cell lysates from transfected S2 cells, immunoprecipitated and probed using the indicated antibodies. (B) CycA immunoprecipitates with each Meru RxL motif mutant, except when Meru 258/9 is mutated to Alanines. (C) CycA depletion by RNAi reduced Meru/Mud association. Source data are available online for this figure.
Figure 5. CycA docking is required for Meru localisation in vivo.

(A) Schematic of RxL motifs and Cdk1 S/T-P phosphorylation sites present in the Meru sequence. (B) Meru mutated at RxL motifs 258, 259 and 258/9 no longer immunoprecipitates with CycA. (B–E) Western blots of co-IP experiment using cell lysates from transfected S2 cells, immunoprecipitated and probed using the indicated antibodies. Mutation of Meru RxL 258 and 259 sites disrupt the interaction between Meru and CycA (B), Mud (C) and Dsh (D), but not Baz (E). (F) Maximum intensity projections from confocal live-imaging of pupal nota at 16 h APF (0 min marking first frame in cytokinesis) show expression of UAS-mK2-meru (magenta) driven by neurG4 and CycA-GFP (green) during SOP mitosis. Scale bar = 10 µm. (G) Graphs showing the intensity of apical–posterior CycA crescent in MeruWΤ and MeruRxL animals at late G2. CycA cortical levels were significantly lower in MeruRxL relative to MeruWΤ cells (P = 2.6 × 10−5). Large and small dashed line represent the median and Q1/Q3, respectively. ****P < 0.0001 using a Mann–Whitney U test. Number of SOPs imaged indicated on the panels from three nota. Source data are available online for this figure.
To determine if CycA binding modulates Meru function, we tested if the RxL 258/9 motif was required for association with other Meru partners. We observed that the Meru RxL motif is necessary for Mud and Dsh binding (Fig. 5C,D), but dispensable for binding to Baz (Fig. 5E). To distinguish between the RxL mutation directly interfering with Mud binding versus Meru/CycA binding being required for Meru to associate with Mud, we depleted CycA by RNAi (Fig. EV2C). CycA expression was required for the Meru/Mud association, consistent with the RxL mutation interfering with Meru/Mud binding indirectly by compromising CycA docking to Meru.
We then generated UAS-meruRxL transgenic flies to test the role of Meru/CycA binding in vivo. Strikingly, mK2-MeruRxL failed to be recruited to the cortex, in agreement with the RxL mutation compromising Dsh binding, and we noted that, after nuclear envelope reassembly, some mK2-MeruRxL accumulated in the nucleus (Figs. 5F and EV3A,A’). Co-expression with Ecad and Dlg showed that mK2-MeruRxL was distributed in both the apical and basal cytoplasm (Fig. EV3B,C’). When we expressed mK2-MeruRxL in a meru1 mutant background, we also observed a decreased CycA posterior crescent compared with MeruWT-expressing animals (Figs. 5F’,G and EV3D,D’), in agreement with the loss of binding demonstrated in Fig. 5B. In summary, CycA docking to Meru via the RxL motif is required for Dsh and Mud binding in cell culture, as well as Meru and CycA cortical localisation in vivo.
Figure EV3. 258/9 RxL motif mutant Meru no longer binds CycA and is mislocalised in vivo.

(A–D) Maximum intensity projections of pupal nota live-imaging at 16 h APF, 0 min marking the first frame indicating cytokinesis, of neurG4 > UAS-mK2-meruRxL and neurG4 > UAS-mK2-meruWT (magenta; A–C and D, respectively), co-expressed with nuclear nls-GFP, apical Ecad-GFP, basolateral Dlg-GFP and CycA-GFP (green; panels A’, B’, C’ and D’, respectively). Scale bars = 10 µm. Source data are available online for this figure.
CycA docking is required for Meru-dependent spindle orientation
We sought to address if the role of Meru in SOP spindle alignment was regulated by CycA. We first tested the effect of blocking Meru/CycA association on the formation of the ternary Dsh/Meru/Mud complex in S2 cells. In contrast to wild-type Meru, the MeruRxL mutant was unable to bridge the Dsh/Mud interaction (Fig. 6A), suggesting that CycA binding to Meru is required for ternary complex formation. Second, we tested if loss of CycA binding impacted Meru’s role in spindle orientation in vivo. We performed a rescue experiment by expressing UAS-mK2-meru constructs in a meru1 mutant background and quantified cortical Mud localisation as in wild-type SOPs above (Fig. 6B–D). In the rescued conditions, both the anterior and posterior cortices had nearly identical levels of Mud when mK2-MeruWT was expressed (Fig. 6B,B’,D). It is interesting to note that, in the absence of Meru overexpression, Mud is more enriched at the anterior than the posterior cortex (Fig. 3C). The equal cortical distribution of Mud upon mK2-MeruWT overexpression is therefore consistent with Meru promoting Mud posterior recruitment. In contrast, mK2-MeruRxL-expressing SOPs had reduced Mud levels at the posterior cortex (Fig. 6C,C’,D), suggesting that CycA binding of Meru is necessary for posterior Mud localisation. We also note that, while MeruWT is enriched at the posterior cortex (Fig. 6B’), we consistently detect some Meru at the anterior cortex (see Fig. 4A for another example). This could be a consequence of overexpression or of the way in which Meru is recruited to the plasma membrane (see “Discussion” for details).
Figure 6. Mutation of the Meru RxL motif results in decreased posterior Mud and abnormal spindle alignment.

(A) Western blot of co-IP experiment using cell lysates from transfected S2 cells, immunoprecipitated and probed using the indicated antibodies. Dsh does not immunoprecipitate with C-terminal Mud in the presence of MeruRxL. (B, C) Maximum intensity projections (6 slices) from confocal live-imaging of pupal nota at 16 h APF expressing Mud-GFP (green) and neurG4 > UAS-mK2-meruWT (B) or neurG4 > UAS-mK2-meruRxL (magenta) (C) in a meru1 background. (D) Graph showing the intensity of cortical Mud at the anterior and posterior crescent of each genotype. Measurements were taken at anaphase. Posterior MeruRxL is significantly lower than MeruWT (P = 1.9 × 10−8). ****P < 0.0001 using a Mann–Whitney U test. Large and small dashed line represent the median and Q1/Q3, respectively. (E, F) Maximum intensity projections from confocal live-imaging of neur-H2B-RFP (magenta) and Jupiter-GFP (green) in a neur > UAS-mK2-meruWT (E) and neur > UAS-mK2-meruRxL background (F). (G) Graphs showing the spindle orientation of each genotype relative to the dorsal midline (G) and epithelial plane (G’). P values indicated for Kolmogorov–Smirnov tests. Scale bars = 10 µm. Number of SOPs imaged from three animals indicated on the panels from three animal. Source data are available online for this figure.
Finally, we analysed the impact of impaired CycA binding on Meru-dependent spindle alignment. mK2-MeruWT expression was sufficient to rescue A–P spindle alignment in the meru1 mutant, however mK2-MeruRxL expression did not rescue A–P spindle angles (Fig. 6E–G), though A–B spindle alignment was not significantly affected (Fig. 6G’). As the A–P spindle defect in the MeruRxL-expressing animals (Fig. 6G) is less pronounced than in meru1 mutants (Fig. 3F), it is possible that some residual CycA binding occurs in this mutant through the other four RxL motifs when overexpressed in vivo. Thus, CycA docking on Meru is required for normal posterior Mud recruitment and consequently SOP mitotic spindle alignment.
Meru Cdk1 phosphorylation sites modulate binding to Mud and Dsh
The best characterised CycA function is Cdk1 regulation, thus we investigated whether Meru phosphorylation by Cdk1 could affect Meru/Mud association. Upon CycA/Cdk1 co-expression in S2 cells, we observed a marked increase in Meru/Mud binding compared to basal levels (Fig. EV4A). The minimal Cdk1 phosphorylation consensus site is S/T-P, while the optimal consensus is S/T-P-x-K/R (Enserink and Kolodner, 2010). We identified five potential phosphorylation sites within the Meru sequence (Fig. 5A). To determine whether these sites are indeed phosphorylated by Cdk1/CycA we carried out proteomic analysis by Mass Spectrometry (MS) to compare samples expressing either MeruWT or MeruRxL in the presence of Cdk1 and CycA. First, Meru, Dsh, CycA, Cdk1 and Mud proteins are all enriched in the MeruWT sample compared to the MeruRxL sample (Fig. EV4B), consistent with our previous data. Our analyses demonstrate the presence of phosphorylation on T496 and S60 in the MeruWT sample (Fig. 7B), the extracted spectrum for the T496 site is shown in Fig. 7A. In contrast, we failed to detect any phosphorylation in the MeruRxL sample, consistent with CycA/Cdk1 dependency of Meru phosphorylation of these sites (Fig. 7B).
Figure EV4. Overexpression of Cdk1 and CycA promotes the Meru/Mud interaction.

(A, C) Western blots of co-IP experiment using cell lysates from transfected S2 cells, immunoprecipitated and probed using the indicated antibodies. (A) Overexpression of Cdk1/CycA increases the amount of Meru binding to Mud. (B) MeruRxL has decreased interaction with CycA, Cdk1, Mud and Dsh relative to MeruWT in a peptide identification analysis by MS (N = 2). (C) No single Meru S/T-P phosphosite is essential to co-IP with C-terminal Mud. Source data are available online for this figure.
Figure 7. Cdk1 phosphorylation sites are required for Meru binding to Mud and phospho-Dsh.

(A) MS/MS spectrum corresponding to modification (phosphorylation) to the Meru T496 site. Fragment ions are indicated in red. Fragment ions with neutral loss are in yellow. (B) Phosphorylation of Meru at S60 and T496 was detected in MeruWT but not in the MeruRxL mutant. Label-free quantification (LFQ) intensities were plotted for each sample and phosphorylation site. (C, D) Western blots of co-IP experiment using cell lysates from transfected S2 cells, immunoprecipitated and probed using the indicated antibodies. Meru mutated to Alanine at five S/T-P phosphorylation sites disrupts co-IP with C-terminal Mud (1452–1961), but not Baz or Dsh (C), unless the Dsh T22 phosphorylation site was also mutated (D). Source data are available online for this figure.
In co-IPs, although several phosphosite mutants affected the Meru/Mud association, no single mutation was sufficient to abolish the interaction (Fig. EV4C). However, when all five sites were mutated (Meru5S/T-A) Meru/Mud binding was strongly reduced (Fig. 7C). In contrast, the Meru5S/T-A mutant still associated with both Baz and Dsh (Fig. 7C). This agrees with Meru not requiring a functional RxL site to interact with Baz (Fig. 5E). The Meru/Dsh interaction, on the other hand, is dependent on Meru’s RxL motif. Thus, we explored the possibility that Dsh is also undergoing Cdk1/CycA phosphorylation. Examining the Dsh sequence, we found a single Cdk1 phosphosite at amino acid 22 (T22) within the DIX (DIshevelled and aXin) domain (Fig. 2A). Interestingly, mutating this site alone (DshT22A) had no impact on binding to wild-type Meru (Fig. 7D). However, when the DshT22A mutant was combined with the Meru5S/T-A mutant, the association was almost completely lost (Fig. 7D). Taken together, this suggests that Meru phosphorylation by Cdk1 is required for Meru/Mud binding, while Meru/Dsh assembly is dependent on phosphorylation of both proteins.
Meru Cdk1 phosphorylation sites are required for proper spindle tethering
To evaluate the impact of Cdk1 phosphorylation of Meru on spindle alignment, we expressed Meru5S/T-A under neurG4 control in a meru1 background. First, we compared the localisation of Meru5S/T-A with MeruWT to the posterior cortex in late G2 (Fig. EV5A–B’). Although unlike MeruRxL (Fig. 5F), Meru5S/T-A could still form a detectable posterior crescent (Fig. EV5B,B’), this was significantly reduced compared with MeruWT (Fig. EV5A’–C). In addition, we observed a reduction in the characteristic Meru puncta that forms at the posterior crescent at this stage (Fig. EV5A’,B’,D). In contrast to meru1 mutants (Fig. 3B,C) and MeruRxL flies (Fig. 6C,D) Meru5S/T-A-expressing SOPs did not display a significant change in cortical Mud levels at anaphase (Fig. EV5H-J). However, there was a clear reduction in Mud at the posterior cortex in Meru5S/T-A animals at both prophase (Fig. EV5E–G) and metaphase (Fig. 8A–C), indicating a delay in Mud posterior recruitment. A–P spindle alignment was perturbed in the Meru5S/T-A mutants (Fig. 8D–F), while A–B alignment was not detectably affected (Fig. 8F’), as in MeruRxL animals (Fig. 6G’). Together, this suggests that Cdk1 phosphorylation of Meru is necessary to correctly orient the mitotic spindle in SOPs.
Figure EV5. Posterior cortical Mud levels return by anaphase in Meru5S/T-A mutants.

(A–B’) Single slice from confocal live-imaging of pupal nota at 16 h APF expressing Mud-GFP (green; A, B) and neurG4 > UAS-mK2-meruWT (magenta; A’) or neurG4 > UAS-mK2-meru5S/T-A (magenta; B’) at late G2. (C) Graph showing the mean intensity of posterior cortical Meru during late G2 in MeruWT and Meru5S/T-A (P = 2.7 × 10−3). (D) Graph displaying the average number of Meru puncta at the posterior cortex in late G2. Puncta of the Meru5S/T-A mutant are significantly decreased relative to MeruWT (P = 9.10 × 10−15). (E–F’) Maximum intensity projections (6 slices; E, F) from confocal live-imaging of pupal nota at 16 h APF expressing Mud-GFP (green; E, F) and neurG4 > UAS-mK2-meruWT (magenta; E’) or neurG4 > UAS-mK2-meru5S/T-A (magenta; F’) in a meru1 background at prophase. (G) Graph showing the intensity of cortical Mud at the anterior and posterior crescent of each genotype at prophase. Posterior Meru5S/T-A is significantly lower than MeruWT (P = 1.61 × 10−7). (H–I’) Maximum intensity projections (6 slices; H, I) from confocal live-imaging of pupal nota at 16 h APF expressing Mud-GFP (green; H, I) and neurG4 > UAS-mK2-meruWT (magenta; H’) or neurG4 > UAS-mK2-meru5S/T-A (magenta; I’) in a meru1 background at anaphase. (J) Graphs showing the intensity of cortical Mud at the anterior and posterior crescent of each genotype at anaphase, where there is no measurable difference at anaphase (P = 0.47). **P < 0.01 and ****P < 0.0001 using a Mann–Whitney U test. Large and small dashed lines in violin plots represent the median and Q1/Q3, respectively. Scale bars = 10 µm. (C, D, G, J) Number of SOPs imaged indicated on the panels from 3 nota. Source data are available online for this figure.
Figure 8. Meru Cdk1 phosphorylation sites promote Mud posterior recruitment and SOP spindle orientation.

(A, B) Maximum intensity projections (6 slices) of confocal live-imaging of pupal nota at 16 h APF expressing Mud-GFP (green) and neurG4 > UAS-mK2-meruWT (A) or neurG4 > UAS-mK2-meru5S/T-A (magenta) (B) in a meru1 background at metaphase (C) Graph showing the intensity of cortical Mud at the anterior and posterior crescent of each genotype. At metaphase, posterior Mud is significantly lower in Meru5S/T-A than MeruWT animals (P = 2.24 × 10−3). **P < 0.01 using a Mann–Whitney U test. Large and small dashed line represent the median and Q1/Q3, respectively. (D, E) Confocal live-imaging of neur-H2B-RFP (magenta) and Jupiter-GFP (green) in a neur > UAS-mK2-meruWT (D) and neur > UAS-mK2-meru5S/T-A background (E). (F) Graphs showing the spindle orientation of each genotype relative to the dorsal midline (F) and epithelial plane (F’). P values indicated for Kolmogorov–Smirnov tests. (C, F, F’) The number of SOPs imaged indicated on the panels from three nota. (G) Diagram showing our proposed model for Meru function in SOP spindle tethering. Meru, which is recruited to the posterior cortex of SOPs by Dsh, promotes posterior Mud recruitment. The Meru/Mud interaction is dependent on the docking of CycA to Meru at the 258/9 RxL motif and is regulated by Cdk1 phosphorylation of Meru. Mud and Dsh may also be Meru-dependent CycA/Cdk1 substrates at the SOP posterior cortex. Data information: Scale bars in (A, B, D, E) = 10 µm. Source data are available online for this figure.
The RRLL motif, a Cyclin-docking site shared by several spindle regulators
The cyclin-binding SLiM we identified in the Meru sequence is striking as it is comprised of two overlapping RxL motifs at amino acids 258/259 (Fig. EV2A). In addition, this site is followed by two optimal Cdk1 consensus phosphorylation sites at amino acids 267 and 274 (Fig. 5A). We used this motif to search for proteins containing overlapping RxLs separated by 7 amino acids from a Cdk1 target site ([RK]-[RK]-L-L-x[7]-[ST]-P, which we termed RRLL motif). Remarkably, we found that Pins, which recruits Mud to the anterior cortex in SOPs and in many other contexts (Bergstralh et al, 2017; di Pietro et al, 2016; Lechler and Mapelli, 2021), possesses a matching site (Fig. EV6A). When we mutated this RRLL motif in Pins, we observed a mislocalisation to the nucleus and delayed localisation to the anterior cortex during SOP mitosis (Fig. EV6B,C). This could be due to competition between cortical localisation and nuclear import of Pins prior to nuclear envelope breakdown. Consistent with this idea, we observed a small amount of nuclear Pins upon expression of wild-type Pins (Fig. EV6B, −24 min and −18 min time points), and loss of the anterior PCP component Vang leads to some Pins nuclear accumulation (Gomes et al, 2009). As Meru and Pins bind Mud through partially overlapping domains (Fig. 2B), this highlights strong mechanistic parallels between Meru and Pins in terms of Mud recruitment. Pins phosphorylation on S436 by Aurora A increases binding to Dlg (Johnston et al, 2009), suggesting that the Dlg/Pins/NuMA complex is also under tight regulation by the mitotic kinases. Indeed, the Dlg binding partner Guk-holder, which has been implicated in spindle tethering in neuroblasts (NBs or neural stem cells) and S2 cells with induced polarisation (Albertson and Doe, 2003; Garcia et al, 2014; Golub et al, 2017), also has an RRLL motif (Fig. EV6A). We found RRLL motifs in other proteins involved in spindle regulation (Aurora borealis (Hutterer et al, 2006), Sister of feo (Vernì et al, 2004)) or cell cycle progression (Double parked (Whittaker et al, 2000)), suggesting it can be used as a predictor of regulation by Cdks (Fig. EV6A).
Figure EV6. Loss of the Pins RxL motif results in delayed cortical localisation.

(A) BLAST search results for novel [R/K]-[R/K]-L-L-x[7]-[S/T]-P (RRLL) motifs resulted in nine proteins of interest, including Meru, Pins, Gukh and AurA, all of which are involved in spindle orientation. (B, C) Maximum intensity projections from confocal live-imaging of pupal nota at 16 h APF, 0 min marking first frame in cytokinesis, shows expression of UAS-mK2-PinsWT (B) and UAS-mK2-PinsRxL (C) driven by neurG4 during SOP mitosis. Scale bars = 10 µm. Source data are available online for this figure.
Discussion
To build and maintain tissues of the appropriate architecture, cells must read external cues from the extracellular matrix and other cells to align their polarity along (PCP) and across (A–B polarity) the tissue plane (Buckley and St Johnston, 2022; Butler and Wallingford, 2017). A key aspect of tissue organisation is the orientation of the mitotic spindle, which is essential not only to determine the position of the daughter cells after division, but also for cell fate determination through asymmetric cell division (Bergstralh et al, 2017; di Pietro et al, 2016; Lechler and Mapelli, 2021). Although the core PCP and A–B polarity pathways are well conserved, cells adopt radically different polarised organisations and cell division orientations to give rise to the diversity of adult tissues, from the epidermis and intestine to the nervous system (Buckley and St Johnston, 2022; Butler and Wallingford, 2017). Understanding how cells use these conserved polarity complexes as landmarks to generate cell type-specific polarity and spindle orientation remains a key challenge in the field, for which few mechanisms and molecular players have been identified. One of the best-understood tissue-specific polarity adaptors is Inscutable (Insc), which was first identified in Drosophila NBs (Kraut et al, 1996; Kraut and Campos-Ortega, 1996). In NBs, Insc connects the apical polarity protein Baz with the spindle tethering machinery via Pins, which promotes spindle orientation perpendicular to the tissue plane, allowing the differentiating NB progeny, the ganglion mother cells, to delaminate (Kraut et al, 1996; Schober et al, 1999; Wodarz et al, 1999; Yu et al, 2000).
We had previously shown that Meru, which is transcriptionally activated in SOPs as part of their differentiation programme (Banerjee et al, 2017; Buffin and Gho, 2010; Reeves and Posakony, 2005), links PCP with A–B polarity by promoting Baz planar polarisation (Banerjee et al, 2017). Here, we show that Meru also couples cell polarity with spindle orientation by assembling a ternary complex with the PCP protein Dsh and the spindle tethering factor Mud (Fig. 2D), promoting Mud localisation to the posterior cortex to orient the SOP spindle along the A–P axis (Fig. 3). In agreement with this model, Mud is depleted from the posterior cortex in meru mutants (Fig. 3A–C) and the spindle is no longer oriented along the A–P axis (Fig. 3D–F’), as in fz and dsh mutants (Bellaïche et al, 2001; Gho and Schweisguth, 1998; Ségalen et al, 2010). Unexpectedly, despite the enrichment of Meru at the posterior cortex, cortical Mud levels were reduced both anteriorly and posteriorly in meru mutant SOPs (Fig. 3C). This could indicate that Meru is also required for Mud stability and in its absence, the overall Mud pool is depleted. This is difficult to verify as low Mud expression precludes reliable measurement of its cytosolic levels. However, in our rescue experiments, MeruWT overexpression in a meru mutant background causes Mud to be deposited equally at the anterior and posterior poles (Fig. 6B,D), in contrast to wild-type SOPs, where we detect more Mud anteriorly than posteriorly (Fig. 3C). Together with the fact that MeruRxL expression causes loss of Mud specifically at the posterior pole (Fig. 6C,D), this strongly argues in favour of our model that Meru promotes posterior Mud recruitment. While Meru is enriched at the posterior SOP cortex, we detect some signal at the anterior pole, especially upon GAL4/UAS-driven overexpression (Fig. 6B’). This could be because overexpressed Meru saturates endogenous Dsh, therefore spilling over to the anterior side, or may indicate a two-step Meru cortical recruitment process where Meru is first recruited to the plasma membrane independently of Dsh, which then biases Meru cortical localisation by preferentially retaining it posteriorly.
Could RASSF proteins such as Meru be implicated in spindle orientation in other contexts? Since the Meru mammalian orthologs RASSF9 and RASSF10 associate with Dvl proteins (Hauri et al, 2013), it would be interesting to test their involvement in oriented cell divisions in contexts in which Dvl1-3 have been implicated, such as HeLa cells (Kikuchi et al, 2010; Yang et al, 2014) and zebrafish gastrulation (Ségalen et al, 2010). Furthermore, similar to Drosophila NBs, mouse Inscutable (mInsc) participates in orientating the spindle perpendicular to the tissue plane to promote differentiation of embryonic epidermal progenitors (Lechler and Fuchs, 2005; Poulson and Lechler, 2010; Williams et al, 2014). As RASSF9 is highly expressed in epidermal keratinocytes and RASSF9 mutant animals show increased proliferation and aberrant differentiation in the epidermis (Lee et al, 2011), it is exciting to speculate that RASSF9 and mInsc could both regulate spindle orientation in this tissue. In Drosophila, the Meru paralog RASSF8 is expressed in many epithelial tissues (Langton et al, 2009), therefore it would also be interesting to investigate its role and that of its mammalian orthologs RASSF7 and RASSF8 in symmetric cell division orientation.
Besides positioning, a second important aspect of spindle orientation is temporally linking progression through the cell cycle with spindle tethering (Bergstralh et al, 2017; di Pietro et al, 2016; Lechler and Mapelli, 2021). Several cell polarity and spindle tethering factors such as Lethal giant larvae, Dvl and NuMA, are regulated through phosphorylation by mitotic kinases, supporting the idea of an intimate relationship between spindle orientation and the cell cycle (di Pietro et al, 2016; Lechler and Mapelli, 2021; Osswald and Morais-de-Sá, 2019). We identified a mechanism whereby Mud cortical localisation is coupled to the cell cycle via Meru phosphorylation, linking cell polarity, the cell cycle and spindle orientation. Darnat et al showed that the Cdk1 partner CycA is recruited to the SOP posterior cortex in a Dsh-dependent manner (Darnat et al, 2022). We find that Meru, but not Dsh, associates with CycA in S2 cells (Fig. 4C). We uncovered in Meru a pair of overlapping Cyclin-binding RxL SLiMs (which we termed the RRLL motif) that promote Dsh/Meru and Mud/Meru binding (Fig. 5C,D) and Meru cortical recruitment (Fig. 6C’). Furthermore, both CycA docking and Cdk1 phosphorylation of Meru promote the localisation of Mud to the posterior cortex (Figs. 6C,D and 8B,C), as well as A–P spindle orientation (Fig. 6F,G). Mutating two of the Meru RxL motifs (amino acids 258/9) is sufficient to disrupt Meru function, but the spindle orientation phenotype of MeruRxL-expressing flies (Fig. 6G,G’) is slightly weaker than that of the meru1 mutant (Fig. 3F,F’), suggesting either that the remaining RxL motifs can provide some functional rescue in vivo, or that CycA binding strongly increases the affinity of Meru for Dsh and Mud, but is not absolutely required for it. We therefore propose a model in which, following its posterior recruitment via Dsh, Meru recruits CycA/Cdk1, which in turn phosphorylates Meru, promoting its stabilisation at the posterior cortex and Mud recruitment (Fig. 8G). Consistent with this model, MeruRxL is impaired in its ability to mediate the assembly of the ternary Dsh/Meru/Mud complex (Fig. 6A), and a form of Meru mutant for its five Cdk1 consensus sites (Meru5S/T-A) displays much reduced Mud binding (Fig. 7C).
Does CycA/Meru association promote the phosphorylation of other posterior cortical proteins? Our data indicate that Dsh may be another target, since mutation of the only Cdk1 consensus site in Dsh causes reduced binding to Meru5S/T-A (Fig. 7D), suggesting phosphorylation of both partners may be required for optimal binding. This may account for the fact that MeruRxL displays a defect in Mud recruitment at anaphase (Fig. 6C,D) whereas the Meru5S/T-A mutant has delayed Mud recruitment but no detectable defect at anaphase (Figs. 8B,C and EV5E–J), as other Meru partners such as Dsh would still be phosphorylated by CycA/Cdk1 in this mutant, partially rescuing complex formation. It is interesting to note that human Dvl2 phosphorylation on T206 by Plk1 is required for spindle orientation in cultured human cells (Kikuchi et al, 2010), therefore Dsh/Dvl proteins may receive inputs from different cell cycle kinases. Finally, it is possible that Mud is also phosphorylated by Cdk1 in a Meru-dependent manner. Indeed, Cdk1 phosphorylates NuMA on four C-terminal residues to control its cortical turnover in human cells and C. elegans, at least in part by inhibiting a NuMA lipid-binding domain (Compton and Luo, 1995; Gehmlich et al, 2004; Hsin-Ling and Ning-Hsing, 1996; Kiyomitsu and Cheeseman, 2013; Kotak et al, 2013; Portegijs et al, 2016; Saredi et al, 1997; Seldin et al, 2013; Zheng et al, 2014).
In summary, we uncover a mechanism whereby Meru allows coupling of the spindle tethering complex both to the cell polarity machinery and cell cycle progression. We propose that similar regulatory logics underpin the diverse spindle orientation mechanisms employed by other symmetrically and asymmetrically dividing cells.
Methods
Reagents and tools table
| Reagent/resource | Reference or source | Identifier or catalogue number |
|---|---|---|
| Experimental models | ||
| w[1118]; P{w[+mW.hs]=GawB}neur[GAL4-A101] Kg[V]/TM3, Sb[1] | Bloomington Drosophila stock center | 6393 |
| y[1] w[*]; TI{TI}shg[GFP] | Bloomington Drosophila stock center | 60584 |
| FlyFos028184(pRedFlp-Hgr)(dlg1[37834]::2XTY1-SGFP-V5-preTEV-BLRP-3XFLAG)dFRT | Vienna Drosophila Resource Center | 318133 |
| w;; GFP::mud(62E1), GFP::mud(65B2) |
Gift from Yohanns Bellaïche Ségalen et al, 2010 10.1016/j.devcel.2010.10.004 |
|
| w;; UAS-Pon-GFP |
Gift from Yohanns Bellaïche Lu et al, 1999 10.1016/s1097-2765(00)80218-x |
|
| y[1] w[*]; P{w[+mC]=UAS-mCD8.mRFP.LG}18a | Bloomington Drosophila stock center | 27398 |
| w;; neur-H2B-RFP |
Gift from François Schweisguth Gomes et al, 2009 10.1371/journal.pone.0004485 |
|
| y[1] v[1]; P{y[+t7.7] v[+t1.8]=TRiP.HMJ21351}attP40 | Bloomington Drosophila stock center | 53968 |
| y[1] w[1118]; P{w[+mC]=PTT-GA}Jupiter[ZCL0931]/TM3, Ser[1] | Bloomington Drosophila stock center | 6825 |
| w;; meru 1 |
Banerjee et al, 2017 10.7554/eLife.25014 |
|
| w;; CycA-GFP |
Gift from Michel Gho and Agnes Audibert Darnat et al, 2022 10.1038/s41467-022-30182-1 |
|
| [1] cv[1] v[1] fs(1)M29[A151] f[1] car[1]/FM7a | Bloomington Drosophila stock center | 4716 |
| w; UAS-mK2-Meru | This paper | |
| w; UAS-mK2-Meru RxL | This paper | |
| w; UAS-mK2-Meru 5S/T-A | This paper | |
| w; UAS-mK2-Pins | This paper | |
| w; UAS-mK2-Pins RxL | This paper | |
| S2 cells | Drosophila Genomics Resource Center | RRID:CVCL_Z232 |
| Recombinant DNA | ||
| Drosophila Gateway Vector Collection | https://carnegiescience.edu/bse/drosophila-gateway-vector-collection | |
| Antibodies | ||
| Mouse anti-FLAG | Sigma | F1804 |
| Rabbit anti-Flag | Sigma | |
| Mouse anti-Myc | Santa Cruz Biotechnology | sc-40 |
| Rabbit anti-HA | Cell Signalling | C29F4 |
| Rat anti-GFP | ChromoTek | 3h9 |
| Rabbit IgG HRP-Linked F(ab′)2 | Sigma | NA9340 |
| Mouse IgG HRP-Linked F(ab′)2 Fragment | Sigma | NA9310 |
| Rat IgG, HRP-linked whole antibody | Sigma | NA935 |
| Oligonucleotides and other sequence-based reagents | ||
| Dsh pCdk1mt fw | CCACATCGACGATGAGACGgCGCCGTATCTGGTGAAGATCCCC | |
| Dsh pCdk1mt rev | GGGGATCTTCACCAGATACGGCGcCGTCTCATCGTCGATGTGG | |
| Pins RxL 390/91 fw | GTGAACATCTCCGATCTAgcAgcGgcAgcCGGAATGCCCGACTCCG | |
| Pins RxL 390/91 rev | CGGAGTCGGGCATTCCGgcTgcCgcTgcTAGATCGGAGATGTTCAC | |
| Meru pCdk1mt 267 fw | GGGTAAAAAGgCGCCACCAAAACCCACAAAAACGCCTCCAAAAGTGCCGAAC | |
| Meru pCdk1mt 267 rev | GTTCGGCACTTTTGGAGGCGTTTTTGTGGGTTTTGGTGGCGcCTTTTTACCC | |
| Meru pCdk1mt 274 fw | GGGTAAAAAGACGCCACCAAAACCCACAAAAgCGCCTCCAAAAGTGCCGAAC | |
| Meru pCdk1mt 274 rev | GTTCGGCACTTTTGGAGGCGcTTTTGTGGGTTTTGGTGGCGTCTTTTTACCC | |
| Meru pCdk1mt 27 fw | CGCTATTGAATCTACCCGAAgcTCCCATTCTTCCGCCGCGAC | |
| Meru pCdk1mt 27 rev | GTCGCGGCGGAAGAATGGGAgcTTCGGGTAGATTCAATAGCG | |
| Meru pCdk1mt 60 fw | GGAGCGTTTGGTGGATGAAgCGCCATCGGTGGGGGAGCCAC | |
| Meru pCdk1mt 60 rev | GTGGCTCCCCCACCGATGGCGcTTCATCCACCAAACGCTCC | |
| Meru pCdk1mt 496 fw | CCTTACAGGAAAATGAAAATgCCCCTCAAGTTGCTAAAAATACGC | |
| Meru pCdk1mt 496 rev | GCGTATTTTTAGCAACTTGAGGGGcATTTTCATTTTCCTGTAAGG | |
| Meru Rxl 258/9 | CGATGTACAAGTGGCTCgcGgcGgcAgcGCATCTGAAAAAGGG | |
| Meru Rxl 258 | CGATGTACAAGTGGCTCgctAAGgctTTGCATCTGAAAAAGGG | |
| Meru Rxl 259 | CGATGTACAAGTGGCTCAAGgcGCTAgcGCATCTGAAAAAGGG | |
| Meru Rxl 154 | CGTGCGATGACATAATCgcGGCGgcGATTGATGACGAACTGCG | |
| Meru Rxl 375 | CATAAGGAGGCGAAAGGATgcGCCCgcGCGAAATAGTGTACG | |
| Meru Rxl 404 | GGAACACGCTCTAACAgcTCAGgcATCGGAAATGTGCCGACTG | |
| Meru Rxl 468 | CGCTGATTAATAACTTAgcGCGTgcGACGTTGGAGGAATCGGAGG | |
| CycA RNAi fw | TAATACGACTCACTATAGGGCGGTGCTGGGCAGAAGGAGCTGG | |
| CycA RNAi rev | TAATACGACTCACTATAGGGCTCGCTCTCCCGGAAATATTCCAG | |
| Chemicals, enzymes and other reagents | ||
| S-trap (Micro) | Protifi | CO2-micro |
| Trypsin | Promega | 90057 |
| Trifluoracetic acid | Thermo Fisher Scientific | LS119-4 |
| Acetonitrile | Thermo Fisher Scientific | A955-212 |
| TEAB | Merck | 18597-100 ML |
| Effectene Transfection Reagent | Qiagen | 301425 |
| Pierce ECL Plus Western Blotting Substrate | Thermo Scientific | 32132 |
| ChromoTek GFP-Trap® Agarose | ChromoTek | gta |
| ChromoTek Myc-Trap® Agarose | ChromoTek | yta |
| Phosphatase Inhibitor Cocktail 2 | Sigma | P5726 |
| Phosphatase Inhibitor Cocktail 3 | Sigma | P0044 |
| cOmplete™, Mini, EDTA-free Protease Inhibitor Cocktail | Roche | 11836170001 |
| MEGAscript™ T7 Transcription Kit | Ambion | AMB13345 |
| Taq PCR Master Mix | Qiagen | 201445 |
| Schneider’s Drosophila Medium | Gibco | 11590576 |
| Penicillin–Streptomycin | Thermo Fisher Scientific | 15070063 |
| Foetal Bovine Serum | Thermo Fisher Scientific | Batch 2503539 |
| Gateway LR Clonase II Enzyme mix | Invitrogen | 10134992 |
| pENTR™/D-TOPO™ Cloning Kit | Invitrogen | 15575730 |
| Quikchange Multi Site-Directed Mutagenesis Kit | Agilent Technologies LDA UK Limited | 200515 |
| Software | ||
| Fiji | 10.1038/nmeth.2019 | http://fiji.sc |
| Python, Numpy, Scipy | Python Software Foundation | |
| Maxquant |
Juergen Cox lab 10.1038/nprot.2016.136 |
v2.6.2.0 |
| Perseus |
Juergen Cox lab 10.1007/978-1-4939-7493-1_7 |
v1.6.14.0 |
| Other | ||
Transgenes and fly stocks
UAS-mK2-meru transgenic lines were created by phiC31-mediated recombination using stock 9723 from Bloomington (as described in Banerjee et al, 2017). The meruRxL and meru5S/T-A mutant constructs were generated under the same protocol but the amino acids sites 258-261 or S27, S60, T267, T274, and S496 were mutated to Alanines in the RxL or 5S/T-A motifs, respectively. The Pins wild-type and RxL mutant were generated by replacing Meru from the construct above with Pins cDNA and mutating the RxL motif at amino acids 390/1 to Alanines at sites 390–393.
The following fly stocks were used: neurG4 (Bellaïche et al, 2001); Ecad-GFP (BL60584); Dlg-GFP (VDRC318133); UAS-Pon-GFP (Lu et al, 1999); GFP::mud(62E1), GFP::mud(65B2) gift from Yohanns Bellaïche (Ségalen et al, 2010); UAS-cd8-mRFP (BL27398); neur-H2B-RFP (Gomes et al, 2009); UAS-pins-RNAi (BL53968); neurG4, UAS-RFP, jupiter::GFP (recombined by Federica Mangione from BL6825); meru1 (Banerjee et al, 2017); CycA-GFP, gift from Michel Gho and Agnes Audibert (Darnat et al, 2022); UAS-nls-GFP (BL4716).
Genotypes
Figure 1B–C’w; UAS-mK2-meru/+; neurG4/+ (D) w; Ecad-GFP/UAS-mK2-meru; neurG4/+ (E–E’) w; UAS-mK2-meru/+; neurG4, UAS-Pon-GFP/+ (F–F”) w; UAS-mK2-meru(WT)/ + ; GFP::mud(62E1), GFP::mud(65B2), neurG4/+ Fig. 3Aw; UAS-cd8-mRFP/+ GFP::mud(62E1), GFP::mud(65B2), neurG4/+ (B) w; UAS-cd8-mRFP/+ GFP::mud(62E1), GFP::mud(65B2), neurG4, meru1/meru1 (D) w;; neur-H2B-RFP, jupiter::GFP (E) w;; neur-H2B-RFP, jupiter::GFP, meru1/meru1 Fig. 4Aw; UAS-mK2-meru/+; CycA::GFP, neruG4/+ Fig. 5Fw; UAS-mK2-meruRxL/+; CycA::GFP, neurG4, meru1/meru1 Fig. 6B,B’w; UAS-mK2-meruWT/+; GFP::mud(62E1), GFP::mud(65B2), neurG4, meru1/meru1 (C–C’) w; UAS-mK2-meruRxL/+; GFP::mud(62E1), GFP::mud(65B2), neurG4, meru1/meru1 (E) w; UAS-mK2-meruWT/+; neurG4, UAS-RFP, jup::GFP, meru1/meru1 (F) w; UAS-mK2-meruRxL/+; neurG4, UAS-RFP, jup::GFP meru1/meru1 Fig. 8A,A’w; UAS-mK2-meruWT/+; GFP::mud(62E1), GFP::mud(65B2), neurG4, meru1/meru1 (B–B’) w; UAS-mK2-meru5S/T-A/ + ; GFP::mud(62E1), GFP::mud(65B2), neurG4, meru1/meru1 (D) w; UAS-mK2-meruWT/+; neurG4, UAS-RFP, jup::GFP, meru1/meru1 (E) w; UAS-mK2-meru5S/T-A/+; neurG4, UAS-RFP, jup::GFP meru1/meru1 Fig. EV1A,A’w; Ecad-GFP/UAS-mK2-meru; neurG4/+ (B–B’) w; UAS-mK2-meru(WT)/ + ; GFP::mud(62E1), GFP::mud(65B2), neurG4/+ (D–D’) w; UAS-mK2-meru/+; neurG4/+ Fig. EV3A,A’w; UAS-nls-GFP/UAS-mK2-meruRxL; neurG4/+ (B–B’) w; Ecad-GFP/UAS-mK2-meruRxL; neurG4/+ (C–C’) w; UAS-mK2-meruRxL/+; neurG4, Dlg-GFP/+ (D–D’) w; UAS-mK2-meruWT/+; CycA::GFP, neurG4, meru1/meru1 Fig. EV5A,A’,E,E’,H,H’)w; UAS-mK2-meruWT/+; GFP::mud(62E1), GFP::mud(65B2), neurG4, meru1/meru1 (B–B’, F–F’, I–I’) w; UAS-mK2-meru5S/T-A/ + ; GFP::mud(62E1), GFP::mud(65B2), neurG4, meru1/meru1 Fig. EV6Bw; UAS-mK2-PinsWT/+; neurG4/+ (C) w; UAS-mK2-PinsRxL/+; neurG4/+ Movie EV1w;; neur-H2B-RFP, jupiter::GFP Movie EV2w;; neur-H2B-RFP, jupiter::GFP, meru1/meru1.
Confocal live-imaging
Drosophila pupae were attached to a Superfrost microscope slide (Thermo Scientific) using double-sided tape so that the A–P axis was positioned along the width of the slide. Once adhered, the pupal case from the head to the start of the abdomen was peeled off. Two stacks of four 22 × 22 mm No. 1.5 Cover Glass (VWR) were glued together and placed on either side of the pupae. A lightly oiled 24 × 50 mm No. 1.5 Cover Glass (VWR) was placed atop the two cover slip stacks so that the oil side rested gently on the exposed notum. The sample was then inverted to image on an inverted Nikon Spinning Disk (×60 or ×100 oil objectives) confocal microscope.
Adult notum and wing imaging
Female adults, raised at 25 °C, were collected based on genotype and were stored at −20 °C or in 70% EtOH at 4 °C for notum and wing imaging, respectively. Adult nota were mounted in 0.8% low melting agarose and imaged on an MZ16 stereomicroscope (Leica) with a QImaging MicroPublisher 6 Colour Camera. Adult wings were mounted onto Superfrost microscope slides (Thermo Scientific) in Euparal covered in 22 × 22 mm No. 1.5 Cover Glass (VWR) for imaging. Wings were imaged with an Axio Scan.Z1 (Zeiss) slide scanner with a ×2.5 and ×10 objective.
Colocalisation analysis, puncta quantification, cortical Mud/Meru/CycA levels and spindle orientation in time-lapse movies
All analyses were carried out using ImageJ. The degree of correlation between Meru and Mud/CycA localisation at the posterior cortex was measured by drawing a 2 μm thick line over the Meru cortical crescent. The line was applied to the single brightest slice (0.6 µm thick) in the stack used in the corresponding figure to the Meru and Mud/CycA channels, and the normalised grey value was plotted versus the distance along the line (in μm). The correlation between the Meru and Mud/CycA grey value levels was calculated by the Pearson’s correlation coefficient (r) in which 1 represents perfect correlation, −1 represents anti-correlation and 0 depicts no correlation. Cortical levels of CycA/Meru were collected by the same means and measured by plotting mean intensity across cells. The number of Mud puncta was quantified by the observation of clustering and manually counting puncta at the cortex.
Levels of cortical Mud were quantified by measuring the sum of the integrated density of a cortex-specific ROI corrected for background fluorescence and ROI area. A mask of the membrane marker (CD8-mRFP or mK2-Meru) was used to outline the cell membrane and placed over the GFP channel to isolate the signal to the dividing cell. As the mitotic spindle in SOPs has a z-tilt (David et al, 2005), the anterior and posterior crescents were analysed separately. The three brightest slices (0.6 µM z-slices) for GFP::Mud signal were max-projected and isolated by drawing around the ROI, ensuring that the signal coming from the centrosome was excluded. The cytoplasmic signal (background fluorescence) was measured using a 10 × 10 pixel square for each cortical max projection. The sum of the anterior and posterior cortical Mud signals, respectively, were corrected for background fluorescence and area of the ROI by the following formula:
To measure spindle angles, movies obtained by confocal microscopy were imported into the Imaris Microscopy Image Analysis Software (Oxford Instruments) to quantify the x- and z axis of every centrosome pair in dividing cells of interest. All measurements were made by manually positioning a point at the center of either centrosome, creating a line through the spindle. The x-, y- and z-coordinates of each point were used to quantify the angle of division relative to a reference position. The first measurement pair was used to define the dorsal midline, identified by visual cues such as anisotropic cell shape, followed by measurement pairs at all dividing cells of interest. The spindle angles in x axis (A–P axis) were calculated as degrees away from the midline. The reference slope (the midline angle) was calculated by the following equation:
To quantify the x axis angle of each centrosome pair, the slope was found with the same equation above (called centrosomem) and calculated as follows:
The z-axis angle (A–B axis) was calculated as degrees away from the plane. The reference plane angle was set to 0. and the absolute z-axis angle of each centrosome pair was calculated with the following equation:
Statistical analysis
Mann–Whitney U tests were performed to test if mean values of cortical Mud/CycA levels were significantly different. The Kolmogorov–Smirnov (K-S) test was used to test if measurements from SOP spindle alignment sample distributions were significantly different. To test the colocalisation of two fluorescently tagged proteins in a time-lapse movie, the Pearson’s correlation test was performed. No blinding was done.
Drosophila cell culture and expression constructs
Drosophila S2 cells were transfected with Effectene transfection reagent (Qiagen) and grown in Drosophila Schneider’s medium (Life Technologies) containing 10% FBS (Sigma), 50 μg/ml penicillin and 50 μg/ml streptomycin. Expression plasmids were generated using Gateway® technology (Life Technologies). Open reading frames (ORF) were PCR-amplified from existing plasmids or cDNA (DGRC, https://dgrc.bio.indiana.edu/vectors/Catalog) and cloned into entry vectors (pENTRTM/D-TOPO). Vectors from the Drosophila Gateway Vector Collection (https://carnegiescience.edu/bse/drosophila-gateway-vector-collection) were used as Destination vectors. Meru, Mud and Baz were tagged N-terminally and Dsh, Cdk1 and all the Cyclins were tagged C-terminally. Point mutations were generated using the Quikchange Multi Site-Directed Mutagenesis Kit (Agilent). All expression plasmids were sequence-verified.
dsRNA production and treatment
dsRNAs were synthesised using the Megascript T7 kit (Thermo Fisher Scientific). DNA templates for dsRNA synthesis were PCR-amplified from genomic DNA using primers that contained the 5’ T7 RNA polymerase-binding site sequence. dsRNAi primers were designed using the DKFZ RNAi design tool (https://e-rnai.dkfz.de/signaling/e-rnai3/). RNAi experiments were carried out in six-well plates using S2 cells. In total, 1–2 × 106 cells were plated per well and cells left to settle. After 3 h, the medium was removed and replaced with 1 mL of serum-free Schneider’s medium containing dsRNAs (20 μg). Cells were soaked for 30 min, and then 2 mL of full Schneider’s medium was added. Transfections were carried out 1 h after dsRNA treatment.
Immunoprecipitation and immunoblot analysis
For immunoprecipitation assays, cells were lysed in lysis buffer (150 mM NaCl, 50 mM HEPES pH 7.5, 0.5% (v/v) Triton X-100) supplemented with phosphatase inhibitor cocktails 2 and 3 (Sigma) and protease inhibitor cocktail (Roche). Cell extracts were cleared at 13,200 rpm for 20 min at 4 °C. GFP and Myc-tagged proteins were purified using agarose beads (Chromotek) according to the manufacturer's protocol. For MS analysis, proteins were eluted using 5% SDS 40 mM in Hepes pH 7.0. Detection of purified proteins and associated complexes was performed by immunoblot analysis using chemiluminescence (Pierce). Western Blots were probed with mouse anti-FLAG (M2, Sigma), rabbit anti-FLAG (Sigma), mouse anti-Myc (9E10, Santa Cruz Biotechnology), rabbit anti-HA (C29F4, NEB), and rat anti-GFP (Chromotek).
Mass spectrometry analysis
Following immunoprecipitation, samples were digested using the S-Trap micro protocol (ProtiFi). Eluted peptides were resuspended in 0.1% trifluoroacetic acid (TFA) and analysed on an Orbitrap Eclipse (Thermo Fisher Scientific) coupled to an UltiMate3000 LC system. In brief, 2 mg of dried peptides were injected onto a trap column (Acclaim PepMap 100; Thermo Fisher Scientific), and peptides were resolved using an Easy-Spray 50 cm column (PepMap RSLC C18) with the following gradient: 0 min—2% solvent B (75% acetonitrile in 0.1% formic acid), 120 min—35% solvent B, 137 min—95% solvent B, 143 min—95% solvent B, 148 min—2% solvent B). The Orbitrap Eclipse was operated in data-dependent acquisition mode using a fixed cycle time of 3 s. The Orbitrap resolution was set to 120,000, the scan range to 300–1500 m/z, standard AGC and auto-injection were enabled, and the fragmentation method was set to HCD at 30. MS2 scans were performed in the Orbitrap with a resolution of 30,000, standard AGC and scan parameters and dynamic injection times. Raw data from duplicate samples was searched in MaxQuant with phosphorylation (STY) and oxidation set as variable modifications, while carbamidomethylation was set as a fixed modification. Label-free quantification (LFQ) intensities corresponding to modified peptides of interest were plotted on the graph.
Supplementary information
Acknowledgements
The authors thank Y Bellaïche, M Gho, F Schweisguth, F Mangione and the Bloomington Drosophila Stock Center for fly stocks and Y Bellaïche for plasmids. The authors are grateful to M Renshaw and members of the Crick Advanced Light Microscopy Facility for advice on confocal imaging and image analysis; J Kurth, S Murray, H Shaw, S Ruiz-Herrera from the Crick Fly Facility for embryo injections and fly stock maintenance; and Tapon lab members for advice. We thank A Moraiti, F Mangione, N Goehring and A Audibert for critical reading of the manuscript. This work was supported by the Francis Crick Institute, which receives its core funding from Cancer Research UK (CC2138, CC1063), the UK Medical Research Council (CC2138, CC1063) and the Wellcome Trust (CC2138, CC1063), and a Wellcome Trust Investigator award (107885/Z/15/Z) to NT. For the purpose of Open Access, the authors have applied a CC BY public copyright licence to any Author Accepted Manuscript version arising from this submission.
Expanded view
Author contributions
Melissa M McLellan: Conceptualisation; Investigation; Writing—original draft; Writing—review and editing. Birgit L Aerne: Conceptualisation; Supervision; Investigation; Writing—original draft; Writing—review and editing. Jennifer J Banerjee Dhoul: Conceptualisation; Investigation; Writing—review and editing. Maxine V Holder: Investigation; Writing—review and editing. Tania Auchynnikava: Investigation; Writing—review and editing. Nicolas Tapon: Conceptualisation; Supervision; Funding acquisition; Writing—original draft; Writing—review and editing.
Source data underlying figure panels in this paper may have individual authorship assigned. Where available, figure panel/source data authorship is listed in the following database record: biostudies:S-SCDT-10_1038-S44318-025-00420-5.
Funding
Open Access funding provided by The Francis Crick Institute.
Data availability
Source data (imaging and biochemistry) are associated with the manuscript. The Mass Spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE (Perez-Riverol et al, 2025) partner repository (https://www.ebi.ac.uk/pride/) with the dataset identifier PXD060813.
The source data of this paper are collected in the following database record: biostudies:S-SCDT-10_1038-S44318-025-00420-5.
Disclosure and competing interests statement
The authors declare no competing interests.
Footnotes
These authors contributed equally: Melissa M McLellan, Birgit L Aerne.
Change history
4/7/2025
In this article, the ORCiD assigned to Melissa M McLellan was inadvertently linked to Birgit L Aerne. This has been corrected.
Supplementary information
Expanded view data, supplementary information, appendices are available for this paper at 10.1038/s44318-025-00420-5.
References
- Albertson R, Chabu C, Sheehan A, Doe CQ (2004) Scribble protein domain mapping reveals a multistep localisation mechanism and domains necessary for establishing cortical polarity. J Cell Sci 117(25):6061–6070 [DOI] [PubMed] [Google Scholar]
- Albertson R, Doe CQ (2003) Dlg, Scrib and Lgl regulate neuroblast cell size and mitotic spindle asymmetry. Nat Cell Biol 5(2):166–170 [DOI] [PubMed] [Google Scholar]
- Axelrod JD, Miller JR, Shulman JM, Moon RT, Perrimon N (1998) Differential recruitment of dishevelled provides signaling specificity in the planar cell polarity and Wingless signaling pathways. Genes Dev 12(16):2610–2622 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banerjee JJ, Aerne BL, Holder MV, Hauri S, Gstaiger M, Tapon N (2017) Meru couples planar cell polarity with apical-basal polarity during asymmetric cell division. eLife 6:1–21 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bastock R, Strutt H, Strutt D (2003) Strabismus is asymmetrically localised and binds to Prickle and Dishevelled during Drosophila planar polarity patterning. Development 130:3007–3014 [DOI] [PubMed] [Google Scholar]
- Bellaïche Y, Gho M, Kaltschmidt JA, Brand AH, Schweisguth F (2001) Frizzled regulates localisation of cell-fate determinants and mitotic spindle rotation during asymmetric cell division. Nat Cell Biol 3(1):50–57 [DOI] [PubMed] [Google Scholar]
- Bellaïche Y, Radovic A, Woods DF, Hough CD, Parmentier ML, O’Kane CJ, Bryant PJ, Schweisguth F (2001) The Partner of Inscuteable/Discs-large complex is required to establish planar polarity during asymmetric cell division in Drosophila. Cell 106(3):355–366 [DOI] [PubMed] [Google Scholar]
- Bergstralh DT, Dawney NS, St Johnston D (2017) Spindle orientation: a question of complex positioning. Development 144(7):1137–1145 [DOI] [PubMed] [Google Scholar]
- Bergstralh DT, Lovegrove HE, Kujawiak I, Dawney NS, Zhu J, Cooper S, Zhang R, St Johnston D (2016) Pins is not required for spindle orientation in the Drosophila wing disc. Development 143(14):2573–2581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Besson C, Bernard F, Corson F, Rouault H, Reynaud E, Keder A, Mazouni K, Schweisguth F (2015) Planar cell polarity breaks the symmetry of PAR protein distribution prior to mitosis in Drosophila sensory organ precursor cells. Curr Biol 25(8):1104–1110 [DOI] [PubMed] [Google Scholar]
- Bilder D, Li M, Perrimon N (2000) Cooperative regulation of cell polarity and growth by Drosophila tumor suppressors. Science 289(5476):113–116 [DOI] [PubMed] [Google Scholar]
- Bosveld F, Markova O, Guirao B, Martin C, Wang Z, Pierre A, Balakireva M, Gaugue I, Ainslie A, Christophorou N, Lubensky DK, Minc N, Bellaïche Y (2016) Epithelial tricellular junctions act as interphase cell shape sensors to orient mitosis. Nature 530(7591):495–498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckley CE, St Johnston D (2022) Apical–basal polarity and the control of epithelial form and function. Nat Rev Mol Cell Biol 23:559–577 [DOI] [PubMed] [Google Scholar]
- Buffin E, Gho M (2010) Laser microdissection of sensory organ precursor cells of Drosophila microchaetes. PLoS ONE 5(2):e9285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butler MT, Wallingford JB (2017) Planar cell polarity in development and disease. Nat Rev Mol Cell Biol 18(6):375–388 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Compton DA, Luo C (1995) Mutation of the predicted p34(cdc2) phosphorylation sites in NuMA impair the assembly of the mitotic spindle and block mitosis. J Cell Sci 108(2):621–633 [DOI] [PubMed] [Google Scholar]
- Corson F, Couturier L, Rouault H, Mazouni K, Schweisguth F (2017) Self-organized Notch dynamics generate stereotyped sensory organ patterns in Drosophila. Science 356:eaai7407 [DOI] [PubMed] [Google Scholar]
- Darnat P, Burg A, Sallé J, Lacoste J, Louvet-vallée S, Gho M, Audibert A (2022) Cortical Cyclin A controls spindle orientation during asymmetric cell divisions in Drosophila. Nat Commun 13:2723 [DOI] [PMC free article] [PubMed] [Google Scholar]
- David NB, Martin CA, Segalen M, Rosenfeld F, Schweisguth F, Bellaïche Y (2005) Drosophila Ric-8 regulates Gαi cortical localization to promote Gαi-dependent planar orientation of the mitotic spindle during asymmetric cell division. Nat Cell Biol 7(11):1083–1090 [DOI] [PubMed] [Google Scholar]
- di Pietro F, Echard A, Morin X, Pietro F, Echard A, Morin X (2016) Regulation of mitotic spindle orientation: an integrated view. EMBO Rep 17(8):1106–1130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enserink JM, Kolodner RD (2010) An overview of Cdk1-controlled targets and processes. Cell Div 5:1–41 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feiguin F, Hannus M, Mlodzik M, Eaton S (2001) The Ankyrin repeat protein diego mediates frizzled-dependent planar polarization. Dev Cell 1(1):93–101 [DOI] [PubMed] [Google Scholar]
- Garcia JD, Dewey EB, Johnston CA (2014) Dishevelled binds the Discs large ‘Hook’ domain to activate GukHolder-dependent spindle positioning in Drosophila. PLoS ONE 9(12):1–17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gehmlich K, Haren L, Merdes A (2004) Cyclin B degradation leads to NuMA release from dynein/dynactin and from spindle poles. EMBO Rep 5(1):97–103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gho M, Schweisguth F (1998) Frizzled signalling controls orientation of asymmetric sense organ precursor cell divisions in Drosophila. Nature 393(6681):178–181 [DOI] [PubMed] [Google Scholar]
- Golub O, Wee B, Newman RA, Paterson NM, Prehoda KE (2017) Activation of discs large by aPKC aligns the mitotic spindle to the polarity axis during asymmetric cell division. eLife 6:1–17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomes JE, Corado M, Schweisguth F (2009) Van Gogh and Frizzled act redundantly in the Drosophila sensory organ precursor cell to orient its asymmetric division. PLoS ONE 4(2):e4485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gómez-Skarmeta JL, Campuzano S, Modolell J (2003) Half a century of neural prepatterning: The story of a few bristles and many genes. Nat Rev Neurosci 4(7):587–598 [DOI] [PubMed] [Google Scholar]
- Goodrich LV, Strutt D (2011) Principles of planar polarity in animal development. Development 138(10):1877–1892 [DOI] [PMC free article] [PubMed]
- Hartenstein V, Posakony JW (1989) Development of adult sensilla on the wing and notum of Drosophila melanogaster. Development 107(2):389–405 [DOI] [PubMed] [Google Scholar]
- Hauri S, Wepf A, Van Drogen A, Varjosalo M, Tapon N, Aebersold R, Gstaiger M (2013) Interaction proteome of human Hippo signaling: Modular control of the co-activator YAP1. Mol Syst Biol 9(1):713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsin-Ling H, Ning-Hsing Y (1996) Dynamic changes of NuMA during the cell cycle and possible appearance of a truncated form of NuMA during apoptosis. J Cell Sci 109:277–288 [DOI] [PubMed] [Google Scholar]
- Hutterer A, Berdnik D, Wirtz-Peitz F, Žigman M, Schleiffer A, Knoblich JA (2006) Mitotic activation of the kinase aurora-A requires its binding partner bora. Dev Cell 11(2):147–157 [DOI] [PubMed] [Google Scholar]
- Izumi Y, Ohta N, Hisata K, Raabe T, Matsuzaki F (2006) Drosophila Pins-binding protein Mud regulates spindle-polarity coupling and centrosome organization. Nat Cell Biol 8(6):586–593 [DOI] [PubMed] [Google Scholar]
- Jenny A, Darken RS, Wilson PA, Mlodzik M (2003) Prickle and Strabismus form a functional complex to generate a correct axis during planar cell polarity signaling. EMBO J 22(17):4409–4420 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenny A, Reynolds-Kenneally J, Das G, Burnett M, Mlodzik M (2005) Diego and Prickle regulate Frizzled planar cell polarity signalling by competing for Dishevelled binding. Nat Cell Biol 7(7):691–697 [DOI] [PubMed] [Google Scholar]
- Johnston CA, Hirono K, Prehoda KE, Doe CQ (2009) Identification of an Aurora-A/PinsLINKER/ Dlg spindle orientation pathway using induced cell polarity in S2 cells. Cell 138(6):1150–1163 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnston CA, Manning L, Lu MS, Golub O, Doe CQ, Prehoda KE (2013) Formin-mediated actin polymerization cooperates with mushroom body defect (Mud)-dynein during frizzled-dishevelled spindle orientation. J Cell Sci 126(19):4436–4444 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kikuchi K, Niikura Y, Kitagawa K, Kikuchi A (2010) Dishevelled, a Wnt signalling component, is involved in mitotic progression in cooperation with Plk1. EMBO J 29(20):3470–3483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiyomitsu T, Cheeseman IM (2013) XCortical dynein and asymmetric membrane elongation coordinately position the spindle in anaphase. Cell 154:391–402 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kotak S, Busso C, Gönczy P (2013) NuMA phosphorylation by CDK1 couples mitotic progression with cortical dynein function. EMBO J 32(18):2517–2529 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kraut R, Campos-Ortega JA (1996) inscrutable, a neural precursor gene of Drosophila, encodes a candidate for a cytoskeleton adaptor protein Dev Biol 174(1):65–81 [DOI] [PubMed] [Google Scholar]
- Kraut R, Chia W, Jan LY, Jan YN, Knoblich JA (1996) Role of inscuteable in orienting asymmetric cell divisions in Drosophila. Nature 383:50–55 [DOI] [PubMed] [Google Scholar]
- Langton PF, Colombani J, Chan EHYY, Wepf A, Gstaiger M, Tapon N (2009) The dASPP-dRASSF8 complex regulates cell-cell adhesion during Drosophila retinal morphogenesis. Curr Biol 19(23):1969–1978 [DOI] [PubMed] [Google Scholar]
- Le Borgne R, Schweisguth F (2003) Unequal segregation of neuralized biases Notch activation during asymmetric cell division. Dev Cell 5(1):139–148 [DOI] [PubMed] [Google Scholar]
- Lechler T, Fuchs E (2005) Asymmetric cell divisions promote stratification and differentiation of mammalian skin. Nature 437(7056):275–280 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lechler T, Mapelli M (2021) Spindle positioning and its impact on vertebrate tissue architecture and cell fate. Nat Rev Mol Cell Biol 22(10):691–708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee C-MM, Yang P, Chen L-CC, Chen C-CC, Wu S-CC, Cheng H-YY, Chang Y-SS (2011) A novel role of RASSF9 in maintaining epidermal homeostasis. PLoS ONE 6(3):e17867 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehner CF, O’Farrell PH (1989) Expression and function of Drosophila cyclin a during embryonic cell cycle progression. Cell 56(6):957–968 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehner CF, O’Farrell PH (1990) The roles of Drosophila cyclins A and B in mitotic control. Cell 61(3):535–547 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu B, Ackerman L, Jan LY, Jan YN (1999) Modes of protein movement that lead to the asymmetric localization of Partner of Numb during Drosophila neuroblast division. Mol Cell 4(6):883–891 [DOI] [PubMed] [Google Scholar]
- Mangione F, Titlow J, Maclachlan C, Gho M, Davis I, Collinson L, Tapon N (2023) Co-option of epidermal cells enables touch sensing. Nat Cell Biol 25(4):540–549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morin X, Bellaïche Y (2011) Mitotic spindle orientation in asymmetric and symmetric cell divisions during animal development. Dev Cell 21(1):102–119 [DOI] [PubMed] [Google Scholar]
- Nakajima YI, Lee ZT, McKinney SA, Swanson SK, Florens L, Gibson MC (2019) Junctional tumor suppressors interact with 14-3-3 proteins to control planar spindle alignment. J Cell Biol 218(6):1824–1838 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakajima YI, Meyer EJ, Kroesen A, McKinney SA, Gibson MC (2013) Epithelial junctions maintain tissue architecture by directing planar spindle orientation. Nature 500(7462):359–362 [DOI] [PubMed] [Google Scholar]
- Neville KE, Finegan TM, Lowe N, Bellomio PM, Na D, Bergstralh DT (2023) The Drosophila mitotic spindle orientation machinery requires activation, not just localization. EMBO Rep 24(3):1–15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osswald M, Morais-de-Sá E (2019) Dealing with apical–basal polarity and intercellular junctions: a multidimensional challenge for epithelial cell division. Curr Opin Cell Biol 60:75–83 [DOI] [PubMed] [Google Scholar]
- Perdigoto CNLR, Gervais L, Overstreet E, Fischer J, Guichet A, Schweisguth F (2008) Overexpression of Partner of Numb induces asymmetric distribution of the PI4P 5-kinase skittles in mitotic sensory organ precursor cells in Drosophila. PLoS ONE 3(8):1–6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez-Riverol Y, Bandla C, Kundu DJ, Kamatchinathan S, Bai J, Hewapathirana S, John NS, Prakash A, Walzer M, Wang S, Vizcaíno JA (2025) The PRIDE database at 20 years: 2025 update. Nucleic Acids Res 53(D1):D543–D553 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Portegijs V, Fielmich L, Galli M, Schmidt R, Mu J, Mourik T Van, Akhmanova A, Heck AJR, Boxem M, Van Den Heuvel S (2016) Multisite phosphorylation of NuMA-related LIN-5 controls mitotic spindle positioning in C. elegans. PLoS Genet 12:e1006291 [DOI] [PMC free article] [PubMed]
- Poulson ND, Lechler T (2010) Robust control of mitotic spindle orientation in the developing epidermis. J Cell Biol 191(5):915–922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ragkousi K, Gibson MC (2014) Cell division and the maintenance of epithelial order. J Cell Biol 207(2):181–188 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reeves N, Posakony JW (2005) Genetic programs activated by proneural proteins in the developing Drosophila PNS. Dev Cell 8(3):413–425 [DOI] [PubMed] [Google Scholar]
- Roegiers F, Younger-Shepherd S, Jan LY, Jan YN (2001a) Bazooka is required for localization of determinants and controlling proliferation in the sensory organ precursor cell lineage in Drosophila. Proc Natl Acad Sci USA 98(25):14469–14474 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roegiers F, Younger-Shepherd S, Jan LY, Jan YN (2001b) Two types of asymmetric divisions in the drosophila sensory organ precursor cell lineage. Nat Cell Biol 3(1):58–67 [DOI] [PubMed] [Google Scholar]
- Saredi A, Howard L, Compton DA (1997) Phosphorylation regulates the assembly of NuMA in a mammalian mitotic extract. J Cell Sci 110(11):1287–1297 [DOI] [PubMed] [Google Scholar]
- Schaefer M, Schevchonko A, Shevchenko A, Knoblich JA (2000) A protein complex containing inscuteable and the Gα-binding protein pins orients asymmetric cell divisions in Drosophila. Curr Biol 10(7):353–362 [DOI] [PubMed] [Google Scholar]
- Schober M, Schaefer M, Knoblich JA (1999) Bazooka recruits Inscuteable to orient asymmetric cell divisions in Drosophila neuroblasts. Nature 402(6761):548–551 [DOI] [PubMed] [Google Scholar]
- Schweisguth F (2015) Asymmetric cell division in the Drosophila bristle lineage: from the polarization of sensory organ precursor cells to Notch-mediated binary fate decision. Wiley Interdiscip Rev: Dev Biol 4(3):299–309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ségalen M, Johnston CA, Martin CA, Dumortier JG, Prehoda KE, David NB, Doe CQ, Bellaïche Y (2010) The Fz-Dsh planar cell polarity pathway induces oriented cell division via Mud/NuMA in Drosophila and Zebrafish. Dev Cell 19(5):740–752 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seldin L, Poulson ND, Foote HP, Lechler T (2013) NuMA localization, stability, and function in spindle orientation involve 4.1 and Cdk1 interactions. Mol Biol Cell 24(23):3651–3662 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siller KH, Cabernard C, Doe CQ (2006) The NuMA-related Mud protein binds Pins and regulates spindle orientation in Drosophila neuroblasts. Nat Cell Biol 8(6):594–600 [DOI] [PubMed] [Google Scholar]
- Siller KH, Doe CQ (2009) Spindle orientation during asymmetric cell division. Nat Cell Biol 11(4):365–374 [DOI] [PubMed] [Google Scholar]
- Simpson P (2007) The stars and stripes of animal bodies: evolution of regulatory elements mediating pigment and bristle patterns in Drosophila. Trends Genet 23(7):350–358 [DOI] [PubMed] [Google Scholar]
- Smith CA, Lau KM, Rahmani Z, Dho SE, Brothers G, She YM, Berry DM, Bonneil E, Thibault P, Schweisguth F, Le Borgne R, McGlade CJ (2007) aPKC-mediated phosphorylation regulates asymmetric membrane localization of the cell fate determinant Numb. EMBO J 26(2):468–480 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sunchu B, Cabernard C (2020) Principles and mechanisms of asymmetric cell division. Development 147(13):dev167650 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tatum NJ, Endicott JA (2020) Chatterboxes: the structural and functional diversity of cyclins. Semin Cell Dev Biol 107(May):4–20 [DOI] [PubMed] [Google Scholar]
- Taylor J, Abramova N, Charlton J, Adler PN (1998) Van Gogh: a new Drosophila tissue polarity gene. Genetics 150(1):199–210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tepass U (2012) The apical polarity protein network in drosophila epithelial cells: regulation of polarity, junctions, morphogenesis, cell growth, and survival. Annu Rev Cell Dev Biol 28(1):655–685 [DOI] [PubMed] [Google Scholar]
- Vernì P, Somma MP, Gunsalus KC, Bonaccorsi S, Belloni G, Goldberg ML, Gatti M (2004) Feo, the Drosophila homolog of PRC1, is required for central-spindle formation and cytokinesis. Curr Biol 14:1569–1575 [DOI] [PubMed] [Google Scholar]
- Whittaker AJ, Royzman I, Orr-Weaver TL (2000) Drosophila double parked: a conserved, essential replication protein that colocalizes with the origin recognition complex and links DNA replication with mitosis and the down-regulation of S phase transcripts. Genes Dev 14(14):1765–1776 [PMC free article] [PubMed] [Google Scholar]
- Williams SE, Ratliff LA, Postiglione MP, Knoblich JA, Fuchs E (2014) Par3-mInsc and Gα i3 cooperate to promote oriented epidermal cell divisions through LGN. Nat Cell Biol 16(8):758–769 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wirtz-Peitz F, Nishimura T, Knoblich JA (2008) Linking cell cycle to asymmetric division: aurora-A phosphorylates the par complex to regulate numb localization. Cell 135(1):161–173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wodarz A, Ramrath A, Kuchinke U, Knust E (1999) Bazooka provides an apical cue for inscuteable localization in Drosophila neuroblasts. Nature 402(6761):544–547 [DOI] [PubMed] [Google Scholar]
- Woods DF, Bryant PJ (1991) The discs-large tumor suppressor gene of Drosophila encodes a guanylate kinase homolog localized at septate junctions. Cell 66(3):451–464 [DOI] [PubMed] [Google Scholar]
- Yang Y, Liu M, Li D, Ran J, Gao J, Suo S, Sun-Cong S, Zhou J (2014) CYLD regulates spindle orientation by stabilizing astral microtubules and promoting dishevelled-NuMA-dynein/ dynactin complex formation. Proc Natl Acad Sci USA 111(6):2158–2163 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y, Mlodzik M (2015) Wnt-frizzled/planar cell polarity signaling: cellular orientation by facing the wind (Wnt). Annu Rev Cell Dev Biol 31:623–646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu F, Morin X, Cai Y, Yang X, Chia W (2000) Analysis of partner of inscuteable, a novel player of Drosophila asymmetric divisions, reveals two distinct steps in inscuteable apical localization. Cell 100(4):399–409 [DOI] [PubMed] [Google Scholar]
- Zheng Z, Wan Q, Meixiong G, Du Q (2014) Cell cycle-regulated membrane binding of NuMA contributes to efficient anaphase chromosome separation. Mol Biol Cell 25(5):606–619 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Source data (imaging and biochemistry) are associated with the manuscript. The Mass Spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE (Perez-Riverol et al, 2025) partner repository (https://www.ebi.ac.uk/pride/) with the dataset identifier PXD060813.
The source data of this paper are collected in the following database record: biostudies:S-SCDT-10_1038-S44318-025-00420-5.