

Abstract
Background
HSV-1 infections are pervasive in the general population of the world, and the gradual development of resistance to conventional antiviral therapies underlines the need to investigate alternative therapies. The present paper investigates the antiviral potential of a bicyclic sesquiterpene, β-caryophyllene (BCP), present in essential oils against Human alpha herpesvirus 1 (HSV-1).
Methods
The antiviral action of β-caryophyllene (BCP) was investigated using plaque reduction assays, molecular docking, molecular dynamics simulations, and surface plasmon resonance (SPR) analysis to explore its interactions with key surface glycoproteins of HSV-1 that are essential for viral entry and fusion: Glycoprotein B (gB), Glycoprotein C (gC), and Glycoprotein D (gD).
Results
BCP exhibited excellent antiviral activity, particularly during the pre-treatment phase, with a high selectivity index (SI) value of 35.86. Molecular docking analysis demonstrated that BCP binds strongly to glycoprotein B (gB) of HSV-1, with a binding energy of − 7.07 kcal/mol. These computational predictions were further validated by surface plasmon resonance (SPR) experiments, which confirmed a strong and specific interaction between BCP and gB, with a calculated equilibrium dissociation constant (KD) of (4.26 ± 0.02) × 10⁻⁶ M, supporting its proposed mechanism of action.
Discussion
Therefore, it was inferred that BCP interferes with viral entry and replication. Further confirmation through molecular dynamics simulation revealed less fluctuation in the Domain II region of gB, which might impede its fusion function. The SPR assay indeed confirmed that the BCP binds strongly to gB with a fast association and a slow dissociation rate, suggesting stability in this interaction.
Conclusion
These findings suggest that BCP is a promising complementary therapy for HSV-1 infection, with the potential to reduce the emergence of drug resistance. However, further preclinical and clinical studies are warranted to fully validate its therapeutic efficacy and safety before patient application.
Supplementary Information
The online version contains supplementary material available at 10.1186/s12906-025-04964-1.
Keywords: HSV-1, β-Caryophyllene, Antiviral, Resistance, Essential oils
Introduction
Human alphaherpesvirus 1 (HuAHV-1), also known as herpes simplex virus type 1 (HSV-1), is one of the most prevalent and highly infectious members of the Orthoherpesviridae. While it primarily presents with mucocutaneous lesions, particularly in the orofacial region, HSV-1 can lead to severe and potentially life-threatening complications such as encephalitis and disseminated infections, especially in immunocompromised individuals [1]. Beyond its classical clinical manifestations, HSV-1 has also been implicated in more severe disease outcomes, including sexually transmitted infections (STIs) and certain malignancies, highlighting its broader significance to public health. Although antiviral agents such as acyclovir (ACV) remain the cornerstone of HSV-1 management, prolonged use, particularly in immunocompromised patients has contributed to the emergence of drug-resistant viral strains, underscoring the need for alternative therapeutic strategies [2] that can adequately address the challenges of resistance [3, 4].
Given the increasing demand for alternative therapeutic options, natural compounds, especially essential oils have attracted considerable interest for their broad-spectrum pharmacological effects, including antiviral, anti-inflammatory, and immunomodulatory properties [5, 6]. Melissa officinalis, commonly known as lemon balm, is a medicinal plant recognized for its antimicrobial and anti-inflammatory properties. Although these bioactivities have been widely reported, their underlying mechanisms, particularly in relation to HSV-1 remain insufficiently elucidated. A deeper understanding of these mechanisms could facilitate the development of novel therapeutic strategies aimed at enhancing existing antiviral treatments and mitigating the emergence of drug-resistant strains.. One compound of particular interest is β-caryophyllene (BCP), a bicyclic sesquiterpene with significant pharmaceutical potential. Melissa officinalis, one of the sources of BCP, has long been recognized for its antiviral properties [7]. Notably, BCP has demonstrated broad-spectrum inhibitory activity against a diverse range of pathogens, including viruses [8–10].
This study investigates the potential therapeutic applications of selected essential oils against HSV-1 through an integrated approach comprising plaque reduction assays, molecular docking, molecular dynamics simulations (MDS), and surface plasmon resonance (SPR) analysis. By focusing on the molecular mechanisms, specifically the interactions between essential oil components and key HSV-1 glycoproteins such as gB, gC, and gD, this research aims to elucidate how these compounds inhibit viral infectivity. A deeper understanding of these interactions may support the use of essential oils or their bioactive constituents as complementary antiviral therapies, with the potential to reduce drug resistance and enhance treatment efficacy in HSV-1 infections.
Material and methods
Computational methods
Protein preparation
The target proteins selected for this study include surface glycoproteins of the herpesvirus, as they are among the main proteins involved in viral entry and cell fusion. The three-dimensional structures of glycoprotein B (gB) and glycoprotein D (gD) were obtained from the Protein Data Bank (PDB IDs: 2GUM and 3U82, respectively) [11]. Since no experimentally resolved structure was available for glycoprotein C (gC), its 3D model was generated using the I-TASSER-MTD server.I-TASSER-MTD employs a deep learning-guided hierarchical approach for multi-domain protein modeling [12]. The structures were prepared for docking using AutoDockTools: removing water molecules, adding polar hydrogen atoms, and assigning Gasteiger charges. Any non-standard residues were mutated to their standard counterparts to ensure the docking conditions are ideal. Next, ModRefiner was used to optimize the stereochemistry of the protein structures to be sure that any disfavored dihedral angles were corrected for optimal geometry [13].
Ligand preparation
The active terpenoid compound, including BCP 3D structures, were retrieved from the PubChem database (PubChem CID: 20,831,623). Converting these to PDB format, ligand was optimized for the process of AutoDock docking using AutoDockTools [14], which includes adding of polar hydrogen atoms, assigning Gasteiger charges, and defining torsional degrees of freedom for each ligand in order to allow flexible docking.
Docking protocol
Thereafter, blind docking was performed to investigate all the potential binding sites on the surface glycoproteins. Since active sites were not pre-defined, the whole surface of the protein was considered a potential binding site. AutoDock Vina 1.2 was used for the docking studies due to the computational efficiency in estimation of binding affinities [15]. Blind docking was conducted to explore the entire surface of the target glycoproteins, with the grid box dimensions set to 82 × 126 × 100 Å3 centered at (x = 43.415, y = 113.979, z = 72.45). The exhaustiveness parameter was set to 16 to ensure comprehensive sampling of potential binding modes. Multiple equilibrated conformations for each ligand were generated for docking analyses.
Molecular dynamics simulation
Eventually, MD simulations were performed for the most favored protein–ligand complexes obtained from the docking studies using Gromacs 2021 software with a force field described by ff99SB + ILDN [16]. The system was solvated in a TIP3P water box and neutralized by adding either sodium or chloride ions. Force field parameters of the ligands were determined using the General Amber Force Field via the Antechamber module in AmberTools [17, 18]. Partial charges were calculated using AM1-BCC and converted into GROMACS format for performing MD. Energy minimization was performed using the steepest descent method for 10,000 steps to eliminate any steric clashes. In two steps, the systems were first equilibrated: first under NVT conditions at 310 K for 200 picoseconds and second under NPT conditions at 1 bar for 200 picoseconds using the V-rescale thermostat and Parrinello-Rahman barostat, respectively. Production was single run for 40 ns with a time step of 2 femtoseconds. Periodic boundary conditions had been applied in order for the system to mimic an infinite system. Electrostatic interactions were calculated using the Particle Mesh Ewald method [19], a bond length constraint through the LINCS algorithm, while simulation data was written every 4 picoseconds for detailed analysis..
Experimental methods
Materials
This study utilized β-Caryophyllene (BCP, CAS Number 87–44-5) and Citral (CAS Number 5392–40-5), sourced from Sigma-Aldrich (Germany) and diluted in ethanol for use in cell culture media. The essential oil from Melissa officinalis was obtained from Primavera Life (Germany) and had previously been analyzed via gas chromatography, revealing 17.31% BCP and 20.13% Citral [20]. Acyclovir (ACV, CAS Number 59277–89-3) was dissolved in DMSO and used as a control in antiviral assays. All experiments were carried out using the A549 cell line under standard growth conditions. A549 cells were obtained from the Pasteur Institute of Iran Cell Bank. As these are commercially available, established cell lines, their use in this study did not require additional ethical approval.The HSV-1 strain and an ACV-resistant laboratory strain, HSV-1/R, were used in the study for testing the antiviral effects of the compounds.
MTT assay
The cytotoxicity of the BCP, Citral, Melissa officinalis essential oil, and a combination of BCP and Citral (17.31%/20.13%) was evaluated using the MTT assay [21]. A549 cells were cultured in 96-well plates with serial dilutions of the tested compounds being added in. Concentrations varied from 10 to 500 µg/ml for BCP and Citral, from 0.0003% to 0.03% (v/v) for Melissa officinalis oil and from 0.003% to 0.075% (v/v) for the BCP-Citral mixture, respectively. The MTT reagent was applied and the resulting formazan crystals were dissolved in DMSO after a 72 h incubation at 37 °C. The optical density was measured at 570 nm to determine the viability of cells relative to control wells.
Plaque Reduction Assay (PRA)
Antiviral activity was assessed by plaque reduction assay (PRA) [8, 22] for BCP, Citral, Melissa officinalis essential oil, and a combination of BCP and Citral (17.31%/20.13%). In general, cells were seeded at 2.5 × 104 cells per well in 12-well culture plates and incubated until reaching approximately 96% confluency. Subsequently, the cells were inoculated with the virus (HSV-1) at an MOI of 0.0002, carried out in three phases, pre-treatment, co-treatment, and post-treatment [8]. After 3 days of incubation, the cells were fixed with 10% formalin and stained with 1% crystal violet, followed by plaque counting. Compound concentrations ranged from 2–60 µg/ml for BCP and Citral, 0.0002–0.005% (v/v) for Melissa officinalis oil, and 0.003–0.075% (v/v) for the BCP–Citral mixture. Three control groups were included in each assay: antiviral controls, mock controls, and virus controls. The dose–response curves provided the inhibitory concentrations (IC50), and the ratio of cytotoxic concentration to IC50 defined the selectivity index.
Combination assay
Post-treatment was performed for both the ACV-sensitive and -resistant strains in a similar type of study was done previously in the PRA section, to understand the combination effect of BCP and ACV. CP was added at final concentrations of 2 to 60 µg/ml, with ACV added at a final concentration of 3 µg/ml. Corresponding SIs were compared to assess the interaction.
Native protein preparation
Glycoprotein B from herpes simplex virus type 1 was isolated and prepared by a series of standardized methods for ensuring the native conformation of the protein for subsequent analysis. HSV-1 was grown in A549 cell lines, upon which the virus was harvested and pelleted down using established centrifugation techniques as in previous studies [23].The viral pellet was then subjected to membrane protein extraction using the MEM-PER™ Plus Membrane Protein Extraction Kit (Thermo Fisher Scientific, Catalog No. 89842) by following the manufacturer's protocol. This protocol achieved good separation of cytoplasmic and membrane protein fractions with a bias toward membrane glycoprotein enrichment [24]. These proteins, which included glycoprotein B, were then separated by Native PAGE on a 4–8% gradient gel under non-denaturing conditions to maintain protein structure. Proteins were separated and visualized following standard procedures [25].
Then, the gel corresponding to the band of gB was excised and further purified by electroelution, which efficiently elutes the proteins out of the gel matrix into a buffer solution. Afterwards, the protein was concentrated by lyophilization [26]. The protein concentration was determined by UV–Vis spectroscopy at 280 nm using a known extinction coefficient for gB according to Gill and von Hippel [27]. Finally, gB expression and purification were verified by Western blot analysis. Standard methods were used for the Western blotting with mouse monoclonal anti-glycoprotein B antibody (Abcam, Catalog No. ab6506) [28]. Indeed, this elaborate procedure further ensured that glycoprotein B would be isolated in its native form and thus befitting structural and functional studies.
Surface Plasmon Resonance (SPR)
Kinetic interactions of glycoproteins of the herpes virus with terpenoid compounds were studied using the multi-parameter SPR device MP-SPR Navi 210 A (BioNavis Ltd., Tampere, Finland) with the Kretschmann prism configuration. Experiments were performed at a fixed angle of incidence under the flow rate of 30 µL/min; the temperature of the sensor was 37 °C. The cleaned gold-coated SPR slides were further modified by immersing them in 1 mM of 11-MUA(11-mercaptoundecanoic acid) for 20 h, obtaining a self-assembled monolayer on the surface, and then rinsed with ethanol and PBS buffer [29].
After establishing a baseline with PBS buffer in the SPR device, the surface was activated using a mixture of EDC and NHS. Glycoprotein (50 µg/mL) was injected for covalent attachment, and unreacted sites were blocked with ethanolamine-HCl. various concentrations BCP (10 nM to 100 µM) were injected over the glycoprotein-immobilized surface, with one flow cell used for sample injection and another as a reference. The surface was regenerated between injections with 10 mM glycine–HCl (pH 2.0). All experiments were performed in triplicate, and kinetic parameters were fitted using a 1:1 Langmuir binding model to calculate the association rate constant (ka), dissociation rate constant (kd), and equilibrium dissociation constant (KD) [29].
Statistical analysis
All experiments were performed in triplicate unless otherwise stated. Data are expressed as mean ± standard deviation (SD). Statistical analyses were conducted using GraphPad Prism version 8.0 (GraphPad Software, USA). Comparisons between two groups were made using Student’s t-test. Comparisons among multiple groups were analyzed using one-way analysis of variance (ANOVA) followed by Tukey’s post-hoc test for multiple comparisons. A p-value less than 0.05 was considered statistically significant.
Results
Computational methods
Molecular docking
Molecular docking for BCP were conducted to estimate its binding affinity toward HSV-1 glycoproteins B (gB), D (gD), and C (gC) with AutoDock Vina because the probable site of action could be the viral surface glycoproteins as results of previous sections have shown its effect in both pre-treatment and co-treatment stages, along with an effect on the reduction in plaque diameter. The binding affinity of BCP to different viral glycoproteins was estimated by the finding that gB showed stronger binding, which was − 7.07 ± 0.33 kcal/mol. This was followed by gD, with a binding energy of − 4.57 ± 0.33 kcal/mol, and the least was gC at − 3.77 ± 0.21 kcal/mol (Fig. 1).
Fig. 1.
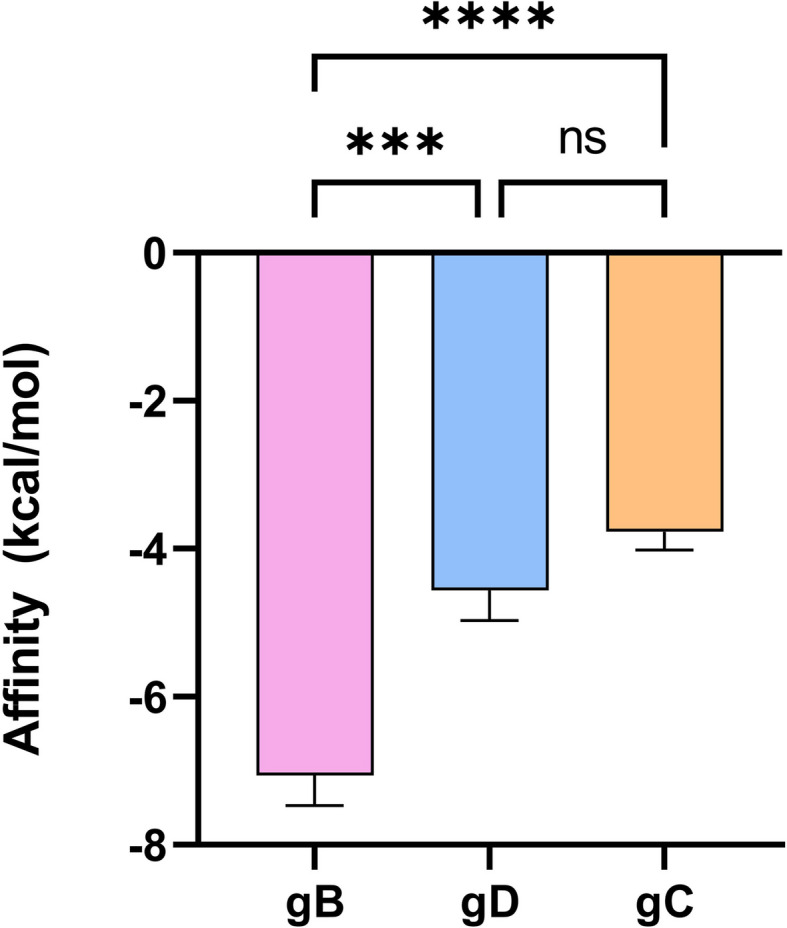
Affinity (kcal/mol) of a BCP for glycoproteins B (gB, pink), D (gD, blue), and C (gC, orange) of HSV-1. BCP shows the highest affinity for gB, followed by gD and gC. The differences in affinity between gB and gD, as well as between gB and gC, are statistically significant (***p < 0.001 and ****p < 0.0001, respectively). However, the affinity between gD and gC is not significantly different (ns), indicating similar binding strength to these glycoproteins
Detailed interaction analysis revealed that β-caryophyllene forms multiple van der Waals contacts with residues such as VAL170, MET183, GLN172, TYR265, and PHE186 across chains A, B, and C of gB. A Pi-Alkyl interaction with PHE186 was observed, contributing significantly to ligand stabilization. The interaction distances ranged from 2.5 to 3.0 Å, indicating a stable binding conformation (Fig. 2) (Supplementary file Fig. 1).
Fig. 2.

Visualization of the interaction network between beta-caryophyllene (BCP) and the gb binding pocket. Van der Waals interactions involve residues such as VAL A:170, MET A:183, GLN A:172, TYR A:265, GLY A:184, GLU A:187, ILE A:185, PHE B:186, and their equivalents in chains B and C. Additionally, a Pi-Alkyl interaction is observed with PHE A:186, which plays a key role in ligand orientation and stabilization. Interaction distances range from 2.5 to 3.0 Å, indicative of a snug fit within the binding pocket, typical for van der Waals forces
Molecular dynamic
To better understand the interaction between BCP and gB, molecular dynamics simulations were performed for this complex. Subsequently, to ensure the structural stability of the protein under different conditions, the RMSD (root mean square deviation) related to the alpha carbon atoms of the protein was calculated and extracted during the simulation time (40 ns) compared to the initial structure.
Figure 3a displays the RMSD changes for the entire gB and each of its chains. Although the RMSD values for all graphs increase at the start of the simulation and stabilize after approximately 3 ns, there are slight differences between them. The graphs of total protein and chain C also stabilize at about 0.3 nm after 3 ns with minor fluctuations until the end of the simulation, whereas chains A (red) and B (green) reach their RMSD value at about 0.24 nm in approximately 2 ns and remain stable for the remaining period of the simulation. Overall, this indicates that the protein achieves stability after around 3 ns, with some variation in the behavior of individual chains.
Fig. 3.

a Protein RMSD changes in the presence of ligand during molecular dynamics simulation time. Black represents the total protein, while red, green, and blue correspond to chains A, B, and C, respectively. b RMSF of protein amino acids in the absence of the ligand. The horizontal axis represents the number of amino acids, and the vertical axis indicates the fluctuation value in nanometers. The flexibility value of each chain was calculated separately. c The graph of eigenvalues against the corresponding eigenvector index obtained from the fluctuation covariance matrix of alpha carbon atoms during the simulation. d PCA diagram showing the protein phase space in the presence and absence of the ligand along PC1 and PC2. e RMSF plot of the protein in PC1 in the presence of the ligand across all three chains
The dynamic behavior of alpha carbon atoms in the structure contains enough information to investigate important movements in proteins and reflects the general movements of the structure. Therefore, the Root Mean Square Fluctuation (RMSF) of alpha carbon atoms was considered to investigate the movements and structural flexibility. In this part, the structural flexibility of the protein in the absence of BCP was analyzed (Fig. 3b). The amount of flexibility in all three chains is completely similar and, in most areas, it is around the value of 0.1 nm and only in the area of amino acids 490–495 the amount of flexibility is high. This region is related to amino acids without regular secondary structure in the gB structure.
Fig. 4.

a Selectivity Index (SI) of Melissa oil (red), terpenoids mixture (blue), beta-caryophyllene (BCP, purple), and citral (green) across pre-treatment, co-treatment, and post-treatment phases against HSV-1, where BCP consistently demonstrated a significantly higher SI in all phases (****p < 0.0001), underscoring its superior antiviral efficacy. b The SI of BCP was compared against acyclovir-susceptible (BCP/S, green) and acyclovir-resistant (BCP/R, purple) HSV-1 strains, both alone and in combination with ACV, revealing that the combination of ACV and BCP significantly enhanced the SI in the acyclovir-susceptible strain (ACV + BCP/S, pink) compared to either treatment alone (****p < 0.0001). c Reduction in plaque diameter of HSV-1 was observed in the presence of BCP alone (Day 3R/BCP, beige) and in combination with ACV (Day 3R/ACV + BCP, red) across varying concentrations (0, 5, 10, 15, and 20 µg/ml), where the combination treatment consistently resulted in a significantly smaller plaque diameter, indicating a synergistic effect that enhanced antiviral activity (*p < 0.05, ****p < 0.0001)
The nature of such movements of various parts of the protein is influenced by a number of factors like temperature changes, mutagenicity, and other influences. Principal Component Analysis was performed to make a more detailed analysis of the simulation of the protein in the presence of the ligand and to predict large-scale collective motions. These are the major elements of the protein, given by the covariance matrix of eigenvectors, representing variations in the path because of the simulation. Those changes, along with the eigenvalue spectrum obtained from the diagonalization of the covariance matrix of alpha carbon atom fluctuations in the presence of the ligand, are shown in Fig. 3c. The eigenvalues reflect coordinated and large-scale motions that decrease in magnitude rapidly. The other eigenvectors are related to more localized and limited movements within the protein. Each eigenvector has a so-called principal component related to it. The first two PCs, PC1 and PC2, have the largest range of movements, with 8.41106 nm2 and 7.29389 nm2, respectively. This indicates that the range of movements in PC1 and PC2 is greater than in the other PCs (Fig. 3c) (Supplementary file Fig. 3).
To further examines the range and extent of protein movement in the presence of the BCP within the phase space, the trajectories from the first two principal components (PC1 and PC2) are depicted as a two-dimensional diagram in Fig. 3d. Phase space is a conceptual space where all possible states of a system (in this case, the gB in the presence of a BCP) are represented. The protein appears in a large cluster and a small cluster along the PC1 axis, spread over a range of −6 to 6 nm, while in PC2, it is concentrated within a single large cluster from −6 to 8 nm (Fig. 3d).
Furthermore, the flexibility of the amino acids in PC1 space across each chain is shown. It is seen that the flexibility is below 0.1 nm for most of the amino acids, whereas notable flexibility is observed in amino acids 490–495. However, it is rather obvious from Fig. 3b and e that the gB, with BCP, has fluctuations far less than in their absence, particularly at domain 2, which is a disordered region where the area of fluctuation reduces by approximately 0.3 nm2 (Fig. 3e).
Experimental methods
MTT and PRA assay
Results of the cytotoxicity assay on A549 cells due to Melissa officinalis essential oil, its terpenoid components, and terpenoid mixture revealed a decrease in cell viability with an increase in the concentration of the reagents applied (Table 1, Fig. 4a). These concentrations were determined as the highest non-cytotoxic in this study: 40 µg/mL for Citral, 50 µg/mL for BCP, 0.005% for Melissa oil, and 0.003% for the Citral/BCP mixture. Antiviral activity evaluation also showed that more than 50% of the inhibitory concentrations of HSV-1 in most phases were below the maximal non-cytotoxic concentration (Table 1). It was also observed that all the tested reagents had concentration-dependent increases in their antiviral activities, and these results expressed as average virus reduction from three independent experiments were: maximum effect seen in the pre-treatment phase, slightly reduced in co- and post-treatment phases, with BCP having a maximal effect throughout the phases.. During the phase of virus treatment, BCP has shown the highest SI value of 35.86 (Fig. 4a). As expected, ACV demonstrated the highest antiviral activity during the infection phase, achieving 98.6% inhibition of viral infectivity, but it exhibited no effect during the pre-treatment and co-treatment phases. On the other hand, post-treatment with the essential oil or its constituent in overlay medium did not significantly decrease plaque formation compared with ACV (The complete dataset, including the results of post-treatment evaluations, is available in the public repository Mendeley Data (10.17632/k8jcgtt4mm.1.).
Table 1.
The cytotoxic concentration 50 (CC50) and inhibitory concentration 50% (IC50) of essential oils and selected terpenes against HSV-1
| Essential oil/Terpenes | Max.nocytotoxic concentration ± SD (%) | CC50 ± SD (%) | IC50 ± SD (%) Pre-treatment |
IC50 ± SD (%) Co-treatment |
IC50 ± SD (%) Post-treatment |
|---|---|---|---|---|---|
| BCP | 50.2 ± 5.2 | 203 ± 9.6 | 5.66 ± 3.5 | 16.25 µg/ml ± 1.76 | 19.92 µg/ml ± 4.82 |
| Citral | 40.34 ± 8.3 | 55.84 ± 2.5 | 9.60 ± 4.6 | 49.71 µg/ml ± 8.38 | 21.23 µg/ml ± 7.36 |
| Melissa oil | 0.004 ± 7.4 | 0.0095 ± 8.4 | 0.00053% ± 2.31 | 0.0044% ± 2.14 | 0.0045% ± 2.1 |
| Terpenoid mixture | 0.006 ± 8.1 | 0.01867 ± 3.6 | 0.002367% ± 7.6 | 0.025% ± 5.16 | 0.0038% ± 7.1 |
| ACV | 998 µg/ml | 1536 µg/ml ± 8.23 | N.D | N.D | 2.48 ± 3.21 |
Combination assay
For BCP, antiviral activity was similarly dose-dependent as obtained for HSV-1/S. Inhibition of HSV-1/R infectivity occurred for different concentrations of BCP, ranging from 1 to 60 µg/mL, with significant inhibition at the higher concentrations. The calculated selectivity index for BCP on HSV-1/R was 13.4 ± 1.71.
The graph represents the mean and standard deviation of SI for the four groups: BCP/S, BCP/R, ACV + BCP/S, and ACV + BCP/R. The mean data collected for SI for each group were as follows: for BCP/S it was 10.20 ± 1.09, for BCP/R it was 12.5 ± 1.07, for ACV + BCP/R it was 13.4 ± 1.71, and for ACV + BCP/S it was 23.38 ± 0.51. There was no significant difference between the values obtained for BCP/S and BCP/R and between BCP/R and ACV + BCP/R. Although the combination of BCP with ACV showed a slight increase in the Selectivity Index of the acyclovir-resistant (ACV-R) virus compared to BCP alone, this increase was not statistically significant. In contrast, in the acyclovir-susceptible (ACV-S) virus group, the combination of BCP and ACV (ACV + BCP/S) resulted in a significantly higher Selectivity Index than all other groups, including BCP/S and BCP/R, indicating that the combination therapy is much more effective against susceptible strains (Fig. 4b).
The data shows the effects of varying BCP concentrations on viral plaque diameter under two different conditions: "Day 3 BCP/R" and "Day 3 ACV + BCP/R."As the concentration of BCP increases from 0 to 20 µg/mL, there is a noticeable reduction in plaque diameters for both conditions. For instance, under the"Day 3 BCP/R"condition, the plaque diameter was 0.38 mm without BCP, which decreased to 0.1263 mm at 20 µg/mL. Under"Day 3 ACV + BCP/R,"the plaque diameter started from 0.44 mm without BCP to 0.09 mm at 20 µg/mL(Table 2). As can be seen, a clear trend is obtained in these results, indicating that the higher concentration of BCP develops smaller viral plaque diameter in both conditions, hence illustrating dose dependence of a BCP inhibition effect on viral plaque formation (Fig. 4c). See supplementary material Fig. 4.
Table 2.
Plaque diameter measurements for day 3R/BCP and day 3R/ACV + BCP across different BCP concentrations
| Concentration of BCP | Day 3R/BCP | Day 3R/ACV + BCP |
|---|---|---|
| 0 | 0.38 | 0.44 |
| 5 | 0.26 | 0.16 |
| 10 | 0.18 | 0.12 |
| 15 | 0.16 | 0.09 |
| 20 | 0.12 | 0.09 |
Native protein preparation
HSV-1 glycoprotein B was purified using a series of standardized operations so that the protein native conformation would be prospectively kept for further analysis. Detailed procedures for virus propagation, extracting the protein, purification, and validation-including the use of membrane protein extraction kits, Native PAGE, and Western blot analysis-can be found in the supplementary materials, Supplementary Section S1. These approaches therefore provided native form glycoprotein B that is suitable for further structural and functional studies (Supplementary Fig. 5, 6 and 7).
Surface plasmon resonance
These kinetic determinations performed at 37 °C give explicit insight into the kinetics of binding. The five overlaid plots for different injections are included in the analysis, showing signals (RU) as a function of time, along with fitted curves for each injection. The derived kinetic parameters include an association rate constant, ka = 5.13949 ± 0.008 × 103 [1/M*s]; a dissociation rate constant, kd = 2.18749 ± 0.01 × 10–2 [1/s]; and an equilibrium dissociation constant, KD = 4.25624 ± 0.02 × 10⁻⁶ M. These are typical for a relatively high affinity involving fast association along with slow dissociation, which in turn implies that the complex between gB and BCP is rather stable. In addition, the consistency of these parameters for all injections has shown negligible standard deviations, further emphasizing the reliability of the measurements (Fig. 5). (Supplementary file Fig. 8 9 and 10).
Fig. 5.

Surface Plasmon Resonance (SPR) sensorgram showing the kinetic evaluation of ligand binding at 37 °C. The data represents five sequential injections at increasing concentrations from 10 nM to 100 µM. The black curve corresponds to 10 nM, red to 100 nM, blue to 1 µM, green to 10 µM, and yellow to 100 µM. Each color represents a different concentration, and the corresponding fitted curves are shown in gray. The gray curves represent individual fits for each injection concentration. Each concentration was injected once during the kinetic run shown here. The experiments were independently repeated three times, and the reported kinetic parameters (ka, kd, KD) represent the average of the triplicate measurements
Discussion
The highest non-cytotoxic concentration among all the tested compounds and the most potent antiviral activity for BCP were defined. Its cytotoxicity profile is more favorable when compared to all the other tested reagents'cytotoxicity profile. Moreover, BCP showed the highest SI, indicating a better balance between antiviral activity and cytotoxicity. By contrast, citral might display higher cytotoxicity, narrowing its therapeutic window [30, 31]. These findings are further appended by the previous works of the particular antiviral property of terpenes. Large-scale antiviral activities of essential oils were reported by Schnitzler et al. [6]. In addition, de Sousa et al. emphasized the broad-spectrum antiviral properties of BCP [32]. Research on determination of the antiviral activity of Melissa officinalis essential oil against HSV-1 infection has also been conducted by Allahverdiyev et a [33]. The high selectivity index obtained in BCP would therefore most probably indicate a therapeutic window within which it can safely be used, similar to other plant compounds [8].
Compared to previous studies that primarily focused on the antiviral effects of essential oils or crude plant extracts, our study provides a significant advancement by specifically β-caryophyllene (BCP) and comprehensively evaluating its direct antiviral potential against HSV-1. Unlike earlier reports that suggested possible antiviral activity without mechanistic confirmation, we employed a multi-faceted approach combining plaque reduction assays, surface plasmon resonance (SPR), molecular docking, and molecular dynamics simulations to validate the binding of BCP to HSV-1 glycoproteins, particularly glycoprotein B (gB). Furthermore, by comparing the effects of BCP on both acyclovir-susceptible and acyclovir-resistant strains, we highlighted its therapeutic promise and limitations.
In addition, our findings indicate that BCP may be the most crucial antiviral component within the studied essential oil. Despite the promising individual performance of BCP, the mixture of citral and BCP in proportions similar to their natural presence exhibited less antiviral activity than BCP alone. This suggests that other constituents in the essential oil may modulate BCP’s efficacy through synergistic or antagonistic interactions, underscoring the complexity of essential oil composition and its impact on overall antiviral effectiveness.
The Selectivity Index (SI) of β-caryophyllene (BCP) observed in our study (35.86) is significantly higher than that of common monoterpenes and essential oils, such as thymol (SI ≈ 7) and eugenol-rich clove oil (SI ≈ 1.4) [34, 35]. Similarly, carvacrol and clove oil have been reported to exhibit low SI values around 1.4 [35]. Studies by Astani et al. further demonstrated that while thymol and carvacrol exhibit notable antiviral activity, their therapeutic window is limited by higher cytotoxicity [8, 36]. In contrast, BCP demonstrates a much stronger therapeutic profile, combining a higher SI with lower cytotoxicity. Although certain plant-derived flavonoids, such as naringin (SI ≈ 400), rutin (SI ≈ 2000), and galangin (SI ≈ 400), exhibit even greater SI values [37], these compounds differ structurally and mechanistically from simple terpenoids. Moreover, eugenol, despite showing antiviral activity against HSV-1, is associated with a narrower therapeutic window due to high cytotoxicity [38], and similar limitations were reported for menthol, another antiviral terpene, which also demonstrated less favorable cytotoxicity profiles [39]. In summary, β-caryophyllene presents a strong balance between antiviral efficacy and cytotoxic safety compared to many commonly studied herbal compounds, reinforcing its potential as an anti-HSV-1 therapeutic agent.
In addition to these comparative findings, the combination therapy involving BCP and ACV did not significantly enhance the selectivity index over BCP alone in the ACV-resistant strain but did so in the ACV-susceptible strain. The absence of a strong synergistic effect between BCP and ACV in acyclovir-resistant (ACV-R) HSV-1 strains can be attributed to the underlying mechanisms of antiviral resistance. Resistance to ACV is primarily mediated through mutations in viral thymidine kinase (TK) or, less commonly, viral DNA polymerase, which reduce the activation or efficacy of ACV. As BCP mainly targets viral entry and fusion processes by interacting with surface glycoproteins rather than interfering with viral DNA replication or nucleotide metabolism, it does not directly overcome TK-associated resistance mechanisms. Consequently, although BCP exhibits independent antiviral activity, it does not synergize significantly with ACV in resistant strains where the ACV activation pathway is already compromised. This mechanistic distinction explains the differential outcomes observed in susceptible versus resistant virus strains.
Additionally, the notable reduction in viral plaque diameters indicates enhanced antiviral activity, suggesting that BCP might improve the overall antiviral effect of ACV through complementary mechanisms. The highly increased SI for the acyclovir-sensitive virus indicates that beta-caryophyllene has augmented the effectiveness of ACV in inhibiting the virus. Since the mechanisms of beta-caryophyllene do not include interference with the replication of the viral genome, its inhibition of viral attachment and fusion and cell-to-cell spread, together with ACV's inhibition of viral genome replication, results in a significant inhibition of virus infectivity. Also, in the use of combination therapy including BCP against ACV-resistant HSV-1 strains, it delays the viral spread and allows more sufficient time for the response of the host immune system [40].
These observations are further supported by previous studies, which have demonstrated that [41–44]. These findings agree with observations in the present study that combination therapy involving BCP and ACV reduced viral plaque diameters; hence, such therapies may hold promise in the enterprise for discovering improved antiviral therapies. Furthermore, it prevents the maturation of plaques, which helps reduce viral load and inhibit the spread of the virus since viral plaques are unable to develop and expand, a beneficial effect for those that have serious or recurrent HSV-1 infections [4, 40, 45].
BCP enhances the antiviral activity of the other compounds by acting on different viral processes and diminishing the inflammatory response due to viral infection [41, 45–47]. This synergistic action may be very useful in combination therapies planning for viral infection treatment. The dose of ACV could be minimized in its combination therapy with BCP to reduce the side effects which normally arise upon higher-dose administration of ACV. The use of a lower dose of ACV might decrease the selective pressure on the virus, thereby decreasing the likelihood of the appearance of resistant mutant strains [4]. This is particularly important in clinical settings where long-term antiviral therapy is needed, as it may help reduce the emergence of resistance and maintain the treatment's effectiveness over time.
Further confirmation of BCP's potential as a potent antiviral agent is given by the molecular docking simulations that it has the highest binding affinity to gB of HSV-1, which may imply interference with viral entry and cell-to-cell spread as its mode of antiviral action. The most significant interactions of BCP with residues at the gB binding site included van der Waals and Pi-alkyl interactions, providing stability and specificity to the binding. Further temporal behavior analysis of the complex BCP-gB was performed using molecular dynamics simulations.
The results obtained from these simulations indeed point out that the overall fluctuation across the gB protein is reduced upon binding of BCP, mostly in the segment of amino acids 490–495 belonging to DII. DII is indeed the disordered and flexible part in the structure of gB and is crucial for the execution of its function. A reduction of about 0.3 nm2 in RMSF at this point clearly indicates a loss of flexibility in this region by binding to BCP. Because DII is intrinsically flexible, this reduction may allow for the stabilization of the protein structure.Therefore, considering that conformational flexibility is a fundamental requirement for the activity of viral fusion proteins—including gB in herpesviruses, the F protein of respiratory syncytial virus (RSV), and hemagglutinin (HA) in influenza virus—such ligand-induced stabilization could interfere with their dynamic behavior, hinder membrane fusion, and ultimately reduce viral infectivity [48].
These interactions between BCP and gB have grave implications for the function of glycoprotein B and thus, in turn, for the virus HSV-1. Glycoprotein B is responsible for the initial attachment and entry of HSV-1 into host cells [49]. A binding of BCP to gB can counteract this process, thus preventing this virus from entering host cells. Glycoprotein B is also implicated in the cell-to-cell spread of the virus [50]. This eventually inhibits gB's function, thereby limiting the total infection of the virus inside the host due to a reduced ability of the virus for propagation inside it [51–53]. The reduction of viral entry along with the cell-to-cell spread is in agreement with the results obtained from the plaque reduction test. The highly reduced viral plaque diameters during combination therapy with ACV indicate that BCP effectively limits viral propagation.
Although gB is critical for viral attachment and entry, herpesvirus seems to use other ways for host cell entry. Our results also support such observation as, despite BCP presenting a good IC50 and SI, it was not able to inhibit the virus in any phase completely. Even with concentrations as low as 50%, it reduced plaque formation; likewise, even at higher concentrations of BCP, complete eradication of plaque was not achieved at any concentration. These findings were thus not included in the manuscript. Thus, herpesvirus uses alternative pathways for attachment, entry, and spread within the environment [54–56].
These results are in accord with the molecular docking and dynamics results of this study, and therefore provide a sound framework for the molecular basis of antiviral action by BCP. That demonstrates the need to use docking and dynamics simulations complementarily, as the former gives an initial snapshot of possible binding poses and interactions, while the latter offers insight about the stability and behavior of the complex in real time. These findings, as mentioned above, which are in concurrence with the strong plaque reduction antiviral properties, affirm the potency of BCP as an agent against HSV-1.
Considering the actual requirement to have native gB for the SPR assay, isolation and preparation of gB from herpes simplex virus type 1 (HSV-1) by using the Thermo Scientific™ Mem-PER™ Plus Membrane Protein Extraction Kit represent an important step forward in viral protein research. This is the first viral system being used; the kit has normally been used for eukaryotic cells. Therefore, this new approach represents not only the versatility and efficiency of the Mem-PER™ Plus kit but also opens new dimensions in studying viral membrane proteins in their native conformation. Isolation of gB in native structure thus set a precedent for other studies in the future and allowed easy investigation of surface glycoproteins by scientists on enveloped viruses. This methodological innovation simplifies the study of these critical viral components and provides a robust, reproducible framework that can be extended to various enveloped viruses, enhancing further our knowledge in viral biology and the mechanisms underlying viral infection.
SPR analysis focused on the interaction between BCP and HSV-1 gB. High binding affinity of BCP to gB was observed in the SPR data, which thus indicates that BCP can well hinder the virus from entering the host cell. The kinetic results showed that BCP manifestation of relatively strong binding kinetics was with a moderate association and dissociation rate constants (ka and kd, respectively). That means BCP binds to gB at an intermediate rate and is also dissociating at an intermediate rate, probably indicating an interaction that is neither too transient nor too stable. Based on these kinetic parameters, the interpretation came out that a high association rate constant, ka, signifies that the drug is associating with its target fairly quickly. In contrast, low values of the dissociation rate constant kd suggest stability in the drug-target complex and a slow process of dissociation. Its low KD value suggests high binding affinity, while its regular character falls within the range of rather strong affinity interactions, such as antibody-antigen bindings, whose KD values lie between 10–6 and 10–9 M. For practical purposes, though, beta-caryophyllene could said to be fast-associating and slow-dissociating. This type of binding behavior is sought in therapeutic contexts because, from a pharmacological point of view, it means that the drug begins to bind quickly to its target in question, hence ensuring rapid onset of action. Besides that, the slow rate of dissociation ensures that the drug lingers on its target longer, sustaining its therapeutic effect over a very long period of action without frequent re-dosing [57].
Despite conducting widespread searches, no studies investigating the interaction between BCP and gB of HSV-1 by SPR analysis were found; thus, this represents the uniqueness of the present study and the novelty of insights into the possible antiviral mechanisms BCP may exert against HSV-1. The binding kinetics of BCP is relatively strong, thus suggesting the possibility of its effective and sustained antiviral action. This balanced kinetic profile may indicate that BCP could stably interact with gB and could thus have the potential to block virus entry into a host cell for a prolonged period. Considering the binding kinetics, together with the effective antiviral properties, the clinical relevance of BCP can be enormous. Given the very high binding to viral glycoproteins and lingering antiviral activity, formulation as topical creams or ointments could be developed for BCP treatment against HSV-1 infection. It could also be combined with previously developed antiviral therapeutics like ACV for enhanced therapeutic action that acts in support of the kinetics from other antiviral agents with a resulting effective suppression of viral infectivity. Given this, a further possibility with BCP is its use as a prophylactic agent that prevents an infection by HSV-1 through the formation of a protective barrier by stable binding to viral glycoproteins and inhibiting viral entry.
Limitations
Although the in vitro findings of this study are promising, several limitations must be acknowledged before translating these results into clinical applications.
First, the pharmacokinetic profiles of β-caryophyllene (BCP) and other essential oil components may differ significantly in vivo, potentially impacting their antiviral efficacy. This highlights the complexity of essential oil interactions and underscores the need to evaluate individual constituents both independently and in combination to fully elucidate their therapeutic potential.
Second, while BCP demonstrated potential against antiviral drug-resistant HSV-1 strains, further validation across a broader panel of resistant isolates is essential to confirm its robustness.
Third, although the extraction of viral proteins such as gB was innovative, further optimization may be necessary to ensure complete structural and functional integrity, which is critical for assay reproducibility and reliability.Fourth, while combination therapy using BCP and acyclovir (ACV) showed promising preliminary results, further validation in animal models and ultimately in human clinical trials is needed to evaluate the long-term safety, pharmacokinetics, and therapeutic efficacy of this approach.Finally, the study lacked a positive control specifically targeting viral surface glycoproteins, which limited the ability to conduct direct comparative interaction analyses. Future studies should incorporate established ligands or inhibitors to strengthen mechanistic interpretations and enhance the translational relevance of the findings.
Future direction
Despite the promising findings demonstrated through plaque reduction assays, molecular docking, molecular dynamics simulations, and SPR analysis, further studies are necessary to fully validate the proposed mechanisms. Future research should include functional assays specifically targeting viral entry or fusion processes, such as ELISA-based entry assays or luciferase-reporter fusion assays, to provide direct evidence of BCP's antiviral action. In addition, although our molecular docking results highlighted the critical role of the PHE A186 residue in ligand stabilization, confirmation through site-directed mutagenesis and subsequent biochemical characterization would be instrumental in substantiating the structural and functional significance of this residue. Moreover, broader investigations should aim to elucidate the molecular mechanisms by which BCP inhibits various phases of the viral life cycle, including interactions with other viral proteins. Evaluating combinations of BCP with other antiviral agents, optimizing dosages, ratios, and administration timing, as well as performing animal studies to confirm in vitro findings and conducting toxicological, efficacy, and pharmacokinetic analyses, will be critical to advancing BCP toward clinical application. Ultimately, the design and implementation of clinical trials will be essential to validate BCP’s therapeutic potential in human subjects, particularly against HSV-1 infections, and to assess long-term outcomes and safety profiles.
Conclusion
This study provides compelling evidence for the antiviral potential of β-caryophyllene (BCP), particularly against HSV-1 during the viral entry phase. The high selectivity index, potent antiviral activity, and strong binding affinity to key viral glycoproteins collectively support the promising role of BCP as a complementary antiviral agent. Although the combination with acyclovir did not significantly enhance the selectivity index, the observed reduction in viral plaque diameters suggests potential therapeutic synergy. Furthermore, molecular docking, molecular dynamics simulations, and surface plasmon resonance analyses offer a solid mechanistic framework for BCP’s antiviral action. Future studies should focus on validating these findings through viral entry inhibition assays and functional analyses of critical glycoprotein residues, as well as in vivo models, to comprehensively elucidate the antiviral mechanisms and therapeutic potential of BCP.
Supplementary Information
{kind=link}
{kind=link}
{kind=link}
Acknowledgements
We would like to thank the Razi Vaccine & Serum Research Institute for their support with the equipment and expertise in electroelution techniques. We also extend our gratitude to Tabriz University of Medical Sciences for providing the surface plasmon resonance (SPR) equipment and expertise. Their expertise and technical support were crucial to the success of this research.
Clinical trial number
Not applicable.
Abbreviations
- ACV
Acyclovir
- BCP
β-Caryophyllene
- HSV-1
Herpes Simplex Virus Type 1
- MDS
Molecular Dynamics Simulations
- PCA
Principal Component Analysis
- SPR
Surface Plasmon Resonance
- gB
Glycoprotein B
- gC
Glycoprotein C
- gD
Glycoprotein D
Glossary
- Selectivity Index (SI)
A measure of the effectiveness of a compound in inhibiting a specific pathogen relative to its toxicity to host cells.
- IC50
The concentration of a substance that inhibits 50% of infectivity.
- CC50
The concentration of a substance that causes a 50% reduction in cell viability or growth.
Authors’ contributions
Mohammad Yasaghi: Conceptualization, Formal analysis, Investigation, Methodology, Writing – original draft.Sahar Dolatabadi: Formal analysis, Investigation, Writing – original draft.Ahad Yamchi: Supervision, Resources, Validation, Writing – review and editing.Alijan Tabarraei: Project administration, Funding acquisition, Methodology, Writing – original draft.Abdolvahab Moradi: Methodology, Validation, Writing – review and editing.Seyedeh Delafruz Hosseini: Software, Writing – review and editing.
Funding
This research was conducted as part of the PhD thesis project of Mohammad Yasaghi, under a research grant (number 113584) approved by Golestan University of Medical Sciences. The funding for this study was provided by Golestan University of Medical Sciences and the Infectious Diseases Research Center, Golestan University of Medical Sciences, Gorgan, Iran.
Data availability
The data presented in this study are openly available in Mendeley Data at:
Bibliographic Details
DOI: 10.17632/k8jcgtt4mm.1
URL ID : https://data.mendeley.com/datasets/k8jcgtt4mm; 10.17632/k8jcgtt4mm.1; 10.17632/k8jcgtt4mm.1; https://data.mendeley.com/datasets/k8jcgtt4mm/1
AUTHOR(S)
Ahad Yamchi; Alijan Tabarraei; Seyyede Delafruz Hosseini; Mohammad Yasaghi; Abdolvahab Wahab Moradi; yasaghi, mohammad; Beryani Dolatabadi, Sahar; Yamchi, Ahad; Tabarraei, Alijan; Moradi, Abdolvahab ; Hosseini , Seyedeh Delafruz
Declarations
Ethics approval and consent to participate
This study was approved by the Ethic Committee of Golestan University of Medical Sciences (Ethic code: IR.Goums.REC.1402.197).
Consent for publication
All authors have read and agreed to the published version of the manuscript.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Ahad Yamchi, Email: Yamchi@gau.ac.ir.
Alijan Tabarraei, Email: alijant@gmail.com.
References
- 1.Preda M, Manolescu LSC, Chivu RD. Advances in alpha herpes viruses vaccines for human. Vaccines. 2023;11(6):1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Piret J, Boivin G. Resistance of herpes simplex viruses to nucleoside analogues: mechanisms, prevalence, and management. Antimicrob Agents Chemother. 2011;55(2):459–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.James SH, Kimberlin DW. Herpes simplex virus infections. Remington and Klein's infectious diseases of the fetus and newborn infant. 2025:745–64. e4.
- 4.Schalkwijk HH, Snoeck R, Andrei G. Acyclovir resistance in herpes simplex viruses: prevalence and therapeutic alternatives. Biochem Pharmacol. 2022;206:115322. [DOI] [PubMed] [Google Scholar]
- 5.Moradi MT, Rafieian-Kopaei M, Karimi A. A review study on the effect of Iranian herbal medicines against in vitro replication of herpes simplex virus. Avicenna J Phytomed. 2016;6(5):506–15. [PMC free article] [PubMed] [Google Scholar]
- 6.Schnitzler P. Essential oils for the treatment of herpes simplex virus infections. Chemotherapy. 2019;64(1):1–7. [DOI] [PubMed] [Google Scholar]
- 7.Petrisor G, Motelica L, Craciun LN, Oprea OC, Ficai D, Ficai A. Melissa officinalis: composition, pharmacological effects and derived release systems-a review. Int J Mol Sci. 2022;23(7):3591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Astani A, Reichling J, Schnitzler P. Comparative study on the antiviral activity of selected monoterpenes derived from essential oils. Phytother Res. 2010;24(5):673–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Klauke AL, Racz I, Pradier B, Markert A, Zimmer AM, Gertsch J, et al. The cannabinoid CB₂ receptor-selective phytocannabinoid beta-caryophyllene exerts analgesic effects in mouse models of inflammatory and neuropathic pain. Eur Neuropsychopharmacol. 2014;24(4):608–20. [DOI] [PubMed] [Google Scholar]
- 10.Gertsch J, Leonti M, Raduner S, Racz I, Chen JZ, Xie XQ, et al. Beta-caryophyllene is a dietary cannabinoid. Proc Natl Acad Sci U S A. 2008;105(26):9099–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Berman HM, Westbrook J, Feng Z, Gilliland G, Bhat TN, Weissig H, et al. The protein data bank. Nucleic Acids Res. 2000;28(1):235–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhou X, Zheng W, Li Y, Pearce R, Zhang C, Bell EW, et al. I-TASSER-MTD: a deep-learning-based platform for multi-domain protein structure and function prediction. Nat Protoc. 2022;17(10):2326–53. [DOI] [PubMed] [Google Scholar]
- 13.Xu D, Zhang Y. Improving the physical realism and structural accuracy of protein models by a two-step atomic-level energy minimization. Biophys J. 2011;101(10):2525–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Morris GM, Huey R, Lindstrom W, Sanner MF, Belew RK, Goodsell DS, et al. AutoDock4 and AutoDockTools4: automated docking with selective receptor flexibility. J Comput Chem. 2009;30(16):2785–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Trott O, Olson AJ. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem. 2010;31(2):455–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Abraham MJ, Murtola T, Schulz R, Páll S, Smith JC, Hess B, et al. GROMACS: high performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX. 2015;1–2:19–25. [Google Scholar]
- 17.Case DA, Aktulga HM, Belfon K, Cerutti DS, Cisneros GA, Cruzeiro VWD, et al. AmberTools. J Chem Inf Model. 2023;63(20):6183–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wang J, Wolf RM, Caldwell JW, Kollman PA, Case DA. Development and testing of a general amber force field. J Comput Chem. 2004;25(9):1157–74. [DOI] [PubMed] [Google Scholar]
- 19.Darden T, York D, Pedersen L. Particle mesh Ewald: An N⋅log(N) method for Ewald sums in large systems. J Chem Phys. 1993;98(12):10089–92. [Google Scholar]
- 20.Schnitzler P, Schuhmacher A, Astani A, Reichling J. Melissa officinalis oil affects infectivity of enveloped herpesviruses. Phytomedicine. 2008;15(9):734–40. [DOI] [PubMed] [Google Scholar]
- 21.Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods. 1983;65(1–2):55–63. [DOI] [PubMed] [Google Scholar]
- 22.Astani A, Reichling J, Schnitzler P. Melissa officinalis extract inhibits attachment of herpes simplex virus in vitro. Chemotherapy. 2012;58(1):70–7. [DOI] [PubMed] [Google Scholar]
- 23.Grosche L, Döhner K, Düthorn A, Hickford-Martinez A, Steinkasserer A, Sodeik B. Herpes simplex virus type 1 propagation, titration and single-step growth curves. Bio Protoc. 2019;9(23):e3441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Qoronfleh MW, Benton B, Ignacio R, Kaboord B. Selective enrichment of membrane proteins by partition phase separation for proteomic studies. J Biomed Biotechnol. 2003;2003(4):249–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wittig I, Braun H-P, Schägger H. Blue native PAGE. Nat Protoc. 2006;1(1):418–28. [DOI] [PubMed] [Google Scholar]
- 26.Tang X, Pikal MJ. Design of freeze-drying processes for pharmaceuticals: practical advice. Pharm Res. 2004;21(2):191–200. [DOI] [PubMed] [Google Scholar]
- 27.Gill SC, von Hippel PH. Calculation of protein extinction coefficients from amino acid sequence data. Anal Biochem. 1989;182(2):319–26. [DOI] [PubMed] [Google Scholar]
- 28.Mahmood T, Yang PC. Western blot: technique, theory, and trouble shooting. N Am J Med Sci. 2012;4(9):429–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fathi F, Ezzati Nazhad Dolatanbadi J, Rashidi MR, Omidi Y. Kinetic studies of bovine serum albumin interaction with PG and TBHQ using surface plasmon resonance. Int J Biol Macromol. 2016;91:1045–50. [DOI] [PubMed]
- 30.Bassolé IH, Juliani HR. Essential oils in combination and their antimicrobial properties. Molecules. 2012;17(4):3989–4006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Burt S. Essential oils: their antibacterial properties and potential applications in foods—a review. Int J Food Microbiol. 2004;94(3):223–53. [DOI] [PubMed] [Google Scholar]
- 32.de Sousa DP, Damasceno ROS, Amorati R, Elshabrawy HA, de Castro RD, Bezerra DP, et al. Essential oils: chemistry and pharmacological activities. Biomolecules. 2023;13(7):1144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Allahverdiyev A, Duran N, Ozguven M, Koltas S. Antiviral activity of the volatile oils of Melissa officinalis L. against Herpes simplex virus type-2. Phytomedicine. 2004;11(7–8):657–61. [DOI] [PubMed] [Google Scholar]
- 34.Sharifi-Rad J, Salehi B, Schnitzler P, Ayatollahi SA, Kobarfard F, Fathi M, et al. Susceptibility of herpes simplex virus type 1 to monoterpenes thymol, carvacrol, p-cymene and essential oils of Sinapis arvensis L., Lallemantia royleana Benth. and Pulicaria vulgaris Gaertn. Cell Mol Biol (Noisy-le-grand). 2017;63(8):42–7. [DOI] [PubMed]
- 35.Kiki MJ. In vitro antiviral potential, antioxidant, and chemical composition of clove (Syzygium aromaticum) essential oil. Molecules. 2023;28(6):2421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Astani A, Reichling J, Schnitzler P. Screening for antiviral activities of isolated compounds from essential oils. Evid Based Complement Alternat Med. 2011;2011:253643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Treml J, Gazdová M, Šmejkal K, Šudomová M, Kubatka P, Hassan STS. Natural products-derived chemicals: breaking barriers to novel anti-HSV drug development. Viruses. 2020;12(2):154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ulanowska M, Olas B. Biological properties and prospects for the application of eugenol-a review. Int J Mol Sci. 2021;22(7):3671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Taylor DJR, Hamid SM, Andres AM, Saadaeijahromi H, Piplani H, Germano JF, et al. Antiviral effects of menthol on coxsackievirus B. Viruses. 2020;12(4):373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Crumpacker CS. Mechanisms of resistance of antiviral drugs active against the human herpes virus. In: Mayers DL, Sobel JD, Ouellette M, Kaye KS, Marchaim D, editors. Antimicrobial drug resistance: mechanisms of drug resistance, vol. 1. Cham: Springer International Publishing; 2017. p. 479–89. [Google Scholar]
- 41.Ma L, Yao L. Antiviral effects of plant-derived essential oils and their components: an updated review. Molecules. 2020;25(11):2627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wani AR, Yadav K, Khursheed A, Rather MA. An updated and comprehensive review of the antiviral potential of essential oils and their chemical constituents with special focus on their mechanism of action against various influenza and coronaviruses. Microb Pathog. 2021;152:104620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Betancur-Galvis L, Jimenez-Jarava OJ, Rivas F, Mendoza-Hernández WE, González-Cardenete MA. Synergistic in vitro antiviral effect of combinations of ivermectin, essential oils, and 18-(Phthalimid-2-yl)ferruginol against arboviruses and herpesvirus. Pharmaceuticals. 2023;16(11):1602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Pourghanbari G, Nili H, Moattari A, Mohammadi A, Iraji A. Antiviral activity of the oseltamivir and Melissa officinalis L. essential oil against avian influenza A virus (H9N2). VirusDisease. 2016;27(2):170–8. [DOI] [PMC free article] [PubMed]
- 45.Chayrov RL, Stylos EK, Chatziathanasiadou MV, Chuchkov KN, Tencheva AI, Kostagianni AD, et al. Tailoring acyclovir prodrugs with enhanced antiviral activity: rational design, synthesis, human plasma stability and in vitro evaluation. Amino Acids. 2018;50(8):1131–43. [DOI] [PubMed] [Google Scholar]
- 46.Francomano F, Caruso A, Barbarossa A, Fazio A, La Torre C, Ceramella J, et al. β-Caryophyllene: a sesquiterpene with countless biological properties. Appl Sci. 2019;9(24):5420. [Google Scholar]
- 47.Jha NK, Sharma C, Hashiesh HM, Arunachalam S, Meeran MN, Javed H, et al. β-caryophyllene, a natural dietary CB2 receptor selective cannabinoid can be a candidate to target the trinity of infection, immunity, and inflammation in COVID-19. Front Pharmacol. 2021;12:590201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Fan Q, Longnecker R, Connolly SA. Herpes simplex virus glycoprotein B mutations define structural sites in domain I, the membrane proximal region, and the cytodomain that regulate entry. J Virol. 2021;95(22):e0105021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Cheshenko N, Herold BC. Glycoprotein B plays a predominant role in mediating herpes simplex virus type 2 attachment and is required for entry and cell-to-cell spread. J Gen Virol. 2002;83(Pt 9):2247–55. [DOI] [PubMed] [Google Scholar]
- 50.Bender FC, Whitbeck JC, Lou H, Cohen GH, Eisenberg RJ. Herpes simplex virus glycoprotein B binds to cell surfaces independently of heparan sulfate and blocks virus entry. J Virol. 2005;79(18):11588–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Cairns Tina M, Fontana J, Huang ZY, Whitbeck JC, Atanasiu D, Rao S, et al. Mechanism of neutralization of herpes simplex virus by antibodies directed at the fusion domain of glycoprotein B. J Virol. 2014;88(5):2677–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Komala Sari T, Gianopulos Katrina A, Nicola Anthony V. Glycoprotein C of herpes simplex virus 1 shields glycoprotein B from antibody neutralization. J Virol. 2020;94(5):10.1128/jvi.01852-19. [DOI] [PMC free article] [PubMed]
- 53.Navarro D, Paz P, Pereira L. Domains of herpes simplex virus I glycoprotein B that function in virus penetration, cell-to-cell spread, and cell fusion. Virology. 1992;186(1):99–112. [DOI] [PubMed] [Google Scholar]
- 54.Campadelli-Fiume G, Menotti L. Entry of alphaherpesviruses into the cell. In: Arvin A, Campadelli-Fiume G, Mocarski E, Moore PS, Roizman B, Whitley R, et al., editors. Human herpesviruses: biology, therapy, and immunoprophylaxis. Cambridge: Cambridge University Press Copyright © Cambridge University Press 2007;2007.
- 55.Nicola AV, McEvoy AM, Straus SE. Roles for endocytosis and low pH in herpes simplex virus entry into HeLa and Chinese hamster ovary cells. J Virol. 2003;77(9):5324–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Spear PG, Longnecker R. Herpesvirus entry: an update. J Virol. 2003;77(19):10179–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Tonge PJ. Drug-target kinetics in drug discovery. ACS Chem Neurosci. 2018;9(1):29–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data presented in this study are openly available in Mendeley Data at:
Bibliographic Details
DOI: 10.17632/k8jcgtt4mm.1
URL ID : https://data.mendeley.com/datasets/k8jcgtt4mm; 10.17632/k8jcgtt4mm.1; 10.17632/k8jcgtt4mm.1; https://data.mendeley.com/datasets/k8jcgtt4mm/1
AUTHOR(S)
Ahad Yamchi; Alijan Tabarraei; Seyyede Delafruz Hosseini; Mohammad Yasaghi; Abdolvahab Wahab Moradi; yasaghi, mohammad; Beryani Dolatabadi, Sahar; Yamchi, Ahad; Tabarraei, Alijan; Moradi, Abdolvahab ; Hosseini , Seyedeh Delafruz