

Abstract
Vibrio species, which include several pathogens, are autochthonous to estuarine and warm coastal marine environments, where biofilm formation bolsters their ecological persistence and transmission. Here, we identify a bicistronic operon, rcbAB, whose products synergistically inhibit motility and promote biofilm maturation post-attachment by modulating intracellular c-di-GMP levels in the human and animal pathogen V. vulnificus. RcbA contains an N-terminal tetratricopeptide repeat (TPR) domain and a structured C-terminal region of unknown function, while RcbB possesses an N-terminal TPR domain and a C-terminal GGDEF domain characteristic of diguanylate cyclases. The TPR domain of RcbB represses its diguanylate cyclase activity, while RcbA’s TPR domain and C-terminal region co-operatively de-repress it. Localization of both proteins to the flagellar pole is TPR-dependent but not co-dependent, although RcbA anchors RcbB to the pole in the absence of polar landmarks such as HubP and flagella. The conservation of rcbAB across diverse bacterial taxa substantiates its fundamental importance in bacterial biology. This work demonstrates how spatio-genetically coordinated TPR domain-containing proteins modulate c-di-GMP signaling, contributing to our understanding of biofilm formation in Vibrio species and potentially other bacteria. It also reveals the first evidence of inter-protein interaction via the TPR domains of both partners, challenging the conventional paradigm in which only one bears the domain.
Author summary
This study identifies the rcbAB operon as a critical regulator of biofilm development in the human and animal pathogen, V. vulnificus. The operon encodes tetratricopeptide repeat (TPR) domain-containing proteins that exhibit novel complex interactions and regulatory mechanisms. The conservation of this operon across diverse bacterial taxa supports its fundamental importance in bacterial biology. These findings expand our understanding of TPR domain versatility in mediating inter-protein interactions and provide new insights into complex formation and cellular signaling pathways in bacteria, which has potential implications for our comprehension of bacterial biofilm formation and may inform future strategies for controlling bacterial infections.
Introduction
Vibrios are Gram-negative, motile, halophilic, facultatively anaerobic bacteria that inhabit estuaries and warm coastal waters [1]. A dozen Vibrio species are pathogenic [2] and their environmental persistence and transmission are bolstered by their ability to form biofilms on biotic and abiotic marine surfaces [3,4]. Infection typically occurs through the consumption of contaminated seafood or exposure to contaminated water [5,6]. As seafood is a widely consumed and economically important food source, contamination by Vibrio species poses a serious public health risk. Among the pathogenic species, V. cholerae, V. parahaemolyticus and V. vulnificus are the primary contributors to the rising global prevalence of seafood-borne infections [7]. V. cholerae, the causative agent of cholera, causes a severe diarrheal illness that can result in fatal outcomes if left untreated [8]. V. parahaemolyticus is predominantly linked to acute gastroenteritis [7] and V. vulnificus causes life-threatening septicemia and wound infections [9,10]. It is an opportunistic pathogen that is responsible for over 95% of seafood-related deaths in the Unites States [11]. V. vulnificus carries the highest death rate and per case economic burden (>$3.3 M) of any foodborne disease agent, with an estimated annual cost of US $320 million [11–13]. It colonizes plankton, shrimp, fish, and eels [14–20] and forms biofilms in bivalves such as oysters that concentrate it from the surrounding waters when filter-feeding [17,21,22].
A biofilm is a complex community of microorganisms where normally free-swimming bacteria exist as part of a sessile, surface-associated community [23] that increases access to nutritional resources, provides protection from environmental insults and predation, and underscores environmental persistence [24], symbiosis [25], lateral gene transfer [26], and the establishment of chronic infections [27]. Biofilm formation is tightly controlled at multiple stages, from initial surface approach and attachment to irreversible adhesion and maturation, culminating in biofilm dispersal. Each stage involves precise molecular signaling and regulatory mechanisms that govern bacterial behavior, colonization, and community development within the biofilm structure [28].
The secondary signaling molecule bis-(3’-5’)-cyclic dimeric guanosine monophosphate (c-di-GMP) is a key regulator of biofilm formation in many bacteria [29]. It is synthesized by diguanylate cyclases (DGCs) bearing a GGDEF domain and degraded by phosphodiesterases (PDEs) containing EAL or HD-GYP domains [30,31]. Increases in c-di-GMP concentration generally repress flagellar-mediated swimming motility and virulence gene expression, while enhancing extracellular polysaccharide (EPS) production and biofilm formation [32–34]. DGC and PDE domains are often found in combination with other domains (e.g., CACHE, PAS, GAF, PBP, CSS, and GCS domains that sense amino acids, nucleotides, light, redox, and oxygen) that enable integration of multiple environmental signals and regulatory inputs to facilitate precise control of c-di-GMP levels in the cell [35,36]. A variety of effectors that bind c-di-GMP and elicit a physiological response have been identified [37]. These include proteins containing degenerate GGDEF and EAL domains [38], the PilZ domain [39], the GIL domain [40], motifs (e.g., I-site, Walker A and W[F/L/M][T/S]R [41–46]) and riboswitches [47–49]. These binding sites can be found alone [50,51] or coupled to output domains, the function of which is regulated by signal binding [52–55].
In V. vulnificus, environmental signals such as changes in calcium, sulfate and temperature can induce biofilm formation by increasing cellular c-di-GMP [56–60]. In response, the regulator BrpR activates expression of brpT, and together these regulators induce the production of BRP EPS [61–65] and calcium-binding matrix proteins that promote biofilm formation [57,58,63,66]. c-di-GMP is also bound by PlzD, a YcgR homolog [67,68] that localizes to the flagellar pole to slows swimming speed and turn frequency of motile cells [69].
Tetratricopeptide repeats (TPRs) represent a structural motif initially identified in yeast [70] that have since been found across a diverse array of proteins in all three domains of life [71], and represent an elegant protein-protein interaction module for the assembly and localization of multiprotein complexes within cells [72,73]. These motifs are characterized by a loosely conserved sequence of 34 amino acids, featuring alternating large and small side-chain residues [72,74]. Despite considerable sequence variability among TPRs, structural analyses reveal a highly conserved three-dimensional conformation [75,76]. Each TPR motif comprises two anti-parallel α-helices connected by a turn. Typically, TPR motifs are arranged in tandem repeats of three to sixteen units aligned at regular angles to form a right-handed superhelix, resulting in a binding groove that provides an extensive surface area for interaction with ligands and other proteins. TPR domain-containing proteins often play vital roles in eukaryotic cell processes, such as gene regulation, steroid receptor function, and protein import [73,77,78]. The importance of TPR-containing proteins is highlighted by the discovery that mutations in the TPR region are linked to several severe human diseases ranging from retinopathy to Down syndrome [79–81]. Strikingly, in most cases the target protein of eukaryotic TPR proteins is unknown. In bacteria, TPR domain-containing proteins have been reported to be directly involved in virulence-associated functions, including the translocation of virulence factors into host cells, adhesion to host cells, and blocking of phagolysosomal maturation [82–86].
Here, we screened a V. vulnificus transposon library for mutants deficient in calcium-induced biofilm formation and identified a two-gene operon, herein designated rcbA and rcbB (regulator of calcium-dependent biofilm development), that regulates biofilm formation and motility behavior. RcbA bears a putative N-terminal TPR domain and structured C-terminal region of unknown function, and RcbB harbors a N-terminal TPR domain and a C-terminal GGDEF domain found in DGCs. We show that RcbA and RcbB co-operatively increase c-di-GMP levels to enhance biofilm maturation post-attachment and slow motility. We demonstrate that the TPR domain of RcbB represses its DGC activity, while RcbA de-represses it. The TPR domains of RcbA and RcbB are crucial for their function, localizing both proteins to the flagellar pole and mediating their interaction. Polar localization of RcbA and RcbB is not co-dependent, but notably, RcbA anchors RcbB to the pole in the absence of polar landmarks such as HubP and flagella. To the best of our knowledge, this observation represents a novel instance of protein-protein interaction mediated by the tetratricopeptide repeat (TPR) domains of both protein partners, contrasting with the conventional paradigm in which only one interacting partner bears a TPR domain. The discovery of this mutual TPR-TPR interaction mechanism expands our knowledge of the versatility of TPR domains in facilitating protein-protein interactions and may have implications for our understanding of protein complex formation and cellular signaling pathways. Finally, the conservation and synteny of the rcbAB operon across diverse bacterial taxa implies that the functions of RcbA and RcbB are fundamentally important for the organisms in which they reside.
Results
A bicistronic operon mediates calcium-induced biofilm development in V. vulnificus
A V. vulnificus transposon (Tn) library was screened to identify mutants deficient in calcium-induced biofilm formation. Whole genome sequencing revealed multiple Tn insertions in an operon comprising two genes, aot11_00230 and aot11_00235, herein designated rcbA and rcbB (Fig 1A). In silico analysis predicted RcbA and RcbB to be cytoplasmic, with RcbA containing an N-terminal TPR domain and C-terminal structured region of unknown function, and RcbB possessing both an N-terminal TPR domain and a C-terminal DGC domain. To ascertain the impact of rcbAB on the production of biofilm matrix components, wildtype and ΔrcbAB strains harboring a PbrpAlacZ reporter fusion (PbrpA is a c-di-GMP responsive promoter that drives the expression of genes responsible for synthesizing BRP, a biofilm-promoting EPS) were cultured in LB media with or without calcium supplementation. Whilst PbrpA expression was induced >12-fold in wildtype cells by the addition of calcium, deletion of rcbAB suppressed this phenotype (Fig 1B). A similar expression pattern was observed for PcabA, a c-di-GMP responsive promoter that drives expression of the calcium-binding matrix protein, CabA (S1 Fig). Collectively, these findings suggested that rcbAB encodes regulators of BRP and CabA production, critical components of V. vulnificus biofilms.
Fig 1. The rcbAB operon regulates calcium-induced brp exopolysaccharide expression.

A, genetic organization of rcbAB. White arrows indicate Tn insertions identified in independent mutants and the bent green arrow represents a putative promoter. The domain structure of the encoded proteins is below. A tetratricopeptide (TPR) domain containing 7 repeats (blue) is present in both proteins, along with a diguanylate cyclase (DGC) domain (green) in RcbB. B, representative images of growth of wildtype V. vulnificus (WT) and ∆rcbAB mutant (Tn) strains bearing a PbrpAlacZ reporter on LB without or supplemented with 15 mM CaCl2 (LB + Ca2+). A plot of the corresponding PbrpAlacZ expression levels is below. Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences between samples (****p < 0.0001; ns, no significant difference) were determined by unpaired Student’s t-test (two-tailed).
To determine the stage at which rcbAB impacted biofilm formation, initial surface attachment and subsequent biofilm development of wildtype and ∆rcbAB strains were monitored under continuous flow conditions in an estuarine medium, IO20YP. After 1 hour of incubation, both wildtype and ∆rcbAB strains exhibited comparable numbers of single cells attached to the surface (Fig 2A). However, at 12- and 24-hours post-attachment, the biomass of the ∆rcbAB strain was significantly reduced, measuring 2-fold and 2.7-fold lower than that of the wildtype strain, respectively. Furthermore, the average and maximum biomass thickness of ∆rcbAB biofilms were 2.7-fold and 3.2-fold lower, respectively, than those observed in wildtype biofilms. Expression of either rcbA or rcbB alone in the ∆rcbAB background did not restore biofilm formation to wildtype levels; however, their co-expression resulted in a 5-fold increase in biofilm formation (Fig 2B), indicating a potential co-operative interaction between RcbA and RcbB in enhancing biofilm formation. These findings suggested that the deletion of rcbAB impedes biofilm development following initial surface attachment and that RcbA and RcbB function co-operatively to enhance biofilm formation during the subsequent stages of biofilm maturation.
Fig 2. RcbA and RcbB co-operatively promote biofilm development post-attachment.

A, representative fluorescence images of biofilms formed by wildtype (WT) and ∆rcbAB strains under constant flow in IO20YP after 1, 12 and 24 hours. The average and maximum biomass thickness is shown below and a plot of the biomass for the same strains is on the right. B, biofilm formation (OD595/OD600) by WT and ∆rcbAB strains expressing (+) rcbA, rcbB or both genes. Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences were determined by an unpaired Student’s t-test (two-tailed) in A and relative to WT cells by one-way ANOVA with a Tukey’s multiple comparisons post-hoc test in B (**p < 0.01, ***p < 0.001, ****p < 0.0001; ns, no significant difference). Expression was induced with 1 µM IPTG.
RcbA and RcbB co-operatively inhibit V. vulnificus swimming motility
Given the observed impact of rcbAB deletion and expression on V. vulnificus biofilm formation, the operon’s influence on swimming motility was investigated. Wildtype, ΔrcbA, ΔrcbB, and ΔrcbAB strains were inoculated into soft agar and motility zones were quantified after overnight incubation. Although an inverse correlation was observed between motility zone size and calcium concentration, there was no significant difference in motility between the strains at any condition tested (S2 Fig), suggesting that the effect of calcium on swimming motility was not rcbAB dependent. Swimming motility was also assessed for wildtype cells expressing rcbA, rcbB, or both genes simultaneously. Relative to wildtype cells carrying empty vector, cells expressing either rcbA or rcbB alone showed no significant change in motility zone size, while cells co-expressing rcbAB exhibited a nearly 2-fold decrease in motility zone diameter regardless of the calcium level (Figs 3A and S3), suggesting no direct calcium modulation of RcbAB activity.
Fig 3. RcbA and RcbB co-operatively inhibit V. vulnificus swimming motility.

A, plot of the swimming motility zones in soft agar by wildtype (WT) cells harboring the empty plasmid (pC2X6HT) or expressing (+) rcbA, rcbB or rcbAB, respectively. Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. B, representative images of motility traces for the same samples in A. Different colored lines in the traces on top represent trajectories for individual cells and the corresponding traces colored by swimming speed are below. C-E, violin plots of the swimming speed, confinement ratio (net displacement/total distance traveled), and turn angles for the cells in B. Dashed horizontal lines denote the median, first and third quartiles for the data range from 100 tracks for each strain. Statistically significant differences (*p < 0.05, **p < 0.01, ****p < 0.0001; ns, no significant difference) relative to WT cells were determined by one-way ANOVA with Dunnett’s multiple comparisons post hoc test. Expression was induced with 10 µM IPTG.
To further characterize the motility phenotype, the swimming speed of individual cells was analyzed (Fig 3B). Vibrio species swim using a distinct, three-step pattern that includes forward runs, reversals of ~180°, and flicks of ~90° (run-reverse-flicks) that re-orient the cell [87,88]. Consistent with the population-level assays, the trajectories for cells expressing either rcbA or rcbB alone showed no significant difference in swimming speed, confinement ratio (a measure of the efficiency of movement of a cell from starting position to endpoint), or turn angles relative to control cells (Fig 3C–3E). In contrast, cells co-expressing rcbA and rcbB exhibited a 50% decrease in average swimming speed (from 53 µm sec-1 to 26 µm sec-1) and a 30% increase in average confinement (from an of 0.45 to 0.32). Moreover, the turn angles, which primarily fell into clusters of ~93° (flicks) and 167° (reversals) for wildtype cells, were skewed toward reversals, with fewer flicks. Collectively, these results suggested a negative synergistic effect of rcbAB expression on V. vulnificus motility behavior that restricted foraging.
The TPR domain of RcbB represses its DGC activity and RcbA de-represses it
Since RcbB contained a predicted GGDEF domain, the cellular levels of c-di-GMP in V. vulnificus that expressed rcbA, rcbB or rcbAB was determined. Relative to wildtype, the level of cellular c-di-GMP was not altered by deletion of the operon, or in ∆rcbAB cells that expressed rcbA, rcbB, rcbBmut (a catalytic mutant bearing the A-to-E mutation, GGAEF) or rcbA∆tpr (which lacked the TPR domain) (Fig 4A). However, modest increases of 1.5-fold were observed for ∆rcbAB cells that expressed rcbB∆tpr or co-expressed rcbA and rcbB, and this effect was not calcium dependent (S4 Fig). Biofilm formation by ∆rcbAB cells expressing rcbA, rcbB, rcbBmut or rcbA∆tpr was similar to ∆rcbAB cells carrying the empty vector (Fig 4B). Additionally, rcbBmut was unable to promote biofilm formation or slow colony spread in the ∆rcbB background (S5 Fig), suggesting that the activity of RcbB was dependent on an intact catalytic motif. Notably, biofilm formation increased 2.5-fold for ∆rcbAB cells expressing rcbB∆tpr, although this was half that observed for cells expressing rcbAB. Likewise, the motility zones of cells expressing rcbA, rcbB, rcbBmut or rcbA∆tpr was similar to wildtype cells and, again, rcbB∆tpr expression was less effective at decreasing motility (1.5-fold) than rcbAB expression (1.9-fold) (Fig 4C). These results suggested that rcbB coded for a functional DGC and the co-expression of rcbA enhanced RcbB activity, resulting in a synergistic increase in c-di-GMP production and concomitant effects on c-di-GMP related phenotypes.
Fig 4. RcbA and RcbB co-operatively regulate c-di-GMP-dependent phenotypes.

Plot of cellular c-di-GMP levels (A), biofilm formation (B) and motility in soft agar (C) of wildtype V. vulnificus (WT) and ∆rcbAB cells that carry the empty expression plasmid or express (+) rcbA, rcbB, rcbBmut (GGAEF active site mutant), rcbAΔtpr (an RcbA TPR deletion mutant), rcbBΔtpr (an RcbB TPR deletion mutant) or the rcbAB operon. Bars show the respective mean values for each strain and error bars represent the standard deviation of triplicate assays. Statistically significant differences relative to wildtype were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test (**p < 0.01, ***p < 0.001, ****p < 0.0001; ns, no significant difference). Expression was induced with 10 µM IPTG in A, 1 µM in B and 10 µM in C.
To mitigate possible complications from the plethora of V. vulnificus gene products predicted to participate in c-di-GMP turnover, cellular c-di-GMP levels were also measured in E. coli MG1655 cells, the motility of which, as observed in V. vulnificus, decreased in response to rcbAB expression (S6 Fig). Relative to control cells, the cellular level of c-di-GMP did not change in cells expressing rcbA alone (Fig 5). Although only a modest increase of 1.5-fold in the intracellular c-di-GMP level was observed for cells expressing rcbB, no increase was observed in cells expressing rcbBmut. Notably, the cellular c-di-GMP level of cells co-expressing rcbA and rcbB increased nearly 4-fold, analogous to the increase observed in V. vulnificus. Cells expressing rcbAΔtpr had c-di-GMP levels similar to control cells. In contrast, rcbBΔtpr expression resulted in an approximately 5-fold increase in c-di-GMP levels, suggesting that the TPR domain of RcbB strongly suppressed its DGC activity. Although rcbAΔtpr-rcbB expression increased c-di-GMP levels 3-fold relative to control cells, this was less than observed for cells expressing rcbAB, and an even greater drop was observed for cells expressing an operon in which rcbA lacked its C-terminal region (rcbAΔC-rcbB). Collectively, these results suggested that the TPR domain of RcbB inhibited its diguanylate cyclase activity and that RcbA relieved this inhibition using both its N-terminal TPR domain and C-terminal region, although we acknowledge that some of the differences in activity of the truncated versions of RcbA and RcbB could be due to changes in stability. Moreover, while removing TPR-mediated inhibition maximized c-di-GMP output by RcbB, this alone did not correlate with maximal phenotypic effect on biofilm formation and motility, and implied that the mechanism by which RcbA modulated RcbB activity extended beyond simple de-repression to, possibly, subcellular localization.
Fig 5. RcbA and RcbB co-operatively regulate c-di-GMP production and the TPR domain of RcbB suppresses its DGC activity.

Plot of intracellular c-di-GMP levels in wildtype E. coli MG1655 cells that carry the empty expression plasmid (control) or express (+) rcbA, rcbB, rcbBmut (GGAEF active site mutant), rcbAB, rcbAΔtpr (an RcbA TPR deletion mutant), rcbBΔtpr (an RcbB TPR deletion mutant), rcbAΔC-rcbB (in which the C-terminus of RcbA is deleted), or rcbAΔtpr-rcbB. (in which the TPR domain of RcbA is deleted). Bars show the respective mean values for each strain and error bars represent the standard deviation of triplicate assays. Statistically significant differences relative to the respective controls were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test (**p < 0.01, ****p < 0.0001; ns, no significant difference). Expression was induced with 10 µM IPTG.
RcbA and RcbB interact via their TPR domains
The impact of RcbA on the activity of RcbB suggested a possible direct interaction between the proteins. AlphaFold 3 was used to predict the joint structure of a RcbA-RcbB complex (Fig 6A). The confidence of the overall predicted fold for the complex (Fig 6C) was high (pTM = 0.56), particularly for the 7 N-terminal TPR motifs of each protein (Fig 6E). A potential interaction surface between the final motif (TPR 7) of both proteins was predicted, as well as self-interaction by RcbA (Fig 6B, 6D and 6F).
Fig 6. RcbA and RcbB interact via their TPR domains.

AlphaFold 3 predicted structure of RcbA and RcbB complexes. Shown are a RcbA-RcbB heterodimer (A, C and E) and RcbA homodimer (B, D and F) colored by chain (A and B), per-atom confidence estimate (pLDDT; C and D) and rainbow for the TPR repeats (E and F). The N-terminal TPR domains of RcbA and RcbB are predicted to contain 7 TPR repeats each (numbered 1-7 and colored red, orange, yellow, green, cyan, light blue and purple, respectively). The C-terminus of RcbA and GGDEF domain of RcbB are colored grey. G, western blot of pull-downs using Ni-coated beads from extracts of E. coli producing either the HIS-tagged TPR domain of RcbA (RcbATPR) or the HA-tagged TPR domain of RcbB (RcbBTPR) alone or together. Anti-HIS antibody (HIS) was used to detect RcbATPR. The blot was stripped for subsequent detection of RcbBTPR with anti-HA antibody (HA). Expression was induced with 50 µM IPTG. H, Bacterial Adenylate Cyclase Two-Hybrid (BACTH) assays of protein-protein interactions. RcbA, RcbB, RcbATPR and RcbBTPR were fused to the C-terminus of Bordetella pertussis adenylate cyclase subunits T18 or T25. The plot shows β-galactosidase activity (Miller units) for the indicated protein combinations. Leucine zipper fusions to T18 and T25 served as a positive control (+), while empty plasmids were used as the negative control (-). Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences (****p < 0.0001; ns, no significant difference) relative to the negative control were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test. Representative images of the respective strains are shown above the plot.
Attempts to determine if RcbA and RcbB interacted in vivo using V. vulnificus cell extracts that contained HIS-tagged RcbA (RcbA), HA-tagged RcbB (RcbB) or both were unsuccessful, as the proteins produced (S7A Fig) were rapidly degraded following cell lysis; similar issues were encountered using extracts containing only the TPR domains of the proteins. Instead, E. coli cell extracts containing the HIS-tagged TPR domain of RcbA (RcbATPR), the HA-tagged TPR domain of RcbB (RcbBTPR) or both were incubated with nickel-coated magnetic beads, and recovered proteins were analyzed by immunoblot. A band was observed with anti-HIS antibody for the two extracts that contained RcbATPR, while no signal was detected for cells producing only RcbBTPR (Fig 6G). The blot was then stripped and re-probed with anti-HA antibody. Notably, no signal was observed for extracts containing only RcbATPR or RcbBTPR, whereas a signal was observed for cells producing both proteins. Collectively, these results suggested that RcbA and RcbB interacted via their TPR domains.
Interaction between RcbA and RcbB was also assessed by Bacterial Adenylate Cyclase Two-Hybrid (BACTH) analysis. RcbA and RcbATPR were fused to the C-terminus of Bordetella pertussis adenylate cyclase subunit T18, and RcbB and RcbBTPR were fused to adenylate cyclase subunit T25. Co-production of T18-RcbA and T25-RcbB resulted in blue colonies and β-galactosidase activity equal to the positive control, suggesting strong interaction between the two proteins (Fig 6H). Co-production of T18-RcbATPR and T25-RcbBTPR also produced blue colonies and high β-galactosidase activity, suggesting that the interaction is mediated via their TPR domains. RcbA self-interaction was also observed, suggesting that it can form dimers or possibly higher order oligomers. However, interaction between T18-RcbATPR and T25-RcbATPR was not detected, arguing against self-interaction being mediated by the TPR domain of RcbA. White colonies and minimal β-galactosidase activity for cells producing T18-RcbB and T25-RcbB or T18-RcbBTPR and T25-RcbBTPR suggested that RcbB does not self-interact. Collectively, these results support the notion that RcbA and RcbB interact with each other via their TPR domains, while RcbA exhibits TPR-independent self-interaction.
RcbA and RcbB localize to the flagellar pole
The previous data raised the possibility of subcellular localization of RcbA and RcbB in the cell. The positions of the proteins were monitored in ∆rcbAB cells using fluorescently tagged fusions (RcbA-mRuby3 and RcbB-mNeonGreen) that were functional and promoted biofilm formation at levels comparable to the wildtype proteins (S8 Fig). Both proteins localized to the same cell pole (Fig 7). Since co-expression of rcbA and rcbB co-operatively slowed swimming motility, we determined if RcbA and RcbB localized to the flagellar pole. FliM that was C-terminally tagged with blue fluorescent protein (BFP) was used as a marker of the flagellar pole. Co-expression of rcbA-mRuby3 and fliM-bfp in a ΔfliMΔrcbA background revealed polar fluorescent foci for the corresponding proteins at the same pole. Since RcbB localizes to the same pole as RcbA, collectively these data suggested that RcbA and RcbB localized to the flagellar pole.
Fig 7. RcbA and RcbB localize to the flagellar pole.

Representative phase, fluorescence and overlay images of ∆rcbAB cells expressing rcbA-mRuby3 and rcbB-mNeonGreen (top panels) and ∆fliM∆rcbA cells expressing rcbA-mRuby3 and fliM-bfp (bottom panels). Scale bars = 1 μm. Expression of rcbA and rcbB was induced with 10 µM IPTG and fliM was induced with 0.05% L-arabinose.
The TPR domain mediates localization of RcbA and RcbB to the cell pole
To elucidate the role of the TPR domain in the polar localization of RcbA and RcbB, fluorescently tagged variants lacking the TPR domain (RcbA∆TPRmRuby3 and RcbB∆TPRmNeonGreen), which were predicted to correctly fold and were produced in V. vulnificus (S7B and S7C Fig), were expressed in ΔrcbA and ΔrcbB cells, respectively, and their subcellular distribution was monitored. In contrast to the wildtype proteins, these truncated variants produced diffuse cytoplasmic signals (Fig 8A and 8B), suggesting that the TPR domain was needed for polar localization of both RcbA and RcbB. The AlphaFold 3-predicted structure suggested that TPR-7 mediated the interaction between RcbA and RcbB (Fig 6E). To test this hypothesis, the localization of RcbA and RcbB variants lacking TPR1-5 (RcbATPR6/7-mRuby3 and RcbBTPR6/7-mNeonGreen) was determined. Fluorescence microscopy revealed that the truncated variants continued to localize to the cell pole (Fig 8C and 8D). However, further truncation to remove TPR6 (RcbATPR7-mRuby3 and RcbBTPR7-mNeonGreen) resulted in diffuse cytoplasmic signals for both proteins (Fig 8E and 8F). These observations suggested that both TPR6 and TPR7 were needed for polar localization of RcbA and RcbB.
Fig 8. TPR6 and TPR7 of RcbA and RcbB are necessary for polar localization.
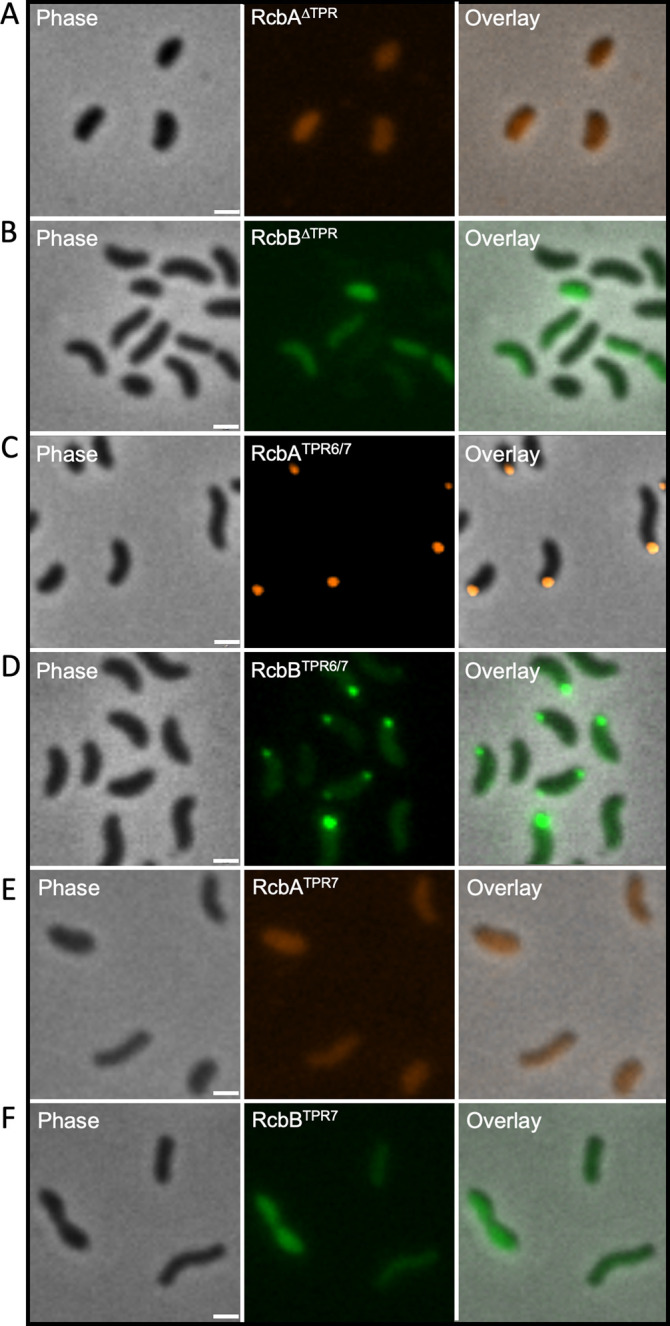
From left to right, representative phase, fluorescence and overlay images of RcbA∆TPR-mRuby3 signal in ∆rcbA cells (A), RcbB∆TPR-mNeonGreen signal in ∆rcbB cells (B), RcbATPR6/7-mRuby3 signal in the ∆rcbA cells (C), RcbBTPR6/7-mNeonGreen signal in ∆rcbB cells (D), RcbATPR7-mRuby3 signal in the ∆rcbA cells (E) and RcbBTPR7-mNeonGreen signal in ∆rcbB cells (F). Scale bar = 1 μm. Expression was induced with 10 µM IPTG.
Polar localization of RcbB and RcbA is not co-dependent but RcbA anchors RcbB in the absence of polar landmarks
To elucidate the interdependence of RcbA and RcbB for polar localization, the subcellular distribution of RcbA-mRuby3 and RcbB-mNeonGreen was separately monitored in ΔrcbAB cells. The signal for RcbA-mRuby3 remained polar in the absence of RcbB and the signal for RcbB-mNeonGreen was polar in the absence of RcbA (Fig 9). This suggested that RcbA and RcbB could independently localize to the cell pole without prior interaction. To investigate the role of polar landmarks in the localization of these proteins, the fluorescence patterns of RcbA-mRuby3 and RcbB-mNeonGreen were monitored in a ΔhubPΔrcbAB background. HubP plays a crucial role in organizing and coordinating various cellular processes at the bacterial cell pole [89–92]. In this genetic context, RcbA-mRuby3 maintained its polar localization, whereas RcbB-mNeonGreen displayed a diffuse cytoplasmic distribution (Fig 10A and 10B). These results indicated that the polar localization of RcbB, but not RcbA, was dependent on HubP. Cells lacking hubP retained the ability to produce polar flagella (S9 Fig). To determine if RcbA and RcbB localization was dependent on polar flagellar synthesis, their subcellular position was monitored in a ∆flrA∆rcbAB background. FlrA is a master regulator of flagellar gene expression in Vibrio species [93–95] and its deletion results in cells that lack flagella (S9 Fig). The same localization pattern for RcbA-mRuby3 and RcbB-mNeonGreen that was observed in ΔhubPΔrcbAB cells was observed in ∆flrA∆rcbAB cells (S10A and S10B Fig). Notably, co-expression of RcbA mitigated the effects of HubP or FlrA deletion on polar localization of RcbB in the ∆hubP∆rcbAB and ∆flrA∆rcbAB backgrounds (Figs 10C and S10C). However, an RcbA variant lacking its TPR domain failed to anchor RcbB at the cell pole (Fig 10D). Since RcbA containing only TPR6/7 successfully tethered RcbB also bearing TPR6/7 to the pole in ∆hubP∆rcbAB cells (Fig 10E), this suggested that these repeats were needed for RcbA-mediated polar anchoring of RcbB. Thus, polar localization of RcbA and RcbB was not co-dependent, but RcbA anchored RcbB to the pole in the absence of the polar landmark protein HubP and flagella.
Fig 9. Polar localization of RcbA and RcbB is not co-dependent.

Representative phase, fluorescence and overlay images of ∆rcbAB cells expressing rcbA-mRuby3 (top panels) or rcbB-mNeonGreen (bottom panels). Scale bars = 1 μm. Expression was induced with 10 µM IPTG.
Fig 10. RcbA anchors RcbB to the pole in the absence of the polar landmark protein HubP.

From left to right, representative phase, fluorescence and overlay images of RcbA-mRuby3 (A), RcbB-mNeonGreen (B), both (C), RcbA∆TPR-mRuby3 and RcbB-mNeonGreen (D) or RcbATPR6/7-mRuby3 and RcbBTPR6/7-mNeonGreen signal (E) in ∆hubP∆rcbAB cells. Scale bar = 1 μm. Expression was induced with 10 µM IPTG.
Phylogenetic distribution of rcbAB
Among the γ-proteobacteria, homologs of the rcbAB operon were identified in 73 out of 91 distinct Vibrio species represented in publicly accessible complete genome assemblies, encompassing a total of 583 out of 617 genomes (S2 Table). Since only one or two genome assemblies were available for some species, sequencing of additional isolates may yet reveal extant strains with rcbAB homologs for these species (e.g., the genomes of two V. alginolyticus isolates lack rcbAB, whereas 43 code for it). In most instances (96%), the operon was situated on the smaller of the two chromosomes within each species, and in all cases, rcbA was found upstream of rcbB. The operon was also detected in other genera within the Vibrionaceae, specifically Allivibrio and Thaumasiovibrio.
The homology of RcbA orthologs varied from 100% in V. vulnificus to a mere 28.5% in V. cincinnatiensis, while the homology of RcbB orthologs ranged from 100% to 43.6% (Fig 11). Each species possessed a single copy of the operon, apart from V. cincinnatiensis, which harbored two copies located more than 230 kb apart on the same chromosome. While the products of one of the operons shared approximately 60% overall identity to those of V. vulnificus, the other, despite exhibiting only about 36% overall identity, retained TPR and DGC domain features, suggesting a common progenitor that evolved by gene duplication and potential sub-functionalization of the paralogs in this species.
Fig 11. Phylogenetic distribution of rcbAB among Vibrio species.

From inside-out, Maximum Likelihood tree (rooted with V. vulnificus) based on 133 common core genes from 617 complete Vibrio genome assemblies, with branch lengths representing substitutions per site. Clades are color-coded for the ten most frequently represented Vibrio species, with the number of genomes harboring rcbAB indicated in parentheses. Taxa names are labeled on the tree. The inner two color-gradient tracks show percent identity to rcbA and rcbB from V. vunificus ATCC27562 of orthologs from the respective strain, while the outer two tracks indicate the number of tetratricopeptide repeats (TPR count) identified in the corresponding proteins.
Notably, despite significant variability in sequence, RcbA homologs consistently displayed a configuration comprising 7 tetratricopeptide motifs (TPRs). Only three species (V. europaeus, V. palustris, and V. tubiashii) harbored homologs with six TPRs. The majority of RcbB homologs also exhibited a configuration of seven TPRs; however, greater variability was observed, with at least ten species possessing homologs with six TPRs (V. atlanticus, V. bathopelagicus, V. campbellii, V. cortegadensis, V. gallaecicus, V. owensii, V. pectenicida, V. pelagius, and V. syngnathi), alongside two species that contained homologs with only five TPRs (V. tritonius and V. poteresiae).
A search among non-Vibrionaceae genomes yielded 257 chromosomes encoding 341 rcbAB homologs in the families Alteromonadaceae (genera Alteromonas and Motilimonas) and Psychromonadaceae (genus Psychromonas), 53 of which had more than one copy and 55 of which had rcbA positioned downstream of rcbB. Among β-proteobacteria, homologs of the rcbAB operon were found within the Chitinibacteraceae, represented by genera such as Andreprevotia, Chitinibacter, Chitinilyticum, Chitiniphilus, Deefgea, Formivibrio, and Jeongeupia (S3 Table). Most homologs shared 10–40% identity with rcbA and rcbB of V. vulnificus and gene order was preserved across most genera; however, rcbB was positioned upstream of rcbA in Jeongeupia and some Chitinibacter species (e.g., C. bivalviorum strain 2T18 had two rcbAB and four rcbBA homologs). The conservation of the operon across diverse bacterial taxa implies that the functions of RcbA and RcbB are fundamentally important for the organisms in which they are found.
Discussion
Biofilm formation is an ancient and universal survival mechanism [23,96]. c-di-GMP is a key regulator of biofilm formation [33,97], increases in which generally enhance EPS production and biofilm formation while repressing flagellar-mediated swimming motility and virulence gene expression. Here, we identify rcbAB, a two-gene operon coding for proteins that partner to regulate c-di-GMP-dependent phenotypes in V. vulnificus and provide insight into the functional dynamics of RcbA and RcbB in the cell. The reduction in post-attachment biofilm biomass of a ΔrcbAB strain supports a significant role for rcbAB in biofilm maturation. Localization of both proteins to the flagellar pole suggested a role for RcbA and RcbB in motility regulation, and co-expression of rcbAB suppressed motility; however, its deletion had no impact on the phenotype. This suggests that the operon primarily influences biofilm formation, while perhaps playing an ancillary role in regulating swimming motility. RcbA contains an N-terminal TPR domain and a structured C-terminal region of unknown function, while RcbB possesses an N-terminal TPR domain and a C-terminal diguanylate cyclase domain. A recent study in V. parahaemolyticus described the function of two separately coded DGCs with predicted periplasmic TPR domains that additively repress the expression of genes required for swarming motility [98]. Both proteins required an intact catalytic diguanylate cyclase motif to function, however, it was not determined if the TPR domains of the DGCs affected their subcellular location or activity, nor were their interacting targets, if any, identified. Removing the TPR domain of RcbB maximized its c-di-GMP output, but this alone did not correlate with maximal phenotypic effect. RcbA and RcbB interact via their N-terminal TPR domains, with RcbA both de-repressing the diguanylate cyclase activity of RcbB and, notably, localizing it to the flagellar pole. The observation that RcbA anchors RcbB at the pole in the absence of polar landmarks such as HubP and flagella suggests a hierarchical regulatory mechanism where RcbA may serve as a scaffold for RcbB. It also suggests that the spatial organization of the RcbA-RcbB complex in the cell is important for function. It is conceivable that polar localization of RcbB’s diguanylate activity generates a high-c-di-GMP microenvironment in the vicinity of the flagellum that can be sensed by nearby proteins so that bacteria can fine tune physiological responses based on environmental cues. The proposed model (Fig 12) illustrates how the interaction and subcellular localization of RcbA and RcbB might regulate biofilm formation and swimming motility by modulating local c-di-GMP levels. The N-terminal TPR domain of RcbB strongly inhibits its C-terminal DGC activity, keeping it in an autoinhibited state. Since DGCs typically function as protein dimers or higher order oligomers [99–101], RcbA, which can self-assemble into homomultimers, facilitates the formation of heterotetramers or higher-order oligomers involving RcbB that are important for de-repression of its diguanylate cyclase activity (S11 Fig). The N-terminal TPR domain of RcbA anchors RcbB to the flagellar pole and, co-operatively with its C-terminal region, relieves RcbB’s autoinhibition to maximize the phenotypic impact of elevated, localized c-di-GMP concentrations. Elevated c-di-GMP levels increase expression of brpR, which encodes a transcriptional activator and potential c-di-GMP effector [64,65,102]. BrpR in turn upregulates brpT expression and together these regulators induce the synthesis of biofilm matrix components such as BRP EPS and calcium-binding proteins to promote biofilm development. Additionally, c-di-GMP produced by RcbB may be sensed by effectors such as PlzD, the positioning of which at the flagellar pole in slow moving, initially adherent and mature biofilm cells, is regulated by c-di-GMP [69]. This outcome alters the foraging behavior of cells by slowing swimming speed and decreasing the number of directional changes, ultimately limiting exploration of the surrounding 3D space. Collectively, these events facilitate the transition from a planktonic to a sessile bacterial lifestyle.
Fig 12. Model for subcellular localization and interaction of RcbA and RcbB in regulating bacterial biofilm formation and motility.

Expression of the bicistronic operon produces RcbA (purple) and RcbB (green), which can independently localize to the flagellar pole. RcbA can self-assemble into homomultimers (purple/red), while RcbB cannot; however, RcbA may facilitate the formation of higher-order oligomers involving RcbB (green/gold) that are functionally important, since DGCs typically function as protein dimers. The N-terminal TPR domain of RcbB strongly inhibits its C-terminal DGC activity, keeping it in an autoinhibited state. RcbA anchors RcbB to the flagellar pole for optimal phenotypic effect and relieves RcbB’s autoinhibition. Elevated c-di-GMP production increases expression of brpR (brown), which encodes a transcriptional activator and potential c-di-GMP effector. BrpR in turn upregulates expression of the regulator encoded by brpT (blue). This cascade induces the synthesis of biofilm matrix components such as BRP exopolysaccharides (EPS) and calcium-binding proteins (dark and light grey, respectively), promoting biofilm development. Additionally, c-di-GMP may be sensed by effector proteins such as PlzD, which localizes to the flagellar pole and reduces the swimming speed and turn frequency of motile cells.
Spatial regulation by targeting diguanylate cyclase activity to the cell pole of bacteria has profound implications for bacterial physiology, as heterogeneity in cellular c-di-GMP levels can lead to diverse phenotypic outcomes within a clonal population, enabling bacteria to better adapt to varying environmental conditions. DGCs, such as YfiN in Escherichia coli, have been shown to relocate to the division site under stress conditions, where they inhibit cell division by preventing septal peptidoglycan synthesis [103]. The DGC PleD exhibits a cell cycle-dependent localization pattern in Caulobacter crescentus. In swarmer cells, PleD remains in an inactive state but undergoes activation during the swarmer-to-stalked cell transition [101]. Its activity is intricately linked to its spatial redistribution to the stalked pole of the cell, suggesting that PleD activates proximal downstream effectors involved in pole remodeling [104]. In Pseudomonas aeruginosa, the DGC DgcP interacts with FimV, a polar peptidoglycan-binding protein, influencing local c-di-GMP production at the cell pole and type IV pilus assembly [105].
Polar localization of RcbA appears to be an intrinsic characteristic, while the localization of RcbB in the absence of RcbA is contingent upon the presence of HubP or polar flagella. These localization dynamics bear resemblance to the interactions of FlhF and FlhG, two proteins that regulate flagellar assembly and positioning in Vibrio species. Specifically, the polar localization of FlhF facilitates flagellar formation and establishes its spatial orientation, whereas FlhG modulates the quantity of flagella produced by interacting with FlhF to inhibit its polar localization, thereby regulating flagellar number [106]. The coordinated activities of these proteins are crucial for the precise determination of flagellar quantity and placement. Historically, it was posited that polar localization of FlhF was intrinsic [107,108], while that of FlhG was dependent on HubP [91,109]. However, recent findings have demonstrated that FlhF can be recruited to the cell pole by the small integral membrane protein, FipA [89]. At this juncture, we cannot exclude the possibility that a similar mechanism may govern polar localization of RcbA and future investigations will aim to ascertain whether RcbA possesses the capability of independent polar localization.
The TPR motif has been found in organisms ranging from bacteria to humans [74]. TPR motif-containing proteins act as scaffolds for the assembly of different multiprotein complexes and play important roles in various cellular processes and pathogenesis [72,110]. Most studies describe TPR domains binding to peptide motifs or other protein regions [73,111], and although multiple TPR-containing proteins may indirectly interact within the same complex, no direct evidence exists of two proteins interacting specifically via their TPR domains (i.e., a TPR-TPR interaction). RcbA and RcbB, and most of their orthologs, were found to harbor 7 repeats in their TPR domains and interacted via these domains to form a heterocomplex. To our knowledge, this is the first evidence supporting an interaction between the TPR domains of two proteins, challenging the conventional paradigm in which only one protein partner bears the domain. Moreover, TPR 6 and 7 were necessary for interaction between RcbA and RcbB, and for their localization to the cell pole. It is tempting to speculate that TPR1-5 of RcbA and RcbB interact with other proteins as part of a larger assemblage at the cell pole. Within the periplasm of E. coli, the TPR domain of the outer membrane porin PgaA interacts with the deacetylation domain of PgaB to form a complex at the outer membrane that increases the deacetylation and glycoside hydrolase activities of PgaB as poly-β(1,6)-N-acetylglucosamine is transported to the cell surface [112]. Additional studies demonstrated that only the TPRs of PgaA that were proximal to its porin moiety interacted with PgaB, leading the authors to propose that its distal TPR motifs spanned the periplasm and interacted with inner membrane components of the secretion apparatus [113]. Additionally, the C-terminal region of RcbA, which bears no identifiable domains, is predicted to adopt extensive secondary structure. Participation of both domains of RcbA in modulating the diguanylate cyclase activity of RcbB hints at a sophisticated regulatory mechanism and nuanced level of control that may be pivotal for RcbA-RcbB function. Further investigation is needed to elucidate the precise nature of the molecular interactions between RcbA and RcbB, as well as their potential association with other proteins at the cell pole.
The rcbAB operon is widely distributed among pathogenic Vibrio species [4,8], symbionts [114–118], other genera within the Vibrionaceae [5], and beyond. This broad distribution suggests an evolutionary advantage conferred by this operon, likely due to its role in biofilm formation and motility - critical factors for survival in diverse aquatic environments. In support of this notion, GefB, a DGC that was recently reported to regulate biofilm formation, swarming and swimming motility in V. parahaemolyticus [119], is a homolog of RcbB and is transcribed from a bicistronic operon that also codes for a homolog of RcbA. It is likely that the functions of RcbA and RcbB have been maintained across many of these species, however, the variability observed among more distant homologs raises the possibility of functional divergence and adaptation among some, warranting further investigation into how these variations impact the ecological fitness of the species in which they are found.
The V. vulnificus genome encodes nearly 100 proteins predicted to make, break, and bind c-di-GMP [120,121], but relatively little is known regarding the environmental signals that regulate c-di-GMP levels and biofilm formation in response to changing environmental conditions. Calcium is a key environmental signal inducing c-di-GMP accumulation and upregulating exopolysaccharide, pilus, and matrix protein expression in V. vulnificus [56–60]. However, important aspects of calcium-dependent mechanisms that govern biofilm formation and motility in V. vulnificus remain unresolved and warrant investigation to address Vibrio biofilm challenges in clinical and environmental contexts. Future studies should focus on identifying environmental factors (e.g., cell density, temperature, nutrient availability) and signaling pathways that modulate RcbAB function and c-di-GMP dynamics. Apropos, the homologous V. parahaemolyticus vpa1477-gefB operon is reportedly regulated by the quorum-sensing regulators AphA and OpaR [119].
Materials and methods
Media and strains
LB, LB agar, Peptone and Yeast Extract were purchased from BD Difco and Instant Ocean was purchased from Instant Ocean. Antibiotics and additives were purchased from Sigma and used at the following concentrations: rifampin (Rf), 100 µg ml-1; kanamycin (Km), 160 µg ml-1; chloramphenicol (Cm), 25 µg ml-1 for E. coli and 2 µg ml-1 for V. vulnificus; ampicillin (Ap), 100 µg ml-1; gentamicin (Gm), 10 µg/ml for E. coli and 35 µg ml-1 for V. vulnificus; trimethoprim (Tp), 10 µg ml-1; 5-bromo-4-chloro-3-indolyl-β-D-galacto-pyranoside (X-Gal), 100 µg ml-1; L-arabinose (ara), 0.05%; and CaCl2 (15 mM); isopropyl-β-D-thiogalactopyranoside (IPTG), see figure legends for concentrations used in each experiment. Instant Ocean was used at 20 parts per thousand (IO20) and was supplemented with 0.5% yeast extract and 1% peptone (IO20YP) for growth of V. vulnificus. A spontaneous Rf resistant isolate of V. vulnificus strain ATCC27562 was used as the parental strain and E. coli S17.1λπ was used for plasmid transfer to V. vulnificus by conjugation.
Construction of the PbrpAlacZ and PcabAlacZ reporter plasmids and screening of a transposon library
We previously constructed a 325 bp PbrpA PCR product that was fused to the E. coli lacZ (lacZEc) and inserted into pTX1CebgA by Gibson assembly [56]. A similar construct was generated using a 271 bp PcabA PCR product. Plasmids were maintained in E. coli S17.1λπ and conjugated to V. vulnificus for integration into ebgA (a cryptic lacZ). Transconjugants were selected on LB Rf Cm plates. To monitor β-galactosidase activity in V. vulnificus without interference due to expression of its endogenous lacZ (lacZVv), the reporter plasmids were conjugated to a V. vulnificus strain bearing a markerless deletion of the promoter for lacZVv that was created using the pRE112 sucrose counterselection plasmid [122].
Tn mutagenesis
The mini-Tn10 delivery vector pNKTXI-SceI [123] was used for transposon mutagenesis as previously described [56]. Briefly, E. coli strains carrying the plasmid were used as donors in mating experiments to transfer the plasmid to V. vulnificus strains bearing the PbrpAlacZ reporter via conjugation. After overnight growth, transconjugants were selected by streaking onto LB plates with Rf, X-Gal, Cm, Km and CaCl2. After 48 hours of growth, white colonies representing loss of brp expression in the presence of CaCl2 (a total of 30,000 mutants were screened) were re-screened on the same type of selective plates. Verified mutants were pooled, collective genomic DNA was prepared and sent for whole-genome sequencing (Eurofins). Geneious (Biomatters) was used to map the reads to the mini-Tn10 reference sequence and recover flanking V. vulnificus genomic sequences that were mapped back against the whole genome of strain ATCC 27562 [121].
Mutant construction and complementation
Allelic replacement of target genes was achieved as follows: PCR fragments corresponding to 1 kb upstream and downstream of the target gene and bearing overlaps with a central Tp, Km or Cm antibiotic resistance cassette were amplified and ligated to the cassette by Gibson reaction. Recipient strains carrying the pMMB-TfoX expression plasmid [124] were induced overnight in LB containing Ap and IPTG at 30°C. A 10 µl aliquot of cells was added to 500 µl of IO20 containing IPTG and 25 µl of the Gibson reaction. The transformation mixture was incubated statically overnight at 30°C. The next day, 1 ml of LB was added to the tube and cells were allowed to outgrow for 3 hours before plating on selective LB containing the appropriate antibiotic. Allelic replacement of the target gene was confirmed by PCR. For complementation, rcbA, rcbB and variants were amplified alone or as an operon and cloned into pC2X6HT [62] by Gibson assembly and expression was induced with IPTG; fliM was cloned into pSU38GT [61] and expression was induced with L-arabinose. Plasmids were conjugated from E. coli S17.1λπ to V. vulnificus and transconjugants were selected on LB Rf Ap plates.
Colony morphology assays and crystal violet staining of biofilms in 96-well plates
For colony morphology assays, strains were grown overnight with agitation in LB containing the appropriate antibiotics at 30°C. Then 3 μl of each bacterial culture was spotted on LB agar plates containing antibiotics. Plates included X-Gal for visualization of PbrpAlacZ expression. Images were captured after 48 hours using a Leica MS5 dissecting scope equipped with a Leica DC300F camera.
For crystal violet staining, strains were grown in IO20YP that contained the appropriate antibiotics at 30°C with shaking until they reached an OD600 of 1. Cultures were adjusted to 1x106 CFU ml-1 in fresh media, and 150 µl was inoculated in triplicate into 96-well microtiter plates. Microtiter plates were incubated statically for 20 hours at 30°C. Following measurement of the planktonic cell density at OD600 on a BioTek Synergy HT microplate spectrophotometer, 50 µl of 0.4% crystal violet (CV) solution was added to each well. After 30 min, the stained wells were washed three times with distilled water and the remaining dye was resolubilized in 200 µl of 30% acetic acid. The OD595 of each well was then recorded and the ratio of OD595/OD600 was calculated to determine the relative level of biofilm formation. All assays were done in triplicate.
Biofilm development in microfluidic chambers
Polydimethylsiloxane (PDMS)-glass flow cell devices containing eight 40 x 5 x 1-mm chambers were fabricated and sterilized [125]. Mid-log-phase wildtype cells (OD600 of 0.1) grown in IO20YP were seeded into separate flow cell chambers. Control chambers for background fluorescence contained medium only. Initial attachment (no flow) proceeded for 20 min, followed by a flow rate of 0.25 ml min-1 in IO20 for 16 hours. The chambers were then flooded with the same medium containing 2.5 µM Syto21 (Molecular Probes) for 10 min. Biofilm images and z-stacks (20 x 1 µm slices) were captured with an Olympus IX83 microscope using a UPLSAPO 40x silicon oil immersion objective (NA, 1.25; WD, 0.3 mm). Background fluorescence was subtracted from each sample, and quantitative analysis to determine biomass was performed using cellSense (Olympus) and Comstat [126,127]. Data from three biological replicates were analyzed for each strain. The images presented are from a single representative experiment.
Soft-agar swimming assays
V. vulnificus cells were grown overnight in IO20 that contained the appropriate antibiotics and inducers at 30°C with agitation. Bacterial culture (2 μl) was inoculated into semisolid IO20 agar (0.3%) plates supplemented with 1% Tryptone and included the appropriate antibiotics and inducers. Plates were incubated overnight at room temperature and the swimming diameter of each strain was measured. All assays were done in triplicate.
Motility tracking
The motility of early-exponential (OD600 of 0.1) V. vulnificus cells that were grown in IO20YP, washed, and resuspended in IO20 was recorded by dark-field microscopy on an Olympus IX83 microscope using a 20x ELWD objective. A stack of at least 150 frames (100 ms exposure time) was recorded for each sample and the movement of 15 randomly sampled motile cells per stack was traced using the MTrackJ47 plugin in Comstat [126,127]. Data from three stacks of three biological replicates were analyzed for each strain. Traces shown are from a single experiment. Violin plots were created with Prism (GraphPad).
Quantification of cellular c-di-GMP concentrations
Intracellular c-di-GMP concentrations were determined with the LUCERNA cyclic-di-GMP Assay Kit (Lucerna, Brooklyn, NY, United States) according to the manufacturer’s instructions. The same procedure was used for both E. coli MG1655 and V. vulnificus. Briefly, cells expressing rcbA, rcbB, rcbAB or their derivatives from pC2X6HT were grown overnight at 37°C with shaking in LB containing the appropriate antibiotics. Samples were diluted 100x into fresh media containing 10 µM IPTG, grown to an OD600 of 1.0 and kept on ice for 30 minutes. The samples were then diluted 1:10 in RNase-free water. Bacterial culture (50 μl) was transferred in triplicate into a 96-well black flat-bottom microtiter plate and 150 μl of reaction mix (50 μl 4X c-di-GMP assay (CA) buffer, 50 μl 4X bacterial compatibility (BC) reagent, 20 μl 10X fluorophore, 10 μl 20X c-di-GMP sensor, and 20 μl RNase-free water) was added to each well. The plate was incubated for 22 hours at room temperature in the dark. After incubation, readings were taken using a BioTek Synergy HT1 microplate reader with excitation set at 469 nm and emission at 501 nm.
Co-immunoprecipitation assays
Motile E. coli DH5α cells that expressed rcbA or rcbB alone or together from pC2X6HT as 6-HIS and HA-tagged fusions, respectively, were used for co-IP assays. Cells were grown overnight in 5 ml LB Ap at 37°C. The next day, cells were diluted 1:100 in 50 ml of fresh media containing IPTG and grown to an OD600 of 0.3. Cells were pelleted and washed twice with 10 ml of ice-cold water. Pellets were resuspended in B-PER Reagent (Thermo Scientific) according to the manufacturer’s instructions and the extracts were centrifuged at 13,000 rpm for 5 minutes to pellet cellular debris. 50 µL of washed HisPur Ni-NTA magnetic beads (Thermo Scientific) was added to the supernatant, which contained the extracted proteins, and the samples were incubated with gentle rolling at room temperature for 30 min. The beads were then washed five times with B-PER to remove non-specifically bound proteins and contaminants. Bound proteins were eluted with buffer containing 250 mM imidazole and separated on NuPAGE 4–12% Bis-Tris pre-cast protein gels (Invitrogen). Western Blot analysis used anti-HIS (BioRad) and anti-HA (Thermo Scientific) HRP antibodies for detection of RcbA and RcbB, respectively.
Fluorescence microscopy
Plasmid expressing mRuby3, mNeonGreen and BFP fusion proteins were conjugated to the indicated V. vulnificus strains and expression was induced with IPTG and/or L-ara. Images of individual cells were captured after cells were mounted on low melting point agarose pads. Images were captured on an Olympus IX83 inverted microscope using a 100 × 1.3 numerical aperture phase contrast objective. Phase contrast and fluorescence images were obtained with a Hamamatsu ORCA-R2 digital charge-coupled device camera, and the light source was the X-cite 120 light-emitting diode (Lumen Dynamics, Mississauga, ON, Canada). Emission filters were purchased from Chroma Technology (Bellows Falls, VT). The specific emission filters were DAPI-5060C-OMF [excitation (EX) filter, 377/50 nm; emission (EM) filter, 447/60 nm; dichroic mirror (DM), 409 nm], GFP-3035D-OMF (EX filter, 473/31 nm; EM filter, 520/35 nm; DM, 495 nm), and mCherry-B-OFF (EX filter, 562/40 nm; EM filter, 641/75 nm; DM, 593 nm). Data from three biological replicates were analyzed for each strain. Images were processed using cellSense software (Olympus) and those presented are from a single representative experiment.
Bacterial-two-hybrid (BACTH) experiments
Two test plasmids (pUT18 and pKNT25) encoding complementary adenylate cyclase fragments fused to full-length RcbA, RcbB or their domains of interest were transformed into E. coli BTH101 cells. The resulting strains were grown overnight in LB that contained the appropriate antibiotics and inducers at 30°C with agitation. Then 3 μl of each bacterial culture was spotted on LB agar plates containing 40 μg ml-1 of X-gal and appropriate antibiotics. The plates were then incubated for 24–48 hours at 30°C and protein interactions were evaluated by both colony growth and development of blue coloration resulting from functional complementation of the split adenylate cyclase protein.
β-Galactosidase assays
Quantification of β-galactosidase activity was carried out following the PopCulture β-gal assay protocol with the following amendments. Briefly, bacteria were grown overnight at 30°C in LB medium with 50 µM IPTG. Bacterial culture (20 μl) was transferred in triplicate into a black 96-well flat-bottom microtiter plate and 180 μl of reaction mix was added to each well (final concentrations of 60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4, 0.1 mg ml-1 lysozyme, 6 mg ml-1 o-nitrophenyl-β-D-galactopyranoside (ONPG), and 4% PopCulture reagent (Millipore). Blank wells contained 20 μl LB medium. The plate was quickly transferred to a BioTek Synergy HT1 microplate reader set to 30°C. OD600 and OD420 readings were taken every 60 s for 30 min. The microtiter plate was shaken at 500 rpm (double orbital) between readings to ensure proper mixing and sample lysis. The slope of OD420 readings over time (OD420 per minute) and the initial OD600 reading were used to calculate Miller units as previously described [128] using the following formula: Miller units = (5,000 × OD420 min−1)/OD600. Data from three technical replicates of three biological replicates were collected for all experiments.
Structural modeling
The structure of RcbA and RcbB complexes were predicted using AlphaFold 3 [129]. Five homology models were generated for each protein, and the model with the best overall agreement score was analyzed further. ChimeraX [130] was used for molecular visualization.
Statistical analyses
Statistical significance was determined by Student's t-test (two-tailed distribution with two-sample, equal variance) when directly comparing two conditions or one-way analysis of variance (ANOVA) followed by pairwise comparisons with a post-hoc adjustment (see figure legends) when comparing data with multiple samples.
Phylogenetic analyses
A protein sequence database was prepared comprising the annotated sequences from the 617 publicly available complete Vibrio genome assemblies. An initial set of potential homologous protein sequence pairs occurring within the same operon were identified by BLASTP [131] (version ncbi-blast-2.13.0+). Multiple sequence alignments (MSA) for these sequences were generated using MUSCLE [132] (version 3.8.31, parameters: -diags -sv -distance1 kbit20_3). Profile HMMs constructed from each set of MSA were searched against the complete Vibrio protein sequence database using HMMER [133] version 3.2.1. The resulting hits were iteratively used to build further profile HMMs while newer hits were being found from the previous iteration. Potential RcbA and RcbB homolog pairs were required to co-occur on the same chromosome or contig sequence and on the same strand, potentially from the same operon. TPR counts per sequence were estimated using TPRpred [134] (with HMM tpr2.8.pp). Single copy core genes were identified from each of the Vibrio assemblies using BUSCO 5.7.1 searching against the vibrionales_odb10 database [135]. MSA was performed on the protein sequences for each of the 133 common core genes from all 617 assemblies using MUSCLE. The individual amino acid residues were substituted with the corresponding codon sequences to obtain the MSA of the coding sequences, while maintaining information on any changes in degenerate positions to preserve codon integrity and the amino acid context. The phylogenetic tree was calculated as the best scoring Maximum Likelihood bootstrap tree using RAxML [136] version 8.2.12 (parameters: -m GTRGAMMA -B 0.03 -# 100). The tree was visualized and exported using the Integrated Tree of Life [137] web service available at https://itol.embl.de.
Supporting information
(DOCX)
(XLSX)
(XLSX)
Plot of wildtype V. vulnificus (WT) and ∆rcbAB mutant (Tn) strains bearing a PcabAlacZ reporter grown in LB without or supplemented with 15 mM CaCl2 (LB + Ca2+). Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences between samples (****p < 0.0001; ns, no significant difference) were determined by unpaired student’s t-test (two-tailed).
(TIFF)
Bar plot of the motility zones for wildtype V. vulnificus and the rcbA, rcbB and rcbAB deletion mutants (∆rcbA, ∆rcbB and ∆rcbAB, respectively) in soft agar lacking (orange) or supplemented with the indicated concentration of CaCl2. Bars show the respective mean values for each strain and error bars represent the standard deviation of triplicate assays. No statistically significant difference (ns) relative to WT was found among strains at the same CaCl2 concentration (one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test).
(TIFF)
Plot of motility by wildtype V. vulnificus and the rcbAB deletion mutant (∆rcbAB) carrying the empty expression plasmid or expressing (+) rcbA, rcbB or rcbAB grown in soft agar lacking (orange) or supplemented with the indicated concentration of CaCl2. Bars show the respective mean values for each strain and error bars represent the standard deviation of triplicate assays. Statistically significant differences relative to wildtype were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test (**p < 0.01; ns, no significant difference). Expression was induced with IPTG (10 µM).
(TIFF)
Plot of intracellular c-di-GMP levels in wildtype V. vulnificus and the rcbAB deletion mutant (∆rcbAB) that carries the empty expression plasmid or express (+) rcbA, rcbB or rcbAB grown in LB lacking (orange) or supplemented with the indicated concentration of CaCl2. Bars show the respective mean values for each strain and error bars represent the standard deviation of triplicate assays. Statistically significant differences relative to wildtype were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test (***p < 0.001; ns, no significant difference). Expression was induced with IPTG (10 µM).
(TIFF)
A, plot of biofilm formation (OD595/OD600) by ∆rcbB cells harboring the empty expression plasmid (control) or plasmids expressing (+) rcbB or a catalytic mutant bearing the A-to-E mutation, GGAEF). B, a plot of swimming motility zones in IO20 soft agar by the same strains in A. Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences (****p < 0.0001; ns, no significant difference) relative to the control were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test. Expression was induced with 1 µM IPTG in A and 10 µM IPTG in B.
(TIFF)
Plot of swimming motility zones in soft agar for several E. coli strains (A) and for MG1655 cells (B) carrying the empty expression vector (Con) or the plasmid with rcbA, rcbB or rcbAB (without or with (+) 10 µM IPTG; separated by the vertical dotted line). Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences (*p < 0.05, ****p < 0.0001; ns, no significant difference) relative to the control were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test.
(TIFF)
A, western blot of ∆rcbA extracts containing HIS-tagged RcbA or ∆rcbB extracts containing HA-tagged RcbB. Expression was induced with the IPTG concentrations indicated below each lane and the proteins were detected with anti-HIS and anti-HA antibodies, respectively. B, alignments of AlphaFold 3 predicted monomer structures of RcbA (purple) with RcbA∆TPR (pink), and RcbB (green) with RcbB∆TPR (blue). C, western blot of ∆rcbA extracts containing HIS-tagged RcbA∆TPR or HA-tagged RcbB∆TPR. Expression and detection conditions were as in A.
(TIFF)
Biofilm formation (OD595/OD600) by ∆rcbAB cells harboring empty plasmids (control) or expressing (+) rcbA-mRuby3 (RcbAmRu), rcbB-mNeonGreen (RcbBmNG) or rcbA-mRuby3-rcbB-mNeonGreen (RcbAmRu/RcbBmNG). Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences (****p < 0.0001; ns, no significant difference) relative to control cells were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test. Expression was induced with 1 µM IPTG.
(TIFF)
Representative fluorescence images of NanoOrange stained ∆hubP and ∆flrA cells. White arrows denote flagella. Scale bar = 1 µm.
(TIFF)
From left to right, representative phase, fluorescence and overlay images of RcbA-mRuby3 (A), RcbB-mNeonGreen (B), or both (C) in ∆flrA∆rcbAB cells. Scale bar = 1 μm. Expression was induced with 10 µM IPTG.
(TIFF)
AlphaFold 3 predicted monomer structures of RcbB alone (blue) and complexed (green) with RcbA. A GTP molecule (stick configuration) is modeled bound at the active site. The grey arrow denotes rotation of the DGC domain relative to the aligned TPR domains.
(TIFF)
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This work was supported by funding from Indiana University to DRM and the IUB-Colonel Bayard Franklin Floyd Memorial Fund in Microbiology to SM. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Sampaio A, Silva V, Poeta P, Aonofriesei F. Vibrio spp.: Life Strategies, Ecology, and Risks in a Changing Environment. Diversity. 2022;14(2):97. [Google Scholar]
- 2.Grimes DJ. The Vibrios: Scavengers, Symbionts, and Pathogens from the Sea. Microb Ecol. 2020;80(3):501–6. doi: 10.1007/s00248-020-01524-7 [DOI] [PubMed] [Google Scholar]
- 3.Yildiz FH, Visick KL. Vibrio biofilms: so much the same yet so different. Trends Microbiol. 2009;17(3):109–18. doi: 10.1016/j.tim.2008.12.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Baker-Austin C, Oliver JD, Alam M, Ali A, Waldor MK, Qadri F, et al. Vibrio spp. infections. Nat Rev Dis Primers. 2018;4(1):1–19. [DOI] [PubMed] [Google Scholar]
- 5.Thompson F, Iida T, Swings J. Biodiversity of vibrios. Microbiol Mol Biol Rev. 2004;68(3):403–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Vezzulli L, Colwell RR, Pruzzo C. Ocean warming and spread of pathogenic vibrios in the aquatic environment. Microb Ecol. 2013;65(4):817–25. doi: 10.1007/s00248-012-0163-2 [DOI] [PubMed] [Google Scholar]
- 7.Bonnin-Jusserand M, Copin S, Le Bris C, Brauge T, Gay M, Brisabois A, et al. Vibrio species involved in seafood-borne outbreaks (Vibrio cholerae, V. parahaemolyticus and V. vulnificus): Review of microbiological versus recent molecular detection methods in seafood products. Crit Rev Food Sci Nutr. 2019;59(4):597–610. doi: 10.1080/10408398.2017.1384715 [DOI] [PubMed] [Google Scholar]
- 8.Morris JG Jr. Cholera and other types of vibriosis: a story of human pandemics and oysters on the half shell. Clin Infect Dis. 2003;37(2):272–80. doi: 10.1086/375600 [DOI] [PubMed] [Google Scholar]
- 9.Haftel A, Sharman T. Vibrio vulnificus Infection. Treasure Island, FL: StatPearls Publishing; 2023. [PubMed] [Google Scholar]
- 10.Coerdt KM, Khachemoune A. Vibrio vulnificus: Review of Mild to Life-threatening Skin Infections. Cutis. 2021;107(2):E12–7. doi: 10.12788/cutis.0183 [DOI] [PubMed] [Google Scholar]
- 11.Oliver JD. The Biology of Vibrio vulnificus. Microbiol Spectr. 2015;3(3):10.1128/microbiolspec.VE-0001–2014. doi: 10.1128/microbiolspec.VE-0001-2014 [DOI] [PubMed] [Google Scholar]
- 12.Archer EJ, Baker-Austin C, Osborn TJ, Jones NR, Martínez-Urtaza J, Trinanes J, et al. Climate warming and increasing Vibrio vulnificus infections in North America. Sci Rep. 2023;13(1):3893. doi: 10.1038/s41598-023-28247-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hoffmann S, Maculloch B, Batz M. Economic Burden of Major Foodborne Illnesses Acquired in the United States. U.S. Department of Agriculture, Economic Research Service; 2015. May. (Economic Information Bulletin). Report No.: 140. [Google Scholar]
- 14.DePaola A, Capers GM, Alexander D. Densities of Vibrio vulnificus in the intestines of fish from the U.S. Gulf Coast. Appl Environ Microb. 1994;60(3):984–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Amaro C, Sanjuan E, Fouz B, Pajuelo D, Lee CT, Hor LI, et al. The fish pathogen Vibrio vulnificus biotype 2: epidemiology, phylogeny, and virulence factors involved in warm-water vibriosis. Microbiol Spectr. 2015;3(3). [DOI] [PubMed] [Google Scholar]
- 16.Marco-Noales E, Milán M, Fouz B, Sanjuán E, Amaro C. Transmission to eels, portals of entry, and putative reservoirs of Vibrio vulnificus serovar E (biotype 2). Appl Environ Microbiol. 2001;67(10):4717–25. doi: 10.1128/AEM.67.10.4717-4725.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hernández‐Cabanyero C, Amaro C. Phylogeny and life cycle of the zoonotic pathogen Vibrio vulnificus. Environ Microbiol. 2020;22(10):4133–48. [DOI] [PubMed] [Google Scholar]
- 18.Montanari MP, Pruzzo C, Pane L, Colwell RR. Vibrios associated with plankton in a coastal zone of the Adriatic Sea (Italy). FEMS Microbiol Ecol. 1999;29(3):241–7. doi: 10.1111/j.1574-6941.1999.tb00615.x [DOI] [Google Scholar]
- 19.Dalsgaa A, Høi L. Prevalence and characterization of Vibrio vulnificus isolated from shrimp products imported into Denmark. J Food Prot. 2016;60(9):1132–4. [DOI] [PubMed] [Google Scholar]
- 20.Teng T, Liang L, Chen K, Xi B, Xie J, Xu P. Isolation, identification and phenotypic and molecular characterization of pathogenic Vibrio vulnificus isolated from Litopenaeus vannamei. PLoS One. 2017;12(10):e0186135. doi: 10.1371/journal.pone.0186135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Froelich B, Ayrapetyan M, Oliver JD. Integration of Vibrio vulnificus into marine aggregates and its subsequent uptake by Crassostrea virginica oysters. Appl Environ Microbiol. 2013;79(5):1454–8. doi: 10.1128/AEM.03095-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wright AC, Hill RT, Johnson JA, Roghman MC, Colwell RR, Morris JG Jr. Distribution of Vibrio vulnificus in the Chesapeake Bay. Appl Environ Microbiol. 1996;62(2):717–24. doi: 10.1128/aem.62.2.717-724.1996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial biofilms: from the natural environment to infectious diseases. Nat Rev Microbiol. 2004;2(2):95–108. doi: 10.1038/nrmicro821 [DOI] [PubMed] [Google Scholar]
- 24.Fisher RA, Gollan B, Helaine S. Persistent bacterial infections and persister cells. Nat Rev Microbiol. 2017;15(8):453–64. doi: 10.1038/nrmicro.2017.42 [DOI] [PubMed] [Google Scholar]
- 25.Raina J-B, Eme L, Pollock FJ, Spang A, Archibald JM, Williams TA. Symbiosis in the microbial world: from ecology to genome evolution. Biol Open. 2018;7(2):bio032524. doi: 10.1242/bio.032524 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Balcázar JL, Subirats J, Borrego CM. The role of biofilms as environmental reservoirs of antibiotic resistance. Front Microbiol. 2015;6:1216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lewis K. Persister cells, dormancy and infectious disease. Nat Rev Microbiol. 2007;5(1):48–56. doi: 10.1038/nrmicro1557 [DOI] [PubMed] [Google Scholar]
- 28.Flemming H-C, Wingender J, Szewzyk U, Steinberg P, Rice SA, Kjelleberg S. Biofilms: an emergent form of bacterial life. Nat Rev Microbiol. 2016;14(9):563–75. doi: 10.1038/nrmicro.2016.94 [DOI] [PubMed] [Google Scholar]
- 29.Valentini M, Filloux A. Biofilms and cyclic di-GMP (c-di-GMP) signaling: lessons from Pseudomonas aeruginosa and other bacteria. J Biol Chem. 2016;291(24):12547–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chou SH, Galperin MY. Diversity of c-di-GMP-binding proteins and mechanisms. J Bacteriol. 2015;198(1):32–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jenal U, Malone J. Mechanisms of cyclic-di-GMP signaling in bacteria. Annu Rev Genet. 2006;40:385–407. doi: 10.1146/annurev.genet.40.110405.090423 [DOI] [PubMed] [Google Scholar]
- 32.Römling U, Gomelsky M, Galperin MY. C-di-GMP: the dawning of a novel bacterial signalling system. Mol Microbiol. 2005;57(3):629–39. [DOI] [PubMed] [Google Scholar]
- 33.Jenal U, Reinders A, Lori C. Cyclic di-GMP: second messenger extraordinaire. Nat Rev Microbiol. 2017;15(5):271–84. doi: 10.1038/nrmicro.2016.190 [DOI] [PubMed] [Google Scholar]
- 34.Römling U, Amikam D. Cyclic di-GMP as a second messenger. Curr Opin Microbiol. 2006;9(2):218–28. doi: 10.1016/j.mib.2006.02.010 [DOI] [PubMed] [Google Scholar]
- 35.Upadhyay AA, Fleetwood AD, Adebali O, Finn RD, Zhulin IB. Cache Domains That are Homologous to, but Different from PAS Domains Comprise the Largest Superfamily of Extracellular Sensors in Prokaryotes. PLoS Comput Biol. 2016;12(4):e1004862. doi: 10.1371/journal.pcbi.1004862 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Randall TE, Eckartt K, Kakumanu S, Price-Whelan A, Dietrich LEP, Harrison JJ. Sensory Perception in Bacterial Cyclic Diguanylate Signal Transduction. J Bacteriol. 2022;204(2):e00433-21. doi: 10.1128/JB.00433-21 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kunz S, Graumann PL. Spatial organization enhances versatility and specificity in cyclic di-GMP signaling. Biol Chem. 2020;401(12):1323–34. doi: 10.1515/hsz-2020-0202 [DOI] [PubMed] [Google Scholar]
- 38.Hengge R. Trigger phosphodiesterases as a novel class of c-di-GMP effector proteins. Philos Trans R Soc Lond B Biol Sci. 2016;371(1707):20150498. doi: 10.1098/rstb.2015.0498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Amikam D, Galperin MY. PilZ domain is part of the bacterial c-di-GMP binding protein. Bioinformatics. 2006;22(1):3–6. doi: 10.1093/bioinformatics/bti739 [DOI] [PubMed] [Google Scholar]
- 40.Fang X, Ahmad I, Blanka A, Schottkowski M, Cimdins A, Galperin MY, et al. GIL, a new c-di-GMP-binding protein domain involved in regulation of cellulose synthesis in enterobacteria. Mol Microbiol. 2014;93(3):439–52. doi: 10.1111/mmi.12672 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Baraquet C, Harwood CS. Cyclic diguanosine monophosphate represses bacterial flagella synthesis by interacting with the Walker A motif of the enhancer-binding protein FleQ. Proc Natl Acad Sci U S A. 2013;110(46):18478–83. doi: 10.1073/pnas.1318972110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hickman JW, Harwood CS. Identification of FleQ from Pseudomonas aeruginosa as a c-di-GMP-responsive transcription factor. Mol Microbiol. 2008;69(2):376–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Krasteva PV, Fong JCN, Shikuma NJ, Beyhan S, Navarro MVAS, Yildiz FH, et al. Vibrio cholerae VpsT regulates matrix production and motility by directly sensing cyclic di-GMP. Science. 2010;327(5967):866–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Leduc JL, Roberts GP. Cyclic di-GMP allosterically inhibits the CRP-like protein (Clp) of Xanthomonas axonopodis pv. citri. J Bacteriol. 2009;191(22):7121–2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lee VT, Matewish JM, Kessler JL, Hyodo M, Hayakawa Y, Lory S. A cyclic-di-GMP receptor required for bacterial exopolysaccharide production. Mol Microbiol. 2007;65(6):1474–84. doi: 10.1111/j.1365-2958.2007.05879.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tschowri N, Schumacher MA, Schlimpert S, Chinnam NB, Findlay KC, Brennan RG, et al. Tetrameric c-di-GMP mediates effective transcription factor dimerization to control Streptomyces development. Cell. 2014;158(5):1136–47. doi: 10.1016/j.cell.2014.07.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Pursley BR, Maiden MM, Hsieh M-L, Fernandez NL, Severin GB, Waters CM. Cyclic di-GMP Regulates TfoY in Vibrio cholerae To Control Motility by both Transcriptional and Posttranscriptional Mechanisms. J Bacteriol. 2018;200(7):e00578-17. doi: 10.1128/JB.00578-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kulshina N, Baird NJ, Ferré-D’Amaŕe AR. Recognition of the bacterial second messenger cyclic diguanylate by its cognate riboswitch. Nat Struct Mol Biol. 2009;16(12):1212–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Pursley BR, Fernandez NL, Severin GB, Waters CM. The Vc2 cyclic di-GMP-dependent riboswitch of Vibrio cholerae regulates expression of an upstream putative small RNA by controlling RNA stability. J Bacteriol. 2019;201(21). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Pecina A, Schwan M, Blagotinsek V, Rick T, Klüber P, Leonhard T, et al. The Stand-Alone PilZ-Domain Protein MotL Specifically Regulates the Activity of the Secondary Lateral Flagellar System in Shewanella putrefaciens. Front Microbiol. 2021;12:668892. doi: 10.3389/fmicb.2021.668892 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kuzmich S, Skotnicka D, Szadkowski D, Klos P, Pérez-Burgos M, Schander E. Three PilZ domain proteins, PlpA, PixA and PixB, have distinct functions in regulation of motility and development in Myxococcus xanthus. Biorxiv. 2021;2021.03.04.433885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ryjenkov DA, Simm R, Römling U, Gomelsky M. The PilZ domain is a receptor for the second messenger c-di-GMP: the PilZ domain protein YcgR controls motility in enterobacteria. J Biol Chem. 2006;281(41):30310–4. doi: 10.1074/jbc.C600179200 [DOI] [PubMed] [Google Scholar]
- 53.Baker AE, Diepold A, Kuchma SL, Scott JE, Ha DG, Orazi G, et al. PilZ domain protein FlgZ mediates cyclic di-GMP-dependent swarming motility control in Pseudomonas aeruginosa. J Bacteriol. 2016;198(13):1837–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ko J, Ryu K-S, Kim H, Shin J-S, Lee J-O, Cheong C, et al. Structure of PP4397 reveals the molecular basis for different c-di-GMP binding modes by Pilz domain proteins. J Mol Biol. 2010;398(1):97–110. doi: 10.1016/j.jmb.2010.03.007 [DOI] [PubMed] [Google Scholar]
- 55.Schumacher MA, Zeng W. Structures of the activator of K. pneumonia biofilm formation, MrkH, indicates PilZ domains involved in c-di-GMP and DNA binding. Proc Natl Acad Sci U S A. 2016;113(36):10067–72. doi: 10.1073/pnas.1607503113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chodur DM, Coulter P, Isaacs J, Pu M, Fernandez N, Waters CM, et al. Environmental Calcium Initiates a Feed-Forward Signaling Circuit That Regulates Biofilm Formation and Rugosity in Vibrio vulnificus. mBio. 2018;9(4):e01377-18. doi: 10.1128/mBio.01377-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lee H, Im H, Hwang S-H, Ko D, Choi SH. Two novel genes identified by large-scale transcriptomic analysis are essential for biofilm and rugose colony development of Vibrio vulnificus. PLoS Pathog. 2023;19(1):e1011064. doi: 10.1371/journal.ppat.1011064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Park JH, Jo Y, Jang SY, Kwon H, Irie Y, Parsek MR, et al. Correction: The cabABC Operon Essential for Biofilm and Rugose Colony Development in Vibrio vulnificus. PLoS Pathog. 2015;11(10):e1005252. doi: 10.1371/journal.ppat.1005252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Pu M, Storms E, Chodur DM, Rowe-Magnus DA. Calcium-dependent site-switching regulates expression of the atypical iam pilus locus in Vibrio vulnificus. Environ Microbiol. 2020;22(10):4167–82. doi: 10.1111/1462-2920.14763 [DOI] [PubMed] [Google Scholar]
- 60.Garrison-Schilling KL, Grau BL, McCarter KS, Olivier BJ, Comeaux NE, Pettis GS. Calcium promotes exopolysaccharide phase variation and biofilm formation of the resulting phase variants in the human pathogen Vibrio vulnificus. Environ Microbiol. 2011;13(3):643–54. doi: 10.1111/j.1462-2920.2010.02369.x [DOI] [PubMed] [Google Scholar]
- 61.Guo Y, Rowe-Magnus DA. Overlapping and unique contributions of two conserved polysaccharide loci in governing distinct survival phenotypes in Vibrio vulnificus. Environ Microbiol. 2011;13(11):2888–990. [DOI] [PubMed] [Google Scholar]
- 62.Chodur DM, Guo L, Pu M, Bruger E, Fernandez N, Waters C, et al. The proline variant of the W[F/L/M][T/S]R cyclic di-GMP binding motif suppresses dependence on signal association for regulator function. J Bacteriol. 2017;199(19):e00344–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Chodur DM, Rowe-Magnus DA. Complex Control of a Genomic Island Governing Biofilm and Rugose Colony Development in Vibrio vulnificus. J Bacteriol. 2018;200(16):e00190-18. doi: 10.1128/JB.00190-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hwang SH, Im H, Choi SH. A master regulator brpR coordinates the expression of multiple loci for robust biofilm and rugose colony development in Vibrio vulnificus. Front Microbiol. 2021;12:679854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Guo Y, Rowe-Magnus DA. Identification of a c-di-GMP-regulated polysaccharide locus governing stress resistance and biofilm and rugose colony formation in Vibrio vulnificus. Infect Immun. 2010;78(3):1390–402. doi: 10.1128/IAI.01188-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Hwang S-H, Park JH, Lee B, Choi SH. A Regulatory Network Controls cabABC Expression Leading to Biofilm and Rugose Colony Development in Vibrio vulnificus. Front Microbiol. 2020;10:3063. doi: 10.3389/fmicb.2019.03063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Han Q, Wang S-F, Qian X-X, Guo L, Shi Y-F, He R, et al. Flagellar brake protein YcgR interacts with motor proteins MotA and FliG to regulate the flagellar rotation speed and direction. Front Microbiol. 2023;14:1159974. doi: 10.3389/fmicb.2023.1159974 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Paul K, Nieto V, Carlquist WC, Blair DF, Harshey RM. The c-di-GMP binding protein YcgR controls flagellar motor direction and speed to affect chemotaxis by a “backstop brake” mechanism. Mol Cell. 2010;38(1):128–39. doi: 10.1016/j.molcel.2010.03.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chen T, Pu M, Subramanian S, Kearns D, Rowe-Magnus D. PlzD modifies Vibrio vulnificus foraging behavior and virulence in response to elevated c-di-GMP. mBio. 2023:e01536-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Sikorski RS, Boguski MS, Goebl M, Hieter P. A repeating amino acid motif in CDC23 defines a family of proteins and a new relationship among genes required for mitosis and RNA synthesis. Cell. 1990;60(2):307–17. doi: 10.1016/0092-8674(90)90745-z [DOI] [PubMed] [Google Scholar]
- 71.Jernigan KK, Bordenstein SR. Tandem-repeat protein domains across the tree of life. PeerJ. 2015;3:e732. doi: 10.7717/peerj.732 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.D’Andrea LD, Regan L. TPR proteins: the versatile helix. Trends Biochem Sci. 2003;28(12):655–62. doi: 10.1016/j.tibs.2003.10.007 [DOI] [PubMed] [Google Scholar]
- 73.Cerveny L, Straskova A, Dankova V, Hartlova A, Ceckova M, Staud F, et al. Tetratricopeptide repeat motifs in the world of bacterial pathogens: role in virulence mechanisms. Infect Immun. 2013;81(3):629–35. doi: 10.1128/IAI.01035-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Goebl M, Yanagida M. The TPR snap helix: a novel protein repeat motif from mitosis to transcription. Trends Biochem Sci. 1991;16(5):173–7. doi: 10.1016/0968-0004(91)90070-c [DOI] [PubMed] [Google Scholar]
- 75.Zeytuni N, Zarivach R. Structural and functional discussion of the tetra-trico-peptide repeat, a protein interaction module. Structure. 2012;20(3):397–405. doi: 10.1016/j.str.2012.01.006 [DOI] [PubMed] [Google Scholar]
- 76.Blatch GL, Lässle M. The tetratricopeptide repeat: a structural motif mediating protein-protein interactions. Bioessays. 1999;21(11):932–9. doi: [DOI] [PubMed] [Google Scholar]
- 77.Sinars CR, Cheung-Flynn J, Rimerman RA, Scammell JG, Smith DF, Clardy J. Structure of the large FK506-binding protein FKBP51, an Hsp90-binding protein and a component of steroid receptor complexes. Proc Natl Acad Sci. 2003;100(3):868–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Abe Y, Shodai T, Muto T, Mihara K, Torii H, Nishikawa S, et al. Structural basis of presequence recognition by the mitochondrial protein import receptor Tom20. Cell. 2000;100(5):551–60. doi: 10.1016/s0092-8674(00)80691-1 [DOI] [PubMed] [Google Scholar]
- 79.Goodarzi MO, Xu N, Cui J, Guo X, Chen YI, Azziz R. Small glutamine-rich tetratricopeptide repeat-containing protein alpha (SGTA), a candidate gene for polycystic ovary syndrome. Hum Reprod. 2008;23(5):1214–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Sohocki MM, Bowne SJ, Sullivan LS, Blackshaw S, Cepko CL, Payne AM, et al. Mutations in a new photoreceptor-pineal gene on 17p cause Leber congenital amaurosis. Nat Genet. 2000;24(1):79–83. doi: 10.1038/71732 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Tsukahara F, Urakawa I, Hattori M, Hirai M, Ohba K- i, Yoshioka T, et al. Molecular characterization of the mouse mtprd gene, a homologue of human tprd: unique gene expression suggesting its critical role in the pathophysiology of down syndrome. J Biochem. 1998;123(6):1055–63. [DOI] [PubMed] [Google Scholar]
- 82.Bröms JE, Edqvist PJ, Forsberg A, Francis MS. Tetratricopeptide repeats are essential for PcrH chaperone function in Pseudomonas aeruginosa type III secretion. FEMS Microbiol Lett. 2006;256(1):57–66. doi: 10.1111/j.1574-6968.2005.00099.x [DOI] [PubMed] [Google Scholar]
- 83.Edqvist PJ, Bröms JE, Betts HJ, Forsberg A, Pallen MJ, Francis MS. Tetratricopeptide repeats in the type III secretion chaperone, LcrH: their role in substrate binding and secretion. Mol Microbiol. 2006;59(1):31–44. doi: 10.1111/j.1365-2958.2005.04923.x [DOI] [PubMed] [Google Scholar]
- 84.Chakraborty S, Monfett M, Maier TM, Benach JL, Frank DW, Thanassi DG. Type IV pili in Francisella tularensis: roles of pilF and pilT in fiber assembly, host cell adherence, and virulence. Infect Immun. 2008;76(7):2852–61. doi: 10.1128/IAI.01726-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Chao J, Wong D, Zheng X, Poirier V, Bach H, Hmama Z, et al. Protein kinase and phosphatase signaling in Mycobacterium tuberculosis physiology and pathogenesis. Biochim Biophys Acta. 2010;1804(3):620–7. doi: 10.1016/j.bbapap.2009.09.008 [DOI] [PubMed] [Google Scholar]
- 86.Mittl PRE, Schneider-Brachert W. Sel1-like repeat proteins in signal transduction. Cell Signal. 2007;19(1):20–31. [DOI] [PubMed] [Google Scholar]
- 87.Taute KM, Gude S, Tans SJ, Shimizu TS. High-throughput 3D tracking of bacteria on a standard phase contrast microscope. Nat Commun. 2015;6(1):8776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Son K, Guasto JS, Stocker R. Bacteria can exploit a flagellar buckling instability to change direction. Nat Phys. 2013;9(8):494–8. [Google Scholar]
- 89.Arroyo-Pérez EE, Hook JC, Alvarado A, Wimmi S, Glatter T, Thormann K, et al. A conserved cell-pole determinant organizes proper polar flagellum formation. eLife. 2024;13(32):RP93004. doi: 10.7554/elife.93004.3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Yamaichi Y, Bruckner R, Ringgaard S, Möll A, Cameron DE, Briegel A, et al. A multidomain hub anchors the chromosome segregation and chemotactic machinery to the bacterial pole. Genes Dev. 2012;26(20):2348–60. doi: 10.1101/gad.199869.112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Altinoglu I, Abriat G, Carreaux A, Torres-Sánchez L, Poidevin M, Krasteva PV, et al. Analysis of HubP-dependent cell pole protein targeting in Vibrio cholerae uncovers novel motility regulators. PLoS Genet. 2022;18(1):e1009991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Takekawa N, Kwon S, Nishioka N, Kojima S, Homma M. HubP, a polar landmark protein, regulates flagellar number by assisting in the proper polar localization of FlhG in Vibrio alginolyticus. J Bacteriol. 2016;198(22):3091–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Homma M, Kobayakawa T, Hao Y, Nishikino T, Kojima S. Function and Structure of FlaK, a Master Regulator of the Polar Flagellar Genes in Marine Vibrio. J Bacteriol. 2022;204(11):e0032022. doi: 10.1128/jb.00320-22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Millikan DS, Ruby EG. FlrA, a sigma54-dependent transcriptional activator in Vibrio fischeri, is required for motility and symbiotic light-organ colonization. J Bacteriol. 2003;185(12):3547–57. doi: 10.1128/JB.185.12.3547-3557.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Prouty MG, Correa NE, Klose KE. The novel sigma54- and sigma28-dependent flagellar gene transcription hierarchy of Vibrio cholerae. Mol Microbiol. 2001;39(6):1595–609. doi: 10.1046/j.1365-2958.2001.02348.x [DOI] [PubMed] [Google Scholar]
- 96.Tolker-Nielsen T. Biofilm Development. Microbiol Spectr. 2015;3(2):MB-0001-2014. [DOI] [PubMed] [Google Scholar]
- 97.Dahlstrom KM, O’Toole GA. A Symphony of Cyclases: Specificity in Diguanylate Cyclase Signaling. Annu Rev Microbiol. 2017;71:179–95. doi: 10.1146/annurev-micro-090816-093325 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Kimbrough JH, McCarter LL. Identification of Three New GGDEF and EAL Domain-Containing Proteins Participating in the Scr Surface Colonization Regulatory Network in Vibrio parahaemolyticus. J Bacteriol. 2021;203(4):e00409-20. doi: 10.1128/JB.00409-20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Ryjenkov DA, Tarutina M, Moskvin OV, Gomelsky M. Cyclic diguanylate is a ubiquitous signaling molecule in bacteria: insights into biochemistry of the GGDEF protein domain. J Bacteriol. 2005;187(5):1792–8. doi: 10.1128/JB.187.5.1792-1798.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Mantoni F, Scribani Rossi C, Paiardini A, Di Matteo A, Cappellacci L, Petrelli R, et al. Studying GGDEF Domain in the Act: Minimize Conformational Frustration to Prevent Artefacts. Life (Basel). 2021;11(1):31. doi: 10.3390/life11010031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Paul R, Abel S, Wassmann P, Beck A, Heerklotz H, Jenal U. Activation of the diguanylate cyclase PleD by phosphorylation-mediated dimerization. J Biol Chem. 2007;282(40):29170–7. doi: 10.1074/jbc.M704702200 [DOI] [PubMed] [Google Scholar]
- 102.Srivastava D, Harris RC, Waters CM. Integration of cyclic di-GMP and quorum sensing in the control of vpsT and aphA in Vibrio cholerae. J Bacteriol. 2011;193(22):6331–41. doi: 10.1128/JB.05167-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Kim HK, Harshey RM. A diguanylate cyclase acts as a cell division inhibitor in a two-step response to reductive and envelope stresses. Mbio. 2016;7(4):e00822-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Paul R, Weiser S, Amiot NC, Chan C, Schirmer T, Giese B, et al. Cell cycle-dependent dynamic localization of a bacterial response regulator with a novel di-guanylate cyclase output domain. Genes Dev. 2004;18(6):715–27. doi: 10.1101/gad.289504 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Nicastro GG, Kaihami GH, Pulschen AA, Hernandez-Montelongo J, Boechat AL, de Pereira TO, et al. c-di-GMP-related phenotypes are modulated by the interaction between a diguanylate cyclase and a polar hub protein. Sci Rep. 2020;10(1):3077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Kondo S, Imura Y, Mizuno A, Homma M, Kojima S. Biochemical analysis of GTPase FlhF which controls the number and position of flagellar formation in marine Vibrio. Sci Rep. 2018;8(1):12115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Kojima S, Terashima H, Homma M. Regulation of the Single Polar Flagellar Biogenesis. Biomolecules. 2020;10(4):533. doi: 10.3390/biom10040533 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Green JCD, Kahramanoglou C, Rahman A, Pender AMC, Charbonnel N, Fraser GM. Recruitment of the earliest component of the bacterial flagellum to the old cell division pole by a membrane-associated signal recognition particle family GTP-binding protein. J Mol Biol. 2009;391(4):679–90. doi: 10.1016/j.jmb.2009.05.075 [DOI] [PubMed] [Google Scholar]
- 109.Arroyo-Pérez EE, Ringgaard S. Interdependent Polar Localization of FlhF and FlhG and Their Importance for Flagellum Formation of Vibrio parahaemolyticus. Front Microbiol. 2021;12:655239. doi: 10.3389/fmicb.2021.655239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Scheufler C, Brinker A, Bourenkov G, Pegoraro S, Moroder L, Bartunik H, et al. Structure of TPR domain-peptide complexes: critical elements in the assembly of the Hsp70-Hsp90 multichaperone machine. Cell. 2000;101(2):199–210. doi: 10.1016/S0092-8674(00)80830-2 [DOI] [PubMed] [Google Scholar]
- 111.Cortajarena AL, Regan L. Ligand binding by TPR domains. Protein Sci. 2006;15(5):1193–8. doi: 10.1110/ps.062092506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Wang Y, Andole Pannuri A, Ni D, Zhou H, Cao X, Lu X, et al. Structural Basis for Translocation of a Biofilm-supporting Exopolysaccharide across the Bacterial Outer Membrane. J Biol Chem. 2016;291(19):10046–57. doi: 10.1074/jbc.M115.711762 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Pfoh R, Subramanian AS, Huang J, Little DJ, Forman A, DiFrancesco BR, et al. The TPR domain of PgaA is a multifunctional scaffold that binds PNAG and modulates PgaB-dependent polymer processing. PLoS Pathog. 2022;18(8):e1010750. doi: 10.1371/journal.ppat.1010750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Visick KL, Stabb EV, Ruby EG. A lasting symbiosis: how Vibrio fischeri finds a squid partner and persists within its natural host. Nat Rev Microbiol. 2021;19(10):654–65. doi: 10.1038/s41579-021-00557-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Nyholm SV, McFall-Ngai MJ. A lasting symbiosis: how the Hawaiian bobtail squid finds and keeps its bioluminescent bacterial partner. Nat Rev Microbiol. 2021;19(10):666–79. doi: 10.1038/s41579-021-00567-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Mandel MJ, Wollenberg MS, Stabb EV, Visick KL, Ruby EG. A single regulatory gene is sufficient to alter bacterial host range. Nature. 2009;458(7235):215–8. doi: 10.1038/nature07660 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Sawabe T, Sugimura I, Ohtsuka M, Nakano K, Tajima K, Ezura Y, et al. Vibrio halioticoli sp. nov., a non-motile alginolytic marine bacterium isolated from the gut of the abalone Haliotis discus hannai. Int J Syst Bacteriol. 1998;48 Pt 2:573–80. doi: 10.1099/00207713-48-2-573 [DOI] [PubMed] [Google Scholar]
- 118.Qiao Y, Wang J, Mao Y, Liu M, Chen R, Su Y, et al. Pathogenic bacterium Vibrio harveyi: an endosymbiont in the marine parasitic ciliate protozoan Cryptocaryon irritans. Acta Oceanol Sin. 2017;36(10):115–9. doi: 10.1007/s13131-017-1050-y [DOI] [Google Scholar]
- 119.Zhou Y, Chang J, Zhang M, Li X, Luo X, Li W, et al. GefB, a GGDEF domain-containing protein, affects motility and biofilm formation of Vibrio parahaemolyticus and is regulated by quorum sensing regulators. Gene. 2025;933:148968. doi: 10.1016/j.gene.2024.148968 [DOI] [PubMed] [Google Scholar]
- 120.Römling U, Gomelsky M, Galperin MY. C-di-GMP: the dawning of a novel bacterial signalling system. Mol Microbiol. 2005;57(3):629–39. doi: 10.1111/j.1365-2958.2005.04697.x [DOI] [PubMed] [Google Scholar]
- 121.Rusch DB, Rowe-Magnus DA. Complete Genome Sequence of the Pathogenic Vibrio vulnificus Type Strain ATCC 27562. Genome Announc. 2017;5(35):e00907–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Edwards RA, Keller LH, Schifferli DM. Improved allelic exchange vectors and their use to analyze 987P fimbria gene expression. Gene. 1998;207(2):149–57. doi: 10.1016/s0378-1119(97)00619-7 [DOI] [PubMed] [Google Scholar]
- 123.Nakhamchik A, Wilde C, Rowe-Magnus DA. Identification of a Wzy polymerase required for group IV capsular polysaccharide and lipopolysaccharide biosynthesis in Vibrio vulnificus. Infect Immun. 2007;75(12):5550–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Dalia AB, Lazinski DW, Camilli A. Identification of a membrane-bound transcriptional regulator that links chitin and natural competence in Vibrio cholerae. mBio. 2014;5(1):e01028-13. doi: 10.1128/mBio.01028-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Pu M, Rowe-Magnus DA. A Tad pilus promotes the establishment and resistance of Vibrio vulnificus biofilms to mechanical clearance. NPJ Biofilms Microbiomes. 2018;4:10. doi: 10.1038/s41522-018-0052-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Heydorn A, Ersbøll BK, Hentzer M, Parsek MR, Givskov M, Molin S. Experimental reproducibility in flow-chamber biofilms. Microbiology (Reading). 2000;146(Pt 10):2409–15. doi: 10.1099/00221287-146-10-2409 [DOI] [PubMed] [Google Scholar]
- 127.Heydorn A, Nielsen AT, Hentzer M, Sternberg C, Givskov M, Ersbøll BK, et al. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology (Reading). 2000;146(Pt 10):2395–407. doi: 10.1099/00221287-146-10-2395 [DOI] [PubMed] [Google Scholar]
- 128.Schaefer J, Jovanovic G, Kotta-Loizou I, Buck M. Single-step method for β-galactosidase assays in Escherichia coli using a 96-well microplate reader. Anal Biochem. 2016;503:56–7. doi: 10.1016/j.ab.2016.03.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Abramson J, Adler J, Dunger J, Evans R, Green T, Pritzel A, et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature. 2024;630(8016):493–500. doi: 10.1038/s41586-024-07487-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Pettersen EF, Goddard TD, Huang CC, Meng EC, Couch GS, Croll TI, et al. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021;30(1):70–82. doi: 10.1002/pro.3943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Camacho C, Coulouris G, Avagyan V, Ma N, Papadopoulos J, Bealer K, et al. BLAST+: architecture and applications. BMC Bioinformatics. 2009;10:421. doi: 10.1186/1471-2105-10-421 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Edgar RC. Muscle5: High-accuracy alignment ensembles enable unbiased assessments of sequence homology and phylogeny. Nat Commun. 2022;13(1):6968. doi: 10.1038/s41467-022-34630-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Potter SC, Luciani A, Eddy SR, Park Y, Lopez R, Finn RD. HMMER web server: 2018 update. Nucleic Acids Res. 2018;46(W1):W200–4. doi: 10.1093/nar/gky448 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Karpenahalli MR, Lupas AN, Söding J. TPRpred: a tool for prediction of TPR-, PPR- and SEL1-like repeats from protein sequences. BMC Bioinformatics. 2007;8:2. doi: 10.1186/1471-2105-8-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Manni M, Berkeley MR, Seppey M, Simão FA, Zdobnov EM. BUSCO update: novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol Biol Evol. 2021;38(10):4647–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30(9):1312–3. doi: 10.1093/bioinformatics/btu033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Letunic I, Bork P. Interactive Tree of Life (iTOL) v6: recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res. 2024;52(W1):W78–82. doi: 10.1093/nar/gkae268 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(XLSX)
(XLSX)
Plot of wildtype V. vulnificus (WT) and ∆rcbAB mutant (Tn) strains bearing a PcabAlacZ reporter grown in LB without or supplemented with 15 mM CaCl2 (LB + Ca2+). Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences between samples (****p < 0.0001; ns, no significant difference) were determined by unpaired student’s t-test (two-tailed).
(TIFF)
Bar plot of the motility zones for wildtype V. vulnificus and the rcbA, rcbB and rcbAB deletion mutants (∆rcbA, ∆rcbB and ∆rcbAB, respectively) in soft agar lacking (orange) or supplemented with the indicated concentration of CaCl2. Bars show the respective mean values for each strain and error bars represent the standard deviation of triplicate assays. No statistically significant difference (ns) relative to WT was found among strains at the same CaCl2 concentration (one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test).
(TIFF)
Plot of motility by wildtype V. vulnificus and the rcbAB deletion mutant (∆rcbAB) carrying the empty expression plasmid or expressing (+) rcbA, rcbB or rcbAB grown in soft agar lacking (orange) or supplemented with the indicated concentration of CaCl2. Bars show the respective mean values for each strain and error bars represent the standard deviation of triplicate assays. Statistically significant differences relative to wildtype were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test (**p < 0.01; ns, no significant difference). Expression was induced with IPTG (10 µM).
(TIFF)
Plot of intracellular c-di-GMP levels in wildtype V. vulnificus and the rcbAB deletion mutant (∆rcbAB) that carries the empty expression plasmid or express (+) rcbA, rcbB or rcbAB grown in LB lacking (orange) or supplemented with the indicated concentration of CaCl2. Bars show the respective mean values for each strain and error bars represent the standard deviation of triplicate assays. Statistically significant differences relative to wildtype were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test (***p < 0.001; ns, no significant difference). Expression was induced with IPTG (10 µM).
(TIFF)
A, plot of biofilm formation (OD595/OD600) by ∆rcbB cells harboring the empty expression plasmid (control) or plasmids expressing (+) rcbB or a catalytic mutant bearing the A-to-E mutation, GGAEF). B, a plot of swimming motility zones in IO20 soft agar by the same strains in A. Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences (****p < 0.0001; ns, no significant difference) relative to the control were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test. Expression was induced with 1 µM IPTG in A and 10 µM IPTG in B.
(TIFF)
Plot of swimming motility zones in soft agar for several E. coli strains (A) and for MG1655 cells (B) carrying the empty expression vector (Con) or the plasmid with rcbA, rcbB or rcbAB (without or with (+) 10 µM IPTG; separated by the vertical dotted line). Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences (*p < 0.05, ****p < 0.0001; ns, no significant difference) relative to the control were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test.
(TIFF)
A, western blot of ∆rcbA extracts containing HIS-tagged RcbA or ∆rcbB extracts containing HA-tagged RcbB. Expression was induced with the IPTG concentrations indicated below each lane and the proteins were detected with anti-HIS and anti-HA antibodies, respectively. B, alignments of AlphaFold 3 predicted monomer structures of RcbA (purple) with RcbA∆TPR (pink), and RcbB (green) with RcbB∆TPR (blue). C, western blot of ∆rcbA extracts containing HIS-tagged RcbA∆TPR or HA-tagged RcbB∆TPR. Expression and detection conditions were as in A.
(TIFF)
Biofilm formation (OD595/OD600) by ∆rcbAB cells harboring empty plasmids (control) or expressing (+) rcbA-mRuby3 (RcbAmRu), rcbB-mNeonGreen (RcbBmNG) or rcbA-mRuby3-rcbB-mNeonGreen (RcbAmRu/RcbBmNG). Bars indicate the respective mean values and error bars represent the standard deviation of triplicate assays. Statistically significant differences (****p < 0.0001; ns, no significant difference) relative to control cells were determined by one-way ANOVA with a Dunnett’s multiple comparisons post-hoc test. Expression was induced with 1 µM IPTG.
(TIFF)
Representative fluorescence images of NanoOrange stained ∆hubP and ∆flrA cells. White arrows denote flagella. Scale bar = 1 µm.
(TIFF)
From left to right, representative phase, fluorescence and overlay images of RcbA-mRuby3 (A), RcbB-mNeonGreen (B), or both (C) in ∆flrA∆rcbAB cells. Scale bar = 1 μm. Expression was induced with 10 µM IPTG.
(TIFF)
AlphaFold 3 predicted monomer structures of RcbB alone (blue) and complexed (green) with RcbA. A GTP molecule (stick configuration) is modeled bound at the active site. The grey arrow denotes rotation of the DGC domain relative to the aligned TPR domains.
(TIFF)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.