

Abstract
Background
The emergence of multicellularity in animals marks a pivotal evolutionary event, which was likely enabled by molecular innovations in the way cells adhere and communicate with one another. β‐Catenin is significant to this transition due to its dual role as both a structural component in the cadherin–catenin complex and as a transcriptional coactivator involved in the Wnt/β‐catenin signaling pathway. However, our knowledge of how this protein functions in ctenophores, one of the earliest diverging metazoans, is limited.
Results
To study β‐catenin function in the ctenophore Mnemiopsis leidyi, we generated affinity‐purified polyclonal antibodies targeting Mlβ‐catenin. We then used this tool to observe β‐catenin protein localization in developing Mnemiopsis embryos. In this article, we provide evidence of consistent β‐catenin protein enrichment at cell–cell interfaces in Mnemiopsis embryos. Additionally, we found β‐catenin enrichment in some nuclei, particularly restricted to the oral pole around the time of gastrulation. The Mlβ‐catenin affinity‐purified antibodies now provide us with a powerful reagent to study the ancestral functions of β‐catenin in cell adhesion and transcriptional regulation.
Conclusions
The localization pattern of embryonic Mlβ‐catenin suggests that this protein had an ancestral role in cell adhesion and may have a nuclear function as well.
Keywords: cell‐adhesion, cellfate specification, ctenophore, fate specification, mnemiopsis, Wnt signaling, β‐catenin
Key Findings
β‐catenin protein is enriched at cell‐cell interfaces in developing Mnemiopsis embryos
β‐catenin protein is enriched in some nuclei, particularly restricted to the oral pole around the time of gastrulation in developing Mnemiopsis embryo
We provide evidence that Mlβ‐catenin protein holds an ancestral role in both cell adhesion and nuclear function
Mlβ‐catenin affinity‐purified antibodies are a powerful reagent to study the ancestral functions of β‐catenin
1. INTRODUCTION
The origin of multicellularity in animals represents a major evolutionary transition, marking the divergence from single‐celled ancestors to complex organisms composed of multiple, specialized cell types. Understanding the mechanisms that evolved to enable this transition will provide insights into the fundamental principles that underpin animal evolution and the diversity of life forms we see today. Critical innovations such as cell–cell adhesion, cell–extracellular matrix interactions, cell–cell communication, and cell fate specification are hypothesized to have facilitated cellular cooperation and the partitioning of roles in the cells of the metazoan last common ancestor. 1 , 2 The β‐catenin protein is a multifunctional molecule central to many of these cellular processes and thus stands out as a key protein for understanding this transition. 3 , 4
β‐Catenin is a structurally and functionally conserved protein found to be important for developmental processes and maintaining tissue homeostasis across many metazoan lineages. It is well established that β‐catenin plays two important roles: acting as a structural component of the cadherin–catenin complex (CCC) that maintains cell–cell adhesion through adherens junctions (AJs) and as a transcriptional coactivator in the Wnt/β‐catenin signaling pathway. 4 Because of its dual purpose, β‐catenin has been identified as a key component involved in cell adhesion, cell fate specification, neurogenesis, spindle orientation, cell migration, cell polarity, and maintenance of stem cells. 4
In its role in cell–cell adhesion, β‐catenin interacts with cadherin, a calcium‐dependent transmembrane glycoprotein that links neighboring cells together by forming dimers in the extracellular space. 5 Cadherins alone are not sufficient in the establishment of a stable AJ. 6 The intracellular domain of classical cadherins includes binding sites for two catenins: p120 and β‐catenin, which are essential for stabilizing this complex at the plasma membrane (Figure 1). 7 , 8 , 9 , 10 In the absence of binding, members of the CCC are degraded through multiple mechanisms, destabilizing the AJs and leading to a loss of adhesion between cells. 11 , 12 , 13 This interaction plays a pivotal role in maintaining tissue integrity and organization in multicellular organisms. 14 Thus, β‐catenin is essential in cell–cell adhesion through the formation and maintenance of AJs by interacting with cadherins.
FIGURE 1.

The dual role of β‐catenin in cell adhesion and transcriptional activation. APC, Adenomatous Polyposis Coli; CK1α, Casein Kinase 1α; Dsh, Disheveled; GSK‐3β, Glycogen Synthase Kinase 3β; LEF, Lymphoid Enhancer Factor; LRP, Low‐density lipoprotein Receptor‐related Protein; TCF, T‐Cell Factor; β‐cat, β‐catenin; α‐cat, α‐catenin.
On the cytoplasmic side of the membrane, β‐catenin has a role in recruiting ⍺‐catenin (⍺‐cat) to the CCC, enabling interaction between the plasma membrane and the actin cytoskeleton. This interaction is necessary for cell adhesion and provides the mechanical force that maintains tissue integrity. 15 In turn, this mechanical force exerted on the CCC is at the heart of the mechano‐transduction process, which is characterized by the translation of mechanical stress into a biochemical message. This mechanically regulated cadherin/β‐catenin signaling pathway is notably involved in the specification of the anterior endoderm in D. melanogaster and in the activation of key genes in mesoderm specification by β‐catenin in zebrafish and Drosophila. 16 , 17 , 18 Furthermore, there is evidence that selective nuclearization of CCC‐associated β‐catenin can be induced through tension‐relaxation events, leading to β‐catenin‐facilitated transcription. 19 , 20
Furthermore, β‐catenin plays an important role in the Wnt/β‐catenin signaling pathway, which can be maintained in two states: “off” and “on” (Figure 1). 21 In the absence of the Wnt ligand, the pathway remains “off.” Here, a multi‐protein complex, known as the destruction complex, regulates cytoplasmic β‐catenin stability. Within this complex, Axin and Adenomatous Polyposis Coli (APC) serve as recruitment and scaffolding proteins while Casein Kinase 1 alpha (CK1α) and Glycogen Synthase Kinase‐3β (GSK‐3β) sequentially phosphorylate β‐catenin, marking it for degradation through the proteasome pathway. 22 , 23 On the other hand, when the Wnt ligand is present, the pathway shifts to the “on” state. The Wnt ligand binds to the Frizzled (Fzd) and Low‐density lipoprotein Receptor‐related Protein (LRP5/6) transmembrane co‐receptors, which then each respectively recruit Disheveled (Dsh) and Axin to the cell membrane. Receptor inhibition of GSK‐3β and Axin phosphorylation interfere with the destruction complex's enzymatic activity, allowing cytoplasmic β‐catenin to accumulate. At a high enough concentration, β‐catenin is able to translocate to the nucleus where it binds to T‐Cell Factor/Lymphoid Enhancer Factor (TCF/LEF) transcription factors and acts as a transcriptional coactivator. The ultimate outcome of β‐catenin nuclearization during the “on” state of the pathway is the transcription of downstream target gene expression. Mutations and altered expression patterns of this pathway have been linked to human diseases including cancer. 24 , 25
During the early development of many bilaterian animals, Wnt/β‐catenin signaling is involved in the establishment of the primary embryonic axis, known as the animal‐vegetal (AV) axis. 26 , 27 , 28 The AV axis is significant because it orients the embryo for future developmental processes, such as germ layer specification. 27 , 28 During normal development of some bilaterians, nuclearization of β‐catenin at the vegetal pole specifies the vegetal blastomeres to give rise to endomesoderm, a bipotential germ layer that will later segregate into endoderm and mesoderm. 28 , 29 Development can be manipulated by inhibiting the nuclearization of β‐catenin in the vegetal micromeres and macromeres in sea urchin and nemertean embryos—disrupting endomesoderm specification and causing ectopic expression of anterior neuroectoderm markers throughout the embryo—resulting in an “anteriorized” phenotype. 29 , 30 , 31 , 32 , 33 Conversely, ectopically activating the Wnt/β‐catenin pathway in the anterior mesomeres breaks the established polarity causing cells typically fated to become ectoderm to become respecified as endomesoderm, giving the embryo a “posteriorized” phenotype. 30 , 34 However, while this pattern is widely observed in bilaterians, not all metazoans specify the endomesoderm at the vegetal pole. 26 , 28 Cnidarians, the closest outgroup to the bilaterians, establish a primary oral‐aboral (OA) axis, which is patterned by Wnt/β‐catenin signaling activation at the oral pole. 35 , 36 Other studies in a more basal metazoan, the ctenophore, have shown that endomesoderm specification also takes place at the oral pole. 37 , 38 However, it is not entirely clear if this specification sets up the OA axis in these animals.
Most of our understanding of β‐catenin's functional role in metazoan biology comes from bilaterian and cnidarian models. 4 , 39 , 40 , 41 While these studies are important for establishing an understanding of these highly conserved processes for β‐catenin, investigating earlier diverging taxa will provide valuable insights into the evolutionary origins of these mechanisms. For instance, the role of β‐catenin in either a cell signaling or cell–cell adhesion context in ctenophores remains poorly characterized. Several lines of evidence now indicate that the ctenophores, or comb jellies, were the earliest emerging metazoan clade. 42 , 43 , 44 , 45 Ctenophores have a small genome (for example, the Mnemiopsis leidyi genome is roughly 150 megabases) with a reduced molecular repertoire relative to other metazoans, which simplifies the identification of conserved proteins and interactions necessary for molecular function. For example, initial in silico analysis suggests that ctenophore proteins such as Cadherin and Axin lack the domain responsible for binding to β‐catenin, 36 , 46 and there is no evidence for a Wnt/Planar Cell Polarity (PCP) signaling pathway. 47 , 48 This raises questions about what type of information will emerge from studying ancestral protein functions in ctenophores. It could either narrow down essential domains required for structural interactions or uncover alternative methods employed for accomplishing developmental and homeostatic processes. Moreover, studying key proteins in ctenophores can enhance our understanding of their evolutionary origins and shed light on how these essential biological processes evolved across Metazoa. Ctenophores, like M. leidyi, occupy a critical phylogenetic position, offering potentially unique insights into the molecular transition from unicellularity to multicellularity in the animal lineage. Therefore, developing tools to study β‐catenin and other critical molecules in ctenophores will be instrumental in uncovering these insights. Here, we report on the successful generation of affinity‐purified rabbit polyclonal antibodies targeting M. leidyi β‐catenin protein and use it to carry out localization studies to determine the subcellular distribution of the protein during early M. leidyi development. Using these antibodies, we were able to observe β‐catenin protein localize to the cell–cell interface in the earliest stages of development and in some subsets of nuclei of developing M. leidyi embryos. This reagent provides a useful tool for future studies on the potential function of β‐catenin in cell–cell adhesion and transcriptional activation in this member of an early emerging metazoan taxon.
2. RESULTS
2.1. β‐Catenin protein structure in M. leidyi
In most metazoans, the core of the β‐catenin protein is composed of 12 Armadillo repeat domains (Arm), essential for the interaction with β‐catenin binding partners for both Wnt/β‐catenin signaling and in AJs. 41 , 49 Those binding sites are overlapping, meaning that β‐catenin can either bind to cadherin or TCF/LEF, but cannot simultaneously accomplish both functions. 8 The M. leidyi genome encodes one single β‐catenin gene of 2688 bps coding for 895 amino acids. 47 Protein domain prediction softwares were only able to identify 9 out of 12 individual Arm repeats conserved in Mlβ‐cat, but also highlighted a larger Arm repeat superfamily that extended past the final recognized domain (Figure 2A). Upon further investigation into the tertiary structure, three additional complete triple alpha helices were identified with relatively strong per‐residue local confidence (Figure 2B,C). Arm domain 7 was predicted with low confidence likely due to only 2 out of the 3 alpha helices being conserved in predicted 3D structure. However, a third, short alpha helix is present immediately before the domain but truncated by a low confidence value region of disorder. Furthermore, the alignment of Mlβ‐cat with structurally characterized orthologs in other animals revealed the conservation of the 2 lysine residues (K312 and K435 in mouse epithelial cadherin) in Mlβ‐cat, crucial for the interaction between a classical cadherin and β‐catenin. 9 Moreover, the phosphorylation sites required by β‐catenin destruction complex components, part of the Wnt/β‐catenin pathway, are also conserved (Serine 112, Serine 116, and Threonine 120 for GSK3β; Serine 124 for CK1α). Additionally, 9 out of the 14 essential binding residues of the ⍺‐catenin binding motif are conserved. 46
FIGURE 2.

The predicted β‐catenin architecture in M. leidyi. (A) Mlβ‐cat protein contains 11 of the 12 Arm domains identified with high confidence flanked by an N‐Terminal and a C‐terminal region. The asterisk above domain 7 indicates predicted partial (2 out of 3 alpha helices) domain conservation. The amino acids necessary for the interaction with GSK3β and CK1α are conserved, as well as the α‐cat binding site. The two crucial lysines (K), essential for the interaction with a classical cadherin, are also conserved. (B) Predicted tertiary structure of Mlβ‐cat protein with Arm domains denoted by color. Red through pink domains were identified through domain prediction software; silver, gray, and white domains fall within the Arm repeat superfamily but were identified manually. The blue Arm domain (domain 7) was identified with low confidence likely due to the missing alpha helix. (C) pLDDT plot for predicted structure. A dip in confidence values for residues between 472 and 478 corresponds with a disordered region that truncates an alpha helix in Arm domain 7.
2.2. Affinity‐purified rabbit polyclonal antibodies generated to Mlβ‐catenin protein specifically targets endogenous Mlβ‐catenin
To confirm that the affinity‐purified rabbit polyclonal antibodies generated were specifically targeting Mlβ‐catenin, whole cell lysates from both embryo and adult M. leidyi were collected and used for Western blot analysis. Only a single band was detected around the expected size (~100 kDa) for both samples (Figure 3A). The discrepancy in size could be explained by the posttranslational modification of Mlβ‐catenin since the protein is known to be phosphorylated in other species, 23 which is known to impact the outcome of protein size on Western blots. Next, fixed whole embryos from multiple stages were immunostained using the Mlβ‐cat antibody and the nuclei were labeled with DAPI. Fixed embryos incubated with the secondary antibody alone were used as a negative control. Samples imaged using scanning confocal microscopy demonstrated robust nuclear staining by DAPI, but showed fluorescence only when embryos were incubated with the primary Mlβ‐cat antibody (Figure 3B).
FIGURE 3.

Mlβ‐cat antibody specifically recognizes endogenous protein. (A) Western blot analysis of M. leidyi cell lysates using affinity‐purified anti‐Mlβ‐cat antibodies. The specificity of the antibody was tested against whole‐cell lysates from both M. leidyi embryos and adults. A single band slightly larger than the expected size (~100 kDa) was detected for both samples. (B) Immunostaining of M. leidyi embryos using affinity‐purified anti‐Mlβ‐cat polyclonal antibodies. The top panels show nuclei stained by DAPI; the bottom panels show results of Mlβ‐cat staining with and without primary antibody (negative control). Images are maximal projection. Scale bar, 50 μM.
2.3. Mlβ‐cat localizes at cell–cell contacts
β‐Catenin is known to play a structural role by its importance in cell–cell adhesion by its association with cadherins at the plasma membrane. To determine if Mlβ‐cat was putatively involved in cell–cell adhesion, we stained M. leidyi embryos at different cell stages from fertilized egg to 8 hpf. We were not able to detect any membrane staining of Mlβ‐cat at the 1‐cell stage, however, a cortical localization of Mlβ‐cat at cell–cell contacts at the 2‐cell stage was visible. This localization continued from the 2‐cell stage through the late stages of the development in M. leidyi (Figure 4). This localization pattern was only present at cell–cell contacts, while the region of cell membranes that lacked contact displayed a diffuse level of Mlβ‐cat staining that more closely resembled the cytoplasm.
FIGURE 4.
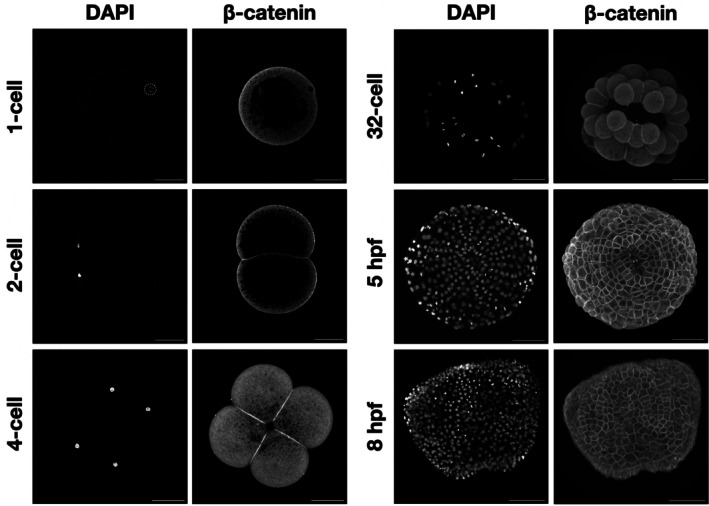
Mlβ‐catenin localizes at cell–cell contacts during M. leidyi embryonic development. Immunostaining of M. leidyi embryos at different stages using affinity‐purified β‐cat antibodies. The left column panels exhibit nuclei stained by DAPI; the right column panels exhibit results of Mlβ‐cat antibody staining. Images for 1 and 2‐cell stages are z‐stack projected images. All other images are maximal projection. Note that enriched localization of β‐catenin only occurs at cell–cell contacts. Scale bar, 50 μM.
2.4. Mlβ‐cat translocates into the nucleus during early embryogenesis
β‐Catenin function in cell signaling is associated with its accumulation into the nucleus to act as a transcriptional cofactor with TCF/LEF. We therefore looked for nuclear staining using the Mlβ‐cat antibody. We first noticed that Mlβ‐cat was uniformly expressed in the cytoplasm at 1‐cell stage, indicating that this protein is likely a maternal component in M. leidyi. However, we did not consistently detect any clear nuclear signal of β‐catenin from 1‐cell to 32‐cell (Figure 5). The first Mlβ‐cat nuclear translocation event that we observed was at the 60‐cell stage in some, but not all, 3E and 2 M\ macromeres as well as some micromeres (Figure 5). Mlβ‐cat nuclear staining continues to be non‐uniform until 4 hpf where nuclearization can be observed in cells at the oral pole, but nuclear staining was absent from the aboral pole.
FIGURE 5.
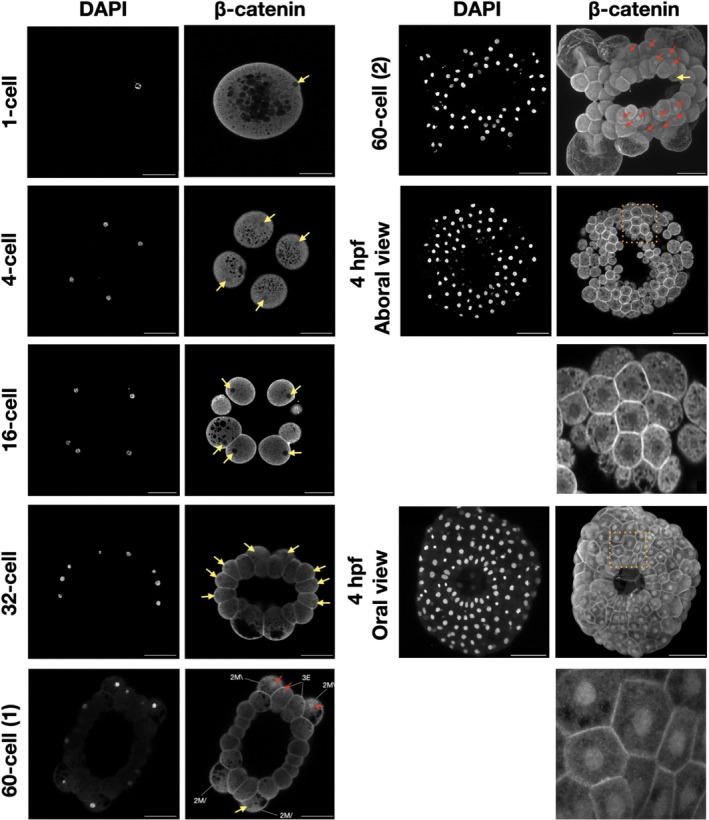
The localization of β‐catenin in M. leidyi embryos. Immunostaining of M. leidyi embryos at different stages using affinity‐purified β‐cat antibodies. The left columns show nuclei stained by DAPI, the right columns show the results of Mlβ‐cat antibody staining. Zoomed‐in insets (dotted boxes) for both the oral and aboral 4 hpf are included below the respective samples. The 1‐cell, 4‐cell, 16‐cell, 32‐cell, 60‐cell (1), and 4 hpf aboral view are z‐stack projected images. The 60‐cell (2) and 4 hpf oral views are maximal projection images. The yellow arrows show the absence of β‐catenin nuclear translocation; while the red arrows indicate β‐catenin nuclear translocation. Scale bar, 50 μM.
3. DISCUSSION
3.1. β‐Catenin robustly localizes to cell–cell interfaces in M. leidyi embryos
β‐Catenin involvement in the CCC is crucial for the adhesion between cells in every bilaterian animal where this process has been experimentally tested. 50 Nevertheless, only a few studies focused on the roles of this complex in early branching lineages. Previous studies on the cnidarian Nematostella vectensis have shown that the CCC is involved in cell–cell adhesion and germ layer formation. 40 , 51 Additionally, a functional CCC has been shown to be involved in cell–cell adhesion in sponges as supported by the co‐immunoprecipitation of interacting CCC components, β‐catenin localizing to cell–cell boundaries in Ephydatia muelleri and furthermore by Yeast 2‐Hybrid screening in another sponge, Oscarella carmela. 52 , 53
As for ctenophores, a prior study using polyclonal Mlβ‐cat antibodies generated against the first 10 amino acids of the protein was unable to capture evidence of the protein localized to the cell–cell interface in M. leidyi. 54 Thus, it was concluded that the CCC is not conserved and hence does not play a role in cell–cell adhesion in ctenophores. However, there is a possibility that the short epitope used to generate the antibodies in the Salinas‐Saavedra et al. 54 study was being blocked by β‐catenin binding partners at the cell surface, impeding the antibodies' ability to bind and hindering the ability to observe this role. Unfortunately, the peptide antibody generated in the 2019 study appears to have lost its activity and we were not able to confirm its expression in embryos or Western blots (data not shown). Our results show that Mlβ‐catenin clearly localizes at cell–cell contacts during M. leidyi embryonic development and therefore likely has a cell‐to‐cell adhesion function in the earliest metazoan lineage (Figure 5). Additionally, transcriptomic and genomic data showed that M. leidyi has a full set of CCC components and the amino acids essential for the interaction between β‐catenin and ⍺‐catenin and between β‐catenin and cadherin are mostly conserved (Figure 2). 46 , 47 The conservation of almost all 12 Arm domains on Mlβ‐catenin leads us to predict that future functional studies will demonstrate that Mlβ‐catenin maintains its ability to interact with CCC partners in ctenophores.
However, it is yet unclear if a functional CCC exists in M. leidyi since the β‐catenin binding site, essential for recruiting β‐catenin at the membrane, is either cryptic or absent from Mnemiopsis cadherin sequences. 46 , 54 An alternative hypothesis is that the CCC emerged from an interaction between β‐catenin and protocadherin instead of classical cadherin. Indeed, it has been shown that a protocadherin can interact with β‐catenin, 55 , 56 , 57 but no study has shown that this interaction is necessary for cell–cell adhesion. A third option is cadherins are entirely unnecessary for cell–cell adhesion in ctenophores. An example of this was shown in the social amoeba Dictyostelium discoideum where in response to starving this organism undergoes the formation of a fruiting body, corresponding to a “multicellular” stage. 58 Analysis of the D. discoideum genome indicates that ⍺‐catenin and β‐catenin genes are present while cadherins are absent. Knocking down the β‐cat ortholog, called Aardvark, demonstrated that the catenins were essential for the establishment of this epithelium‐like structure. 59 Clarifying the interacting partners of Mlβ‐cat at the cell membrane will provide insights into the evolutionary origin of the CCC across metazoa.
3.2. β‐Catenin is nuclearized in cells around the blastopore in M. leidyi embryos
β‐Catenin can be found in three locations inside cells: in the cytoplasm, at the cell surface on the cytoplasmic side of the cell membrane, and in the nucleus. In each of these locations, it interacts with different proteins and maintains different roles. In order to fulfill its role as a cell signaling molecule, β‐catenin must avoid being marked for degradation by the destruction complex in the cytoplasm and then translocate into the nucleus. In the vast majority of organisms where this process has been studied, this begins to occur in the early stages of development and facilitates transcription of genes involved in specifying cell fate. 27 , 28 A prior study in Mnemiopsis leidyi tracked the spatiotemporal expression of the Mlβ‐catenin protein during development and found nuclearization beginning at the zygote stage. 54 Furthermore, this study reported that there was continued nuclear expression of Mlβ‐cat in macromeres at the animal half in the endomesodermal progenitors through 11 hpf, arguing that this is substantial evidence to conclude that Mlβ‐cat has a role in specifying endomesoderm in early M. leidyi embryogenesis. When trying to replicate these results using the affinity‐purified antibodies we generated to a Mlβ‐catenin polypeptide, we were unable to find clear and consistent evidence of nuclear Mlβ‐cat in embryos prior to the 60‐cell stage. From the 60‐cell to 3 hpf, we find nuclear Mlβ‐cat to be patchy and without a clear pattern. This is unexpected due to the highly stereotyped cleavage program and precocious specification of cell fate seen in ctenophores. 38 Early stage ctenophore embryos develop at an accelerated pace making it difficult to capture specific stages for fixation and staining. This rapid cell cycle could also explain why visualizing nuclear β‐catenin in these early embryos is difficult since the nuclear envelope is being disassembled and reassembled to accommodate mitotic processes. Improving methods for fixing may make obtaining spatial‐temporal protein expression data more consistent in these early stages.
Scanning confocal analysis of Mlβ‐cat antibody stained Mnemiopsis embryos at around 4 hpf started to show a clear pattern of nuclear staining restricted blastomeres at the oral pole, while cells at the aboral pole did not show any nuclear staining. This is evidence that Mlβ‐cat may maintain its role as a co‐activator of transcription, but does not align with the previously described expression pattern. The fact that this pattern is restricted to one pole does indicate the possibility for differential gene expression or stabilization, although patchy nuclear translocation during early development and a restriction to the ectodermal cells at the oral pole leaves us with questions about this protein's functional role in cell type specification. Future functional experiments are required to determine if β‐catenin maintains its role as a transcriptional co‐activator in the Wnt/β‐catenin pathway in ctenophores.
Furthermore, the transportation of β‐catenin into the nucleus is not well understood in all metazoans. There are several proposed mechanisms for transport including direct nuclear pore complex interaction, various piggyback or chaperone candidates, and post‐translational modification. 60 With their ancestral molecular repertoire and favorable optical properties, ctenophore embryos might be a powerful model to investigate the mechanisms behind β‐catenin nuclearization.
3.3. Developing tools to study ctenophores
The combination of Western blot analysis and immunohistochemistry results gives us high confidence that the newly generated affinity‐purified Mlβ‐cat antibody specifically targets β‐catenin without any detectable non‐specific interactions. As such, this antibody can now be used as a tool to ask follow‐up questions about the functional roles of β‐catenin in the earliest metazoan branch. The evolution and development community has come to recognize that ctenophores sit at a crucial point for understanding the early evolutionary history of metazoans and has been working to develop experimental methods and resources like assembled reference genomes, transcriptomes, morpholino oligonucleotides, and CRISPR‐Cas9. 61 , 62 , 63 , 64 , 65 Hopefully, with this expanding toolkit, we can begin to answer questions about transition from unicellularity to multicellularity in the animal lineage as well as better understand the molecular components the earliest metazoans maintain that are necessary for cell–cell adhesion and cell fate specification. As we develop new tools and techniques to study ctenophores, we will not only better understand how these animals operate on a molecular level but also gain insights into how conserved mechanisms have evolved over time.
4. EXPERIMENTAL PROCEDURES
4.1. Structural analysis of Mlβ‐catenin protein domains
The protein domains of Mlβ‐catenin (Mlβ‐cat, ML073715a) have been predicted and checked with Interpro 100.0 (https://www.ebi.ac.uk/interpro/) 66 and SMART 9.0 (http://smart.embl-heidelberg.de/). 67 The tertiary structure was predicted using AlphaFold v2.3.1 on the COSMIC 2 platform (https://cosmic2.sdsc.edu/). 68
4.2. Maintenance and spawning
The M. leidyi adults were caught in a marina at Flagler Beach (Florida, United States, 29°30′60.0″ N 81°08′47.1″ W) and were maintained in 40 L kreisels with constant seawater flow, temperature, and salinity at the Whitney Laboratory for Marine Bioscience of the University of Florida (Saint Augustine, Florida, USA). Care and maintenance of adults including spawning, fertilization, and embryo culturing have been carried out as previously described, 69 however, the salinity the adult animals were kept in was adjusted to ⅔ strength filtered sea water (FSW, 25 parts per thousand) to improve spawning efficiency. Spawning was induced by incubating the adults for 3 h in the dark at RT and then moving them to glass bowls approximately 1 h before spawning to collect embryos. Embryos were kept in glass dishes in ⅔ FSW until the desired stage.
4.3. Molecular cloning of Mlβ‐cat
The total RNA was extracted from mixed embryonic stages using Trizol (Sigma, cat.# T9424). A partial length Mlβ‐cat coding sequence was amplified from cDNA using the Advantage RT for PCR kit (Clontech, cat #639506) using the following primers:
Mlβ‐cat pGEX forward: ATAGGATCCATGGAAACGCCAGTATAT.
Mlβ‐cat pGEX reverse: TATAAGAATTCTCAGGTCGTTGTGACGGG.
The amplified cDNA was then ligated into a pGEX‐6P‐1 vector for cloning and inducing a GST‐tagged Mlβ‐cat protein. The first 200 amino acids of Mlβ‐cat were selected to be used as the antigen for the polyclonal antibodies due to their predicted high immunogenicity. BLAST search against the M. leidyi Protein Model 2.2 (Mnemiopsis Genome Project Portal, https://research.nhgri.nih.gov/mnemiopsis/) using the antigen sequence only recovered the β‐catenin sequence at a significant E‐value.
4.4. Antigen preparation and antibody generation
BL21(DE3) competent E. coli (New England Biolabs cat.# C2530H) were transformed with the Mlβ‐cat pGEX‐6P‐1 construct. Five milliliters of inoculated terrific broth (TB) medium was incubated overnight (12–18 h) in a shaking incubator at 37°C at 200 rpm. Overnight cultures were used to inoculate 500 mLs of TB medium, which were then incubated until they reached an OD600 between 0.2 and 0.3, at which point the bacteria were induced by adding 0.5 mM isopropyl β‐d‐1‐thiogalactopyranoside (IPTG). Cultures were allowed to incubate for an additional 3 h before bacteria were collected by centrifuging at 5000 g for 15 min at 4°C. The induced GST‐tagged Mlβ‐cat protein was affinity purified on a glutathione sepharose beads matrix as described. 70 Approximately, 1.5 mg of Mlβ‐cat protein was collected via PreScission Protease (GenScript cat.# Z02799) digest and 6 mg of Mlβ‐cat‐GST protein collected via 10 mM glutathione whole protein elution was sent to Labcorp Early Development Laboratories Inc. (Denver, PA). There, the Mlβ‐cat N‐terminal polypeptide was used as the antigen for single rabbit polyclonal antibody generation and the Mlβ‐cat‐GST protein was used for affinity purification of the antibody from final bleed serum.
4.5. Western blot analysis
M. leidyi embryo protein samples were collected by centrifuging mixed‐stage embryos (between 8 and 12 h post fertilization, hpf) in an Eppendorf tube to pellet, removing as much sea water as possible, and adding minimum volume (between 30 and 50 μL) of 1X SDS gel‐loading buffer (50 mM Tris‐Cl pH 6.8, 100 mM dithiothreitol, 2% sodium dodecyl sulfate, 0.1% bromophenol blue, and 10% glycerol) to fully lyse the pellet. M. leidyi adult protein samples were collected by dissecting the lobes off, using a homogenizer to dissociate cells, centrifuging to pellet, removing as much sea water as possible, and adding a minimum volume (between 100 and 500 μL) of 1X SDS gel‐loading buffer to fully lyse the sample. Lysates were boiled for 5 min and 30 μL was then separated on 10% SDS‐PAGE gels, transferred onto 0.45 μm nitrocellulose membranes (Bio‐Rad Laboratories, Inc. cat. # 1620115) and blocked overnight in a 5% milk powder 1X tris buffered saline solution. The immunoblot was probed with the rabbit anti‐Mlβ‐cat (1:500). Blots were developed using an IRDye 800 (1:10,000) donkey anti‐rabbit secondary antibody (LI‐COR, Inc. cat.# 926–32,213) and imaged using a LI‐COR Odyssey Infrared Imaging System.
4.6. Fixing and immunofluorescence analysis of embryos
The vitelline membranes of M. leidyi embryos were mechanically removed using forceps before fixation. Embryos were kept in plastic dishes treated with 3% Bovine Serum Albumin (BSA) in ⅔ FSW to prevent them from sticking to the dish. Embryos of different developmental stages were fixed (4% paraformaldehyde, 0.2% glutaraldehyde in 1X phosphate buffered saline) for 5 min at RT, before being fixed in a second fixative solution (4% PFA in 1X phosphate buffered saline) for 10 min at RT. Embryos were washed three times with 1X PBS and then permeabilized in 0.2% Triton in PBS. Embryos were rinsed several times in 1X PBT (0.1% BSA, 0.2% Triton in 1X phosphate buffered saline) before the blocking step in 5% Normal Goat Serum (NGS in 1X PBT) for 1 h at RT on a rocker. The blocking solution was then replaced by the primary antibody (anti‐Mlβ‐cat; 1:500 in blocking reagent) and incubated overnight at 4° with gentle rocking. Fixed embryos were washed several times with 1X PBT. A goat anti‐rabbit IgG AlexaFluor 647 (Invitrogen, cat.# A‐21245) secondary antibody was diluted in 5% NGS (1:250) and embryos were incubated in the dark at 4° overnight on a rocker. Negative controls were incubated with the secondary antibody only. Samples were washed with 1X PBT for a total period of 2 h. Embryos were finally stained with DAPI (1:1000 in PBT; Invitrogen, Inc. Cat. #D1306) to allow nuclear visualization. Stained samples were rinsed two times with 1X phosphate‐buffered saline before mounting in the same solution. The embryos were imaged under a Zeiss 710 scanning confocal microscope. All the images were processed through ImageJ (z‐stack and maximal intensity projection). More than 30 embryos were examined at each embryonic stage.
ACKNOWLEDGMENTS
This work was partially supported by NASA (80NSSC18K1067) to AHW, and MQM and NSF (1755364) to MQM.
Walters BM, Guttieres LJ, Goëb M, Marjenberg SJ, Martindale MQ, Wikramanayake AH. β‐Catenin localization in the ctenophore Mnemiopsis leidyi suggests an ancestral role in cell adhesion and nuclear function. Developmental Dynamics. 2025;254(9):1055‐1067. doi: 10.1002/dvdy.70004
Brian M. Walters and Lucas J. Guttieres contributed equally to this study.
REFERENCES
- 1. Richter DJ, King N. The genomic and cellular foundations of animal origins. Annu Rev Genet. 2013;47:509‐537. doi: 10.1146/annurev-genet-111212-133456 [DOI] [PubMed] [Google Scholar]
- 2. Ros‐Rocher N, Pérez‐Posada A, Leger MM, Ruiz‐Trillo I. The origin of animals: an ancestral reconstruction of the unicellular‐to‐multicellular transition. Open Biol. 2021;11(2):200359. doi: 10.1098/rsob.200359 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Shapiro L. The multi‐talented β‐catenin makes its first appearance. Structure. 1997;5(10):1265‐1268. doi: 10.1016/S0969-2126(97)00278-5 [DOI] [PubMed] [Google Scholar]
- 4. Valenta T, Hausmann G, Basler K. The many faces and functions of Î 2‐catenin. EMBO J. 2012;31(12):2714‐2736. doi: 10.1038/emboj.2012.150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Nollet F, Kools P, Van Roy F. Phylogenetic analysis of the cadherin superfamily allows identification of six major subfamilies besides several solitary members. J Mol Biol. 2000;299(3):551‐572. doi: 10.1006/jmbi.2000.3777 [DOI] [PubMed] [Google Scholar]
- 6. Ishiyama N, Ikura M. The three‐dimensional structure of the cadherin–catenin complex. Adherens Junctions: from Molecular Mechanisms to Tissue Development and Disease. Springer; 2012:39‐62. doi: 10.1007/978-94-007-4186-7_3 [DOI] [PubMed] [Google Scholar]
- 7. Peifer M, McCrea PD, Green KJ, Wieschaus E, Gumbiner BM. The vertebrate adhesive junction proteins β‐catenin and plakoglobin and the drosophila segment polarity gene armadillo form a multigene family with similar properties mark. J Cell Biol. 1992;181(3):681‐691. doi: 10.1083/jcb.118.3.681 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Orsulic S, Huber O, Aberle H, Arnold S, Kemler R. E‐cadherin binding prevents β‐catenin nuclear localization and β‐catenin/LEF‐1‐mediated transactivation. J Cell Sci. 1999;112(8):1237‐1245. doi: 10.1242/jcs.112.8.1237 [DOI] [PubMed] [Google Scholar]
- 9. Huber AH, Weis WI. The structure of the β‐catenin/E‐cadherin complex and the molecular basis of diverse ligand recognition by β‐catenin. Cell. 2001;105(3):391‐402. doi: 10.1016/S0092-8674(01)00330-0 [DOI] [PubMed] [Google Scholar]
- 10. Davis MA, Ireton RC, Reynolds AB. A core function for p120‐catenin in cadherin turnover. J Cell Biol. 2003;163(3):525‐534. doi: 10.1083/jcb.200307111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Hinck L, Näthke IS, Papkoff J, Nelson WJ. Dynamics of cadherin/catenin complex formation: novel protein interactions and pathways of complex assembly. J Cell Biol. 1994;125(6):1327‐1340. doi: 10.1083/jcb.125.6.1327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Wu WJ, Hirsch DS. Mechanism of E‐cadherin lysosomal degradation. Nat Rev Cancer. 2009;9(2):143. doi: 10.1038/nrc2521-c1 [DOI] [PubMed] [Google Scholar]
- 13. Kourtidis A, Ngok SP, Anastasiadis PZ. P120 catenin: an essential regulator of cadherin stability, adhesion‐induced signaling, and cancer progression. Prog Mol Biol Transl Sci. 2013;116:409‐432. doi: 10.1016/B978-0-12-394311-8.00018-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Meng W, Takeichi M. Adherens junction: molecular architecture and regulation. Cold Spring Harb Perspect Biol. 2009;1:1‐13. doi: 10.1098/rstb.1989.0070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Lecuit T, Yap AS. E‐cadherin junctions as active mechanical integrators in tissue dynamics. Nat Cell Biol. 2015;17(5):533‐539. doi: 10.1038/ncb3136 [DOI] [PubMed] [Google Scholar]
- 16. Farge E. Mechanical induction of twist in the drosophila foregut/Stomodeal primordium. Curr Biol. 2003;13:1365‐1377. doi: 10.1016/S0960-9822(03)00576-1 [DOI] [PubMed] [Google Scholar]
- 17. Desprat N, Supatto W, Pouille PA, Beaurepaire E, Farge E. Tissue deformation modulates twist expression to determine anterior midgut differentiation in drosophila embryos. Dev Cell. 2008;15(3):470‐477. doi: 10.1016/j.devcel.2008.07.009 [DOI] [PubMed] [Google Scholar]
- 18. Brunet T, Bouclet A, Ahmadi P, et al. Evolutionary conservation of early mesoderm specification by mechanotransduction in Bilateria. Nat Commun. 2013;4(2821):1‐15. doi: 10.1038/ncomms3821 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Röper J‐C, Mitrossilis D, Stirnemann G, et al. The major β‐catenin/E‐cadherin junctional binding site is a primary molecular mechano‐transductor of differentiation in vivo. eLife. 2018;32(3):152‐160. doi: 10.1016/S1553-7250(06)32020-X [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Gayrard C, Bernaudin C, Déjardin T, Seiler C, Borghi N. Src‐ and confinement‐dependent FAK activation causes E‐cadherin relaxation and β‐catenin activity. J Cell Biol. 2018;217(3):1063‐1077. doi: 10.1083/jcb.201706013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Nusse R, Clevers H. Wnt/β‐catenin signaling, disease, and emerging therapeutic modalities. Cell. 2017;169(6):985‐999. doi: 10.1016/j.cell.2017.05.016 [DOI] [PubMed] [Google Scholar]
- 22. Aberle H, Bauer A, Stappert J, Kispert A, Kemler R. Β‐catenin is a target for the ubiquitin‐proteasome pathway. EMBO J. 1997;16(13):3797‐3804. doi: 10.1093/emboj/16.13.3797 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Stamos JL, Weis WI. The β‐catenin destruction complex. Cold Spring Harb Perspect Biol. 2013;5(1):1‐16. doi: 10.1101/cshperspect.a007898 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Clevers H. Wnt/β‐catenin signaling in development and disease. Cell. 2006;127(3):469‐480. doi: 10.1016/j.cell.2006.10.018 [DOI] [PubMed] [Google Scholar]
- 25. Klaus A, Birchmeier W. Wnt signalling and its impact on development and cancer. Nat Rev Cancer. 2008;8(5):387‐398. doi: 10.1038/nrc2389 [DOI] [PubMed] [Google Scholar]
- 26. Goldstein B, Freeman G. Axis specification in animal development. Bioessays. 1997;19(2):105‐116. doi: 10.1002/bies.950190205 [DOI] [PubMed] [Google Scholar]
- 27. Petersen CP, Reddien PW. Wnt signaling and the polarity of the primary body Axis. Cell. 2009;139(6):1056‐1068. doi: 10.1016/j.cell.2009.11.035 [DOI] [PubMed] [Google Scholar]
- 28. Martindale MQ. The evolution of metazoan axial properties. Nat Rev Genet. 2005;6(12):917‐927. doi: 10.1038/nrg1725 [DOI] [PubMed] [Google Scholar]
- 29. Loh KM, van Amerongen R, Nusse R. Generating cellular diversity and spatial form: Wnt signaling and the evolution of multicellular animals. Dev Cell. 2016;38(6):643‐655. doi: 10.1016/j.devcel.2016.08.011 [DOI] [PubMed] [Google Scholar]
- 30. Wikramanayake AH, Huang L, Klein WH. β‐Catenin is essential for patterning the maternally specified animal‐vegetal axis in the sea urchin embryo. Proc Natl Acad Sci U S A. 1998;95(16):9343‐9348. doi: 10.1073/pnas.95.16.9343 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Logan CY, Miller JR, Ferkowicz MJ, McClay DR. Nuclear β‐catenin is required to specify vegetal cell fates in the sea urchin embryo. Development. 1999;126(2):345‐357. doi: 10.1242/dev.126.2.345 [DOI] [PubMed] [Google Scholar]
- 32. Range RC, Angerer RC, Angerer LM. Integration of canonical and noncanonical Wnt signaling pathways patterns the neuroectoderm along the anterior–posterior Axis of sea urchin embryos. PLoS Biol. 2013;11(1):e1001467. doi: 10.1371/journal.pbio.1001467 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Henry JQ, Perry KJ, Wever J, Seaver E, Martindale MQ. β‐Catenin is required for the establishment of vegetal embryonic fates in the nemertean, Cerebratulus Lacteus. Dev Biol. 2008;317(1):368‐379. doi: 10.1016/j.ydbio.2008.02.042 [DOI] [PubMed] [Google Scholar]
- 34. Sun H, Peng CFJ, Wang L, Feng H, Wikramanayake AH. An early global role for Axin is required for correct patterning of the anterior–posterior axis in the sea urchin embryo. Dev. 2021;148(7):1‐13. doi: 10.1242/dev.191197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Wikramanayake AH, Hong M, Lee PN, et al. An ancient role for nuclear β‐catenin in the evolution of axial polarity and germ layer segregation. Nature. 2003;426(6965):446‐450. doi: 10.1038/nature02113 [DOI] [PubMed] [Google Scholar]
- 36. Sun H, Zidek R, Martindale MQ, Wikramanayake AH. The Nematostella Wnt/β‐catenin destruction complex provides insight into the evolution of the regulation of a major metazoan signal transduction pathway. bioRxiv. 2024;1‐57. doi: 10.1101/2024.10.27.620500 [DOI] [Google Scholar]
- 37. Freeman G. The establishment of the oral‐aboral axis in the ctenophore embryo. J Embryol Exp Morphol. 1977;42:237‐260. doi: 10.1242/dev.42.1.237 [DOI] [Google Scholar]
- 38. Martindale MQ, Henry JQ. Intracellular fate mapping in a basal metazoan, the ctenophore Mnemiopsis leidyi, reveals the origins of mesoderm and the existence of indeterminate cell lineages. Dev Biol. 1999;214(2):243‐257. doi: 10.1006/dbio.1999.9427 [DOI] [PubMed] [Google Scholar]
- 39. Martindale MQ, Hejnol A. A developmental perspective: changes in the position of the blastopore during Bilaterian evolution. Dev Cell. 2009;17(2):162‐174. doi: 10.1016/j.devcel.2009.07.024 [DOI] [PubMed] [Google Scholar]
- 40. Clarke DN, Lowe CJ, Nelson WJ. The cadherin‐catenin complex is necessary for cell adhesion and embryogenesis in Nematostella vectensis. Dev Biol. 2019;447(2):170‐181. doi: 10.1016/j.ydbio.2019.01.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. van der Wal T, van Amerongen R. Walking the tight wire between cell adhesion and WNT signalling: a balancing act for β‐catenin. Open Biol. 2020;10(12):267. doi: 10.1098/rsob.200267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Dunn CW, Hejnol A, Matus DQ, et al. Broad phylogenomic sampling improves resolution of the animal tree of life. Nature. 2008;452(7188):745‐749. doi: 10.1038/nature06614 [DOI] [PubMed] [Google Scholar]
- 43. Hejnol A, Obst M, Stamatakis A, et al. Assessing the root of bilaterian animals with scalable phylogenomic methods. Proc R Soc B Biol Sci. 2009;276(1677):4261‐4270. doi: 10.1098/rspb.2009.0896 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Whelan NV, Kocot KM, Moroz TP, et al. Ctenophore relationships and their placement as the sister group to all other animals. Nat Ecol Evol. 2017;1(11):1737‐1746. doi: 10.1038/s41559-017-0331-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Schultz DT, Haddock SHD, Bredeson JV, Green RE, Simakov O, Rokhsar DS. Ancient gene linkages support ctenophores as sister to other animals. Nature. 2023;618(7963):110‐117. doi: 10.1038/s41586-023-05936-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Belahbib H, Renard E, Santini S, et al. New genomic data and analyses challenge the traditional vision of animal epithelium evolution. BMC Genomics. 2018;19(1):1‐15. doi: 10.1186/s12864-018-4715-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Ryan JF, Pang K, Schnitzler CE, et al. The genome of the ctenophore Mnemiopsis leidyi and its implications for cell type evolution. Science. 2013;342(6164):1242592. doi: 10.1126/science.1242592 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Moroz LL, Kocot KM, Citarella MR, et al. The ctenophore genome and the evolutionary origins of neural systems. Nature. 2014;510(7503):109‐114. doi: 10.1038/nature13400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Schneider SQ, Finnerty JR, Martindale MQ. Protein evolution: structure–function relationships of the oncogene Beta‐catenin in the evolution of multicellular animals. J Exp Zool B Mol Dev Evol. 2003;295(1):25‐44. doi: 10.1002/jez.b.6 [DOI] [PubMed] [Google Scholar]
- 50. Halbleib JM, Nelson WJ. Cadherins in development: cell adhesion, sorting, and tissue morphogenesis. Genes Dev. 2006;20(23):3199‐3214. doi: 10.1101/gad.1486806 [DOI] [PubMed] [Google Scholar]
- 51. Pukhlyakova EA, Kirillova AO, Kraus YA, Zimmermann B, Technau U. A cadherin switch marks germ layer formation in the diploblastic sea anemone Nematostella vectensis. Dev. 2019;146(20):1‐15. doi: 10.1242/dev.174623 [DOI] [PubMed] [Google Scholar]
- 52. Schippers KJ, Nichols SA. Evidence of signaling and adhesion roles for β‐catenin in the sponge ephydatia muelleri. Mol Biol Evol. 2018;35(6):1407‐1421. doi: 10.1093/molbev/msy033 [DOI] [PubMed] [Google Scholar]
- 53. Nichols SA, Roberts BW, Richter DJ, Fairclough SR, King N. Origin of metazoan cadherin diversity and the antiquity of the classical cadherin/β‐catenin complex. Proc Natl Acad Sci U S A. 2012;109(32):13046‐13051. doi: 10.1073/pnas.1120685109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Salinas‐Saavedra M, Wikramanayake A, Martindale M. Β‐catenin has an ancestral role in cell fate specification but not cell adhesion. bioRxiv. 2019;520957. doi: 10.1101/520957 [DOI] [Google Scholar]
- 55. Chen MW, Vacherot F, De la Taille A, et al. The emergence of protocadherin‐PC expression during the acquisition of apoptosis‐resistance by prostate cancer cells. Oncogene. 2002;21(51):7861‐7871. doi: 10.1038/sj.onc.1205991 [DOI] [PubMed] [Google Scholar]
- 56. Parenti S, Ferrarini F, Zini R, et al. Mesalazine inhibits the β‐catenin signalling pathway acting through the upregulation of μ‐protocadherin gene in colo‐rectal cancer cells. Aliment Pharmacol Ther. 2010;31(1):108‐119. doi: 10.1111/j.1365-2036.2009.04149.x [DOI] [PubMed] [Google Scholar]
- 57. de Nys R, Gardner A, van Eyk C, et al. Proteomic analysis of the developing mammalian brain links PCDH19 to the Wnt/β‐catenin signalling pathway. Mol Psychiatry. 2024;29(7):1‐12. doi: 10.1038/s41380-024-02482-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Urushihara H. Developmental biology of the social amoeba: history, current knowledge and prospects. Dev Growth Differ. 2008;50:277‐281. doi: 10.1111/j.1440-169X.2008.01013.x [DOI] [PubMed] [Google Scholar]
- 59. Dickinson DJ, Nelson WJ, Weis WI. A polarized epithelium organized by β‐ and α‐catenin predates cadherin and metazoan origins. Science. 2011;331(6022):1336‐1339. doi: 10.1126/science.1199633 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Anthony CC, Robbins DJ, Ahmed Y, Lee E. Nuclear regulation of Wnt/β‐catenin signaling: It's a complex situation. Genes Basel. 2020;11(8):1‐11. doi: 10.3390/genes11080886 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Moreland RT, Nguyen AD, Ryan JF, et al. A customized web portal for the genome of the ctenophore Mnemiopsis leidyi . BMC Genomics. 2014;15(1):1‐13. doi: 10.1186/1471-2164-15-316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Davidson PL, Koch BJ, Schnitzler CE, et al. The maternal‐zygotic transition and zygotic activation of the Mnemiopsis leidyi genome occurs within the first three cleavage cycles. Mol Reprod Dev. 2017;84(11):1218‐1229. doi: 10.1002/mrd.22926 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Yamada A, Martindale MQ, Fukui A, Tochinai S. Highly conserved functions of the Brachyury gene on morphogenetic movements: insight from the early‐diverging phylum Ctenophora. Dev Biol. 2010;339(1):212‐222. doi: 10.1016/j.ydbio.2009.12.019 [DOI] [PubMed] [Google Scholar]
- 64. Presnell JS, Browne WE. Krüppel‐like factor gene function in the ctenophore Mnemiopsis leidyi assessed by CRISPR/Cas9‐mediated genome editing. Development. 2021;148(17):199771. doi: 10.1242/DEV.199771 [DOI] [PubMed] [Google Scholar]
- 65. Presnell JS, Bubel M, Knowles T, Patry W, Browne WE. Multigenerational laboratory culture of pelagic ctenophores and CRISPR–Cas9 genome editing in the lobate Mnemiopsis leidyi . Nat Protoc. 2022;17(8):1868‐1900. doi: 10.1038/s41596-022-00702-w [DOI] [PubMed] [Google Scholar]
- 66. Paysan‐Lafosse T, Blum M, Chuguransky S, et al. InterPro in 2022. Nucleic Acids Res. 2023;51(D1):D418‐D427. doi: 10.1093/nar/gkac993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Schultz J, Milpetz F, Bork P, Ponting CP. SMART, a simple modular architecture research tool: identification of signaling domains. Proc Natl Acad Sci U S A. 1998;95(11):5857‐5864. doi: 10.1073/pnas.95.11.5857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Cianfrocco MA, Wong‐Barnum M, Youn C, Wagner R, Leschziner A. COSMIC2: a science gateway for cryo‐electron microscopy structure determination. ACM Int Conf Proceeding Ser. 2017;F1287:13‐17. doi: 10.1145/3093338.3093390 [DOI] [Google Scholar]
- 69. Salinas‐Saavedra M, Martindale MQ. Improved protocol for spawning and immunostaining embryos and juvenile stages of the ctenophore Mnemiopsis leidyi . Protoc Exch. 2018;1‐10. doi: 10.1038/protex.2018.092 [DOI] [Google Scholar]
- 70. Kielkopf CL, Bauer W, Urbatsch IL. Purification of fusion proteins by affinity chromatography on glutathione resin. Cold Spring Harb Protoc. 2020;2020(6):217‐224. doi: 10.1101/pdb.prot102202 [DOI] [PubMed] [Google Scholar]