

SUMMARY
Plastoglobuli (PG) are plant lipoprotein compartments, present in plastid organelles. They are involved in the formation and/or storage of lipophilic metabolites. FIBRILLINs (FBNs) are one of the main PG‐associated proteins and are particularly abundant in carotenoid‐enriched chromoplasts found in ripe fruits and flowers. To address the contribution of different FBNs, independently and in combination, to isoprenoid formation and sequestration, a multiplex gene editing approach was undertaken in tomato. This approach generated a suite of single and high‐order fbn mutants that were shown to lack transcripts and respective protein products. The major PG‐related FBNs in tomato chosen for this study are SlFBN1, SlFBN2a, SlFBN4 and SlFBN7a. When knocked out independently, functional redundancy was revealed. However, paralog‐specific roles were detected regulating specific isoprenoids (e.g. plastochromanol 8) or plastidial esterification capability. In addition, high‐order fbn mutants displayed altered isoprenoid chromoplast sequestration patterns, notably with a significant reduction in carotenes (phytoene and phytofluene) in the PG fraction. Proteomic analysis confirmed the absence of PG‐core associated proteins, including NAD(P)H‐ubiquinone oxidoreductase C1, tocopherol cyclase (VTE1) and phytol esterase (PES1/PYP). Perturbations to the ultrastructure of the plastid were revealed, with aberrant PG formation and morphology predominating in high‐order mutants. Global lipidome profiles also highlighted broader changes directly affecting storage and plastid membrane lipids, for example, tri‐ and diacylglycerides and galactolipid species. Collectively, these results support both structural and metabolic roles of SlFBNs in PGs. The findings expose fundamental aspects of metabolic compartmentalisation in plant cells and the importance of lipoprotein particles for plastid integrity and functionality.
Keywords: plastoglobuli, carotenoids, isoprenoids, chromoplast, esterification, lipid homeostasis, fibrillin, multiplexing gene editing
Significance Statement
A multiplex CRISPR/Cas approach has been used to characterise members of the FIBRILLIN (FBN) multigene family, through the generation and characterisation of independent and high‐order mutants. Molecular and biochemical characterisation has been performed, showing functional redundancy between members and the roles of FBN in plastoglobuli (PG) biogenesis and lipid metabolism.

A multiplex gene editing methodology has been used to generate independent and higher order mutants for the FBN gene family. Characterisation of these valuable resources has revealed an involvement of FBNs in PG biogenesis, lipid remodelling and isoprenoid formation. This genetic resource will now aid the elucidation of the role of FBNs and PG in carotenoid‐enriched tissues, with potential applications in improving crops for nutritional quality and resilience to abiotic stresses associated with climate change.
INTRODUCTION
The role of lipid droplets and lipoproteins in maintaining lipid homeostasis has been studied extensively in animal systems due to their involvement in cardiovascular diseases and obesity (Goldstein & Brown, 2009). Similar lipoprotein structures are also present in the plastids of plants. However, their function remains comparatively poorly understood (Lundquist et al., 2020). Plant plastids, including photosynthetically active chloroplasts and other types, contain dynamic lipid storage sub‐compartments termed plastoglobuli (PGs). PG number, size and composition vary depending on plastid differentiation and type, being linked to plant developmental transitions. PG characteristics can also change as a result of stress responses (Lichtenthaler, 2013; van Wijk & Kessler, 2017). Recent evidence has supported that the PG is not only a sink for lipids but also an active metabolic hub containing key metabolites of biotechnological importance (Rottet et al., 2015; Spicher & Kessler, 2015).
Structurally, PGs are surrounded by a membrane lipid monolayer, encapsulating a neutral lipid core. Triacylglycerides (TAGs) can predominate in the core, depending on plastid type, but also present are isoprenoids (carotenoids, tocopherols and other prenylquinones) (Bréhélin et al., 2007). A specific set of proteins coat the PG surface, which also greatly vary depending on cell type (van Wijk & Kessler, 2017). A major structural protein class found in PGs is FIBRILLINs (FBNs). These plastid‐specific proteins are encoded by a multigene family whose members contain a typical plastid lipid‐associated protein (PAP) domain. They do, however, lack integral transmembrane domains but possess amphipathic helices predicted to bind to the PG lipid monolayer (Shanmugabalaji et al., 2013; Singh & McNellis, 2011). FBNs also contain a conserved lipocalin‐like motif which has been suggested to participate in the binding and transport of small hydrophobic molecules (Kim et al., 2017; Singh & McNellis, 2011).
In the model flowering plant Arabidopsis, 14 FBNs have been identified; seven of which are considered PG‐associated proteins, while the others are located primarily in thylakoid membranes or stroma of chloroplasts (Lundquist et al., 2012; van Wijk & Kessler, 2017). Functionally, FBNs are involved in different plant processes ranging from photosynthesis to colour acquisition during organ development and response to stress conditions (Langenkämper et al., 2001; Leitner‐Dagan et al., 2006; Youssef et al., 2010). Moreover, FBNs can fulfil key structural roles associated with metabolic pathways (Kim et al., 2015, 2017).
FBNs are best known for their role in flower and fruit chromoplasts, where they are involved in metabolism for the synthesis and storage of carotenoids and isoprenoid‐derived compounds with high nutritional and industrial value (Nogueira et al., 2018; Sun et al., 2020). FBN1/PAP and its homologues in different species are required for the formation of carotenoid‐sequestering structures occurring during fruit ripening such as PGs and fibrils (Deruere et al., 1994; Fang et al., 2020; Kilambi et al., 2013; Leitner‐Dagan et al., 2006; Suzuki et al., 2015). Enhanced‐carotenoid plant genotypes produced by biotechnological interventions typically exhibit altered chromoplast structure with increased PG number, highlighting the metabolic composition influence on plastid morphology (Berry et al., 2019; Enfissi et al., 2010, 2019; Nogueira et al., 2013). Despite multiple studies implicating FBNs with PG formation and dynamics, the underlying mechanisms of how FBNs fulfil this role remain poorly understood. It has been proposed that PG‐targeted FBNs may be involved in globule size, preventing PG coalescence and favouring their clustering (Rey et al., 2000). Given the multigene feature of FBNs, questions remain as to which FBN members act on PG formation, and/or which affect PG core lipid remodelling, leading to changes in plant metabolism. Indeed, individual FBN contributions are challenging to interpret because several members occur simultaneously in plant cells.
In the present study, the independent and complementary roles of the major PG‐related FBNs have been investigated by adopting a multiplexing gene editing approach in tomato. Metabolomic, proteomic, and cellular ultrastructural analysis has provided a deeper insight into the functional diversity of the fbn gene family and how they can be exploited in future biotechnological applications.
RESULTS
Putative plastoglobular FBNs associated with tomato chromoplast development
Using the Arabidopsis thaliana FBN (AtFBN) protein sequences (van Wijk & Kessler, 2017), 15 tomato fbn (Slfbn) genes were identified in total covering all AtFBN groups. For comparison, homologous FBN sequences from pepper (Capsicum annuum, CaFBNs), another Solanaceae species with fruits highly specialised for carotenoid storage, were also retrieved. Slfbn orthologs of Atfbns encoding PG‐targeted proteins (Lundquist et al., 2012), namely Slfbn1/chrc, ‐2a, ‐2b, ‐4, ‐7a, ‐7b, ‐8 were determined by phylogenetic analysis. Notably, paralogous pairs primarily clustered within each plant family, supporting lineage‐specific expansions (Figure 1a).
Figure 1.
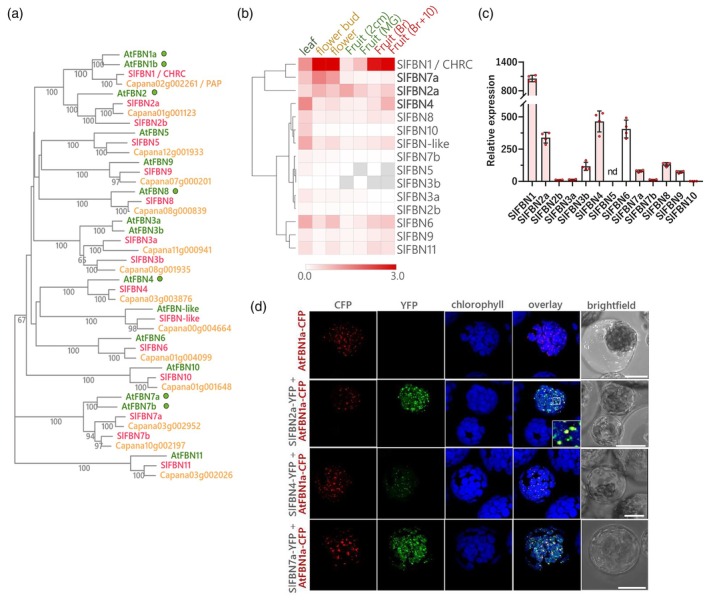
Identification of tomato FIBRILLINs (SlFBNs), expression profile and subcellular localisation.
(a) Phylogenetic analysis of FBN proteins from Arabidopsis thaliana (AtFBN), Solanum lycopersicum (SlFBN), Capsicum annuum (cv. Zunla) inferred using the Neighbour‐Joining method with 1000 bootstrap replicates. Numbers in the nodes denote bootstrap confidence levels (only values >50 are shown). Green circles indicate plastoglobuli (PG)‐related AtFBNs (van Wijk & Kessler, 2017). Solanum lycopersicum gene IDs are listed in Table S9.
(b) Heatmap representation of Slfbn expression profile. Normalised expression levels were obtained from the TomExpress database (http://tomexpress.toulouse.inra.fr).
(c) Relative expression of Slfbn genes in fruits at 1 day post‐breaker (B1) stage by quantitative polymerase chain reaction. Values are expression levels normalised to cac and act2 reference genes (mean ± SEM, n = 4 biological replicates). Tomato orthologues of PG‐related FBNs were highlighted in red. Nd, not detected.
(d) Transient co‐expression of full‐length SlFBN tagged to YFP (SlFBN2a‐YFP, SlFBN4‐YFP and SlFBN7a‐YFP), with PG marker (AtFBN1a‐CFP) into protoplasts isolated from Arabidopsis (Ler0) leaves. The CFP, YFP, chlorophyll and merged fluorescence, obtained from a confocal microscope, are shown in columns of the panel from the left to the right. Bright field images of protoplasts were shown in the last column. Co‐expression of all SlFBN‐YFP and AtFBN1a‐CFP led, at least, to partial co‐localisation. Scale bars = 20 μm.
Of the seven PG‐related Slfbn transcripts, levels of five members (Slfbn1, ‐2a, ‐7a, ‐4, ‐8) were highest at carotenoid‐enriched stages of flower and fruit development, based on transcriptome database queries (Figure 1b). Their expression profile was further confirmed by quantitative polymerase chain reaction (qPCR) analysis in fruit (Figure 1c). Slfbn1/chrc was confirmed as the most highly chromoplast‐expressed Slfbn (Kilambi et al., 2013) and correlated to Slfbn2a and ‐7a expression (Figure 1b). Besides, Slfbn4 transcripts also accumulated as tomato fruit ripening advanced. The qualitative expression profiles of the poorly characterised Slfbn6 transcripts were observed to be similar to the characterised Slfbn1 transcripts and, therefore, for logistical priorities, the Slfbn1 was included for the functional characterisation of higher order mutants.
Pepper transcriptomic data (Liu et al., 2017) was also interrogated to check Cafbn transcript abundance across fruit development. Interestingly, PG‐related Cafbn homologues had expression patterns similar to tomato (highly expressed at chromoplast‐enriched tissues/stages) (Figure S1). Cafbn1/pap was the predominant Cafbn expressed in fruits. Despite differences in flowers due to the lack of chromoplasts in pepper petals, high expression rates of Cafbn4, Cafbn2 and Cafbn7a were detected at the late stages of pepper fruit ripening, arguing for their role in carotenoid sequestration.
Based on the above orthologues and expression profiles, SlFBN2a, SlFBN4 and SlFBN7a emerged as candidates involved in PG biogenesis and carotenoid sequestration during fruit ripening, potentially complementing SlFBN1/CHRC function in chromoplasts. To first check their subcellular localisation, each SlFBN candidate, with a yellow fluorescent protein tag (SlFBN‐YFP), was transiently co‐expressed with the plastoglobular marker AtFBN1a (Vidi et al., 2006) in protoplasts. The signal fluorescence of all SlFBN‐YFP fusions was restricted to the plastids. While SlFBN4‐YFP fluorescence greatly co‐localised with the PG marker, the patterns of SlFBN2a‐YFP and SlFBN7a‐YFP suggested a more diffuse distribution of these proteins within the plastid, not restricted to PG (Figure 1d). Together, the results suggest that SlFBN2a, ‐4, ‐7a can be associated with PGs and likely have a role in PG formation and composition.
Generation of single and high‐order fbn mutants by CRISPR‐Cas
To address the contribution of SlFBN2a, ‐4, ‐7a in PG formation and isoprenoid sequestration, single loss‐of‐function mutants for the candidate SlFBNs were generated via CRISPR‐Cas technology (Figure 2; Table S1). To assess functional redundancy within the FBN family, a multiplexing approach was also undertaken to simultaneously deliver combinations of mutant alleles for SlFBN1, ‐2a, ‐4 (triple mutants) and SlFBN1, ‐2a, ‐4, ‐7a (quadruple mutants) (Figure S2).
Figure 2.

Gene‐edited fbn mutants generated by CRISPR‐Cas.
(a) Intron/exon structure of Slfbn1, Slfbn2a, Slfbn4, Slfbn7a locus. PAP/FIBRILLIN domain is indicated in grey.
(b) Nucleotide sequences showing induced mutations in Slfbn inherited mutations in T2 transgene null‐segregant plants. Indels are highlighted in red characters.
(c) Relative expression of Slfbn1/chrc and Slfbn2a in ripe fruits (B7, 7 days post‐breaker) by quantitative polymerase chain reaction. Values are relative expression levels (mean ± SD, n ≥ 3 biological replicates) to Azy control. Asterisks indicate statistically significant differences (one‐way anova, Dunnett's multiple post‐test, *P < 0.05, compared to Azy).
(d) Immunoblotting analysis of SlFBN1. Proteins extracted from B7 ripe fruits (10 μg) were separated on SDS/PAGE, blotted and membranes probed for the presence of the FBN1 with antiserum raised against Capsicum FBN1/PAP. An extra T1 Cas + line (fbn1,2a,4#60) harbouring in‐frame indel was included in the analysis for comparison.
Sequencing of the targeted genomic regions confirmed edited alleles in primary T0 transformants obtained through single and multiplexing strategies. Editing efficiency ranged from 67 to 100% (Table S2). Mutations occurred in different combinations: chimera, biallelic and homozygous, being mostly short indels upstream of PAM sites. Large out‐of‐frame deletions (>100 bp) were also detected at a lower frequency. Edited alleles harbouring frameshift or nonsense (premature stop codon, PTC) mutations were predicted as knock‐outs. Lines carrying in‐frame deletion alleles, which could either retain functionality or be loss‐of‐function, were also selected for comparison when available. Possible off‐target sites for the gRNAs examined in T0 and T1 plants (Table S1) were undetected, supporting the high specificity of the gRNAs designed. T2 Cas‐free homozygous/biallelic Slfbn plants were successfully established and used for further comprehensive phenotyping and molecular characterisation (Figure 2; Table S3). Since transcripts with PTC introduced by gene editing can be targeted by nonsense‐mediated decay pathways, by which nonsense mRNAs are destroyed (Popp & Maquat, 2016), expression of Slfbn1/chrc and Slfbn2a, both highly expressed in fruits, was checked by qPCR. Levels of PTC‐containing mutant transcripts were reduced in fbn fruits compared to their control (Figure 2c). Additionally, FBN1 protein levels were checked by immunoblotting using an antibody to CaFBN1/PAP (Deruere et al., 1994). SlFBN1 was exclusively detected in fruits from edited lines carrying either wild‐type or in‐frame deletion alleles (Figure 2d).
Visual inspection of fbn mutants revealed no obvious difference in fruit colour but altered flower pigmentation. The typical bright yellow colour failed to develop, particularly in fbn mutants harbouring Slfbn7a defective alleles (single fbn7a and quadruple fbn1,2a,4,7a) (Figure 5a). Triple and quadruple mutants, under non‐stressed growth conditions, had some altered plant morphological traits. These morphological changes consisted of a shorter internode length compared to their controls, and visually, leaf areas were reduced in the quadruple mutants (Figure S3b). No significant changes were detected in maximum quantum efficiency of PSII (Fv/Fm) of fbn mutants, except for the fbn1,2a,4,7a‐1 quadruple mutant (Figure S3d).
Figure 5.

SlFBN7a regulates carotenoid esterification driven colour retention in tomato flowers, while SlFBN1, ‐2a, 4 affect plastoglobuli (PG) morphology.
(a) HPLC chromatogram traces for control (Azy), fbn7a, triple (fbn1,2a,4) and quadruple (fbn1,2a,4,7a) mutants. Peaks corresponding to carotenoids in free forms, mono‐ and di‐esters are indicated.
Carotenoid composition and total levels of single (b) and multiple (c) fbn mutants. Data (n ≥ 4, biological replicates) were analysed by one‐way anova with Dunnett's post‐tests; asterisks denote statistically significant differences (*P < 0.05, **P < 0.01, ***P < 0.001) compared to azygous (Azy) control. Error bars indicate ±SD. D, di‐esters; F, free forms; M, mono‐esters.
(d) Representative transmission electron (TEM) micrographs of flower chromoplasts from Azy and fbn mutants. Detailed view of PG is shown on the right panels. Scale bars = 500 nm (left panels), and 50 nm (right panels).
(e) Irregular shapes assumed by aberrant triple and quadruple PGs. Red head arrows indicate PG features found in mutants as crystalloid‐like structure, physical continuity with thylakoid remnants and irregular PG morphology.
(f) Scatter plots showing diameter of PGs found in TEM petal micrographs. Yellow area highlights supersized PGs.
(g) PG number per chromoplast area.
SlFBN homologues influence the accumulation of carotenoids and other isoprenoids in plastids
To assess how Slfbn mutations influence plastidial isoprenoid metabolism, fruit and leaf isoprenoid profiles were first examined by ultra‐performance liquid chromatography‐photo diode array (UPLC‐PDA) (Figure 3; Table S4). Three fruit stages were analysed: mature green (MG), red ripe (B7) and late red ripe (B10). No significant changes in carotenoids or chlorophylls in leaf or MG tissues were observed (Table S4), except foliar phytoene. Carotenoid changes occurred predominantly in ripe fruits from high‐order fbn mutants (Figure 3b; Table S4). The early carotene intermediates (phytoene, phytofluene and ζ‐carotene) were most affected and reduced by 60% in the mutants compared to control (Azygous, Azy) at the B10 stage. Unexpectedly, the decrease in lycopene, the main carotenoid accumulating in red fruit, was less pronounced (levels of 80% retained). β‐Carotene levels, however, were unaffected despite lower levels of its immediate precursor γ‐carotene. In single fbn fruits, carotenoid content remained similar to controls at all fruit stages.
Figure 3.

SlFBNs altered isoprenoid profile in tomato fruits and leaves of fbn mutants.
(a) Schematic representation of the plastidial biosynthetic pathway of isoprenoid compounds determined by ultra‐performance liquid chromatography‐photo diode array and the corresponding sample set: fruits at mature green, ripe (B7 and B10) stages and leaves.
(b) Scatter‐plot showing representative carotenoids in ripe fruit (B10) and leaves. Each dot represents a single biological replicate. Data (n ≥ 4, biological replicates) were analysed by one‐way anova with Dunnett's post‐tests; asterisks denote statistically significant differences (*P < 0.05, **P < 0.01, ***P < 0.001) compared to azygous (Azy) control. Error bars indicate ±SD.
(c) α‐tocopherol and PC‐8 levels. AC, wild‐type; GGPP, geranylgeranyl pyrophosphate; PC‐8, plastochromanol; PQ‐9, plastoquinone. Full data set available in Table S4.
Other isoprenoids known to be sequestered in the PG core, such as tocopherol and quinone derivatives, responded differently to single or multiple losses of SlFBNs (Figure 3b; Table S4). Despite similar levels of α‐tocopherol and plastoquinone (PQ‐9), plastochromanol‐8 (PC‐8) significantly changed in fbn mutants. PC‐8 is a PQ‐9 derivative predominantly found in PG associated with stress responses (Zbierzak et al., 2010). While the lack of SlFBN2a approximately halved PC‐8 levels compared to the control, deficiency in SlFBN7a had an opposite effect, favouring PC‐8 accumulation with up to a twofold increase. Of note, similar PC‐8 levels observed among fbn7a lines carrying either in‐frame (fbn7a‐3) or nonsense alleles (fbn7a‐1, fbn7a‐2) suggested that all Slfbn7a editions likely derived from Slfbn7a null alleles. In high‐order fbn mutants, Slfbn2a and Slfbn7a defective alleles displayed compensatory interaction to modulate PC‐8 levels, though outcomes were specific to the organ and developmental stages. In triple mutants, PC‐8 levels were decreased in chloroplast‐containing tissues, MG fruits (50%) and leaves (20%), compared to the control, while no consistent effect was found in red fruits. In quadruple mutants, the addition of a defective SlFBN7a restored PC‐8 levels in fruits at all stages. Together, these findings define mutation‐specific effects of SlFBNs on levels of prenylquinones. While SlFBN2a favours PC‐8 accumulation, SlFBN7a prevents it. To further substantiate these findings, an extra set of T1 Slfbn‐edited lines harbouring Cas9‐gRNA transgene (Cas +) with the following genotypes: triple (Cas + /fbn1,2a,4), quadruple (Cas + /fbn1,2a,4,7a) and double (Cas + /fbn2a,4) mutant (Table S3) were analysed. This additional set of fbn mutants allowed us not only to confirm the fruit profile found in T2 Cas‐free progeny but also to evaluate SlFBN1‐specific contribution to the isoprenoid profile by comparing Cas + /fbn2a,4 and Cas + /fbn1,2a,4 (Table S4). PC‐8 levels were similar between double and triple mutants, implicating SlFBN2a as the major positive regulator of PC‐8 levels. The levels of PQ‐9 were also analysed in the tissue to complement the determinations from sub‐plastidial fractions and no change was observed (Table S4).
Changes in isoprenoid sequestration in fruit chromoplasts of fbn mutants
SlFBN‐dependent accumulation of isoprenoids may be linked to how these compounds are preferentially sequestered across plastid sub‐compartments (Nogueira et al., 2013). To test this hypothesis, sub‐plastidial chromoplast fractionations were isolated and analysed for isoprenoid contents (Figure 4). A differential sequestration pattern was observed in high‐order fbn mutants (Figure 4b) compared to the fbn7a line and their controls. For example, the proportion of phytoene and phytofluene was decreased in the PG fraction of higher order fbn mutants compared to the control, and instead carotenes and other carotenoids preferentially accumulated in membrane and crystal fractions. In fbn7a, reduction in the proportion of β‐carotene and lycopene content in the membrane was observed (Figure 4b). Altered deposition of α‐tocopherol and PC‐8 across sub‐chromoplast compartments was also observed (Figure 4b). Accordingly, the relative composition of PG fractions greatly changed compared to the control when multiple SlFBN were perturbed (Figure 4c). For example, in comparison to the single mutant fbn7a and its control, the proportion of α‐tocopherol and PC‐8 was reduced in the PG fractions but increased in the membranes. Together, these results suggest a reduced capacity of isoprenoid sequestration in the PG caused by multiple SlFBN disruptions.
Figure 4.

Sequestration of isoprenoids into sub‐chromoplast compartments in fbn mutants.
(a) Separation of membranes from isolated tomato chromoplasts (fruits at 3–5 days post breaker stage) by flotation on a discontinuous sucrose gradient. Plastoglobuli (PG)‐enriched buoyant fractions (F1+F2) form a red layer at the top of the gradient.
(b) Isoprenoid profile of sub‐chromoplast fractions obtained from triple (fbn1,2a,4), quadruple (fbn1,2a,4,7a) and fbn7a mutants. Values represent % of the total found in each fraction. Red arrows indicate reduced isoprenoid sequestration into PG sub‐compartment.
(c) Isoprenoid composition (% nmol) of PG fractions; for this comparison, lycopene was excluded from analysis.
(d) Representative proteins found in PG‐enriched (F1+F2) and top membrane/envelope‐enriched (F15) fractions based on proteomic analysis. Proteins were isolated from fractions, separated by SDS‐PAGE; excised gel fragments were trypsinised, and the resulting peptides analysed by mass spectrometry. Asterisks denote proteins found only in one of the control lines (AC or Azy).
Proteome analysis of fbn mutants exposes abnormal protein targeting to PG surface
To investigate the direct changes caused by lack of functional PG‐related FBNs on PGs, proteomic analysis was performed on fruit sub‐plastidial chromoplast fractions, focusing on PG‐enriched fractions. The composition of tomato fruit PG proteome was defined based on plastid proteins consistently identified in control and azygous samples, with potential contaminants filtered out according to subcellular localisation data (Table S7). From the SlFBN family, four members, SlFBN1, ‐2a, ‐4 and ‐8, were readily identified as PG‐associated proteins (Figure 4d). Surprisingly, SlFBN7a was not detected in either the PG fraction or further membrane‐related fractions. Other PG‐associated proteins (Lundquist et al., 2012) including NAD(P)H‐ubiquinone oxidoreductase C1 (NDC1), tocopherol cyclase (VTE1), PYP/PES1, plastoglobular protein of 18 kD (PG18) and flavin‐reductase related (FRR) were detected. Moreover, the vesicle‐inducing protein in plastids 1 (VIPP1) was found. This protein is known for its roles in the biogenesis and repair of thylakoid membrane protein complexes and conferring tolerance to membrane stress (Theis et al., 2019), and therefore considered a putative PG‐associated protein.
Both in triple and quadruple fbn mutants, target SlFBN1, ‐2a, and ‐4 were absent in all analysed fractions. Indeed, only a few PG representatives were detected in fbn mutant proteomes. This included FRR, a PG unknown for the triple and LOXB for the quadruple mutants. Thus, PG protein composition is influenced by mechanisms that require functional FBNs.
SlFBN7a modulates esterification capability in flower chromoplasts
The yellow pigmentation of tomato flowers is determined by xanthophylls in their esterified form conjugated to medium‐length fatty acids (C14:0–C16:0), forming mono‐ and di‐esters (Ariizumi et al., 2014; Neuman et al., 2014). While fbn2a and fbn4 were typically bright yellow, fbn7a petals appeared pale (Figure 5a). Quadruple mutants also failed to develop yellow pigmentation, showing a more severe phenotype than fbn7a. Triple fbn mutant flowers only exhibited an apparently less intense yellow colour. To better understand the biochemical basis of the pale phenotype, detailed compositional analysis of carotenoids in petal extracts was performed using high‐performance liquid chromatography (HPLC‐PDA) to resolve carotenoids in free, mono‐ and di‐ester forms (Figure 5a–c; Table S5). Carotenoid levels and composition significantly changed in fbn7a petals. The total levels were decreased by ~50% relative to the control. The free carotenoids became predominant in the fbn7a compositional profile (threefold increase), followed by a decrease in the proportion of mono‐ and di‐esters (~70 and 65% of the values found in control, respectively) (Figure 5b). No significant changes in content or the composition of fbn2a and fbn4 petals were detected. Analysis of high‐order fbn mutants further confirmed the major role of SlFBN7a in modulating the esterification capability of flower chromoplasts. Pale yellow petals of fbn1,2a,4,7a were similar to fbn7a and primarily accumulated free xanthophylls (>4‐fold higher versus control) while having lower levels of esterified xanthophylls. The decrease in mono‐ and di‐ester proportions (45 and 30% of the values found in control) exceeded those observed in fbn7a solely (Table S5), suggesting cumulative disruption in the esterification capability when multiple SlFBNs are defective. Importantly, fbn1,2a,4,7a flowers had noticeable β‐carotene quantities, which are often analytically masked by xanthophyll esters in non‐saponified extracts (Price et al., 2018). Meanwhile, triple mutants (e.g. fbn1, 2a, 4), which had an unedited FBN7a protein, retained their quantitative and qualitative carotenoid content, supporting the FBN7a role in the regulation of esterification activity in flower chromoplasts.
To explore potential compensatory molecular mechanisms triggered by the targeted loss of SlFBNs, the expression of other Slfbns encoding PG‐associated proteins in flowers was compared (Figure S8). In petals, lower mRNA levels of the target Slfbn7a, highly expressed in flowers, were found in fbn7a and quadruple fbn1,2a,4,7a mutants. Importantly, transcript levels of Slfbn7b and Slfbn8 displayed no significant perturbation in fbn mutants, suggesting a lack of compensatory transcriptional response. Moreover, expression of the genes encoding PG‐located biosynthetic enzymes, including Slccd4B and Slpes1/pyp remained largely unchanged, except for a low level decrease of Slpes1 mRNA levels found in fbn7a mutants. This latter gene transcript encodes the acyltransferase responsible for xanthophyll esterification in tomato flowers (Ariizumi et al., 2014). Expression of psy1, the gene encoding the first enzyme of the carotenoid pathway, also remained unchanged in fbn mutants. These results suggested a lack of major transcriptional compensatory mechanisms triggered by the loss of functional SlFBNs.
A similar compositional profile was also confirmed in the extra T1 fbn mutants (Figure S4; Table S5), although in this case, the total carotenoid content of triple (Cas + /fbn1,2a,4) mutants was significantly reduced compared to wild‐type Ailsa Craig (AC), which may reflect the absence of azygous comparators at this stage. Furthermore, the total carotenoid amounts found in triple (Cas +/fbn1,2a,4) and quadruple (Cas +/fbn1,2a,4,7a) mutants were equivalent, indeed lower than those found in the double (Cas +/fbn2a,4) mutant, supporting SlFBN1/CHRC as one of the main contributors to xanthophyll sequestration in flower chromoplasts.
FBNs are required for PG formation and chromoplast development
To investigate the effect of PG‐targeted SlFBN deficiency on PG formation and chromoplast architecture, petals and fruit pericarps were examined under transmission electron microscopy (TEM). The plastids in petals were present in different forms (Ariizumi et al., 2014), ranging from chloroplast‐like structure (CLS) (early stage) to the fully re‐differentiated carotenoid‐rich chromoplasts. Early stage plastids had thylakoids organised in grana and low numbers of electron‐dense PGs (Figure 5d). Mature chromoplasts instead displayed the typical abundant small PGs (diameter ~60 nm), with a less electron‐dense core where carotenoids are sequestered. In pale yellow fbn7a petals, PGs were similar to the control in size and morphology but appeared less in number (Figure 5d–g). High‐order fbn mutants had chromoplasts with great variation in sub‐structures, including thylakoid remnants, with few and supersized PGs (diameter >140 nm; Figure 5f). In some cases, their abnormal PGs seemed to be associated with the persistent dismantled thylakoids and showed a discontinuous lipid monolayer instead of a smooth surface. Additionally, they displayed crystalloid‐like inclusions (e.g. lightly stained zones) in the form of filaments (Figure 5e). Particularly in quadruple mutants, with predominantly non‐esterified xanthophylls, PGs even assumed elongated to spindle‐shaped forms, instead of round structures, filled with homogeneous electron‐dense filaments. Interestingly, early stage plastids of high‐order fbn mutants exhibited a typical CLS with only a few supersized PGs (Figure S5).
Unlike flowers, ripe fruits in tomato contain more diverse carotenoid‐sequestering structures including endomembranes, carotenoid crystals and bigger PGs (Nogueira et al., 2016). The chromoplasts found in control samples displayed crystals of lycopene and β‐carotene and several high‐electron‐dense PGs ~100 nm in diameter. PGs of single fbn2a and fbn7a mutants changed in number and morphology, whereas fbn4 chromoplasts remained similar to control (Figure 6). In fbn2a mutants, supersized PGs (diameter >200 nm) were observed at high frequency, with some of them displaying electron‐lucent areas at their core. In fbn7a mutants, fruit PGs were found with a less electron‐dense core, and only a small proportion corresponded to supersized PGs (at least 25%). The changes in PG electron density are likely to be due to altered chemical composition, affecting extraction during preparation (Hempel et al., 2014).
Figure 6.

Altered fruit chromoplast ultrastructure of fbn mutants.
(a) Two representative transmission electron (TEM) micrographs of fruit chromoplasts (5 days post‐breaker stage) from Azy and fbn mutants; scale bars = 0.5 μm. Red arrows indicate plastoglobulis (PGs) with a zoomed‐in view shown in the bottom panels; scale bars = 0.2 μm. Giant PG formation was observed in some chromoplasts of triple (fbn1,2a,4) and quadruple (fbn1,2a,4,7a) mutants.
(b) Scatter plots showing the diameter of PGs found in TEM fruit micrographs. Yellow area highlights supersized PGs.
(c) PG number per chromoplast area.
Triple and quadruple mutants displayed significant alterations in fruit chromoplast structure consistent with their carotenoid profile. They frequently failed to develop typical globular PGs. Those PGs detected were found to be unusually big, with some even reaching giant size (diameter >500 nm). Lycopene crystals seemed to be less developed in comparison with the control, consistent with lower lycopene levels (Figure 6).
Metabolome and lipidome alterations associated with FBN deficiency
The impact of PG‐associated FBNs on the plant metabolome focused on high‐order fbn mutants. Changes in the metabolic composition of primary metabolites were addressed by GC–MS profiling of polar and non‐polar‐saponified extracts of fruits and leaves (Table S6). In total, 117 metabolites were identified across the genotypes. Principal component analysis (PCA) using the combined dataset revealed little variation in the primary/intermediate metabolite composition of the fbn mutants compared to their respective controls. The exception was green fruits (MG), where a separation between fbn mutants and controls was attained (Figure S6). A limited number of metabolites were found to be altered in the fbn mutants compared to their azygous controls. In leaves, MG and ripe fruits, 18, 12, and 8 metabolite differentiators were detected, respectively, and included representative amino acids, sugars, organic acids, isoprenoids and fatty acids (Table S6). For example, SlFBN deficiency led to unexpected changes in amino acid content of fbn fruits; these changes included increases in glutamine and threonine at the MG stage (Figure S6b) and significant decreases in phenylalanine in ripe fruits (Figure S6c). Among the fatty acids released through saponification, myristic acid (C14:0) decreased consistently in the fbn samples of ripe fruit (Figure 6c), while in leaves, other minor saturated species were also significantly altered, such as pentadecanoic acid (C15:0) and heptadecanoic acid (C17:0) (Figure S6d).
PGs are known to be implicated in lipid remodelling (van Wijk & Kessler, 2017); therefore, a deeper lipidomic analysis was performed based on LC–MS: (i) an untargeted lipidome and (ii) a specific targeted analysis for triacylglycerol (TAG) and its derivatives. The score plot of the PCA from the untargeted lipidome illustrates the clustering of the fbn mutants and their controls (Figure 7a). Orthogonal partial least squares discriminant analysis was implemented to identify differences between the control, triple and quadruple fbn mutant lipidomes. The highest variable on projection (VIP) scores, which rank the importance of features in distinguishing genotypes, were retrieved. These combined analyses revealed SlFBN deficiency had the strongest effects on plastidial membrane galactolipids (monogalactosyldiacylglycerol, MGDG; digalactosyldiacylglycerol; DGDG) and TAG; the latter presumably representing pools derived from the cytosol and plastids. In leaves, while several MGDG and DGDG‐related species increased in both triple and quadruple mutants, several TAG species were found depleted, particularly in quadruple mutants. The same trend for the TAG lipid class was found in petals, but in contrast to leaf tissues, MGDG and DGDG‐related lipid species featured among the top representatives decreased in fbn mutants. Targeted TAG analysis (Figure S7) allowed further confirmation of impaired esterification capability associated with defective fbn7a alleles; several TAG‐related species were found exclusively depleted in quadruple mutants either in leaves or flowers. Notably, lowered TAG‐related lipid species were found both in triple and quadruple mutants, suggesting that aberrant PG morphologies associated with SlFBN deficiency per se are sufficient to trigger changes in lipid metabolism. Additionally, TAG‐specific fbn1,2a,4,7a changes imply that FBN7a may modulate broader lipid esterification, not only carotenoids.
Figure 7.

Untargeted lipidomic analysis of high‐order fbn mutants. Data was obtained from LC–MS based on the hydrophilic interaction liquid chromatography method.
Panels (a–d) correspond to leaf tissue, and panels (e–h) correspond to flower tissue analysis. Score plots of principal component analysis (a, e) and orthogonal partial least squares discriminant analysis (OPLS‐DA) (b, f) based on untargeted LC–MS chemical features. Data were log‐transformed and Pareto scaled. Hierarchical clustering of the top 50 variables (c, g) with the highest VIP scores identified by OPLS‐DA. Heat maps are coloured by red and blue colours indicating higher and lower abundance, respectively, as measured by row standardised z‐scores derived from LC–MS peak intensity (n = 3 biological replicates). Genotypes are annotated as coloured boxes on the top‐right of the heatmap. The boxes on the right of the heatmap indicate the lipid classes. The graphs representing quantitative changes (peak area normalised by dry weight) (d, h) of selected chemical features based on VIP scores and statistical significance (anova followed by Fisher's LSD, α = 0.05).
DISCUSSION
The present work has utilised the CRISPR/Cas‐based technology to systematically interrogate the functionality of the FBN multigene family. The repertoire of edited alleles generated (Figure 2; Table S3) both as individual and high‐order mutants has been confirmed by transcript and protein analysis. Collectively, this suite of mutants represents a valuable genetic resource (Figures 3 and 5). The fbn mutants have been phenotyped, isoprenoid composition determined, analysis of the plastid ultrastructure carried out and the metabolome and lipidome profiled. Overall, the comprehensive characterisation of the fbn mutants clearly indicated that FBNs are essential components of the PG, and although functional redundancy is evident with respect to isoprenoid biosynthesis, tissue‐specific roles linked to lipid storage or redox homeostasis‐related lipophilic compounds exist.
Cellular ultrastructure analysis revealed dramatic effects on PGs present in the fbn mutants. For example, deficiency in multiple SlFBNs such as the combination of SlFBN1, 2a, 4 resulted in heterogeneous and supersized PGs, while unaffected plastoglobular structures were found in individual fbn mutants (Figures 5 and 6; Figure S5). FBNs have been previously proposed to prevent PG coalescence (Rey et al., 2000) and promote PG formation via thylakoid budding. This hypothesis is supported by the present study where the high‐order fbn mutants presented dismantled thylakoids and discontinuous monolayers resulting in irregular PG structures (Figure 5d). The mechanism by which FBN proteins are targeted towards PGs and participate in their biogenesis and stability remains to be determined. Certain hypotheses suggest biophysical mechanisms driving the spatial partitioning (Olzmann & Carvalho, 2019), while other arguments favour post‐translational modifications influencing the sub‐organelle location (Lohscheider et al., 2016). However, a dedicated molecular machinery targeting and transporting FBN into PGs has not been described to date.
The cellular and spatial proteome analysis of isolated PG fractions performed in this work confirms the partitioning of PG‐localised SlFBN1, ‐2a, ‐4 and ‐8, but mixed results were obtained for SlFBN7a. The imaging of SlFBN7a‐YFP fusion expression indicates co‐localisation with PG structures in leaves (Figure 1), but undetectable levels of trypsin‐derived peptides in fruit tissue were found, contrasting previous results reported in tomato (Szymanski et al., 2017). Technical differences (GEL‐LC versus in‐solution digestion) and/or analytical sensitivity limitations could be attributed, but also tissue‐specific implications may be involved since Slfbn7a transcript levels are (i) significantly higher in flower tissue than in fruit and (ii) different coloured flower phenotypes were observed in the collection of fbn mutants. The carotenoid analysis obtained here indicated that SlFBN7a deficiency led to a substantial loss of carotenoid esters in petals. The enzyme catalysing carotenoid esterification in tomato flowers is PES1 (Ariizumi et al., 2014; Lewis et al., 2021), a non‐specific acyltransferase that is PG localised. However, the nature of the interaction between PES and FBN7a in tomato remains understudied. Candidate mechanisms have considered complex formation, as recently reported in rapeseed flowers (Li et al., 2023), or the assembly of enriched FBN‐substrate (carotenoid and lipids) condensates. Further hypotheses arise from the data collected in the present study. For example, the further decrease in carotenoid esters observed in the triple and quadruple mutants containing deficient SlFBN7a suggests that a specific macromolecular configuration would be favourable for optimal activity, beyond bilateral interactions. In addition, the effect of SlFBN7a on esterification activity is also extended to other lipid classes as several TAGs species were significantly decreased in both leaves and flowers of quadruple mutants carrying defective fbn7a alleles (Figures S7 and S8). Specific paralog functionality is also inferred from the data generated. For example, in the case of SlFBN2a, the deficient mutants present higher levels of PC‐8. The AtFBN2 has been shown to interact with NDC1 (Torres‐Romero et al., 2021). This enzyme reduces the oxidised pool of PQ‐9 in PG, thus regenerating the substrate (plastoquinol‐9, PQH2‐9) required by the tocopherol cyclase VTE1 to produce PC‐8 (Piller et al., 2011; Torres‐Romero et al., 2021). The SlFBN2a could have a positive effect on PC‐8 accumulation in a similar manner, either by direct association with NDC1 or indirectly by facilitating access/exposure to substrate.
In both cases, SlFBN7a and SlFBN2a, it is suggested that the molecular mechanisms of FBNs may go beyond protein–protein interaction, and functional macromolecular assemblies are potentially involved. Biosynthetic metabolons and similar molecular scaffolds have been previously described for different biosynthetic pathways, including sectors of the isoprenoids family (Fraser, Schuch, et al., 2000; Wurtzel, 2019). The evidence from the SlFBN2a and SlFBN7a reported here, together with those of AtFBN2 (Piller et al., 2011), AtFBN5 (Kim et al., 2015), the algal chloroplast SEC14‐like protein (CPSFL1) (Hertle et al., 2020) or the tomato tocopherol binding protein (SlTBP) (Bermúdez et al., 2018), suggests a putative role of FBNs as biosynthetic ancillary proteins that could bind and/or transfer substrates to enzymes directly or indirectly by operating as large macromolecular structures or biosynthetic hubs. The overall cellular and metabolic effect of the FBNs studied here is summarised in Figure 7 and a mechanistic model is proposed.
Concomitant with ultrastructural, proteome, and isoprenoid profiling changes in the PG of the fbn mutants, pronounced effects on the global lipidome in different tissues (fruit, flower, leaf) also occur. Depletion of several fatty acids in ripe fruit and leaf (Figure S6), alterations in galactolipids and TAGs composition in triple and quadruple mutants (Figures S7 and S8) such as reciprocal levels of MGDG and DGDG between flower and leaf, illustrate the impact on the plant's lipidome. In particular, the effect in leaf and flowers on plastidial galactolipids and their turnover into TAGs (Higashi et al., 2018; Lippold et al., 2012), which appears compromised by alterations in the PG structure and its proteome. The differential metabolic demands between respective plastid types should also not be ignored. Interestingly, several examples in the scientific literature report alterations in metabolite composition and metabolic reprogramming in engineered outputs targeting isoprenoid production (Enfissi et al., 2019; Fraser et al., 2007; Nogueira et al., 2023), also accompanied by profound perturbations in plastid structure and transcriptome. However, the collection of fbn mutants studied here presented subtle global effects on the steady state metabolome or key related transcripts. Collectively, this would infer that changes in metabolite composition have a greater effect on orchestrating the adaptation of cellular structures (Figure 8).
Figure 8.

Summary of changes associated with SlFBN deficiency and proposed model.
(a) Knockout of SlFBNs alters morphology and number of plastoglobuli (PGs), followed by changes in lipid metabolism and sequestration. Functional redundancies of SlFBN were exposed by aberrant PG phenotypes found in high‐order fbn mutants. SlFBN functional divergence, instead, was dissected through metabolite profiling of fbn mutants. Specifically, SlFBN7a influences esterification capability having a key role in flower colour retention. SlFBN2a together with SlFBN7a, in turn, regulate PG‐related metabolite accumulation having opposite effects on PC‐8 levels.
(b) Proposed model. As PG composition is affected by SlFBN functionality, we propose that during PG formation and growth, their surface area may serve as docking sites for FBN proteins to bind and recruit enzymes that sustain PG growth and core lipid remodelling, forming complexes such as SlFBN2a/NDC1 (based on previously shown interaction between FBN2 and NDC1, Torres‐Romero et al., 2021). Thus, SlFBNs may shape PG lipid core via local isoprenoid biosynthesis. Alternatively, other members such as FBN7a might support PG formation as part of the biosynthetic machinery for ester production, where it may selectively facilitate the availability/transfer of the newly synthesised lipids by acyltransferases (TAG and/or carotenoid esters), from plastidial membranes to PG. In the absence of functional SlFBN7a, transfer of other prenyl lipids such as PQ‐9 could be favoured, which in turn boosts PG‐local production of PC‐8. Collectively, we propose that SlFBNs can shape PG composition not only by supporting PG local isoprenoid synthesis but also by influencing the lipid transfer from thylakoids to PG. VTE1, tocopherol cyclase.
In summary, multiplex gene editing tools have been used to decipher the functionality of the FBN multigene family. Functional redundancy is evident, but collectively FBNs play a major role in PG biogenesis and stability. In addition, the results reveal tissue‐specific roles of individual FBNs, for example, SlFBN7a and SlFBN2a in carotenoid and TAG esterification and PC‐8 formation, respectively. It must be emphasised, though, that PG alterations and related lipophilic compositional changes are more pronounced when multiple FBNs have been knocked out. The findings of the present work expand the knowledge on fundamental aspects of metabolic compartmentalisation and tissue specialisation in plant cells, emphasising the role of lipoprotein particles in plastid physiology. Considering the relevance of these sub‐plastid structures in plant development and adaptation to environmental cues (Coulon et al., 2024; van Wijk & Kessler, 2017), and the interactive/dynamic nature of the FBNs discussed above, genetic resources based on multiplexed mutants represent a valuable strategy for elucidating the role of FBNs and their individual and collective mechanisms of action.
MATERIALS AND METHODS
Phylogenetic analysis
For phylogenetic analysis, BlastP searches were performed using the protein sequences of AtFBN (van Wijk & Kessler, 2017) as queries against the tomato genome (Solanaceae Genomic network, http://solgenomics.net). For pepper (Capsicum annuum, cv. Zunla), sequences were retrieved from the PepperHub database (Liu et al., 2017). The sequences were aligned using the MUSCLE package available in the MEGA X software with default parameters, and Neighbour‐Joining phylogenies with 1000 bootstrap replications were created with the distances calculated according to the best model indicated by MEGA (Kumar et al., 2018).
Subcellular localisation and confocal microscopy
For SlFBN subcellular localisation, the full‐length CDS of Slfbn2a, Slfbn4, Slfbn7a (without stop codon) were cloned into the p2YGW7, resulting in a C‐terminally tagged YFP protein. The PG marker used, Atfbn1a fused in‐frame with cfp, was from the vector p2CGW7 (Vidi et al., 2006). Arabidopsis mesophyll protoplasts were isolated as previously described (Wu et al., 2009). Protoplast transfection assay was performed using a polyethylene glycol‐based method (Yoo et al., 2007) immediately after the protoplast isolation. After 24 h, fluorescence emitted from the protoplasts was observed using a TCS‐SP8 laser scanning confocal microscope (Leica Microsystems CMS, Wetzlar, Germany) as follows: excitation 514 nm, emission 520–570 nm for YFP; excitation 458 nm, emission 465–505 nm for CFP; excitation 514 nm, emission 650–740 nm for chlorophyll autofluorescence. Emitted fluorescence was false‐coloured in green (YFP), red (CFP) and blue (chlorophyll). The confocal images shown are maximal projections of selected planes of a z‐stack.
Plant material and growth conditions
Tomato plants (Solanum lycopersicum, cv. AC) were grown under greenhouse conditions with a 16/8‐h day night photoperiod at 25°C/19°C, respectively. For experiments, homozygous/biallelic and azygous segregants were grown at the same time in the same glasshouse room.
CRISPR‐Cas9 constructs for generating tomato fbn mutants
Genomic sequences 5′‐G(N)19NGG were selected to design two gRNA spacer sequence targeting specifically Slfbn1, Slfbn2a, Slfbn4, Slfbn7a, avoiding off‐target effects, using CRISPR‐P software (Lei et al., 2014). Secondary structure analysis of target‐sgRNA sequences was performed with the programme RNA folding tool (Hamada et al., 2016). The standardised modular cloning system Golden Gate MoClo was used (Engler et al., 2014; Weber et al., 2011). The Cas9 expression cassette consists of a 2x35S Cauliflower Mosaic Virus promoter (pICH51288) and a plant codon‐optimised Cas9 coding sequence, pcoCas9 (Fauser et al., 2014), which was domesticated for removal of internal BbsI and BsaI restriction sites, both parts used to assemble a Level 1 transcriptional unit (pICH47742::2x35S::Cas9) according to Lawrenson et al. (2015). For the gRNAs, a 5′‐tailed primer harbouring the individual Slfbn spacer (guide sequence), BsaI recognition site and compatible vector overhang was used to generate specific gRNA amplicons by PCR from a gRNA scaffold template (AddGene no. 46966; for primers; see Table S9). Each gRNA PCR fragment was individually assembled with Arabidopsis U6 small RNA promoter (pICSL01009::AtU6p, AddGene no. 46968) in the appropriate Level 1 vector (plCH47751, plCH47761, plCH47772, plCH47781, pICH47791, pICH47732, pICH47742, pICH47751) depending on the position of each gRNA in the final binary vector as described by Lawrenson et al. (2015). A neomycin phosphotransferase II (NPTII) cassette (pICH47732::NOSp::NPTII, Addgene no. 51144) was used as a selectable marker for plant transformation. For single fbn mutants, NOSp::NPTII; 2x35Sp:Cas9; Level 1 AtU6p::gRNA1, Level 1 AtU6p::gRNA2 were assembled into the final Level 2‐based binary vector. For building multiplex Slfbn gRNA vectors, NOSp::NPTII; 2x35Sp:Cas9; Level 1 AtU6p::gRNA1 to AtU6p::gRNA(n) were assembled into Level P vector and produced multiplex vectors P1‐M1 (designed for targeting Slfbn1, Slfbn2a, Slfbn4) and P1‐M2 (designed for targeting Slfbn1, Slfbn2a, Slfbn4, Slfbn7a). Restriction‐ligation reactions were performed in 20 μl volume in a thermocycler for 40 cycles of 37°C for 3 min followed by 16°C for 4 min, with a final incubation of 5 min at 50°C and 5 min at 80°C. All assembled plasmids were validated by restriction enzyme analysis and confirmed by Sanger sequencing. Final binary vectors were transformed into Agrobacterium tumefaciens (strain LBA4404).
Agrobacterium tumefaciens‐mediated transformation and selection of transformants
Agrobacterium tumefaciens‐mediated transformations of the S. lycopersicum cv AC were performed as previously described in Pino et al. (2010) using cotyledon segments from 8‐day‐old seedlings precultured with Agrobacterium containing the CRISPR/Cas9 constructs of interest. Selective regeneration medium containing kanamycin (100 mg L−1) was used for explant selection. After 8 weeks, well‐rooted T0 plantlets were acclimatised to greenhouse conditions and their leaves sampled for genotyping.
Detection and determination of targeted mutagenesis
Genomic DNA was purified from leaves using the DNeasy 96 plant kit (Qiagen, Venlo, the Netherlands) following the manufacturer's instructions. The presence/absence of the transgene in regenerated primary plants (T0) and offspring (T1 and T2 plants) was detected by PCR in the genomic DNA using NPTII and Cas9‐specific primers (Table S9). To characterise the Cas9‐induced mutations in the transformed plantlets, specific primers for Slfbn, whose binding positions are about 250 bp upstream of the target site, were used for PCR amplification. Amplicons were analysed by Sanger sequencing. For those with multiple peaks in chromatograms, the PCR product was cloned into TOPO‐TA (Thermo Fisher Scientific, Waltham, MA, USA) and, at least five clones per sample were sequenced. T1 progeny of the confirmed T0 gene‐edited lines were PCR‐genotyped for screening of inherited mutant alleles (either in homozygous or biallelic state) and Cas9 null segregants. The azygous segregant line descended from an original CRISPR‐single and from a CRISPR‐multiplex vector transformant was selected as a control line. Phenotyping was performed on T2 Cas9 null segregant homozygous offspring for fbn singles, triple and quadruple mutants, unless stated otherwise. For multiplex CRISPR constructs, an extra set of gene‐edited lines for triple (fbn1,2a,4), quadruple (fbn1,2a,4,7a) as well as double mutant (fbn2a,4), the latter generated through a P1‐M1 transformant whose Slfbn1 edition failed, was obtained in AC background lacking green shoulder (glk2/uniform recessive). Their corresponding T1 progeny harboured Cas9‐gRNA expression cassette Cas+.
Isoprenoid determination and quantification
Isoprenoids (carotenoids, tocochromanols and chlorophylls) were extracted from lyophilised tissue powder (15 mg) as described by Enfissi et al. (2010). From homogenised (powdered) freeze‐dried tomato material, isoprenoids (carotenoids, chlorophylls, quinones and tocopherols) were extracted using methanol (250 ml) and chloroform (500 ml) with 50 mM Tris–HCl pH 7.5 (250 ml) added subsequently to aid the formation of a liquid: liquid partition. The resulting hypophase after centrifugation at 3000 × g for 3 min was removed and the suspension re‐extracted with chloroform. The pooled chloroform extract was dried under nitrogen gas and resuspended in ethyl acetate (50 ml) for HPLC separation. HPLC analysis was carried out with a Waters Alliance system; the separation column used was a C30 bonded silica‐based reverse‐phase column 250 × 4.6 mm i.d. 5 μm (YMC [Lancaster, UK], Thermo Fisher Scientific, Phenomenex [Macclesfield, UK]) and corresponding C30 guard column (20 × 4.6 mm, 5 μm). Mobile phase solvents used were (A) methanol, (B) methanol:water (80:20) (vol.) containing 0.2% (w/vol.) of ammonium acetate and (C) methyl‐tert butyl ether. A flow rate of 1 ml min−1 was used and the gradient ran over 60 min. At 0–6 min the mobile phase composition is 95% A, 5% B; at 7 min: 80% A, 5% B, 15% C; 12 min: 80% A, 5% B, 15% C; 32 min: 30% A, 5% B, 65% C; 54 min: 30% A, 5% B, 65% C; 56 min: 95% A, 5% B; 62 min: 95% A, 5% B. An injection volume of 10 μl is used and the column oven is maintained at 25°C. The elute is monitored continuously across the wavelength range of 200–600 nm using an on‐line PDA (Fraser et al., 2000).
A similar extraction of isoprenoids was carried out for UPLC with an online PDA. Compounds were analysed by reverse‐phase chromatography using a UPLC system (Acquity, Waters Company, Wilmslow, UK) equipped with a PDA detector (Acquity, Waters Company). A UPLC BEH‐C18 column (100 mm × 2.1 mm; 1.7 μm, Acquity, Waters Company) was used for separation as described in Nogueira et al. (2013). In both cases, HPLC and UPLC peak identification was achieved by comparison of characteristic UV/Vis spectrum with authentic standards, reference spectra and retention times (Fraser et al., 2007). For xanthophyll esters in flowers, a spectra table is available in Table S10. Quantification was performed using dose–response curves obtained from authentic standards when available.
Sub‐chromoplast fractionation
Chromoplasts were isolated from fruits (90 g) at B3–B4 stage, and sub‐compartments were fractionated using a discontinuous gradient of sucrose, according to Nogueira et al. (2013). In brief, ripening tomato fruit (150 g) were deseeded and cut into pieces (ca 1 cm squares), then soaked in extraction buffer 50 mM Tris–HCl, 1 mM DTT, 1 mM EDTA with 0.4 m sucrose and adjusted to pH 7.8. Homogenisation was performed with a Waring blender (2 × 3 sec blasts). The slurry was filtered through 2–4 layers of muslin. The suspension was centrifuged at 5000 × g for 10 min at 4°C. The resulting pellet was resuspended in extraction buffer and centrifuged at 9000 × g for 10 min at 4°C. The crude plastid pellet was resuspended in 50 mM Tricine pH 7.9, containing 2 mM EDTA and 2 mM DTT with 5 mM sodium bisulphite. Lysis of the plastids was carried out with a Potter‐Elvehjem tissue lyser. The lysed fraction was separated on a discontinuous gradient with 38, 20 and 5% w/v sucrose present in the buffer used to resuspend plastids.
qPCR expression analyses
Total RNA was extracted from flowers and fruit pericarps using the RNeasy kit (Qiagen) according to the manufacturer's instructions. RNA quality was assessed by agarose gel electrophoresis. Total RNA (1 μg) was treated with DNaseI and converted into cDNA using the QuantiTect Reverse Transcription kit (Qiagen), according to the manufacturer's protocols. Real‐time qPCR assays were performed in duplicate using the RotorGene SYBR green PCR kit (Qiagen) on the Rotor‐Gene Q, with approximately 10 ng of reverse‐transcribed RNA. Primer sequences are listed in Table S1. Relative expression was calculated as described by Quadrana et al. (2013). For reference gene selection, expression stability of four known reference genes—cac, exp, act1 and act2 (Cheng et al., 2017; Expósito‐Rodríguez et al., 2008), were evaluated using GeNorm (Vandesompele et al., 2002) with act2 and cac selected based on the lowest expression stability values (M) of 0.362 and 0.438, respectively.
Transmission electron microscopy
Segments from pericarp fruit and mature, opened flowers were fixed at room temperature in solution (3% [v/v] glutaraldehyde, 4% [v/v] formaldehyde buffered with 0.1 m PIPES buffer pH 7.2) and then stored at 4°C for at least 24 h until processing. Samples were post‐fixed in buffered 1% (w/v) osmium tetroxide and uranyl acetate, washed, dehydrated in a graded series of acetone and embedded in resin. Ultrathin sections were stained with Reynolds lead citrate and imaged on a Tecnai T12 Transmission Electron Microscope (Field Electron and Ion Company, Hillsboro, OR, USA). Measurements were made in ImageJ v.2.0.0‐rc‐68/1.52e (National Institutes of Health, Bethesda, MD, USA).
Immunoblotting and proteomic analysis
For immunoblotting, ripe fruit homogenates (150 mg) were extracted with extraction buffer containing 0.1 m Tris‐Cl (pH 7.5–8.0), 6 m urea, 2 m thiourea, 0.2% (v/v) Triton X‐100, 0.2% (w/v) sarcosyl, and 2 mM DTT. Total protein content was quantified using the Bradford method (Bradford, 1976), and 10 μg of total protein was separated on a 12.5% w/v polyacrylamide gel and blotted to PVDF membrane. Blots were probed for the presence of the FBN1 using a polyclonal antibody raised against Capsicum FBN1/PAP (Deruere et al., 1994) and analysed using the method described by Fraser et al. (1994).
For proteomic analysis, total protein was extracted from isolated fractions, run in the SDS‐PAGE gel, subsequently digested with trypsin and analysed into nano LC–MS/MS as described by Nogueira et al. (2013). Analyses were conducted in an AdvanceBio Peptide Map column (2.1 × 100 mm, 2.7 μm, Agilent Technologies Inc., Santa Clara, CA, USA) and Infinity II 1290 UHPLC coupled to a 6550 iFunnel QTof (Agilent Technologies Inc.). Data analyses were performed using Spectrum Mill MS Proteomics Workbench (Rev B.06.00.201, Agilent Technologies Inc.) and Mascot Distiller (v2.4.2.0, Matrix Science, London, UK) as described in Nogueira et al. (2013).
Metabolite profiling by GC–MS
Polar and non‐polar saponified extracts from fruits and leaves were prepared and analysed as previously described in Almeida et al. (2021). For polar and non‐polar metabolite analysis, freeze‐dried tomato tissue (10 mg) was extracted with methanol (400 μl); then, ultrapure water was added (400 μl). The suspension was mixed by rotary inversion for 1 h. Chloroform (800 μl) was then added, and the mixture was centrifuged for 5 min at 20 000 × g . From the polar extract (upper phase) a 10 μl aliquot was removed, and D4‐Succinic acid (10 μl of a 0.1 mg ml−1 stock). From the non‐polar (lower phase), 700 μl was transferred and D27‐Myristic acid (10 μl) was added. The polar and non‐polar extracts were dried. Derivatisationwas performed using methoxyamine hydrochloride (MEOX) solution (1 mg ml−1) in pyridine; 30 μl of the MEOX solution was added to the dried polar or non‐polar extracts. Following heating for 1 h at 40°C, N‐methyl‐N‐trimethylsilyl (MSTFA; 70 μl) was added for a further 2 h at 40°C. Using 1 ml injections, GC/MS separations were conducted using a temperature gradient starting at 70°C for 1 min, followed by an 8°C min−1 increase to 325°C, holding the final temperature for 2 min. Data analysis was carried out using AMDIS (version 2.73), while confirmation of metabolites can be achieved through analysis of authentic standards or a database (e.g. NIST11, Bethesda, MD, USA; http://chemdata.nist.gov/mass‐spc/ms‐search/).
Lipid analysis by liquid chromatography (LC)–MS/MS
Leaf, fruit and flower petals (5–10 mg) were extracted with chloroform: methanol (1 ml, 2:1, v/v). Analysis was performed as previously described by Drapal et al. (2022) for phospho‐ and galactosyl lipids and neutral lipids. Samples were processed with Agilent Profinder (v10.0 SP1, Agilent Technologies Inc.) and identification was performed through comparison to authentic standards and MS/MS fragmentation pattern.
Statistical analyses
Univariate statistical analysis for comparison between control and fbn mutants was performed by Student's t‐test or anova followed by a post hoc test (as stated in figure and tables) with the level of significance set to 0.05, using GraphPad Prism 8 software. For multivariate statistical analysis, MetaboAnalyst 4.0 (Chong et al., 2019) and Simca®17 (Sartorius, Gottingen, Germany) software were utilised.
CONFLICT OF INTEREST
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
AUTHOR CONTRIBUTIONS
Conceptualisation: JA and PDF; methodology and experimentation: JA, KL, LP‐F, MD and PDF; formal analysis: JA; investigation: JA; resources: PDF; data curation: JA; writing—original draft: JA; writing—review and editing: JA, LP‐F, MD, KL, EMAE and PDF; visualisation: JA; project administration: PDF and EMAE; funding acquisition: PDF and EMAE.
Supporting information
Figure S1. Expression profile of FIBRILLIN (FBN) loci in pepper. Heatmap representation of Cafbn expression profile in pepper. Expression data were obtained from PepperHub (http://www.hnivr.org/), represented as FPKM‐normalised values.
Figure S2. CRISPR/Cas9 constructs for target gene editing of Slfbn genes. Scheme of final binary vectors containing the NPTII selectable marker, Cas9 driven by the 2x35S promoter and transcriptional units for gRNA expression (under control of U6 promoter). LB, left border; RB, right border.
Figure S3. Plant morphology of multiple knockout fbn mutants. (a) Representative fully expanded leaves (cv. Ailsa Craig) of 10‐weeks‐old Azy, AC and triple and quadruple fbn mutants. All plants were cultivated together in the same growth chamber. Orange arrows indicate lesions caused by fungal pathogens. Bar = 5 cm. (b) Internode length of the four equivalent internodes. (c) Maximum quantum efficiency of Photosystem II (Fv/Fm). Data (n > 5, biological replicates) are means and error bars indicate standard deviation (SD). Statistically significant differences between Azy and fbn mutant are indicated by P‐values (one‐way anova with Dunnett's post‐tests, P < 0.05).
Figure S4. Flower carotenoid composition of Cas + /Slfbn‐edited lines. T1 Slfbn‐edited lines profiling by HPLC‐PDA. Carotenoid composition and total levels of double (Cas+/fbn2a,4), triple (Cas+/fbn1,2a,4) and quadruple (Cas+/fbn1,2a,4,7a) fbn mutants. Data (n = 3, biological replicates) were analysed by one‐way anova with Dunnett's post‐test; asterisks denote statistically significant differences (*P < 0.05, **P < 0.01, ***P < 0.001) compared to AC control. Error bars indicate ±SD. D, di‐esters; F, free forms; M, mono‐esters.
Figure S5. Aberrant plastoglobuli (PG) morphology found in flower plastids of high‐order fbn mutants. Representative transmission electron micrographs (TEM) of early stage plastids (bearing thylakoids organised in grana and a few electron dense PGs) from petals from Azy and fbn mutants. Detailed view of PGs was shown on the inset panels. Giant PG formation was observed in some plastids of quadruple (fbn1,2a,4,7a) mutants. Scale bars = 500 nm (main panels), and 50 nm (inset).
Figure S6. Effect of SlFBN deficiency on tomato primary metabolism. (a) Scores plot obtained by principal component analysis (PCA) for metabolite levels measured in polar and non‐polar extracts in fruits at mature green (MG) and ripe (B7) stage, and in leaves (from top to bottom). Changes in metabolites associated with SlFBN deficiency in MG (b), ripe fruits (c) and leaves (d). Quantification was determined relative to the internal standard and values are presented as mean ± SD from four biological replicates. Only significant changes compared to respective Azy control are shown (pair‐wise t‐test corrected for multiple comparison using Holm–Sidak's post‐test; Adjusted *P < 0.05, **P < 0.01, ***P < 0.001). Full data set available in Table S6. C14:0, myristic acid; C15:0, pentadecanoic acid; C17:0, heptadecanoic acid.
Figure S7. Targeted lipidomic analysis of triacylglycerol (TAG) and its derivatives. (a, c) Heat map of top 100 features identified by anova (Post hoc Fisher's LSD, α = 0.05, corrected by multiple comparisons) in leaf and flower, respectively. Red and blue colours indicate higher and lower abundance, respectively, as measured by column standardised z‐scores derived from LC–MS peak intensity (n = 3 biological replicates). Genotypes are indicated on the right of the heatmap. The boxes on the top of the heatmap indicate the lipid classes, and whether the variable is found among the highest VIP scores (top 100) identified by OPLS‐DA. (b, d) Selected chemical features from heat maps of leaf and petal, respectively, that show statistically significant changes on fbn mutants (anova followed by Fisher's LSD, α = 0.05).
Figure S8. Relative expression of Slfbns and other genes encoding metabolic enzymes associated with PG by qPCR. Values are expression levels normalised to cac and act2 reference genes (mean ± SEM; n ≥ 3 biological replicates). Significant differences (Student's t‐test, *P < 0.05, **P < 0.01, ***P < 0.001) between fbn mutants and control are indicated.
Table S1. gRNA recognition sequences in Slfbn genes and off‐targets.
Table S2. Number of plants recovered from Agrobacterium‐mediated transformation harbouring mutations in the Slfbn target gene.
Table S3. Slfbn edited lines phenotyped in this study.
Table S4. Isoprenoid profile of fruits and leaves from fbn mutants determined by UPLC‐PDA.
Table S5. Isoprenoid profile of petals from fbn mutants determined by HPLC‐PDA.
Table S6. Metabolite levels measured in fruits (MG and B7 stage) and leaves by GC–MS.
Table S7. Proteome associated with PG fraction of tomato fruits from control and fbn mutant genotypes.
Table S8. Proteome associated with membrane fraction of tomato fruits from control and fbn mutant genotypes.
Table S9. Primers used in this study.
Table S10. Peak identification of isoprenoids analysed by HPLC‐PDA.
ACKNOWLEDGEMENTS
This work is supported by the H2020 programme TomGEM/679766; a holistic multiactor approach towards the design of new tomato varieties and management practices to improve yield and quality in the face of climate change, and Biotechnology and Biological Sciences Research Council (BBSRC)OPTICAR Project (optimisation of tomato fruit carotenoid content for nutritional improvement and industrial exploitation; Project BB/P001742/1). Group members at RHUL are thanked for their constructive advice. Dr. Marcel Kuntz, Grenoble University for the initial antibody to FBN. pICSL4723‐P1 and pAGM472 were kindly provided by Mark Youles, Sainsbury Laboratory, UK. For the purpose of open access, the author has applied a Creative Commons Attribution (CCBY) licence to any Author Accepted Manuscript version arising from this submission. BioRender is acknowledged for the genration of Figure 8.
DATA AVAILABILITY STATEMENT
Raw data files for lipidomics and proteomics are available through Mendeley data (DOI: 10.17632/9xmnybzgnd.1 and DOI: 10.17632/3hxz7c77r5.1). All other data generated or analysed during this study are included in this article (Appendix S1).
REFERENCES
- Almeida, J. , Perez‐Fons, L. & Fraser, P.D. (2021) A transcriptomic, metabolomic and cellular approach to the physiological adaptation of tomato fruit to high temperature. Plant, Cell & Environment, 44, 2211–2229. [DOI] [PubMed] [Google Scholar]
- Ariizumi, T. , Kishimoto, S. , Kakami, R. , Maoka, T. , Hirakawa, H. , Suzuki, Y. et al. (2014) Identification of the carotenoid modifying gene PALE YELLOW PETAL 1 as an essential factor in xanthophyll esterification and yellow flower pigmentation in tomato (Solanum lycopersicum). Plant Journal, 79, 453–465. [DOI] [PubMed] [Google Scholar]
- Bermúdez, L. , del Pozo, T. , Silvestre Lira, B. , de Godoy, F. , Boos, I. , Romanò, C. et al. (2018) A tomato tocopherol‐binding protein sheds light on intracellular α‐tocopherol metabolism in plants. Plant & Cell Physiology, 59, 2188–2203. [DOI] [PubMed] [Google Scholar]
- Berry, H.M. , Rickett, D.V. , Baxter, C.J. , Enfissi, E.M.A. & Fraser, P.D. (2019) Carotenoid biosynthesis and sequestration in red chilli pepper fruit and its impact on colour intensity traits. Journal of Experimental Botany, 70, 2637–2650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradford, M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein‐dye binding. Analytical Biochemistry, 72, 248–254. [DOI] [PubMed] [Google Scholar]
- Bréhélin, C. , Kessler, F. & van Wijk, K.J. (2007) Plastoglobules: versatile lipoprotein particles in plastids. Trends in Plant Science, 12, 260–266. [DOI] [PubMed] [Google Scholar]
- Cheng, Y. , Bian, W. , Pang, X. , Yu, J. , Ahammed, G.J. , Zhou, G. et al. (2017) Genome‐wide identification and evaluation of reference genes for quantitative RT‐PCR analysis during tomato fruit development. Frontiers in Plant Science, 8, 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chong, J. , Wishart, D.S. & Xia, J. (2019) Using MetaboAnalyst 4.0 for comprehensive and integrative metabolomics data analysis. Current Protocols in Bioinformatics, 68, 1–128. [DOI] [PubMed] [Google Scholar]
- Coulon, D. , Nacir, H. , Bahammou, D. , Jouhet, J. , Bessoule, J.J. , Fouillen, L. et al. (2024) Roles of plastoglobules and lipid droplets in leaf neutral lipid accumulation during senescence and nitrogen deprivation. Journal of Experimental Botany, 75, 6542–6562. [DOI] [PubMed] [Google Scholar]
- Deruere, J. , Romer, S. , d'Harlingue, A. , Backhaus, R.A. , Kuntz, M. & Camara, B. (1994) Fibril assembly and carotenoid overaccumulation in chromoplasts: a model for supramolecular lipoprotein structures. Plant Cell, 6, 119–133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drapal, M. , Gerrish, C. & Fraser, P.D. (2022) Changes in carbon allocation and subplastidal amyloplast structures of specialised Ipomoea batatas (sweet potato) storage root phenotypes. Phytochemistry, 203, 113409. [DOI] [PubMed] [Google Scholar]
- Enfissi, E.M.A. , Barneche, F. , Ahmed, I. , Lichtlé, C. , Gerrish, C. , McQuinn, R.P. et al. (2010) Integrative transcript and metabolite analysis of nutritionally enhanced DE‐ETIOLATED1 downregulated tomato fruit. Plant Cell, 22, 1190–1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enfissi, E.M.A. , Nogueira, M. , D'Ambrosio, C. , Stigliani, A.L. , Giorio, G. , Misawa, N. et al. (2019) The road to astaxanthin production in tomato fruit reveals plastid and metabolic adaptation resulting in an unintended high lycopene genotype with delayed over‐ripening properties. Plant Biotechnology Journal, 17, 1501–1513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Engler, C. , Youles, M. , Gruetzner, R. , Ehnert, T.‐M. , Werner, S. , Jones, J.D.G. et al. (2014) A golden gate modular cloning toolbox for plants. ACS Synthetic Biology, 3, 839–843. [DOI] [PubMed] [Google Scholar]
- Expósito‐Rodríguez, M. , Borges, A.A. , Borges‐Pérez, A. & Pérez, J.A. (2008) Selection of internal control genes for quantitative real‐time RT‐PCR studies during tomato development process. BMC Plant Biology, 8, 131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fang, X. , Liu, S. , Gao, P. , Liu, H. , Wang, X. , Luan, F. et al. (2020) Expression of ClPAP and ClPSY1 in watermelon correlates with chromoplast differentiation, carotenoid accumulation, and flesh color formation. Scientia Horticulturae, 270, 109437. [Google Scholar]
- Fauser, F. , Schiml, S. & Puchta, H. (2014) Both CRISPR/Cas‐based nucleases and nickases can be used efficiently for genome engineering in Arabidopsis thaliana . Plant Journal, 79, 348–359. [DOI] [PubMed] [Google Scholar]
- Fraser, P.D. , Enfissi, E.M.A. , Halket, J.M. , Truesdale, M.R. , Yu, D. , Gerrish, C. et al. (2007) Manipulation of phytoene levels in tomato fruit: effects on isoprenoids, plastids, and intermediary metabolism. The Plant Cell, 19, 3194–3211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fraser, P.D. , Pinto, M.E.S. , Holloway, D.E. & Bramley, P.M. (2000) Application of high‐performance liquid chromatography with photodiode array detection to the metabolic profiling of plant isoprenoids. The Plant Journal, 24, 551–558. [DOI] [PubMed] [Google Scholar]
- Fraser, P.D. , Schuch, W. & Bramley, P.M. (2000) Phytoene synthase from tomato (Lycopersicon esculentum) chloroplasts—partial purification and biochemical properties. Planta, 211, 361–369. [DOI] [PubMed] [Google Scholar]
- Fraser, P.D. , Truesdale, M.R. , Bird, C.R. , Schuch, W. & Bramley, P.M. (1994) Carotenoid biosynthesis during tomato fruit development (evidence for tissue‐specific gene expression). Plant Physiology, 105, 405–413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein, J.L. & Brown, M.S. (2009) The LDL receptor. Arteriosclerosis, Thrombosis, and Vascular Biology, 29, 431–438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamada, M. , Ono, Y. , Kiryu, H. , Sato, K. , Kato, Y. , Fukunaga, T. et al. (2016) Rtools: a web server for various secondary structural analyses on single RNA sequences. Nucleic Acids Research, 44, W302–W307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hempel, J. , Amrehn, E. , Quesada, S. , Esquivel, P. , Jiménez, V.M. , Heller, A. et al. (2014) Lipid‐dissolved γ‐carotene, β‐carotene, and lycopene in globular chromoplasts of peach palm (Bactris gasipaes Kunth) fruits. Planta, 240, 1037–1050. [DOI] [PubMed] [Google Scholar]
- Hertle, A.P. , García‐Cerdán, J.G. , Armbruster, U. , Shih, R. , Lee, J.J. , Wong, W. et al. (2020) A Sec14 domain protein is required for photoautotrophic growth and chloroplast vesicle formation in Arabidopsis thaliana . Proceedings of the National Academy of Sciences, 117, 9101–9111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Higashi, Y. , Okazaki, Y. , Takano, K. , Myouga, F. , Shinozaki, K. , Knoch, E. et al. (2018) HEAT INDUCIBLE LIPASE1 remodels chloroplastic monogalactosyldiacylglycerol by liberating α‐linolenic acid in Arabidopsis leaves under heat stress. The Plant Cell, 30, 1887–1905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilambi, H.V. , Kumar, R. , Sharma, R. & Sreelakshmi, Y. (2013) Chromoplast‐specific carotenoid‐associated protein appears to be important for enhanced accumulation of carotenoids in hp1 tomato fruits. Plant Physiology, 161, 2085–2101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, E.H. , Lee, D.W. , Lee, K.R. , Jung, S.J. , Jeon, J.S. & Kim, H.U. (2017) Conserved function of fibrillin5 in the plastoquinone‐9 biosynthetic pathway in Arabidopsis and rice. Frontiers in Plant Science, 8, 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, E.H. , Lee, Y. & Kim, H.U. (2015) Fibrillin 5 is essential for plastoquinone‐9 biosynthesis by binding to solanesyl diphosphate synthases in Arabidopsis. Plant Cell, 27, 2956–2971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar, S. , Stecher, G. , Li, M. , Knyaz, C. & Tamura, K. (2018) MEGA X: molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution, 35, 1547–1549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langenkämper, G. , Manac'h, N. , Broin, M. , Cuiné, S. , Becuwe, N. , Kuntz, M. et al. (2001) Accumulation of plastid lipid‐associated proteins (fibrillin/CDSP34) upon oxidative stress, ageing and biotic stress in Solanaceae and in response to drought in other species. Journal of Experimental Botany, 52, 1545–1554. [DOI] [PubMed] [Google Scholar]
- Lawrenson, T. , Shorinola, O. , Stacey, N. , Li, C. , Østergaard, L. , Patron, N. et al. (2015) Induction of targeted, heritable mutations in barley and Brassica oleracea using RNA‐guided Cas9 nuclease. Genome Biology, 16, 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lei, Y. , Lu, L. , Liu, H.Y. , Li, S. , Xing, F. & Chen, L.L. (2014) CRISPR‐P: a web tool for synthetic single‐guide RNA design of CRISPR‐system in plants. Molecular Plant, 7, 1494–1496. [DOI] [PubMed] [Google Scholar]
- Leitner‐Dagan, Y. , Ovadis, M. , Shklarman, E. , Elad, Y. , David, D.R. & Vainstein, A. (2006) Expression and functional analyses of the plastid lipid‐associated protein CHRC suggest its role in chromoplastogenesis and stress. Plant Physiology, 142, 233–244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis, E.R. , Nogueira, M. , Enfissi, E.M.A. & Fraser, P.D. (2021) The esterification of xanthophylls in Solanum lycopersicum (tomato) chromoplasts; the role of a non‐specific acyltransferase. Phytochemistry, 191, 112912. [DOI] [PubMed] [Google Scholar]
- Li, R. , Zeng, Q. , Zhang, X. , Jing, J. , Ge, X. , Zhao, L. et al. (2023) Xanthophyll esterases in association with fibrillins control the stable storage of carotenoids in yellow flowers of rapeseed (Brassica juncea). New Phytologist, 240, 285–301. [DOI] [PubMed] [Google Scholar]
- Lichtenthaler, H.K. (2013) Plastoglobuli, thylakoids, chloroplast structure and development of plastids. In: Advances in photosynthesis and respiration, Vol. 36. Switzerland: Springer, pp. 337–361. [Google Scholar]
- Lippold, F. , vom Dorp, K. , Abraham, M. , Hölzl, G. , Wewer, V. , Yilmaz, J.L. et al. (2012) Fatty acid phytyl ester synthesis in chloroplasts of Arabidopsis. Plant Cell, 24, 2001–2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, F. , Yu, H. , Deng, Y. , Zheng, J. , Liu, M. , Ou, L. et al. (2017) PepperHub, an informatics hub for the chili pepper research community. Molecular Plant, 10, 1129–1132. [DOI] [PubMed] [Google Scholar]
- Lohscheider, J.N. , Friso, G. & Van Wijk, K.J. (2016) Phosphorylation of plastoglobular proteins in Arabidopsis thaliana . Journal of Experimental Botany, 67, 3975–3984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lundquist, P.K. , Poliakov, A. , Bhuiyan, N.H. , Zybailov, B. , Sun, Q. & van Wijk, K.J. (2012) The functional network of the Arabidopsis plastoglobule proteome based on quantitative proteomics and genome‐wide coexpression analysis. Plant Physiology, 158, 1172–1192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lundquist, P.K. , Shivaiah, K.K. & Espinoza‐Corral, R. (2020) Lipid droplets throughout the evolutionary tree. Progress in Lipid Research, 78, 101029. [DOI] [PubMed] [Google Scholar]
- Neuman, H. , Galpaz, N. , Cunningham, F.X. , Zamir, D. & Hirschberg, J. (2014) The tomato mutation nxd1 reveals a gene necessary for neoxanthin biosynthesis and demonstrates that violaxanthin is a sufficient precursor for abscisic acid biosynthesis. The Plant Journal, 78, 80–93. [DOI] [PubMed] [Google Scholar]
- Nogueira, M. , Berry, H. , Nohl, R. , Klompmaker, M. , Holden, A. & Fraser, P.D. (2016) Subchromoplast fractionation protocol for different Solanaceae fruit species. Bio‐Protocol, 6, e1861. [Google Scholar]
- Nogueira, M. , Enfissi, E.M. , Almeida, J. & Fraser, P.D. (2018) Creating plant molecular factories for industrial and nutritional isoprenoid production. Current Opinion in Biotechnology, 49, 80–87. [DOI] [PubMed] [Google Scholar]
- Nogueira, M. , Enfissi, E.M.A. , Price, E.J. , Menard, G.N. , Venter, E. , Eastmond, P.J. et al. (2023) Ketocarotenoid production in tomato triggers metabolic reprogramming and cellular adaptation: the quest for homeostasis. Plant Biotechnology Journal, 22, 427–444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nogueira, M. , Mora, L. , Enfissi, E.M. , Bramley, P.M. & Fraser, P.D. (2013) Subchromoplast sequestration of carotenoids affects regulatory mechanisms in tomato lines expressing different carotenoid gene combinations. The Plant Cell, 25, 4560–4579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olzmann, J.A. & Carvalho, P. (2019) Dynamics and functions of lipid droplets. Nature Reviews Molecular Cell Biology, 20, 137–155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piller, L.E. , Besagni, C. , Ksas, B. , Rumeau, D. , Bréhélin, C. , Glauser, G. et al. (2011) Chloroplast lipid droplet type II NAD(P)H quinone oxidoreductase is essential for prenylquinone metabolism and vitamin K 1 accumulation. Proceedings of the National Academy of Sciences of the United States of America, 108, 14354–14359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pino, L.E. , Lombardi‐Crestana, S. , Azevedo, M.S. , Scotton, D.C. , Borgo, L. , Quecini, V. et al. (2010) The Rg1 allele as a valuable tool for genetic transformation of the tomato ‘micro‐Tom’ model system. Plant Methods, 6, 23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popp, M.W. & Maquat, L.E. (2016) Leveraging rules of nonsense‐mediated mRNA decay for genome engineering and personalized medicine. Cell, 165, 1319–1322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price, E.J. , Bhattacharjee, R. , Lopez‐Montes, A. & Fraser, P.D. (2018) Carotenoid profiling of yams: clarity, comparisons and diversity. Food Chemistry, 259, 130–138. [DOI] [PubMed] [Google Scholar]
- Quadrana, L. , Almeida, J. , Otaiza, S.N. , Duffy, T. , Corrêa da Silva, J. , de Godoy, F. et al. (2013) Transcriptional regulation of tocopherol biosynthesis in tomato. Plant Molecular Biology, 81, 309–325. [DOI] [PubMed] [Google Scholar]
- Rey, P. , Gillet, B. , Romer, S. , Eymery, F. , Massimino, J. , Peltier, G. et al. (2000) Over‐expression of a pepper plastid lipid‐associated protein in tobacco leads to changes in plastid ultrastructure and plant development upon stress. The Plant Journal, 21, 483–494. [DOI] [PubMed] [Google Scholar]
- Rottet, S. , Besagni, C. & Kessler, F. (2015) The role of plastoglobules in thylakoid lipid remodeling during plant development. Biochimica et Biophysica Acta ‐ Bioenergetics, 1847, 889–899. [DOI] [PubMed] [Google Scholar]
- Shanmugabalaji, V. , Besagni, C. , Piller, L.E. , Douet, V. , Ruf, S. , Bock, R. et al. (2013) Dual targeting of a mature plastoglobulin/fibrillin fusion protein to chloroplast plastoglobules and thylakoids in transplastomic tobacco plants. Plant Molecular Biology, 81, 13–25. [DOI] [PubMed] [Google Scholar]
- Singh, D.K. & McNellis, T.W. (2011) Fibrillin protein function: the tip of the iceberg? Trends in Plant Science, 16, 432–441. [DOI] [PubMed] [Google Scholar]
- Spicher, L. & Kessler, F. (2015) Unexpected roles of plastoglobules (plastid lipid droplets) in vitamin K1 and E metabolism. Current Opinion in Plant Biology, 25, 123–129. [DOI] [PubMed] [Google Scholar]
- Sun, T. , Yuan, H. , Chen, C. , Kadirjan‐Kalbach, D.K. , Mazourek, M. , Osteryoung, K.W. et al. (2020) ORHis, a natural variant of OR, specifically interacts with plastid division factor ARC3 to regulate chromoplast number and carotenoid accumulation. Molecular Plant, 13, 864–878. [DOI] [PubMed] [Google Scholar]
- Suzuki, M. , Takahashi, S. , Kondo, T. , Dohra, H. , Ito, Y. , Kiriiwa, Y. et al. (2015) Plastid proteomic analysis in tomato fruit development. PLoS One, 10, 1–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szymanski, J. , Levin, Y. , Savidor, A. , Breitel, D. , Chappell‐Maor, L. , Heinig, U. et al. (2017) Label‐free deep shotgun proteomics reveals protein dynamics during tomato fruit tissues development. The Plant Journal, 90, 396–417. [DOI] [PubMed] [Google Scholar]
- Theis, J. , Gupta, T.K. , Klingler, J. , Wan, W. , Albert, S. , Keller, S. et al. (2019) VIPP1 rods engulf membranes containing phosphatidylinositol phosphates. Scientific Reports, 9, 8725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torres‐Romero, D. , Gómez‐Zambrano, Á. , Serrato, A.J. , Sahrawy, M. & Mérida, Á. (2021) Arabidopsis fibrillin 1‐2 subfamily members exert their functions via specific protein–protein interactions. Journal of Experimental Botany, 73, 1–914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Wijk, K.J. & Kessler, F. (2017) Plastoglobuli: plastid microcompartments with integrated functions in metabolism, plastid developmental transitions, and environmental adaptation. Annual Review of Plant Biology, 68, 253–289. [DOI] [PubMed] [Google Scholar]
- Vandesompele, J. , De Preter, K. , Filip, P. , Poppe, B. , Van Roy, N. , De Paepe, A. et al. (2002) Accurate normalization of real‐time quantitative RT‐PCR data by geometric averaging of multiple internal control genes. Genome Biology, 3, 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vidi, P.A. , Kanwischer, M. , Baginsky, S. , Austin, J.R. , Csucs, G. , Dörmann, P. et al. (2006) Tocopherol cyclase (VTE1) localization and vitamin E accumulation in chloroplast plastoglobule lipoprotein particles. Journal of Biological Chemistry, 281, 11225–11234. [DOI] [PubMed] [Google Scholar]
- Weber, E. , Engler, C. , Gruetzner, R. , Werner, S. & Marillonnet, S. (2011) A modular cloning system for standardized assembly of multigene constructs. PLoS One, 6, e16765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu, F.‐H. , Shen, S.‐C. , Lee, L.‐Y. , Lee, S.‐H. , Chan, M.‐T. & Lin, C.‐S. (2009) Tape‐Arabidopsis sandwich ‐ a simpler Arabidopsis protoplast isolation method. Plant Methods, 5, 16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wurtzel, E.T. (2019) Changing form and function through carotenoids and synthetic biology. Plant Physiology, 179, 830–843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoo, S.‐D. , Cho, Y.‐H. & Sheen, J. (2007) Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nature Protocols, 2, 1565–1572. [DOI] [PubMed] [Google Scholar]
- Youssef, A. , Laizet, Y. , Block, M.A. , Maréchal, E. , Alcaraz, J.P. , Larson, T.R. et al. (2010) Plant lipid‐associated fibrillin proteins condition jasmonate production under photosynthetic stress. The Plant Journal, 61, 436–445. [DOI] [PubMed] [Google Scholar]
- Zbierzak, A.M. , Kanwischer, M. , Wille, C. , Vidi, P.‐A. , Giavalisco, P. , Lohmann, A. et al. (2010) Intersection of the tocopherol and plastoquinol metabolic pathways at the plastoglobule. The Biochemical Journal, 425, 389–399. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. Expression profile of FIBRILLIN (FBN) loci in pepper. Heatmap representation of Cafbn expression profile in pepper. Expression data were obtained from PepperHub (http://www.hnivr.org/), represented as FPKM‐normalised values.
Figure S2. CRISPR/Cas9 constructs for target gene editing of Slfbn genes. Scheme of final binary vectors containing the NPTII selectable marker, Cas9 driven by the 2x35S promoter and transcriptional units for gRNA expression (under control of U6 promoter). LB, left border; RB, right border.
Figure S3. Plant morphology of multiple knockout fbn mutants. (a) Representative fully expanded leaves (cv. Ailsa Craig) of 10‐weeks‐old Azy, AC and triple and quadruple fbn mutants. All plants were cultivated together in the same growth chamber. Orange arrows indicate lesions caused by fungal pathogens. Bar = 5 cm. (b) Internode length of the four equivalent internodes. (c) Maximum quantum efficiency of Photosystem II (Fv/Fm). Data (n > 5, biological replicates) are means and error bars indicate standard deviation (SD). Statistically significant differences between Azy and fbn mutant are indicated by P‐values (one‐way anova with Dunnett's post‐tests, P < 0.05).
Figure S4. Flower carotenoid composition of Cas + /Slfbn‐edited lines. T1 Slfbn‐edited lines profiling by HPLC‐PDA. Carotenoid composition and total levels of double (Cas+/fbn2a,4), triple (Cas+/fbn1,2a,4) and quadruple (Cas+/fbn1,2a,4,7a) fbn mutants. Data (n = 3, biological replicates) were analysed by one‐way anova with Dunnett's post‐test; asterisks denote statistically significant differences (*P < 0.05, **P < 0.01, ***P < 0.001) compared to AC control. Error bars indicate ±SD. D, di‐esters; F, free forms; M, mono‐esters.
Figure S5. Aberrant plastoglobuli (PG) morphology found in flower plastids of high‐order fbn mutants. Representative transmission electron micrographs (TEM) of early stage plastids (bearing thylakoids organised in grana and a few electron dense PGs) from petals from Azy and fbn mutants. Detailed view of PGs was shown on the inset panels. Giant PG formation was observed in some plastids of quadruple (fbn1,2a,4,7a) mutants. Scale bars = 500 nm (main panels), and 50 nm (inset).
Figure S6. Effect of SlFBN deficiency on tomato primary metabolism. (a) Scores plot obtained by principal component analysis (PCA) for metabolite levels measured in polar and non‐polar extracts in fruits at mature green (MG) and ripe (B7) stage, and in leaves (from top to bottom). Changes in metabolites associated with SlFBN deficiency in MG (b), ripe fruits (c) and leaves (d). Quantification was determined relative to the internal standard and values are presented as mean ± SD from four biological replicates. Only significant changes compared to respective Azy control are shown (pair‐wise t‐test corrected for multiple comparison using Holm–Sidak's post‐test; Adjusted *P < 0.05, **P < 0.01, ***P < 0.001). Full data set available in Table S6. C14:0, myristic acid; C15:0, pentadecanoic acid; C17:0, heptadecanoic acid.
Figure S7. Targeted lipidomic analysis of triacylglycerol (TAG) and its derivatives. (a, c) Heat map of top 100 features identified by anova (Post hoc Fisher's LSD, α = 0.05, corrected by multiple comparisons) in leaf and flower, respectively. Red and blue colours indicate higher and lower abundance, respectively, as measured by column standardised z‐scores derived from LC–MS peak intensity (n = 3 biological replicates). Genotypes are indicated on the right of the heatmap. The boxes on the top of the heatmap indicate the lipid classes, and whether the variable is found among the highest VIP scores (top 100) identified by OPLS‐DA. (b, d) Selected chemical features from heat maps of leaf and petal, respectively, that show statistically significant changes on fbn mutants (anova followed by Fisher's LSD, α = 0.05).
Figure S8. Relative expression of Slfbns and other genes encoding metabolic enzymes associated with PG by qPCR. Values are expression levels normalised to cac and act2 reference genes (mean ± SEM; n ≥ 3 biological replicates). Significant differences (Student's t‐test, *P < 0.05, **P < 0.01, ***P < 0.001) between fbn mutants and control are indicated.
Table S1. gRNA recognition sequences in Slfbn genes and off‐targets.
Table S2. Number of plants recovered from Agrobacterium‐mediated transformation harbouring mutations in the Slfbn target gene.
Table S3. Slfbn edited lines phenotyped in this study.
Table S4. Isoprenoid profile of fruits and leaves from fbn mutants determined by UPLC‐PDA.
Table S5. Isoprenoid profile of petals from fbn mutants determined by HPLC‐PDA.
Table S6. Metabolite levels measured in fruits (MG and B7 stage) and leaves by GC–MS.
Table S7. Proteome associated with PG fraction of tomato fruits from control and fbn mutant genotypes.
Table S8. Proteome associated with membrane fraction of tomato fruits from control and fbn mutant genotypes.
Table S9. Primers used in this study.
Table S10. Peak identification of isoprenoids analysed by HPLC‐PDA.
Data Availability Statement
Raw data files for lipidomics and proteomics are available through Mendeley data (DOI: 10.17632/9xmnybzgnd.1 and DOI: 10.17632/3hxz7c77r5.1). All other data generated or analysed during this study are included in this article (Appendix S1).