

Abstract
In each round of nuclear pre-mRNA splicing, the U4/U6·U5 tri-snRNP must be assembled from U4/U6 and U5 snRNPs, a reaction that is at present poorly understood. We have characterized a 61 kDa protein (61K) found in human U4/U6·U5 tri-snRNPs, which is homologous to yeast Prp31p, and show that it is required for this step. Immunodepletion of protein 61K from HeLa nuclear extracts inhibits tri-snRNP formation and subsequent spliceosome assembly and pre-mRNA splicing. Significantly, complementation with recombinant 61K protein restores each of these steps. Protein 61K is operationally defined as U4/U6 snRNP-specific as it remains bound to this particle at salt concentrations where the tri-snRNP dissociates. However, as shown by two-hybrid analysis and biochemical assays, protein 61K also interacts specifically with the U5 snRNP-associated 102K protein, indicating that it physically tethers U4/U6 to the U5 snRNP to yield the tri-snRNP. Interestingly, protein 61K is encoded by a gene (PRPF31) that has been shown to be linked to autosomal dominant retinitis pigmentosa. Thus, our studies suggest that disruptions in tri-snRNP formation and function resulting from mutations in the 61K protein may contribute to the manifestation of this disease.
Keywords: pre-mRNA splicing/retinitis pigmentosa/snRNP protein function/spliceosome assembly/ U4/U6·U5 tri-snRNPs
Introduction
The splicing of pre-mRNA in the nucleus is catalyzed by a large ribonucleoprotein complex, the spliceosome (for reviews see Krämer, 1996; Burge et al., 1999). The spliceosome consists of the pre-mRNA substrate and several small nuclear ribonucleoproteins (snRNPs), along with splicing factors not integrated into snRNPs. Each snRNP is a stable complex of a single RNA molecule (snRNA) and several proteins; some of these proteins are common to all snRNPs (the Sm proteins), while others are specific to a given snRNP (Will and Lührmann, 2001). The individual snRNPs interact during the splicing cycle in a highly dynamic manner. For example, at the start of the splicing cycle the U4 and U6 snRNPs are tightly associated by extensive RNA–RNA base-pairing, forming a single particle termed U4/U6. This complex associates in turn with U5 snRNP to yield the U4/U6·U5 tri-snRNP. The latter undergoes further rearrangements during the assembly of the catalytically active spliceosome. These include the partial or complete dissociation of U4 snRNP from the complex, the formation of new base-pairing between U6 and U2, and the interaction of U6 and U5 snRNAs with pre-mRNA sequences. After splicing, the spliceosome dissociates to the snRNP level, and the tri-snRNP must be re-assembled from U4, U5 and U6 snRNPs in order to take part in a new round of splicing (reviewed in Nilsen, 1998; Staley and Guthrie, 1998).
Little is known about the interactions that mediate U4/U6 and U5 snRNP association. However, protein– protein interactions are believed to play a major part in tri-snRNP formation. Indeed, the tri-snRNP particle is particularly protein-rich. In humans it contains at least 29 proteins, most of which are evolutionarily conserved, and orthologous proteins have also been identified in purified yeast tri-snRNPs (Gottschalk et al., 1999; Stevens and Abelson, 1999). However, aside from the observation of a crosslink between the yeast U5 protein Prp8p and U6 snRNA (Vidal et al., 1999), there is a paucity of information about interactions that potentially mediate the association of the U5 and U4/U6 snRNPs. In yeast, Prp24p plays a major role in the formation of the U4/U6 snRNP, thereby facilitating the regeneration of U4/U6·U5 tri-snRNPs (Raghunathan and Guthrie, 1998). However, Prp24p is under-represented, if present at all, in the yeast tri-snRNP complex (Gottschalk et al., 1999; Stevens and Abelson, 1999). Prp6p appears to be crucial for the interaction of the two snRNP subunits in the tri-snRNP. That is, mutation of the PRP6 gene inhibits tri-snRNP accumulation, while the stability and integrity of the U4/U6 and U5 snRNPs is not affected, indicating that Prp6p is solely required for tri-snRNP formation (Galisson and Legrain, 1993). Indirect evidence for a similar role in bridging U5 and U4/U6 snRNPs has recently been provided also for the human counterpart of Prp6p, i.e. the U5 snRNP-specific 102 kDa (102K) protein (Makarov et al., 2000). Very little is known, however, about the role of U4/U6-specific proteins in the formation of human tri-snRNPs, except that the 10S U4/U6 snRNP particle, lacking all U4/U6-specific proteins, does not bind to 20S U5 snRNPs (Behrens and Lührmann, 1991; Utans et al., 1992).
According to present knowledge, the following proteins have been found by biochemical separation and/or by immunoprecipitation to be associated with the human U4/U6 snRNP: (i) seven Sm proteins, which are bound to the Sm site of U4 snRNA; (ii) seven LSm (‘like-Sm’) proteins (LSm 2–8) are associated with U6 snRNA (Achsel et al., 1999; Salgado-Garrido et al., 1999); (iii) three proteins with respective molecular weights of 20, 60 and 90 kDa are present (Horowitz et al., 1997; Lauber et al., 1997; Teigelkamp et al., 1998), but the nature of their association with U4 and/or U6 snRNA has not yet been determined; and finally (iv) a 15.5 kDa protein in the HeLa tri-snRNP has been characterized recently (Nottrott et al., 1999), and is the first U4-specific protein shown to interact directly with the U4 snRNA.
In this article, we describe the molecular and functional characterization of a human tri-snRNP protein, which has an apparent molecular weight of 61 kDa and is, henceforth, termed protein 61K. Operationally we classify the 61K protein as U4/U6 snRNP-specific because it remains predominantly bound to this particle at elevated salt concentrations where the tri-snRNP dissociates into U4/U6 and U5 snRNPs. Immunodepletion and complementation studies demonstrate that protein 61K plays a crucial role in the formation of the U4/U6·U5 tri-snRNP and subsequent spliceosome formation and splicing. Moreover, our results suggest that 61K bridges U4/U6 and U5 snRNPs by specific interaction with the U5 snRNP protein 102K. Interestingly, protein 61K is the expression product of a gene (PRPF31), which was recently demonstrated to underlie autosomal dominant retinitis pigmentosa (adRP) linked to chromosome 19q13.4 (Vithana et al., 2001). Thus, the studies described here pave the way to investigate the function in tri-snRNP formation and splicing of the various 61K mutants detected in patients suffering from adRP (Vithana et al., 2001).
Results
The human tri-snRNP 61K protein is encoded by a gene linked to adRP
A putative 61K cDNA was isolated as described in Materials and methods. It encodes a protein 499 amino acids in length with a predicted molecular mass of 55.4 kDa. The complete amino acid sequence of this protein is shown in Figure 1. The cDNA encoded protein is identical to the protein encoded by the gene PRPF31 on human chromosome 19q13.4, which has recently been shown to be linked to adRP (Vithana et al., 2001; see also Discussion).

Fig. 1. The human 61K protein present in U4/U6·U5 tri-snRNPs is homologous to the yeast splicing factor Prp31p and the box C/D snoRNP proteins NOP56 and NOP58. (A) Alignment of the 61K protein sequence with S.cerevisiae Prp31p (DDBJ/EMBL/GenBank accession No. Z72876). (B) Alignment of the central part of the 61K protein encompassing amino acids 93–328 with the homologous sequences of the human NOP56 (aa 174–411, DDBJ/EMBL/GenBank accession No. Y12065) and NOP58 (aa 183–395, DDBJ/EMBL/GenBank accession No. AF123534) proteins. Sequence alignments were performed using the Clustal method.
The following criteria were used to verify that the protein obtained by translation of this cDNA was identical to the human tri-snRNP 61K protein. (i) The cDNA encoded protein contains all six peptides obtained by microsequencing the endogenous protein (see Materials and methods); the discrepancy in five of 72 amino acids is attributed to experimental error in microsequencing. (ii) Antibodies raised against a C-terminal peptide of the 61K protein, but not the corresponding pre-immune serum, specifically recognized the native 61K protein in nuclear extracts (Figure 2A, lanes 2 and 3) and in purified 25S U4/U6·U5 tri-snRNPs (lane 4). (iii) The full-length transcription/translation product of the cDNA co-migrated on SDS–polyacrylamide gels with protein 61K present in purified HeLa tri-snRNPs (Figure 2A, compare lanes 1 and 5); the lower molecular weight bands are assumed to result either from internal translation initiation or from degradation. (iv) Moreover, the translation product was efficiently immunoprecipitated by anti-61K antibodies (Figure 2A, lanes 6 and 7).

Fig. 2. (A) Confirmation of the identity of the 61K cDNA. Lane 1: proteins of purified tri-snRNPs were separated by 10% SDS–PAGE and visualized by Coomassie Blue staining. The identity of the proteins is indicated on the left. Lanes 2–4: immunodetection of protein 61K in HeLa nuclear extract and tri-snRNPs. Proteins of purified U4/U6·U5 tri-snRNPs (lane 4) or HeLa nuclear extract (lanes 2 and 3) were separated by 10% SDS–PAGE, blotted onto a membrane and immunostained with affinity-purified anti-61K antibodies (lanes 3 and 4) or corresponding pre-immune serum (NIS, lane 2). Lanes 5–7: characterization of the protein generated by in vitro translation of the 61K protein cDNA. The 61K cDNA-derived translation product, labeled with [35S]methionine was immunoprecipitated with protein A– Sepharose-bound anti-61K antibodies; the bound material was fractionated by 10% SDS–PAGE and detected by fluorography (lane 7). Lanes 5 and 6 show an aliquot of the translation reaction and a mock immunoprecipitation performed in the absence of anti-61K antibodies, respectively. The position of molecular weight markers is indicated on the right. (B) Association of protein 61K with U4/U6 snRNPs. snRNP particles from HeLa nuclear extracts were immunoprecipitated with anti-61K (lanes 2–8) or anti-116K (lanes 9–11) antibodies, at the salt concentration indicated above each lane. Precipitation with the Sm-specific antibody Y12 at 150 mM NaCl (lane 1) served as a positive control. The co-precipitated RNAs were extracted, fractionated by denaturing 10% PAGE, transferred to a membrane and hybridized with probes specific for the U1, U2, U4, U5 and U6 snRNAs. The positions of the snRNAs are indicated on the left.
Protein 61K is evolutionarily conserved and shares a homology domain with box C/D snoRNP proteins NOP56 and NOP58
Database searches with the complete amino acid sequence of the human 61K protein revealed that it is a homolog of the Prp31p protein of Saccharomyces cerevisiae (25% identity, 60% similarity) and Schizosaccharomyces pombe (68% similarity). Both proteins have been shown to be essential splicing factors in yeast (Weidenhammer et al., 1996; Bishop et al., 2000). The regions of homology are found throughout the entire sequence and are not restricted to one particular domain (Figure 1; data not shown); thus, these proteins are potentially orthologs. Moreover, homologs of protein 61K are also found in Drosophila (77% similarity, AAF49655), Caenorhabditis elegans (69% similarity, AAF60425) and Arabidopsis thaliana (56.5% similarity, AC004473).
In addition, database searches revealed a family of more distant homologs, including nucleolar proteins NOP56 and NOP58 from box C/D snoRNPs (Lyman et al., 1999). The similarity between yeast Nop56p, Nop58p and Prp31p was noted previously for a region comprising one-third of each protein (Gautier et al., 1997). In the case of the human proteins, a central domain of protein 61K (amino acids 93–328) corresponding to a half of the protein length, shares homology with the NOP proteins (57% similarity, Figure 1B). Since all of these proteins are components of RNP complexes, this raises the possibility that the conserved central domain may be an RNA binding element (see Discussion).
Association of protein 61K with the U4/U6 snRNP at high salt concentrations
Our observation that protein 61K was not present in purified U5 snRNPs (Bach et al., 1989; data not shown), implied that it initially associated with the U4/U6 snRNP or, alternatively, only with tri-snRNPs. To address this question, we carried out immunoprecipitations with nuclear extract and antibodies against 61K or against protein 116K, which is stably integrated into the U5 snRNP (Fabrizio et al., 1997). While the anti-61K antibodies precipitated the tri-snRNP complex at 150 mM NaCl, at salt concentrations >300 mM they precipitated only U4 and U6 snRNAs, and not U5 (Figure 2B, lanes 2–8). Only background levels of U1 and U2 snRNPs were co-precipitated by the anti-61K antibody in a salt independent manner (lanes 2–8). Conversely, anti-116K antibodies precipitated only U5 snRNAs at high salt concentrations (lanes 9–11). Equivalent results were obtained when the snRNA content of the immunoprecipitates was analyzed by [32P]pCp-labeling instead of northern blotting (data not shown). These data show that protein 61K remains associated with the U4/U6 snRNP, at least in part, after U5 snRNP has dissociated from the tri-snRNP complex at elevated salt concentrations.
Although the 61K protein is clearly a constituent of the tri-snRNP, this need not mean that most of the 61K in the nucleus is found in this particle. To determine what portion of protein 61K is associated with snRNPs and what portion is free, we investigated the distribution of this protein by glycerol-gradient centrifugation of nuclear extracts. After centrifugation, the gradient was fractionated, and aliquots from each fraction were analyzed for RNA (see Figure 5B, upper panel) and for protein 61K by immunoblotting with anti-61K antibodies (see Figure 5B, lower panel). Significantly, the vast majority of the 61K protein peaked together with the 25S U4/U6·U5 tri-snRNP (lanes 16–19, compare lower and upper panels) indicating that most of the 61K protein is associated with the tri-snRNP; only a minor amount of 61K migrated at the top of the gradient.

Fig. 5. Protein 61K is required for the formation of the U4/U6·U5 tri-snRNP. (A) Anti-Sm (α-Sm), anti-116K (α-116K) and anti-60K (α-60K) antibodies were used to immunoprecipitate snRNPs from mock-depleted (M), 61K-depleted (Δ) or 61K-depleted extract complemented with native protein 61K (Figure 4B) that was eluted from the anti-61K beads (Δ+). RNA was isolated from the immunoprecipitated snRNPs and characterized by northern blot analysis. (B and C) Destabilization of tri-snRNPs in the absence of protein 61K. Mock-depleted (B) or 61K-depleted (C) nuclear extract was fractionated on a 10–30% glycerol gradient (run in parallel). RNAs from each fraction were extracted, fractionated by denaturing 10% PAGE, and visualized by silver staining. The identities of the snRNAs (confirmed by northern hybridization, data not shown) are indicated on the right. For the mock-depleted extract, the distribution of protein 61K (detected by 10% SDS–PAGE, blotting onto a membrane and immunostaining with anti-61K) is shown in the lower part of (B).
Protein 61K is localized in splicing factor compartments of HeLa cells
The relatedness in sequence of protein 61K and the nucleolar NOP56 and NOP58 proteins (see Figure 1) raised the question whether protein 61K would reside exclusively in the nucleoplasm (as predicted by its association with the tri-snRNP) or whether a certain fraction would also localize in the nucleolus. We investigated the subcellular distribution of protein 61K by indirect immunofluorescence microscopy employing anti-61K antibodies. Labeling in situ of HeLa cells with the anti-61K antibody resulted in a predominantly nuclear punctate staining pattern, whereby the nucleoli and cytoplasm remained largely unstained (Figure 3A). To determine whether protein 61K is located in nuclear speckles and Cajal bodies, which have been identified as typical snRNP-containing structures (for a review see Sleeman and Lamond, 1999), we double-stained HeLa cells with monoclonal antibody Y12, which reacts with the snRNP Sm proteins (Lerner et al., 1981). As shown in Figure 3A and B, the pattern produced by anti-61K antibodies is identical to that obtained with Y12, in which 20–30 speckles, as well as Cajal bodies, are observed over a less intense diffuse staining of the nucleoplasm. As seen in an overlay image (Figure 3C), the signals from snRNPs and protein 61K indeed predominantly localize in the speckles and Cajal bodies, demonstrating that protein 61K exhibits a nuclear distribution characteristic of splicing factors in vivo and may thus be a part of snRNP-containing structures.

Fig. 3. Subcellular localization of the 61K protein in HeLa cells. HeLa cells were double-stained with affinity-purified anti-61K antibodies (A) and with monoclonal antibody Y12 (B). The red (A) and green fluorescence (B) was recorded independently and combined in an overlay image (C), leading to the yellow staining of structures decorated by both antibodies. The bars represent 10 µm.
Protein 61K is required for pre-mRNA splicing in vitro
To investigate the possible role of the 61K protein in pre-mRNA splicing, we used anti-61K antibodies to prepare immunodepleted nuclear extracts. The depletion reaction was carried out at 700 mM NaCl, i.e. under conditions where protein 61K is completely dissociated from U4/U6 snRNPs (Figure 2B and Makarov et al., 2000), to ensure that only protein 61K is depleted and not entire U4/U6 particles. As shown by immunoblotting (Figure 4A), protein 61K was efficiently removed (>95%) under these conditions. Moreover, the antibody-bound 61K could be eluted under native conditions by an excess of the antigenic peptide. Analysis of the peptide eluate (Figure 4B) showed that it contained almost exclusively protein 61K, demonstrating that the antibodies remove primarily the targeted protein from nuclear extracts. As shown in Figure 4C, both steps of the splicing reaction were strongly inhibited in the 61K-depleted extracts (lanes 6–10), with the amounts of reaction products greatly reduced compared with the mock-depleted nuclear extract (lanes 1–5). Residual splicing activity (Figure 4C, lane 10) is probably due to the small amount of protein 61K remaining in the depleted extract (Figure 4A, lane 2), which could not be removed by repeated immunodepletion steps. Complementation of the depleted extract with recombinant 61K protein completely restored splicing activity (lanes 11–15). Thus, the catalytic steps of splicing depend upon the presence of protein 61K.

Fig. 4. Pre-mRNA splicing and spliceosome formation require the presence of the 61K protein. (A and B) Nuclear extract is efficiently immunodepleted of the 61K protein. (A) Western blot of mock-depleted (lane 1) or 61K-depleted extract (lane 2) probed with anti-61K antibody. (B) Material eluted from the respective columns with antigenic peptide was analyzed by 10% SDS–PAGE and stained with Coomassie Blue. (C) The time course of splicing of 32P-labeled pre-mRNA was monitored in mock-depleted (mock) and 61K-depleted (Δ61) extracts, or 61K-depleted extract complemented with recombinant 61K protein (Δ61+61K). Reaction products were analyzed on a denaturing polyacrylamide gel, and the splicing substrate/products detected by autoradiography. The positions of the bands corresponding to the pre-mRNA, intermediates and spliced products are indicated on the left. (D) Spliceosome assembly in mock-depleted extracts (mock), 61K-depleted extracts (Δ61) and 61K-depleted extracts complemented with the recombinant 61K (Δ61+61K) was analyzed by native gel electrophoresis followed by autoradiography. The bands corresponding to the H, A, B and C complexes, as well as the gel origin, are indicated on the left.
Immunodepletion of nuclear extract of 61K protein stalls spliceosome assembly at the pre-spliceosome level
Next we examined the influence of protein 61K on spliceosome formation. For this purpose we analyzed splicing complexes by agarose gel electrophoresis under native conditions. As shown in Figure 4D, removal of protein 61K blocks the transition from the pre-spliceosome (U1 and U2 snRNPs plus pre-mRNA; complex A) to the mature spliceosome (complex B), which additionally contains the U4/U6·U5 tri-snRNP. After the addition of recombinant 61K protein to the immunodepleted splicing extract, mature spliceosomes were formed (Figure 4D), which correlates with the restoration of splicing activity.
The 61K protein is required for U4/U6·U5 tri-snRNP formation
The defect in the formation of mature spliceosomes (complex B) in 61K-depleted HeLa nuclear extract could reflect a requirement of protein 61K either for the recruitment of the tri-snRNP to the pre-spliceosome or for tri-snRNP stability. To distinguish between these possibilities, immunoprecipitations were performed with 61K-depleted or mock-depleted nuclear extract using antibodies specific for the integral U5–116K protein. Prior to immunoprecipitation, both extracts were dialyzed against D buffer containing 100 mM KCl in order to restore conditions that allow tri-snRNP formation. As expected, anti-116K antibodies efficiently precipitated re-formed U4/U6·U5 tri-snRNP from mock-depleted extracts (Figure 5A, lane 4). In contrast, exclusively U5 snRNPs were precipitated from 61K-depleted nuclear extracts (lane 5), indicating that the absence of protein 61K prevented tri-snRNPs from forming. Significantly, addition of protein 61K to the depleted extract fully restored tri-snRNP complex formation (lane 6). Thus, protein 61K is essential for the formation of the tri-snRNP complex in vitro.
The lack of stable tri-snRNP formation in the absence of protein 61K was further demonstrated by glycerol-gradient centrifugation of mock versus depleted nuclear extract (Figure 5B and C). In contrast to the mock-depleted extract where the majority of U4 and U6 snRNAs migrated as part of intact 25S tri-snRNPs (Figure 5B, fractions 16–19), most of U4/U6 sedimented as a 13S particle in the 61K-depleted extract (Figure 5C, fractions 7–10).
Several findings indicate that the removal of protein 61K from nuclear extract does not affect the stability of the accumulating U4/U6 or U5 snRNP particles. First, the Sm-specific monoclonal antibody Y12 precipitated equivalent amounts of the U4, U6 and U5 snRNPs, in addition to U1 and U2 snRNPs, irrespective of whether 61K-depleted, mock-depleted or depleted/complemented nuclear extract was used (Figure 5A, lanes 1–3). Secondly, antibodies against the U4/U6-specific 60K protein (Lauber et al., 1997) precipitated U4/U6 snRNPs from 61K-depleted extract (Figure 5A, lane 7), indicating that the U4/U6 snRNA interaction remains intact and that the binding of protein 60K and thus of the 20K/60K/90K heterotrimer (Horowitz et al., 1997; Lauber et al., 1997), is not dependent on protein 61K. Finally, glycerol-gradient centrifugation of 61K-depleted nuclear extract led to the accumulation of 20S U5 and 13S U4/U6 snRNP particles (Figure 4C). No increase in the amount of free U4 or U6 snRNP was observed after 61K-depletion. Taken together, these results demonstrate that protein 61K plays a critical and specific part in the formation of the tri-snRNP from U4/U6 and U5 snRNPs. This is a prerequisite for the integration of the tri-snRNP into spliceosomes and thus also for subsequent splicing catalysis.
Protein 61K interacts with the U5 snRNP-specific 102K protein
The 61K protein could promote tri-snRNP formation in at least two ways. First, the binding of 61K to the U4/U6 snRNP could promote a conformation of this particle compatible with docking to U5 snRNPs but without contacting directly U5 snRNP. Alternatively, protein 61K could bridge U4/U6 and U5 snRNPs by interacting with components of both U4/U6 and U5 particles. This model would predict a specific interaction of 61K with one or more U5 snRNP component. Consistent with this idea, in vitro translated 35S-labeled 61K protein binds to purified 20S U5 snRNPs, as demonstrated by co-immunoprecipitation of 61K with U5 snRNPs at low salt concentration using anti-Sm (Y12) antibodies (Figure 6A, lane 4). The interaction of 61K with the U5 snRNP is specific and required the presence of U5-specific proteins since no binding was observed with 10S U5 or U1 snRNP core particles (lanes 5 and 6, respectively) or with 12S U1 and U2 snRNPs (lane 3).
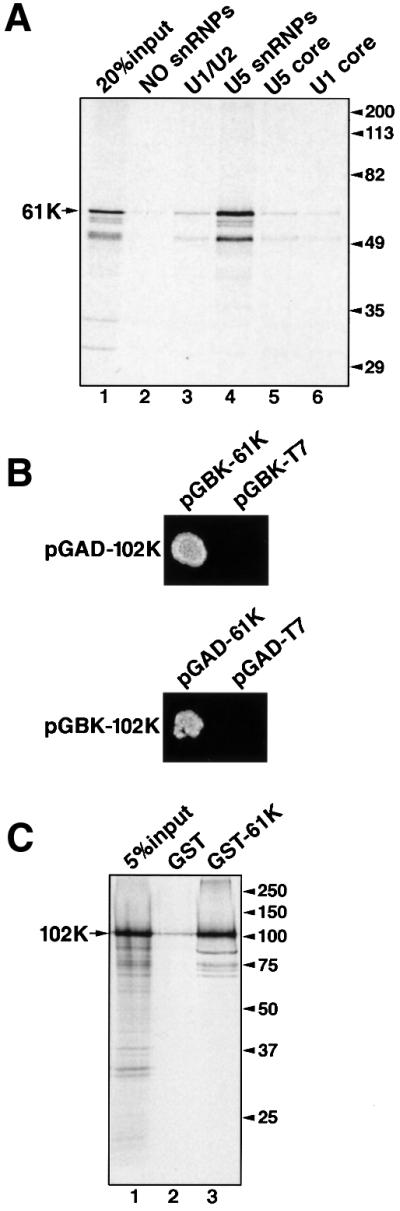
Fig. 6. (A) Protein 61K interacts with U5 snRNP. Protein 61K produced by translation in vitro was incubated with buffer only (lane 2), 12S U1 and U2 snRNPs (lane 3), U5 snRNPs (lane 4), or U5 and U1 core particles (lanes 5 and 6). The snRNP particles were then precipitated with the Sm-specific antibody Y12. The co-precipitated proteins were fractionated by SDS–PAGE and visualized by fluorography. (B) Protein 61K interacts with the U5-specific 102K protein in a yeast two-hybrid assay. The yeast strain AH109 was co-transformed with the indicated plasmid pairs and protein–protein interaction was identified on minimal medium lacking tryptophan, leucine, histidine and adenine. DNA-binding-domain vector (pGBKT7) and activation-domain vector (pGADT7) served as negative controls. (C) Co-precipitation of protein 102K with the 61K protein. 35S-labeled, in vitro translated protein 102K was incubated with GST (lane 2) or a protein 61K–GST fusion protein (lane 3). Proteins were precipitated using glutathione–Sepharose, fractionated by SDS–PAGE and visualized by fluorography.
We next searched for proteins that physically interact with the 61K protein by performing two-hybrid interaction screens with the U5-specific 15K, 40K, 52K, 100K, 102K, 116K, 200K and 220K proteins. In addition, the tri-snRNP proteins 65K and 110K, as well as the U4/U6-specific proteins 15.5K, 60K and 90K, were also included in the screen; in each case 61K was always used as a bait. Interestingly, only one out of the 13 tested proteins interacted with protein 61K, namely the U5 snRNP-specific protein 102K (Figure 6B; data not shown). This interaction was also observed in the reciprocal two-hybrid screen (with 102K as bait), indicating a tight interaction between these two proteins (Figure 6B). Physical and specific interaction between the 61K and 102K proteins was independently confirmed using a biochemical assay. In glutathione S-transferase (GST)-pulldown experiments in vitro translated, 35S-labeled 102K protein efficiently interacts with a purified recombinant fusion protein of 61K with GST (Figure 6C). This interaction is specific, as only a low level of 102K protein was precipitated with beads containing GST alone (Figure 6C). In summary, our results indicate that protein 61K functions as a bridge in the tri-snRNP, physically interacting with its U4/U6 and U5 snRNP subunits.
Discussion
Requirement of protein 61K for tri-snRNP formation and splicing
A crucial step in the assembly pathway of the spliceosomal U4/U6·U5 tri-snRNP is the interaction between the U4/U6 and U5 snRNPs. The results presented here demonstrate that the 61K protein present in tri-snRNPs is required for this step. This conclusion is corroborated by the following observations: (i) immunodepletion of 61K protein from HeLa cell nuclear extract inhibited tri-snRNP formation while U4/U6 and U5 snRNPs accumulated (Figure 5); (ii) the removal of protein 61K did not affect the stability of the U4/U6 or the U5 snRNPs as indicated by the findings that both particles retain their particle-specific proteins and that there was no increase in free U4 or U6 snRNPs or 10S U5 core snRNPs (Figure 5); and (iii) the formation of U4/U6·U5 tri-snRNPs was restored by complementation of the depleted extract with recombinant 61K protein, confirming the specificity of the effect of protein 61K depletion on tri-snRNP stability (Figure 5).
Major steps in the splicing process are the formation of spliceosome complex A, which consists of the U1 and U2 snRNPs bound to the pre-mRNA, and its subsequent transformation into complex B upon addition of the tri-snRNP. It is generally believed that the latter takes place in one step, i.e. by the binding of a pre-formed tri-snRNP to complex A (Cheng and Abelson, 1987; Konarska and Sharp, 1987; Utans et al., 1992), rather than by the sequential and/or independent addition of the U5 and U4/U6 snRNPs. The results described here provide additional direct evidence that it is indeed the pre-formed tri-snRNP that binds to complex A; in the absence of protein 61K the tri-snRNP fails to form and neither of its snRNP components becomes stably bound to complex A, as investigated by native gel electrophoresis (Figure 4D).
The function of protein 61K bears a certain resemblance to that of the U4/U6·U5 tri-snRNP 65K and 110K proteins (Makarova et al., 2001), but differs in several important aspects. As is the case for 61K, the removal of either the 65K or 110K protein prevents binding of the tri-snRNP to complex A. However, in the absence of these two proteins the stability of the tri-snRNP is unaffected. Thus, while a major function of the 61K protein is to promote U4/U6·U5 tri-snRNP formation, the 65K and 110K proteins play a major part in recruiting the pre-formed tri-snRNP to the pre-spliceosome.
The human protein 61K clearly exhibits significant similarity in sequence to the yeast Prp31p protein (Figure 1), which is also classified as a U4/U6-specific protein in yeast (Weidenhammer et al., 1997). There appears to be a mechanistic difference in the function of the two proteins, however. Evidence was presented that Prp31p mediates the binding of the pre-formed tri-snRNP to complex A (Weidenhammer et al., 1997), while our data clearly point to a crucial role for protein 61K at the earlier step of tri-snRNP formation. The conclusions drawn by Weidenhammer et al. (1997) rest to a large part on the observation that heat inactivation of extracts from a temperature-sensitive strain inhibits the assembly of mature spliceosomes while significant amounts of U4/U6·U5 tri-snRNPs were maintained. These experiments do not exclude the possibility, however, that at the non-permissive temperature the inactivated mutant Prp31p protein may induce a conformation of the tri-snRNP incompatible with docking to the pre-spliceosome, while it is still able to tether U4/U6 to U5 snRNPs. It will therefore be interesting to test whether, similar to the situation in HeLa extracts, physical depletion of Prp31p from yeast extracts prevents stable formation of the U4/U6·U5 tri-snRNP.
Evidence that protein 61K promotes tri-snRNP formation by tethering U4/U6 to U5 snRNPs
The 61K protein could facilitate the formation of the U4/U6·U5 tri-snRNP either indirectly, by promoting a conformation of the U4/U6 snRNP compatible with docking to U5 snRNP or, more directly, by physically tethering U4/U6 to U5 snRNP. The observations described here that the 61K protein specifically interacts with both U4/U6 and U5 snRNPs strongly favor a bridging role for this protein in tri-snRNP formation. A strong interaction of 61K with U4/U6 snRNPs was initially documented with anti-61K antibodies, which precipitated U4/U6·U5 tri-snRNP at low salt concentrations (where the tri-snRNP is stable), but only U4/U6 snRNPs at higher salt concentrations (where the tri-snRNP dissociates into U5 and U4/U6 snRNPs, see Figure 2B). Several lines of evidence indicate that there is also a specific binding site for protein 61K on the U5 snRNP. At low salt concentrations in vitro translated 61K protein also binds to purified 20S U5 snRNPs and this binding requires U5-specific proteins (Figure 6A). Most importantly, using two-hybrid screens and biochemical pulldown experiments we could demonstrate that 61K specifically interacts with the 102K protein of the 20S U5 snRNP in vivo and in vitro (Figure 6B and C). These data indicate that the U5–102K protein is the major interaction partner of protein 61K on the U5 snRNP subunit of the tri-snRNP.
Interestingly, independent evidence for a role of the 61K’s interaction partner U5–102K and its yeast ortholog Prp6p in bridging U5 and U4/U6 snRNPs in the tri-snRNP has been provided previously. For example, antibodies directed against the C-terminal region of the 102K protein specifically immunoprecipitated free U5 snRNPs, and not U4/U6·U5 tri-snRNPs, from HeLa nuclear extract, suggesting that in the tri-snRNP, the C-terminal region of the 102K protein is covered by U4/U6-specific proteins (Makarov et al., 2000). Consistent with a bridging function for the 102K protein, it could be shown that in vitro translated U5–102K binds to purified 13S U4/U6 snRNPs, which contained all U4/U6-specific proteins, including protein 61K (Makarov et al., 2000). In yeast, mutation of the PRP6 gene inhibits tri-snRNP accumulation, while accumulation of the individual U4/U6 and U5 snRNPs is not affected (Galisson and Legrain, 1993). This suggests that the Prp6p protein is not needed for the integrity and hence stability of the U4/U6 and U5 snRNPs, but only for tri-snRNP formation. In summary, the results described in this paper together with those cited above, indicate that the 61K protein tethers U4/U6 to U5 snRNP by interaction with the U5-associated 102K protein.
Structural similarity between U4/U6 snRNPs and box C/D snoRNPs
Sequence similarity searches also revealed significant homology between human protein 61K and the nucleolar NOP56 and NOP58 proteins of the box C/D snoRNPs. The similarity is greatest in the central region, comprising amino acids 97–328 of the 61K protein (Figure 1). Indeed, this resemblance adds to a growing body of evidence that there may be a general similarity, or at least some points of very close similarity, between the nuclear U4/U6 snRNP and nucleolar box C/D snoRNPs. Specifically, it has recently been observed that the 15.5K protein that directly binds the U4 snRNA is also associated with the box C/D snoRNAs, where it recognizes a stem–internal loop–stem structural motif almost identical to its binding site on the U4 snRNA (Nottrott et al., 1999; Watkins et al., 2000). Thus, the spliceosomal U4 snRNP and the nucleolar box C/D snoRNPs share a similar RNP core.
Mutations in protein 61K and dominant autosomal retinitis pigmentosa
Recently it has been reported that mutations in a human gene termed PRPF31, whose gene product is identical with the 61K tri-snRNP protein characterized in this present study underlie adRP on chromosome 19q13.4 (Vithana et al., 2001). The mutations detected in afflicted individuals from four families included missense substitutions, deletions and insertions in one allele of PRPF31. Thus, the pathophysiological basis of adRP linked to the PRPF31 gene is the functional loss of one allele resulting in haplo-insufficiency. As most of the observed mutations would lead to severely truncated 61K protein variants, it is likely that these mutants would no longer promote tri-snRNP formation with the result that the concentration of functional tri-snRNP in the cell would decrease by ∼50%. Since we have shown in this present study that inhibition of U4/U6·U5 tri-snRNP formation—due to the absence of 61K protein—abolishes pre-mRNA splicing, the capacity of cells expressing low amounts of functional 61K protein to perform pre-mRNA splicing would be severely compromised. In view of the crucial role that protein 61K plays in tri-snRNP formation and thus splicing (this paper), as well as the fact that the PRPF31 gene is ubiquitously expressed (Vithana et al., 2001), the question remains why mutations in this gene would produce a disease phenotype only in rod photoreceptors. As pointed out by Vithana et al. (2001), rod cells have a particularly high demand for spliced opsin mRNA, and thus these cells may be particularly sensitive to a reduced level of functional tri-snRNPs and thus a kinetically compromised splicing machinery.
The studies described here pave the way to investigate the functional activity in tri-snRNP formation and splicing of the various 61K mutants detected by Vithana et al. (2001) in patients suffering from adRP. Such studies should also allow us to determine the effects of different mutations on 61K protein function. Finally, once the interaction of the 61K protein with the U4/U6 and U5 snRNP is understood at the molecular level, it may be possible to restore the function of certain 61K mutants expressed in patients with adRP in a rational way.
Materials and methods
cDNA cloning of protein 61K
Twenty-five S U4/U6·U5 tri-snRNPs were isolated from HeLa nuclear extracts according to Laggerbauer et al. (1996). Proteins were extracted and fractionated by SDS–PAGE. Partial amino acid sequences of tryptic peptides of protein 61K were determined as described previously (Lauber et al., 1996). The peptide sequences obtained were: RFPELESLVPNALDYIRTVK, YFSEMAEFLERK, AEFLKVKGEKSGLM, MKIFEYISGQAKASE, GVAGGLTNLSK, FAEI (amino acids shown in bold type deviate from the sequence of the cDNA translation product). Database searches at the National Center for Biotechnology Information (NCBI) using these peptide sequences yielded several expressed tag sequences (ESTs), of which R87935 and AA101050 from human cDNA libraries, and AA087405 from a mouse cDNA library, were obtained commercially from RZPD (Berlin) and sequenced. As the human ESTs contained only partial cDNA sequences, two ESTs were ligated together using a unique restriction site in their overlapping regions to produce a full-length cDNA. The cDNA sequences of human and mouse protein 61K have been submitted to the DDBJ/EMBL/GenBank database under accession Nos AY040822 and AY040823, respectively.
Antibodies and production of recombinant protein 61K
Anti-61K antibodies were raised in a rabbit against a C-terminal peptide (amino acid residues 484–497). The antibodies were affinity purified on a SulfoLink column containing the immobilized peptide as described by the manufacturer (Pierce) and diluted 1:1000 for western blot analysis.
The full-length coding region of protein 61K was cloned into the pET-28a expression vector (Novagen) and N-terminally His-tagged protein 61K was expressed in Escherichia coli. The 61K protein was found in inclusion bodies that were washed with B-PERTM Bacterial Protein Extraction Reagent (Pierce). The protein was solubilized in the presence of 6 M guanidine hydrochloride and refolded with the non-detergent sulfobetaine, NDSB-201 (Calbiochem) (Vuillard et al., 1998). It was subsequently purified on a Ni-NTA column (Qiagen) under native conditions (50 mM Tris–HCl pH 7.8, 250 mM NaCl), dialyzed against buffer containing 20 mM HEPES–KOH pH 7.9, 250 mM KCl, 1.5 mM MgCl2, 0.5 mM dithiothreitol, 10% glycerol, and stored at –70°C at a concentration of 0.1 mg/ml.
Immunoprecipitation, immunodepletion and in vitro splicing
Immunoprecipitations were performed with nuclear extract or purified snRNPs and antibodies raised against protein 61K, the U5-specific 116K protein (Fabrizio et al., 1997) or the Sm-proteins (monoclonal antibody Y12; kindly provided by J.Steitz, Yale University) essentially as described previously (Makarova et al., 2001). Co-immunoprecipitated RNAs were analyzed by northern blot hybridization with 32P-labeled probes generated from plasmids encoding individual snRNAs, as described previously (Fabrizio et al., 1989). 35S-labeled protein 61K was generated by in vitro translation of the 61K cDNA using a TNT–T3 coupled reticulocyte system as described by the manufacturer (Promega). For immunodepletion of HeLa nuclear extracts, the salt concentration of 400 µl extract (stored in C buffer; Dignam et al., 1983) was first adjusted to 700 mM NaCl. The extract was then incubated with 60 µl of protein A–Sepharose charged with affinity-purified antibodies for 2 h at 4°C with end-over-end rotation. Beads with bound material were removed by centrifugation and the resulting depleted nuclear extract was then dialyzed for 4 h against D buffer (Dignam et al., 1983) in a Slide-A-Lyzer 3.5K (Pierce). The bound 61K protein was eluted from the protein A–Sepharose in 200 µl of D buffer containing 0.5 mg/ml of the antigenic peptide. Mock-depleted extracts were treated in a manner identical to that of the depleted extracts, except that protein A beads were either pre-blocked with bovine serum albumin and tRNA, or incubated with pre-immune serum; both methods yielded very similar results.
Analysis of pre-mRNA splicing was performed as described by Makarova et al. (2001). For complementation studies, 2 µg of native or recombinant 61K protein were added to 125 µl of depleted extract at 700 mM NaCl. Two micrograms of recombinant 61K protein correspond approximately to the amount of endogenous 61K protein in 125 µl of extract. The latter value was determined by quantitating on SDS gels the 61K protein which was affinity purified from a defined amount of nuclear extract and retrieved from the anti-61K affinity column by elution with the antigenic peptide. The extracts were dialyzed for 4 h against D buffer and then added to the splicing reaction. Alternatively, protein 61K was added to the dialyzed extract to a final concentration of 0.32 µM and the mixture was incubated at 0°C for 10 min before addition of pre-mRNA. For analysis of spliceosomal complexes, 5 µl aliquots of the splicing reaction were mixed with 1 µl of heparin (5 mg/ml) at defined time intervals and placed on ice. Splicing complexes were analyzed by 2% agarose gel electrophoresis according to Das and Reed (1999).
Glycerol-gradient fractionation of nuclear extract
After dialysis, mock- and 61K-depleted extracts were diluted 1:1 with gradient buffer (20 mM HEPES–KOH pH 7.9, 150 mM NaCl, 1.5 mM MgCl2, 0.5 mM dithiothreitol) and loaded onto a linear 4 ml 10–30% (w/v) glycerol gradient. After centrifugation for 14 h at 29 000 r.p.m. in a Beckman SW60 rotor, the gradient was fractionated manually from the top. RNAs were extracted from gradient fractions with phenol– chloroform, precipitated and analyzed by denaturing 10% PAGE followed by silver staining. Proteins were subjected to 10% SDS–PAGE followed by blotting and immunostaining with anti-61K antibody.
Indirect immunofluorescence
Immunolocalization of the 61K protein was performed as described previously (Makarov et al., 2000). HeLa cells were grown on cover slips and fixed for 10 min with 2% (w/v) paraformaldehyde (Merck) in phosphate-buffered saline (PBS). The cells were rendered permeable by incubation for 5 min in PBS with 0.2% Triton X-100 (Sigma). The primary antibodies, affinity-purified anti-61K and monoclonal Y12, were used at 1:20 and 1:1000 dilution, respectively. The secondary antibodies, Cy3-conjugated goat anti-rabbit IgG (Amersham) and fluorescein isothiocyanate (FITC)-conjugated sheep anti-mouse (Sigma), were diluted to a final concentration of 0.002 mg/ml, respectively. Images were recorded with a Zeiss confocal laser scanning microscope with excitation wavelength of 488 nm (for FITC) and 543 nm (for Cy3).
Two-hybrid analysis and GST-pulldown
Protein–protein interaction was analyzed using the MATCHMARKER-III Two-Hybrid System (Clontech). The full-length coding regions of proteins 61K and 102K were subcloned into DNA binding domain vector pGBKT7 and activation domain vector pGADT7, resulting in pGBKp61, pGBKp102, pGADp61, pGADp102, respectively. Plasmid pairs were co-transformed into the yeast strain AH109 by the lithium acetate method and transformants were selected on minimal synthetic dropout (SD) medium lacking tryptophan and leucine at 30°C for 3–4 days. For analysis of interaction, each transformant was replicated on SD medium lacking tryptophan, leucine, histidine and adenine and allowed to grow for 7 days at 30°C.
To confirm the interaction detected by two hybrid analysis, a GST-pulldown experiment was performed as described previously (Einarson, 2001). Aliquots of the in vitro translated 35S-labeled protein 102K were incubated in 400 µl of TBST buffer (20 mM Tris pH 8.0, 150 mM NaCl, 0.1% Triton X-100) with either a recombinant fusion protein of 61K and GST or recombinant GST alone. After incubation for 2 h at 4°C, glutathione–Sepharose was added to the mixtures and incubated for another 2 h. Sepharose was recovered by centrifugation, washed, and the bound material was eluted in SDS sample buffer and analyzed by SDS–PAGE.
Acknowledgments
Acknowledgements
We thank P.Kempkes for HeLa cell preparation and I.Öchsner and A.Badouin for U snRNP preparation. We also thank S.Nottrott for advice on the purification of recombinant protein 61K and C.L.Will for helpful comments and discussions on the manuscript. We are grateful to the Resource Centre of the German Genome Project (RZPD) at the Max Planck Institute of Molecular Genetics for providing EST clones. This work was supported by the Gottfried Wilhelm Leibniz Program and grants from the Deutsche Forschungsgemeinschaft (SFB 397, SFB 523) and Fonds der Chemischen Industrie (to R.L.).
References
- Achsel T., Brahms,H., Kastner,B., Bachi,A., Wilm,M. and Lührmann,R. (1999) A doughnut-shaped heteromer of human Sm-like proteins binds to the 3′-end of U6 snRNA, thereby facilitating U4/U6 duplex formation in vitro. EMBO J., 18, 5789–5802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bach M., Winkelmann,G. and Lührmann,R. (1989) 20S small nuclear ribonucleoprotein U5 shows a surprisingly complex protein composition. Proc. Natl Acad. Sci. USA, 86, 6038–6042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrens S.E. and Lührmann,R. (1991) Immunoaffinity purification of a [U4/U6·U5] tri-snRNP from human cells. Genes Dev., 5, 1439–1452. [DOI] [PubMed] [Google Scholar]
- Bishop D.T., McDonald,W.H., Gould,K.L. and Forsburg,S.L. (2000) Isolation of an essential Schizosaccharomyces pombe gene, prp31+, that links splicing and meiosis. Nucleic Acids Res., 28, 2214–2220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burge C.B., Tuschl,T. and Sharp,P.A. (1999) Splicing of precursors to mRNAs by the spliceosomes. In Gesteland,R.F. and Atkins,J.F. (eds), The RNA World. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 525–560.
- Cheng S.C. and Abelson,J. (1987) Spliceosome assembly in yeast. Genes Dev., 1, 1014–1027. [DOI] [PubMed] [Google Scholar]
- Das R. and Reed,R. (1999) Resolution of the mammalian E complex and the ATP-dependent spliceosomal complexes on native agarose mini-gels. RNA, 5, 1504–1508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dignam J.D., Lebovitz,R.M. and Roeder,R.G. (1983) Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res., 11, 1475–1489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Einarson M.B. (2001) Detection of protein–protein interactions using the GST fusion protein pulldown technique. In Sambrook,J. and Russel,D.W. (eds), Molecular Cloning, a Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 18.55–18.59.
- Fabrizio P., McPheeters,D.S. and Abelson,J. (1989) In vitro assembly of yeast U6 snRNP: a functional assay. Genes Dev., 3, 2137–2150. [DOI] [PubMed] [Google Scholar]
- Fabrizio P., Laggerbauer,B., Lauber,J., Lane,W.S. and Lührmann,R. (1997) An evolutionarily conserved U5 snRNP-specific protein is a GTP-binding factor closely related to the ribosomal translocase EF-2. EMBO J., 16, 4092–4106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galisson F. and Legrain,P. (1993) The biochemical defects of prp4-1 and prp6-1 yeast splicing mutants reveal that the PRP6 protein is required for the accumulation of the [U4/U6·U5] tri-snRNP. Nucleic Acids Res., 21, 1555–1562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gautier T., Berges,T., Tollervey,D. and Hurt,E. (1997) Nucleolar KKE/D repeat proteins Nop56p and Nop58p interact with Nop1p and are required for ribosome biogenesis. Mol. Cell. Biol., 17, 7088–7098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gottschalk A., Neubauer,G., Banroques,J., Mann,M., Lührmann,R. and Fabrizio,P. (1999) Identification by mass spectrometry and functional analysis of novel proteins of the yeast [U4/U6·U5] tri-snRNP. EMBO J., 18, 4535–4548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horowitz D.S., Kobayashi,R. and Krainer,A.R. (1997) A new cyclophilin and the human homologues of yeast Prp3 and Prp4 form a complex associated with U4/U6 snRNPs. RNA, 3, 1374–1387. [PMC free article] [PubMed] [Google Scholar]
- Konarska M.M. and Sharp,P.A. (1987) Interactions between small nuclear ribonucleoprotein particles in formation of spliceosomes. Cell, 49, 763–774. [DOI] [PubMed] [Google Scholar]
- Krämer A. (1996) The structure and function of proteins involved in mammalian pre-mRNA splicing. Annu. Rev. Biochem., 65, 367–409. [DOI] [PubMed] [Google Scholar]
- Laggerbauer B., Lauber,J. and Lührmann,R. (1996) Identification of an RNA-dependent ATPase activity in mammalian U5 snRNPs. Nucleic Acids Res., 24, 868–875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lauber J., Fabrizio,P., Teigelkamp,S., Lane,W.S., Hartmann,E. and Lührmann,R. (1996) The HeLa 200 kDa U5 snRNP-specific protein and its homologue in Saccharomyces cerevisiae are members of the DEXH-box protein family of putative RNA helicases. EMBO J., 15, 4001–4015. [PMC free article] [PubMed] [Google Scholar]
- Lauber J., Plessel,G., Prehn,S., Will,C.L., Fabrizio,P., Gröning,K., Lane,W.S. and Lührmann,R. (1997) The human U4/U6 snRNP contains 60 and 90kD proteins that are structurally homologous to the yeast splicing factors Prp4p and Prp3p. RNA, 3, 926–941. [PMC free article] [PubMed] [Google Scholar]
- Lerner E.A., Lerner,M.R., Janeway,C.A.,Jr and Steitz,J.A. (1981) Monoclonal antibodies to nucleic acid-containing cellular constituents: probes for molecular biology and autoimmune disease. Proc. Natl Acad. Sci. USA, 78, 2737–2741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyman S.K., Gerace,L. and Baserga,S.J. (1999) Human Nop5/Nop58 is a component common to the box C/D small nucleolar ribonucleoproteins. RNA, 5, 1597–1604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makarov E.M., Makarova,O.V., Achsel,T. and Lührmann,R. (2000) The human homologue of the yeast splicing factor Prp6p contains multiple TPR elements and is stably associated with the U5 snRNP via protein–protein interactions. J. Mol. Biol., 298, 567–575. [DOI] [PubMed] [Google Scholar]
- Makarova O.V., Makarov,E.M. and Lührmann,R. (2001) The 65 and 110 kDa SR-related proteins of the U4/U6·U5 tri-snRNP are essential for the assembly of mature spliceosomes. EMBO J., 20, 2553–2563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nilsen T.W. (1998) RNA–RNA interactions in nuclear pre-mRNA splicing. In Simons,R.W. and Grunberg-Manago,M. (eds), RNA Structure and Function. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 279–307.
- Nottrott S., Hartmuth,K., Fabrizio,P., Urlaub,H., Vidovic,I., Ficner,R. and Lührmann,R. (1999) Functional interaction of a novel 15.5kD [U4/U6·U5] tri-snRNP protein with the 5′ stem–loop of U4 snRNA. EMBO J., 18, 6119–6133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raghunathan P.L. and Guthrie,C. (1998) A spliceosomal recycling factor that reanneals U4 and U6 small nuclear ribonucleoprotein particles. Science, 279, 857–860. [DOI] [PubMed] [Google Scholar]
- Salgado-Garrido J., Bragado-Nilsson,E., Kandels-Lewis,S. and Seraphin, S. (1999) Sm and Sm-like proteins assemble in two related complexes of deep evolutionary origin. EMBO J., 18, 3451–3462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sleeman J.E. and Lamond,A.I. (1999) Nuclear organization of pre-mRNA splicing factors. Curr. Opin. Cell Biol., 11, 372–377. [DOI] [PubMed] [Google Scholar]
- Staley J.P. and Guthrie,C. (1998) Mechanical devices of the spliceosome: motors, clocks, springs and things. Cell, 92, 315–326. [DOI] [PubMed] [Google Scholar]
- Stevens S.W. and Abelson,J. (1999) Purification of the yeast U4/U6·U5 small nuclear ribonucleoprotein particle and identification of its proteins. Proc. Natl Acad. Sci. USA, 96, 7226–7231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teigelkamp S., Achsel,T., Mundt,C., Gothel,S.F., Cronshagen,U., Lane,W.S., Marahiel,M. and Lührmann,R. (1998) The 20kD protein of human [U4/U6·U5] tri-snRNPs is a novel cyclophilin that forms a complex with the U4/U6-specific 60kD and 90kD proteins. RNA, 4, 127–141. [PMC free article] [PubMed] [Google Scholar]
- Utans U., Behrens,S., Lührmann,R., Kole,R. and Krämer,A. (1992) A splicing factor that is inactivated during in vivo heat shock is functionally equivalent to the [U4/U6·U5] triple snRNP-specific proteins. Genes Dev., 6, 631–641. [DOI] [PubMed] [Google Scholar]
- Vidal V.P., Verdone,L., Mayes,A.E. and Beggs,J.D. (1999) Characterization of U6 snRNA–protein interactions. RNA, 5, 1470–1481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vithana E.N. et al. (2001) A human homolog of yeast pre-mRNA splicing gene, PRP31, underlies autosomal dominant retinitis pigmentosa on chromosome 19q13.4 (RP11). Mol. Cell, 8, 375–381. [DOI] [PubMed] [Google Scholar]
- Vuillard L., Rabilloud,T. and Goldberg,M.E. (1998) Interactions of non-detergent sulfobetaines with early folding intermediates facilitate in vitro protein renaturation. Eur. J. Biochem., 256, 128–135. [DOI] [PubMed] [Google Scholar]
- Watkins N.J. et al. (2000) A common core RNP structure shared between the small nucleoar box C/D RNPs and the spliceosomal U4 snRNP. Cell, 103, 457–466. [DOI] [PubMed] [Google Scholar]
- Weidenhammer E.M., Singh,M., Ruiz-Noriega,M. and Woolford,J.L.,Jr (1996) The PRP31 gene encodes a novel protein required for pre-mRNA splicing in Saccharomyces cerevisiae. Nucleic Acids Res., 24, 1164–1170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weidenhammer E.M., Ruiz-Noriega,M. and Woolford,J.L.,Jr (1997) Prp31p promotes the association of the U4/U6·U5 tri-snRNP with prespliceosomes to form spliceosomes in Saccharomyces cerevisiae. Mol. Cell. Biol., 17, 3580–3588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Will C.L. and Lührmann,R. (2001) Spliceosomal UsnRNP biogenesis, structure and function. Curr. Opin. Cell Biol., 13, 290–301. [DOI] [PubMed] [Google Scholar]