

Abstract
Streptococcus equi subsp. equi is the causative agent of strangles, a disease of the upper respiratory tract in horses. The initiation of S. equi subsp. equi infection is likely to involve cell surface-anchored molecules mediating bacterial adhesion to the epithelium of the host. The present study describes the cloning and characterization of FNEB, a fibronectin-binding protein with cell wall-anchoring motifs. FNEB can thus be predicted as cell surface located, contrary to the two previously characterized fibronectin-binding proteins in S. equi subsp. equi, FNE and SFS. Assays of antibody titers in horses and in experimentally infected mice indicate that the protein is immunogenic and expressed in vivo during S. equi subsp. equi infection. Using Western ligand blotting, it was shown that FNEB binds to the N-terminal 29-kDa fragment of fibronectin, while SFS and FNE both bind to the adjacent 40-kDa fragment. S. equi subsp. equi is known to bind fibronectin to a much lower degree than the closely related S. equi subsp. zooepidemicus, but the binding is primarily directed to the 29-kDa fragment. Inhibition studies using S. equi subsp. equi cells indicate that FNEB mediates cellular binding to fibronectin in this species.
Streptococcus equi, a beta-hemolytic horse pathogen belonging to the Lancefield group C, includes three subspecies, S. equi subsp. equi, S. equi subsp. zooepidemicus, and the recently described S. equi subsp. ruminatorum (3). Infection by S. equi subsp. equi causes strangles, a serious and highly contagious disease of the upper respiratory tract of the horse (23). S. equi subsp. zooepidemicus is considered an opportunistic commensal, often occurring in the upper respiratory tract of healthy horses, but it can also cause disease, e.g., in the uterus and in wounds. While S. equi subsp. equi is essentially confined to equids, S. equi subsp. zooepidemicus has also been found to occur in a wide range of other animals and in humans.
Some of the factors that are assumed to be important in the virulence of S. equi subsp. equi include the hydrophobic antiphagocytic capsule (1), the M-like proteins SeM and SzPSe (14, 24), secreted toxins such as streptolysin S (4), and at least four pyrogenic mitogens (2, 19). The initiation of S. equi infection is likely to involve several surface-anchored proteins (adhesins) binding to the tonsil epithelium of the host. Adhesins that could contribute to these interactions include the fibrinogen-binding proteins SzPSe and SeM; the immunoglobulin G (IgG)-, serum albumin-, and α2-macroglobulin-binding protein ZAG (10); the collagen-binding protein CNE (7); and the collagen-like protein SclC (6).
A group of bacterial adhesins that have received much attention are proteins targeting fibronectin (Fn), a glycoprotein found in the extracellular matrix and body fluids of vertebrates. These proteins are found in Streptococcus pyogenes (SfbI/F1), Staphylococcus aureus (FnBPA and FnBPB), Streptococcus dysgalactiae (FnBA and FnBB), and other bacterial species (20). In S. pyogenes and S. aureus, these Fn-binding proteins have been reported to mediate the invasion of host cells without the need for other bacterial factors (17, 22).
So far, two Fn-binding extracellular proteins of S. equi subsp. equi have been reported, FNE (11) and SFS (8). Since neither of these contains cell wall-anchoring motifs and FNE has been found secreted in growth media, they are not likely to contribute to bacterial adherence.
In the present study, we describe a novel protein called FNEB, containing conserved Fn-binding repeats and cell wall-anchoring motifs. Furthermore, the binding specificities of FNEB, FNE, and SFS are analyzed, and the immunological responses in horses to the different Fn-binding proteins are compared.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions.
S. equi subsp. equi strain 1866 was obtained from NordVacc Läkemedel AB, Stockholm, Sweden, and strain DSM 20561 was obtained from DSM, Braunschweig, Germany. Other S. equi subsp. equi (n = 6) and S. equi subsp. zooepidemicus (n = 10) strains used in this study were obtained from the National Veterinary Institute (SVA), Uppsala, Sweden. The Escherichia coli strain ER2566 and the plasmid vector pTYB4 were obtained from New England Biolabs Inc. (NEB), MA. Streptococcal strains were grown on horse blood agar plates or in Todd-Hewitt broth (Oxoid, Basingstoke, Hampshire, United Kingdom) supplemented with 0.5% yeast extract. E. coli was cultured in Luria-Bertani broth supplemented with ampicillin (100 μg ml−1) or on LAA plates (Luria-Bertani broth with ampicillin and agar [15 g liter −1]). Incubations were at 37°C unless otherwise stated.
Proteins, sera, and reagents.
Bovine serum Fn was obtained from Sigma, Steinheim, Germany. Horse sera were obtained from the Swedish Veterinary Institute (SVA), Uppsala, Sweden, and NordVacc, Stockholm, Sweden. The NEB IMPACTT7 system was used to produce and purify recombinant FNEB proteins.
Protein SFS (GenBank accession number AF136451) from S. equi subsp. equi has previously been described (8). Protein FNE (GenBank accession number AF360373) from S. equi subsp. equi and protein FNZ (GenBank accession number X99995) from S. equi subsp. zooepidemicus have also previously been described (9, 11). The production of the N-terminal half (amino acids 32 to 337) of FNZ, in this study called FNE, is described in reference 11.
Chymotryptic fragments of Fn, corresponding to the N-terminal 29-kDa fibrin-binding domain, the 40-kDa collagen-binding domain, and the 105-kDa integrin-binding domain, were isolated as described previously (18).
125I was obtained from Amersham Biosciences AB, Uppsala, Sweden, and used to label whole bovine Fn and the three Fn fragments according to the Iodo-Beads labeling method described in the manual provided by the manufacturer (Pierce, Rockford, IL).
DNA sequencing and similarity studies.
The nucleotide sequences of the inserts in pFNEB S and pFNEB L were determined using a DYEnamic ET terminator cycle sequencing premix kit, a model 377 Perkin-Elmer DNA sequencer, and software from the Vector NTI suite (Informax, Bethesda, MD). The NCBI BLAST2 program (www.ncbi.nlm.nih.gov/BLAST/bl2seq/bl2.html) was used to analyze sequence similarities. To analyze the structure and properties of FNEB, the following web-based tools were used: ProtParam (us.expasy.org/tools/protparam.html), DAS (www.sbc.su.se/∼miklos/DAS/), and SignalP (www.cbs.dtu.dk/services/SignalP/).
Construction of clones and purification of recombinant proteins.
To express and purify FNEB, two different constructs were made, pFNEB S, encoding amino acids 36 to 237 (23 kDa), and pFNEB L, encoding amino acids 36 to 396 (40 kDa) of FNEB. The two constructs were made as follows. Primer OFE7:5, 5′-CATGCCATGGAGCAGTATTACGGGTGGAGTGAC-3′, combined with primer OFE8:3, 5′-CCGCTCGAGAGGCTCTTCGGGAACAATAATTGA-3′ (pFNEB S), or with primer OFE4:3, 5′-CCGCTCGAGATGGAATTCCAGCCTAGGAC-3′ (pFNEB L), was used to PCR amplify the corresponding DNA fragments using S. equi subsp. equi strain 1866 DNA as a template. The nucleotide sequences of the primers that hybridize to the fneB gene are underlined, and the introduced restriction endonuclease cleavage sites (NcoI and XhoI) are in bold. Both PCR amplifications were performed using ReadyToGo PCR beads (Amersham) with the following program repeated for 25 cycles: step 1, 95°C, 30 s; step 2, 46°C, 15 s; and step 3, 72°C, 2 min. The PCR products were digested with NcoI and XhoI and ligated into the pTYB4 vector (NEB) previously digested with the same restriction endonucleases. After ligation, the plasmids were electrotransformed into E. coli ER2566 and spread on LAA plates. Transformants were transferred to nitrocellulose (NC) membranes, and expression of the FNEB constructs was induced by incubation on LAA plates with 0.4 mM IPTG (isopropyl-β-d-thiogalactopyranoside). The pFNEB L clone was identified by colony screening using horse sera and horseradish peroxidase (HRP)-labeled anti-Fn IgG (Dako Cytomation, Glostrup, Denmark). The inserts of pFNEB L and pFNEB S were sequenced, and the clones were used to produce recombinant proteins according to the manufacturer's (NEB) recommendations.
A clone of the sfs gene, encoding the full-length mature protein, was made and used to express recombinant proteins according to the same procedure. The following primers were used: OSFS1:5, 5′-CTGGCCATGGCACTTGTCTTTGGAGGTTTA-3′, and OSFS2:3, 5′-CAGACTCGAGGTCGGGATTGTAAGAATAG-3′.
Reverse transcription (RT)-PCR.
Streptococcal cells were grown to an optical density at 600 nm (OD600) of 0.6 and then harvested, and total RNA was prepared using the FastRNA Pro Blue kit (Qbiogene, CA). The preparation was DNase treated and purified using an RNeasy mini kit (QIAGEN, West Sussex, United Kingdom). First-strand cDNAs of the fneB, fne, sfs, and gyrA transcripts were synthesized with SuperScript III (Invitrogen, Carlsbad CA) according to the procedure specified by the manufacturer. One microgram of total RNA was used as a template with the following primers: FNEB R, 5′-TCAAGATCATTCTGAGGCTCTTC-3′; FNE R, 5′-CCTCCATGGAAACCTATAAGTCC-3′; SFS R, 5′-GCCTGATGCTGATGATTAGTAGG-3′; and GYR R, 5′-ATCAATTCTGCTTGTGCGATAAT-3′. In the PCR, the abovementioned gene-specific primers were used in combination with FNEB F (5′-AAAAGTCAAGCGCTAAAGGAGTT-3′), FNE F (5′-GGGCATGACCATATGAAAGATTA-3′), SFS F (5′-TGCACTTGTCTTTGGAGGTTTAT-3′), and GYR F (5′-GACTCAATTTGACAAGGACAAGG-3′), in the following program repeated for 22 to 30 cycles: step 1, 95°C, 30 s; step 2, 46°C, 15 s; and step 3, 72°C, 1 min.
PCR products were analyzed by gel electrophoresis and sequenced. The RNA preparation was confirmed to be DNA free by running PCRs without the RT step for 35 cycles.
HiTrap affinity column purification of FNEB ligands.
FNEB L (0.5 mg) was coupled to a 1-ml HiTrap NHS-activated high-performance column according to the instructions provided by the manufacturer (Amersham). Horse serum (20 ml) was filtered through a 0.45-μm filter and then passed through the column. The column was washed with phosphate-buffered saline (PBS) supplemented with Tween 20 (0.05%, vol/vol) (PBS-T), followed by washing with PBS before bound ligands were eluted in 5 ml of 0.5 M glycine-HCl (pH 2.0). The pH of the 0.5-ml fractions was neutralized using Tris-HCl (pH 8.0).
SDS-PAGE, Western ligand blot analysis, and Western blot analysis.
The purified recombinant proteins FNEB L, FNEB S, SFS, and FNE were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) analysis under reduced conditions using the PhastSystem (Amersham) with precast 8-to-25% gradient gels. To analyze the Fn-binding specificity, the proteins were diffusion blotted onto an NC membrane for 30 min at 65°C. The NC membrane was subsequently blocked with PBS-T and casein (0.5%, wt/vol) for 1 h at room temperature. The NC membrane was incubated with 125I-labeled Fn fragments overnight at 4°C. After extensive washing with PBS-T, the NC membrane was exposed to Biomax MS (Kodak) film for 48 h.
A 2-μl sample of the eluted material from the HiTrap affinity column purification was analyzed by Western blotting, using HRP-labeled anti-Fn IgG (Dako Cytomation, Glostrup, Denmark).
Binding and inhibition analysis.
The ability of FNEB L, SFS, and FNE in solution to inhibit the binding of the iodinated 29-kDa fragment to immobilized FNEB L was studied. Microtiter wells (Immulon 2 Removawell; Dynatech Laboratories Inc.) were coated with 100 μl FNEB L (4 μg ml−1) in 0.05 M NaHCO3 (pH 9.7) for 1 h at room temperature. After being washed with PBS-T, the wells were blocked for 30 min with casein (0.1%) in PBS-T. FNEB L, SFS, and FNE (approximately 0.01 to 10 μg ml−1) were incubated with the 125I-labeled 29-kDa fragment (∼7,000 cpm) for 1 h at room temperature. The mixed samples were added to the coated wells and incubated for 1 h at room temperature. After the wells were washed with PBS-T, the radioactivity bound to the wells was measured in a γ-counter (LKB Wallac, Turku, Finland). In the same type of experiment, FNEB L, SFS, and FNE were tested for their ability to inhibit the binding of the iodinated 40-kDa fragment (∼7,000 cpm) to wells coated with SFS (6.5 μg ml−1) or FNE (5.5 μg ml−1).
The ability of streptococcal cells to bind whole Fn, the 29-kDa fragment, and the 40-kDa fragment was measured. Streptococcal cells were grown to an OD600 of 0.6 and then harvested and washed with PBS. Cells (∼106 CFU) were incubated with iodinated Fn (∼16,000 cpm), the 29-kDa fragment (∼17,000 cpm), or the 40-kDa fragment (∼11,000 cpm) in 1 ml PBS-T with casein (0.1%, wt/vol) for 2 hours using an end-over-end mixer and then pelleted and washed twice in PBS-T. The radioactivity bound to the cells was measured in a γ-counter. FNEB L was also used to inhibit the binding of the iodinated 29-kDa fragment to S. equi subsp. equi cells. A fixed concentration of the iodinated 29-kDa fragment (∼16,000 cpm) was mixed with dilutions of FNEB L (or FNEB S as a negative control) and casein (0.1%, wt/vol) to achieve a volume of 200 μl. After 1 h of incubation using an end-over-end mixer, 300 μl cells (total, ∼106 CFU) was added. After 2 h of incubation, the cells were pelleted and washed, and the radioactivity was measured. The same experiment was performed using iodinated whole Fn (∼22,000 cpm). Here, 300 μl cells corresponding to 3 × 106 CFU was added for the second incubation.
Determination of titers of antibody from mice and horses by enzyme-linked immunosorbent assay (ELISA).
Sera from NMRI mice (n = 10) were taken before and after an experimental S. equi subsp. equi infection that had proceeded for 2 weeks (5). Sera were taken from horses with clinical signs of strangles and positive cultivation of S. equi subsp. equi (n = 10) and from horses without any previous or present signs of strangles (n = 16).
Microtiter wells (Costar) were coated overnight with 100 μl of FNEB L (10 μg ml−1) in PBS. The plates were then blocked with bovine serum albumin (2%, wt/vol) for 1 h at 37°C. After the plates were washed, mouse or horse serum samples in PBS-T were added to the wells, followed by twofold serial dilutions. After incubation for 1 h at 37°C, the plates were washed, and antibody binding was detected with antibodies (1:1,000 dilution) against mouse or horse IgG (DAKO, Glostrup, Denmark) raised in rabbits and conjugated with HRP (Sigma Chemical Co.). The development of a colorimetric reaction was determined with o-phenylenediamine dihydrochloride tablets (DAKO). The absorbance was determined spectrophotometrically at 492 nm.
ELISA was also performed following the same procedure to determine levels of antibody against FNEB S, FNE, and SFS in serum samples from horses. Endpoint dilutions of antibodies are expressed as log10 values of the serum dilutions required to give an A492 of 1.0.
Nucleotide sequence accession number.
The GenBank accession number for the fneB gene is AY898649.
RESULTS
Identification of the fneB gene.
The genome of S. equi subsp. equi has been determined by shotgun sequencing, and finishing/gap closure is in progress (http://www.sanger.ac.uk/Projects/S_equi/). The S. equi subsp. equi genome was searched for open reading frames encoding proteins with Fn-binding repeats. A 1,425-nucleotide-long gene was found and has been given the name fneB. The fneB gene encodes a protein of 475 amino acids, termed FNEB (Fig. 1A). FNEB displays structures typical for gram-positive cell surface proteins, such as an N-terminal signal sequence (amino acids 1 to 35), a proline-rich cell wall-spanning region (amino acids 403 to 445), and a C-terminal hydrophobic transmembrane region. Furthermore, the transmembrane region is preceded by the amino acids LPKTH, resembling the common cell wall-anchoring LPXTG motif found in surface proteins of gram-positive bacteria (15). After export to the cell surface, which results in the removal of the signal sequence and cleavage in the LPXTG motif, the deduced mature cell wall-anchored protein should consist of 410 amino acids with a molecular mass of 45.6 kDa.
FIG. 1.

(A) Schematic presentation of FNEB and alignment to FNE and FNZ. The signal sequence (SS), wall-spanning region (W), membrane-spanning region (M), and cell wall-binding motif (LPKTH) are indicated. The C-terminal Fn-binding regions of FNEB and FNZ are in gray. The two horizontal bars represent recombinant proteins FNEB S and FNEB L. The numbers in parentheses refer to amino acid positions in FNEB. The similarities of the regions defined by horizontal lines are presented as percentages of identical amino acid residues. (B) Alignment of the repetitive Fn-binding C-terminal part of FNEB. Gaps (indicated by dashes) were inserted to obtain optimal alignment. The numbers in parentheses refer to amino acid positions. Amino acids fitting the conserved 5F1- to 2F1-binding motif ED(T/S)(X9,10)GG(X3,4)(I/V)DF are written in bold. The conserved amino acids that are assumed to mediate binding to the 1F1 module are underlined.
The presence of the fneB gene was confirmed in six out of six strains of S. equi subsp. equi by PCR, using primer OFE1:5 combined with OFE8:3 and OFE4:3. Fragments of the same length were also found in 3 out of 10 S. equi subsp. zooepidemicus strains tested.
Similarity of FNEB to related Fn-binding proteins.
Of the previously studied Fn-binding proteins in S. equi, FNZ of S. equi subsp. zooepidemicus is the only protein covalently attached to the cell surface. The C-terminal half of FNZ contains an Fn-binding region with sequence repeats typical of Fn-binding proteins in gram-positive cocci. This region has been shown to bind to the 29-kDa fragment of Fn. In the N-terminal part of FNZ, another hitherto uncharacterized Fn-binding motif is present (11). In S. equi subsp. equi, a frameshift mutation is present in the corresponding gene, fne, and hence, only the N-terminal part of the protein is translated. The protein, in this subspecies called FNE, thus contains the signal peptide and the N-terminal Fn-binding domain of FNZ. Without the C-terminal cell wall-anchoring motif, the native protein is secreted into the growth medium (11).
SFS is another secreted Fn-binding protein, found in both S. equi subsp. equi and S. equi subsp. zooepidemicus. The binding site of SFS has been mapped to a centrally located 21-amino-acid-long motif, repeated twice. These repeats show no similarities to any previously characterized Fn-binding proteins, but similar sequences are found in collagen. SFS was found to inhibit the binding between collagen and Fn, suggesting that collagen and SFS compete for the same site on Fn (8).
The similarities of FNEB to FNE and FNZ are outlined in Fig. 1A. The N-terminal half of FNEB displays 29% identical residues with FNE/FNZ. Otherwise, this part of the protein shows no significant similarities to the translated products of any genes in the GenBank database, with the exception of fnz2, a so far unpublished gene found in S. equi subsp. zooepidemicus (GenBank accession number BAD00708). The C-terminal repetitive Fn-binding region of the protein shows 61% identity to the corresponding region in FNZ. The same part of the protein shares significant similarities to a number of Fn-binding proteins in gram-positive bacteria, including SfbI/F1 (S. pyogenes), FnBPA (S. aureus), and FnBB (S. dysgalactiae) (20), among others.
The Fn-binding region of FNEB.
In the majority of the Fn-binding proteins of gram-positive cocci, the binding is directed to the N-terminal 29-kDa fragment of the Fn molecule. The 29-kDa fragment is composed of five tightly folded type 1 (F1) modules, 1F1 to 5F1 (20). Assays of chemically modified peptide fragments from bacterial proteins have shown that a conserved core sequence, defined as ED(T/S)(X9,10)GG(X3,4)(I/V)DF, mediates binding to the 29-kDa fragment (13). The three-dimensional structure of a streptococcal FnBP peptide (B3) in complex with the 1F1-to-2F1 module pair has been revealed (21). On binding to this module pair, the unfolded B3 peptide forms an antiparallel strand to the β-sheet structure of the F1 modules in a tandem β-zipper interaction. The study (21) also revealed that the previously known (I/F)HFDNX(X)P motif in streptococcal FnBPs mediates additional binding to the 1F1 module.
A similar organization of the putative Fn-binding region in FNEB can be predicted by sequence alignment. Figure 1B shows this region, with the conserved 2F1- to 5F1-binding residues and 1F1-binding motif indicated.
Binding specificities of FNEB, FNE, and SFS.
To confirm the Fn-binding activity of FNEB, horse sera were passed through a HiTrap NHS-activated HP column coupled with the FNEB L protein (Fig. 1A). The bound ligands were eluted and analyzed using SDS-PAGE. In Fig. 2, Western blot analysis confirms that the eluted fractions contain Fn. Furthermore, iodinated FNEB L was shown to bind to microtiter wells coated with Fn in a concentration-dependent manner (data not shown), which demonstrates that FNEB binds to both immobilized Fn and Fn in solution.
FIG. 2.
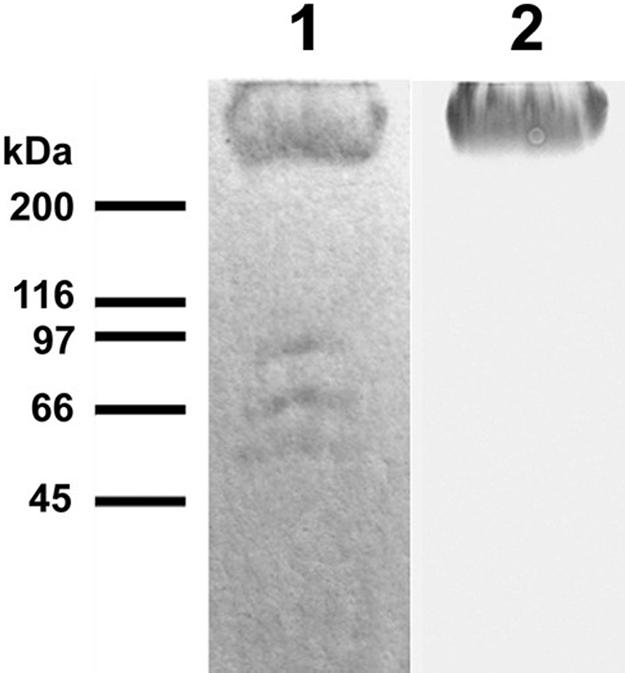
Lane 1, SDS-PAGE analysis of ligands to FNEB isolated from horse sera. Two of the bands between 45 and 116 kDa were identified as the IgG heavy chain and the IgM constant region using Western blotting and tandem mass spectrometry sequencing. These antibodies were not specific for FNEB but did bind directly to Fn, presumably due to denaturation, and were therefore ignored. Lane 2 shows anti-Fn Western blot results of the same gel. Molecular mass markers are indicated.
To determine the binding specificity of FNEB, a Western ligand blot analysis was performed with the iodinated 29-kDa-, 40-kDa-, and 105-kDa fragments. The results demonstrate that FNEB L specifically binds to the 29-kDa fragment (Fig. 3B). Since FNEB S shows no binding, it can be concluded that the binding domain is located in the C-terminal half of the protein. SFS is known to inhibit collagen binding to Fn, suggesting that collagen and SFS bind to the same site in Fn (8). The Western ligand blot analysis confirms that SFS binds to the 40-kDa collagen/gelatin-binding region of Fn (Fig. 3C). It has previously been shown that FNE and the N-terminal half of FNZ do not bind to the 29-kDa fragment of Fn (10). The Western ligand blot analysis shows that FNE binds to Fn within the 40-kDa fragment (Fig. 3C). None of the recombinant proteins bound to the 105-kDa fragment.
FIG. 3.

(A) SDS-PAGE gel with SFS (lane 1), FNE (lane 2), FNEB L (lane 3), and FNEB S (lane 4). Molecular mass markers are indicated. (B) Western ligand blot results showing binding of the iodinated 29-kDa fragment to FNEB L. (C) Western ligand blot results of the same gel, showing binding of the iodinated 40-kDa fragment to SFS and FNE. No binding to the 105-kDa fragment was detected.
To further analyze the binding specificities of the proteins, a series of inhibition assays were performed. In the first experiment, dilutions of SFS, FNE, and FNEB L were mixed with the iodinated 40-kDa fragment and incubated in microtiter wells coated with SFS. The results show that SFS, but not FNEB L, could inhibit the binding of the 40-kDa fragment to SFS (Fig. 4A). Interestingly, FNE could inhibit the same binding but to a lesser extent than SFS. In the second experiment, proteins were mixed with the iodinated 40-kDa fragment and incubated in wells coated with FNE. A cross inhibition between FNE and SFS was also seen here (Fig. 4B). In the third experiment, the iodinated 29-kDa fragment was used in wells coated with FNEB L. This binding was inhibited by FNEB L but not by FNE or SFS (Fig. 4C).
FIG. 4.

Inhibition studies using Fn-binding proteins. A fixed concentration of the iodinated 40-kDa or 29-kDa fragment (∼7,000 cpm and ∼15,000 cpm, respectively) was incubated with SFS (□), FNE (▵), or FNEB L (⧫). After 1 h, the samples were transferred to microtiter wells coated with SFS, FNE, or FNEB L, and after 2 h, the wells were washed, and the radioactivity bound to the wells was measured. Mean values ± standard deviations (SDs) (n = 3) are given. (A) SFS and FNE, but not FNEB L, inhibit the binding of the 40-kDa fragment to immobilized SFS. (B) FNE and SFS, but not FNEB L, inhibit the binding of the 40-kDa fragment to immobilized FNE. (C) FNEB L, but not SFS or FNE, inhibits the binding of the 29-kDa fragment to immobilized FNEB L.
Together, these experiments demonstrate that FNE and SFS both bind to the 40-kDa fragment of Fn but that FNEB binds to the 29-kDa fragment. The binding of SFS and FNE to Fn appears to be overlapping or located sufficiently close for a sterical inhibition.
Fn binding to whole cells of S. equi.
In all isolates of S. equi subsp. equi studied, the binding to Fn was considerably lower than in isolates of S. equi subsp. zooepidemicus. The binding of the 29-kDa fragment was found to be very low, and S. equi subsp. equi was therefore considered to be a nonbinder (10). However, a low but observable binding of the 29-kDa fragment to cells of S. equi subsp. equi could be demonstrated, as shown in Fig. 5. The binding of whole Fn, the 29-kDa fragment, and the 40-kDa fragment to cells of S. equi subsp. equi strain 1866 and DSM 20561 was compared to the binding to cells of S. equi subsp. zooepidemicus strain ZV. In this assay, S. equi subsp. equi showed a relatively low binding to the 29-kDa fragment and no binding to the 40-kDa fragment, while S. equi subsp. zooepidemicus bound to both.
FIG. 5.

Binding of Fn (A), the 29-kDa fragment (B), and the 40-kDa fragment (C) to two S. equi subsp. equi strains (1866 and DSM 20561) and the S. equi subsp. zooepidemicus strain ZV (9). Cells harvested at log phase (total, 3 × 106 CFU) in solution were incubated with the iodinated ligands, spun down, and washed, and the radioactivity was measured. Mean values ± SDs (n = 3) of the specific binding of the added ligand (given as percentages) are shown. No significant differences were observed when cells from overnight cultures were used (data not shown).
The binding of S. equi subsp. equi cells to the 29-kDa fragment, as well as to whole Fn, was inhibited when the ligand was preincubated with FNEB L (Fig. 6). In both experiments, FNEB S was used as a negative control. No binding between S. equi subsp. equi cells and the 105-kDa fragment could be detected.
FIG. 6.

Inhibition tests using whole cells. S. equi subsp. equi cells (total, 106 CFU) were mixed with a fixed concentration of the iodinated 29-kDa fragment or whole Fn (∼16,000 cpm and ∼22,000 cpm, respectively) and an increasing concentration of FNEB L (⧫) or FNEB S (□). After incubation, the cells were spun down and washed, and the radioactivity bound to the cells was measured. Mean values ± SDs (n = 3) are given. (A) FNEB L, but not FNEB S, inhibits the binding of the iodinated 29-kDa fragment to S. equi subsp. equi cells. (B) FNEB L, but not FNEB S, inhibits the binding of iodinated whole Fn to S. equi subsp. equi cells.
fneB, fne, and sfs are expressed in vitro.
Attempts to release the native FNEB protein from the cell surface by mechanical methods have so far not been successful. Instead, RT-PCR was used to confirm the expression of fneB, fne, and sfs in log-phase cells grown under standard laboratory conditions (Fig. 7). Following cDNA synthesis, the PCRs were performed with increasing numbers of cycles to quantify the differences in transcription levels of the three genes. The gyrA gene, here used as a positive control, gave clear bands after 22 cycles. fne required 24 cycles, fneB required 27 cycles, and sfs required 29 cycles. The highest expression is thus seen in fne, followed by fneB and sfs.
FIG. 7.
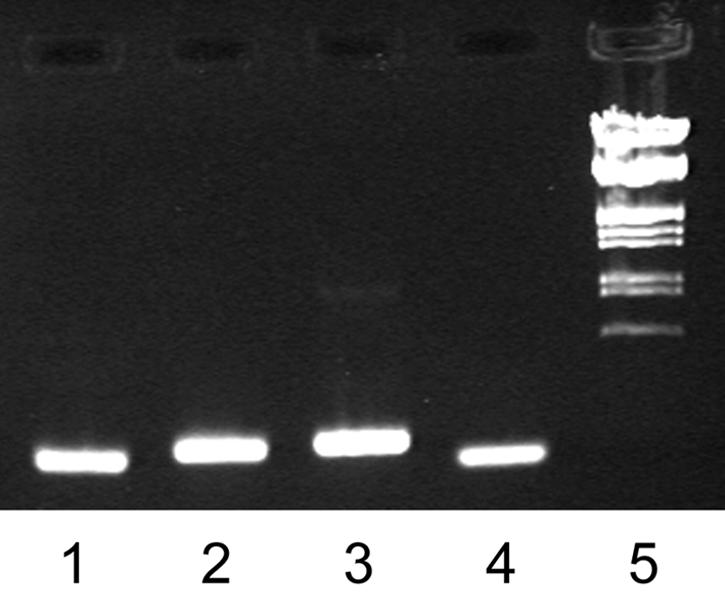
Gel electrophoresis of fneB (lane 1), fne (lane 2), sfs (lane 3), and gyrA (lane 4) RT-PCR products and EcoRI/HindIII-digested λ DNA marker (lane 5). Total RNA was prepared from cells grown to an OD600 of 0.6, and first-strand cDNA of the fneB, fne, sfs, and gyrA transcripts was prepared by reverse transcription. In the second reaction, the transcripts were amplified by gene-specific PCR.
IgG antibody response in mice and horses against FNEB.
To assess whether FNEB is expressed during an ongoing infection, a mouse model of S. equi subsp. equi infection was employed (5). Mouse sera were collected before and after the experimental infection. These sera were analyzed with an ELISA for IgG recognizing FNEB L. Endpoint dilutions are shown in Fig. 8. Antibody titers for mice infected with S. equi subsp. equi were higher (P < 0.05, by paired t test) than antibody titers for the same animals taken before infection, indicating that FNEB is expressed by S. equi subsp. equi during this experimental infection.
FIG. 8.

Titers of IgG antibody against FNEB L in sera from mice and horses with and without strangles. The log10 dilution of sera required to give an absorbance value at a cutoff of 1.0 was calculated for each individual serum sample. Mean values and standard errors of serum dilutions are shown. Mouse sera (n = 10) are from an experimental S. equi subsp. equi infection, and horse sera are from natural infections (n = 10) and from healthy horses (n = 16).
The expression of FNEB during infection was also demonstrated by a similar analysis of antibodies against FNEB L in horses. Serum samples were collected from horses with clinically verified strangles and from horses without any history of strangles. Figure 8 shows that there was a significant (P < 0.05, by unpaired t test) difference between these two groups, pointing to the fact that FNEB is expressed during infection and exposed to the immune system.
The same serum samples from horses were also tested in an ELISA for the presence of IgG antibodies against FNE and SFS. It has previously been shown (5) that there are significantly higher levels of IgG antibodies against FNE and SFS in sera from horses with strangles than in sera from healthy horses. In this study, titers of antibody in normal sera against FNE correlated with titers of antibody against FNEB L (r = 0.77) (Fig. 9B), but no correlation was found when sera from horses with strangles were analyzed (r = 0.34) (Fig. 9E). This is presumably due to different immune responses in different animals during infection. There was no correlation between IgG antibodies against SFS and FNEB L in any of the groups (Fig. 9C and F). However, there was a correlation between titers of antibody against FNEB S and FNEB L, both for healthy horses and for horses with strangles (r = 0.74 and 0.72) (Fig. 9A and D). This shows that the region that differs between FNEB S and FNEB L, the Fn-binding domain, does not strongly contribute to the immunogenicity of the protein.
FIG. 9.

Correlation between different horse IgG antibody titers. y axis, titers of antibody (expressed as log10 of dilution required to give an A of 1.0) against FNEB L; x axis, titers of antibody against FNEB S (A and D), FNE (B and E), and SFS (C and F). Panels A, B, and C show results for healthy horses without strangles. Panels D, E, and F show results for horses with strangles.
The levels of antibody against FNEB and FNE in healthy horses are presumably due to colonization with S. equi subsp. zooepidemicus. Individuals with a high level of colonization would have higher levels of antibody to both proteins, explaining the correlation in levels of antibody against FNEB L and FNE seen in Fig. 9B. However, upon infection with S. equi subsp. equi, a stronger humoral immune response is triggered, and different individuals respond differently to FNEB and FNE, eliminating the correlation seen in the noninfected group.
DISCUSSION
The present work describes the identification and cloning of FNEB, a protein in S. equi subsp. equi with conserved Fn-binding repeats commonly found in FnBPs of gram-positive cocci. Using PCR, fragments corresponding to the fneB gene were found in 6 out of 6 S. equi subsp. equi isolates and 3 out of 10 S. equi subsp. zooepidemicus isolates, in the latter subspecies probably due to the similar, as yet unpublished, fnz2 gene which has nearly identical primer sites.
Western ligand blotting was used to demonstrate that FNEB binds to the N-terminal 29-kDa fibrin-binding fragment of Fn. In binding assays using whole cells, S. equi subsp. equi showed a relatively low binding to the 29-kDa fragment and no binding to the 40-kDa fragment, while S. equi subsp. zooepidemicus bound to both. This is in accordance with the hypothesis that FNEB is the protein mediating Fn binding to cells of S. equi subsp. equi but that FNZ contributes to the Fn binding to cells of S. equi subsp. zooepidemicus. The observed binding of S. equi subsp. equi cells to the 29-kDa fragment, as well as to whole Fn, could be inhibited by the addition of recombinant FNEB, which further indicates that FNEB mediates cellular binding to Fn in this species.
In S. pyogenes and S. aureus, the importance of bacterial adhesion to Fn during infection is better understood. The binding of plasma Fn to these pathogens has been shown to trigger integrin-mediated internalization by epithelial and endothelial cells (17, 22). In S. equi subsp. equi, no intracellular survival has been reported in nonphagocytic cells, but since the bacterium has been known to cause long-term infection in asymptomatic carriers, the possibility of Fn-mediated S. equi subsp. equi internalization would be an interesting hypothesis to evaluate in future research.
The two previously studied Fn-binding proteins of S. equi subsp. equi, SFS and FNE, are both known to bind outside of the 29-kDa fragment. The present study shows that they bind to the adjacent 40-kDa gelatin/collagen-binding domain. Interestingly, cross-inhibition tests showed that the two proteins could prevent each other from binding to this part of Fn. This result indicates that the binding sites on Fn are overlapping or sufficiently close in location for a sterical inhibition. Although no known cell wall-anchoring motifs could be found in SFS and FNE, there is a possibility that these proteins reattach to the bacterial surface by an unknown mechanism. However, the lack of binding to the 40-kDa fragment in S. equi subsp. equi cells grown under conditions where FNE and SFS are expressed suggests that these proteins are not attached to the cell. The expression of SFS in cells grown in vitro might be too low to provide any detectable 40-kDa fragment binding, which leaves open the possibility for cell association, but a reattachment of FNE seems improbable considering that the protein has previously been detected in the media of laboratory cultures (11). The biological functions of these two secreted Fn-binding proteins remain to be investigated.
Serological studies of horses suffering from strangles, as well as a strangles model in mice, show that FNEB is expressed during infection, resulting in an IgG antibody response. However, in horses with strangles, titers of antibody against FNEB, FNE, and SFS show no correlation, indicating that no general antibody response against Fn-binding proteins is generated during S. equi subsp. equi infection. Instead, individual horses respond differently to these antigens. Even though strains of S. equi subsp. equi are genetically highly homogeneous, another possible explanation is a variation in the expression of the antigens in different isolates. The broad divergence in antibody responses against FNEB L and FNE in horses with strangles indicates that antibodies against these proteins do not cross react.
Considering the high homology between the genomes of S. equi subsp. equi and S. equi subsp. zooepidemicus, an interesting issue is the molecular basis of the differences in pathogenicity between these subspecies. Suggested factors include the S. equi subsp. equi-specific exotoxins SePE-I and -H, the antiphagocytic M protein, and the constitutive capsule synthesis, which in S. equi subsp. zooepidemicus is tightly regulated (23). As seen in this and previous studies, the level of Fn binding exemplifies another difference. In a recent study on the role of Fn adhesion in the virulence of S. pyogenes (16), the gene for protein F1 was introduced into a strain lacking this gene. Surprisingly, these F1-expressing bacteria were less virulent than the F1-negative strain in a mouse model. The virulence of these bacteria was partially restored when the bacteria were used to infect mice lacking plasma Fn. A similar enhancement of virulence of S. aureus lacking FnBPs, compared to that of its isogenic parental strain, has also been observed (12). In light of these findings, it is tempting to speculate that the lower ability of S. equi subsp. equi to bind to Fn than of S. equi subsp. zooepidemicus is a contributing factor to its increased virulence.
Acknowledgments
This work was supported by the AgriFunGen program at the Swedish University of Agricultural Sciences, Carl Tryggers Foundation (CTS 02:90), FORMAS (22.1/2001-0911), Stiftelsen Svensk Hästforskning (SSH:0447057), and NordVacc Läkemedel AB.
We also thank L. Frykberg, K. Jacobsson, and E. Ljunggren for critical comments and advice.
Editor: J. T. Barbieri
REFERENCES
- 1.Anzai, T., J. F. Timoney, Y. Kuwamoto, Y. Fujita, R. Wada, and T. Inoue. 1999. In vivo pathogenicity and resistance to phagocytosis of Streptococcus equi strains with different levels of capsule expression. Vet. Microbiol. 67:277-286. [DOI] [PubMed] [Google Scholar]
- 2.Artiushin, S. C., J. F. Timoney, A. S. Sheoran, and S. K. Muthupalani. 2002. Characterization and immunogenicity of pyrogenic mitogens SePE-H and SePE-I of Streptococcus equi. Microb. Pathog. 32:71-85. [DOI] [PubMed] [Google Scholar]
- 3.Fernandez, E., V. Blume, P. Garrido, M. D. Collins, A. Mateos, L. Dominguez, and J. F. Fernandez-Garayzabal. 2004. Streptococcus equi subsp. ruminatorum subsp. nov., isolated from mastitis in small ruminants. Int J. Syst. Evol. Microbiol. 54:2291-2296. [DOI] [PubMed] [Google Scholar]
- 4.Flanagan, J., N. Collin, J. Timoney, T. Mitchell, J. A. Mumford, and N. Chanter. 1998. Characterization of the haemolytic activity of Streptococcus equi. Microb. Pathog. 24:211-221. [DOI] [PubMed] [Google Scholar]
- 5.Flock, M., K. Jacobsson, L. Frykberg, T. R. Hirst, A. Franklin, B. Guss, and J.-I. Flock. 2004. Recombinant Streptococcus equi proteins protect mice in challenge experiments and induce immune response in horses. Infect. Immun. 72:3228-3236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Karlstrom, A., K. Jacobsson, M. Flock, J. I. Flock, and B. Guss. 2004. Identification of a novel collagen-like protein, SclC, in Streptococcus equi using signal sequence phage display. Vet. Microbiol. 104:179-188. [DOI] [PubMed] [Google Scholar]
- 7.Lannergard, J., L. Frykberg, and B. Guss. 2003. CNE, a collagen-binding protein of Streptococcus equi. FEMS Microbiol. Lett. 222:69-74. [DOI] [PubMed] [Google Scholar]
- 8.Lindmark, H., and B. Guss. 1999. SFS, a novel fibronectin-binding protein from Streptococcus equi, inhibits the binding between fibronectin and collagen. Infect. Immun. 67:2383-2388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lindmark, H., K. Jacobsson, L. Frykberg, and B. Guss. 1996. Fibronectin-binding protein of Streptococcus equi subsp. zooepidemicus. Infect. Immun. 64:3993-3999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lindmark, H., P. Jonsson, E. Engvall, and B. Guss. 1999. Pulsed-field gel electrophoresis and distribution of the genes zag and fnz in isolates of Streptococcus equi. Res. Vet. Sci. 66:93-99.10208886 [Google Scholar]
- 11.Lindmark, H., M. Nilsson, and B. Guss. 2001. Comparison of the fibronectin-binding protein FNE from Streptococcus equi subspecies equi with FNZ from S. equi subspecies zooepidemicus reveals a major and conserved difference. Infect. Immun. 69:3159-3163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.McElroy, M. C., D. J. Cain, C. Tyrrell, T. J. Foster, and C. Haslett. 2002. Increased virulence of a fibronectin-binding protein mutant of Staphylococcus aureus in a rat model of pneumonia. Infect. Immun. 70:3865-3873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.McGavin, M. J., S. Gurusiddappa, P. E. Lindgren, M. Lindberg, G. Raucci, and M. Hook. 1993. Fibronectin receptors from Streptococcus dysgalactiae and Staphylococcus aureus. Involvement of conserved residues in ligand binding. J. Biol. Chem. 268:23946-23953. [PubMed] [Google Scholar]
- 14.Meehan, M., P. Nowlan, and P. Owen. 1998. Affinity purification and characterization of a fibrinogen-binding protein complex which protects mice against lethal challenge with Streptococcus equi subsp. equi. Microbiology 144:993-1003. [DOI] [PubMed] [Google Scholar]
- 15.Navarre, W. W., and O. Schneewind. 1999. Surface proteins of gram-positive bacteria and mechanisms of their targeting to the cell wall envelope. Microbiol. Mol. Biol. Rev. 63:174-229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Nyberg, P., T. Sakai, K. H. Cho, M. G. Caparon, R. Fassler, and L. Bjorck. 2004. Interactions with fibronectin attenuate the virulence of Streptococcus pyogenes. EMBO J. 23:2166-2174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ozeri, V., I. Rosenshine, D. F. Mosher, R. Fassler, and E. Hanski. 1998. Roles of integrins and fibronectin in the entry of Streptococcus pyogenes into cells via protein F1. Mol. Microbiol. 30:625-637. [DOI] [PubMed] [Google Scholar]
- 18.Perris, R., and S. Johansson. 1987. Amphibian neural crest cell migration on purified extracellular matrix components: a chondroitin sulfate proteoglycan inhibits locomotion on fibronectin substrates. J. Cell Biol. 105:2511-2521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Proft, T., P. D. Webb, V. Handley, and J. D. Fraser. 2003. Two novel superantigens found in both group A and group C Streptococcus. Infect. Immun. 71:1361-1369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schwarz-Linek, U., M. Hook, and J. R. Potts. 2004. The molecular basis of fibronectin-mediated bacterial adherence to host cells. Mol. Microbiol. 52:631-641. [DOI] [PubMed] [Google Scholar]
- 21.Schwarz-Linek, U., J. M. Werner, A. R. Pickford, S. Gurusiddappa, J. H. Kim, E. S. Pilka, J. A. Briggs, T. S. Gough, M. Hook, I. D. Campbell, and J. R. Potts. 2003. Pathogenic bacteria attach to human fibronectin through a tandem beta-zipper. Nature 423:177-181. [DOI] [PubMed] [Google Scholar]
- 22.Sinha, B., P. P. Francois, O. Nusse, M. Foti, O. M. Hartford, P. Vaudaux, T. J. Foster, D. P. Lew, M. Herrmann, and K. H. Krause. 1999. Fibronectin-binding protein acts as Staphylococcus aureus invasin via fibronectin bridging to integrin alpha5beta1. Cell. Microbiol. 1:101-117. [DOI] [PubMed] [Google Scholar]
- 23.Timoney, J. F. 2004. The pathogenic equine streptococci. Vet. Res. 35:397-409. [DOI] [PubMed] [Google Scholar]
- 24.Timoney, J. F., S. C. Artiushin, and J. S. Boschwitz. 1997. Comparison of the sequences and functions of Streptococcus equi M-like proteins SeM and SzPSe. Infect. Immun. 65:3600-3605. [DOI] [PMC free article] [PubMed] [Google Scholar]