

Abstract
Inhibitors of the 3-hydroxy-3-methylglutaryl coenzyme A reductase enzyme have recently been shown to stimulate bone formation in rodents both in vitro and in vivo. In bone cells, these inhibitors increase the gene expression of bone morphogenetic protein-2, which is an autocrine-paracrine factor for osteoblast differentiation.
The findings that statins increase bone formation and bone mass in rodents suggest a potential new action for these compounds, which may be beneficial in patients with established osteoporosis where marked bone loss has occurred. Recent clinical data suggest that they may reduce the risk of fracture in patients taking these drugs.
Keywords: bone formation, HMG-CoA reductase inhibitors, osteoblasts, osteoporosis, statins
Introduction
There has been a remarkable increase in knowledge in the area of osteoporosis during the last 25 years. Patients with established osteoporosis have lost more than 50% of bone mass at critical sites in the skeleton, with marked disruption of trabecular bone microarchitecture. Anabolic therapies, therefore, are desperately needed.
Current drugs to treat osteoporosis include bisphosphonates, calcitonin, estrogen and related compounds, vitamin D analogues and ipriflavone. These are all bone resorption inhibitors, which act mainly to stabilize bone mass by inhibiting the activity of osteoclasts (the cells responsible for bone loss). The ability of these drugs to increase bone mass is relatively small, certainly no more than 2% per year. It is desirable, therefore, to have a satisfactory and universally acceptable drug that would stimulate new bone formation and correct the disturbance of trabecular microarchitecture, which is a characteristic of established osteoporosis.
Chemical nature of statins
Statins are specific, competitive inhibitors of the 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase enzyme. These include naturally occurring lovastatin, chemically modified simvastatin and pravastatin [1-3] and the synthetically derived atorvastatin, fluvastatin and cerivastatin. All of these agents are widely used for lowering cholesterol, and they provide an important and effective approach to the treatment of hyperlipidemia and arteriosclerosis [4].
Because the primary site of cholesterol synthesis is the liver, these agents have been designed to be hepatoselective. The enzyme HMG-CoA reductase catalyzes the rate-limiting step in cholesterol biosynthesis, and while cholesterol is the bulk product of the pathway controlled by this enzyme, its direct product, mevalonate, is a precursor to a number of non-sterol compounds that are vital to a variety of cellular functions.
Bone metabolism
Bone is a metabolically active organ in which the organizational pattern of the mineral and organic components determines the successful mechanical function of the skeleton [5,6]. Bone turnover is controlled by defined agents and mechanisms that regulate bone formation and bone resorption, which are the two major processes of bone remodeling. Disturbances in these mechanisms can lead to either bone loss, resulting in osteoporosis, or an overgrowth of bone, leading to osteosclerosis. Since new bone formation is primarily a function of the osteoblast, agents regulating bone formation can act by either increasing/decreasing the replication of cells of the osteoblastic lineage or modifying the differentiated function of the osteoblast.
It would, therefore, be beneficial to stimulate the osteoblastic activity at local sites in bone by an oral anabolic agent, resulting in bone formation where needed.
Discovery of the effects of statins on bone
In attempts to identify small molecular weight bone anabolic compounds, attention has focused on the growth regulatory factors responsible for the control of normal bone remodeling. The bone morphogenetic proteins (BMPs) have bone-forming activity and account for the major proportion of the osteoinductive potential of bone extracts [7-9]. The BMP-2 promoter has been characterized, and based on the properties of BMP-2, this promoter was utilized as a target to identify new compounds that stimulate its transcription and subsequent osteoblast differentiation.
Identification of small molecules that enhance BMP-2 transcription utilized a cell-based screening assay [10]. Screening a collection of natural products led to the identification of an extract, containing lovastatin as the active constituent, that specifically stimulated the BMP-2 promoter. Further investigations found that statins stimulate bone formation both in vitro and in vivo in animal models of osteoporosis associated with increased expression of the BMP-2 gene in bone cells [11].
In vitro effects
Simvastatin, mevastatin and atorvastatin (but not pravastatin) were found to have identical effects to those seen with lovastatin. Cerivastatin, however, was 10-100-fold more potent than the other statins. These agents stimulated BMP-2 transcription and also increased endogenous BMP-2 mRNA and protein expression in human MG63 osteoblastic cells two-fold. These findings have been confirmed by other groups [12,13].
It has been shown that statins cause a marked increase in osteoblast accumulation and new bone formation in cultures of neonatal murine calvaria (Fig. 1). Transient exposure of bone cultures to statins was enough to initiate a cascade of bone formation, probably induced by the local production of the osteogenic protein BMP-2. Interestingly, pravastatin was unable to stimulate the BMP-2 promoter activity and it did not stimulate new bone formation in neonatal murine calvaria.
Figure 1.
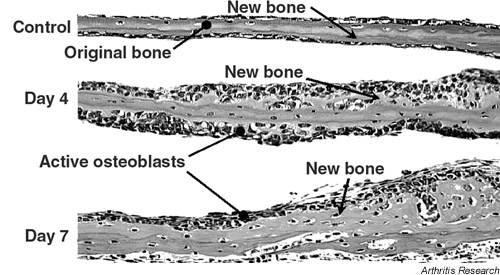
Cultures of murine neonatal calvaria incubated for either 4 or 7 days in the presence of simvastatin at 1 μM. Small amounts of new bone are present in control cultures whereas cultures exposed to simvastatin for 4 days show marked new bone formation and osteoblast accumulation. Cultures exposed for 7 days show further enhancement of bone formation.
In vivo effects
Initial in vivo experiments have shown that statins injected locally over the calvaria of normal mice result in a 30–50% increase in calvarial width. This indicates that statins have a direct effect on bone formation when applied locally.
There is a requirement, however, for an oral bone anabolic agent that stimulates systemic new bone formation for the treatment of bone loss diseases, such as osteoporosis. Ovariectomized rats, treated systemically with statins, showed marked increases in bone density when compared to untreated rats. Bones of rats treated orally with cerivastatin showed a 43% increase in tibial trabecular volumes, and rats treated orally with simvastatin showed a 38% increase in tibial trabecular volumes compared to the controls (Fig. 2). Fibroblast growth factor has been previously shown to stimulate bone formation in vivo and was used as a positive control. The anabolic effect of statins was confirmed with significant increases in both bone formation rate and mineral apposition rate in the tibiae of rats treated with cerivastatin at 0.1 mg/kg/day. Statins, therefore, have the potential to stimulate bone formation both in vitro and in vivo in rats. Cerivastatin improved cortical bone strength in ovariectomized rats when used in doses as low as 0.1 mg/kg/day, and it significantly increased bone mineral density, bone formation rate, osteocalcin mRNA levels as well as resistance to fracture [14]. Further studies have shown that simvastatin given orally to rats significantly increased cancellous bone compressive strength in the vertebral bodies of these rats [15].
Figure 2.
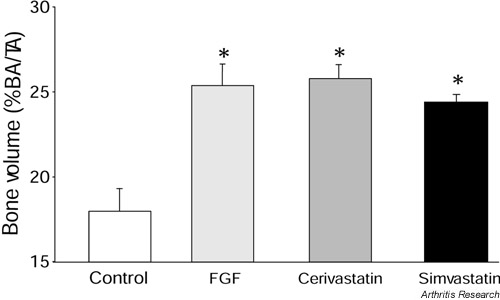
Bone volume in the tibia of ovariectomized rats treated either by daily oral lavage with cerivastatin (0.1 mg/kg/day) or simvastatin (10 mg/kg/day) or subcutaneously with fibroblast growth factor (FGF) (80 μg/kg/day). The bone volume is expressed as per cent bone area (BA)/ total area measured (TA). *P < 0.05, versus control group.
Mechanism of action
The reduction in mevalonate pathway intermediates with a subsequent inhibition of prenylation by statins is responsible for a large proportion of the pleiotropic effects of these drugs. Mevalonate, farnesyl pyrophosphate and geranylgeranyl pyrophosphate all inhibited statin-stimulated bone formation. Furthermore, because geranylgeranyl pyrophosphate inhibited statin stimulated bone formation, inhibition of prenylation due to geranylgeranylation must play a major role in the stimulation of bone formation by this drug.
There are many proteins known to require this form of prenylation for their activity, including guanosine triphosphatases such as Rho, Rac and Rap. These proteins play important roles in cellular proliferation and differentiation, and, therefore, any perturbation of their activity influences cellular activity. A number of the pleiotropic effects of statins result from their effects on prenylation. One of the particular roles prenylation plays in cellular activity is its control of endothelial nitric oxide synthase [16-18] and it has subsequently been shown to play a major role in the effects of statins on bone formation.
Clinical findings
Statins are bone anabolic agents, which have been orally administered to rats, and which have relatively low toxicity in humans. They could provide an important treatment for osteoporosis, particularly when significant amounts of trabecular bone have been lost. Current therapies for the treatment of osteoporosis, including estrogen replacement therapy, selective estrogen receptor modulators, and bisphosphonates, are primarily based on blunting the resorption component of bone remodeling.
Of course, the major question that arises is whether statins will have similar effects on human bone. Based on previous findings [13], Bauer and Cummings examined their large databases to determine if there was any previously unrecognized association between statin usage and skeletal status. They found that there was a possible relationship between statin use, bone mineral density and subsequent fractures [19]. Since then a study in post-menopausal women has been published that indicated a significant increase in bone mineral density associated with taking statins [20]. Statins have also been shown to exhibit a protective effect against non-pathological fracture among older women [21-25]. Conversely, several preliminary reports (one using the same database as a positive published report) have suggested that statins do not show these effects [26-28]. There are major issues with all of these studies: they are retrospective; the compliance of patients taking is statins unknown; and the dose of statin used varied considerably.
All of the statins that are currently available have been selected for their capacity to target the liver and decrease cholesterol biosynthesis, but they are poorly distributed to bone. It is uncertain, therefore, that oral administration of currently available statins will have beneficial effects on bone in humans; however, there are several possibilities for improving biodistribution to bone.
The more recent potent statins such as cerivastatin or atorvastatin may get past the liver and reach the bone. Alternative modes of administration of the statins, such as topical application through a skin patch, may improve biodistribution. Furthermore, there may be other drugs of this class that have not been selected for development as cholesterol-lowering agents because of their relatively greater biodistribution to peripheral tissues. These may be ideal drugs for use as bone-active agents.
Conclusion
Perhaps the most important consequence of these findings is not that the statins themselves may be effective drugs for diseases of bone loss, but rather that these results focus attention on the pathway of cholesterol biosynthesis and its relationship to BMP-2 expression and bone formation. This has been further emphasized by recent observations that the nitrogen-containing bisphosphonates (drugs that reduce bone resorption and have a large market for osteoporosis) also target enzymes in this pathway. This may lead to the identification of other potential molecular targets for drug discovery as well as other therapeutic approaches to enhance bone formation, and thus produce the ideal anabolic agent for osteoporosis.
Abbreviations
BMP = bone morphogenetic protein; HMG-CoA = 3-hydroxy-3-methylglutaryl coenzyme A.
References
- Todd PA, Goa Kl. Simvastatin. A review of its pharmacological properties and therapeutic potential in hypercholesterolaemia. Drugs. 1990;40:583–607. doi: 10.2165/00003495-199040040-00007. [DOI] [PubMed] [Google Scholar]
- Kishida Y, Naito A, Iwado S, Terahara A, Tsujita Y. Research and development of pravastatin. Yakugaku Zasshi. 1991;111:469–487. doi: 10.1248/yakushi1947.111.9_469. [DOI] [PubMed] [Google Scholar]
- Henwood JM, Heel RC. Lovastatin. A preliminary review of its pharmacodynamic properties and therapeutic use in hyperlipidaemia. Drugs. 1988;36:429–454. doi: 10.2165/00003495-198836040-00003. [DOI] [PubMed] [Google Scholar]
- Hunninghake DB. Therapeutic efficacy of the lipid-lowering armamentarium: the clinical benefits of aggressive lipid-lowering therapy. Am J Med. 1998;104:9s–13s. doi: 10.1016/s0002-9343(98)00040-0. [DOI] [PubMed] [Google Scholar]
- Glimcher MJ, Krane SM. The organizational and structure of bone and the mechanism of calcification. In: Ramachandran GN, Goudl BS, editor. A treatise on collagen. Biology of collagens. New York: Academic Press; 1968. pp. 68–91. [Google Scholar]
- Glimcher MJ. Comparison, structure and organization of bone and other mineralized tissues and the mechanism of calcification. In: Aubarch GD, editor. Handbook of physiology, endocrinology, parathyroid gland. Washington: American Physiological Society; 1976. pp. 25–48. [Google Scholar]
- Mundy GR, Boyce B, Hughes D, Wright K, Bonewald L, Dallas S, Harris S, Ghosh-Choudhury N, Chen D, Dunstan C, Izbicka E, Yoneda T. The effects of cytokines and growth factors on osteoblastic cells. Bone. 1995;17:71s–75s. doi: 10.1016/8756-3282(95)00182-d. [DOI] [PubMed] [Google Scholar]
- Wozney JM, Rosen V. Bone morphogenetic proteins. In: Mundy GR, Martin TJ, editor. Physiology and Pharmacology of Bone. New York: Springer-Verlag; 1998. pp. 725–748. [Google Scholar]
- Harris SE, Bonewald LF, Harris MA, Sabatini M, Dallas S, Feng J, Ghosh-Choudhury N, Wozney J, Mundy GR. Effects of transforming growth factor beta on bone nodule formation and expression of bone morphogenetic protein 2, osteocalcin, osteopontin, alkaline phosphatase, and type I collagen mRNA in long-term cultures of fetal rat calvarial osteoblasts. J Bone Miner Res. 1994;9:855–863. doi: 10.1002/jbmr.5650090611. [DOI] [PubMed] [Google Scholar]
- Ghosh-Choudhury N, Windle JJ, Koop BA, Harris MA, Guerrero DL, Wozney JM, Mundy GR, Harris SE. Immortalized murine osteoblasts derived from BMP 2-T-antigen expressing transgenic mice. Endocrinology. 1996;137:331–339. doi: 10.1210/endo.137.1.8536632. [DOI] [PubMed] [Google Scholar]
- Mundy G, Garrett R, Harris S, Chan J, Chen D, Rossini G, Boyce B, Zhao M, Gutierrez G. Stimulation of bone formation in vitro and in rodents by statins. Science. 1999;286:1946–1949. doi: 10.1126/science.286.5446.1946. [DOI] [PubMed] [Google Scholar]
- Sugiyama M, Kodama T, Konishi K, Abe K, Asami S, Oikawa S. Compactin and simvastatin, but not pravastatin, induce bone morphogenetic protein-2 in human osteosarcoma cells. Biochem Biophys Res Commun. 2000;271:688–692. doi: 10.1006/bbrc.2000.2697. [DOI] [PubMed] [Google Scholar]
- Maeda T, Matsunuma A, Kawane T, Horiuchi N. Simvastatin promotes osteoblast differentiation and mineralization in MC3T3-E1 cells. Biochem Biophys Res Commun. 2001;280:874–877. doi: 10.1006/bbrc.2000.4232. [DOI] [PubMed] [Google Scholar]
- Wilkie D, Bowman B, Lyga A, Bagi CM, Miller SC, Ranges GE, Carley W. Cerivastatin increases cortical bone formation in OVX rats. J Bone Min Res. 2000;15(suppl):s549. [Google Scholar]
- Oxlund H, Andreassen TT. Simvastatin given orally to adult rats increases the compressive strength of vertebral bodies. J Bone Min Res. 2000;15(suppl):s549. [Google Scholar]
- Laufs U, Fata VL, Liao JK. Inhibition of 3-hydroxy-3-methylglutaryl (HMG)-CoA reductase blocks hypoxia-mediated down-regulation of endothelial nitric oxide synthase. J Biol Chem. 1997;272:31725–31729. doi: 10.1074/jbc.272.50.31725. [DOI] [PubMed] [Google Scholar]
- Laufs U, Liao JK. Post-transcriptional regulation of endothelial nitric oxide synthase mRNA stability by Rho GTPase. J Biol Chem. 1998;273:24266–24271. doi: 10.1074/jbc.273.37.24266. [DOI] [PubMed] [Google Scholar]
- Garrett IR, Esparza J, Chen D, Zhao M, Gutierrez G, Escobedo A, Horn D, Mundy GR. Statins mediate their effects on osteoblasts by inhibition of HMG-CoA reductase and ultimately BMP2. J Bone Min Res. 2000;15(suppl):s225. [Google Scholar]
- Bauer DC, Mundy GR, Jamal SA, Black DM, Cauley JA, Harris F, Duong T, Cummings SR. Statin use, bone mass and fracture: an analysis of two prospective studies. J Bone Min Res. 1999;14:1188. [Google Scholar]
- Edwards CJ, Hart DJ, Spector TD. Oral statins and increased bone-mineral density in postmenopausal women. Lancet. 2000;355:2218–2219. doi: 10.1016/s0140-6736(00)02408-9. [DOI] [PubMed] [Google Scholar]
- Chan KA, Andrade SE, Boles M, Buist DS, Chase GA, Donahue JG, Goodman MJ, Gurwitz JH, Lacroix AZ, Platt R. Inhibitors of hydroxymethylglutaryl-coenzyme A reductase and risk of fracture among older women. Lancet. 2000;355:2185–2188. doi: 10.1016/S0140-6736(00)02400-4. [DOI] [PubMed] [Google Scholar]
- Wang PS, Solomon DH, Mogun H, Avorn J. HMG-CoA reductase inhibitors and the risk of hip fractures in elderly patients. JAMA. 2000;283:3211–3216. doi: 10.1001/jama.283.24.3211. [DOI] [PubMed] [Google Scholar]
- Meier CR, Schlienger RG, Kraenzlin ME, Schlegel B, Jick H. HMG-CoA reductase inhibitors and the risk of fractures. JAMA. 2000;283:3205–3210. doi: 10.1001/jama.283.24.3205. [DOI] [PubMed] [Google Scholar]
- Meier CR, Schlienger RG, Kraenzlin ME, Schlegel B, Jick H. Statin drugs and the risk of fracture. JAMA. 2000;284:1921–1922. [PubMed] [Google Scholar]
- Chung YS, Lee MD, Lee SK, Kim HM, Fitzpatrick LA. HMG-CoA reductase inhibitors increase BMD in type 2 diabetes mellitus patients. J Clin Endocrinol Metab. 2000;85:1137–1142. doi: 10.1210/jcem.85.3.6476. [DOI] [PubMed] [Google Scholar]
- Lacroix AZ, Cauley JA, Jackson R, McGowan J, Pettinger M, Hsia J, Chen 7, Lewis C, Bauer DC, Daugherty 8, McNeeley SG, Passaro M. Does statin use reduce risk of fracture in post-menopausal women? Results from the Women's Health Initiative Observational Study [abstract 1066]. J Bone Min Res. 2000;15(suppl):s155. [Google Scholar]
- van Staa TP, Wegman SLJ, de Vries F, Leufkens HGM, Cooper C. Use of statins and risk of fractures [abstract 1067]. J Bone Min Res. 2000;15(suppl):s155. [Google Scholar]
- Cauley JA, Jackson R, Pettinger M, Lacroix A, Bauer D, Chen Z, Daugherty S, Hsia J, Lewis CE, McGowan J, McNeeley SG, Passaro M. Statin use and bone mineral density (BMD) in older women: The Women's Health Initiative Observational Study (WH I-OS) [abstract 1068]. J Bone Min Res. 2000;15(suppl):s155. [Google Scholar]