

Abstract
A comprehensive technique was developed for using three-dimensional 17O magnetic resonance spectroscopic imaging at 9.4T for rapidly imaging the cerebral metabolic rate of oxygen consumption (CMRO2) in the rat brain during a two-min inhalation of 17O2. The CMRO2 value (2.19 ± 0.14 μmol/g/min, n = 7) was determined in the rat anesthetized with α-chloralose by independent and concurrent 17O NMR measurements of cerebral H217O content, arterial input function, and cerebral perfusion. CMRO2 values obtained were consistent with the literature results for similar conditions. Our results reveal that, because of its superior sensitivity at ultra-high fields, the 17O magnetic resonance spectroscopic imaging approach is capable of detecting small dynamic changes of metabolic H217O during a short inhalation of 17O2 gas, and ultimately, for imaging CMRO2 in the small rat brain. This study provides a crucial step toward the goal of developing a robust and noninvasive 17O NMR approach for imaging CMRO2 in animal and human brains that can be used for studying the central role of oxidative metabolism in brain function under normal and diseased conditions, as well as for understanding the mechanisms underlying functional MRI.
In the brain, the majority of energy consumption occurs by neuronal activity (1). This energy need is met predominantly through oxygen consumption mediated by the mitochondrial respiratory chain, coupled to oxidation of glucose as the main carbon source (2). The oxidative energy need of the human brain to sustain neuronal activity constitutes a large fraction of the energy requirements (≈20%) in the entire body. The central role played by oxygen consumption is also evident in pathologies associated with the brain; perturbations in brain oxidative metabolism have been closely linked to many brain diseases such as schizophrenia, Alzheimer's disease, Huntington's disease, Parkinson's disease, and mitochondrial dysfunction, as well as aging problems (3–7). One line of evidence linking these diseases and cerebral oxidative metabolism is the histopathological finding that the activity of cytochrome oxidase, the key mitochondrial enzyme that catalyzes the reduction of oxygen to form water, is impaired in patients with schizophrenia (3) and Alzheimer's disease (4, 5).
Basal cerebral oxygen consumption is not uniform across different regions in the brain. It is recognized that capillary density in the brain is inhomogeneous (ref. 8 and refs. therein), suggesting that mitochondrial density, and hence, oxygen consumption are also likely to be inhomogeneous because a good correlation exists between capillarity and mitochondrial density in tissues (9). Alterations in neuronal activity due to stimulation or performance of a task increases this spatial nonuniformity by inducing regional changes in oxygen utilization as well as in blood flow and glucose consumption (10–12). In addition, the impairment in cytochrome oxidase content in the diseased brain was also found to affect different regions in the brain selectively (3–5). These considerations imply that the ability to image the cerebral metabolic rate of oxygen consumption (CMRO2) in vivo is essential for efforts aimed at investigating cerebral oxidative metabolism under normal and pathological conditions.
The earliest approach of CMRO2 measurement is based on the Kety-Schmidt method for measuring cerebral blood flow (CBF) together with arteriovenous differences of oxygen content (13). This method, thought to be the most accurate one in the literature, provides only global CMRO2 information without spatial differentiation within the brain. Positron emission tomography (PET) has been widely used for imaging CMRO2 in humans (14–16). By introducing oxygen gas enriched with the isotope 15O into the human body and monitoring the spatial distribution and accumulation rate of metabolic water (H215O), PET can determine regional CMRO2 and provide a CMRO2 image. However, the 15O-PET is unable to distinguish the radioactive signals between the 15O2 molecule bound to hemoglobin and the 15O atom incorporated into H215O. To overcome this drawback, a measurement of cerebral blood volume using inhalation of C15O has to be performed in addition to the CBF measurement based on i.v. injection of H215O. These PET requirements make CMRO2 measurements difficult to execute experimentally and complicate the calculation of CMRO2 values.
The 13C NMR methods based on monitoring the cerebral metabolism of intravenously infused [1-13C] glucose (e.g., (10, 11, 17, 18)) provide an alternative approach for CMRO2 measurements. The labeling kinetics for several intracellular metabolites, most importantly glutamate, can be unequivocally measured by these methods; extraction of CMRO2 from experimental data relies on extensive modeling (10, 11, 18, 19). In addition, the 13C NMR methods require a long measurement time (60–120 min) because of relatively slow turnover rates of cerebral metabolites, and it is difficult, although not impossible, to achieve three-dimensional (3D) CMRO2 imaging because of a relatively long repetition time required for signal acquisitions.
The possibility of using the 17O NMR spectroscopy/imaging techniques for monitoring labeled H217O as a tracer of oxygen utilization or tissue perfusion have been introduced a decade ago (20–30). These early studies can be divided into two distinct groups. The first group involved the simple 17O NMR approach for detecting H217O directly (20, 21, 24–27). Nonlocalized 17O NMR was used for most studies to estimate CMRO2 in the whole brain of experimental animals as well as in the human occipital lobe (31) after inhalation of oxygen gas enriched with the 17O isotope (17O2). Few studies attempted to obtain a coarse CMRO2 image (≥800 μl voxel size) in the cat brain with a long measurement time (25, 27). These previous efforts, all of which were conducted at relatively low fields compared with what is available currently, demonstrated the dramatic limitations imposed by the low inherent sensitivity of 17O NMR due to its low gyromagnetic ratio. These sensitivity limitations have hindered further developments of the 17O NMR approach and lead to the exploration of a second group of methods that attempt to circumvent the sensitivity limitations of 17O NMR by using indirect detection through 17O-coupled protons.
The indirect 17O detection approach relies on the effect of 1H-17O scalar coupling on the transverse relaxation time (T2) of the water protons, which is considerably shortened by the presence of the 17O nucleus in the same water molecule (22). It has also been shown that the water proton T2 of H217O can be restored to that of H216O when 17O decoupling is applied; in this case, the difference between proton signals obtained with and without 17O decoupling is related to the 17O content of the water sample (28–30). However, the indirect 17O measurements present difficulties for quantitatively correlating the proton signal changes to the concentrations of H217O because the water T2 is sensitive to many physiologic parameters such as pH and temperature. In addition, the fractional change in 1H signal induced by variations in metabolic H217O concentration is small at basal conditions. These indirect 17O detection approaches have so far not been successful in imaging CMRO2 quantitatively.
Recently, we have investigated the longitudinal and transverse relaxation times [T1, T2, and apparent T2 (T2*)] as well as the signal-to-noise ratio (SNR) of 17O spins in water by using the 17O magnetic resonance spectroscopic (MRS) imaging method at 4.7T and 9.4T field strengths (32). Our results showed that the relaxation times of 17O water are field-independent, and the 17O NMR sensitivity increased approximately fourfold at 9.4T compared with a field strength of 4.7T. With this sensitivity gain, we were able to obtain two-dimensional 17O MRS images of natural abundance H217O in the rat brain at 9.4T with a temporal resolution of 12 s (32). These results demonstrated the potential of 17O MRS imaging at ultra-high field for mapping the spatial distribution of metabolic H217O dynamically in the rat brain.
In this study, we investigated the feasibility for developing the 17O NMR approach for imaging CMRO2 in a small animal model during a brief inhalation of 17O2 gas. We have successfully applied the 3D 17O MRS imaging method to determine CBF after a bolus injection of H217O through an internal carotid artery and the accumulation rate of H217O generated by oxidative metabolism in the rat brain during a two-min 17O2 inhalation with superior spatial (27 μl) and temporal (11 s per 3D image) resolution. Furthermore, we performed experiments where such images were obtained together with continuous experimental monitoring of the arterial input function related to recirculated H217O via an implanted carotid artery MR coil. These measurements were used to calculate CMRO2 values rigorously in image voxels by using a complete modeling that accounts for all contributions to the cerebral H217O content accumulated during an inhalation of 17O2 and demonstrate the feasibility for imaging CMRO2. Based on these results, we present high-resolution 3D CMRO2 images obtained with 17O NMR under the basal condition in the rat brain anesthetized with α-chloralose.
Theory
The dynamic change of cerebral H217O concentration accumulated during an inhalation of 17O2 is determined by three processes: oxygen consumption generating metabolic H217O in the brain, perfusion resulting in H217O washout from the brain, and flow recirculation bringing extra H217O into the brain. The mass balance equation of labeled H217O concentrations during an inhalation of 17O2 is given by
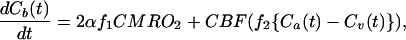 |
1 |
where Ca(t), Cb(t), and Cv(t) are the time-dependent H217O concentrations expressed as [H217O] in excess of the natural abundance [H217O] level in the arterial blood, brain tissue, and venous blood, respectively. The constant α is the enrichment fraction of 17O-labeled O2 gas. Proper use of Eq. 1 requires consistencies of units among all terms used in the equation. The determinations of Cb, Ca, and Cv are based on the 17O NMR measurements and calibrated by using the natural abundance H217O concentration (20.35 μmol/(g of brain water) for brain tissue, and μmol/(g of blood water) for blood, calculated from natural abundance H217O enrichment of 0.037% and the molecular weight of H217O = 19.0. Therefore, we prefer to use convenient units of μmol/(g of brain water) for Cb(t) and μmol/(g of blood water) for Ca(t) and Cv(t). CMRO2 is expressed in the conventional unit of μmol/min/(g brain tissue) and is converted from the unit of μmol/min/(g brain water) by using the conversion constant f1 given by f1 = (g of brain tissue)/(g of brain water) = 1/βbrain = 1.266, where βbrain is 0.79 (14, 27). The constant f2 is also a unit conversion factor and is given by f2 = ρ blood⋅βblood/βbrain = 1.05 × 0.81/0.79 = 1.077, where ρblood = (g blood)/(ml blood) = 1.05 (33), and βblood = (g blood water)/(g blood) = 0.81 (14). If water in brain tissue is in equilibrium with water in venous blood (i.e., fast water exchange across capillaries), f2Cv(t) = Cb(t)/λ where λ = 0.90 is the brain/blood partition coefficient with the unit of (ml blood)/(g of brain tissue) (34). Substituting this relation and introducing two new correction parameters (n and m) into Eq. 1 leads to
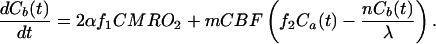 |
2 |
The solution of Eq. 2 is
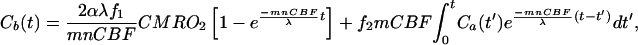 |
3 |
where m is a correction factor accounting for the limited permeability of water because water is not freely diffusible across the brain-blood barrier; the value of m is 0.84 for the CBF range that we studied herein (35). The constant n is another correction factor that accounts for the additional restriction on the permeability of the H217O generated through oxidative metabolism in the mitochondria (“metabolic” H217O). This additional restriction is included in this modeling because we have observed that the washout rate of the metabolic H217O after the cessation of 17O2 inhalation was significantly slower than the washout rate of the H217O that permeates brain tissue subsequent to a bolus injection of H217O through the internal carotid artery in the rat.‡ The cause of this difference could be permeability restrictions imposed by the inner and outer mitochondrial membranes that must be traversed by all water molecules generated by oxygen consumption. The ratio of the washout rate of the metabolic H217O after the cessation of 17O2 inhalation vs. the cerebral H217O after a bolus injection of H217O through the internal carotid artery gives the value of n. Limited permeability for the metabolic water has been reported in the human brain (36).
The CMRO2 value can be precisely calculated by using Eq. 3 if all parameters of Cb(t), CBF, and Ca(t) can be experimentally measured for the same animal brain. The Cb(t) and CBF values can be determined by using 17O NMR approaches. To measure Ca(t), we designed an implanted 17O rf coil that permits the continuous measurement of Ca(t) in a rat carotid artery. This measurement can be simultaneously performed with Cb(t) measurement during an inhalation of 17O2 by using separate rf channels on the MR spectrometer equipped with multiple receivers. In principle, the Ca(t) function is determined by the metabolic H217O generated in the entire animal body and the equilibration of this H217O with the blood compartment. Considering that the oxygen consumption rate is constant (i.e., time independent) in all tissues during the short inhalation time, one can approximate Ca(t) as a linear function of time [i.e., Ca(t) = At, where A is a constant]. This approximation was supported by our experimental results (see Fig. 5 presented later) and the literature. Then, the solution of Eq. 3 for CMRO2 is
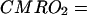 |
4 |
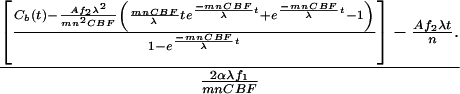 |
Therefore, according to Eq. 4, the CMRO2 value can be calculated by experimental data on CBF, A, n, Cb(t), and other known constants (f1, f2, m, α, and λ) for each data point measured at different inhalation times for each voxel. All CMRO2 values reported (except for those in Fig. 5) are averages of the values calculated as a function of time, excluding the first two time points because of their relatively large measurement errors and the fact that they may represent transient values during the approach to a steady-state rate of H217O production by oxidative phosphorylation. Such transient values are expected during the build-up of 17O2 in the brain at values that exceed the Km of cytochrome oxidase for oxygen. Ability to obtain measurements as a function of time and the use of only the time independent values avoids this complication.
Figure 5.

(A) Natural abundance 17O spectrum of H217O in the rat carotid artery blood (7 μl) obtained before inhalation of 17O2. (B) Time course of 17O MR signals of Ca(t) in one carotid artery (●) and Cb(t) from a representative voxel in the rat brain (□) during inhalation of 17O2. (C) Plot of the calculated CMRO2 values using the complete modeling as described by Eq. 4 as a function of time.
CBF values can be determined from the “washout” rate of the tracer H217O in the brain tissue following a rapid bolus injection of H217O through one carotid artery (32) according to Eq. 5:
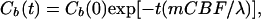 |
5 |
where Cb(0) is the cerebral H217O concentration after the arrival of all labeled water after the bolus injection. Fitting the exponential decay of the H217O washout curve according to Eq. 5 gives the CBF value.
Materials and Methods
NMR Methods.
All NMR experiments were performed on a 9.4T/31-cm bore magnet (Magnex Scientific, Abingdon, U.K.) interfaced to a Unity INOVA console (Varian). A multinuclear head surface-coil probe consisting of a four-turn oval-shaped 17O coil (≈ 1 cm × 2 cm; 54.25 MHz) and a large butterfly-shaped 1H coil (400 MHz) was used.
A TURBOFLASH (fast low-angle shot) image sequence was used to acquire scout images [TR/TE = 8 ms/4 ms, field of view (FOV) = 3 cm × 3 cm, and image matrix size = 128 × 128]. The spatial localization of 17O MRS was achieved by using the 3D Fourier Series Window MRS imaging technique (32, 37). In this method, the k-space sampling is weighted according to the Fourier coefficients of a predetermined voxel shape to achieve an optimal filter applied at the acquisition stage. A cylindrical voxel shape (circular shape on the coronal orientation) was used in this study (37). Short T1 value (4.5 ms) of cerebral H217O in the rat allows rapid signal acquisition, and ultimately, more signal averages within the same sampling time (32). Therefore, a short TR of 12 ms (≈3T1) was used for gaining SNR. The acquisition time for each 3D 17O-MRS image was 11 s (total scan number = 948; rf pulse width = 50 μs, spectral width = 30 kHz; FOV = 18 × 18 × 15 mm3, 9 × 9 × 5 phase encodes). Phase-encoding gradient was 0.35 ms long and had a half-sine waveform, which allowed for a short gradient echo time (TE = 0.4 ms). This short TE is critical for minimizing signal loss because of the relatively fast transverse relaxation of the 17O magnetization (T2 = 3.0 ms at 9.4T; ref. 32). The peak strength of the phase-encoding gradient increment was 0.09–0.11 G/cm. The image voxel size was 27 μl. A 17 × 17 × 9 matrix of free induction decay (FIDs) were generated from the original 9 × 9 × 5 phase encode data for each 3D 17O image. The FIDs were zero-filled, and a 100-Hz line broadening (LB) was used before fast Fourier transformation for SNR enhancement. The 17O NMR signals from each voxel were quantified by measuring the H217O resonance peak intensities.
General Animal Preparation.
Male Sprague–Dawley rats (250–300 g body weight) were anesthetized with ≈2% (vol/vol) isoflurane in a mixture of O2 and N2O gases (2:3) during surgery. After oral intubations, femoral artery and vein were catheterized for physiological monitoring, blood sampling, and chemical administration. After surgery, anesthesia was switched to α-chloralose by using continuous infusion of 0.4 ml/hr of 15 mg/ml α-chloralose solution. Blood gases were sampled for monitoring physiological conditions [pH = 7.35 ± 0.03, pCO2 = 42.1 ± 3.0 mmHg, pO2 = 124 ± 16 mmHg (1 mmHg = 133 Pa)]. The arterial blood pressure (80–110 mmHg) and EtCO2 level (≈3%) were monitored throughout the experiment. The rectal temperature of the rats was maintained at 37 ± 1°C by using a heated water blanket. All animal surgical procedures and experimental protocols were conducted under the guidelines of the National Institutes of Health and the Institutional Animal Care and Use Committee of the University of Minnesota.
CBF Measurements.
For CBF measurements, one external carotid artery was catheterized with a PE-50 tubing for gaining access to the internal carotid artery without interrupting blood circulation into the brain (32). A 0.05–0.1 ml 50%-enriched H217O was rapidly injected into the brain through the internal carotid artery for CBF measurements using 17O MRS imaging (32).
Ca(t) Measurements.
An implanted vascular rf coil was designed and constructed for continuously detecting the 17O NMR signal changes of H217O in the rat carotid artery at 9.4T. This coil was based on a modified solenoid coil design combined with an rf shielding.§ The rf shielding ensured that the NMR signal detected by the implanted coil was only attributed by the artery blood (≈7 μl) without contaminations from surrounding tissues. Therefore, additional spatial localization was not necessary for determining Ca(t). In addition, the rf shielding minimized the electromagnetic coupling between the implanted 17O coil and the head 17O surface coil tuned at the same operating frequency (two-coil arrangement), which allowed simultaneous measurements of both Ca(t) and Cb(t) by using two receiver channels with the same temporal resolution (11 s).
17O2 Inhalation Experiments.
For 17O2 inhalation studies, the 17O-labeled O2 gas with 58.2–72.1% enrichment (Isotec) was mixed with N2O gas (≈2:3) and then stored in a cylindrical gas reservoir. The 3D 17O MRS imaging acquisitions were started with the normal gas mixture first for 2 min; these natural abundance 3D 17O MRS images were used as references for quantifications of metabolic H217O concentrations in the rat brain during and after the 17O2 inhalation. Subsequently, the respiration gas was quickly switched to the 17O2-labeled gas mixture. After 2 min of 17O2 inhalation, the gas line was switched back to the normal gas mixture, and the 3D 17O MRS imaging acquisitions were continued throughout the procedure; a total of 100 images were collected in about 19 min.
Determination of Constant n.
The ratio between the washout rate of the metabolic H217O after the cessation of 17O2 inhalation and the washout rate of the H217O trace from the H217O bolus injected through the rat internal carotid artery gave the value of constant n for each voxel.
All results are presented as mean ± SD.
Results
The multinuclear surface-coil probe was used to collect a series of global 17O and 1H spectra from the natural abundance water in the rat brain to estimate the relative signal fluctuations between 17O and 1H NMR. Two stacked plots of 25 17O and 1H spectra acquired consecutively by using the 17O coil and the much larger 1H coil, respectively, are shown in Fig. 1. Although the number of acquisitions (NT) for each spectrum was different for the 17O and 1H nuclei, the repetition time (TR) was ≈3T1 in both cases; the total acquisition time was 6.2 s for all spectra. Under this condition, we found that the signal fluctuations in the consecutively acquired spectra for the two nuclei were comparable (SD = 0.20% for 1H spectra; SD = 0.35% for 17O spectra). This result demonstrates that with respect to the capability for detecting fractional changes in NMR signal intensity, the 17O method is comparable to the 1H method.
Figure 1.

(A) Stacked plot of 25 1H NMR spectra of natural abundance water in the rat brain acquired and processed with the following parameters: NT = 1, TR = 6.2s, LB = 25Hz (≈ half linewidth). The SD of the 1H water peak height in these spectra is 0.20%. (B) Stacked plot of 25 17O NMR spectra of natural abundance water in the rat brain acquired and processed with the following parameters: NT = 512, TR = 12ms for each scan, LB = 100Hz (≈ half linewidth); the SD of the 17O water peak height in these spectra is 0.35%. The total acquisition times for each 1H and 17O spectra were the same (6.2 s).
The 3D 17O MRS imaging technique was applied to measure the natural abundance H217O distribution in the rat brain with high temporal resolution of 11 s and spatial resolution (27 μl voxel size). Fig. 2A shows representative data obtained from three adjacent coronal images in a rat brain showing anatomical images acquired by the 1H coil and the 17O MRS images of natural abundance H217O acquired by the 17O coil. Fig. 2B displays one 17O chemical shift image from the middle image slice as shown in Fig. 2A. The spatial distributions of 17O NMR signal intensity are not uniform because of the inhomogeneous B1 of the 17O surface coil. Nevertheless, excellent SNR was achieved with a relatively short acquisition time at 9.4T, especially for the central voxels where SNR was optimized by the B1 profile of the 17O coil (SNR ≈ 40:1).
Figure 2.

(A) 3D 17O brain images of natural abundance H217O from three adjacent slices (Left, color images) and corresponding anatomical images (Right, gray images) in the coronal orientation from a representative rat. (B) Chemical shift image of natural abundance H217O from Middle as shown in A.
The large sensitivity gained at ultra-high field revealed the potential of 17O MRS imaging for detecting small dynamic changes of cerebral H217O in small animal brains during a short inhalation of 17O2. Fig. 3 demonstrates stacked plots of 17O spectra of cerebral H217O from one representative voxel before (natural abundance), during, and after a 2-min inhalation of 17O2 (72.1% 17O enrichment). It is clearly evident that the 17O signal intensity of cerebral H217O in the entire measurement is characterized by three distinct phases: (i) constant before the 17O2 inhalation; (ii) approximately linearly increasing during the 17O2 inhalation; and (iii) decreasing after the cessation of 17O2 inhalation, approximately exponentially, and reaching a new steady state within a short time (<10 min).
Figure 3.

Stacked plots of the cerebral H217O spectra from one representative voxel (27 μl voxel size) as indicated in the anatomical image (Left Inset) acquired before (natural abundance), during (as indicated by the dark bar under the stacked plots) and after a 2-min 17O2 inhalation.
When the cerebral H217O concentrations reached a new steady state after the 17O2 inhalation experiment, the CBF measurement was performed by a rapid bolus injection of H217O into one internal carotid artery. The dynamic distribution of cerebral H217O signal during the washout period was monitored by using 3D 17O MRS imaging. The washout curves excluding the first data point were fitted to an exponential decay function according to Eq. 5, and these fits yielded CBF values. Fig. 4 demonstrates one example of CBF measurement obtained from the same rat and image voxel used in Fig. 3 showing the stacked plot of 17O spectra and the exponential fit. Because the H217O bolus was injected into one of the internal carotid arteries, and the H217O tracer was mainly distributed into the ipsilateral hemisphere, CBF and, subsequently, CMRO2 quantifications were performed only for the 17O image voxels in the ipsilateral hemisphere of the rat brain.
Figure 4.

(A) Stacked plots of the cerebral H217O spectra from one voxel (used in Fig. 3) after a fast bolus injection of H217O into the rat brain through an internal carotid artery. (B) Exponential decay fitting of the H217O washout curve for calculating CBF (0.56 ml/g/min for this voxel).
Fig. 5A illustrates the natural abundance H217O spectrum from the blood in the rat carotid artery. Fig. 5B plots the results of simultaneous measurements of Ca(t) change in one carotid artery and Cb(t) change in a voxel in the ipsilateral hemisphere brain during an inhalation of 17O2. The Ca(t) data were fitted by a linear function for calculating the value of constant A. Finally, the values of Cb(t), CBF, and n from each voxel and the value of A measured from each 17O inhalation measurement were used to calculate CMRO2 according to Eq. 4. Fig. 5C demonstrates one example of CMRO2 calculations as a function of time from a single voxel. It is evident in Fig. 5C that the CMRO2 value is independent of the inhalation time if the initial two points are excluded. The averaged CMRO2 values (excluding two initial points) and CBF values from seven measurements are 2.19 ± 0.14 μmol/g/min and 0.53 ± 0.07 ml/g/min, respectively, in the rat brains anesthetized with α-chloralose.
The mean CBF and n values obtained from the entire hemisphere ipsilateral to the internal carotid artery where a H217O bolus was injected were subsequently used to calculate CMRO2 values for both ipsilateral and contralateral hemispheres approximately and to obtain 3D CMRO2 images. This approximation was made based on the fact that the CBF distribution in the hemisphere where it was measured was relatively uniform and that two hemispheres should be equivalent with respect to CBF. Fig. 6 illustrates three adjacent CMRO2 images in the coronal orientation from a representative rat brain. The inhomogeneous signal intensity caused by the surface coil sensitivity profile (Fig. 2) is not reflected in the CMRO2 maps, as expected.
Figure 6.

3D coronal CMRO2 images of rat brain measured during a 2-min 17O2 inhalation.
Discussion
The 17O T1 value in H217O is less than 5 ms in the rat brain at 9.4T (32). This short T1 permits much faster data acquisitions, more signal averages, and, consequently, a high SNR per unit of time. This advantage is critical for the success of CMRO2 measurement using direct 17O detection. The comparable fractional signal fluctuations between consecutively acquired 1H and 17O NMR spectra (Fig. 1) indicated that direct 17O detection of changes in metabolic H217O signal is likely to be more sensitive than the indirect 17O detection through 1H NMR. This notion is based on the fact that the fractional change of water proton signal caused by a 2-min 17O2 inhalation at basal condition is estimated to be 0.15–0.30% (29), which is difficult to detect reliably because this change is comparable to the signal fluctuations in consecutively acquired 1H spectra or images. These signal fluctuations are not dominated by the inherent SNR of a single spectrum or image but are mainly determined by the physiological processes of breathing, heart pulsation, and vasomotion, as well as NMR scanner instability. In contrast, 17O NMR has a much larger dynamic range for detecting the changes of metabolic H217O (20–40%) during the same 17O2 inhalation. This difference is the major merit that makes 17O NMR more suitable for CMRO2 imaging.
The 17O approach has several major advantages for CMRO2 imaging in comparison with other existing methods. First, 17O NMR can specifically detect metabolic H217O without confounding signals from the 17O2 bound to hemoglobin or 17O2 dissolved in tissue space. When bound to hemoglobin, the 17O2 resonance is broadened beyond detection because of the slow rotational motion for the large molecular weight oxyhemoglobin complex. 17O2 as an unbound molecule in gas phase or dissolved in water is strongly paramagnetic because of its unpaired electrons and, hence, undetectable because of the dipolar coupling between the electrons and the nucleus. As a result, 17O NMR avoids the complication in the calculation of CMRO2 encountered in the PET methodology. In addition, unlike PET, the 17O isotope is stable and nonradioactive, and the experimental procedure of 17O2 inhalation does not introduce any physiological perturbation and complexities in the animal. These merits can significantly simplify the modeling of CMRO2 calculation as well as provide an approach for imaging CMRO2 in living organs with minimal biological risk. Second, the natural abundance signal of H217O, which can be accurately imaged, provides an internal reference for rigorously calibrating and calculating the absolute H217O concentration changes for both Cb(t) and Ca(t) during 17O2 inhalation; such a quantification is crucial for determining absolute CMRO2 values and is independent of the inhomogeneities of 17O coil sensitivity. Third, there is only a single and well defined 17O resonance peak of H217O, thus making spectral analysis simple. Fourth, the 17O resonance peak is relatively broad and has a low resonance frequency (only 14% of the 1H resonance frequency); hence, it is significantly less sensitive to static magnetic field inhomogeneities. Fifth, after the cessation of 17O2 inhalation, the cerebral H217O concentration reaches a new steady state within a short time (6–10 min); this fast recovery should allow repeated CMRO2 measurements in the same subject and same experimental session. This capability is essential for studying the oxidative metabolism changes related to perturbations in physiology and function, where at least two measurements are required under control and perturbation conditions. Finally, at ultra-high fields the 17O NMR sensitivity is adequate to obtain relatively high-resolution images, even relative to the small size of the rat brain, with an 11-s acquisition time; this sensitivity makes the 17O approach possible for imaging CMRO2 within a short measurement duration (2 min).
Because of the technical challenges faced for most existing CMRO2 imaging methods, there is virtually no literature reporting direct measurements of CMRO2 in the rat brain. For instance, the image voxel size of most conventional PET scanners is on the order of several milliliters (e.g., ref. 38); this size is already larger than the size of the entire rat brain. Nevertheless, we compared our result of CMRO2 measurement by using the 17O NMR approach with the literature results obtained by indirect methods under similar condition. Based on an autoradiographic approach (39), cerebral metabolic rate of glucose (CMRglc) was recently determined to be ≈0.37 μmol/g/min in the somatosensory and motor cortices of rats anesthetized with α-chloralose. By using the CMRO2/CMRglc ratio of 5.5 (2), a CMRO2 value of 2.04 μmol/g/min is calculated from this CMRglc measurement, which is in excellent agreement with our 17O-based CMRO2 result (2.19 ± 0.14 μmol/g/min). The CBF value of 0.58–0.68 ml/g/min reported in the same autoradiographic study (39) is also close to the mean CBF value of 0.53 ± 0.07 ml/g/min measured in our study. The heteronuclear editing MRS and 1H MRI methods covering a large range of cortical activity at different anesthesia conditions had been used to measure CMRO2 and CBF values in the sensory motor cortex of mature rats, and a linear correlation between CMRO2 and CBF was observed (CMRO2 = 3.76⋅CBF + 0.18, R2 = 0.99; ref. 40). The estimated CMRO2 from this relation is 2.16 μmol/g/min if the CBF value observed in our study (0.53 ml/g/min) is applied. This CMRO2 value is again in excellent agreement with our result. These comparisons provide mutual support between our direct CMRO2 measurements and other indirect measurements reported in the literature and support the validity of the 3D CMRO2 imaging using 17O NMR.
The CMRO2 measurements presented in this paper involved two invasive procedures: one for determining Ca(t) by means of the implanted 17O rf coil, and the second for measuring CBF by an intra-arterial catheter and bolus injection of H217O. The CBF measurement can be performed noninvasively by using the arterial spin tagging MR approaches (41). The implanted rf coil, on the other hand, is not practical for routine measurements of CMRO2 by using small experimental animal models (e.g., rat or mouse), nor is it suitable for human applications. Therefore, it would be important to explore further the feasibility of the 17O NMR approach for reliably imaging CMRO2 without resorting to invasive Ca(t) measurements. A successful outcome from such a study will establish a completely noninvasive 17O approach for CMRO2 imaging.
In comparison with most existing approaches, the high-field 17O NMR method requires a significantly short measurement time for imaging CMRO2 and can be performed within a 2-min 17O2 inhalation period. However, this method assumes that CMRO2 is constant during the measurement. SNR considerations suggest that the CMRO2 measurement time can further shorten at 9.4T or higher magnetic fields. Although rapid dynamic changes of CMRO2 that occur faster than this time-scale will be beyond the reach of this method, this time-scale is faster than any other technique currently available for assessing CMRO2, especially at the spatial resolution available from the 17O NMR approach.
Conclusion
We have successfully developed a comprehensive technique by using 17O MRS imaging at ultra-high field for fast imaging of CMRO2 in the rat brain and validated it with complementary measurements. The overall results from our study provide an essential step toward the goal for developing a robust, fast, reliable, and noninvasive 17O NMR approach for imaging the rate of oxidative metabolism for both animal and human brains and, potentially, for other living organs. Such an approach would be useful for studying the central role of oxidative metabolism in brain function under normal and diseased conditions, as well as for understanding the mechanisms underlying functional MRI (42).
Acknowledgments
We thank Dr. Jae-Hwan Kwag and John Strupp for their technical assistance and Dr. Haiying Liu for scientific discussions. This work was supported by National Institutes of Health Grants NS41262, NS38070, NS39043, and EB00329; National Research Resource Grant P41 RR08079 (from the National Institutes of Health); the MIND Institute; and the W. M. Keck Foundation.
Abbreviations
- CMRO2
cerebral metabolic rate of oxygen
- CBF
cerebral blood flow
- PET
positron emission tomography
- 3D
three-dimensional
- MRS
magnetic resonance spectroscopic
- SNR
signal-to-noise ratio
Footnotes
Zhu, X. H., Lei, H., Zhang, Y., Zhang, X. L., Zhang, N. Y., Ugurbil, K. & Chen, W. (2002) Proc. Int. Soc. Mag. Res. Med. 10, 1094 (abstr.).
Zhang, X. L., Tian, R. X., Zhu, X. H, Zhang, Y., Merkle, H. & Chen, W. (2002) Proc. Int. Soc. Mag. Res. Med. 10, 889 (abstr.).
References
- 1.Attwell D, Laughlin S B. J Cereb Blood Flow Metab. 2001;21:1133–1145. doi: 10.1097/00004647-200110000-00001. [DOI] [PubMed] [Google Scholar]
- 2.Siesjo B K. Brain Energy Metabolism. New York: Wiley; 1978. [Google Scholar]
- 3.Maurer I, Zierz S, Moller H. Schizophr Res. 2001;48:125–136. doi: 10.1016/s0920-9964(00)00075-x. [DOI] [PubMed] [Google Scholar]
- 4.Wong-Riley M, Antuono P, Ho K C, Egan R, Hevner R, Liebl W, Huang Z, Rachel R, Jones J. Vision Res. 1997;37:3593–3608. doi: 10.1016/S0042-6989(96)00210-6. [DOI] [PubMed] [Google Scholar]
- 5.Maurer I, Zierz S, Moller H J. Neurobiol Aging. 2000;21:455–462. doi: 10.1016/s0197-4580(00)00112-3. [DOI] [PubMed] [Google Scholar]
- 6.Frackowiak R S, Herold S, Petty R K, Morgan-Hughes J A. Brain. 1988;111:1009–1024. doi: 10.1093/brain/111.5.1009. [DOI] [PubMed] [Google Scholar]
- 7.Beal M F. Ann Neurol. 1992;31:119–130. doi: 10.1002/ana.410310202. [DOI] [PubMed] [Google Scholar]
- 8.Harrison R V, Harel N, Panesar J, Mount R J. Cereb Cortex. 2002;12:225–233. doi: 10.1093/cercor/12.3.225. [DOI] [PubMed] [Google Scholar]
- 9.Hoppeler H, Kayar S R. News Physiol Sci. 1988;3:113–116. [Google Scholar]
- 10.Hyder F, Chase J R, Behar K L, Mason G F, Siddeek M, Rothman D L, Shulman R G. Proc Natl Acad Sci USA. 1996;93:7612–7617. doi: 10.1073/pnas.93.15.7612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chen W, Zhu X H, Gruetter R, Seaquist E R, Ugurbil K. Magn Reson Med. 2001;45:349–355. doi: 10.1002/1522-2594(200103)45:3<349::aid-mrm1045>3.0.co;2-8. [DOI] [PubMed] [Google Scholar]
- 12.Raichle M E. In: Handbook of Physiology–The Nervous System. Mountcastle V B, Plum F, Geiger S R, editors. Washington, DC: Am. Physiol. Soc.; 1987. pp. 643–674. [Google Scholar]
- 13.Kety S S, Schmidt C F. J Clin Invest. 1948;27:476–483. doi: 10.1172/JCI101994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ter-Pogossian M M, Eichling J O, Davis D O, Welch M J. J Clin Invest. 1970;49:381–391. doi: 10.1172/JCI106247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mintun M A, Raichle M E, Martin W R, Herscovitch P. J Nucl Med. 1984;25:177–187. [PubMed] [Google Scholar]
- 16.Lenzi G L, Frackowiak R S, Jones T, Heather J D, Lammertsma A A, Rhodes C G, Pozzilli C. Eur Neurol. 1981;20:285–290. doi: 10.1159/000115248. [DOI] [PubMed] [Google Scholar]
- 17.Rothman D L, Novotny E J, Shulman G I, Howseman A M, Petroff O A, Mason G, Nixon T, Hanstock C C, Prichard J W, Shulman R G. Proc Natl Acad Sci USA. 1992;89:9603–9606. doi: 10.1073/pnas.89.20.9603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gruetter R, Seaquist E R, Ugurbil K. Am J Physiol. 2001;281:E100–E112. doi: 10.1152/ajpendo.2001.281.1.E100. [DOI] [PubMed] [Google Scholar]
- 19.Mason G F, Rothman D L, Behar K L, Shulman R G. J Cereb Blood Flow Metab. 1992;12:434–447. doi: 10.1038/jcbfm.1992.61. [DOI] [PubMed] [Google Scholar]
- 20.Mateescu G D, Yvars G M, Dular T. In: Water and Ions in Biological Systems. Lauger P, Packer L, Vasilescu V, editors. Basel: Birkhauser; 1988. pp. 239–250. [Google Scholar]
- 21.Mateescu G D, Yvars G, Pazara D I, Alldridge N A, LaManna J C, Lust D W, Mattingly M, Kuhn W. In: Synthesis and Application of Isotopically Labeled Compounds. Baillie T A, Jones J R, editors. Amsterdam: Elsevier; 1989. pp. 499–508. [Google Scholar]
- 22.Hopkins A L, Barr R G. Magn Reson Med. 1987;4:399–403. doi: 10.1002/mrm.1910040413. [DOI] [PubMed] [Google Scholar]
- 23.Kwong K K, Hopkins A L, Belliveau J W, Chesler D A, Porkka L M, McKinstry R C, Finelli D A, Hunter G J, Moore J B, Barr R G, Rosen B R. Magn Reson Med. 1991;22:154–158. doi: 10.1002/mrm.1910220116. [DOI] [PubMed] [Google Scholar]
- 24.Arai T, Nakao S, Mori K, Ishimori K, Morishima I, Miyazawa T, Fritz-Zieroth B. Biochem Biophys Res Commun. 1990;169:153–158. doi: 10.1016/0006-291x(90)91447-z. [DOI] [PubMed] [Google Scholar]
- 25.Pekar J, Ligeti L, Ruttner Z, Lyon R C, Sinnwell T M, van Gelderen P, Fiat D, Moonen C T, McLaughlin A C. Magn Reson Med. 1991;21:313–319. doi: 10.1002/mrm.1910210217. [DOI] [PubMed] [Google Scholar]
- 26.Fiat D, Ligeti L, Lyon R C, Ruttner Z, Pekar J, Moonen C T, McLaughlin A C. Magn Reson Med. 1992;24:370–374. doi: 10.1002/mrm.1910240218. [DOI] [PubMed] [Google Scholar]
- 27.Pekar J, Sinnwell T, Ligeti L, Chesnick A S, Frank J A, McLaughlin A C. J Cereb Blood Flow Metab. 1995;15:312–320. doi: 10.1038/jcbfm.1995.36. [DOI] [PubMed] [Google Scholar]
- 28.Ronen I, Navon G. Magn Reson Med. 1994;32:789–793. doi: 10.1002/mrm.1910320616. [DOI] [PubMed] [Google Scholar]
- 29.Ronen I, Merkle H, Ugurbil K, Navon G. Proc Natl Acad Sci USA. 1998;95:12934–12939. doi: 10.1073/pnas.95.22.12934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Reddy R, Stolpen A H, Leigh J S. J Magn Reson. 1995;108:276–279. doi: 10.1006/jmrb.1995.1133. [DOI] [PubMed] [Google Scholar]
- 31.Fiat D, Dolinsek J, Hankiewicz J, Dujovny M, Ausman J. Neurol Res. 1993;15:237–248. doi: 10.1080/01616412.1993.11740143. [DOI] [PubMed] [Google Scholar]
- 32.Zhu X H, Merkle H, Kwag J H, Ugurbil K, Chen W. Magn Reson Med. 2001;45:543–549. doi: 10.1002/mrm.1073. [DOI] [PubMed] [Google Scholar]
- 33.Altman P L. Biological Handbook Series. Washington, DC: Fed. Am. Soc. Exper. Biol.; 1961. p. 14. [Google Scholar]
- 34.Herscovitch A, Raichle M E. J Cereb Blood Flow Metab. 1985;5:65–69. doi: 10.1038/jcbfm.1985.9. [DOI] [PubMed] [Google Scholar]
- 35.Herscovitch P, Raichle M E, Kilbourn M R, Welch M J. J Cereb Blood Flow Metab. 1987;7:527–542. doi: 10.1038/jcbfm.1987.102. [DOI] [PubMed] [Google Scholar]
- 36.Ohta S, Meyer E, Thompson C J, Gjedde A. J Cereb Blood Flow Metab. 1992;12:179–192. doi: 10.1038/jcbfm.1992.28. [DOI] [PubMed] [Google Scholar]
- 37.Hendrich K, Hu X, Menon R, Merkle H, Camarata P, Heros R, Ugurbil K. J Magn Reson. 1994;105:225–232. doi: 10.1006/jmrb.1994.1128. [DOI] [PubMed] [Google Scholar]
- 38.Fox P T, Raichle M E, Mintun M A, Dence C. Science. 1988;241:462–464. doi: 10.1126/science.3260686. [DOI] [PubMed] [Google Scholar]
- 39.Nakao Y, Itoh Y, Kuang T Y, Cook M, Jehle J, Sokoloff L. Proc Natl Acad Sci USA. 2001;98:7593–7598. doi: 10.1073/pnas.121179898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hyder F, Kennan R P, Kida I, Mason G F, Behar K L, Rothman D. J Cereb Blood Flow Metab. 2000;20:485–498. doi: 10.1097/00004647-200003000-00007. [DOI] [PubMed] [Google Scholar]
- 41.Detre J A, Zhang W, Roberts D A, Silva A C, Williams D S, Grandis D J, Koretsky A P, Leigh J S. NMR Biomed. 1994;7:75–82. doi: 10.1002/nbm.1940070112. [DOI] [PubMed] [Google Scholar]
- 42.Barinaga M. Science. 1997;276:196–198. doi: 10.1126/science.276.5310.196. [DOI] [PubMed] [Google Scholar]