

Abstract
Using a specific recombination assay, we show in the plant Arabidopsis thaliana that AtRad1 protein plays a role in the removal of non-homologous tails in homologous recombination. Recombination in the presence of non-homologous overhangs is reduced 11-fold in the atrad1 mutant compared with the wild-type plants. AtRad1p is the A. thaliana homologue of the human Xpf and Saccharomyces cerevisiae Rad1 proteins. Rad1p is a subunit of the Rad1p/Rad10p structure-specific endonuclease that acts in nucleotide excision repair and inter-strand crosslink repair. This endonuclease also plays a role in mitotic recombination to remove non-homologous, 3′-ended overhangs from recombination intermediates. The Arabidopsis atrad1 mutant (uvh1), unlike rad1 mutants known from other eukaryotes, is hypersensitive to ionizing radiation. This last observation may indicate a more important role for the Rad1/Rad10 endonuclease in recombination in plants. This is the first direct demonstration of the involvement of AtRad1p in homologous recombination in plants.
Introduction
Plants need sunlight for photosynthesis; however, they are thus exposed to the harmful effects of solar ultraviolet (UV) radiation. The two major DNA lesions produced by UV irradiation are cyclobutane pyrimidine dimers (CPDs) and pyrimidine (6,4) pyrimidinone dimers (reviewed by Mitchell and Nairn, 1989; Friedberg et al., 1995). If the lesions are not removed, they both block DNA replication and transcription (Protic-Sabljic and Kraemer, 1985) and also cause mutations.
Nucleotide excision repair (NER) is responsible for processing DNA lesions that generate major distortion in the helical DNA structure, such as these UV photoproducts and bulky covalent chemical adducts (reviewed by Friedberg et al., 1995). The biochemical steps constituting this repair pathway have been characterized in Escherichia coli, yeast and mammalian systems (Aboussekhra and Wood, 1994; Laat et al., 1999), but much less information is available for plants (for reviews, see Britt, 1999; Tuteja et al., 2001). Following specific recognition of the damage and formation of a pre-incision complex, a singlestrand oligonucleotide containing the damage site is excised and the resulting gap filled in and sealed (Friedberg et al., 1995). In Saccharomyces cerevisiae, two structurespecific endonucleases nick the damaged DNA strand, Rad1p/Rad10p 5′ and Rad2p 3′ of the damaged site (Davies et al., 1995). The Rad1p/Rad10p endonuclease consists of a heterodimer of Rad1p and Rad10p (Bailly et al., 1992; Bardwell et al., 1992, 1993) and has been shown to incise DNA specifically at 5′-doublestrand/3′-single-strand junctions (Tomkinson et al., 1993; Bardwell et al., 1994).
Rad1 and Rad10 proteins exhibit significant amino acid conservation with the mammalian Xpf and Ercc1 proteins, respectively (Brookman et al., 1996; Sijbers et al., 1996). The gene RAD1 also has homologues in Schizosaccharomyces pombe (rad16; Carr et al., 1994) and Drosophila melanogaster (MEI-9; Sekelsky et al., 1995). AtRAD1, a homologue of RAD1, has recently been described as a single-copy gene in the plant Arabidopsis thaliana (Fidantsef et al., 2000; Gallego et al., 2000; Liu et al., 2000) and shown to be important for the removal of 6-4 photoproducts from UV-irradiated plants (Liu et al., 2000). These authors have also shown that the Arabidopsis uvh1 mutant carries a single base pair substitution in the atrad1 gene, leading to missplicing of the atrad1 mRNA in mutant plants (Liu et al., 2000). In vitro assays have confirmed this role in NER and also indicated a possible role for AtRad1 protein (AtRad1p) in the repair of oxidative DNA damage (Li et al., 2002). A single RAD10 homologue is found in the fully sequenced Arabidopsis genome (locus At3g05210), and a RAD10 homologue has also been described in lily (Xu et al., 1998).
The Ercc1/Xpf endonuclease plays an essential role in NER. Ercc1−/− mice show a more severe phenotype than other NER-deficient mouse models, apparently associated with a premature replicative senescence in the Ercc1−/− cells (Weeda et al., 1997). Moreover, Xpf-deficient mammalian cells and AtRad1p-deficient plants, unlike other NER-deficient cells, are sensitive to crosslinking agents (Westerveld et al., 1984; Hoy et al., 1985; Gallego et al., 2000). Unlike xpf and yeast rad1 and rad10 mutants, the atrad1 mutant (uvh1) is also hypersensitive to ionizing radiation (Harlow et al., 1994; Jenkins et al., 1995; Jiang et al., 1997; reviewed by Paques and Haber, 1997). These observations underline the importance of a potential role for AtRad1p in recombination, as well as its role in NER. A number of reports have shown Rad1p and Rad10p to have a role in mitotic recombination (reviewed by Paques and Haber, 1997). Mutations in RAD1 and RAD10 reduce intra-chromosomal recombination between directly repeated sequences 8-fold and also decrease the efficiency of homologous integration of linear DNA fragments and circular plasmids (Klein, 1988; Schiestl and Prakash, 1988, 1990; Prado and Aguilera, 1995). The rad1 and rad10 mutations are epistatic to one another (Schiestl and Prakash, 1988, 1990; Saffran et al., 1994), so the recombination role depends upon the heterodimer formation and most likely on its endonuclease function. Yeast excision repair mutants other than rad1 and rad10 do not show these recombinational phenotypes, further demonstrating the specific role of Rad1p/Rad10p in recombination (Ivanov and Haber, 1995).
Intra-chromosomal recombination between tandem, direct repeats occurs most efficiently via single-strand annealing (SSA) following the formation of a DNA double-strand break (DSB) between, or within, the repeated sequences (discussed by Paques and Haber, 1997). Exonucleolytic resection of the 5′-ended strands at the DSB produces two complementary, 3′-ended single strands. These strands anneal and, after excision of any non-homologous 3′-ended overhangs and new DNA synthesis, ligation restores two continuous strands. Rad1p and Rad10p are required for the removal of these 3′ non-homologous tails and for the removal of non-homologous sequences during DSB-induced recombination between plasmid inverted repeats (Fishman-Lobell and Haber, 1992; Ivanov and Haber, 1995; Prado and Aguilera, 1995; Saparbaev et al., 1996; Paques and Haber, 1997; Sugawara et al., 1997; Colaiacovo et al., 1999). Similarly, Ercc1p is required to remove non-homologous tails from ends-in gene targeting constructs in CHO cells (Adair et al., 2000; Sargent et al., 2000) and for targeted gene replacement in mouse embryonic stem cells (Niedernhofer et al., 2001). Interestingly, Niedernhofer et al. (2001) suggest that the role of Rad1p/Rad10p in nicking synapsed recombination intermediates leads to the formation of a DSB in the recipient chromosomal DNA, thus permitting insertion of non-homologous DNA into the chromosome.
As part of our analysis of DNA recombination mechanisms in plants, we present here results of a specific assay showing that AtRad1p plays a role in permitting productive recombination between DNA molecules in A. thaliana in the presence of non-homologous tails.
Results and discussion
Over the last decade, a number of studies have contributed to our understanding of the apparent conservation of recombination-associated proteins and recombination mechanisms between plants and other eukaryotes (reviewed in Britt, 1999; Gorbunova and Levy, 1999; Mengiste and Paszkowski, 1999; Vergunst and Hooykaas, 1999; Tuteja et al., 2001). However, the lack of identified mutant plants for recombination functions has meant that experimental verification of the roles of specific proteins in recombination in plants is lagging behind the identification, based on analogy, of the corresponding genes. In the present study, we tested specifically the involvement of the AtRad1p endonuclease in removing non-homologous overhangs to permit productive homologous recombination between linear DNA molecules in A. thaliana. Our assay involves observing transient expression of recombined plasmids following biolistic transformation of Arabidopsis leaves.
AtRad1p is not needed for homologous recombination in the absence of non-homologous overhangs
In this 'no-tails' experiment, we addressed the question of whether AtRad1p is required for recombination between overlapping homologous DNA regions in the absence of non-homologous overhangs (as indicated in Figure 1).
Figure 1.
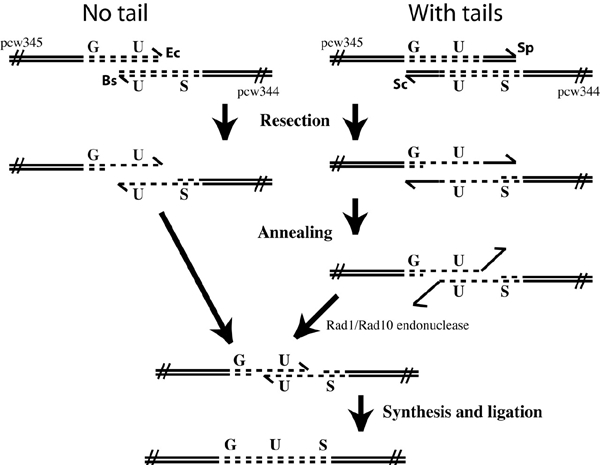
Recombination assay. Linear recombination substrates were prepared from pcw344 and pcw345 plasmids by restriction with enzymes that leave ('with tails') or do not leave ('no tails') non-homologous overhangs. The 'no tails' test was carried out with pcw345/EcoRI and pcw344/BsmI and the 'with tails' test with pcw345/SapI and pcw344/ScaI. The relevant linearized plasmids were used to co-transform leaves from either wild-type or atrad1 plants and the number of recombinant spots per leaf counted. Resection of the linear molecules permits annealing of complementary homologies. The annealed 3′-ended strands of 'with tails' plasmids have unpaired, non-homologous overhangs. The Rad1/Rad10 endonuclease removes these, permitting re-synthesis and ligation to generate the recombined product. Both DNA strands are shown with arrowheads indicating the 3′ ends. Ec, EcoRI; Bs, BsmI; Sp, SapI; Sc, ScaI.
In order to monitor inter-molecular recombination, plasmids pcw344 and pcw345 were linearized with EcoRI and BsmI, respectively, and used to co-transform leaves from wild-type and atrad1 plants (as described in detail in Methods and shown in Figure 1). Plasmid pcw344 contains only the region encoding the C-terminal region of the β-glucuronidase (GUS) gene and the nopaline synthetase (NOS) terminator, whereas pcw345 contains the 35S promoter and the region encoding the N-terminal region of GUS. The overlapping homology region is 590 bp long, which has previously been shown to be sufficient for efficient recombination in analogous assays (Puchta and Hohn, 1991). GUS activity was subsequently detected as blue spots following histochemical treatment. Examples of transformed leaves showing blue GUS spots are shown in Figure 2.
Figure 2.
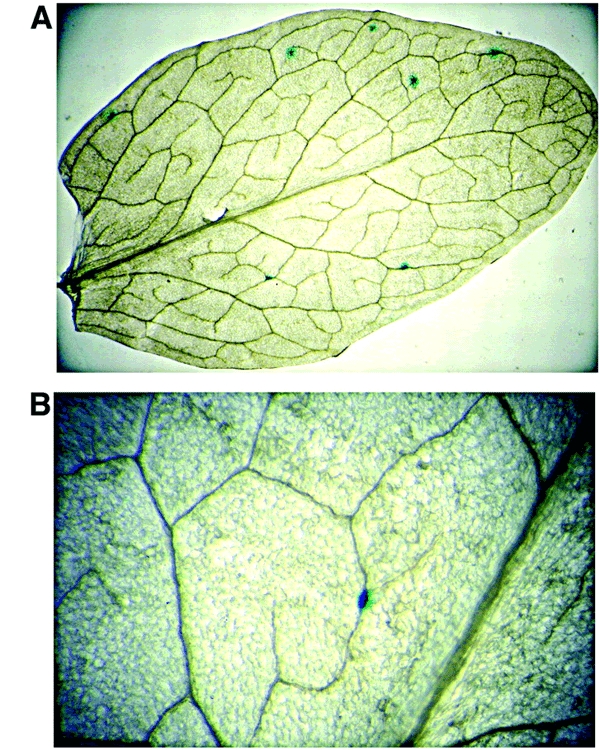
Examples of blue spots after GUS staining: (A) atrad1 leaf transformed by pGUS23 and (B) an example of a recombinant blue spot arising from recombination (atrad1 'no tails').
As shown in Tables 1 and 2 and Figure 3, the frequencies of recombination events do not differ significantly between the two lines. Thus AtRad1p is not required for DSB repair by homologous recombination in the absence of non-homologous overhangs ('no tails').
Table 1.
AtRad1p removes overhanging tails for homologous recombination: numbers of GUS+ blue spots per leaf
| pGUS23 |
No tails |
With tails |
|||
|---|---|---|---|---|---|
| wild type | atrad1 | wild type | atrad1 | wild type | atrad1 |
| 17 | 35 | 7 | 8 | 11 | 1 |
| 24 | 28 | 3 | 7 | 13 | 2 |
| 28 | 23 | 11 | 4 | 10 | 2 |
| 57 | 41 | 6 | 3 | 16 | 0 |
| 30 | 23 | 12 | 9 | 18 | 1 |
| 30 | 31 | 6 | 5 | 14 | 0 |
| 36 | 45 | 7 | 6 | 16 | 5 |
| 31 | 21 | 4 | 4 | 15 | 0 |
| 44 | 43 | 5 | 4 | 18 | 0 |
| 21 | 31 | 5 | 5 | 20 | 2 |
| 23 | 2 | 1 | 24 | 2 | |
| 2 | 2 | 12 | 2 | ||
| 1 | 3 | 16 | 0 | ||
| 11 | 4 | 14 | 2 | ||
| 3 | 3 | 10 | 1 | ||
| 3 | 2 | 13 | 1 | ||
| 5 | 1 | 16 | 2 | ||
| 6 | 11 | 1 | |||
| 2 | 8 | 2 | |||
| 4 | 6 | 0 | |||
| 14 | |||||
Wild-type and atrad1 mutant leaves were transformed with pGUS23 (transformation efficiency control) or with 'with tails' or 'no tails' recombination substrates. After histochemical staining, blue spots were counted under a dissecting microscope. Counts are from at least three different transformations.
Table 2.
Atrad1p removes overhanging tails for homologous recombination: χ2 (1 degree of freedom) values for the null hypothesis that all leaves from a given line are from the same population with respect to number of blue spots
| Wild type |
atrad1 |
Chi squared |
|||||||
|---|---|---|---|---|---|---|---|---|---|
| DNA | n | N | X | n | N | X | Reduction | wild type | atrad1 |
| pGUS23 | 11 | 341 | 31.0 | 10 | 321 | 32.0 | 0.97 | 0.82 | 0.40 |
| No tails | 17 | 93 | 5.5 | 20 | 83 | 4.2 | 1.3 | 0.53 | 0.80 |
| With tails | 21 | 295 | 14.1 | 20 | 26 | 1.3 | 10.85 | 17.19 | 20.00 |
Non-parametric statistical analysis was carried out as described in Methods. In the 'with tails' transformation, numbers of recombinants (blue spots) in the atrad1 mutant are significantly (10.85-fold) reduced, permitting the rejection of the null hypothesis. No significant differences are observed in the transformation efficiency (pGUS23) or the 'no tails' control transformations.
n, number of leaves analysed; N, total number of blue spots; X, mean number of recombination events per leaf.
Figure 3.
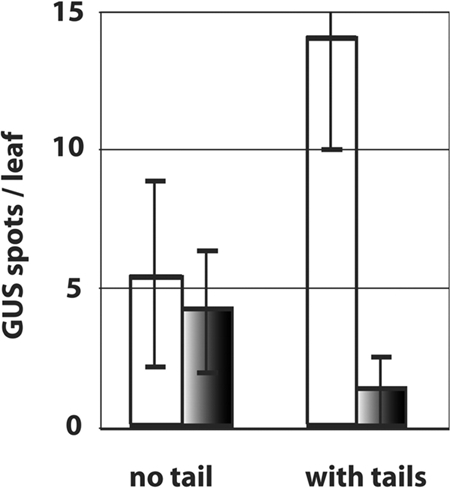
Recombination in atrad1 and wild-type leaves. Effect of the non-homologous tails on the recombination rate in wild-type (white bars) and atrad1 mutant (grey bars) plants. The presence of non-homologous overhangs ('with tails') specifically reduces the frequency of recombinant spots in the atrad1 mutant. No significant difference between mutant and wild-type plants is seen in the absence of the non-homologous overhangs ('no tails'). Error bars are ± 1 standard deviation.
No recombinant spots were detected in control transformations carried out in parallel with the same quantities of the two plasmids individually (data not shown). Transformation efficiency was controlled in parallel by transforming with pGUS23, which contains the entire GUS transcription cassette. In order to produce a quantifiable number of spots, 10-fold less plasmid DNA was used (5 mg versus 50 mg in the recombination assays). The numbers of GUS+ spots obtained with pGUS23 transformation of atrad1 and wild-type leaves were similar; thus, these two plant lines were transformed with similar efficiency (Tables 1 and 2).
AtRad1p is required for homologous recombination in the presence of non-homologous overhangs
In S. cerevisiae, the Rad1p/Rad10p endonuclease is required for the removal of 3′-ended, non-homologous single-stranded tails from SSA intermediates during DSB-induced recombination between direct repeats (Fishman-Lobell and Haber, 1992; Ivanov and Haber, 1995; Prado and Aguilera, 1995; Sugawara et al., 1997).
To test the role of AtRad1p in the elimination of non-homologous tails during SSA, a second recombination test was carried out. atrad1 and wild-type plants were co-transformed with pcw344/ScaI and pcw345/SapI (as described in Methods and shown in Figure 1). The overlapping homology region is 590 bp in both cases, to which pcw344/ScaI adds a non-homologous tail of 816 bp and pcw345/SapI a tail of 233 bp.
The atrad1 mutant plants show significantly (11-fold) less recombinant spots than the wild-type plants ('with tails', Tables 1 and 2 and Figure 3). Thus, AtRad1p is implicated in DSB repair by homologous recombination in the presence of non-homologous tails. We note that, although the absence of AtRad1p strongly reduces recombination in the presence of non-homologous tails, it does not eliminate it altogether in our assay. Thus, a minor, AtRad1p-independent pathway for these events must exist. Another possible factor contributing to these AtRad1-independent events might be due to exonucleolytic action on the linear DNA substrates. This possibility is supported by our observation that, in the wild-type leaves, the 'with tails' constructs give more recombinant spots than the 'no tails' constructs. Thus, the tails may act as a 'buffer' protecting the homologous GUS sequences from exonuclease degradation prior to recombination. That such an exonuclease activity does not affect our conclusions concerning the role of AtRad1p in our experiments is demonstrated by the equivalent recombination efficiencies in the wild-type and mutant leaves with the 'no tails' substrates (Figure 3).
In conclusion, we show here that the Arabidopsis Rad1p homologue acts on recombining DNA molecules to permit productive homologous recombination in the presence of non-homologous DNA overhangs. We are now testing the implication of AtRad1p in other forms of homologous recombination in Arabidopsis, such as gene targeting and spontaneous and induced intra-chromosomal recombination.
Methods
Strains and growth conditions.
Arabidopsis thaliana were ecotype Columbia (Col). The atrad1 UV-hypersensitive mutant (uvh1-1; Harlow et al., 1994) was obtained from the Nottingham Arabidopsis Stock Center (CS3819; NASC: online catalogue available at http://nasc/nott.ac.uk/home.html). After sterilization with sodium hypochlorite (7%, w/v), atrad1 and wild-type seeds (ecotype col4) were sown on germination medium [Murashige and Skoog basal medium with vitamins (Sigma), supplemented with 30 g/l sucrose, pH 5.8]. Plates were incubated in a growth chamber at 22°C (16 h light, 8 h dark). atrad1 mutants were verified both by Southern hybridization and UV hypersensitivity (data not shown).
Plasmid constructs.
As the control for transformation efficiency, we used plasmid pGUS23 (Puchta and Hohn, 1991), which contains the cauliflower mosaic virus (CaMV) 35S promoter, the GUS gene and the NOS transcription terminator.
The plasmid pcw344 was constructed by digesting pGUS23 with NdeI and SnaBI to eliminate the 35S promoter and the GUS N-terminal region. The resulting plasmid was blunt-ended with T4 DNA polymerase, purified through low-melting-point agarose and re-circularized with T4 DNA ligase.
The plasmid pcw345 was obtained by SfuI–EcoRI digestion of plasmid pGUS23C1 (a deletion derivative of pGUS23; Puchta and Hohn, 1991), which contains the 35S promoter, the GUS N-terminal region and the NOS terminator. This eliminated the NOS terminator. The resulting plasmid was purified, blunt-ended and circularized as above
Biolistic transformation and recombination assay.
Leaves from 3- to 4-week-old wild-type and atrad1 mutant plants were placed adaxial side up on solid germination medium in Petri dishes. Gold beads (Sigma; 0.6 μm diameter) were washed in water then rinsed and resuspended in ethanol at 0.1 μg/μl. To 50 μl of beads, 50 μl (50 μg) of plasmid DNA, 250 μl of 2.5 M CaCl2 and 100 μl of 0.1 M spermidine (Sigma) were added. After 5 min on ice, the beads were pelleted with a brief centrifugation and 400 μl of supernatant removed.
For each transformation, 5 μl (10%) of the resuspended, DNA-coated gold beads were used with a helium particle gun (6 bars, 28 kPa, 13 cm from the leaves; Finer et al., 1992). Leaves were then placed in a growth chamber at 22°C (16 h light, 8 h dark) for 1 or 2 days and the GUS activity assay carried out as described by Jefferson et al. (1987). The number of blue spots per leaf was determined by visual observation using a dissecting microscope.
Transformation efficiency was controlled by transforming wild-type and mutant plants with 5 μl (5 μg) of pGUS23. The lower quantity of plasmid DNA (5 μg versus 50 μg in the recombination assays) was used to reduce the number of spots per leaf in order to facilitate counting in the controls.
Plasmids pcw344 and pcw345 were linearized with BsmI or EcoRI, respectively, to prepare the 'no-tail' recombination substrate. 'With tail' substrates were prepared by digesting pcw344 and pcw345 with ScaI or SapI, respectively. In all cases, the overlapping homology region is 590 bp, to which pcw344/ScaI adds a non-homologous tail of 816 bp and pcw345/SapI a tail of 233 bp. In both 'with-tails' and 'no-tails' experiments, the transformation was carried out with 25 μl (25 μg) of each plasmid (pcw344 and pcw345).
Statistical analysis.
The data on the number of recombination events per transformed leaf were analysed non-parametrically (with the kind advice of Dr E.J. Louis, University of Leicester, UK). All leaves analysed from a given line were ranked by the number of blue spots for each set of leaves analysed. On the null hypothesis that the plants analysed from each line belong to the same distribution, half should be below the mid-point of the ranking and half above. χ2 (1 degree of freedom) values were calculated for the wild-type and atrad1 mutants from each type of transformation (Table 2).
Acknowledgments
We thank Biogemma and the laboratory of Michel Bernard (INRA) for kindly letting us use their helium canons.
References
- Aboussekhra A. and Wood R.D. (1994) Repair of UV-damage DNA by mammalian cells and Saccharomyces cerevisiae. Curr. Opin. Genet. Dev., 4, 212–220. [DOI] [PubMed] [Google Scholar]
- Adair G.M., Rolig R.L., Moore-Faver D., Zabelshansky M., Wilson J.H. and Nairn R.S. (2000) Role of ERCC1 in removal of long non-homologous tails during targeted homologous recombination. EMBO J., 19, 5552–5561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailly V., Sommers C.H., Sung P., Prakash L. and Prakash S. (1992) Specific complex formation between proteins encoded by the yeast DNA repair and recombination genes RAD1 and RAD10. Proc. Natl Acad. Sci. USA, 89, 8273–8277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bardwell A.J., Bardwell L., Johnson D.K. and Friedberg E.C. (1993) Yeast DNA recombination and repair proteins Rad1 and Rad10 constitute a complex in vivo mediated by localized hydrophobic domains. Mol. Microbiol., 8, 1177–1188. [DOI] [PubMed] [Google Scholar]
- Bardwell A.J., Bardwell L., Tomkinson A.E. and Friedberg E.C. (1994) Specific cleavage of model recombination and repair intermediates by the yeast Rad1–Rad10 DNA endonuclease. Science, 265, 2082–2085. [DOI] [PubMed] [Google Scholar]
- Bardwell L., Cooper A.J. and Friedberg E.C. (1992) Stable and specific association between the yeast recombination and DNA repair proteins RAD1 and RAD10 in vitro. Mol. Cell. Biol., 12, 3041–3049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Britt A.B. (1999) Molecular genetics of DNA repair in higher plants. Trends Plant Sci., 4, 20–25. [DOI] [PubMed] [Google Scholar]
- Brookman K.W. et al. (1996) ERCC4 (XPF) encodes a human nucleotide excision repair protein with eukaryotic recombination homologs. Mol. Cell. Biol., 16, 6553–6562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr A.M. et al. (1994) The rad16 gene of Schizosaccharomyces pombe: a homolog of the RAD1 gene of Saccharomyces cerevisiae. Mol. Cell. Biol., 14, 2029–2040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colaiacovo M.P., Paques F. and Haber J.E. (1999) Removal of one nonhomologous DNA end during gene conversion by a RAD1- and MSH2-independent pathway. Genetics, 151, 1409–1423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies A.A., Friedberg E.C., Tomkinson A.E., Wood R.D. and West S.C. (1995) Role of the Rad1 and Rad10 proteins in nucleotide excision repair and recombination. J. Biol. Chem., 270, 24638–24641. [DOI] [PubMed] [Google Scholar]
- Fidantsef A.L., Mitchell D.L. and Britt A.B. (2000) The Arabidopsis UVH1 gene is a homolog of the yeast repair endonuclease RAD1. Plant Physiol., 124, 579–586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finer J., Vain P., Jones M. and MacMullen M. (1992) Developement of the particle inflow gun for DNA delivery to plant cells. Plant Cell Rep., 11, 323–328. [DOI] [PubMed] [Google Scholar]
- Fishman-Lobell J. and Haber J.E. (1992) Removal of nonhomologous DNA ends in doublestrand break recombination—the role of the yeast ultraviolet repair gene RAD1. Science, 258, 480–484. [DOI] [PubMed] [Google Scholar]
- Friedberg E.C., Walker G.C. and Siede W. (1995) DNA Repair and Mutagenesis. American Society for Microbiology, Washington, DC, USA. [Google Scholar]
- Gallego F., Fleck O., Li A., Wyrzykowska J. and Tinland B. (2000) AtRAD1, a plant homologue of human and yeast nucleotide excision repair endonucleases, is involved in dark repair of UV damages and recombination. Plant J., 21, 507–518. [DOI] [PubMed] [Google Scholar]
- Gorbunova V. and Levy A.A. (1999) How plants make ends meet: DNA doublestrand break repair. Trends Plant Sci., 4, 263–269. [DOI] [PubMed] [Google Scholar]
- Harlow G.R., Jenkins M.E., Pittalwala T.S. and Mount D.W. (1994) Isolation of uvh1, an Arabidopsis mutant hypersensitive to ultraviolet light and ionizing radiation. Plant Cell, 6, 227–235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoy C.A., Thompson L.H., Mooney C.L. and Salazar E.P. (1985) Defective DNA cross-link removal in Chinese hamster cell mutants hypersensitive to bifunctional alkylating agents. Cancer Res., 45, 1737–1743. [PubMed] [Google Scholar]
- Ivanov E.L. and Haber J.E. (1995) RAD1 and RAD10, but not other excision repair genes, are required for doublestrand break-induced recombination in Saccharomyces cerevisiae. Mol. Cell. Biol., 15, 2245–2251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jefferson R.A., Kavanagh T.A. and Bevan M.W. (1987) GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J., 6, 3901–3907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenkins M.E., Harlow G.R., Liu Z., Shotwell M.A., Ma J. and Mount D.W. (1995) Radiationsensitive mutants of Arabidopsis thaliana. Genetics, 140, 725–732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang C.Z., Yen C.N., Cronin K., Mitchell D. and Britt A.B. (1997) UV- and γ-radiation sensitive mutants of Arabidopsis thaliana. Genetics, 147, 1401–1409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein H.L. (1998) Different types of recombination events are controlled by the RAD1 and RAD52 genes of Saccharomyces cerevisiae. Genetics, 120, 367–377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laat W.L.D., Jaspers N.G. and Hoeijmakers J.H. (1999) Molecular mechanism of nucleotide excision repair. Genes Dev., 13, 768–785. [DOI] [PubMed] [Google Scholar]
- Li A., Schuermann D., Gallego F., Kovalchuk I. and Tinland B. (2002) Repair of damaged DNA by Arabidopsis cell extract. Plant Cell, 14, 263–273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Z., Hossain G.S., Islas-Osuna M.A., Mitchell D.L. and Mount D.W. (2000) Repair of UV damage in plants by nucleotide excision repair: Arabidopsis UVH1 DNA repair gene is a homolog of Saccharomyces cerevisiae Rad1. Plant J., 21, 519–528. [DOI] [PubMed] [Google Scholar]
- Mengiste T. and Paszkowski J. (1999) Prospects for the precise engineering of plant genomes by homologous recombination. Biol. Chem., 380, 749–758. [DOI] [PubMed] [Google Scholar]
- Mitchell D.L. and Nairn R.S. (1989) The biology of the (6-4) photoproduct. Photochem. Photobiol., 49, 805–819. [DOI] [PubMed] [Google Scholar]
- Niedernhofer L.J., Essers J., Weeda G., Beverloo B., de Wit J., Muijtjens M., Odijk H., Hoeijmakers J.H. and Kanaar R. (2001) The structurespecific endonuclease Ercc1-Xpf is required for targeted gene replacement in embryonic stem cells. EMBO J., 20, 6540–6549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paques F. and Haber J.E. (1997) Two pathways for removal of nonhomologous DNA ends during doublestrand break repair in Saccharomyces cerevisiae. Mol. Cell. Biol., 17, 6765–6771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prado F. and Aguilera A. (1995) Role of reciprocal exchange, one-ended invasion crossover and singlestrand annealing on inverted and direct repeat recombination in yeast: different requirements for the RAD1, RAD10, and RAD52 genes. Genetics, 139, 109–123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Protic-Sabljic and Kraemer K.H. (1985) One pyrimidine dimer inactivates expression of a transfected gene in xeroderma pigmentosum cells. Proc. Natl Acad. Sci. USA, 82, 6622–6626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puchta H. and Hohn B. (1991) A transient assay in plant cells reveals a positive correlation between extrachromosomal recombination rates and length of homologous overlap. Nucleic Acids Res., 19, 2693–2700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saffran W.A., Greenberg R.B., Thalerscheer M.S. and Jones M.M. (1994) Single strand and double strand DNA damage-induced reciprocal recombination in yeast. Dependence on nucleotide excision repair and RAD1 recombination. Nucleic Acids Res., 22, 2823–2829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saparbaev M., Prakash L. and Prakash S. (1996) Requirement of mismatch repair genes MSH2 and MSH3 in the RAD1–RAD10 pathway of mitotic recombination in Saccharomyces cerevisiae. Genetics, 142, 727–736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sargent R.G., Meservy J.L., Perkins B.D., Kilburn A.E., Intody Z., Adair G.M., Nairn R.S. and Wilson J.H. (2000) Role of the nucleotide excision repair gene ERCC1 in formation of recombination-dependent rearrangements in mammalian cells. Nucleic Acids Res., 28, 3771–3778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schiestl R.H. and Prakash S. (1988) RAD1, an excision repair gene of Saccharomyces cerevisiae, is also involved in recombination. Mol. Cell. Biol., 8, 3619–3626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schiestl R.H. and Prakash S. (1990) RAD10, an excision repair gene of Saccharomyces cerevisiae, is involved in the RAD1 pathway of mitotic recombination. Mol. Cell. Biol., 10, 2485–2491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sekelsky J.J., McKim K.S., Chin G.M. and Hawley R.S. (1995) The Drosophila meiotic recombination gene mei-9 encodes a homologue of the yeast excision repair protein Rad1. Genetics, 141, 619–627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sijbers et al., 1996Sijbers A.M. (1996) Xeroderma pigmentosum group F caused by a defect in a structurespecific DNA repair endonuclease. Cell, 86, 811–822. [DOI] [PubMed] [Google Scholar]
- Sugawara N., Paques F., Colaiacovo M. and Haber J.E. (1997) Role of Saccharomyces cerevisiae Msh2 and Msh3 repair proteins in doublestrand break-induced recombination. Proc. Natl Acad. Sci. USA, 94, 9214–9219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomkinson A.E., Bardwell A.J., Bardwell L., Tappe N.J. and Friedberg E.C. (1993) Yeast DNA repair and recombination proteins Rad1 and Rad10 constitute a singlestranded-DNA endonuclease. Nature, 362, 860–862. [DOI] [PubMed] [Google Scholar]
- Tuteja N., Singh M.B., Misra M.K., Bhalla P.L. and Tuteja R. (2001) Molecular mechanisms of DNA damage and repair: progress in plants. Crit. Rev. Biochem. Mol. Biol., 36, 337–397. [DOI] [PubMed] [Google Scholar]
- Vergunst A.C. and Hooykaas P.J.J. (1999) Recombination in the plant genome and its application in biotechnology. Crit. Rev. Plant Sci., 18, 1–31. [Google Scholar]
- Weeda G. et al. (1997) Disruption of mouse ERCC1 results in a novel repair syndrome with growth failure, nuclear abnormalities and senescence. Curr. Biol., 7, 427–439. [DOI] [PubMed] [Google Scholar]
- Westerveld A., Hoeijmakers J.H., van Duin M., de Wit J., Odijk H., Pastink A., Wood R.D. and Bootsma D. (1984) Molecular cloning of a human DNA repair gene. Nature, 310, 425–429. [DOI] [PubMed] [Google Scholar]
- Xu H., Swoboda I., Bhalla P.L., Sijbers A.M., Zhao C., Ong E.K., Hoeijmakers J.H. and Singh M.B. (1998) Plant homologue of human excision repair gene ERCC1 points to conservation of DNA repair mechanisms. Plant J., 13, 823–829. [DOI] [PubMed] [Google Scholar]