

Abstract
A diverse range of conditions, from mitogenic stimuli to cytotoxic stress, can induce cell senescence. Here, I propose that simultaneous stimulation of mitogen-activated pathways and downstream inhibition of cyclin-dependent kinases leads, ultimately, to cell senescence. This model distinguishes between two types of growth arrest: first, exit to G0 phase, which is caused by the withdrawal of mitogens and can lead to apoptosis; and second, hypermitogenic arrest, which is stimulated by mitogens and can lead to senescence. The concept of hypermitogenic arrest defines cell senescence as a functionally active, stable and conditionally reversible state.
Introduction
Cell senescence has been defined as a form of replicative cell death or, in other words, a state in which proliferation is irreversibly arrested (Mathon & Lloyd, 2001). Nevertheless, senescent cells are metabolically active and can be maintained in culture for several years. Senescence may result from telomere shortening, which occurs during each cell cycle in some cell types, but senescence does not occur in others unless they are cultured under inadequate conditions (Mathon et al., 2001; Tang et al., 2001). These states are referred to as spontaneous and premature senescence, respectively, and some excellent recent reviews discuss their common and divergent features (for example, see Serrano & Blasco, 2001; Hanh, 2002). It has been suggested that premature senescence is triggered by the cumulative trauma of culturing cells in vitro (Sherr & DePinho, 2000), and also by various mitogenic, oncogenic, oxidative, cytostatic and toxic compounds, including DNA-damaging agents (McConnell et al., 1998; Lundberg et al., 2000; Roninson et al., 2001). What do the mitogenic and anti-mitogenic stimuli listed above have in common? It seems that two simultaneous events are required and sufficient to cause senescence. First, strong stimulation of mitogen-activated pathways is required. Second, cyclin-dependent kinases (CDKs) must be blocked, either directly or through the induction of CDK inhibitors (CDKIs).
Senescence-inducing 'mitogens' inhibit CDKs
In normal cells, mitogens (growth factors) initiate and maintain the transition from G1 phase to S phase in the cell cycle. Mitogen-activated protein kinase (MAPK) signalling pathways induce cyclin D1, which results in the activation of either CDK4 or CDK6 (henceforth referred to as CDK4/6), after which the cell no longer requires mitogens to complete the cell cycle. This point in late G1 phase is known as the 'restriction point' (Pardee, 1974). CDK4/6 phosphorylates the retinoblastoma protein (Rb), causing the release of E2F. This in turn transactivates cyclin E, which then activates CDK2. The upstream, mitogen-activated pathways thus stimulate the downstream cell-cycle machinery by inducing cyclins, which are the activators of CDKs. However, MAPK pathways can also induce CDKIs, offering the cell two possibilities: proliferation or growth arrest (Fig. 1). For example, the classical MAPK signalling cascade involves the sequential activation of Ras, Raf1, MAPK kinase (MEK) and extracellularsignal-regulated kinase (ERK), which stimulates activators (such as cyclin D) and inhibitors (such as p21, p16, p15 and p57) of CDKs (Marshall, 1995; Sewing et al., 1997; Woods et al., 1997; Chang et al., 2002). The same is true of MAPK pathways that act through the JNK (Jun kinase) and p38 kinases. Furthermore, both p21 and p27 can have opposing effects on CDK4/6 and CDK2 (Sherr & Roberts, 1999). What determines which of the two options the cell takes? It has been suggested that it is the strength or duration of the signal that is important; strong and/or sustained activation of the MAPK pathways arrests the cell cycle, whereas transient activation induces cell-cycle progression (Marshall, 1995). In support of this, low levels of Raf1 activity induce cyclin D1 and therefore proliferation, whereas high levels lead to p21 induction and growth arrest (Sewing et al., 1997; Woods et al., 1997). In addition, the simple explanation—that the proliferative status of the cell predetermines its response—should not be overlooked. Whereas a 'mitogen' may arrest a cycling cell or stimulate G0–G1 phase progression in a resting cell, it cannot possibly arrest a cell that is already resting. Thus, the overall cellular response may be predetermined by whether resting or cycling cells are targeted.
Figure 1.
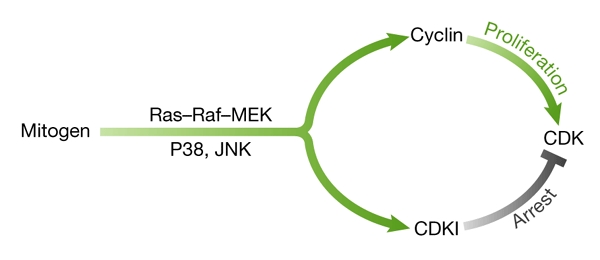
Dual mitogenic signalling. Mitogens simultaneously induce activators (such as cyclin D1) and cyclin-dependent-kinase inhibitors (CDKIs, such as p21, p16 and p15) through mitogen-activated pathways (Raf1/mitogen-activated protein kinase kinase (MEK)/extracellular-signal-related kinase (ERK), p38 and JNK (Jun kinase)), leading ultimately to cellular proliferation or arrest.
Regardless of what determines the choice between growth arrest and proliferation, cell senescence occurs only when mitogenic stimuli lead to CDK inhibition. In primary cells, Ras and the downstream MAPK pathways can induce senescence due to the induction of CDKIs (Missero et al., 1996; Serrano et al., 1997; Lin et al., 1998; Zhu et al., 1998; Malumbres et al., 2000; Wang et al., 2002; Brookes et al., 2002). All CDKIs induce senescence when ectopically expressed in fibroblasts (McConnell et al., 1998). Either the overexpression of positive regulators acting downstream of cyclin D1 (for example, CDK4/6, E2F1 and c-Myc) or the inactivation of tumour suppressors (such as Rb, p53 and p16) can block Ras-induced senescence. Even spontaneous senescence can be delayed by the overexpression of CDK4/6 (Morris et al., 2002; Holland et al., 1998). We can therefore conclude that the inhibition of pathways, either at the level of CDK4/6 or downstream of CDK4/6, is essential for all forms of senescence.
Senescence-inducing cytostatic stress
Ionizing radiation, DNA-damaging drugs, the p53 tumour suppressor, microtubule-active drugs (such as Taxol), oxidative stress and hypoxia-mimicking iron chelators, inhibitors of histone acetylase, transforming growth factor-β (TGF-β) and retinoids are all able to trigger premature cell senescence (McConnell et al., 1998; Chang et al., 1999; Roninson et al., 2001; Terao et al., 2001; Itahana et al., 2001). Most of these agents (especially in high doses) can also induce apoptosis, particularly in cells with 'ready-to-run' apoptotic machinery (that is, caspases). In such cells, the inhibition of apoptosis may allow cytostatic stress to induce senescence instead. For example, by inhibiting apoptosis, Bcl2 can promote p53-dependent senescence (Rincheval et al., 2002). All anti-mitogens directly or indirectly inhibit CDKs, causing G1-phase arrest and/or G2-phase arrest. For example, DNA-damaging agents cause the induction of CDKIs (such as p21) and changes in the phosphorylation states of CDKs and, thereby, their inhibition. Nevertheless, MAPKs are not inhibited, and these agents activate mitogenic signalling. Radiation stimulates Ras and numerous mitogenic kinases, including Raf1 and Akt (Kasid et al., 1996; Liu et al., 1996; Shaulian et al., 2000; Lee et al., 2000; Fang et al., 2001). Furthermore, p53 can induce growth factors that activate MAPK and Akt signalling, hypoxia causes autocrine secretion of mitogens and activation of mitogenic kinases, and DNA damage and hypoxia may, paradoxically, induce proliferation (Ishii et al., 1995; Adelman et al., 2000; Fang et al., 2001; Das et al., 2001; Salnikow et al., 2002). This suggests that, while inhibiting CDKs, senescence-inducing cytostatic stress may actually activate mitogenic pathways.
As is the case for premature senescence, spontaneous senescence occurs in the presence of mitogens. In the latter case, senescence is caused by the stress due to telomere shortening as the cells proliferate. In mitogen-induced senescence, hyperactive MAPK pathways inhibit CDKs. The elegance of cell senescence is that one stimulus can both activate upstream mitogenic signalling and arrest the cell cycle downstream.
Upstream and downstream signalling
It is assumed that cellular senescence evolved to suppress tumorigenesis (Lin et al., 1998; Campisi, 2001); that is, normal cells can respond to potentially oncogenic stimuli by entering the senescent state. This is reminiscent of the conflicting signal model, in which 'unscheduled' activation of potentially oncogenic stimuli leads to apoptosis, “ensuring that cell growth is restricted to the correct paracrine environment” (Raff, 1992; Evan & Littlewood, 1998). Does this mean that potentially oncogenic stimuli can cause both apoptosis and senescence? If so, how are the two outcomes determined? Here, we view them in the light of the upstream and downstream signalling pathways. During normal proliferation, the upstream mitogen-activated pathways stimulate downstream cell-cycle signalling (Fig. 2A). In the absence of upstream mitogenic signalling, the downstream cell-cycle pathways are inhibited (Fig. 2B), and the cells are in G0 arrest. Conversely, forced, isolated overexpression of downstream oncogenes (for example, viral E1A, c-Myc and E2F) can induce apoptosis (Fig. 2C). However, cell-cycle progression and apoptosis are not permanently connected, and can be separated; for example, apoptosis can be suppressed by mitogens (Evan & Littlewood, 1998). Finally, in the presence of mitogenic signalling, 'unscheduled' inhibition of the cell cycle, such as by CDKIs, results in senescence (Fig. 2D). Therefore, whereas oncogene-induced apoptosis is associated with 'forced' proliferation, senescence is associated with 'forced' arrest (Fig. 2C versus 2D). In this view, oncogene-induced apoptosis and senescence are mirror images, each involving the isolated activation of either upstream or downstream mitogenic pathways.
Figure 2.
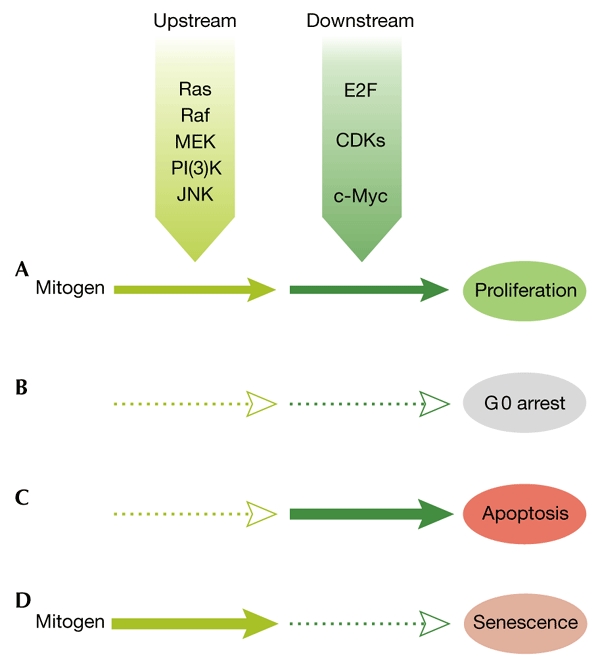
Sequential and reciprocal activation of upstream and downstream signalling pathways. (A) Normal mitogenic signalling involves the sequential activation of both upstream and downstream pathways. Note that, due to feedback loops, the distinctions between 'upstream' and 'downstream' are not always maintained, and the molecules indicated in the boxes are merely examples. (B) Classical growth arrest in G0 occurs when there is no activation of either upstream or downstream pathways. (C) The conflicting signal model of apoptosis involves the isolated overactivation of downstream pathways. (D) The conflicting signal model of senescence involves the isolated overactivation of upstream pathways.
Cell-cycle arrest: hypomitogenic and hypermitogenic
The model of mitogen-induced senescence presented here leads to an unexpected prediction: there are two opposing types of growth arrest. Classical growth arrest, or G0 arrest, which is caused by growth-factor withdrawal, is essentially an exit from the cell cycle (Fig. 3), and is characterized by low levels of cyclin D1. After mitogenic stimulation, cells in the G0 state enter G1, during which they induce cyclin D1 and progress through the restriction point (Blagosklonny & Pardee, 2002). It is generally assumed that all resting cells ultimately exit the cell cycle. However, when cells arrest in response to mitogens, the consequences are different. In this state of hypermitogenic arrest, cells arrest at or beyond the restriction point, in either G1 or G2 phase (Fig. 3), and numerous anticancer drugs arrest cells when the levels of cyclins D, E, A or B are elevated (Gong et al., 1995; Darzynkiewicz, 2002). The hypermitogenic arrest induced by phorbol esters is characterized by CDK inhibition in the presence of cyclin D1 (Blagosklonny, 1998), and iron chelators, contact inhibition and transforming growth factor-β induce a senescence-like late G1 arrest that is often associated with the induction of p27 (Kletsas et al., 1995; Yoon et al., 2002). DNA damage can induce permanent growth arrest in G2, which is dependent on Rb (Naderi et al., 2002). Also, hyperactivated Raf1 can arrest cells in G2 (Kerkhoff & Rapp, 1998). In cycling cells, Ras induces cyclin D1 in G2 phase (Sherr, 2002). Therefore, forced hypermitogenic arrest is associated with elevated cyclin D1.
Figure 3.
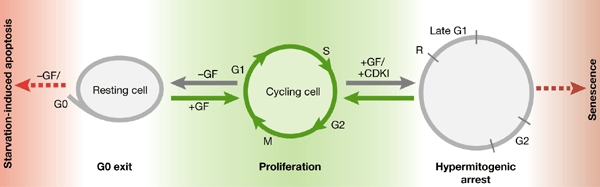
From proliferation to either G0 arrest or hypermitogenic arrest. 'Cycling cells' proliferate in the presence of growth factors (GFs). The withdrawal of GFs causes a reversible exit from the cell cycle, known as G0, producing a 'resting cell'. In normal cells, a complete lack of GFs may cause apoptosis. GF stimulation in the presence of cyclin-dependent-kinase inhibitors (+CDKI) can cause hypermitogenic arrest, and continuous stimulation can lead to senescence.
The existence of G0 has been questioned time after time, precisely because the idea of G0 arrest has been overgeneralized. The distinction between G0 (hypomitogenic arrest) and hypermitogenic arrest may explain much of this controversy.
Hypermitogenic arrest, cell morphology and function
Intriguingly, the model of hypermitogenic arrest predicts a 'large-cell' morphology, and high levels of autocrine and/or paracrine secretion by senescent cells. Hypermitogenic arrest must be accompanied by protein synthesis—as mitogens stimulate not only DNA synthesis, but also RNA and protein synthesis—and by metabolism. It is an arrest of the cell cycle, not of growth. In fact, high-intensity Raf signals are known to convert mitotic cell cycling into cellular growth (Kerkhoff & Rapp, 1998). Thus, hypermitogenic arrest can explain the large-cell morphology that has been associated with senescence.
It is also possible that a cell may attempt to compensate for the cell-cycle block by the overinduction of mitogenic signalling. Senescent cells are highly active in the autocrine and paracrine secretion of mitogens (Roninson et al., 2001). Unlike the hypomitogenic arrest at G0, hypermitogenic arrest is enhanced by stimulation by mitogens. Therefore, hypermitogenic arrest and cell senescence are stable conditions, which are often seen as irreversible. For example, consistent with the senescent phenotype, p38-induced G1 cell-cycle arrest is irreversible after four days (Haq et al., 2002). As another example, when p16 expression was induced for six days, DNA synthesis remained inhibited and the cells acquired the morphological features of senescence. If p16 induction was interrupted after six days, most cells retained these morphological features (Dai & Enders, 2000).
Overturning hypermitogenic arrest and senescence
The concept presented here predicts that, unlike G0 arrest, hypermitogenic arrest cannot be blocked by mitogenic stimuli and, indeed, senescent cells have been shown to be resistant to mitogens (Mathon & Lloyd, 2001). However, as illustrated by the following examples, it can be overcome by induction of downstream 'oncogenes'. Both c-Myc and E1A override the hypermitogenic arrest caused by PMA (phorbol-12-myristate-13-acetate) (Blagosklonny, 1998). Reactivation of cyclin E1, which acts downstream of p16, is sufficient to trigger escape from Ras-induced senescence (Peeper et al., 2002). In contrast to stimulation by mitogens, stimulation by viral proteins can reactivate terminally differentiated cells of the muscle system: for example, the large-T viral oncoprotein induces the cycling of terminally differentiated skeletal muscle myotubes (Ohkubo et al., 1994), E2F1 can override 'irreversible' arrest in post-mitotic ventricular myocytes (Agah et al., 1997), and the E1A oncogene can reactivate the cell cycle in terminally differentiated skeletal muscle cells (Tiainen et al., 1996). Also, the inactivation of either p53 or simian virus 40 (SV40) induces senescent cells to re-enter S phase and to revert to their 'young' morphology (Gire & Wynford-Thomas, 1998). Finally, oncogenes that act downstream of the restriction point (such as c-Myc and CDK4), and thus may overcome senescence, cooperate with hypermitogenic oncogenes (such as Ras), leading to malignant transformation (Land et al., 1983).
The physiological role of cell senescence
The activation of apoptosis, by 'unscheduled' oncogenic signalling, is thought to contribute to the prevention of cancer. Similarly, cell senescence has been proposed as a mechanism to block immortalization and tumorigenesis. For example, it may limit the transforming potential of excessive Ras mitogenic signalling. However, this view may be too focused on cancer. Similar to apoptosis, which has many physiological roles in development and tissue turnover, senescence may have other physiological functions. By analogy to apoptosis, it is generally perceived as a pre-death stage that results in the elimination of cells. The fact that senescence is a stable and metabolically active state, however, argues that the functions of senescence and apoptosis oppose one another: senescence does not eliminate a cell, but instead preserves it in a maximally stable and metabolically active state. For example, senescent fibroblasts have a useful purpose: they express proteins that are normally induced on wounding, including those that remodel the extracellular matrix (Benanti et al., 2002). Furthermore, senescence in fibroblasts is associated with resistance to apoptosis caused by radiation (Yeo et al., 2000). By contrast, mitogen deprivation results in an unstable condition that can lead to apoptosis because mitogens are also survival factors.
Many terminally differentiated cells may be arrested in a hypermitogenic (senescence-like) state, rather than in the unstable and apoptosis-prone G0 arrest, owing to phenomena such as contact inhibition. One can speculate that muscle cells, dermal fibroblasts and certain neuronal cells may be preserved in this way. For example, contact inhibition of growth may be accompanied by p27 accumulation and cellular senescence (Munro et al., 2001). Unlike arrest due to growth-factor withdrawal, arrest due to contact inhibition occurs in the presence of mitogens. Interestingly, cell–cell interactions between biliary cells and hepatocytes result in the mid-G1-phase arrest of hepatocytes, which are insensitive to mitogens (Loyer et al., 1996). If quiescence in 'contact-inhibited' organs, such as the liver, is controlled by CDKIs (such as p27), then the downregulation of these inhibitors might be necessary for liver regeneration, for example after partial hepatectomy.
It is possible that certain mitogens secreted by contact-inhibited tissue might maintain hypermitogenic arrest, essentially acting like growth inhibitors. More than 30 years ago, it was postulated that tissues produce inhibitors, known as chalones, to control cell homeostasis (Bullough & Laurence, 1968). Despite thousands of publications describing putative chalones, these mysterious inhibitors have never been purified. Perhaps these elusive inhibitors are simply ordinary mitogens, acting as mitogens in G0 cells and as chalones in cycling cells. Finally, if cells in our body are arrested in a hypermitogenic state, this explains why mammalian viral oncoproteins (for example, E1A, E7, E6 and T antigen) act downstream of the restriction point (Fig. 2, downstream pathways). In fact, oncoproteins that act upstream would simply strengthen the senescent state. The sequence of oncogenic events that lead to cancer may therefore vary depending on the prevalence of hypermitogenic versus G0 arrest in different tissues.
Acknowledgments
I apologize to those authors whose work was not cited due to page limitations.
References
- Adelman D.M., Gertsenstein M., Nagy A., Simon M.C. & Maltepe E. (2000) Placental cell fates are regulated in vivo by HIF-mediated hypoxia responses. Genes Dev., 14, 3191–3203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agah R., Kirshenbaum L.A., Abdellatif M., Truong L.D., Chakraborty S., Michael L.H. & Schneider M.D. (1997) Adenoviral delivery of E2F-1 directs cell cycle reentry and p53-independent apoptosis in postmitotic adult myocardium in vivo. J. Clin. Invest., 100, 2722–2728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benanti J.A., Williams D.K., Robinson K.L., Ozer H.L. & Galloway D.A. (2002) Induction of extracellular matrix-remodeling genes by the senescence-associated protein APA-1. Mol. Cell. Biol., 22, 7385–7397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blagosklonny M.V. (1998) The mitogen-activated protein kinase pathway mediates growth arrest or E1A-dependent apoptosis in SKBr3 human breast cancer cells. Int. J. Cancer, 78, 511–517. [DOI] [PubMed] [Google Scholar]
- Blagosklonny M.V. & Pardee A.B. (2002) The restriction point of the cell cycle. Cell Cycle, 1, 103–110. [PubMed] [Google Scholar]
- Brookes S. et al. (2002) INK4a-deficient human diploid fibroblasts are resistant to RAS-induced senescence. EMBO J., 21, 2936–2945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bullough W.S. & Laurence E.B. (1968) Epidermal chalone and mitotic control in the Vx2 epidermal tumour. Nature, 220, 134–135. [DOI] [PubMed] [Google Scholar]
- Campisi J. (2001) Cellular senescence as a tumorsuppressor mechanism. Trends Cell Biol., 11, S27–S31. [DOI] [PubMed] [Google Scholar]
- Chang B.D., Broude E.V., Dokmanovic M., Zhu H., Ruth A., Xuan Y., Kandel E.S., Lausch E., Christov K. & Roninson I.B. (1999) A senescence-like phenotype distinguishes tumor cells that undergo terminal proliferation arrest after exposure to anticancer agents. Cancer Res., 59, 3761–3767. [PubMed] [Google Scholar]
- Chang F., Steelman L.S. & McCubrey J.A. (2002) Raf-induced cell cycle progression in human TF-1 hematopoietic cells. Cell Cycle, 1, 220–226. [PubMed] [Google Scholar]
- Dai C.Y. & Enders G.H. (2000) p16 INK4a can initiate an autonomous senescence program. Oncogene, 19, 1613–1622. [DOI] [PubMed] [Google Scholar]
- Darzynkiewicz Z. (2002) One more reason to get arrested (in G1). Cell Cycle, 1, 318–319. [DOI] [PubMed] [Google Scholar]
- Das M., Bouchey D.M., Moore M.J., Hopkins D.C., Nemenoff R.A. & Stenmark K.R. (2001) Hypoxia-induced proliferative response of vascular adventitial fibroblasts is dependent on G protein-mediated activation of mitogen-activated protein kinases. J. Biol. Chem., 276, 15631–15640. [DOI] [PubMed] [Google Scholar]
- Evan G. & Littlewood T. (1998) A matter of life and cell death. Science, 281, 1317–1322. [DOI] [PubMed] [Google Scholar]
- Fang L., Li G.N., Liu G.Z., Lee S.W. & Aaronson S.A. (2001) p53 induction of heparin-binding EGF-like growth factor counteracts p53 growth suppression through activation of MAPK and P13K/Akt signaling cascades. EMBO J., 20, 1931–1939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gire V. & Wynford-Thomas D. (1998) Reinitiation of DNA synthesis and cell division in senescent human fibroblasts by microinjection of anti-p53 antibodies. Mol. Cell. Biol., 18, 1611–1621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gong J., Traganos F. & Darzynkiewicz Z. (1995) Growth imbalance and altered expression of cyclins B1, A, E and D3 in MOLT-4 cells synchronized in the cell cycle by inhibitors of DNA replication. Cell Growth Differ., 6, 1485–1493. [PubMed] [Google Scholar]
- Hanh W.C. (2002) Senescence, telomere shortening and telomere maintenance. Cancer Biol. Ther., 1, 398–400. [DOI] [PubMed] [Google Scholar]
- Haq R., Brenton J.D., Takahashi M., Finan D., Finkielsztein A., Damaraju S., Rottapel R. & Zanke B. (2002) Constitutive p38HOG mitogen-activated protein kinase activation induces permanent cell cycle arrest and senescence. Cancer Res., 62, 5076–5082. [PubMed] [Google Scholar]
- Holland E.C., Hively W.P., Gallo V. & Varmus H.E. (1998) Modeling mutations in the G1 arrest pathway in human gliomas: overexpression of CDK4 but not loss of INK4a–ARF induces hyperploidy in cultured mouse astrocytes. Genes Dev., 12, 3644–3649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishii K., Yamaoka K., Hosoi Y., Ono T. & Sakamoto K. (1995) Enhanced mitogen-induced proliferation of rat splenocytes by low-dose whole-body X-irradiation. Physiol. Chem. Phys. Med. NMR, 27, 17–23. [PubMed] [Google Scholar]
- Itahana K., Dimri G. & Campisi J. (2001) Regulation of cellular senescence by p53. Eur. J. Biochem., 268, 2784–2791. [DOI] [PubMed] [Google Scholar]
- Kasid U., Suy S., Dent P., Ray S., Whiteside T.L. & Sturgill T.W. (1996) Activation of Raf by ionizing radiation. Nature, 382, 813–816. [DOI] [PubMed] [Google Scholar]
- Kerkhoff E. & Rapp U.R. (1998) High-intensity Raf signals convert mitotic cell cycling into cellular growth. Cancer Res., 58, 1636–1640. [PubMed] [Google Scholar]
- Kletsas D., Stathakos D., Sorrentino V. & Philipson L. (1995) The growth-inhibitory block of TGF-β is located close to the G1/S border in the cell cycle. Exp. Cell Res., 217, 477–483. [DOI] [PubMed] [Google Scholar]
- Land H., Parada L.F. & Weinberg R.A. (1983) Tumorigenic conversion of primary embryo fibroblasts requires at least two cooperating oncogenes. Nature, 304, 596–602. [DOI] [PubMed] [Google Scholar]
- Lee S.W., Fang L., Igarashi M., Ouchi T., Lu K.P. & Aaronson S.A. (2000) Sustained activation of Ras/Raf/mitogen-activated protein kinase cascade by the tumor suppressor p53. Proc. Natl Acad. Sci. USA, 97, 8302–8305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin A.W., Barradas M., Stone J.C., van Aelst L., Serrano M. & Lowe S.W. (1998) Premature senescence involving p53 and p16 is activated in response to constitutive MEK/MAPK mitogenic signaling. Genes Dev., 12, 3008–3019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Z.G., Baskaran R., Lea-Chou E.T., Wood L.D., Chen Y., Karin M. & Wang J.Y. (1996) Three distinct signalling responses by murine fibroblasts to genotoxic stress. Nature, 384, 273–276. [DOI] [PubMed] [Google Scholar]
- Loyer P., Ilyin G., Cariou S., Glaise D., Corlu A. & Guguen-Guillouzo C. (1996) Progression through G1 and S phases of adult rat hepatocytes. Prog. Cell Cycle Res., 2, 37–47. [DOI] [PubMed] [Google Scholar]
- Lundberg A.S., Hahn W.C., Gupta P. & Weinberg R.A. (2000) Genes involved in senescence and immortalization. Curr. Opin. Cell Biol., 12, 705–709. [DOI] [PubMed] [Google Scholar]
- Malumbres M., De Castro I.P., Hernandez M.I., Jimenez M., Corral T. & Pellicer A. (2000) Cellular response to oncogenic Ras involves induction of the Cdk4 and Cdk6 inhibitor p15(INK4b). Mol. Cell. Biol., 20, 2915–2925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marshall C.J. (1995) Specificity of receptor tyrosine kinase signaling: transient versus sustained extracellular signal-regulated kinase activation. Cell, 80, 179–185. [DOI] [PubMed] [Google Scholar]
- Mathon N.F. & Lloyd A.C. (2001) Cell senescence and cancer. Nature Rev. Cancer, 1, 203–213. [DOI] [PubMed] [Google Scholar]
- Mathon N.F., Malcolm D.S., Harrisingh M.C., Cheng L. & Lloyd A.C. (2001) Lack of replicative senescence in normal rodent glia. Science, 291, 872–875. [DOI] [PubMed] [Google Scholar]
- McConnell B.B., Starborg M., Brookes S. & Peters G. (1998) Inhibitors of cyclin-dependent kinases induce features of replicative senescence in early passage human diploid fibroblasts. Curr. Biol., 8, 351–354. [DOI] [PubMed] [Google Scholar]
- Missero C., Di Cunto F., Kiyokawa H., Koff A. & Dotto G.P. (1996) The absence of p21Cip1/WAF1 alters keratinocyte growth and differentiation and promotes Ras-tumor progression. Genes Dev., 10, 3065–3075. [DOI] [PubMed] [Google Scholar]
- Morris M., Hepburn P. & Wynford-Thomas D. (2002) Sequential extension of proliferative lifespan in human fibroblasts induced by over-expression of CDK4 or 6 and loss of p53 function. Oncogene, 21, 4277–4288. [DOI] [PubMed] [Google Scholar]
- Munro J., Steeghs K., Morrison V., Ireland H. & Parkinson E.K. (2001) Human fibroblast replicative senescence can occur in the absence of extensive cell division and short telomeres. Oncogene, 20, 3541–3552. [DOI] [PubMed] [Google Scholar]
- Naderi S., Hunton I.C. & Wang J.Y. (2002) Radiation dose-dependent maintenance of G(2) arrest requires retinoblastoma protein. Cell Cycle, 1, 193–200. [PubMed] [Google Scholar]
- Ohkubo Y., Kishimoto T., Nakata T., Yasuda H. & Endo T. (1994) SV40 large T antigen reinduces the cell cycle in terminally differentiated myotubes through inducing Cdk2, Cdc2, and their partner cyclins. Exp. Cell Res., 214, 270–278. [DOI] [PubMed] [Google Scholar]
- Pardee A.B. (1974) A restriction point for control of normal animal cell proliferation. Proc. Natl Acad. Sci. USA, 71, 1286–1290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peeper D.S., Shvarts A., Brummelkamp T., Douma S., Koh E.Y., Daley G.Q. & Bernards R. (2002) A functional screen identifies hDRIL1 as an oncogene that rescues RAS-induced senescence. Nature Cell Biol., 4, 148–153. [DOI] [PubMed] [Google Scholar]
- Raff M.C. (1992) Social controls on cell survival and cell death. Nature, 356, 397–400. [DOI] [PubMed] [Google Scholar]
- Rincheval V., Renaud F., Lemaire C., Godefroy N., Trotot P., Boulo V., Mignotte B. & Vayssiere J.L. (2002) Bcl-2 can promote p53-dependent senescence versus apoptosis without affecting the G1/S transition. Biochem. Biophys. Res. Commun., 298, 282–288. [DOI] [PubMed] [Google Scholar]
- Roninson I.B., Broude E.V. & Chang B.D. (2001) If not apoptosis, then what? Treatment-induced senescence and mitotic catastrophe in tumor cells. Drug Resist. Update, 4, 303–313. [DOI] [PubMed] [Google Scholar]
- Salnikow K., Kluz T., Costa M., Piquemal D., Demidenko Z.N., Xie K. & Blagosklonny M.V. (2002) The regulation of hypoxic genes by calcium involves c-Jun/AP-1, which cooperates with hypoxia-inducible factor 1 in response to hypoxia. Mol. Cell. Biol., 22, 1734–1741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serrano M. & Blasco M.A. (2001) Putting the stress on senescence. Curr. Opin. Cell Biol., 13, 748–753. [DOI] [PubMed] [Google Scholar]
- Serrano M., Lim A.W., McCurrach M.E., Beach D. & Lowe S.W. (1997) Oncogenic Ras provokes premature cell senescence associated with accumulation of p53 and p16INK1A. Cell, 88, 593–602. [DOI] [PubMed] [Google Scholar]
- Sewing A., Wiseman B., Lloyd A.C. & Land H. (1997) High-intensity Raf signal causes cell cycle arrest mediated by p21Cip1. Mol. Cell. Biol., 17, 5588–5597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaulian E., Schreiber M., Piu F., Beeche M., Wagner E.F. & Karin M. (2000) The mammalian UV response: c-Jun induction is required for exit from p53-imposed growth arrest. Cell, 103, 897–907. [DOI] [PubMed] [Google Scholar]
- Sherr C.J. (2002) D1 in G2. Cell Cycle, 1, 36–38. [PubMed] [Google Scholar]
- Sherr C.J. & DePinho R.A. (2000) Cellular senescence: mitotic clock or culture shock? Cell, 102, 407–410. [DOI] [PubMed] [Google Scholar]
- Sherr C.J. & Roberts J.M. (1999) CDK inhibitors: positive and negative regulators of G1-phase progression. Genes Dev., 13, 1501–1512. [DOI] [PubMed] [Google Scholar]
- Tang D.G., Tokumoto Y.M., Apperly J.A., Lloyd A.C. & Raff M.C. (2001) Lack of replicative senescence in cultured rat oligodendrocyte precursor cells. Science, 291, 868–871. [DOI] [PubMed] [Google Scholar]
- Terao Y. et al. (2001) Sodium butyrate induces growth arrest and senescence-like phenotypes in gynecologic cancer cells. Int. J. Cancer, 94, 257–267. [DOI] [PubMed] [Google Scholar]
- Tiainen M., Spitkovsky D., Jansen-Durr P., Sacchi A. & Crescenzi M. (1996) Expression of E1A in terminally differentiated muscle cells reactivates the cell cycle and suppresses tissuespecific genes by separable mechanisms. Mol. Cell. Biol., 16, 5302–5312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang W., Chen J.X., Liao R., Deng Q., Zhou J.J., Huang S. & Sun P. (2002) Sequential activation of the MEK-extracellular signal-regulated kinase and MKK3/6-p38 mitogen-activated protein kinase pathways mediates oncogenic ras-induced premature senescence. Mol. Cell. Biol., 22 3389–3403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woods D., Parry D., Cherwinski H., Bosch E., Lees E. & McMahon M. (1997) Raf-induced proliferation or cell cycle arrest is determined by the level of Raf activity with arrest mediated by p21Cip1. Mol. Cell. Biol., 17, 5598–5611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeo E.J., Hwang Y.C., Kang C.M., Choy H.E. & Park S.C. (2000) Reduction of UV-induced cell death in the human senescent fibroblasts. Mol. Cell, 10, 415–422. [PubMed] [Google Scholar]
- Yoon G., Kim H.J., Yoon Y.S., Cho H., Lim I.K. & Lee J.H. (2002) Iron chelation-induced senescence-like growth arrest in hepatocyte cell lines: association of transforming growth factor β1 (TGF-β1)-mediated p27Kip1 expression. Biochem. J., 366, 613–621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu J.Y., Woods D., McMahon M. & Bishop J.M. (1998) Senescence of human fibroblasts induced by oncogenic Raf. Genes Dev., 12, 2997–3007. [DOI] [PMC free article] [PubMed] [Google Scholar]