

Abstract
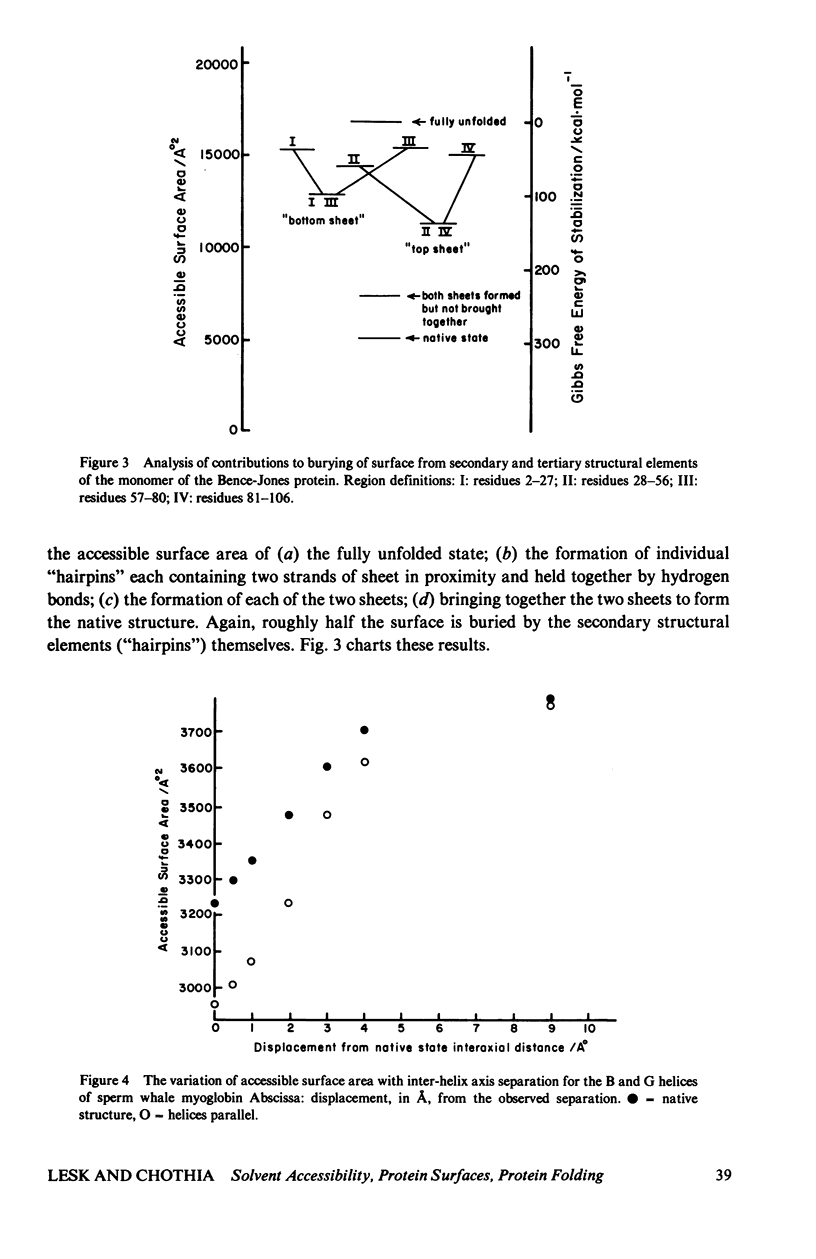
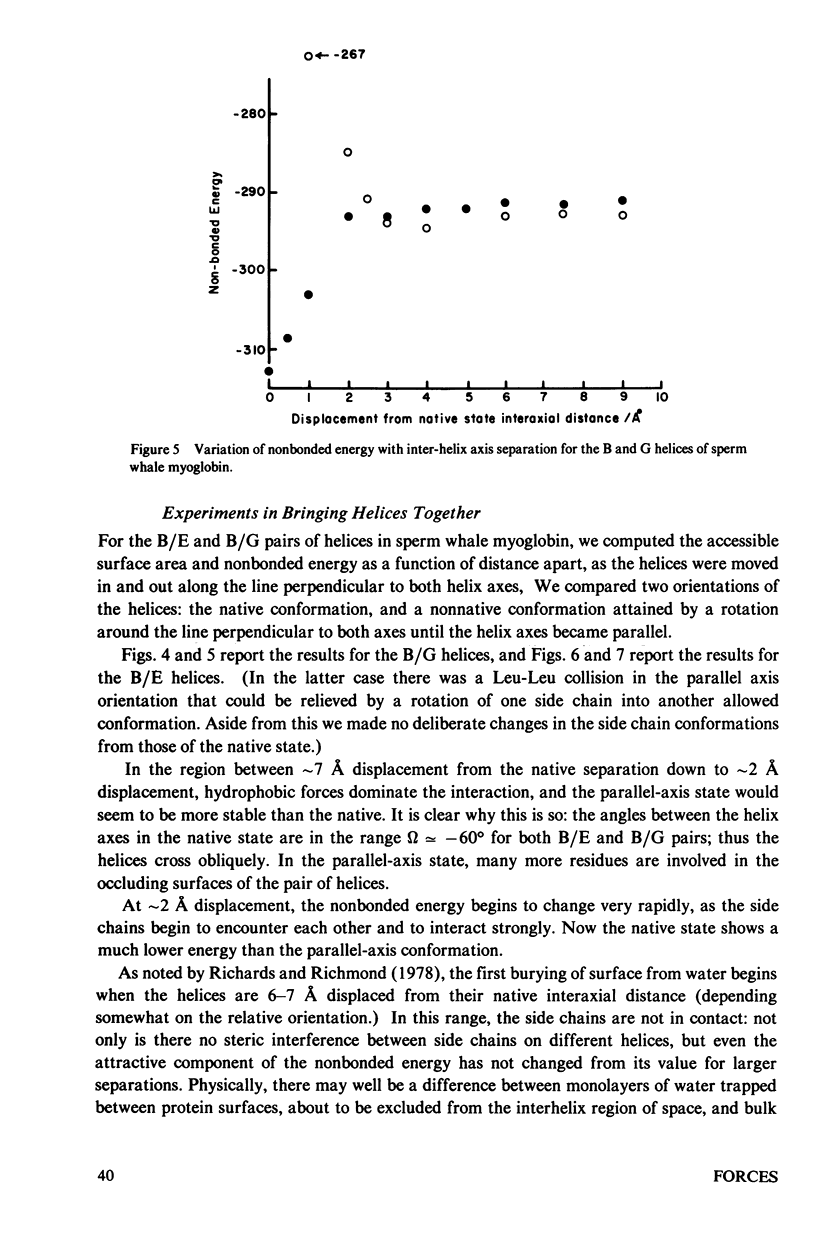
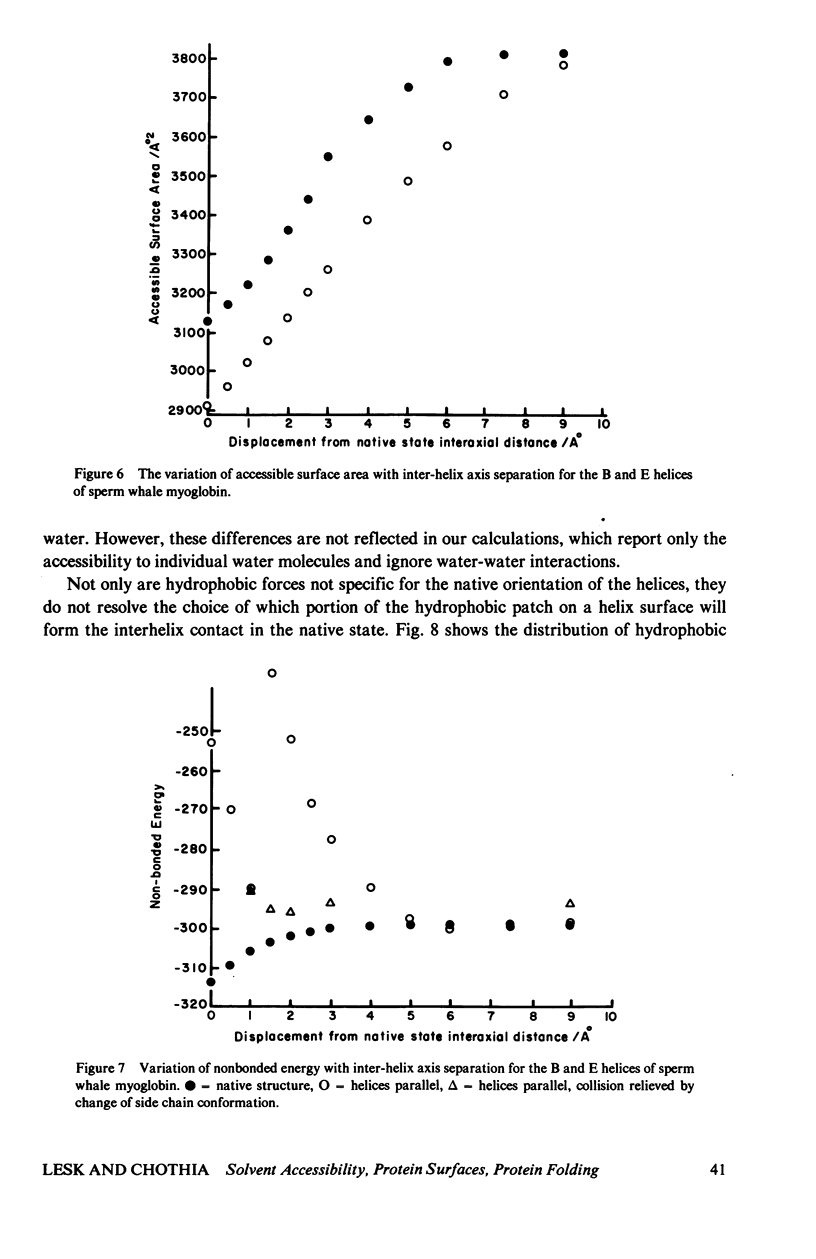
Studies of the native structures of proteins, together with measurements of the thermodynamic properties of the transition between unfolded and native states, have defined the major components of the forces that stabilize native protein structures. However, the nature of the intermediates in the folding process remains largely hypothetical. It is a fairly widespread and not implausible assumption that the intermediates in the folding of a monomeric protein contain the same kinds of secondary and tertiary structures that appear in the native conformation, and that, although unstable, their lifetimes are prolonged by forces similar to those that stabilize the native structure. We wished to examine what happens if, during the folding of a monomeric protein, regions of secondary structure come together to form an intermediate of reduced instability. We applied calculations of accessible surface area (a measure of hydrophobic stabilization) and parameterized nonbonded energy calculations (measuring the strengths of van der Waals forces) to identify the kinds of stabilizing interactions that might be available to such an intermediate. First, we analyzed the total buried surface area of two types of proteins into contributions from formation of secondary structure alone, interaction of pairs of secondary-structural elements, the formation of the structure alone, interaction of pairs of secondary-structural elements, the formation of the complete secondary structure without the turns, and the complete native structure. The formation of secondary structure alone, without tertiary-structural interactions, buries roughly half the surface that the complete structure does. We then analyzed in more detail the approach of two alpha-helices to form a complex, as an illustrative example of the nature of the interaction between compact structural units which remain fairly rigid during their interaction. Many features of the results are not limited to the interaction of alpha-helices. (The results therefore neither confirm nor refute the hypothesis that alpha-helices are intermediates in the folding proteins). We find that the first forces to be felt upon approach arise from solvent conditions on the relative position and orientation of the two helices as does the close packing which optimizes the van der Waals interactions at shorter distances apart. Therefore there appears to be a range of distances in which hydrophobic interactions could create a nonspecific complex between two helices in which the side chains might have sufficient time to seek the proper interdigitation observed in the native structure, where the two helices are in intimate contact. Indeed, we find that only in the final stages of approach is the native geometry the most stable; in the region in which solvent-exclusion forces predominate, the conformation with helix axes parallel is more stable than the native conformation, in the cases we examined...
Full text
PDF






Selected References
These references are in PubMed. This may not be the complete list of references from this article.
- Chothia C., Levitt M., Richardson D. Structure of proteins: packing of alpha-helices and pleated sheets. Proc Natl Acad Sci U S A. 1977 Oct;74(10):4130–4134. doi: 10.1073/pnas.74.10.4130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chothia C. The nature of the accessible and buried surfaces in proteins. J Mol Biol. 1976 Jul 25;105(1):1–12. doi: 10.1016/0022-2836(76)90191-1. [DOI] [PubMed] [Google Scholar]
- Creighton T. E. Experimental studies of protein folding and unfolding. Prog Biophys Mol Biol. 1978;33(3):231–297. doi: 10.1016/0079-6107(79)90030-0. [DOI] [PubMed] [Google Scholar]
- Karplus M., Weaver D. L. Protein-folding dynamics. Nature. 1976 Apr 1;260(5550):404–406. doi: 10.1038/260404a0. [DOI] [PubMed] [Google Scholar]
- Lee B., Richards F. M. The interpretation of protein structures: estimation of static accessibility. J Mol Biol. 1971 Feb 14;55(3):379–400. doi: 10.1016/0022-2836(71)90324-x. [DOI] [PubMed] [Google Scholar]
- Lesk A. M., Chothia C. How different amino acid sequences determine similar protein structures: the structure and evolutionary dynamics of the globins. J Mol Biol. 1980 Jan 25;136(3):225–270. doi: 10.1016/0022-2836(80)90373-3. [DOI] [PubMed] [Google Scholar]
- Levitt M. Energy refinement of hen egg-white lysozyme. J Mol Biol. 1974 Jan 25;82(3):393–420. doi: 10.1016/0022-2836(74)90599-3. [DOI] [PubMed] [Google Scholar]
- Privalov P. L., Khechinashvili N. N. A thermodynamic approach to the problem of stabilization of globular protein structure: a calorimetric study. J Mol Biol. 1974 Jul 5;86(3):665–684. doi: 10.1016/0022-2836(74)90188-0. [DOI] [PubMed] [Google Scholar]
- Richards F. M., Richmond T. Solvents, interfaces and protein structure. Ciba Found Symp. 1977;(60):23–45. doi: 10.1002/9780470720424.ch2. [DOI] [PubMed] [Google Scholar]