

Abstract
1. The sensitivity of detecting modulation in a test tone sinusoidally frequency-modulated at a rate ϕtest is diminished after exposure to a conditioning tone more deeply frequency-modulated at a rate ϕcond provided that ϕcond is not very different from ϕtest, the sound amplitude being kept constant for each tone at a comfortable hearing level 40-45 dB above threshold.
2. When ϕcond = ϕtest the frequency deviation in the modulated test tone must be increased to about three times the unconditioned threshold magnitude to be detectable immediately after exposure to the conditioning tone. Detection sensitivity returns to normal in about one minute.
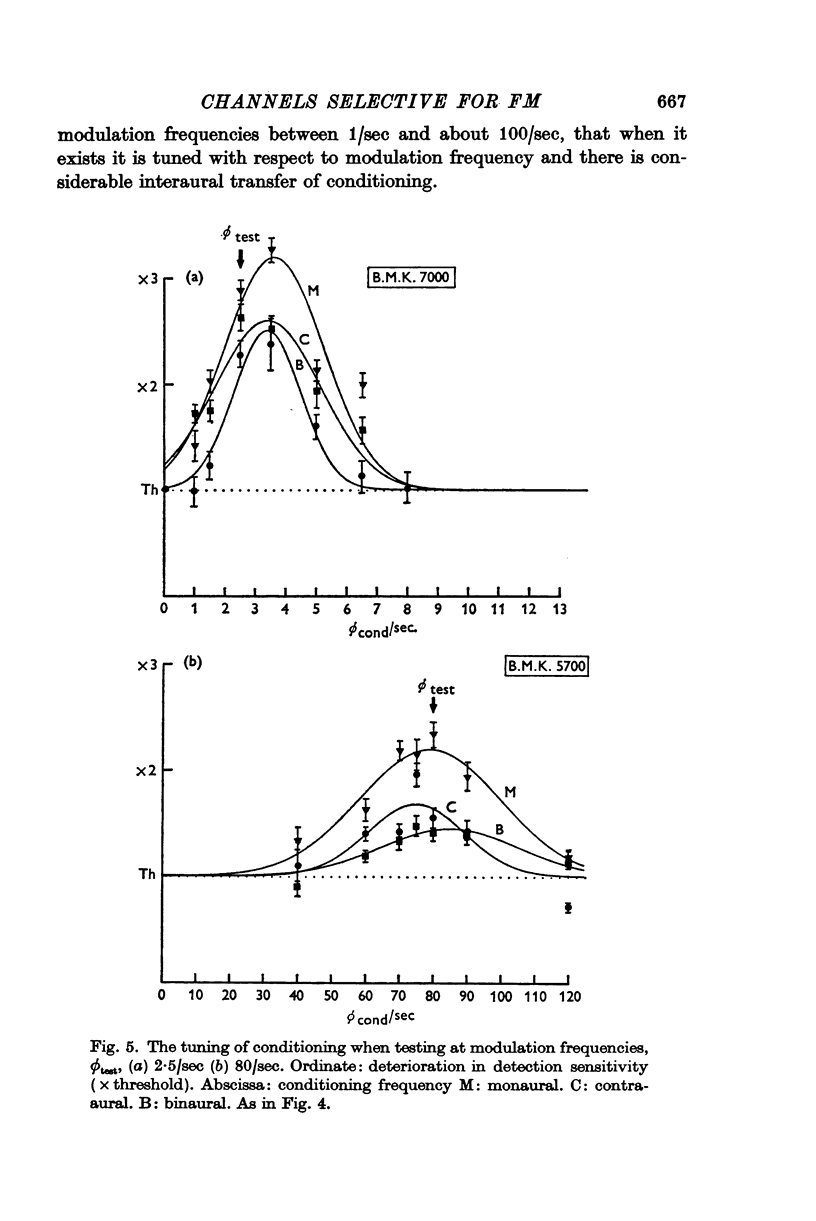
3. At low modulation frequencies the conditioning effects are tuned, being much diminished when ϕcond differs from ϕtest by a few cycles per second.
4. Comparing monaural with contra-aural conditioning demonstrates a considerable interaural transfer of about 60-80% of the effect, indicating that the conditioning and its selectivity are predominantly central phenomena.
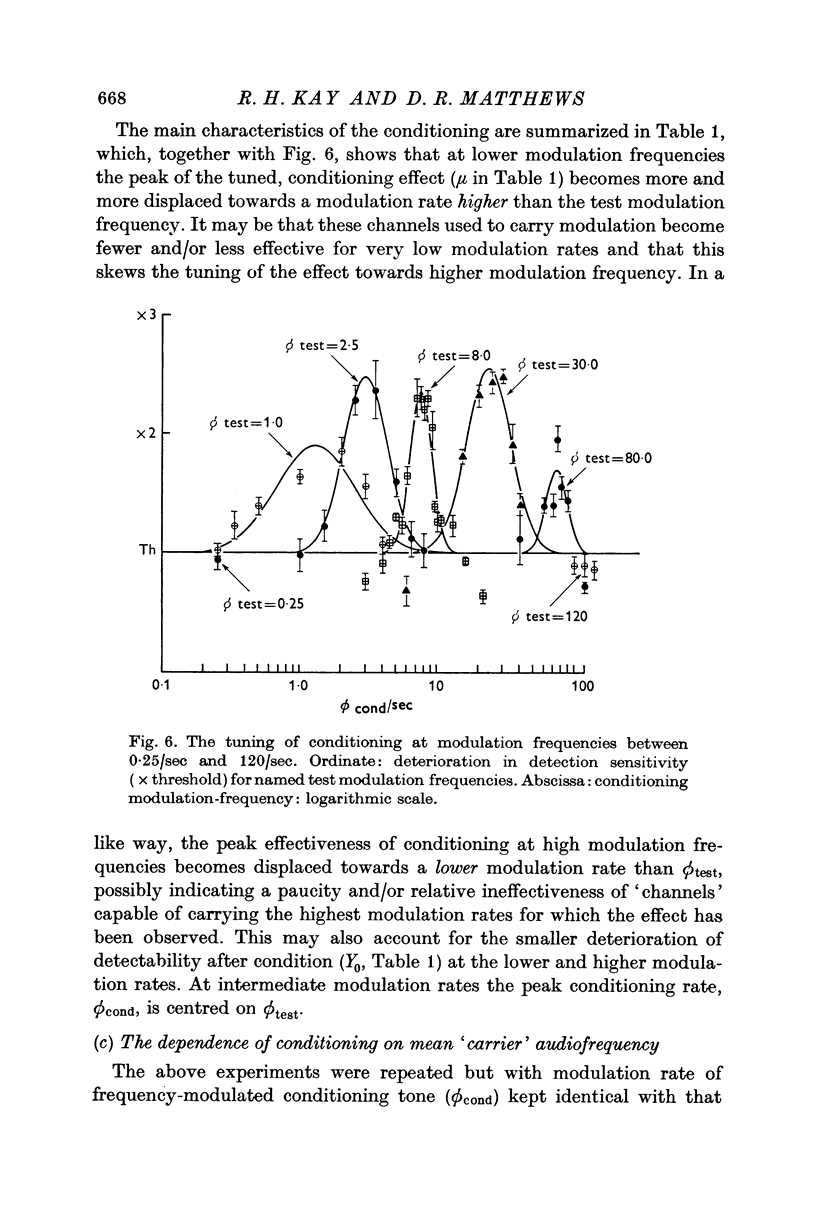
5. The magnitude of the deterioration in detection sensitivity after conditioning is about 3 × at modulation frequencies between about 3/sec and 30/sec. It diminishes at lower and higher modulation frequencies and is effectively absent at 100/sec modulation. The bandwidth of the effect increases from a few cycles per second at the lower end of this range, to some tens of cycles per second at the upper end.
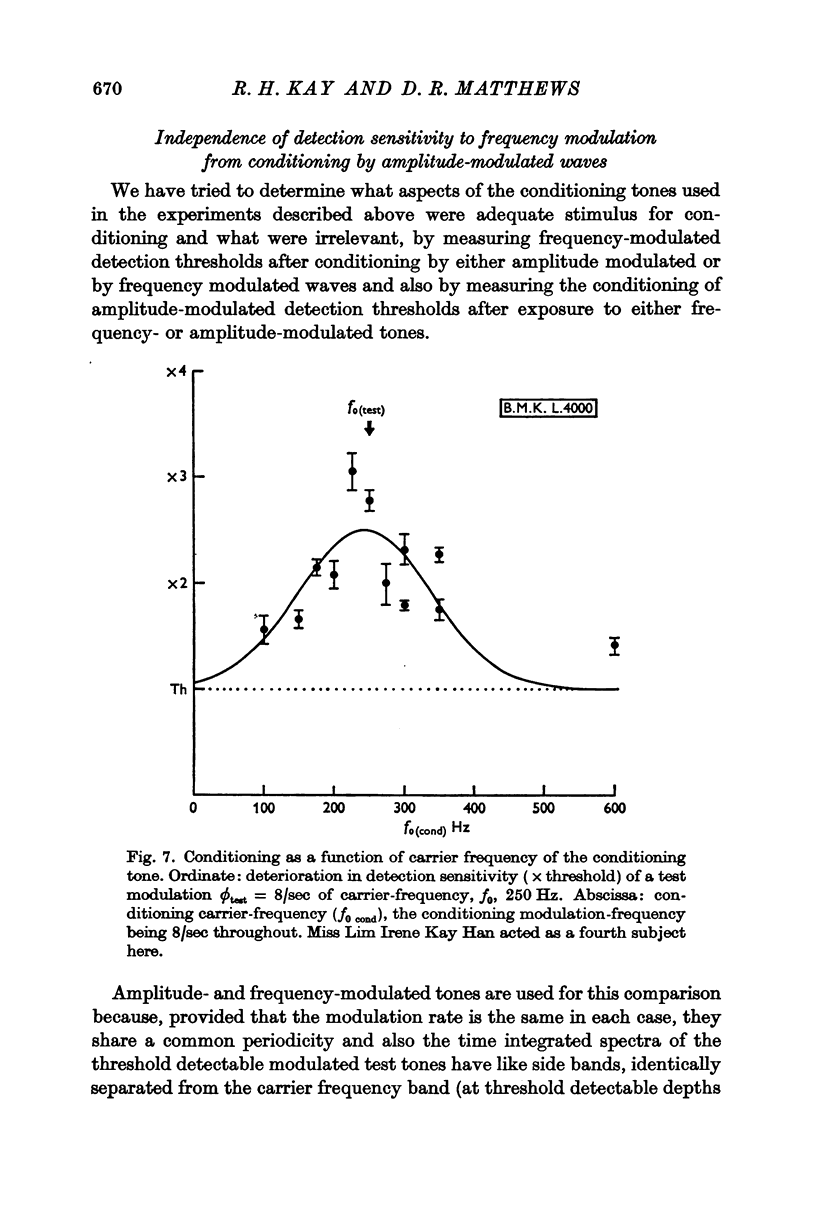
6. For the same modulation frequency, the conditioning is relatively insensitive to the mean `carrier' audiofrequency, f0. The band width in terms of carrier frequency is at least as wide as `critical bands'. With a test signal f0 = 250 Hz, ϕtest = 8/sec, conditioning is still appreciable for a conditioning tone of ϕcond = 8/sec but centred upon f0 = 150 Hz or = 350 Hz. Conditioning is thus not explicable in terms of coincidences between particular spectral components in the conditioning and test tones.
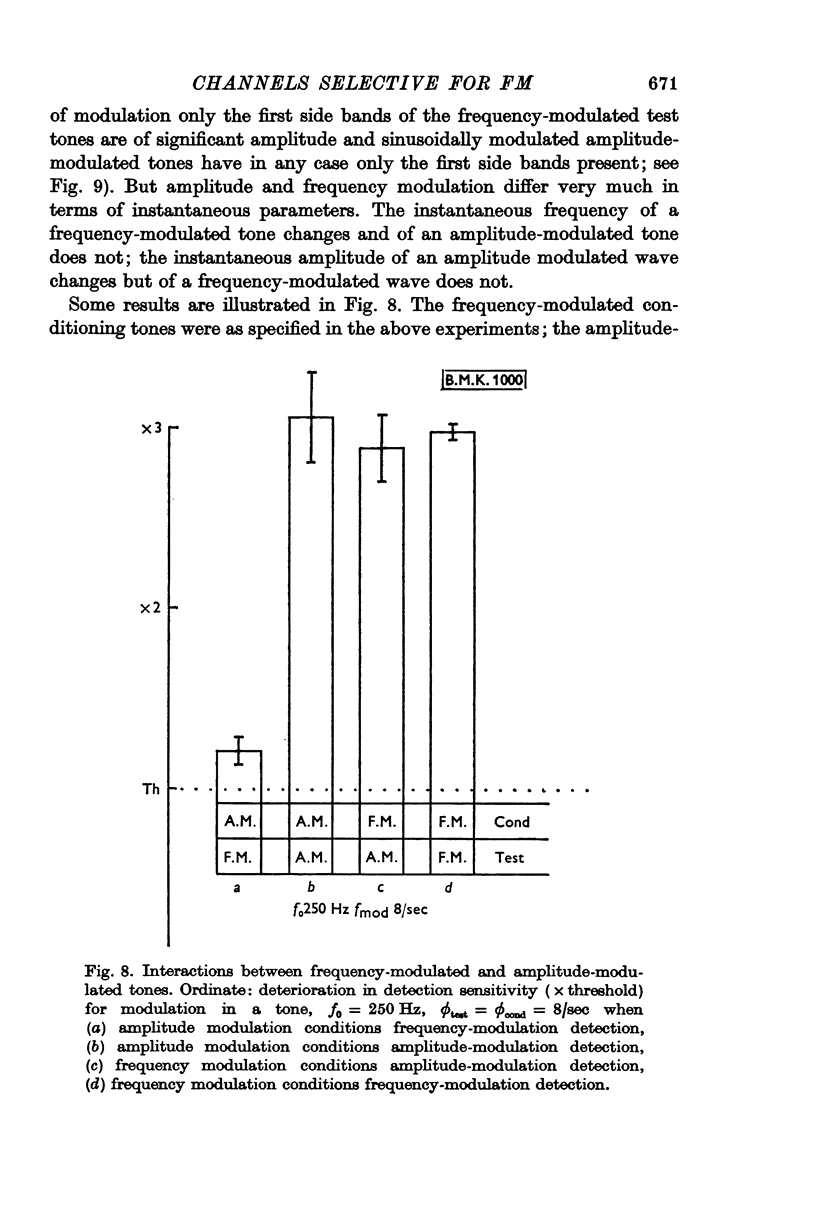
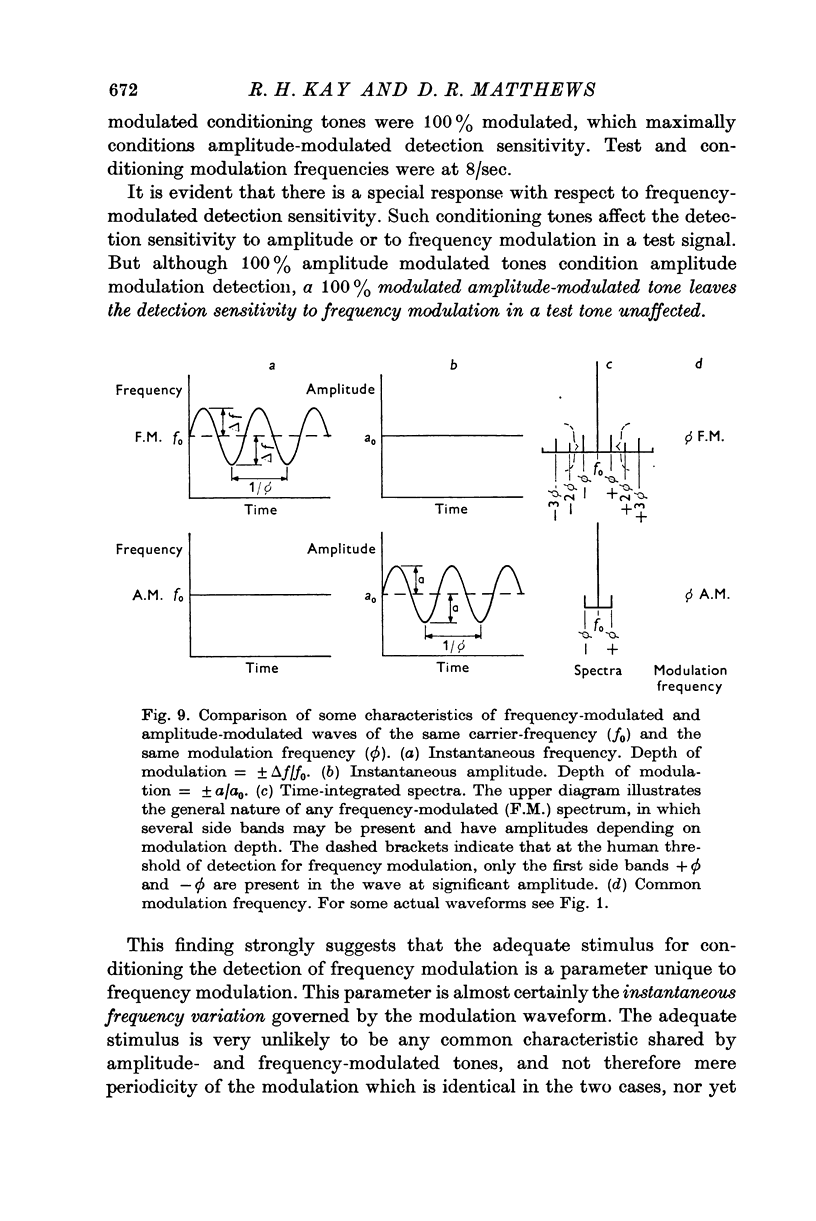
7. Whereas the sensitivity of detecting 8/sec amplitude modulation in a tone is conditioned by prior exposure to either amplitude- or frequency-modulated tones, in contrast the detectability of 8/sec frequency-modulated signals is conditioned only by prior exposure to frequency-modulated tones and not by amplitude-modulated conditioning tones. This underlines the special place of frequency modulation in human audition and emphasizes that the operative stimulus cannot be some aspect common to amplitude modulation and frequency modulation, like identical periodicity or coincident positioning of bands in the integrated spectra of the tones, but points to the instantaneous frequency-modulated wave form as the adequate stimulus.
8. These findings strongly suggest that the human auditory pathways contain `channels' in their organization which determine a final response selectively tuned to particular frequency-modulations. Periodicity coding alone cannot adequately explain this effect which may well only be understood in terms of a `place' theory of frequency selectivity.
9. This organization is well suited to subserve the recognition of frequency-modulation patterns in acoustic signals rather independently of the mean audiofrequency that carries the frequency modulation.
Full text
PDF


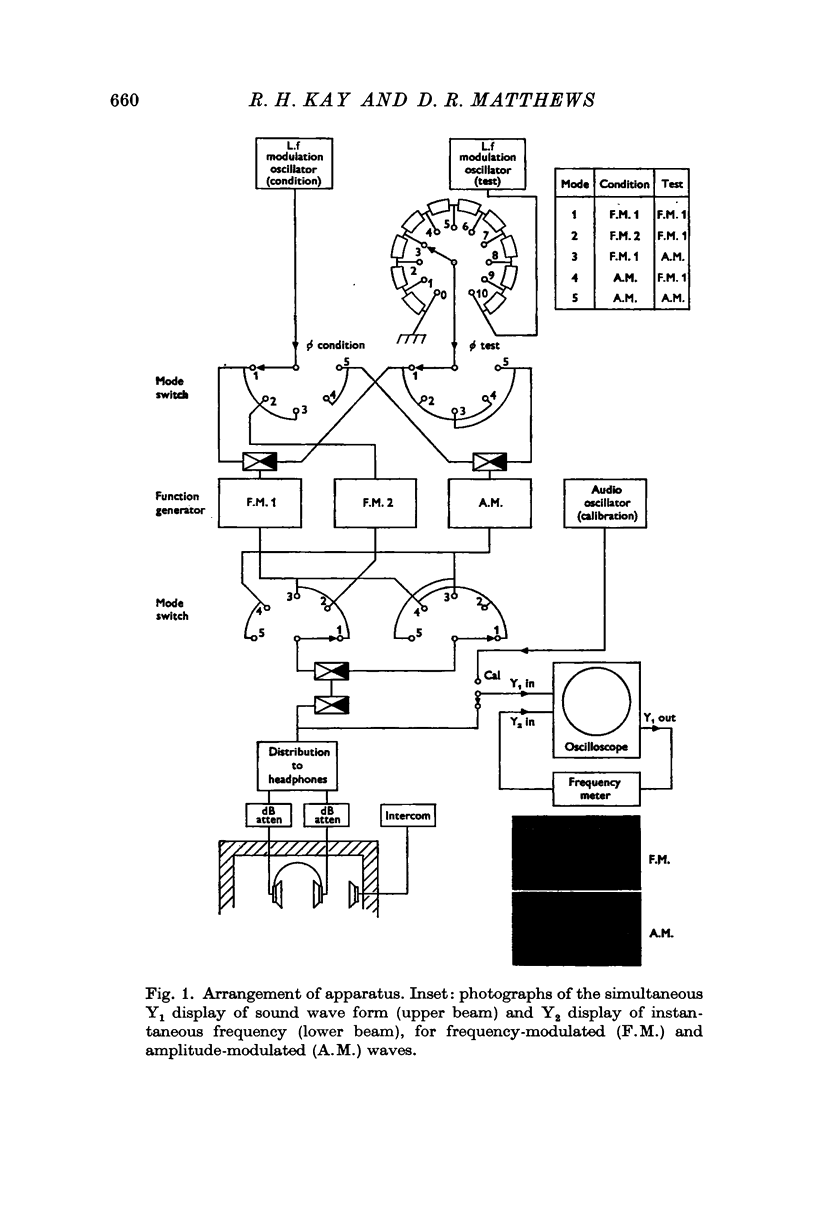


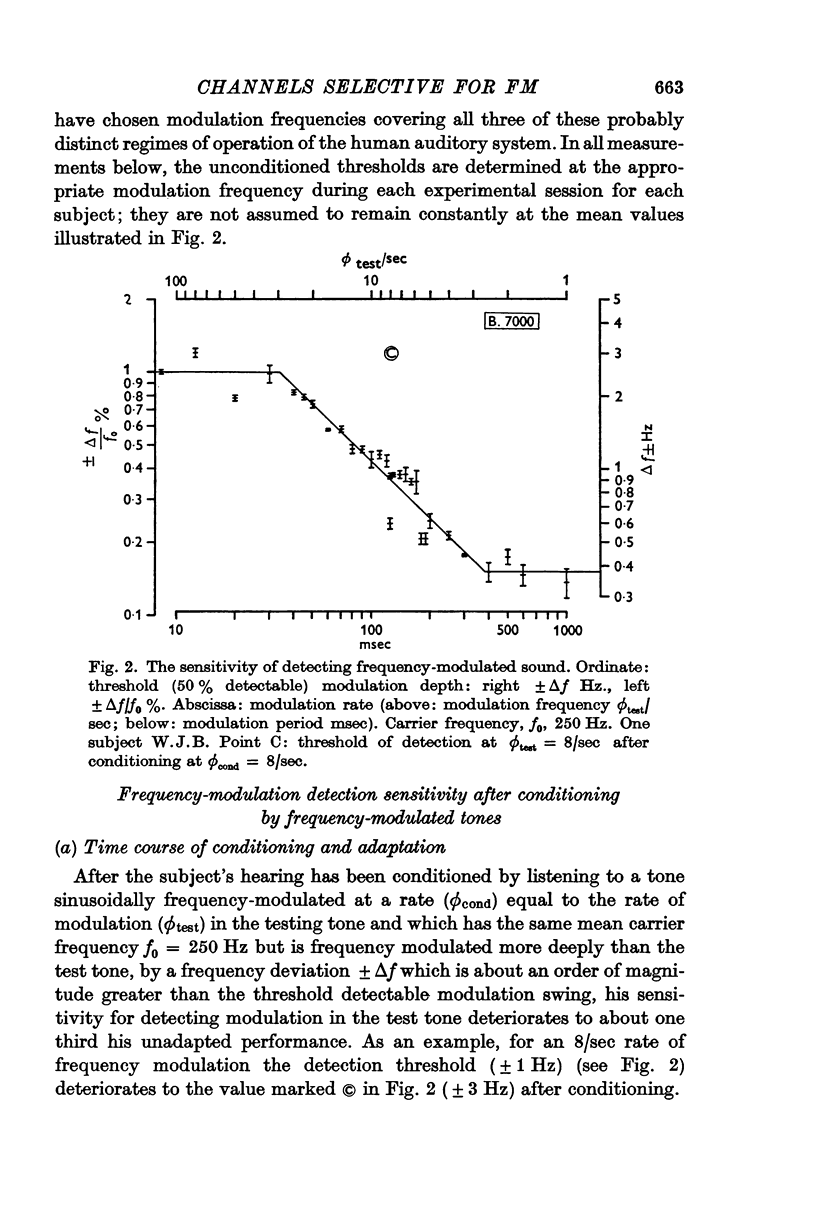
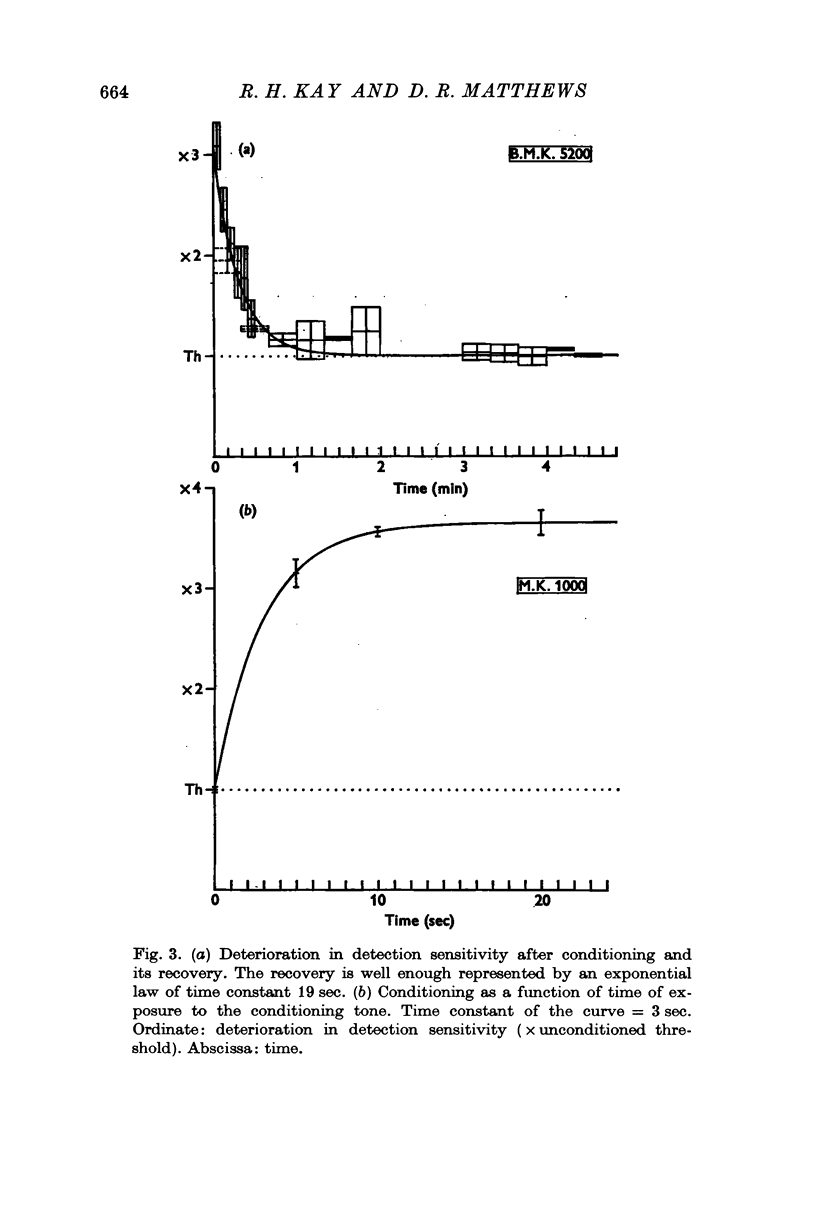

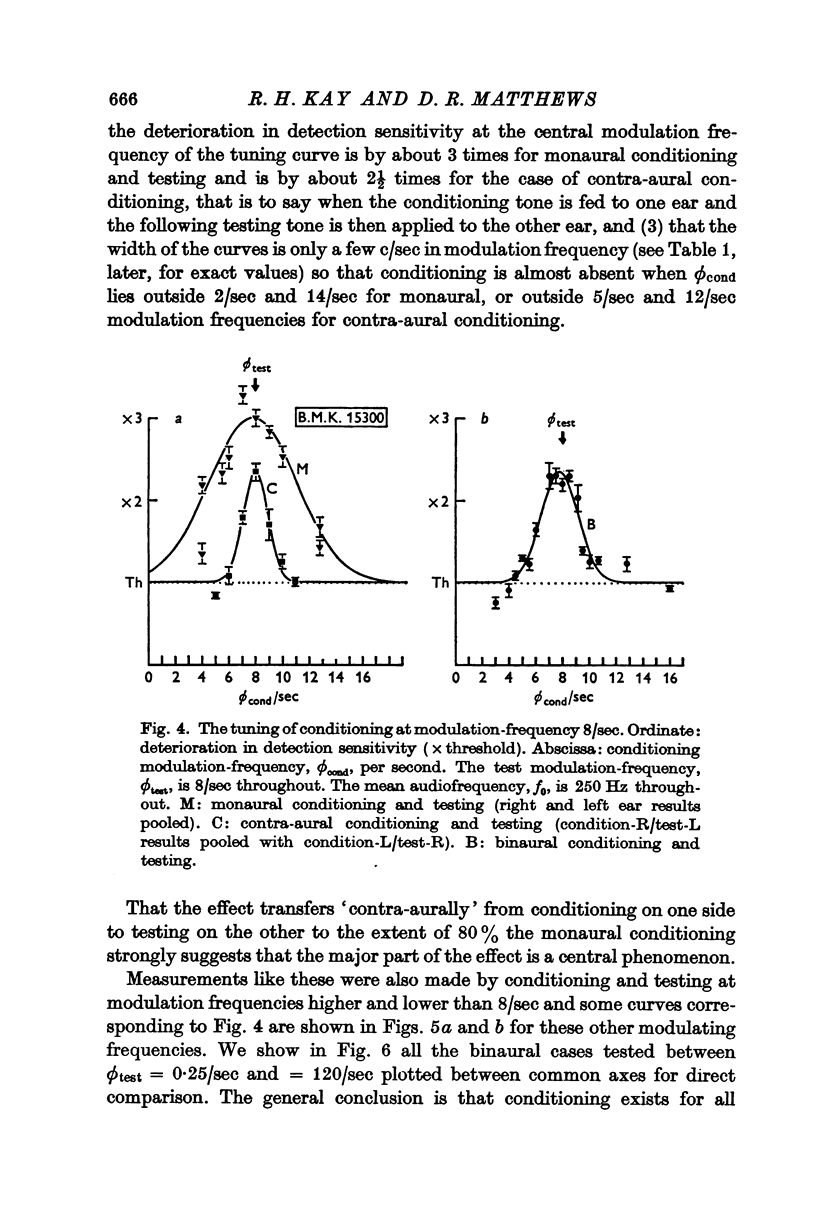






Selected References
These references are in PubMed. This may not be the complete list of references from this article.
- BARLOW H. B., HILL R. M., LEVICK W. R. RETINAL GANGLION CELLS RESPONDING SELECTIVELY TO DIRECTION AND SPEED OF IMAGE MOTION IN THE RABBIT. J Physiol. 1964 Oct;173:377–407. doi: 10.1113/jphysiol.1964.sp007463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barlow H. B., Levick W. R. The mechanism of directionally selective units in rabbit's retina. J Physiol. 1965 Jun;178(3):477–504. doi: 10.1113/jphysiol.1965.sp007638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blakemore C., Campbell F. W. On the existence of neurones in the human visual system selectively sensitive to the orientation and size of retinal images. J Physiol. 1969 Jul;203(1):237–260. doi: 10.1113/jphysiol.1969.sp008862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HUBEL D. H., WIESEL T. N. Receptive fields of single neurones in the cat's striate cortex. J Physiol. 1959 Oct;148:574–591. doi: 10.1113/jphysiol.1959.sp006308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KONIG E. Pitch discrimination and age. Acta Otolaryngol. 1957 Nov-Dec;48(5-6):475–489. doi: 10.3109/00016485709126909. [DOI] [PubMed] [Google Scholar]
- Kay R. H., Matthews D. R. Temporal specificity in human auditory conditioning by frequency-modulated tones. J Physiol. 1971 Oct;218 (Suppl):104P–106P. [PubMed] [Google Scholar]
- Kelly J. B., Whitfield I. C. Effects of auditory cortical lesions on discriminations of rising and falling frequency-modulated tones. J Neurophysiol. 1971 Sep;34(5):802–816. doi: 10.1152/jn.1971.34.5.802. [DOI] [PubMed] [Google Scholar]
- Moller A. R. Unit responses in the cochlear nucleus of the rat to sweep tones. Acta Physiol Scand. 1969 Aug;76(4):503–512. doi: 10.1111/j.1748-1716.1969.tb04497.x. [DOI] [PubMed] [Google Scholar]
- Nelson P. G., Erulkar S. D., Bryan J. S. Responses of units of the inferior colliculus to time-varying acoustic stimuli. J Neurophysiol. 1966 Sep;29(5):834–860. doi: 10.1152/jn.1966.29.5.834. [DOI] [PubMed] [Google Scholar]
- Smith R. A., Jr Studies of temporal frequency adaptation in visual contrast sensitivity. J Physiol. 1971 Aug;216(3):531–552. doi: 10.1113/jphysiol.1971.sp009539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHITFIELD I. C., EVANS E. F. RESPONSES OF AUDITORY CORTICAL NEURONS TO STIMULI OF CHANGING FREQUENCY. J Neurophysiol. 1965 Jul;28:655–672. doi: 10.1152/jn.1965.28.4.655. [DOI] [PubMed] [Google Scholar]