

Abstract
Decapping is a key step in general and regulated mRNA decay. In Saccharomyces cerevisiae it constitutes a rate-limiting step in the nonsense-mediated decay pathway that rids cells of mRNAs containing premature termination codons. Here two human decapping enzymes are identified, hDcp1a and hDcp2, as well as a homolog of hDcp1a, termed hDcp1b. Transiently expressed hDcp1a and hDcp2 proteins localize primarily to the cytoplasm and form a complex in human cell extracts. hDcp1a and hDcp2 copurify with decapping activity, an activity sensitive to mutation of critical hDcp residues. Importantly, coimmunoprecipitation assays demonstrate that hDcp1a and hDcp2 interact with the nonsense-mediated decay factor hUpf1, both in the presence and in the absence of the other hUpf proteins, hUpf2, hUpf3a, and hUpf3b. These data suggest that a human decapping complex may be recruited to mRNAs containing premature termination codons by the hUpf proteins.
Regulation of mRNA stability plays an important role in cellular modulation of gene expression (reviewed in references 4, 42, 50, and 65). mRNAs that are normally rapidly turned over can be stabilized upon cellular cues to cause accumulation of the protein product. For example, in mammals many proto-oncogene and interleukin mRNAs contain destabilizing AU-rich elements (AREs) in their 3′ untranslated regions (reviewed in reference 10). When the proteins are needed, specific cellular signals override the ARE destabilizing elements, resulting in stabilization of the mRNAs. mRNAs that are normally stable can also be destabilized to repress protein expression. For example, histone mRNAs are rapidly destabilized upon exit of the cell cycle S phase when DNA replication ceases and histone production is no longer needed (48). Proteins that regulate mRNA turnover do so by interaction with the cellular mRNA decay machinery, a machinery which is currently poorly understood for mammals. To understand how mRNA turnover is regulated, it is of fundamental importance to dissect the general mRNA decay pathways.
mRNA decapping is a key step in general and regulated mRNA decay in eukaryotes. In Saccharomyces cerevisiae the predominant pathway of mRNA decay proceeds via slow removal of the poly(A) tail by deadenylation, followed by decapping and 5′-to-3′ exonucleolytic decay (5, 14, 25, 43, 56, 58, 59). Alternatively, deadenylated transcripts can be degraded from the 3′ end by the exosome, a complex of 3′-to-5′ exonucleases (1, 26). Although deadenylated decapped mRNA species can be detected (12), exosome-mediated mRNA decay may be the predominant pathway in mammalian cells (9, 45, 63).
Eukaryotes possess specific mRNA surveillance pathways that serve to deplete the cell of irregular mRNAs (reviewed in reference 61). One recently discovered mRNA surveillance process, called nonstop decay, uses an EF1A-like GTPase, Ski7, that recruits the exosome to degrade mRNAs that lack translation termination codons (20, 60). Such mRNAs are believed to arise mainly from premature polyadenylation within the open reading frame (ORF) (20). Other irregular mRNAs, which have acquired premature termination codons (PTCs) by mutation or erroneous processing, are degraded by the process of nonsense-mediated decay (NMD) (reviewed in references 19, 23, 24, 27, 34, 36, 39, and 61). In mammals, PTCs are recognized by their position relative to the last mRNA exon-exon junction (46). Recent studies have shown that a multisubunit exon-junction complex (EJC) is deposited 20 to 24 nucleotides upstream of exon-exon junctions after pre-mRNA splicing (32). A translation termination event upstream of one or more EJCs triggers NMD (38). This is mediated by three hUpf proteins, hUpf1, -2, and -3, which interact with both the EJC (29, 31, 38) and translation termination factors eRF1 and eRF3 (13, 62). How the Upf proteins that mediate the NMD process trigger decay is largely unknown. In S. cerevisiae the first step in NMD is decapping, which is followed by 5′-to-3′ exonucleolytic decay (44).
Decapping is thus a key step in both NMD and general mRNA decay. In S. cerevisiae, two interacting proteins involved in decapping have been identified, called Dcp1p and Dcp2p (5, 16, 30). Dcp1p appears to be responsible for the actual decapping activity, whereas Dcp2p is necessary for Dcp1p activity. Very little is known about decapping in mammals. A decapping activity that is activated on mRNAs lacking a poly(A) tail or containing AREs has been detected in mammalian cell extracts (6, 21). Moreover, a separate decapping activity associated with the exosome is responsible for ridding the cell of the m7GpppN product of a complete 3′-to-5′ exonucleolytic degradation (63).
To gain insight into the process of mRNA decapping in mammals, human homologs of S. cerevisiae decapping enzymes Dcp1p and Dcp2p, termed hDcp1a, hDcp1b, and hDcp2, were identified. hDcp1a and hDcp2 proteins interact with each other and coimmunopurify with decapping activity in vitro, an activity sensitive to mutation of critical hDcp residues. Transiently expressed hDcp1a and hDcp2 localize to the cytoplasm of human HeLa cells. Moreover, hDcp1a and hDcp2 coimmunoprecipitate with the NMD protein hUpf1, providing a possible link between NMD and decapping.
MATERIALS AND METHODS
Plasmids.
Plasmids pcDNA3-FLAG-hDcp1a and pcDNA3-Myc-hDcp1a contain the full-length ORF of hDcp1a cDNA inserted between EcoRI and NotI sites of plasmids pcDNA3-FLAG and pcDNA3-Myc, which are derivatives of pcDNA3 (Invitrogen) containing a FLAG or a Myc tag, respectively, upstream of a modified polylinker (sequences available upon request). pcDNA3-FLAG-hDcp1a D20A and R59A contain GAC-to-GCC and AGG-to-GCG mutations at codons 20 and 59, respectively. pcDNA3-FLAG-hDcp1a 1-411, 1-251, and 1-146 contain N-terminal fragments of hDcp1a of 411, 251, and 146 codons, respectively. pcDNA3-FLAG-hDcp2 and pcDNA3-Myc-hDcp2 contain the full-length ORF of hDcp2 cDNA inserted between BamHI and NotI sites of pcDNA3-FLAG and pcDNA3-Myc. pcDNA3-FLAG-hDcp2 E148Q contains a GAA-to-CAA mutation at codon 148. Plasmids pcDNA3-FLAG-RNPS1, -hUpf3b, and -hUpf1 have been described earlier (37, 38). pcDNA3-Myc-hUpf1 contains the hUpf1 ORF inserted between BamHI and NotI sites of pcDNA3-Myc.
In vitro decapping assays.
Human embryonic kidney 293 (HEK293) cells were grown in Dulbecco modified Eagle medium (Invitrogen) containing 10% fetal bovine serum (Invitrogen) in 150-mm-diameter dishes and transiently transfected with a total of 20 μg of plasmid expressing FLAG-tagged hDcp proteins, by using Lipofectamine according to the manufacturer's directions (Invitrogen). Forty hours after transfection, ∼2.5 × 107 cells were washed and scraped in phosphate-buffered saline, and after pelleting, they were lysed in 1 ml of hypotonic gentle lysis buffer (10 mM Tris-HCl [pH 7.5], 10 mM NaCl, 2 mM EDTA, 0.5% Triton X-100, 0.5 mM phenylmethylsulfonyl fluoride, 2 μg of leupeptin/ml, 2 μg of aprotinin/ml) for 10 min on ice. NaCl was added to 150 mM, and the extracts were incubated for another 5 min on ice. Cell debris was removed by centrifugation, and the supernatant was nutated with 50 μl of anti-FLAG M2 agarose (Sigma) for 4 h at 4°C. Beads were washed eight times with 1 ml of NET-2 (50 mM Tris-HCl [pH 7.5], 150 mM NaCl, 0.05% Triton X-100), and FLAG-tagged protein was eluted by gently shaking the beads in 50 μl of NET-2 containing 1 mg of FLAG peptide/ml and 0.1 mg of bovine serum albumin/ml at 4°C for 2 h. Eluates were stored at −80°C.
A 5′ m7cap-labeled RNA substrate was produced by incubating an 88-nucleotide T7 RNA polymerase transcript with [α-32P]GTP, S-adenosyl-l-methionine (Sigma), and bacterially expressed vaccinia virus capping enzyme (construct kindly provided by Stewart Shuman) as described by Zhang et al. (67). Decapping reactions were carried out at 30°C for 2 h in a total of 10 μl of 50 mM Tris-HCl (pH 8.0)-30 mM (NH4)2SO4-1 mM MgCl2 containing ∼30 nCi of cap-labeled RNA and 0.2 to 10 μl of immunopurified decapping enzyme. In one experiment, the decapping reaction mixture was subsequently incubated with 0.5 mM ATP and 0.5 U of nucleoside diphosphate kinase (Sigma) at 30°C for 30 min. All reactions were terminated by addition of EDTA to 10 mM, and 3 μl was separated by thin-layer chromatography as described by Zhang et al. (67).
Coimmunoprecipitation assays.
RNase-treated cell lysates of ∼107 transiently transfected HEK293 cells from one 100-mm-diameter dish, generated as described above, were incubated for 4 h at 4°C with 25 μl of anti-FLAG M2 agarose (Sigma). Beads were washed 10 times with NET-2. Bound protein was eluted by incubation with 20 μl of NET-2 containing 1 mg of FLAG peptide/ml for 2 h at 4°C, followed by addition of 20 μl of sodium dodecyl sulfate (SDS) sample buffer to the eluate. In some experiments, the cell lysates were immunodepleted prior to immunoprecipitation, by incubation for 4 h at 4°C with 10 mg of protein A-Sepharose (Amersham/Pharmacia) preincubated with 40 μl of rabbit anti-hUpf serum (37), followed by clearance of the extract by incubation for 30 min at 4°C with 5 mg of protein A-Sepharose. Immunoprecipitates were separated by SDS-polyacrylamide gel electrophoresis followed by Western blotting.
Nucleotide sequence accession numbers.
GenBank accession numbers for hDcp1a, hDcp1b, and hDcp2 are AY146651, AY146652, and AY146650, respectively.
RESULTS
Human homologs of S. cerevisiae decapping enzymes.
In order to understand the process of mRNA decapping in mammals, the expressed sequence tag and human genome databases were searched for homologs of S. cerevisiae decapping enzymes Dcp1p and Dcp2p. Two distant human homologs of S. cerevisiae Dcp1p, called hDcp1a and hDcp1b, and a single homolog of S. cerevisiae Dcp2p, called hDcp2, were identified (Fig. 1). hDcp1a and hDcp1b each have in the N terminus two regions of 26% identity to the N and C termini of the 231-amino-acid S. cerevisiae Dcp1p protein, respectively (Fig. 1A). Importantly, the regions of similarity include residues important for S. cerevisiae Dcp1p activity as determined by alanine mutation scanning (57). In contrast, the C-terminal ∼450 amino acids of hDcp1 proteins show no similarity to S. cerevisiae Dcp1p. hDcp1a and hDcp1b share 31% identity over their entire length and 68% identity within the N-terminal conserved region. hDcp2 exhibits 36% identity to a 218-amino-acid region of S. cerevisiae Dcp2p (Fig. 1B), including a mutT domain which is essential for Dcp2p function (16). Moreover, a C-terminal region of ∼50 amino acids shows 30% identity between hDcp2 and S. cerevisiae Dcp2p. hDcp1a was recently identified as a Smad4-interacting protein, called SMIF (2), whereas hDcp1b and hDcp2 are of unknown function.
FIG. 1.

Human homologs of S. cerevisiae decapping enzymes. (A) Schematics of S. cerevisiae Dcp1p (yDcp1) and the human homologs hDcp1a and hDcp1b. (B) Schematics of S. cerevisiae Dcp2p (yDcp2) and the human homolog hDcp2. Gray bars indicate regions of similarity (percent identity given below) between human and S. cerevisiae Dcps. The black bars in yDcp2 and hDcp2 indicate the conserved mutT domains. Residues critical for decapping activity are indicated (see Fig. 4). aa, amino acids.
hDcp1a and hDcp2 interact in an RNA-independent manner.
S. cerevisiae Dcp1p and Dcp2p are interacting proteins (16). To test if hDcp1a and hDcp2 also interact, HEK293 cells were transiently cotransfected with plasmids expressing FLAG-tagged hDcp2 and Myc-tagged hDcp1a and, as a negative control, Myc-tagged hnRNPA1. Forty hours after transfection, cell extracts were prepared, RNase treated, and subjected to an anti-FLAG resin. Immunoprecipitates were subsequently tested for the presence of Myc-hDcp1a by Western blotting. As seen in Fig. 2A, ∼3 to 4% of total Myc-hDcp1a copurifies with FLAG-hDcp2 (compare the precipitate in lane 3 to 5% of total extract in lane 1), whereas no Myc-hDcp1a is found in immunoprecipitates from cells transfected with an empty vector (lane 2). Under the conditions used here, hDcp1a is expressed at about 2-fold-lower levels than hnRNPA1 but at about 10- to 15-fold-higher levels than hDcp2. Therefore, about 30 to 60% of FLAG-hDcp2 is complexed with Myc-hDcp1a. In contrast, no Myc-hnRNPA1 was observed in the precipitate (lower panel). To confirm these results, Myc-tagged hDcp2 was also tested for copurification with FLAG-hDcp1a. In this case, about 30 to 40% of Myc-hDcp2 copurified with FLAG-hDcp1a (Fig. 2B, lane 3). hDcp1a consistently appears as a doublet in denaturing gels (Fig. 2; see also Fig. 6), perhaps due to protein modification. It is concluded that hDcp1a and hDcp2 interact in an RNA-independent manner.
FIG. 2.
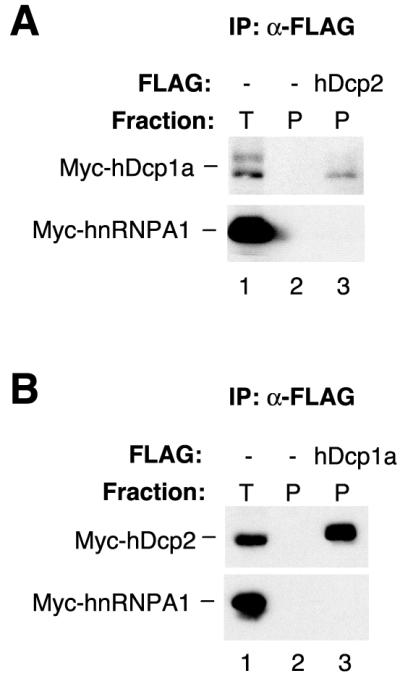
hDcp1a and hDcp2 interact in cell extracts. (A) Western blot for Myc-tagged hDcp1a and hnRNPA1 proteins, which were transiently expressed in HEK293 cells together with an empty vector (lanes 1 and 2) or a plasmid encoding FLAG-tagged hDcp2 (lane 3) and subjected to anti-FLAG immunoprecipitation after RNase treatment. (B) Western blot for Myc-tagged hDcp2 and hnRNPA1 proteins, transiently expressed in HEK293 cells together with an empty vector (lanes 1 and 2) or a plasmid encoding FLAG-tagged hDcp1a (lane 3) and subjected to anti-FLAG immunoprecipitation after RNase treatment. Immunoprecipitates (P; lanes 2 and 3) are compared to 5% of total extract (T; lane 1).
FIG. 6.

hDcp1a and hDcp2 interact with the NMD protein hUpf1. (A) Western blot for Myc-tagged hDcp1a, hDcp2, or hnRNPA1, transiently expressed in HEK293 cells together with an empty vector (lanes 1 and 2) or a plasmid encoding FLAG-tagged hUpf1 (lane 3) and subjected to anti-FLAG immunoprecipitation after RNase treatment. (B) Western blot for Myc-tagged hUpf1 or hnRNPA1, transiently expressed in HEK293 cells together with an empty vector (lanes 1, 2, 4, and 5) or a plasmid encoding FLAG-tagged hDcp1a (lane 3) or hDcp2 (lane 6) and subjected to anti-FLAG immunoprecipitation after RNase treatment. Immunoprecipitates (P; lanes 2, 3, 5, and 6) are compared to 5% of the total extract (T; lanes 1 and 4) in panels A and B. (C) RNase-treated extracts from HEK293 cells transiently expressing Myc-hDcp1a, Myc-hnRNPA1, and FLAG-hUpf1 were immunodepleted with preimmune serum (pi; lanes 1, 2, and 3) or anti-hUpf1 (α1; lane 4), anti-hUpf2 (α2; lane 5), or anti-hUpf3b (α3b; lane 6) serum and subsequently immunoprecipitated with anti-FLAG antibody. Immunoprecipitates (upper two panels) were probed for the presence of Myc-hDcp1a and Myc-hnRNPA1 by Western blotting. Five percent of the total extract (T; lane 1) was compared to the pellets (P; lanes 2 to 5). Depleted extracts (D; lower five panels, lanes 3 to 6) were probed with monoclonal anti-Myc antibody or polyclonal rabbit antibodies against hUpf1, hUpf2, hUpf3a, and hUpf3b as indicated.
Decapping activity of hDcp proteins in vitro.
To test if hDcp proteins are associated with decapping activity, N-terminally FLAG-tagged hDcp1a and hDcp2 were immunopurified from transiently transfected HEK293 cells by passing cell extracts over an anti-FLAG antibody column followed by elution with a FLAG peptide (see Materials and Methods). The eluates were tested for decapping activity in vitro by incubation with an m7cap-labeled RNA substrate. As indicated by release of m7GDP, both FLAG-hDcp1a and FLAG-hDcp2 copurify with decapping activity (Fig. 3, lanes 4 to 7). Eluates from cells transfected with an empty vector served as a negative control and showed no decapping (lanes 2 and 3). The products released by immunopurified FLAG-hDcps correspond to m7GDP, because they can be chased into m7GTP by using nucleoside diphosphate kinase and ATP as shown for hDcp1a in lane 10.
FIG. 3.

Decapping assays with immunopurified and recombinant hDcp1a and hDcp2. α-32P-m7cap-labeled RNA was incubated with immunopurified FLAG-tagged hDcp1a (lanes 4, 5, 9, and 10; corresponding to 0.2 × 106, 1 × 106, 1 × 106, and 1 × 106 cells' worth of protein, respectively), hDcp2 (lanes 6 and 7; 1 × 106 and 5 × 106 cells' worth, respectively), mock-purified cell extract (lanes 2 and 3; 1 × 106 and 5 × 106 cells' worth, respectively), or no cell extract (lanes 1, 8, and 11) or with purified bacterially expressed GST (lanes 12 and 13; 10 and 100 ng, respectively), GST-hDcp1a (lanes 14 and 15; 10 and 100 ng, respectively), GST-hDcp2 (lanes 16 and 17; 10 and 100 ng, respectively), or GST-hDcp1a and GST-hDcp2 (lanes 18 and 19; 10 and 100 ng each, respectively), and products were separated by thin-layer chromatography. The reaction mixture in lane 10 was treated with nucleoside diphosphate kinase (NDPK) and ATP. The migration of unlabeled GMP, GDP, m7GMP, m7GDP, and m7GTP (25 μg) is indicated on the left.
Purified, bacterially produced hDcp1a and hDcp2 proteins, fused with glutathione S-transferase (GST), were also tested for decapping activity in vitro. As seen in Fig. 3, GST-hDcp2 exhibits decapping activity (lanes 16 and 17). In contrast, GST-hDcp1a showed no detectable activity (lanes 14 and 15) and was not capable of stimulating the activity of GST-hDcp2 (compare lanes 18 and 19 to lanes 16 and 17). Similar observations were originally reported by E. van Dijk and coworkers (B. Séraphin, personal communication).
Critical mutations in hDcp1a and hDcp2 disrupt decapping activity.
The decapping activity of immunopurified hDcp1a and hDcp2 could originate from either the hDcps themselves or unknown associated factors. To render it likely that the hDcp proteins are responsible for the observed decapping activity, immunopurified hDcp1a and hDcp2 proteins mutated at residues previously shown to be critical for S. cerevisiae Dcp activity (16, 57) (Fig. 1) were tested for decapping activity (Fig. 4). The hDcp1a D20A and R59A immunoprecipitates were both impaired in decapping by 4- to 12-fold (lanes 4 and 5). Interestingly, it was observed by Western blotting that both hDcp1a D20A and R59A proteins were expressed at 10- to 20-fold-lower levels than wild-type hDcp1 (data not shown), but the amount of added immunoprecipitate was adjusted to use a similar amount of FLAG-hDcp in each decapping reaction (Fig. 4, lower panel). Mutation of a conserved mutT domain glutamate residue (E148Q) in hDcp2, which was previously demonstrated to be critical in other mutT domain proteins including S. cerevisiae Dcp2p (16, 35, 51), results in 20-fold-lower activity (Fig. 4, lane 10). Unlike the hDcp1a mutants, hDcp2 E148Q is as stable as the wild-type protein (data not shown). Immunoprecipitates containing FLAG-tagged hDcp1a deletion mutants were also tested for decapping activity. Although only the N-terminal 133-amino-acid region of hDcp1a is similar to S. cerevisiae Dcp1p, each of the hDcp1a C-terminal deletion mutants tested were impaired in decapping (Fig. 4, lanes 6 to 8), even though they were all found to copurify with Myc-hDcp2 (data not shown).
FIG. 4.

Mutation of critical residues in hDcp1a and hDcp2 disrupts decapping activity. The figure shows the results of decapping assays with immunopurified wild-type and mutant hDcp1a and hDcp2 proteins as indicated. The amount of protein corresponds to 5 × 106, 0.5 × 106, 5 × 106, 5 × 106, 1 × 106, 0.5 × 106, 0.5 × 106, 5 × 106, and 5 × 106 cells in lanes 2 to 10, respectively. The relative amount of m7GDP generated by each hDcp mutant was quantified with a phosphorimager and is given below relative to that of wild-type hDcps, which was set to 100%. A fraction of each assay (lanes 3 to 10) was subjected to Western blotting with anti-FLAG antibodies, shown in the lower panel. wt, wild type.
Transiently expressed hDcp1a and hDcp2 localize primarily to the cytoplasm.
If hDcp proteins are responsible for mRNA decapping in human cells, they would be expected to primarily localize to the cytoplasm, where most mRNA decay is believed to take place. It was therefore tested where transiently expressed FLAG-tagged hDcp1a and hDcp2 proteins localize by indirect immunofluorescence. Human HeLa cells were transiently transfected with plasmids expressing FLAG-hDcp1a or FLAG-hDcp2. Forty hours after transfection, the cells were fixed with paraformaldehyde and permeabilized, and the localization of the FLAG-tagged proteins was assessed by indirect immunofluorescence (Fig. 5). Both FLAG-hDcp1a and FLAG-hDcp2 are found primarily in localized dots in the cytoplasm. However, for both FLAG-hDcp1a and FLAG-hDcp2, some nuclear staining is evident.
FIG. 5.

Transiently expressed hDcp1a and hDcp2 proteins localize primarily in the cytoplasm. The figure shows the results of immunocytochemical staining of fixed, permeabilized HeLa cells, transiently expressing FLAG-tagged hDcp1a (panels 1 to 3) or hDcp2 (panels 4 to 6). Cells were stained with anti-FLAG M2 monoclonal antibody and visualized with Texas red-conjugated anti-mouse immunoglobulin G antibody (panels 1 and 4). Nuclei are visualized with 4′,6′-diamidino-2-phenylindole (DAPI) (panels 2 and 5). Panels 1 to 6 show one cell, representative of at least 100 observed transfected cells.
hDcp1a and hDcp2 proteins interact with the NMD protein hUpf1.
Decapping may play a role in several mRNA decay pathways in human cells. In S. cerevisiae, the process of NMD triggers decapping by an unknown mechanism (44). One possibility is that decapping enzymes are recruited to nonsense-containing mRNAs by the Upf proteins that are central players in the NMD pathway. Supporting this view is the finding that S. cerevisiae Upf1p interacts with Dcp2p (also called NMD1) in a two-hybrid screen (22). To test if hDcp1a and hDcp2 interact with hUpf1, HEK293 cells were transiently cotransfected with plasmids expressing FLAG-tagged hUpf1 and Myc-tagged hDcp proteins. Myc-hnRNPA1 served as a negative control. In this experiment FLAG-tagged, as opposed to endogenous, hUpf1 was used for immunoprecipitations because of an observed low, but significant, cross-reaction between anti-hUpf1 antibodies and FLAG-hDcp1 (data not shown). Anti-FLAG immunoprecipitates from RNase-treated cell extracts were tested for the presence of Myc-tagged proteins by Western blotting. In contrast to Myc-hnRNPA1, which was undetectable in the precipitates, ∼2 to 4% of total Myc-hDcp1a and ∼50% of total Myc-hDcp2 copurified with FLAG-hUpf1 (Fig. 6A; compare the precipitates in lane 3 to 5% of total extract in lane 1). In this experiment, both hUpf1 and hnRNPA1 were expressed at about twofold-higher levels than hDcp1a, which in turn was expressed at 10- to 15-fold-higher levels than hDcp2 (data not shown). To confirm this result, Myc-hUpf1 was also tested for its ability to coimmunopurify with FLAG-tagged hDcp1a and hDcp2. As seen in Fig. 6B, ∼10% of total Myc-hUpf1 copurified with FLAG-hDcp1a and FLAG-hDcp2.
hUpf1 interacts with the other hUpf proteins, hUpf2, hUpf3a, and hUpf3b. To test if the interaction between hUpf1 and hDcp1a requires the presence of the other hUpf proteins, the ability of Myc-hDcp1a to coimmunoprecipitate with FLAG-hUpf1 was tested after depletion of endogenous hUpf proteins. Passage of cell extracts over anti-hUpf1 or anti-hUpf2 antibody columns resulted in complete depletion of endogenous hUpf1 or hUpf2 proteins, respectively, whereas the other hUpf proteins were affected twofold or less (Fig. 6C, lanes 4 and 5, lower five panels). In contrast, anti-hUpf3b antibodies depleted both of the homologous hUpf3b and hUpf3a proteins as well as hUpf2 (Fig. 6C, lane 6, lower four panels). This is most likely a result of cross-reaction between anti-hUpf3b antibodies and hUpf3a (data not shown) and because the equally abundant hUpf2 protein interacts strongly with the hUpf3 proteins (37, 40, 54). None of the antibodies significantly depleted Myc-hDcp1a from the extracts (Fig. 6C, third panel from top). As seen in Fig. 6C (upper panels) Myc-hDcp1a, but not Myc-hnRNPA1, coimmunoprecipitates with FLAG-hUpf1 even in the absence of hUpf2 (lane 5) or hUpf2, hUpf3a, and hUpf3b (lane 6). As expected, no Myc-hDcp1a is observed in the immunoprecipitate when hUpf1 has first been depleted (lane 4). Interestingly, the amount of Myc-hDcp1a coprecipitating with FLAG-hUpf1 is reduced three- to fivefold after depletion with anti-hUpf2 or anti-hUpf3b antibodies, even though hUpf1, which is 10- to 20-fold more abundant than hUpf2 and hUpf3 proteins (40), was not significantly affected by the depletions (compare lanes 5 and 6 with lane 3). This suggests that, even though a maximum of 5 to 10% of cellular hUpf1 can be complexed with hUpf2 and/or hUpf3 proteins at any given time, the majority of hUpf1 protein that interacts with hDcp1a is in a complex with hUpf2 and/or hUpf3. These data suggest a link between NMD and human decapping enzymes.
DISCUSSION
Several lines of evidence indicate that hDcp1a and hDcp2 are involved in mRNA decapping in human cells and that they may be implicated in NMD. First, hDcp1a and hDcp2 are interacting proteins, similar to their S. cerevisiae homologs (Fig. 2). Second, hDcp1a and hDcp2 copurify with decapping activity from human cells, and point mutations in critical hDcp1a and hDcp2 residues impair the decapping activity (Fig. 3 and 4). Third, recombinant GST-hDcp2 exhibits decapping activity (Fig. 3) (E. van Dijk and B. Séraphin, personal communication). Fourth, transiently expressed hDcp1a and hDcp2 proteins localize primarily in the cytoplasm, as would be expected for proteins involved in general mRNA decay (Fig. 5). Fifth, both hDcp1a and hDcp2 interact with hUpf1, a protein central to the NMD pathway (Fig. 6). Sequential depletion and immunoprecipitation experiments demonstrated that, although hUpf1 does not require the presence of hUpf2 and hUpf3 proteins for interaction with hDcp1a, most of hUpf1 complexed with hDcp1a is also in complex with hUpf2 and hUpf3 (Fig. 6C). Unfortunately, an attempt to directly implicate hDcps in NMD was unsuccessful, because expression of inactive hDcp mutants described for Fig. 4, as well as hDcp2 mutants with C-terminal deletions of 60, 120, or 180 amino acids, in HEK293 or HeLa cells had no dominant negative effects on a coexpressed β-globin NMD substrate (data not shown). Therefore, there is currently no direct evidence for the significance of the strong interaction between hUpf1 and the hDcp proteins.
Characteristics of hDcp proteins.
Two human homologs of S. cerevisiae Dcp1p were identified, hDcp1a and hDcp1b. It is possible that they function redundantly in decapping in human cells, although there is so far no evidence that hDcp1b is involved in decapping other than the global similarity to hDcp1a. Point mutations of the conserved D20 and R59 residues of hDcp1a led to impaired decapping activity (Fig. 4). Importantly, both of these residues are conserved in hDcp1b (Fig. 1A). The observation that hDcp1a D20A and R59A proteins were expressed at 10- to 20-fold-lower levels than wild-type hDcp1a indicates that they are unstable. Perhaps the poor decapping activity of the hDcp1a mutants is a result of impaired folding or of a loss of interaction with an important protein partner that would also lead to protein instability. The corresponding S. cerevisiae Dcp1p mutants do not appear to be unstable (57). Interestingly, hDcp1a (also called SMIF) may play a role in Smad-mediated transforming growth factor β signal transduction, because it was reported earlier to interact with Smad4 and to partially translocate to the nucleus in response to transforming growth factor β stimulation (2).
hDcp2 contains a mutT domain which is found in proteins that catalyze pyrophosphatase reactions (7). Since decapping involves cleavage of a pyrophosphate, the observation that mutation of a critical glutamate residue in the mutT domain of Dcp2 strongly impairs decapping activity (Fig. 4) (16) suggests that the Dcp2 protein is the enzyme responsible for catalysis. Moreover, recombinant GST-hDcp2, but not GST-hDcp1a, shows decapping activity (Fig. 3). However, purified S. cerevisiae Dcp1p exhibits decapping activity after separation from Dcp2p (17, 30), and immunopurified hDcp1a deletion mutants that maintain their ability to copurify with hDcp2 are impaired in decapping activity (Fig. 4, lanes 6 to 8). Most likely Dcp1 and Dcp2 form a complex required for decapping.
Recruitment of decapping enzymes to nonsense-containing mRNAs?
Although it remains to be established if NMD in mammals, as in S. cerevisiae, triggers decapping, a link between human Upf proteins and decapping enzymes has now been identified (Fig. 6). The data presented here suggest that hDcp1 and hDcp2 may be recruited to mRNAs by hUpf1 in complex with hUpf2 and hUpf3. Perhaps selective recruitment of decapping enzymes to PTC-containing mRNAs triggers NMD. This model is depicted in Fig. 7.
FIG. 7.

Model for how the interaction between hUpf1 and hDcp proteins may play a role in NMD. The EJC is deposited onto the mRNA upstream of exon-exon junctions after pre-mRNA splicing. hUpf3 associates with the EJC, followed by hUpf2 and hUpf1. A translation termination event upstream of the EJC/hUpf complex may allow hUpf1 to recruit the hDcp proteins to trigger mRNA decapping. RF, translation release factor.
How could recruitment of hDcp proteins lead to decapping? Several lines of evidence indicate that decapping does not occur on an mRNA without prior dissociation of the cap-binding complex eIF4F. For example, decapping by S. cerevisiae Dcp1p is inhibited by the cap-binding protein eIF4E in vitro (53), and mutations in eIF4F components that inhibit translation initiation lead to an increased rate of decapping in S. cerevisiae (3, 8, 52). Therefore, decapping may not be triggered by simply recruiting hDcps to the mRNA. In S. cerevisiae, the RNA helicase Dhh1p is implicated in decapping of deadenylated mRNAs, and it was proposed that Dhh1p may function by dissociating the cap-binding complex from the mRNA (11, 18). Importantly, it was demonstrated earlier that an RNA helicase can dissociate an RNA-protein complex (28). In the case of NMD, perhaps the conserved RNA helicase function of Upf1, which is critical for NMD (55, 64), could play a similar role. Interestingly, Upf2 contains three eIF4G-like repeats (37, 41, 49, 54), and hUpf2 can interact with eIF4A (41). Therefore, it is possible that Upf2 assists in breaking up the eIF4F complex by interacting with individual components (41).
If the model in Fig. 7 is correct, some mechanism must ensure that decapping enzymes are only recruited and/or activated on mRNAs containing PTCs, because hUpf proteins are believed to associate with all intron-containing mRNAs (31, 33, 37). Perhaps phosphorylation of hUpf1, which has been shown elsewhere to be crucial for NMD (15, 47, 66), is the trigger to recruit hDcps. Alternatively, phosphorylation of hDcps themselves may be necessary for activation. Note that hDcp1a produces a doublet in SDS-polyacrylamide gels (Fig. 2 and 6) and that S. cerevisiae Dcp1p is a phosphoprotein (30). These events may be triggered only if a complex of EJC, hUpf proteins, and eRFs is formed on an mRNA by a translation termination event upstream of an exon-exon junction (Fig. 7). The details of the possible significance of the interaction between hUpf1 and hDcp proteins for NMD of PTC-containing mRNAs are an important problem for future studies.
Acknowledgments
Norm Pace, Elsebet Lund, Morgan Tucker, and Eileen Wagner are thanked for advice on the manuscript. Morgan Tucker is also thanked for advice on in vitro decapping assays. Stewart Shuman is thanked for a vaccinia virus capping enzyme expression plasmid. Robert M. Ross is thanked for technical assistance.
REFERENCES
- 1.Allmang, C., E. Petfalski, A. Podtelejnikov, M. Mann, D. Tollervey, and P. Mitchell. 1999. The yeast exosome and human PM-Scl are related complexes of 3′→5′ exonucleases. Genes Dev. 13:2148-2158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bai, R. Y., C. Koester, T. Ouyang, S. A. Hahn, M. Hammerschmidt, C. Peschel, and J. Duyster. 2002. SMIF, a Smad4-interacting protein that functions as a co-activator in TGFβ signalling. Nat. Cell Biol. 4:181-190. [DOI] [PubMed] [Google Scholar]
- 3.Barnes, C. A. 1998. Upf1 and Upf2 proteins mediate normal yeast mRNA degradation when translation initiation is limited. Nucleic Acids Res. 26:2433-2441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Beelman, C. A., and R. Parker. 1995. Degradation of mRNA in eukaryotes. Cell 81:179-183. [DOI] [PubMed] [Google Scholar]
- 5.Beelman, C. A., A. Stevens, G. Caponigro, T. E. LaGrandeur, L. Hatfield, D. M. Fortner, and R. Parker. 1996. An essential component of the decapping enzyme required for normal rates of mRNA turnover. Nature 382:642-646. [DOI] [PubMed] [Google Scholar]
- 6.Bergman, N., M. Opyrchal, E. J. Bates, and J. Wilusz. 2002. Analysis of the products of mRNA decapping and 3′-to-5′ decay by denaturing gel electrophoresis. RNA 8:959-965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bessman, M. J., D. N. Frick, and S. F. O'Handley. 1996. The MutT proteins or “Nudix” hydrolases, a family of versatile, widely distributed, “housecleaning” enzymes. J. Biol. Chem. 271:25059-25062. [DOI] [PubMed] [Google Scholar]
- 8.Cereghino, G. P., D. P. Atencio, M. Saghbini, J. Beiner, and I. E. Scheffler. 1995. Glucose-dependent turnover of the mRNAs encoding succinate dehydrogenase peptides in Saccharomyces cerevisiae: sequence elements in the 5′ untranslated region of the Ip mRNA play a dominant role. Mol. Biol. Cell 6:1125-1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chen, C. Y., R. Gherzi, S. E. Ong, E. L. Chan, R. Raijmakers, G. J. Pruijn, G. Stoecklin, C. Moroni, M. Mann, and M. Karin. 2001. AU binding proteins recruit the exosome to degrade ARE-containing mRNAs. Cell 107:451-464. [DOI] [PubMed] [Google Scholar]
- 10.Chen, C. Y., and A. B. Shyu. 1995. AU-rich elements: characterization and importance in mRNA degradation. Trends Biochem. Sci. 20:465-470. [DOI] [PubMed] [Google Scholar]
- 11.Coller, J. M., M. Tucker, U. Sheth, M. A. Valencia-Sanchez, and R. Parker. 2001. The DEAD box helicase, Dhh1p, functions in mRNA decapping and interacts with both the decapping and deadenylase complexes. RNA 7:1717-1727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Couttet, P., M. Fromont-Racine, D. Steel, R. Pictet, and T. Grange. 1997. Messenger RNA deadenylylation precedes decapping in mammalian cells. Proc. Natl. Acad. Sci. USA 94:5628-5633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Czaplinski, K., M. J. Ruiz-Echevarria, S. V. Paushkin, X. Han, Y. Weng, H. A. Perlick, H. C. Dietz, M. D. Ter-Avanesyan, and S. W. Peltz. 1998. The surveillance complex interacts with the translation release factors to enhance termination and degrade aberrant mRNAs. Genes Dev. 12:1665-1677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Decker, C. J., and R. Parker. 1993. A turnover pathway for both stable and unstable mRNAs in yeast: evidence for a requirement for deadenylation. Genes Dev. 7:1632-1643. [DOI] [PubMed] [Google Scholar]
- 15.Denning, G., L. Jamieson, L. E. Maquat, E. A. Thompson, and A. P. Fields. 2001. Cloning of a novel phosphatidylinositol kinase-related kinase: characterization of the human SMG-1 RNA surveillance protein. J. Biol. Chem. 276:22709-22714. [DOI] [PubMed] [Google Scholar]
- 16.Dunckley, T., and R. Parker. 1999. The DCP2 protein is required for mRNA decapping in Saccharomyces cerevisiae and contains a functional MutT motif. EMBO J. 18:5411-5422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dunckley, T., M. Tucker, and R. Parker. 2001. Two related proteins, Edc1p and Edc2p, stimulate mRNA decapping in Saccharomyces cerevisiae. Genetics 157:27-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fischer, N., and K. Weis. 2002. The DEAD box protein Dhh1 stimulates the decapping enzyme Dcp1. EMBO J. 21:2788-2797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Frischmeyer, P. A., and H. C. Dietz. 1999. Nonsense-mediated mRNA decay in health and disease. Hum. Mol. Genet. 8:1893-1900. [DOI] [PubMed] [Google Scholar]
- 20.Frischmeyer, P. A., A. van Hoof, K. O'Donnell, A. L. Guerrerio, R. Parker, and H. C. Dietz. 2002. An mRNA surveillance mechanism that eliminates transcripts lacking termination codons. Science 295:2258-2261. [DOI] [PubMed] [Google Scholar]
- 21.Gao, M., C. J. Wilusz, S. W. Peltz, and J. Wilusz. 2001. A novel mRNA-decapping activity in HeLa cytoplasmic extracts is regulated by AU-rich elements. EMBO J. 20:1134-1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.He, F., and A. Jacobson. 1995. Identification of a novel component of the nonsense-mediated mRNA decay pathway by use of an interacting protein screen. Genes Dev. 9:437-454. [DOI] [PubMed] [Google Scholar]
- 23.Hentze, M. W., and A. E. Kulozik. 1999. A perfect message: RNA surveillance and nonsense-mediated decay. Cell 96:307-310. [DOI] [PubMed] [Google Scholar]
- 24.Hilleren, P., and R. Parker. 1999. Mechanisms of mRNA surveillance in eukaryotes. Annu. Rev. Genet. 33:229-260. [DOI] [PubMed] [Google Scholar]
- 25.Hsu, C. L., and A. Stevens. 1993. Yeast cells lacking 5′→3′ exoribonuclease 1 contain mRNA species that are poly(A) deficient and partially lack the 5′ cap structure. Mol. Cell. Biol. 13:4826-4835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jacobs, J. S., A. R. Anderson, and R. P. Parker. 1998. The 3′ to 5′ degradation of yeast mRNAs is a general mechanism for mRNA turnover that requires the SKI2 DEVH box protein and 3′ to 5′ exonucleases of the exosome complex. EMBO J. 17:1497-1506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jacobson, A., and S. W. Peltz. 2000. Destabilization of nonsense-containing transcripts in Saccharomyces cerevisiae, p. 827-847. In N. Sonenberg, J. W. B. Hershey, and M. B. Mathews (ed.), Translational control of gene expression. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
- 28.Jankowsky, E., C. H. Gross, S. Shuman, and A. M. Pyle. 2001. Active disruption of an RNA-protein interaction by a DExH/D RNA helicase. Science 291:121-125. [DOI] [PubMed] [Google Scholar]
- 29.Kim, V. N., N. Kataoka, and G. Dreyfuss. 2001. Role of the nonsense-mediated decay factor hUpf3 in the splicing-dependent exon-exon junction complex. Science 293:1832-1836. [DOI] [PubMed] [Google Scholar]
- 30.LaGrandeur, T. E., and R. Parker. 1998. Isolation and characterization of Dcp1p, the yeast mRNA decapping enzyme. EMBO J. 17:1487-1496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Le Hir, H., D. Gatfield, E. Izaurralde, and M. J. Moore. 2001. The exon-exon junction complex provides a binding platform for factors involved in mRNA export and nonsense-mediated mRNA decay. EMBO J. 20:4987-4997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Le Hir, H., E. Izzauralde, L. E. Maquat, and M. J. Moore. 2000. The spliceosome deposits multiple proteins 20-24 nucleotides upstream of mRNA exon-exon junctions. EMBO J. 19:6860-6869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lejeune, F., Y. Ishigaki, X. Li, and L. E. Maquat. 2002. The exon junction complex is detected on CBP80-bound but not eIF4E-bound mRNA in mammalian cells: dynamics of mRNP remodeling. EMBO J. 21:3536-3545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Li, S., and M. F. Wilkinson. 1998. Nonsense surveillance in lymphocytes? Immunity 8:135-141. [DOI] [PubMed] [Google Scholar]
- 35.Lin, J., C. Abeygunawardana, D. N. Frick, M. J. Bessman, and A. S. Mildvan. 1996. The role of Glu 57 in the mechanism of the Escherichia coli MutT enzyme by mutagenesis and heteronuclear NMR. Biochemistry 35:6715-6726. [DOI] [PubMed] [Google Scholar]
- 36.Lykke-Andersen, J. 2001. mRNA quality control: marking the message for life or death. Curr. Biol. 11:R88-R91. [DOI] [PubMed] [Google Scholar]
- 37.Lykke-Andersen, J., M. D. Shu, and J. A. Steitz. 2000. Human Upf proteins target an mRNA for nonsense-mediated decay when bound downstream of a termination codon. Cell 103:1121-1131. [DOI] [PubMed] [Google Scholar]
- 38.Lykke-Andersen, J., M. D. Shu, and J. A. Steitz. 2001. Communication of the position of exon-exon junctions to the mRNA surveillance machinery by the protein RNPS1. Science 293:1836-1839. [DOI] [PubMed] [Google Scholar]
- 39.Maquat, L. E. 2000. Nonsense-mediated RNA decay in mammalian cells: a splicing-dependent means to down-regulate the levels of mRNAs that prematurely terminate translation, p. 849-868. In N. Sonenberg, J. W. B. Hershey, and M. B. Mathews (ed.), Translational control of gene expression. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
- 40.Maquat, L. E., and G. Serin. 2001. Nonsense-mediated mRNA decay: insights into mechanism from the cellular abundance of human Upf1, Upf2, Upf3, and Upf3X proteins. Cold Spring Harbor Symp. Quant. Biol. 66:313-319. [DOI] [PubMed] [Google Scholar]
- 41.Mendell, J. T., S. M. Medghalchi, R. G. Lake, E. N. Noensie, and H. C. Dietz. 2000. Novel Upf2p orthologues suggest a functional link between the translation initiation and nonsense surveillance complexes. Mol. Cell. Biol. 20:8944-8957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Mitchell, P., and D. Tollervey. 2001. mRNA turnover. Curr. Opin. Cell Biol. 13:320-325. [DOI] [PubMed] [Google Scholar]
- 43.Muhlrad, D., C. J. Decker, and R. Parker. 1995. Turnover mechanisms of the stable yeast PGK1 mRNA. Mol. Cell. Biol. 15:2145-2156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Muhlrad, D., and R. Parker. 1994. Premature translational termination triggers mRNA decapping. Nature 370:578-581. [DOI] [PubMed] [Google Scholar]
- 45.Mukherjee, D., M. Gao, J. P. O'Connor, R. Raijmakers, G. Pruijn, C. S. Lutz, and J. Wilusz. 2002. The mammalian exosome mediates the efficient degradation of mRNAs that contain AU-rich elements. EMBO J. 21:165-174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nagy, E., and L. E. Maquat. 1998. A rule for termination-codon position within intron-containing genes: when nonsense affects RNA abundance. Trends Biochem. Sci. 23:198-199. [DOI] [PubMed] [Google Scholar]
- 47.Page, M. F., B. Carr, K. R. Anders, A. Grimson, and P. Anderson. 1999. SMG-2 is a phosphorylated protein required for mRNA surveillance in Caenorhabditis elegans and related to Upf1p of yeast. Mol. Cell. Biol. 19:5943-5951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Pandey, N. B., and W. F. Marzluff. 1987. The stem-loop structure at the 3′ end of histone mRNA is necessary and sufficient for regulation of histone mRNA stability. Mol. Cell. Biol. 7:4557-4559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ponting, C. P. 2000. Novel eIF4G domain homologues linking mRNA translation with nonsense-mediated mRNA decay. Trends Biochem. Sci. 25:423-426. [DOI] [PubMed] [Google Scholar]
- 50.Ross, J. 1996. Control of messenger RNA stability in higher eukaryotes. Trends Genet. 12:171-175. [DOI] [PubMed] [Google Scholar]
- 51.Safrany, S. T., S. W. Ingram, J. L. Cartwright, J. R. Falck, A. G. McLennan, L. D. Barnes, and S. B. Shears. 1999. The diadenosine hexaphosphate hydrolases from Schizosaccharomyces pombe and Saccharomyces cerevisiae are homologues of the human diphosphoinositol polyphosphate phosphohydrolase. Overlapping substrate specificities in a MutT-type protein. J. Biol. Chem. 274:21735-21740. [DOI] [PubMed] [Google Scholar]
- 52.Schwartz, D. C., and R. Parker. 1999. Mutations in translation initiation factors lead to increased rates of deadenylation and decapping of mRNAs in Saccharomyces cerevisiae. Mol. Cell. Biol. 19:5247-5256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Schwartz, D. C., and R. Parker. 2000. mRNA decapping in yeast requires dissociation of the cap binding protein, eukaryotic translation initiation factor 4E. Mol. Cell. Biol. 20:7933-7942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Serin, G., A. Gersappe, J. D. Black, R. Aronoff, and L. E. Maquat. 2001. Identification and characterization of human orthologues to Saccharomyces cerevisiae Upf2 protein and Upf3 protein (Caenorhabditis elegans SMG-4). Mol. Cell. Biol. 21:209-223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Sun, X., H. A. Perlick, H. C. Dietz, and L. E. Maquat. 1998. A mutated human homologue to yeast Upf1 protein has a dominant-negative effect on the decay of nonsense-containing mRNAs in mammalian cells. Proc. Natl. Acad. Sci. USA 95:10009-10014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Tharun, S., W. He, A. E. Mayes, P. Lennertz, J. D. Beggs, and R. Parker. 2000. Yeast Sm-like proteins function in mRNA decapping and decay. Nature 404:515-518. [DOI] [PubMed] [Google Scholar]
- 57.Tharun, S., and R. Parker. 1999. Analysis of mutations in the yeast mRNA decapping enzyme. Genetics 151:1273-1285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Tharun, S., and R. Parker. 2001. Targeting an mRNA for decapping: displacement of translation factors and association of the Lsm1p-7p complex on deadenylated yeast mRNAs. Mol. Cell 8:1075-1083. [DOI] [PubMed] [Google Scholar]
- 59.Tucker, M., and R. Parker. 2000. Mechanisms and control of mRNA decapping in Saccharomyces cerevisiae. Annu. Rev. Biochem. 69:571-595. [DOI] [PubMed] [Google Scholar]
- 60.van Hoof, A., P. A. Frischmeyer, H. C. Dietz, and R. Parker. 2002. Exosome-mediated recognition and degradation of mRNAs lacking a termination codon. Science 295:2262-2264. [DOI] [PubMed] [Google Scholar]
- 61.Wagner, E., and J. Lykke-Andersen. 2002. mRNA surveillance: the perfect persist. J. Cell Sci. 115:3033-3038. [DOI] [PubMed] [Google Scholar]
- 62.Wang, W., K. Czaplinski, Y. Rao, and S. W. Peltz. 2001. The role of Upf proteins in modulating the translation read-through of nonsense-containing transcripts. EMBO J. 20:880-890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wang, Z., and M. Kiledjian. 2001. Functional link between the mammalian exosome and mRNA decapping. Cell 107:751-762. [DOI] [PubMed] [Google Scholar]
- 64.Weng, Y., K. Czaplinski, and S. W. Peltz. 1996. Genetic and biochemical characterization of mutations in the ATPase and helicase regions of the Upf1 protein. Mol. Cell. Biol. 16:5477-5490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wilusz, C. J., M. Wormington, and S. W. Peltz. 2001. The cap-to-tail guide to mRNA turnover. Nat. Rev. Mol. Cell. Biol. 2:237-246. [DOI] [PubMed] [Google Scholar]
- 66.Yamashita, A., T. Ohnishi, I. Kashima, Y. Taya, and S. Ohno. 2001. Human SMG-1, a novel phosphatidylinositol 3-kinase-related protein kinase, associates with components of the mRNA surveillance complex and is involved in the regulation of nonsense-mediated mRNA decay. Genes Dev. 15:2215-2228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Zhang, S., C. J. Williams, M. Wormington, A. Stevens, and S. W. Peltz. 1999. Monitoring mRNA decapping activity. Methods 17:46-51. [DOI] [PubMed] [Google Scholar]