

Abstract
Hookworm infection is associated with growth delay and iron deficiency anemia in developing countries. A series of experiments were designed in order to test the hypothesis that host dietary iron restriction mediates susceptibility to hookworm infection using the hamster model of Ancylostoma ceylanicum. Animals were maintained on diets containing either 10 ppm iron (iron restricted) or 200 ppm iron (standard/high iron), followed by infection with A. ceylanicum third-stage larvae. Infected animals fed the standard diet exhibited statistically significant growth delay and reduced blood hemoglobin levels compared to uninfected controls on day 20 postinfection. In contrast, no statistically significant differences in weight or hemoglobin concentration were observed between infected and uninfected animals fed the iron-restricted diet. Moreover, iron-restricted animals were observed to have reduced intestinal worm burdens on day 10 and day 20 postinfection compared to those of animals maintained on the standard/high-iron diet. In a subsequent study, animals equilibrated on diets containing a range of iron levels (10 ppm, 40 ppm, 100 ppm, or 200 ppm) were infected with A. ceylanicum and followed for evidence of hookworm disease. Infected animals from the intermediate-dietary iron (40- and 100-ppm) groups exhibited greater weight loss and anemia than those in the low (10-ppm)- or high (200-ppm)-iron diet groups. Mortality was also significantly higher in the intermediate-dietary-iron groups. These data suggest that severe dietary iron restriction impairs hookworm development in vivo but that moderate iron restriction enhances host susceptibility to severe disease.
Hookworm infection and iron deficiency anemia occur commonly throughout much of the developing world (3, 28, 32, 33). Hookworms contribute to iron deficiency by actively feeding on blood from lacerated capillaries in the intestinal mucosa, resulting in significant gastrointestinal hemorrhage, loss of serum proteins, and intestinal inflammation (19-21). It is also well recognized that the cumulative effect of chronic hookworm infection and iron deficiency in children and women of reproductive age can be devastating (13, 23, 27, 34, 36), with detrimental effects on growth, as well as physical and cognitive development (1, 31, 35). As a result, strategies aimed at controlling the impact of hookworm infection on childhood nutrition often include combination therapy with anthelminthics and iron supplements (2, 9, 12, 15, 25, 33).
While deficiencies in essential micronutrients like iron are thought to modulate the risk of infection caused by a variety of pathogens, to date little is known about the role host iron status might play in disease caused by hookworms (10, 11). Previous studies have shown that Syrian hamsters infected by oral gavage with infective A. ceylanicum hookworm larvae in the third stage (L3) experience growth delay and anemia, beginning approximately 14 days postinfection (5-7, 16, 17). Interestingly, despite resolution of anemia and reduction in intestinal worm burden, animals do not reach the weights of uninfected, age-matched controls as far out as 100 days postinfection (5), similar to growth kinetics observed in children who acquire hookworm infection in areas of endemicity (8, 30, 31). Thus, the hamster model of A. ceylanicum is an appropriate system for characterizing the pathogenesis of hookworm infection and is particularly well suited for probing host-parasite interactions in vivo.
In order to characterize the impact of host nutritional status on hookworm pathogenesis, experiments were designed to test the hypothesis that dietary iron impacts susceptibility to hookworm-associated anemia and growth delay. We report here evidence to support a bidirectional role for host iron in mediating hookworm pathogenesis, influencing both parasite development and host susceptibility to severe disease. Ultimately, these studies may provide new insights into the evolutionary mechanisms through which host and parasite compete for specific micronutrients.
(This work was presented in part at the 53rd Annual Meeting of the American Society of Tropical Medicine and Hygiene, Miami, FL, in 2004.)
MATERIALS AND METHODS
Parasite and host species.
All animal studies were approved by the Yale University Animal Care and Use Committee. The A. ceylanicum life cycle was maintained as described previously (5). Male Syrian hamsters of the Lak:LVG(SYR)BR outbred strain (Charles River Laboratories, Charles River, MA) were infected with A. ceylanicum L3 by oral gavage. Adult worms were harvested manually from the intestinal mucosa approximately 21 days postinfection. Animals were fed ad libitum a specially formulated chow (Harlan Teklad, Madison, WI) varying only in iron content (formulation of diet available upon request).
Measurement of blood hemoglobin and serum iron concentration.
Blood was collected from the orbital plexus of hamsters into heparinized capillary tubes (Fisher Scientific, Pittsburgh, PA) and assayed within 4 h of collection (5). Hemoglobin was measured using a total hemoglobin assay kit (Sigma Diagnostics, St. Louis, MO) by following the manufacturer's protocol with the following modifications: 8 μl whole blood was mixed into 2 ml Drabkin's solution (prepared as directed using reagents provided in the kit) in glass test tubes, and the samples were incubated for 15 min at room temperature. Following incubation, sample tubes were vortexed and 200 μl was transferred to duplicate wells of a 96-well microtiter plate. The optical density of each sample was measured at 530 nm using a microplate reader (Molecular Devices, Sunnyvale, CA). Sample values were determined using a hemoglobin standard curve prepared from reagents provided in the kit.
For measurement of serum iron concentration, 10 μl of hamster serum was added to 125 μl citric acid buffer in individual wells of a 96-well plate. The samples were then incubated for 5 min at room temperature. The initial absorbance was measured at 450 nm, followed by addition of 25 μl sodium ascorbate buffer (with Ferrozine). After 5 min, a second absorbance measurement was taken at 450 nm, and the difference between the two values was used to calculate serum iron levels using a standard-iron solution (Roche Molecular Systems, Inc., Alameda, CA).
Measurement of fecal egg counts.
Feces from infected animals were collected on day 23 postinfection. Pooled samples from each animal group were mixed thoroughly, and 1 g of feces was placed in a tube containing 10 ml saturated NaCl2. The sample was then vortexed for 1 min to allow complete mixing and filtered through two layers of gauze. After being mixed, 0.5 ml of the filtrate was added to each chamber of a McMaster slide (Hausser Scientific, Horsham, PA) and the total number of eggs in each chamber was determined using light microscopy. Three separate samples were analyzed in order to determine a mean egg count per gram of feces for each infected animal group.
In vivo studies of hookworm pathogenesis. (i) Effect of dietary iron restriction on hookworm pathogenesis and parasite development.
Fifteen hamsters were maintained on a standard-iron diet (200 ppm iron in the form of ferric citrate) (Harlan Teklad, Madison, WI), while fifteen were fed an iron-restricted (10-ppm) diet ad libitum. The nutrient contents of the diets were otherwise equivalent. After 21 days on either diet, 10 animals from each dietary group were infected by oral gavage with 100 A. ceylanicum L3 (5). Weight, blood hemoglobin, and serum iron measurements were obtained every 10 days, during which time animals were continued on their preinfection diets. Five animals from each dietary group were sacrificed on day 10, and the other five were sacrificed on day 20 postinfection; the intestinal worm burden was recorded for each animal (5).
(ii) Intermediate-dietary-iron restriction and hookworm pathogenesis.
Four groups of 10 hamsters were placed on diets containing 10 ppm, 40 ppm, 100 ppm, or 200 ppm iron. After 21 days, five animals from each dietary group were infected with 100 Ancylostoma ceylanicum L3 by oral gavage. Fecal egg counts were measured on day 23 postinfection as described above.
Statistical methods.
All results are presented as means ± standard errors. Comparisons of measurements between groups of animals were carried out using Student's t test. Alternatively, if F testing indicated unequal variances between groups, a Welch t test was employed. A Mann-Whitney U test was used to calculate the significance of worm burden data, and analysis of variance with Tukey-Kramer multiple comparisons was used to evaluate fecal egg counts. A log-rank test was used to calculate significance for survival data. In each case, P values of less than 0.05 were considered statistically significant.
RESULTS
Host iron is required for hookworm development and pathogenesis.
In order to characterize the effect of host iron status on hookworm pathogenesis, an initial experiment was conducted in which weanling hamsters were maintained on diets containing either 200 ppm (standard) or 10 ppm (restricted) iron. Animals from each dietary group were infected with A. ceylanicum L3 and followed for evidence of anemia and growth delay compared to the blood hemoglobin level and growth of uninfected controls. As shown in Fig. 1A, the weights of uninfected animals on the 200-ppm diet increased steadily throughout the study period, reaching a mean body weight of 114 ± 5 g at day 20 postinfection. Infected animals maintained on this standard diet exhibited growth delay beginning at approximately day 10 postinfection (5), reaching a mean weight of 88 ± 5 g, a difference that was statistically significant (P = 0.017). In contrast, there was no significant difference in day 20 postinfection weights between uninfected (90 ± 2 g) and infected (94 ± 8 g) animals maintained on the iron-restricted diet (P = 0.6).
FIG. 1.

Dietary iron restriction reduces clinical sequelae of hookworm infection. The effects of primary infection with A. ceylanicum on weight (A), blood hemoglobin (B), and serum iron levels (C) were determined in groups of hamsters maintained on a standard-iron (200 ppm) (left panels) or restricted-iron (10 ppm) (right panels) diet. * indicates statistical significance (P < 0.05). See Results for numerical values.
Blood hemoglobin levels in infected animals maintained on the 200-ppm-iron diet decreased between day 10 and day 20 postinfection, falling from a mean of 16.7 ± 0.5 g/dl to a mean of 11.1 ± 0.8 g/dl (P was 0.0002 versus values for uninfected controls) (Fig. 1B). This drop in blood hemoglobin correlates with the transition of A. ceylanicum to the adult blood-feeding stage, which occurs at days 10 to 14 postinfection (5, 16). In contrast, at day 20 postinfection there was no statistically significant difference in blood hemoglobin levels between infected (10.2 ± 1.2 g/dl) and uninfected (11.8 ± 0.5 g/dl) animals maintained on the iron-restricted diet (P = 0.3).
As shown in Fig. 1C, infected animals maintained on the 200-ppm-iron diet showed a decline in serum iron levels during the course of the study, although the difference in serum iron concentration between infected (299 ± 62 μg/dl) and uninfected (489 ± 31 μg/dl) animals measured on day 20 postinfection did not reach statistical significance (P = 0.06). By contrast, serum iron concentrations in hamsters maintained on the 10-ppm diet were very low (<50 μg/dl) at all time points during the study, with no statistically significant difference between infected and uninfected animals.
Intestinal worm burdens were measured on day 10 and day 20 postinfection for all dietary groups (Fig. 2). At day 10 postinfection, the mean worm burden in the iron-restricted animals was 0.8 ± 0.4, compared to a mean of 24.8 ± 13.8 worms harvested from the 200-ppm-iron diet group (P = 0.032). A statistically significant difference in worm burden was also observed at day 20 postinfection. At this time point, animals maintained on the standard diet yielded a mean of 28.0 ± 5.7 worms, while those on the iron-restricted diet harbored a mean of 3.6 ± 1.2 worms (P = 0.008). Of note, this study was repeated two additional times with similar results, thus confirming the reproducibility of these observations.
FIG. 2.
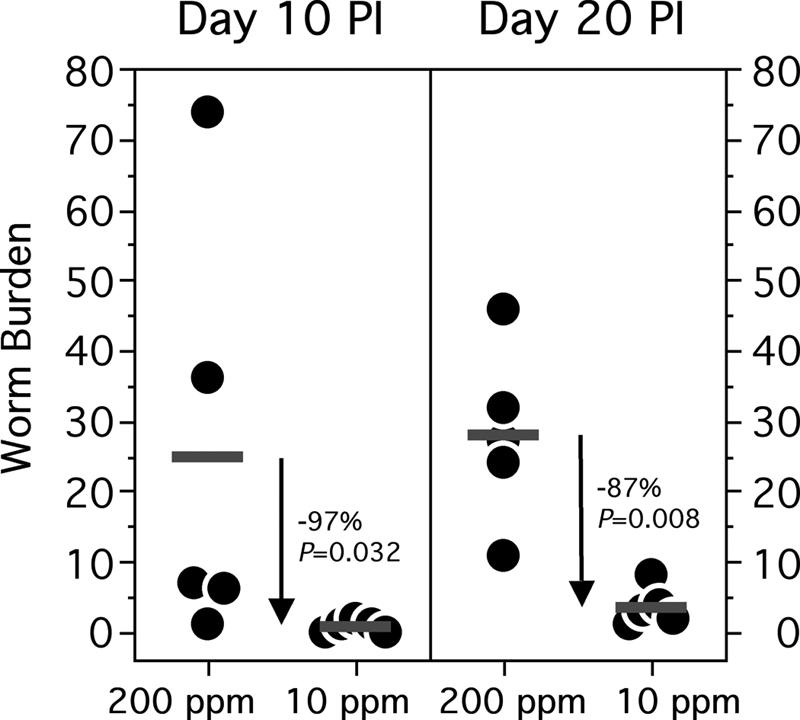
Dietary iron restriction reduces intestinal hookworm burden. Individual worm burdens are indicated by filled circles and means of each group by horizontal bars. Arrows indicate statistically significant reductions. PI, postinfection.
Moderate restriction of dietary iron exacerbates hookworm disease in vivo.
In order to define a threshold level of dietary iron that might mediate host susceptibility to hookworm infection, groups of five hamsters were equilibrated on diets containing various levels of iron (10 ppm, 40 ppm, 100 ppm, and 200 ppm) prior to infection with A. ceylanicum L3. Data on weight and blood hemoglobin concentration, measured at day 22 postinfection, are shown in Fig. 3. As previously observed (Fig. 1A), there was no statistically significant difference in weight between uninfected and infected animals maintained on the iron-restricted diet (10 ppm) over the course of the study (P = 0.8) (Fig. 3A). By comparison, at day 22 postinfection there was a difference in the average percentages of weight gained between infected (8.9 ± 17.3%) and uninfected (29.3 ± 6.5%) animals maintained on a diet containing 40 ppm iron, although substantial variation within the infected group may have contributed to this difference not being statistically significant (P = 0.3). In contrast, however, infected animals maintained on the 100-ppm diet lost 9.6% ± 6.3% of their preinfection body weight, while the weight of uninfected animals in this diet group increased by 29.9% ± 4.9%. This difference was highly significant (P = 0.001). In this experiment, there was no significant difference in percentages of weight gain observed between infected and uninfected animals maintained on the standard-iron (200-ppm) diet (P = 0.35).
FIG. 3.

Intermediate dietary iron restriction increases the pathogenicity of primary hookworm infection. Animals were equilibrated on diets containing intermediate levels of iron (10, 40, 100, or 200 ppm) prior to infection with A. ceylanicum. Shown are data from measurements of percentages of the change in weight relative to weight on day 0 (A), blood hemoglobin concentration (B), and serum iron concentration (C) taken on day 22 postinfection. P values for statistically significant differences are shown above brackets.
Measurement of blood hemoglobin concentrations demonstrated statistically significant differences between infected (10.1 ± 2.1 g/dl) and uninfected (18.0 ± 0.2 g/dl) animals maintained on diets containing 40 ppm iron at day 22 postinfection (P = 0.021). The difference between infected (12.3 ± 1.5 g/dl) and uninfected (18.0 ± 0.5 g/dl) animals maintained on the 100-ppm-iron diet was also statistically significant (P = 0.021). The difference in blood hemoglobin concentrations at day 22 postinfection between infected (16. 1 ± 0.3 g/dl) and uninfected (19.0 ± 0.9 g/dl) animals receiving the 200-ppm-iron diet was less striking, but also reached statistical significance (P = 0.026) (Fig. 3B). As was noted in the prior experiment, there was no difference in blood hemoglobin levels between infected (11.0 ± 0.7 mg/dl) and uninfected (12.1 ± 1.0 mg/dl) animals maintained on the iron-restricted (10-ppm) diet (P = 0.4).
Statistically significant differences in serum iron concentrations were noted between infected and uninfected animals maintained on either the 40-ppm or 100-ppm-iron diet, as measured on day 22 postinfection (Fig. 3C). Serum iron levels in the infected 40-ppm-iron diet group averaged 114 ± 44 μg/dl, compared to 400 ± 22 μg/dl in the uninfected group (P < 0.001). Likewise, the mean serum iron concentrations of infected animals maintained on the 100-ppm-iron diet was 51 ± 10 μg/dl by day 22 postinfection, compared to a mean of 280 ± 19 μg/dl in the uninfected group (P < 0.001). As demonstrated in the previous experiment, there was no statistically significant difference in serum iron concentrations between the infected and uninfected animals maintained on the iron-restricted diet (10 ppm) at day 22 postinfection (120 ± 32 μg/dl versus 90 ± 21 μg/dl). Infected animals on the 200-ppm-iron diet had somewhat higher mean serum iron concentrations (325 ± 24 μg/dl) than uninfected animals (254 ± 20 μg/dl) at day 22 postinfection, although this difference was not statistically significant (P = 0.056).
Consistent with the greater pathology as measured by blood hemoglobin levels and weight, animals maintained on the intermediate-iron diets (40 ppm and 100 ppm) exhibited a significant increase in fecal egg counts performed on day 23 postinfection. (Fig. 4). Mean counts of 0, 731 ± 40, 865 ± 67, and 228 ± 22 eggs per gram were obtained from animals in the 10-ppm-, 40-ppm-, 100-ppm-, and 200-ppm-iron diet groups, respectively. Animals maintained on the intermediate-dietary-iron regimens also exhibited significantly greater mortality following infection than animals on the iron-restricted (10 ppm) and standard-iron (200 ppm) diets (Fig. 5). By day 35 postinfection, three out of five animals from the 100-ppm diet group and one out of five animals from the 40-ppm diet group had died, presumably as a result of infection. By day 60 postinfection, three out of five animals in the 40-ppm group had died, while four out of five animals in the 100-ppm group had died. In contrast, none of the animals in the iron-restricted (10-ppm) or standard-iron (200-ppm) diet groups died during the course of the study. These differences between groups with regard to mortality was found to be statistically significant (P = 0.008). None of the uninfected animals from any of the dietary groups (10, 40, 100, or 200 ppm iron) died during the course of the observation period.
FIG. 4.

Intermediate dietary iron restriction is associated with increased fecal egg excretion. Feces from infected animals was collected on day 23 postinfection, and hookworm eggs were quantified as described in Materials and Methods. Bars represent the mean numbers of eggs/gram counted in three separate samples of feces. Brackets indicate statistically significant differences between groups.
FIG. 5.

Intermediate dietary iron restriction is associated with increased mortality following hookworm infection. A survival curve is shown for animals maintained on intermediate (10-, 40-, 100-, 200-ppm) iron diets. Statistical significance of survival differences was analyzed using a log-rank test, with an overall significance level of 0.008. Pairwise comparisons also revealed significant differences between both groups B (40 ppm) and C (100 ppm) and group A (10 ppm), as well as between groups B and C and group D (200 ppm).
DISCUSSION
The studies described here were designed to characterize the role of host iron status in mediating hookworm pathogenesis in vivo. First, it was demonstrated that severe dietary iron restriction leads to a significant reduction in intestinal worm burden following infection with A. ceylanicum. This attrition occurs at a point in the hookworm life cycle that predates blood feeding, suggestive of an essential role for iron in parasite development from the infectious larval stage (L3) to the adult stage. A second study demonstrated a statistically significant correlation between host dietary iron content and objective measures of hookworm pathogenicity, including weight, blood hemoglobin levels, fecal egg counts, and host survival using an animal model of A. ceylanicum infection. To our knowledge, these are the first in vivo studies to define a potential role for a specific dietary micronutrient, in this case iron, in either hookworm development or host susceptibility to infection. It is worthwhile noting that these studies were carried out using the oral route of hookworm infection, and it is unknown whether results might differ in animals infected percutaneously.
The capacity of adult hookworms to cause significant losses of blood and iron during feeding has been well established using in vivo models (4, 5, 21). Hookworm infection has long been associated with iron deficiency anemia, and data from human studies generally confirm an inverse correlation between intensity of infection, as estimated by fecal egg counts, and blood hemoglobin levels (9, 22, 33-35). It has generally been presumed that individuals who are iron deficient are likely more susceptible to hookworm anemia, due to reduced total body iron stores in the face of iron loss from gastrointestinal hemorrhage. However, it has also been suggested that hookworm infection modulates iron metabolism in the host, resulting in enhanced reabsorption from the gut as a means of compensating for hookworm-associated blood loss (10, 11). Such a compensatory mechanism was originally put forth to explain the fact that death from overwhelming hookworm anemia is rare, despite calculations that show that iron losses from hookworm infection are likely to far exceed dietary intake in many communities of high endemicity (11, 24).
The data presented here offer another potential explanation for this apparent contradiction, namely, that severe iron deficiency may directly influence the ability of hookworms to establish and/or maintain infection. Our findings are supportive of the recent observations that Caenorhabdis elegans and related helminths, including hookworms, lack the ability to synthesize heme and thus depend on exogenously acquired heme as an iron source (26). It is plausible that animals fed a low-iron diet reveal diminished hookworm pathogenicity due to inadequate heme availability for hookworm growth and development. Moreover, the effect of host iron restriction on parasite burden was evident as early as day 10 postinfection, at which time significantly fewer worms were recovered from animals in the iron-restricted group than from infected animals maintained on the 200-ppm-iron diet (Fig. 2). This unexpected finding raises the possibility that in the setting of severe dietary iron restriction, hookworms are incapable of establishing infection in the intestine.
Having established that host dietary iron is necessary for establishment of hookworm infection, a subsequent experiment was carried out in order to define a threshold of iron necessary for parasite survival and host susceptibility. Intermediate levels of dietary iron appeared sufficient to support the establishment of a patent infection, with animals maintained on the 40-ppm- or 100-ppm-iron diets demonstrating more severe anemia and growth delay than those animals fed a severely iron-restricted diet (Fig. 3). The animals maintained on the intermediate iron diets also demonstrated a significant reduction in survival, which correlated with intensity of infection as measured by fecal egg counts (Fig. 4 and 5). In contrast, the animals maintained on the 200-ppm diet exhibited evidence of both less-severe disease and a lower intensity of infection (as measured by fecal egg counts). These data confirm that moderate dietary iron restriction is, in fact, associated with enhanced susceptibility to disease, which may explain similar effects noted in prior studies of nutritional anemia and hookworm infection (11, 14). This finding is also consistent with evidence suggesting an important role for host nutritional status in resistance to other nematode infections (18, 29, 31). The mechanism through which moderate dietary iron restriction might directly impair innate and acquired immune responses to hookworm infection is currently being investigated.
These observations on the role of host dietary iron on hookworm pathogenesis may ultimately have an impact on how control strategies in areas of endemicity are implemented and evaluated. In particular, the degree to which dietary iron supplementation may directly impact hookworm development on the one hand and host susceptibility on the other needs to be more carefully evaluated in laboratory- and field-based studies. Ultimately, by making severely iron-deficient individuals only marginally less so, iron supplementation may enhance the potential for hookworm larvae to successfully establish infection in the intestine. Conversely, supplementation may improve the iron status of some individuals so that they are no longer susceptible to the most severe form of the disease. In other words, the effect of iron supplementation should no longer be viewed solely from the perspective of the host, but also as a potential effector of hookworm development and/or virulence.
These studies illuminate what has likely evolved to be a complex relationship between the hookworm and its mammalian host centering on the availability of a micronutrient essential to both, namely, iron. Work is under way to define the role of iron (and iron-containing compounds) in parasite development, as well as the molecular and immunologic mechanism(s) through which moderate dietary iron restriction impairs host defenses against hookworm. Because of the substantial overlap in the global prevalence of hookworm infection and iron deficiency, a characterization of the nature of the relationship between host and parasite, specifically as it relates to the utilization of iron, may lead to the identification of new targets for hookworm drug and vaccine development.
Acknowledgments
This work was supported by grants A107210 and A147929 from the National Institutes of Health and a Clinical Research Grant from the March of Dimes Birth Defects Foundation.
We thank Cindy Roy, Nancy Andrews, Leonard Milstone, Brian Adams, and Barbara Mickelson for helpful comments and technical assistance.
Editor: W. A. Petri, Jr.
REFERENCES
- 1.Adams, E. J., L. S. Stephenson, M. C. Latham, and S. N. Kinoti. 1994. Physical activity and growth of Kenyan school children with hookworm, Trichuris trichiura and Ascaris lumbricoides infections are improved after treatment with albendazole. J. Nutr. 124:1199-1206. [DOI] [PubMed] [Google Scholar]
- 2.Atukorala, T. M., L. D. de Silva, W. H. Dechering, T. S. Dassenaeike, and R. S. Perera. 1994. Evaluation of effectiveness of iron-folate supplementation and anthelminthic therapy against anemia in pregnancy—a study in the plantation sector of Sri Lanka. Am. J. Clin. Nutr. 60:286-292. [DOI] [PubMed] [Google Scholar]
- 3.Bungiro, R., and M. Cappello. 2004. Hookworm infection: new developments and prospects for control. Curr. Opin. Infect. Dis. 17:421-426. [DOI] [PubMed] [Google Scholar]
- 4.Bungiro, R. D., Jr., B. R. Anderson, and M. Cappello. 2003. Oral transfer of adult Ancylostoma ceylanicum hookworms into permissive and nonpermissive host species. Infect. Immun. 71:1880-1886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bungiro, R. D., Jr., J. Greene, E. Kruglov, and M. Cappello. 2001. Mitigation of hookworm disease by immunization with soluble extracts of Ancylostoma ceylanicum. J. Infect. Dis. 183:1380-1387. [DOI] [PubMed] [Google Scholar]
- 6.Bungiro, R. D., Jr., C. V. Solis, L. M. Harrison, and M. Cappello. 2004. Purification and molecular cloning of and immunization with Ancylostoma ceylanicum excretory-secretory protein 2, an immunoreactive protein produced by adult hookworms. Infect. Immun. 72:2203-2213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chu, D., R. D. Bungiro, M. Ibanez, L. M. Harrison, E. Campodonico, B. F. Jones, J. Mieszczanek, P. Kuzmic, and M. Cappello. 2004. Molecular characterization of Ancylostoma ceylanicum Kunitz-type serine protease inhibitor: evidence for a role in hookworm-associated growth delay. Infect. Immun. 72:2214-2221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Crompton, D. W. 1986. Nutritional aspects of infection. Trans. R. Soc. Trop. Med. Hyg. 80:697-705. [DOI] [PubMed] [Google Scholar]
- 9.Crompton, D. W. 2000. The public health importance of hookworm disease. Parasitology 121(Suppl.):S39-S50. [DOI] [PubMed] [Google Scholar]
- 10.Crompton, D. W., and M. C. Nesheim. 2002. Nutritional impact of intestinal helminthiasis during the human life cycle. Annu. Rev. Nutr. 22:35-59. [DOI] [PubMed] [Google Scholar]
- 11.Crompton, D. W., and R. R. Whitehead. 1993. Hookworm infections and human iron metabolism. Parasitology 107(Suppl.):S137-S145. [DOI] [PubMed] [Google Scholar]
- 12.Dossa, R. A., E. A. Ategbo, F. L. de Koning, J. M. van Raaij, and J. G. Hautvast. 2001. Impact of iron supplementation and deworming on growth performance in preschool Beninese children. Eur. J. Clin. Nutr. 55:223-228. [DOI] [PubMed] [Google Scholar]
- 13.Dreyfuss, M. L., R. J. Stoltzfus, J. B. Shrestha, E. K. Pradhan, S. C. LeClerq, S. K. Khatry, S. R. Shrestha, J. Katz, M. Albonico, and K. P. West, Jr. 2000. Hookworms, malaria and vitamin A deficiency contribute to anemia and iron deficiency among pregnant women in the plains of Nepal. J. Nutr. 130:2527-2536. [DOI] [PubMed] [Google Scholar]
- 14.Foster, A. O., and W. W. Cort. 1935. Further studies on the effect of a generally deficient diet upon the resistance of dogs to hookworm infestation. Am. J. Hyg. 21:302-318. [Google Scholar]
- 15.Friis, H., D. Mwaniki, B. Omondi, E. Muniu, F. Thiong'o, J. Ouma, P. Magnussen, P. W. Geissler, and K. F. Michaelsen. 2003. Effects on haemoglobin of multi-micronutrient supplementation and multi-helminth chemotherapy: a randomized, controlled trial in Kenyan school children. Eur. J. Clin. Nutr. 57:573-579. [DOI] [PubMed] [Google Scholar]
- 16.Garside, P., and J. M. Behnke. 1989. Ancylostoma ceylanicum in the hamster: observations on the host-parasite relationship during primary infection. Parasitology 98:283-289. [DOI] [PubMed] [Google Scholar]
- 17.Garside, P., J. M. Behnke, and R. A. Rose. 1989. The immune response of male DSN hamsters to a primary infection with Ancylostoma ceylanicum. J. Helminthol. 63:251-260. [DOI] [PubMed] [Google Scholar]
- 18.Ing, R., Z. Su, M. E. Scott, and K. G. Koski. 2000. Suppressed T helper 2 immunity and prolonged survival of a nematode parasite in protein-malnourished mice. Proc. Natl. Acad. Sci. USA 97:7078-7083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Jones, B. F., and M. Cappello. 2004. Hookworm infection: molecular mechanisms of disease and targets for control. Drug Discov. Today 1:217-222. [Google Scholar]
- 20.Kalkofen, U. P. 1970. Attachment and feeding behavior of Ancylostoma caninum. Z. Parasitenkd. 33:339-354. [DOI] [PubMed] [Google Scholar]
- 21.Kalkofen, U. P. 1974. Intestinal trauma resulting from feeding activities of Ancylostoma caninum. Am. J. Trop. Med. Hyg. 23:1046-1053. [DOI] [PubMed] [Google Scholar]
- 22.Latham, M. C., L. S. Stephenson, A. Hall, J. C. Wolgemuth, T. C. Elliot, and D. W. Crompton. 1983. Parasitic infections, anaemia and nutritional status: a study of their interrelationships and the effect of prophylaxis and treatment on workers in Kwale District, Kenya. Trans. R. Soc. Trop. Med. Hyg. 77:41-48. [DOI] [PubMed] [Google Scholar]
- 23.Montresor, A., R. J. Stoltzfus, M. Albonico, J. M. Tielsch, A. L. Rice, H. M. Chwaya, and L. Savioli. 2002. Is the exclusion of children under 24 months from anthelmintic treatment justifiable? Trans. R. Soc. Trop. Med. Hyg. 96:197-199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Pawlowski, Z. S. 1995. Anthelmintic therapy and iron supplementation of pregnant women. Am. J. Clin. Nutr. 62:1023-1024. [DOI] [PubMed] [Google Scholar]
- 25.Pena-Rosas, J. P., M. C. Nesheim, M. N. Garcia-Casal, D. W. Crompton, D. Sanjur, F. E. Viteri, E. A. Frongillo, and P. Lorenzana. 2004. Intermittent iron supplementation regimens are able to maintain safe maternal hemoglobin concentrations during pregnancy in Venezuela. J. Nutr. 134:1099-1104. [DOI] [PubMed] [Google Scholar]
- 26.Rao, A. U., L. K. Carta, E. Lesuisse, and I. Hamza. 2005. Lack of heme synthesis in a free-living eukaryote. Proc. Natl. Acad. Sci. USA 102:4270-4275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Santiso, R. 1997. Effects of chronic parasitosis on women's health. Int. J. Gynaecol. Obstet. 58:129-136. [DOI] [PubMed] [Google Scholar]
- 28.Savioli, L., and M. Albonico. 2004. Soil-transmitted helminthiasis. Nat. Rev. Microbiol. 2:618-619. [DOI] [PubMed] [Google Scholar]
- 29.Scott, M. E., and K. G. Koski. 2000. Zinc deficiency impairs immune responses against parasitic nematode infections at intestinal and systemic sites. J. Nutr. 130:1412S-1420S. [DOI] [PubMed] [Google Scholar]
- 30.Stephenson, L. S. 2001. Optimising the benefits of anthelmintic treatment in children. Paediatr. Drugs 3:495-508. [DOI] [PubMed] [Google Scholar]
- 31.Stephenson, L. S., M. C. Latham, and E. A. Ottesen. 2000. Malnutrition and parasitic helminth infections. Parasitology 121(Suppl.):S23-S38. [DOI] [PubMed] [Google Scholar]
- 32.Stoltzfus, R. J. 2001. Iron-deficiency anemia: reexamining the nature and magnitude of the public health problem. Summary: implications for research and programs. J. Nutr. 131:697S-701S. [DOI] [PubMed] [Google Scholar]
- 33.Stoltzfus, R. J., H. M. Chway, A. Montresor, J. M. Tielsch, J. K. Jape, M. Albonico, and L. Savioli. 2004. Low dose daily iron supplementation improves iron status and appetite but not anemia, whereas quarterly anthelminthic treatment improves growth, appetite and anemia in Zanzibari preschool children. J. Nutr. 134:348-356. [DOI] [PubMed] [Google Scholar]
- 34.Stoltzfus, R. J., H. M. Chwaya, A. Montresor, M. Albonico, L. Savioli, and J. M. Tielsch. 2000. Malaria, hookworms and recent fever are related to anemia and iron status indicators in 0- to 5-y old Zanzibari children and these relationships change with age. J. Nutr. 130:1724-1733. [DOI] [PubMed] [Google Scholar]
- 35.Stoltzfus, R. J., H. M. Chwaya, J. M. Tielsch, K. J. Schulze, M. Albonico, and L. Savioli. 1997. Epidemiology of iron deficiency anemia in Zanzibari schoolchildren: the importance of hookworms. Am. J. Clin. Nutr. 65:153-159. [DOI] [PubMed] [Google Scholar]
- 36.Stoltzfus, R. J., J. D. Kvalsvig, H. M. Chwaya, A. Montresor, M. Albonico, J. M. Tielsch, L. Savioli, and E. Pollitt. 2001. Effects of iron supplementation and anthelmintic treatment on motor and language development of preschool children in Zanzibar: double blind, placebo controlled study. BMJ 323:1389-1393. [DOI] [PMC free article] [PubMed] [Google Scholar]