

Abstract
The up-regulation of various matrix metalloproteinases (MMP), certain cell receptors such as integrins and CD44, and the SIBLING family of integrin-binding glycophosphoproteins have been reported separately and in various combinations for many types of tumors. The mechanisms by which these different proteins may be interacting and enhancing a cancer cell’s ability to survive and metastasize has become an interesting issue in cancer biology. Dentin matrix protein 1 (DMP1) has been known for a number of years to bind to CD44 and RGD-dependent integrins. This SIBLING was recently shown to be able to specifically bind and activate proMMP-9 and to make MMP-9 much less sensitive to inhibition by tissue inhibitors of metalloproteinases (TIMP) and synthetic inhibitors. In this study we used a modified Boyden chamber assay to show that DMP1 enhanced the invasiveness of the MMP-9 expressing colon cancer cell line, SW480, through Matrigel™ in a dose-dependant manner. 100 nM DMP1 increased invasion 4-fold over controls (86.1 ± 13.9 vs. 22.3 ± 9.8, P<0.001). The enhanced invasive potential required the presence of MMP-9 and at least one of the cell surface receptors, CD44, αvβ3, or αvβ5 integrin. The bridging of MMP-9 to the cell surface receptors was shown by both pull-down and fluorescence activated cell sorter (FACS) experiments. Because all of these proteins were also shown by immunohistochemistry to be expressed in serial sections of a colon adenocarcinoma, we have hypothesized that the MMP-9/DMP1/cell surface complexes observed to enhance cell invasion in vitro may be aiding metastatic events in vivo.
INTRODUCTION
Dentin matrix protein 1 (DMP1) is an acidic phosphoglycoprotein and member of the integrin-binding SIBLING protein family (1). All of the SIBLINGs have been shown over the years to be up-regulated in many different primary tumors but curiously are often not expressed in high-passage cell lines derived from these tumors. DMP1, historically thought to be expressed in only in bones and teeth, has recently been shown to be made in some normal ductal epithelial tissues as well (2, 3). Although the precise functions of the protein have not been explicated, it has been associated with the regulation of postnatal chondrogenesis, and subsequent osteogenesis and matrix mineralization (2, 4–6). DMP1 has been shown to bind both αvβ3-integrin and CD44 cell surface molecules (7). Several studies have shown significantly elevated expression of DMP1 in a number of cancerous tissues including lung (8), breast, uterus, thyroid, and colon (9). DMP1 has been shown to bind with nM affinity and to activate latent matrix metalloproteinase-9 (proMMP-9) apparently without removal of its propeptide. It can bind active MMP-9 without changing the protease’s ability to digest natural substrates and causes the protease to remain active in the presence of normally inhibitory amounts of TIMP1 in vitro (10). Up-regulation of MMP-9 has been described for many types of cancers including colon cancer (11) and it has been used in a portal blood assay to determine colorectal metastases in liver (12). Furthermore, the increased expression of MMP-9 in primary colon tumors has been associated with liver metastases (13) and was increased in biopsy samples from patients with colon cancer (14). We have recently shown by array analysis that there is a coordinate increase in MMP-9 and DMP1 expression in lung and kidney cancer (9). The αvβ3 and αvβ5 integrins have also been associated with colon cancer metastasizing to liver in an orthotopic murine model (15). CD44 expression has been linked to transendothelial migration of colon cancer cell lines (16) and to the invasion of colorectal cancer cell lines that also showed increased MMP-9 expression (17). Another member of the SIBLING family, bone sialoprotein (BSP), promotes invasion of several osteotropic cancer cell lines by localizing MMP-2 on the cell surface through αvβ3-integrin (18). DMP1 had no effect on the invasion of the recombinant BSP-responsive cancer cell lines suggesting that MMP-2 was the rate-limiting protease for those particular cell lines in the modified Boyden chamber assay.
Matrix metalloproteinases (MMPs) are a class of zinc-dependent endopeptidases that are collectively capable of digesting all extracellular matrix components. These proteases play an important role in remodeling the ECM, and their regulation is implicated in a variety of diseases including cancer invasion and metastasis (19). The regulation of MMP activity in tumor cell invasion and metastasis involves selective expression as well as a balance of activators, inhibitors, and regulators. The first step in invasion generally requires digestion of basement membrane proteins, including type IV collagen. MMP-2, 3, 7, 9, and 10 are all able to degrade this collagen (20). MMP-2 (gelatinase A) and MMP-9 (gelatinase B) have long been known to degrade extracellular matrix macromolecules in basement membranes and other interstitial connective tissues (21). It has been shown that the localization of active MMP-9 on the cell membrane of the invasive cell through association with CD44 can be critical for tumor cell invasion and metastasis (22). Similarly, MMP-2 localization on the cancer cell surfaces through αvβ3-integrin has been reported to be important in the initiation of invasion of several cancer cells (18, 23, 24). Therefore, understanding the interaction of MMPs, cell adhesion molecules, and any modifying proteins could offer insight into cancer biology.
We hypothesized that DMP1 may play a role in cancer metastasis by bridging MMP-9 to the cell surface through αvβ3-integrin, αvβ5-integrin, and/or CD44. To test this hypothesis, both fluorescence activated cell sorting (FACS) analysis and a previously described modified Boyden chamber invasion assay were used (25) to measure the enhancement of invasion by DMP1 for the colon cancer cell line, SW480, cells known to expresses significant amounts of MMP-9 in vitro.
MATERIALS AND METHODS
Materials
The human colon cancer cell line SW480 (CCL-228) was obtained from the American Type Culture Collection (Manassas, VA). Fetal bovine serum was purchased from Equitech-Bio, Inc. (Kerrville, TX). RPMI 1640 medium, L-glutamine, 2-mercaptoethanol, sodium pyruvate, modified Eagle medium (MEM) non-essential amino acids, Hanks’ balanced salt solution (HBSS), phosphate-buffered saline (PBS), Versene (0.53 mM EDTA in PBS), and 10% zymogram gelatin gels were from Invitrogen, Inc. (Carlsbad, CA). Matrigel™ was from Collaborative Research Inc. (Bedford, MA). Transwell inserts and companion plates were purchased from BD Biosciences Discovery Labware (Bedford, MA). Calcein acetoxymethyl ester dye and the Alexa Fluor 488 protein-labeling kit were purchased from Molecular Probes, Inc. (Eugene, OR). Mouse anti-human αvβ3-integrin monoclonal antibody immobilized on immunoaffinity gel matrix (GEM1976), mouse anti-human αvβ5-integrin monoclonal antibody immobilized on immunoaffinity gel matrix (GEM1961), anti-human αvβ3-integrin monoclonal antibody (LM609, MAB1976Z), anti-human αvβ5-integrin monoclonal antibody (P1F6, MAB1961Z), recombinant αvβ3-integrin in Triton X-100 (CC1018), and recombinant αvβ5-integrin in Triton X-100 (CC1025) were obtained from Chemicon International, Inc. (Temecula, CA). Anti human CD44 monoclonal antibody (BU75) was purchased from Ancell Corporation (Bayport, MN). ProMMP-9 was from Oncogene Research Products (Boston, MA). ProbeQuant G-50 microcolumns were from Amersham Pharmacia Biotech Inc. (Piscataway, NJ). Microcon YM-30 centrifugal filter devices were from Millipore Corporation (Bedford, MA).
Cell Culture
The SW480 cells were first grown in RPMI 1640 medium supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 5 mM 2-mercaptoethanol, 2 mM sodium pyruvate, and 0.1 mM MEM non-essential amino acids in a humidified atmosphere of 5% CO2/95% air at 37°C. When the cells were about 80% confluent, they were either used in the experiments as described in appropriate sections below or subcultured with the split ratio of 1:10 for up to 20 passages.
SIBLING Production and Purification
Recombinant DMP1 with good posttranslational modifications (including glycosylation, sulfation and possibly phosphorylation) was made as described previously (7). Briefly, full-length bovine DMP1 cDNA (26) was subcloned into a high-expression, replication-deficient adenovirus type 5 with transcription under the control of the cytomegalovirus (CMV) promoter. The DMP1-KAE construct was made by in situ mutagenesis of the RGD domain into KAE while in pBluescript. The entire insert was checked for fidelity and then shuttled to the adenovirus plasmid (27). The adenoviruses were selected, purified, and expressed as previously described (28). Recombinant SIBLINGs were generated by infecting mid-passage, subconfluent normal human bone marrow stromal cells (a gift of Dr. Pamela Gehron Robey, NIDCR, NIH). Harvested serum-free medium was subjected to anion-exchange chromatography, as described (7, 28), to isolate SIBLINGs. Purity of each SIBLING was greater than 95%, as measured after sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis.
Preparation of Matrigel™ Coated Inserts
Matrigel™-coated inserts placed in 24-well plates were prepared as described previously (18). Briefly, Matrigel™ was diluted with ice-cold sterile H2O and mixed well. The solution was dispensed as 5 μg/50 μL/insert/well. The plates were left in a laminar flow tissue culture hood overnight to dry. The inserts were then rinsed with sterile Hank’s balanced salt solution (HBSS) to rehydrate films and the plates were immediately used as described below.
Modified Boyden Chamber Cell Invasion Assay
Invasiveness of each cancer cell line was measured by using a UV-opaque transwell polycarbonate membrane (diameter of 6.4 mm and pore size of 8 μm) in a modified Boyden chamber cell invasion assay. Transwell inserts precoated with Matrigel™ as described above were placed in a 24-well plate. Pre-confluent cells were removed from culture dishes with 0.53 mM EDTA in PBS, washed twice in HBSS, and resuspended in serum-free RPMI 1640 culture medium at a final density of 4 x 105 cells per mL. Quadruplicate cultures of cells were briefly pretreated in a final volume of 250 μL of serum-free medium (containing 0.1% bovine serum albumin) with either buffer or SIBLINGs in sterile 1.5-mL microcentrifuge tubes for 10 minutes and then placed in the upper compartment of a Boyden chamber. In some cases, cells were first treated for 20 minutes with blocking antibodies, or control IgGs in the tube and then placed in the upper chamber. In the latter cases, buffer or recombinant SIBLING was then added directly to the chamber. To induce migration through the Matrigel layer, the lower chambers were filled with 750 μL of serum-free medium conditioned by mouse NIH 3T3 fibroblastic cells and containing 0.1% bovine serum albumin. Cells were then incubated in humidified incubator at 37°C for 24 hours. Cells that had not migrated through the barrier were removed from the top compartment and inserts were moved to another 24-well plate in which each well contained 0.5 mL the fluorescent dye, Calcein acetoxymethyl ester, at 4 μg/mL. The plate was incubated at 37°C for 45 minutes to allow the living cells to take up and activate the dye before the fluorescent intensity was read from the bottom of the insert with a fluorescence plate reader (Wallac 1420 VICTOR2 Multilabel Reader, PerkinElmer Life Sciences Inc., Boston, MA). Fluorescence intensity was proportional to the number of cells having migrated to the bottom of the UV-opaque membrane.
Pull-Down Experiments
Commercial mouse anti-human αvβ3-integrin or αvβ5-integrin monoclonal antibody immobilized on immunoaffinity gel matrix beads were washed three times in ice-cold Triton buffer (TB = 20 mM Tris-HCl [pH 7.4], 150 mM NaCl, 0.2% Triton X-100, 2 mM MgCl2, and 0.1 mM CaCl2) and incubated in 1 mL TB containing 1% bovine serum albumin at 4°C for 30 minutes with gentle shaking. After washing three times with 1 mL of ice-cold TB, the beads were gently shaken with or without 10 μg of αvβ3-integrin or αvβ5-integrin complex in 50 μL TB at 4°C for 10 minutes. The beads were then gently pelleted, the liquid was carefully removed, and the beads were washed in 1 mL TB. The beads were then resuspended in 1 mL buffer and separated into equal volume aliquots. An aliquot was gently shaken with buffer alone or buffer containing 500 nM DMP1 or 500 nM DMP1-KAE (in a final volume of 50 μL) at 4°C for 10 minutes. The beads were then pelleted, washed in 1 mL of TB, and incubated in 50 μL of TB containing 1 μg of proMMP-9 at 4°C for 10 minutes. The beads were pelleted and washed with 1 mL TB, and the MMP-9 was eluted from the beads with 80 μL of 1X zymogram sample buffer (2.5 mL 0.5 M Tris-HCl, pH 6.8 + 2 mL of glycerol + 4 mL 10% (w/v) Sodium Dodecyl Sulfate + 0.5 mL 0.1% Bromophenol Blue adjusted to 20 mL with distilled water) and resolved by electrophoresis on a 10% (gelatin) zymogram gel.
SDS-Polyacrylamide Gel Electrophoresis and Zymography
Samples in zymogram gel sample buffer were loaded on a 10% (gelatin) zymogram gel, subjected to electrophoresis, and processed as recommended by the manufacturer. Resulting Coomassie-stained gels were visualized by dynamic integrated exposure using an EagleEye II imaging system (Stratagene Corp., La Jolla, CA).
Labeling of Purified Human MMP-9 with Alexa 488 Dye
Latent protease (proMMP-9) was fluorescently labeled with the Alexa Fluor 488 protein-labeling kit according manufacturer’s protocol but adjusted to the smaller amount of protein being labeled. All steps were performed at 4 °C. Briefly, shipping buffer from 50 μg of proMMP-9 was changed into the reaction buffer (PBS) on ProbeQuant G-50 microcolumns, and the resulting elulant concentrated to approximately 50 μL with a prewashed Microcon YM-30 centrifugal filter device. Five μL of 0.1 M sodium bicarbonate was added to raise the pH to 7.5–8.5 for efficient labeling. The reactive dye was dissolved in 0.5 mL of PBS containing 0.1 M sodium bicarbonate, 50 μL of Alexa Fluor 488 dye was added to the proMMP-9 solution, and the reaction mixture was stirred for 2 hours. The labeled proMMP-9 protein was then separated from the unreacted dye on a ProbeQuant G-50 microcolumn (in PBS) and stored as aliquots at −80°C until use.
Flow Cytometry
Cells were detached from culture dishes with PBS containing 0.53 mM EDTA, washed twice in HBSS, and then incubated at 2 x 106 cells per mL at room temperature for 10 minutes with buffer alone or buffer containing either 500 nM DMP1 or 500 nM DMP1-KAE. For the studies involving the blocking antibodies, cells were incubated with buffer alone or buffer containing αvβ3-integrin antibody (LM609, 4 μg/mL), αvβ5-integrin antibody (P1F6, 4 μg/mL), CD44 antibody (BU75, 4 μg/mL), or control IgG (4 μg/mL) at room temperature for 10 minutes, and then the mixture was incubated with 500 nM DMP1 or 500 nM DMP1-KAE for another 10 minutes. In the final step for all samples, cells were pelleted, washed once in HBSS, and then incubated at room temperature with Alexa Fluor 488-labeled human proMMP-9 at 5 μg/mL for 10 minutes. The cells were pelleted, washed once, resuspended in HBSS, and analyzed immediately with a FACSCalibur cell sorter equipped with a 488-nm argon laser and Cellquest software (BD PharMingen, Bedford, MA).
Rabbit Anti-Human MMP-9 polyclonal Antibody (LF-184) Production
The carboxy-terminal 23 aminoacids of human MMP-9 (RSELNQVDQVGYVTYDILQCPED), a region with relatively low homology to human MMP-2, was synthesized and conjugated to maleimide-activated keyhole limpet hemocyanin (#77606, Pierce Biotechnology, Inc., Rockford, IL) through the cysteinyl residue. A New Zealand white rabbit was injected four times with ~1.0 mg of conjugate each under an approved animal protocol at a contract facility (Covance Inc., Denver, PA). ELISA showed good activity to authentic human MMP-9 but no cross reactivity with authentic human MMP-2 or MMP-3. 1.6 mg of synthetic peptide was conjugated to 2 ml column of SulfoLink agarose beads (#44895, Pierce Biotechnology, Inc., Rockford, IL) according to the manufacturer’s instructions. Antibodies were bound to the affinity column, washed sequentially with PBS, PBS+T, PBS, and then eluted in 0.1 M glycine, pH 2.4. The fractions were immediately neutralized, dialyzed against PBS, and stored frozen at −80°C until use.
Immunohistochemistry
Immunohistochemistry was performed on human colon adenocarcinoma serial sections cut from blocks obtained from the Cooperative Human Tissue Network (CHTN), Mid-Atlantic Division, Charlottesville, VA under IRB approval. The Zymed ST5050 automated system (Zymed Lab Inc., San Fransisco, CA) was used with following primary antibodies: mouse anti-human DMP1 monoclonal antibody (LFMb-31, 1:100) (2), mouse anti-human CD44 (H-CAM, 1:100) monoclonal antibody (MAB2140, Chemicon International, Inc., Temecula, CA), mouse anti-human αv-integrin (CD51, 1:50) monoclonal antibody (407286), mouse anti-human β3-integrin (CD61, 1:50) monoclonal antibody (407314), mouse anti-human β5-integrin monoclonal antibody (407316, 1:50) (all from EMD Biosciences, Inc., San Diego, CA), and the affinity purified rabbit anti-human MMP-9 polyclonal antibody (LF-184, 1:200). Briefly, the paraffin block containing a moderately differentiated colon adenocarcinoma was serially sectioned (5 μm thickness) and dried on silane-coated slides (#418, Statlab Medical Products Inc., Lewisville, TX). The sections were manually deparaffinized in xylene and rehydrated through a graded ethanol series (100%, 95%, and 70%). After the endogenous peroxidase activity was blocked with 3% hydrogen peroxide in methanol for 20 minutes, the sections were washed 3 times in phosphate-buffered saline (PBS) for 5 minutes each and covered with 0.05% Tween-20 in PBS (PBS-T) before loading the slides onto the preprogrammed ST5050 automated immunohistochemistry machine. The ST5050 was programmed to incubate each slide for one hour with appropriate primary antibody diluted in 10% normal goat serum in PBS. The slides thereafter went through a 4x1 minute wash cycle with PBS-T before incubation with SuperPicTure Polymer HRP-conjugated broad-spectrum secondary antibody (Zymed Lab Inc., San Fransisco, CA) for 10 minutes. The slides were processed through another wash cycle and then were developed with AEC Single Solution chromogen (Zymed Lab Inc., San Fransisco, CA) for 2 minutes. Sections were lightly counterstained manually with Mayer’s hematoxylin for 10–20 seconds before applying an overlay of Clearmount glaze (both from Zymed Lab Inc., San Fransisco, CA) over the sections. After air-drying, slides were coverslipped with Histomount (Zymed Lab Inc., San Fransisco, CA). All steps were performed at room temperature. Negative controls included the substitution of primary antibody with non-immune rabbit serum or mouse IgG control (Zymed Lab Inc., San Fransisco, CA). Sections were photographed with an AxioCam MR-MRGrab camera imaging system (Carl Zeiss Vision, Munchen, Germany), which included an Axioplan2 microscope, an AxioCam MRm camera, and AxioVision 3.1 software.
Statistical Analysis
Data are the mean of quadruplicate determinations and 95% confidence interval (CI). Each experiment has been repeated at least 2 times. In each case, data from a single representative experiment is shown. Multiple comparisons were performed with a one-way analysis of variance followed by Dunnett's test for treatment vs. control comparisons. Pairwise comparisons were carried out by performing nonparametric Mann-Whitney U test. In each analysis, differences were considered statistically significant for P values less than 0.05. All statistical tests were two-sided.
RESULTS
DMP1 enhances invasiveness of colon cancer cell line, SW480, in vitro.
Previous work showed that many human cancer cell lines that produced MMP-2 demonstrated enhanced invasion by BSP/MMP-2 interactions but that addition of recombinant DMP1 had no such effect. Therefore, the invasiveness of SW480 colon cancer cells, which are known to express DMP1’s partner protease, MMP-9, but not MMP-2 (29) was measured using a modified Boyden-chamber invasion assay. SW480 cells showed a clear dose-response increase in their invasiveness through the Matrigel™ barrier upon the addition of recombinant DMP1, with a maximum 3.9-fold increase at 100 nM DMP1 (86.1 ± 13.9 vs. 22.3 ± 9.8 for untreated cells, P<0.001) (Fig. 1). The increase in the invasiveness of the SW480 cell line by DMP1 was not due to enhancement of integrin-mediated cell attachment/migration alone because the same amount of recombinant human BSP or OPN, two other members of the SIBLING family that can support cell attachment/migration but cannot bind and activate MMP-9, did not increase the invasiveness of the SW480 cells (data not shown).
Fig. 1.
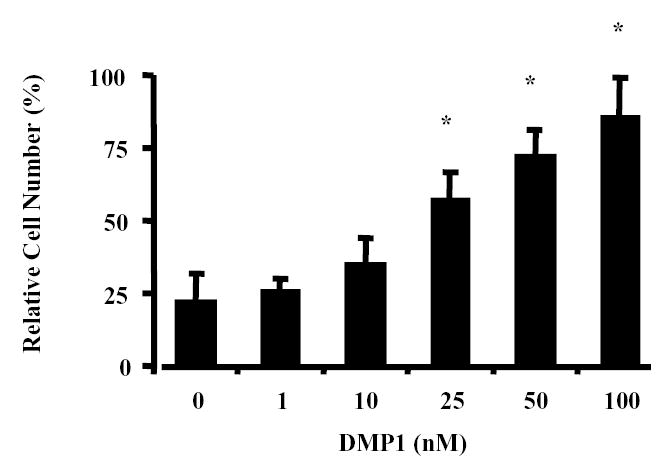
Dentin matrix protein 1 (DMP1) and the invasion of the colon cancer cell line SW480 in vitro. Untreated or DMP1-treated SW480 colon cancer cells were placed in the top wells of separate Boyden chambers that each had Matrigel™-coated UV-opaque transwell inserts. The lower chambers contained serum-free conditioned medium as a chemoattractant. The cells were incubated at 37 °C for 24 h. Invasive cells that penetrated the Matrigel™ artificial basement membrane barrier and moved into the lower chamber were then detected by Calcein acetoxymethyl ester (AM) fluorescent dye activated by the live cells. The relative fluorescence (RF) in the lower chamber corresponded directly to the number of cells that digested their way through the barrier and migrated into to the lower chamber. Data are the means of triplicate samples, and error bars are the 95% confidence intervals. A P value of <0.001 was obtained for multiple comparisons within the panel by use of one-way analysis of variance. Each treatment group was also individually compared with the control (0 nM DMP1) group by use of the Dunnett test. *P<0.001, compared with untreated cells by the Dunnett test. All statistical tests were two-sided.
DMP1-enhanced invasion requires MMP-9 and cell surface receptors αvβ3-integrin, αvβ5-integrin, or CD44.
Invasion enhancement studies were also performed with DMP1-KAE, a recombinant DMP1 protein whose integrin-binding RGD (ArgGlyAsp) sequence was replaced with the chemically similar but inactive tripeptide, KAE (LysAlaGlu). SW480 cells treated with DMP1-KAE showed a 2.4–fold increase at 100 nM DMP1-KAE (83.6± 16.3 vs. 34.4 ± 20.2 for untreated cells, P<0.01) (Fig. 2A.). While significantly less robust than the enhancement from normal DMP1, the mutant DMP1-KAE clearly aided the invasion by these cells without the ability to bind to RGD-dependent integrins. Addition of a blocking CD44 antibody completely inhibited the DMP1-KAE-enhanced invasion (12.7 ± 4.7 vs. 11.1 ± 2.9) (Fig. 2B.) showing that DMP1 can act through both RGD-dependent integrins and CD44. To complete the study of cell surface proteins, blocking antibodies to the αvβ3 and αvβ5-integrins (both RGD-dependent) as well as to CD44 were separately added to SW480 cells prior to the addition of normal DMP1 in the invasion assay. Each lowered the efficiency of the DMP1-enhanced invasion but not to DMP1-free control levels suggesting that each was contributing to a portion of the DMP1-enhancement (Fig 2C). Simultaneous addition of all three antisera reduced the DMP1-enhanced invasion to control levels (Fig. 2C) suggesting that both αvβ3 and αvβ5-integrins contributed to the integrin-related DMP1 enhancement of invasion by SW480 cells.
Fig. 2.
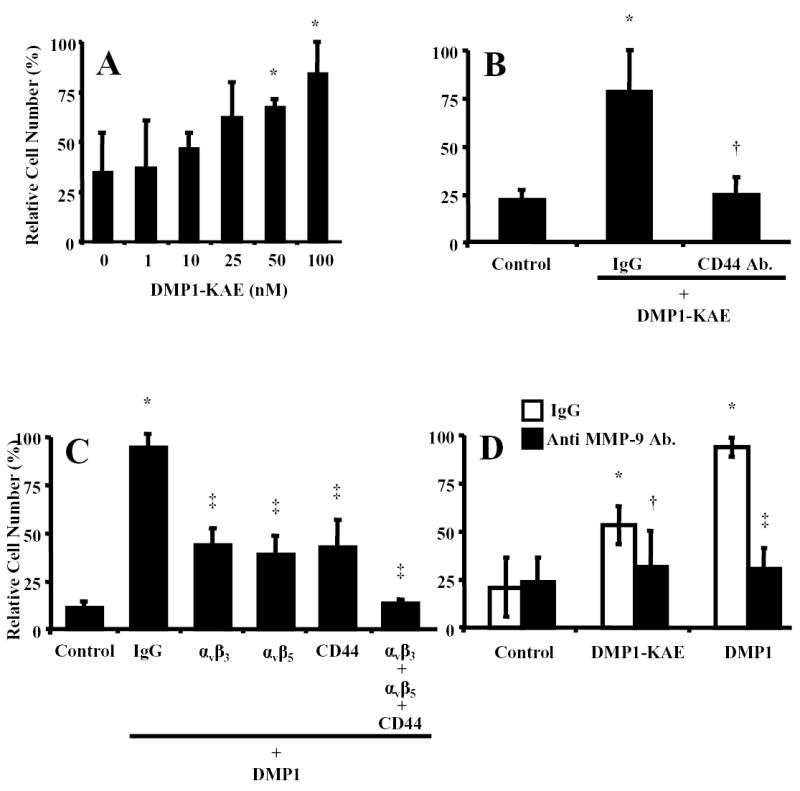
RGD-dependent integrins and CD44 are involved in the DMP1-enhanced invasion of colon cancer cells in vitro. A) SW480 cells treated with a 0–100 nM DMP1-KAE (in which the RGD domain of DMP1 was replaced with the integrin-inactive tripeptide, KAE), were placed in the upper portion of a modified Boyden chamber coated with Matrigel™. The addition of DMP1-KAE resulted in a positive but less robust dose-response pattern compared to the integrin-binding native DMP1 (Fig. 1), showing that DMP1 does not have an absolute requirement to bind to RGD-dependent integrins to enhance invasion. B) Addition of CD44 antibodies (20 μg/ml) to the DMP1-KAE treatment completely negates the enhanced invasion showing that CD44 is the alternate cell surface-binding partner for DMP1. C) Blocking monoclonal antibodies to αvβ3 or αvβ5 integrins, or CD44 (20 μg/ml each antibody) were each able to partially block DMP1-enhanced invasion but not to control levels, suggesting that DMP1 can act through each of these cell surface receptors. Combined, the antibodies completely block the DMP1-enhanced invasion. Nonspecific IgGs had no affect on the invasion enhancement. D) Inhibition of MMP-9 activity by addition of blocking antibodies (20 μg/ml) returned both the DMP1- and DMP1-KAE-enhanced invasion to control levels. Relative fluorescence (RF), which corresponds to the number of cells that migrated through the Matrigel™, is as described in Fig. 1. Panel A data are the means of quadruplicate samples, and error bars are 95% confidence intervals. A P value of <0.01 was obtained for multiple comparisons within each panel, by use of one-way analysis of variance. Each treatment group was also individually compared with the control, untreated group by use of the Dunnett test. *P<0.01, compared with untreated cells by the Dunnett test. All statistical tests were two-sided. Data for panels B–D are the means of quadruplicate samples from a representative experiment, and error bars are 95% confidence intervals. A Mann-Whitney U test was used for the pairwise comparisons. A P value of <0.01 was obtained for SW480 cells treated DMP1-KAE or DMP1 vs. untreated respective control cells (*). A P value of <0.01 was obtained for SW480 cells treated with antibody+DMP−KAE (†) or DMP1 (‡) vs. isotype control IgG+DMP−KAE or DMP1, respectively. All statistical tests were two-sided.
Since integrins, CD44, and MMP-9 have previously been shown to be associated with cancer cell invasiveness (9, 16), and because DMP1 specifically binds to MMP-9 with nanomolar affinity (10), we hypothesized that DMP1 may bridge MMP-9 to the cell surface via CD44, αvβ3 and/or αvβ5 integrins where it could become more effective in digesting the Matrigel during the invasion process. Both DMP1 and DMP1-KAE-enhanced SW480 cell invasion were almost completely inhibited by the addition of a blocking antibody to MMP-9 (Fig. 2D.).
DMP1 bridges MMP-9 to CD44, αvβ3, and αvβ5 integrins.
Because DMP1 can separately form complexes with specific cell surface proteins and with MMP-9, DMP1 may be able to bridge MMP-9 to αvβ3, or αvβ5 integrins in vitro. The hypothesis was tested with pull-down experiments using purified components. Only a small amount of proMMP-9 was detected using just the beads, showing that the pull-down assay had a low background (first lane, Fig. 3). Beads with bound αvβ3 or αvβ5 integrin but no DMP1 resulted in a similar, small amount of proMMP-9 pulled down with the beads (second lane, Fig. 3). Addition of DMP1-KAE also gave background levels of bound proMMP-9 (third lane, Fig. 3). However, addition of DMP1 to either αvβ3 or αvβ5 integrin-coated beads, greatly increased the amount of bead-associated proMMP-9, indicating the bridging of proMMP-9 to each integrin (fourth lane, Fig. 3). BSP and OPN are also known to be able to bind to αvβ3 and possibly αvβ5 integrins but substitution of recombinant BSP or OPN for DMP1 in the pull-down experiments resulted in only background levels of MMP-9, again illustrating the specificity of the DMP1 for proMMP-9 (data not shown). Because we were unable to obtain recombinant CD44, we could not test the formation of a proMMP-9/DMP1/CD44 complex in vitro using highly purified proteins in the pull-down experiments.
Fig. 3.
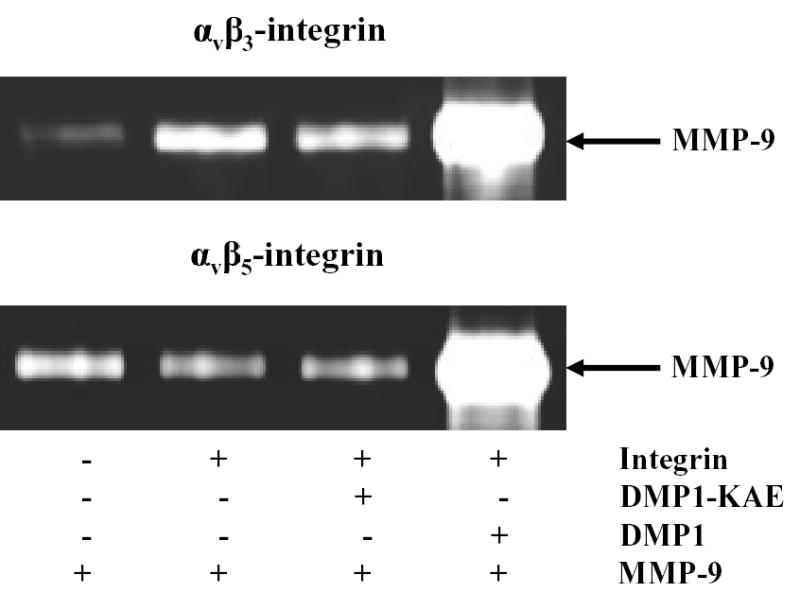
Pull-down experiments showing bridging of matrix metalloproteinase 9 (MMP-9) to αvβ3 integrin and αvβ5 integrin by dentin matrix protein 1 (DMP1). The αvβ3 or αvβ5 integrins were first bound to their respective monoclonal antibodies previously attached to beads by the manufacturer. After washing, the beads were incubated with buffer alone or buffer containing 500 nM DMP1-KAE or 500 nM DMP1, washed, and subsequently treated with recombinant proMMP-9. The washed samples were then electrophoresed on 10% zymogram gelatin gels and examined by Coomassie blue staining after digestion conditions were performed. Beads alone have low background level of MMP-9 binding (Lane 1). Note that the addition of DMP1 (Lane 4) but not DMP1-KAE (lane 3) enabled proMMP-9 to be pulled down with both sets of integrin-bound beads. Control levels of proMMP-9 were observed without addition of DMP1 (Lane 2).
We were, however, able to detect proMMP-9/DMP1/CD44 complexes as well as proMMP-9/DMP1/integrin complexes on living cells through FACS analysis. Purified proMMP-9 was covalently labeled with the fluorescent dye, AlexaFluor-488. Flow cytometry was used to determine the amount of labeled proMMP-9 bound to SW480 cells with and without DMP1 (or DMP1-KAE) treatment. Addition of DMP1 produced an 81% increase in the mean value of the signal of labeled proMMP-9 bound, compared with that of cells not pretreated with DMP1 (Fig. 4A). The addition of DMP1-KAE produced a lower but significant increase of proMMP-9 binding (57%) (Fig. 4B) verifying the earlier results showing that RGD-binding integrins were not the only cell surface protein that could be used by DMP1. Separate addition of blocking antibodies to αvβ3-integrin (Fig. 4D), αvβ5-integrin (Fig. 4E), or CD44 (Fig. 4F) before the treatment with DMP1 each partially blocked the DMP1-enhanced binding of the labeled proMMP-9 to SW480 cells. However, when cells were treated with the blocking antibodies against both integrins and CD44 together, labeled proMMP-9 binding decreased to the DMP1-free control levels (Fig. 4G). Furthermore, when cells were first treated with anti-CD44 monoclonal antibody and then with DMP1-KAE, the MMP-9 binding was reduced to the untreated control levels (Fig. 4H) supporting the hypothesis of the formation of an MMP-9/DMP1/CD44 complex on cell surfaces.
Fig. 4.
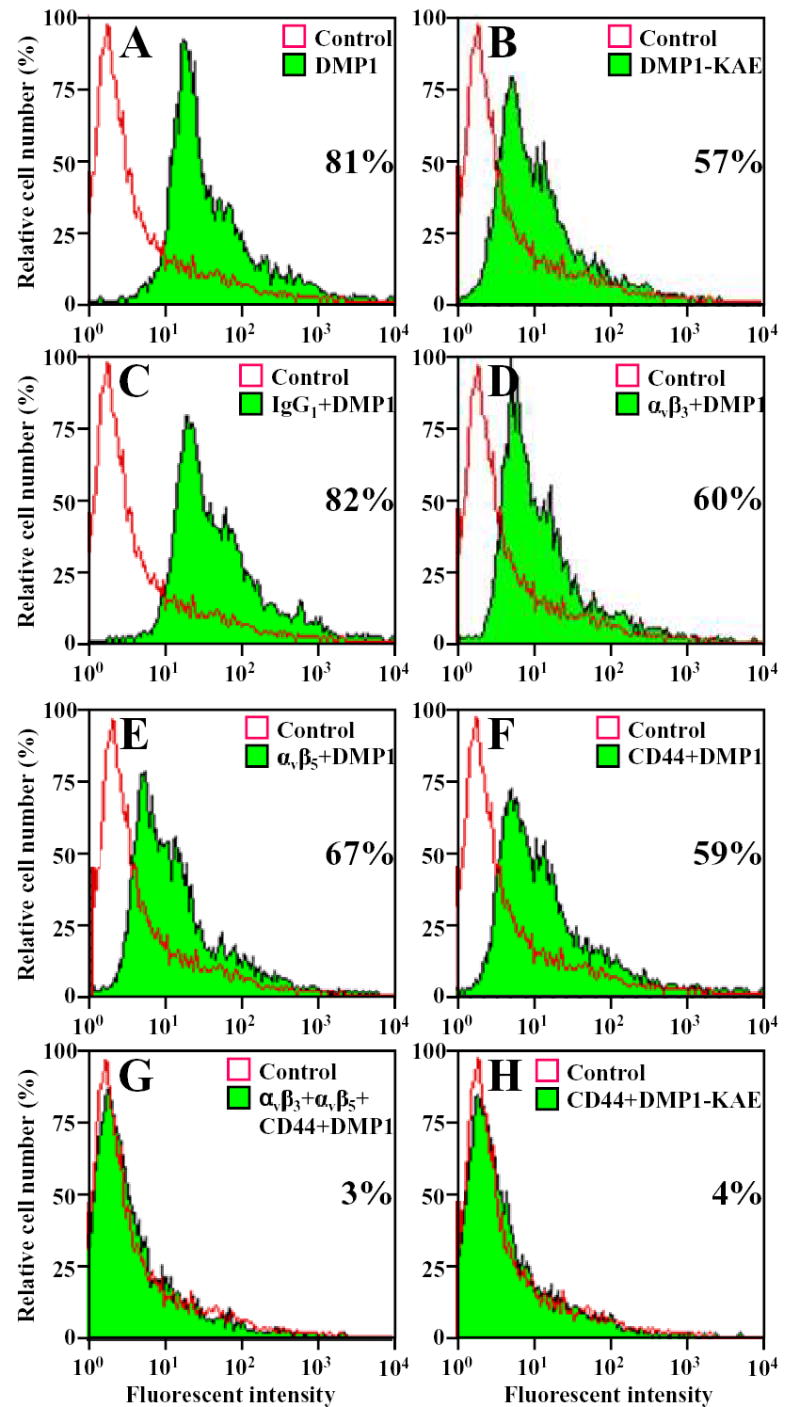
Dentin matrix protein 1 (DMP1) enhanced binding of fluorescently labeled latent matrix metalloproteinase 9 (proMMP-9) to SW480 cells. ProMMP-9 was labeled with Alexa Fluor-488 and incubated with cells treated as indicated or left untreated cells (Control). Pretreating the cells with DMP1 (shaded area) increased the amount of labeled proMMP-9 bound to the living cells, compared with untreated cells as analyzed by FACS (A). Pretreating cells with DMP1-KAE (shaded area) showed increased binding of labeled proMMP-9, compared with that of untreated cells (open area) (B). Blocking cell-surface molecules αvβ3 integrin (D), αvβ5 integrin (E), or CD44 (F) with their respective monoclonal antibodies decreased the DMP1-enhanced binding of the labeled proMMP-9, but not to control levels. However, when all three antisera were added together (αvβ3 + αvβ5 + CD44) the DMP1-enhanced binding of labeled proMMP-9 was completely blocked (G). Treating cells with a nonimmune IgG had no affect on the ability of DMP1 to enhance the binding of proMMP-9 (C). DMP1-KAE-enhanced binding of labeled proMMP-9 was essentially blocked by CD44 monoclonal antibody alone (H). Numbers represent the percent of cells bound labeled proMMP-9.
Indirect fluorescent immunocytochemistry experiments were performed to show that added DMP1 and MMP-9 both localize to the same cells. DMP1 and MMP-9 were co-localized on the SW480 cells (Fig. 5, A–C) suggesting that DMP1 rapidly localizes MMP-9 to the cell surface in vitro by binding to possibly both integrins and CD44 cell surface molecules. Added recombinant DMP1-KAE and MMP-9 were also detected at the same locations in these cells (Fig. 5, D–F) suggesting that DMP1 also localizes MMP-9 to the cell surface in vitro through CD44. Like the SIBLINGs in most other established cancer cell lines, DMP1 was not detected in untreated SW480 cells, but endogenous MMP-9 was weakly positive (data not shown).
Fig. 5.
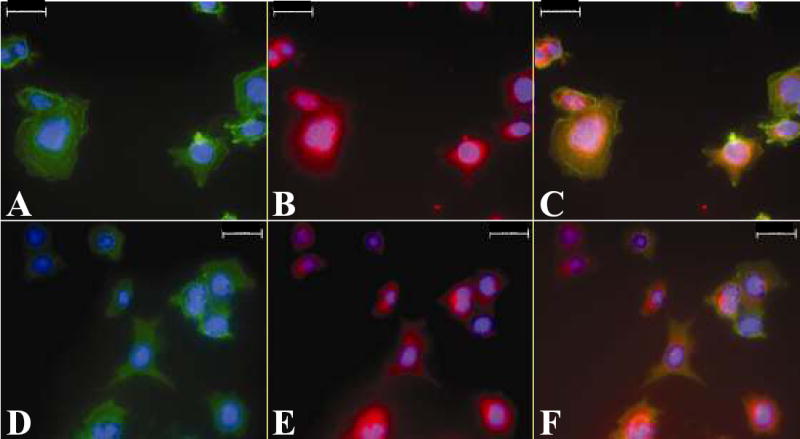
Co-localization of dentin matrix protein 1 (DMP1) and RGD-mutant DMP1 (DMP1-KAE) with matrix metalloproteinase 9 (MMP-9) on SW480 colon cancer cells. Cells were treated first with DMP1 and then with proMMP-9. The cells were incubated with 1) a mouse monoclonal antibody that can detect DMP1 as well as DMP1-KAE and 2) an affinity purified rabbit polyclonal antibody against MMP-9. Bound antibodies were then detected by indirect immunofluorescence with Cy2-conjugated AffiniPure goat anti-mouse IgG for DMP1 or DMP1-KAE, and Cy5-conjugated Affini-Pure goat anti-rabbit IgG for MMP-9. The green color in panel A shows the location of DMP1, the red color in panel B shows the location of MMP-9, and the yellow color in panel C shows that the two proteins co-localize. Similarly, the green color in panel D shows the location of DMP1-KAE, the red color in panel E shows the location of MMP-9, and the yellow color in panel F shows that the two proteins co-localize. 4, 6-Diamidino-2-phenylindole (DAPI) was used as a nuclear stain (blue color). Scale bar = 20 μm.
Expression of DMP1, MMP-9, CD44, and integrin subunits αv, β3, and β5 in colon adenocarcinoma in vivo.
The expression of MMP-9, CD44 as well as the αv, β3, and β5 integrin subunits in tumors have been shown separately and in some combinations many times previously. However, in order to illustrate that the MMP-9/DMP1/cells surface receptors are made by the same cells and therefore logically may form the complexes describe above in vivo, immunohistochemistry reactions were performed on serial paraffin sections of a human colon adenocarcinoma. The results show that all six proteins are co-expressed in the same cells/areas, suggesting that the complexes could easily be locally formed by simple interactions (Fig 6). The red/brown staining indicates the expression of DMP1 (Fig. 6A), CD44 (Fig. 6B), αv integrin subunit (Fig. 6C), β3 integrin subunit (Fig. 6D), β5 integrin subunit (Fig. 6E), and MMP-9 (Fig. 6F). The negative controls, mouse IgG (Fig. 6G), and rabbit non-immune serum (Fig. 6H), illustrate the low background for these assays.
Fig. 6.
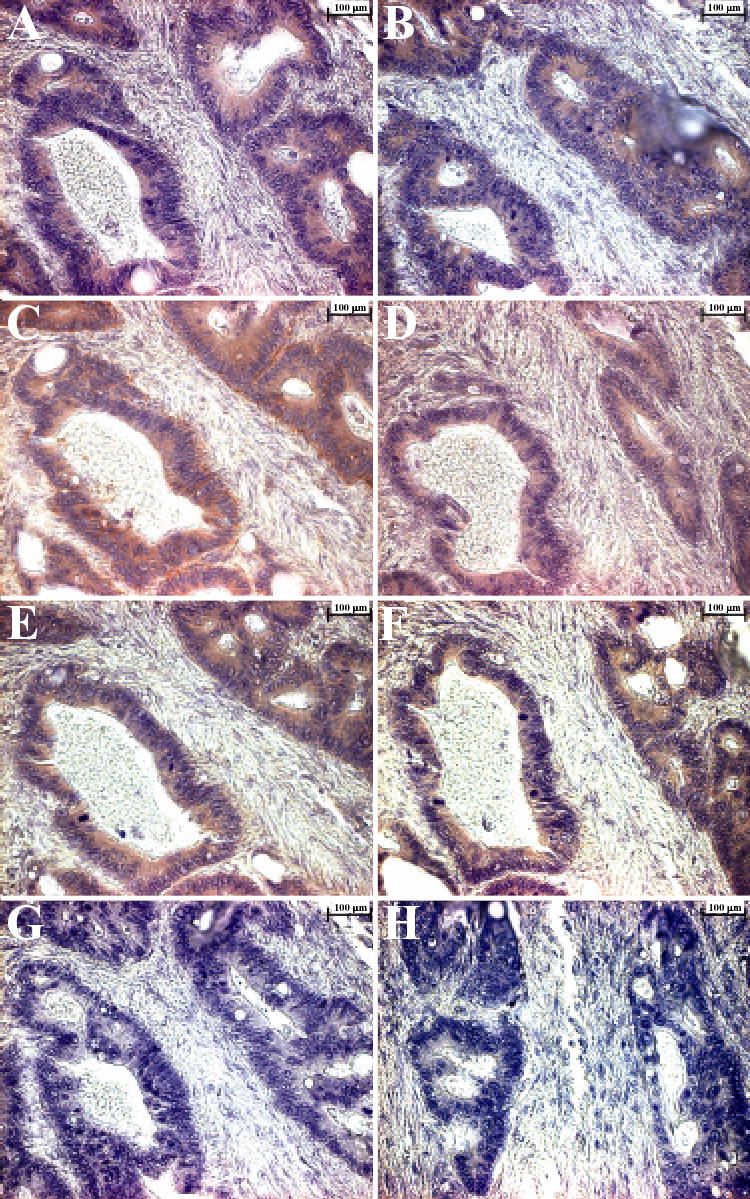
The expression of dentin matrix protein 1 (DMP1), matrix metalloproteinase 9 (MMP-9), CD44, and all three subunits of integrin αvβ3 and αvβ5 in near-serial sections of moderately differentiated colon adenocarcinoma. Deparaffinized sections of a colon adenocarcinoma were incubated separately with antibodies specific for DMP1, MMP-9, CD44, and the αv, β3, and β5 integrin chains. Localization of the antibodies was determined with SuperPicture Polymer HRP-conjugated broad-spectrum secondary antibody and AEC Single Solution chromogen. Red/brown color indicates that DMP1 (Panel A), CD44 (Panel B), αv integrin (Panel C), β3 integrin(Panel D), β5 integrin(Panel E), MMP-9 (Panel F) are all associated with the same cells, suggesting that all proteins required for formation of complexes of CD44, or integrins αvβ3 or αvβ5, DMP1, and MMP-9 are expressed in the same cells. The negative controls: mouse IgG (Panel G), and rabbit immune serum (Panel H) revealed no signal. Mayer’s hematoxylin was used as a nuclear counterstain (purple-blue). Scale bar = 100 μm.
DISCUSSION
Cancer progression reaches a critical period when a tumor starts to invade and metastasize. One of the initial steps in metastasis requires invasion of tumor cells through basement membrane and nearby stroma. The presence of MMPs, various cell adhesion molecules, and SIBLING proteins during tumor development, growth, invasion, metastasis, and angiogenesis have been well documented. However, the mechanisms by which these molecules work in the coordination of tumor progression are less well elucidated. Several co-localization studies have reported that αvβ3 integrin may function not only as an adhesion and/or migration receptor but also to activate and properly distribute proteases that degrade the extracellular matrix during invasion (23, 24, 30). Three members of the SIBLING family (BSP, OPN, and DMP1) bind and activate proMMP-2, proMMP-3, and proMMP-9, respectively (10). Furthermore, the same pairings of SIBLINGs and active MMPs cause their respective TIMPs to be much less effective at inhibiting the proteases. Recombinant BSP has been shown to promote invasion of many MMP-2 expressing osteotropic cancer cell lines through formation of a MMP-2/BSP/αvβ3-integrin complex (18). DMP1, which has recently been shown to be expressed by various tumors (8, 9), had no effect on these same MMP-2 producing cell lines.
In this paper, the role of DMP1 in an invasion assay was studied using a colon cancer cell line, SW480, that has been reported to express significant levels of MMP-9, but no MMP-2 (29). Recombinant DMP1 added to the Boyden chamber wells caused a dose-response increase in the invasiveness of the cells through a Matrigel barrier. Addition of two other SIBLING family members with similar cell attachment properties, BSP or OPN, did not increase the invasiveness of the SW480 cells suggesting that DMP1’s action was not due to simple attachment and/or migration effects. This result is similar to the specific enhancement of invasion of several MMP-2 expressing cancer cell lines in response to the addition of MMP-2’s SIBLING partner, BSP, but not to either OPN (MMP-3’s partner) or DMP1. Negating DMP1’s ability to bind to RGD-dependent integrins by mutation of the RGD to the inactive KAE or by the addition of antibodies that block αvβ3 integrin and αvβ5 integrin only partially affected DMP1’s ability to enhance invasion. This suggests that DMP1 may also be using another of its known cell surface receptors, CD44. Antibodies to CD44 resulted in a partial block in the DMP1-enhanced invasion and a complete return to control levels when used with the DMP1-KAE protein. Used together, antibodies against αvβ3-integrin, αvβ5-integrin, and CD44 reduced the DMP1-enhanced invasion to nearly control levels.
The DMP1-enhanced invasion by the SW480 cells was also inhibited by a specific antibody for it protease partner, MMP-9. Pull-down studies using purified proteins showed the formation of MMP-9/DMP1/αvβ3-integrin and MMP-9/DMP1/αvβ5-integrin complexes in vitro. Since purified recombinant CD44 was not available, we did not look at the formation of the hypothesized MMP-9/DMP1/CD44 complex by this approach. However, the formation of the DMP1/MMP-9/CD44 as well as the other two integrin-related complexes at the cellular level by FACS analysis was shown. Together, these results suggest that cells use DMP1 as a bridge to link MMP-9 to its cell surface receptors, αvβ3 integrin, αvβ5 integrin, and/or CD44 thereby increasing their ability to invade through basement membranes and other connective tissues particularly in the absence of MMP-2. The modified Boyden chamber assay can measure only certain rate-limiting steps in this model system, but one can hypothesize that in vivo, a cell may use a BSP/MMP-2/integrin complex to digest some matrix components and a DMP1/MMP-9/integrin or DMP1/MMP-9/CD44 complex to digest other proteins during the metastasis process. This may help to explain why some invasive tumors are found to express a combination of SIBLINGs and their MMP partners.
In conclusion, it was shown that DMP1 can enhance the invasiveness of a model colon cancer cell in vitro by bridging MMP-9 to the cell surface in an RGD-dependent (with αvβ3 integrin, or αvβ5 integrin) and RGD-independent (with CD44) manner. Furthermore, it was shown that intact tumors can express all of these proteins within the same cellular structures suggesting that these complexes studied in vitro will likely be formed in vivo.
Acknowledgments
The authors express their gratitude for the assistance of Ms Li Li of NIDCR during the sectioning and immunostaining of the paraffin tissue blocks. This research was supported by the Intramural Research Program of the NIH, National Institute of Dental and Craniofacial Research.
References
- 1.Fisher LW, Torchia DA, Fohr B, Young MF, Fedarko NS. Flexible structures of SIBLING proteins, bone sialoprotein, and osteopontin. Biochem Biophys Res Commun. 2001;280(2):460–5. doi: 10.1006/bbrc.2000.4146. [DOI] [PubMed] [Google Scholar]
- 2.Ogbureke KU, Fisher LW. Expression of SIBLINGs and Their Partner MMPs in Salivary Glands. J Dent Res. 2004;83(9):664–70. doi: 10.1177/154405910408300902. [DOI] [PubMed] [Google Scholar]
- 3.Ogbureke KU, Fisher LW. Renal expression of SIBLING proteins and their partner matrix metalloproteinases (MMPs) Kidney Int. 2005;68(1):155–66. doi: 10.1111/j.1523-1755.2005.00389.x. [DOI] [PubMed] [Google Scholar]
- 4.George A, Sabsay B, Simonian PA, Veis A. Characterization of a novel dentin matrix acidic phosphoprotein. Implications for induction of biomineralization. J Biol Chem. 1993;268(17):12624–30. [PubMed] [Google Scholar]
- 5.MacDougall M, Gu TT, Luan X, Simmons D, Chen J. Identification of a novel isoform of mouse dentin matrix protein 1: spatial expression in mineralized tissues. J Bone Miner Res. 1998;13(3):422–31. doi: 10.1359/jbmr.1998.13.3.422. [DOI] [PubMed] [Google Scholar]
- 6.Ye L, Mishina Y, Chen D, Huang H, Dallas SL, Dallas M, et al. Dentin matrix protein 1 (Dmp1) deficient mice display severe defects in cartilage formation responsible for a chondrodysplasia-like phenotype. J Biol Chem. 2004 doi: 10.1074/jbc.M412911200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jain A, Karadag A, Fohr B, Fisher LW, Fedarko NS. Three SIBLINGs (small integrin-binding ligand, N-linked glycoproteins) enhance factor H's cofactor activity enabling MCP-like cellular evasion of complement-mediated attack. J Biol Chem. 2002;277(16):13700–8. doi: 10.1074/jbc.M110757200. [DOI] [PubMed] [Google Scholar]
- 8.Chaplet M, De Leval L, Waltregny D, Detry C, Fornaciari G, Bevilacqua G, et al. Dentin matrix protein 1 is expressed in human lung cancer. J Bone Miner Res. 2003;18(8):1506–12. doi: 10.1359/jbmr.2003.18.8.1506. [DOI] [PubMed] [Google Scholar]
- 9.Fisher LW, Jain A, Tayback M, Fedarko NS. Small integrin binding ligand N-linked glycoprotein gene family expression in different cancers. Clin Cancer Res. 2004;10(24):8501–11. doi: 10.1158/1078-0432.CCR-04-1072. [DOI] [PubMed] [Google Scholar]
- 10.Fedarko NS, Jain A, Karadag A, Fisher LW. Three small integrin binding ligand N-linked glycoproteins (SIBLINGs) bind and activate specific matrix metalloproteinases. Faseb J. 2004;18(6):734–6. doi: 10.1096/fj.03-0966fje. [DOI] [PubMed] [Google Scholar]
- 11.Mook OR, Frederiks WM, Van Noorden CJ. The role of gelatinases in colorectal cancer progression and metastasis. Biochim Biophys Acta. 2004;1705(2):69–89. doi: 10.1016/j.bbcan.2004.09.006. [DOI] [PubMed] [Google Scholar]
- 12.Ishida H, Murata N, Tada M, Okada N, Hashimoto D, Kubota S, et al. Determining the levels of matrix metalloproteinase-9 in portal and peripheral blood is useful for predicting liver metastasis of colorectal cancer. Jpn J Clin Oncol. 2003;33(4):186–91. doi: 10.1093/jjco/hyg035. [DOI] [PubMed] [Google Scholar]
- 13.Matsuyama Y, Takao S, Aikou T. Comparison of matrix metalloproteinase expression between primary tumors with or without liver metastasis in pancreatic and colorectal carcinomas. J Surg Oncol. 2002;80(2):105–10. doi: 10.1002/jso.10106. [DOI] [PubMed] [Google Scholar]
- 14.Roeb E, Dietrich CG, Winograd R, Arndt M, Breuer B, Fass J, et al. Activity and cellular origin of gelatinases in patients with colon and rectal carcinoma differential activity of matrix metalloproteinase-9. Cancer. 2001;92(10):2680–91. doi: 10.1002/1097-0142(20011115)92:10<2680::aid-cncr1622>3.0.co;2-7. [DOI] [PubMed] [Google Scholar]
- 15.Reinmuth N, Liu W, Ahmad SA, Fan F, Stoeltzing O, Parikh AA, et al. Alphavbeta3 integrin antagonist S247 decreases colon cancer metastasis and angiogenesis and improves survival in mice. Cancer Res. 2003;63(9):2079–87. [PubMed] [Google Scholar]
- 16.Fujisaki T, Tanaka Y, Fujii K, Mine S, Saito K, Yamada S, et al. CD44 stimulation induces integrin-mediated adhesion of colon cancer cell lines to endothelial cells by up-regulation of integrins and c-Met and activation of integrins. Cancer Res. 1999;59(17):4427–34. [PubMed] [Google Scholar]
- 17.Murray D, Morrin M, McDonnell S. Increased invasion and expression of MMP-9 in human colorectal cell lines by a CD44-dependent mechanism. Anticancer Res. 2004;24(2A):489–94. [PubMed] [Google Scholar]
- 18.Karadag A, Ogbureke KU, Fedarko NS, Fisher LW. Bone sialoprotein, matrix metalloproteinase 2, and alpha(v)beta3 integrin in osteotropic cancer cell invasion. J Natl Cancer Inst. 2004;96(12):956–65. doi: 10.1093/jnci/djh169. [DOI] [PubMed] [Google Scholar]
- 19.Matrisian LM. Cancer biology: extracellular proteinases in malignancy. Curr Biol. 1999;9(20):R776–8. doi: 10.1016/S0960-9822(00)80011-1. [DOI] [PubMed] [Google Scholar]
- 20.Forget MA, Desrosiers RR, Beliveau R. Physiological roles of matrix metalloproteinases: implications for tumor growth and metastasis. Can J Physiol Pharmacol. 1999;77(7):465–80. [PubMed] [Google Scholar]
- 21.Kleiner DE, Stetler-Stevenson WG. Matrix metalloproteinases and metastasis. Cancer Chemother Pharmacol. 1999;43(Suppl):S42–51. doi: 10.1007/s002800051097. [DOI] [PubMed] [Google Scholar]
- 22.Yu Q, Stamenkovic I. Localization of matrix metalloproteinase 9 to the cell surface provides a mechanism for CD44-mediated tumor invasion. Genes Dev. 1999;13(1):35–48. doi: 10.1101/gad.13.1.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Deryugina EI, Bourdon MA, Luo GX, Reisfeld RA, Strongin A. Matrix metalloproteinase-2 activation modulates glioma cell migration. J Cell Sci. 1997;110(Pt 19):2473–82. doi: 10.1242/jcs.110.19.2473. [DOI] [PubMed] [Google Scholar]
- 24.Deryugina EI, Bourdon MA, Jungwirth K, Smith JW, Strongin AY. Functional activation of integrin alpha V beta 3 in tumor cells expressing membrane-type 1 matrix metalloproteinase. Int J Cancer. 2000;86(1):15–23. doi: 10.1002/(sici)1097-0215(20000401)86:1<15::aid-ijc3>3.0.co;2-b. [DOI] [PubMed] [Google Scholar]
- 25.Albini A, Iwamoto Y, Kleinman HK, Martin GR, Aaronson SA, Kozlowski JM, et al. A rapid in vitro assay for quantitating the invasive potential of tumor cells. Cancer Res. 1987;47(12):3239–45. [PubMed] [Google Scholar]
- 26.Hirst KL, Ibaraki-O'Connor K, Young MF, Dixon MJ. Cloning and expression analysis of the bovine dentin matrix acidic phosphoprotein gene. J Dent Res. 1997;76(3):754–60. doi: 10.1177/00220345970760030701. [DOI] [PubMed] [Google Scholar]
- 27.Becker TC, Noel RJ, Coats WS, Gomez-Foix AM, Alam T, Gerard RD, et al. Use of recombinant adenovirus for metabolic engineering of mammalian cells. Methods Cell Biol. 1994;43(Pt A):161–89. doi: 10.1016/s0091-679x(08)60603-2. [DOI] [PubMed] [Google Scholar]
- 28.Fedarko NS, Fohr B, Robey PG, Young MF, Fisher LW. Factor H binding to bone sialoprotein and osteopontin enables tumor cell evasion of complement-mediated attack. J Biol Chem. 2000;275(22):16666–72. doi: 10.1074/jbc.M001123200. [DOI] [PubMed] [Google Scholar]
- 29.Giambernardi TA, Grant GM, Taylor GP, Hay RJ, Maher VM, McCormick JJ, et al. Overview of matrix metalloproteinase expression in cultured human cells. Matrix Biol. 1998;16(8):483–96. doi: 10.1016/s0945-053x(98)90019-1. [DOI] [PubMed] [Google Scholar]
- 30.Brooks PC, Stromblad S, Sanders LC, von Schalscha TL, Aimes RT, Stetler-Stevenson WG, et al. Localization of matrix metalloproteinase MMP-2 to the surface of invasive cells by interaction with integrin alpha v beta 3. Cell. 1996;85(5):683–93. doi: 10.1016/s0092-8674(00)81235-0. [DOI] [PubMed] [Google Scholar]